thesis for the degree ראותהתלבקםשלרובח doctor of ... · thesis for the degree...
TRANSCRIPT

1
Thesis for the degree Doctor of Philosophy
חבור לשם קבלת התואר
דוקטור לפילוסופיה
By Nava Levit Binnun
Advisor Prof. Elisha Moses
January 2007
Submitted to the Scientific Council of the Weizmann Institute of Science
Rehovot, Israel
חקירת רשת קוגניטיבית במוח בריא ובמוח עם סכיזופרניה באמצעות סטימולציה מגנטית טראנס גולגלתית
Probing a Cognitive Network in the Brain with and without Schizophrenia Using Transcranial Magnetic
Stimulation
מאת
'
נאוה לויט בן נון
מנחהאלישע מוזס פרופ
של המדעית מוגש למועצהלמדע מכון ויצמן
ישראל, רחובות
ז"תשסשבט


Acknowledgments
This work is dedicated with love
To my mother, Shoshana Levit To my mother- in-law, Avigail Binnun
and
To my son Uri
Watching you grow and develop Taught me more than anything
About the greatness of the human mind
I would like to thank warmly the many people who contributed to my project.
To my advisor, Elisha Moses, for enabling me much freedom, for encouraging me to
be creative and for supporting me at difficult stages.
To Dr. Avi Peled, for discussions that were the starter of all this work and for all the
time and support in the experiments.
To Dr. Nestor Handzy, for his assistance and central contribution.
To all the lab members over the years: Stephan Thiberge, Ilan Breskin, Ofer
Feinerman, Dudu Biron, Assaf Rotem, Shimhson Jacobi, Jordi Soriano-Fradera and
Enric Alvarez-Lacalle – for all the help and pleasant atmosphere. Special thanks to
Jordi who volunteered his brain for me.
To Amos Arieli for lending me the EEG system and spending much time listening to
my ideas and assisting me with analysis.
To Ehud Ahissar and Moshe Fried for letting me use their EEG equipment and for the
flexibility they enabled me in its use.
To Assaf Pressman that helped in the first stages to understand the EEG signals, with
programming and finally with getting the EMG system.
To Enrico Segre for constructing the Labview software used in the experiments in a
very user friendly manner.
To the medical team in Sha'ar Menashe for the support in experiments with patients
with Schizophrenia.
To all the people that participated as subjects in my experiments for trying as best as
they could to do the tasks.
To the people supporting the research in our departments: Israel Gonen, Rachel
Goldman, Malka Paz, Perla Zalcberg, Yuri Shimoni, Yossi Shopen, Gershon Elazar,
2

Yossi Leibovitz, Yossi Drier - for the pleasant atmosphere, for always giving a hand,
solving problems with all there hearts, and always with a smile and good will. I wish
myself that I will have the fortune to work in such atmosphere also in the future.
To Yohai Kroovi for the supply of tasty Guavas...
To Yuri Shimoni for the bulding of a TMS from scrap upon my request and providing
SOS help.
A special thanks to Yossi Drier for always being there to help with a smile and for all
the insightful discussions about life. You are a model for me.
To Assaf Weiss, Roy Taragan and Yuval Hart for assisting with the programming and
analysis.
To Edna Schechtman for assistance with the statistics even outside office hours.
To Prof. Tamar Flash and Dr. Dahlia Sharon for fruitful discussions.
To my PhD committee, Prof. Rafi Malach and Dr. Roy Bar Ziv for the time.
To Ami Shalit for the flexible attitude.
To my Yoga and Meditation teachers throughout the years, Nira Yorav, Naomi Eini,
Tanisara, Kitisaro, Patricia and Charles Jeneou, for the opportunity to understand
deeply what I believe is the true human experience.
To my family:
To my mother, Shoshana: this Phd is yours.
To my father, Shimon, for the support and the good advice that helped me finish this
long journey.
To my mother in law, Avigail, for all the invaluable help with the kids that gave me
the time and quiet state of mind to work.
To my father in law, Meno, for the curiosity and interest and for the great help with
Uri.
To my flowers, Uri and Yoni, who were born during this work, for giving me the
greatest experience of watching a person develop.
And last but not least -
To my dear husband Yariv, for all the support, freedom and space to make my
decisions.
3

Summary
Schizophrenia is a devastating disease which affects 1% of the world population, at
the beginning of adult life, creating an enormous economic burden on society and a
major grief to the surrounding family. Leading theories in Schizophrenia research
suggest that brain connectivity deficits, both over-connectivity and under-
connectivity, underlie this disease. Schizophrenia is sometimes described as a
‘breakdown of consciousness’, affecting almost every aspect involved in the complex
experience of being human. Thus understanding the pathology of Schizophrenia may
provide insight to how the ‘normal’ human experience arises from the biological
architecture of the brain.
The main goal of this study was to utilize Transcranial Magnetic Stimulation (TMS)
perturbations to study the connectivity and stability of a distributed cognitive network
in the brain of both healthy people and people with Schizophrenia. Transcranial
Magnetic Stimulation is a non-invasive method in which a strong pulsed magnetic
field applied at the scalp produces electric currents inside the brain. The magnetic
field decreases rapidly with distance enabling effective stimulation only of areas
relatively close to the surface of the cortex.
TMS was used in an externally paced finger tapping task, perturbing the internal
network oscillations invoked by the finger motion as it keeps pace with a metronome.
Although a relatively simple task, it induces activation of a distributed neural network
of frontal, sensorimotor, cerebellar and deep brain areas that underlie brain functions
of volition, awareness of action, attention, timing, motor planning, motor execution,
sensory-feedback and error monitoring mechanisms. TMS perturbations were
synchronized to the metronome and applied to the network at the level of the primary
motor cortex (M1).
4

First we were able to show, in both healthy and Schizophrenia subjects, that TMS
perturbation could separate different components of the network. TMS perturbations
profoundly affected the motion of the finger, resulting in two abnormal behaviors that
subjects were unaware of; a doubling of the frequency of the tap and a stalling of the
finger for half the period. Tapping accuracy itself was surprisingly not affected,
suggesting that the timing process involved in controlling the tapping movement is
separate from the motor processes in charge of execution of the motor commands.
In subjects with Schizophrenia TMS perturbations permeated to other components of
the network, altering perception of timing accuracy and invoking high-level
deficiencies related to attention and volition in the form of lapses. These lapses,
defined as little to no movement of the finger during a metronome period, were
observed when these subjects were tapping with the TMS perturbation.
We then performed the TMS perturbations described above while manipulating in
parallel putative components of the network underlying the finger tapping task. In one
condition attention was deviated away from the tapping task into a parallel mental
arithmetic task. In the second condition the left dorsolateral prefrontal cortex (L-
DLPFC) was inhibited by repetitive TMS prior to the tapping task, and lasted
presumably during the whole experiment.
In healthy subjects the effect of TMS perturbations to M1 following these
manipulations was no more only on the motor component, but also on timing and
awareness of finger. This suggests that our manipulations changed the stability of the
network causing the TMS perturbations to the motor component to diffuse to other
components.
In people with Schizophrenia, these manipulations similarly enhanced the effect of the
TMS perturbations on different components of the system. The lapses, which
appeared only in patients with Schizophrenia, were significantly aggravated by the
attention shifting condition. L-DLPFC inhibition caused a significant improvement in
the performance of the finger when it tapped together with the TMS perturbations and
seemed to reduce the probability of occurrence of lapses. The DLPFC has previously
5

been associated with high executive functions and altered activation in it appears to be
specific to the disease process of Schizophrenia. The fact that lesioning it improved
finger performance of subjects with Schizophrenia and affected possible error
correction, timing and attention processes, suggests that the L-DLPFC is a link in a
network governing finger tapping, which is mal-functional in Schizophrenia, and that
removal of the L-DLPFC component alleviates its dysfunction.
Using EEG (ElectroEncephaloGram) to measure the response of global brain activity
to TMS perturbations in healthy subjects and subjects with Schizophrenia, we found
differences in both amplitude and timing of the EEG responses evoked by TMS.
These differences are in line with recent results (Massimini et al., 2005) obtained with
more sophisticated tools and provides further support that TMS evoked potentials can
be used to study changes in brain connectivity and responsiveness in Schizophrenia.
The results obtained in both setups support the disconnection hypothesis of
Schizophrenia (Friston, 1998; Andreasen et al., 1999) and demonstrate the ability to
manipulate and probe distinct components of a distributed cognitive network using
TMS.
Results are discussed in terms of their contribution to the understanding how the
balance and stability of interactions between networks in the brain can account for the
vast spectrum of human conditions.
6


Table of Contents
LIST OF ABBREVIATIONS................................................................................................................9
INTRODUCTION AND MOTIVATION...........................................................................................10
Wiring patterns in the brain resemble a complex process of tinkering ..................................................10 Observations of abnormal human conditions.........................................................................................12 Research motivation ...............................................................................................................................13 BACKGROUND...................................................................................................................................14
NETWORKS AND COGNITION ...................................................................................................................14 Two principles of organization...............................................................................................................14 Anatomical networks ..............................................................................................................................14 Functional connectivity ..........................................................................................................................15 PERTURBATIONS AS A TOOL TO STUDY BRAIN-BEHAVIOUR RELATIONSHIPS............................................15 Brain lesions, electrical stimulation, local cooling, anaesthetic microinjection....................................15 Transcranial Magnetic Stimulation........................................................................................................16 SCHIZOPHRENIA ......................................................................................................................................16 Defining symptoms .................................................................................................................................17 Possible causes.......................................................................................................................................17 Conscious integration and Schizophrenia – the 'disconnection hypothesis'...........................................19 Medication..............................................................................................................................................20 FINGER TAPPING .....................................................................................................................................20 Synchronization task activates distributed areas in the brain ................................................................21 Models ....................................................................................................................................................21 Cerebellum is a key player in event timing.............................................................................................21 Perturbations to finger tapping ..............................................................................................................22 TRANSCRANIAL MAGNETIC STIMULATION..............................................................................................22 Pulsed magnetic field induces electric field in the brain........................................................................22 The effects of TMS on neuronal tissue....................................................................................................22 Spatial and temporal resolution .............................................................................................................23 Controlling for sensory effects ...............................................................................................................24 TMS IN THE STUDY OF BRAIN DYNAMICS ................................................................................................25 Extensive use of TMS..............................................................................................................................25 Studying cortical connectivity and dynamics .........................................................................................25 TMS IN THE STUDY OF SCHIZOPHRENIA..................................................................................................26 MAIN RESEARCH GOALS...............................................................................................................27
MAIN RESULTS..................................................................................................................................28
CHAPTER 1: TRANSCRANIAL MAGNETIC STIMULATION IN A FINGER TAPPING
TASK SEPARATES MOTOR FROM TIMING MECHANISMS AND INDUCES FREQUENCY
DOUBLING ..........................................................................................................................................30
INTRODUCTION........................................................................................................................................30 MATERIALS AND METHODS ....................................................................................................................31 Subjects...................................................................................................................................................31 Design.....................................................................................................................................................31 Measurement of finger motion................................................................................................................33 TMS parameters .....................................................................................................................................33 Recording and Analysis..........................................................................................................................35 Statistical Analysis .................................................................................................................................36 RESULTS..................................................................................................................................................36 DISCUSSION.............................................................................................................................................42 FIGURES AND TABLES ............................................................................................................................49
7

CHAPTER 2: TRANSCRANIAL MAGNETIC STIMULATION AT M1 DISRUPTS
COGNITIVE NETWORKS IN PATIENTS OF SCHIZOPHRENIA.............................................59
INTRODUCTION........................................................................................................................................59 MATERIALS AND METHODS ....................................................................................................................60 Subjects...................................................................................................................................................60 Statistical analysis ..................................................................................................................................62 RESULTS..................................................................................................................................................62 DISCUSSION.............................................................................................................................................67 FIGURES AND TABLES .............................................................................................................................71 CHAPTER 3: PROBING CONNECTIVITY AND STABILITY IN THE SCHIZOPHRENIA
BRAIN USING TMS PERTURBATIONS.........................................................................................77
INTRODUCTION........................................................................................................................................77 MATERIALS AND METHODS ....................................................................................................................79 I. Finger Tapping Experiment ................................................................................................................79 Subjects...................................................................................................................................................80 Design.....................................................................................................................................................80 Creating a virtual lesion with TMS ........................................................................................................82 Recording and Analysis..........................................................................................................................82 Statistical analysis ..................................................................................................................................82 II. EEG Experiment. ...............................................................................................................................83 Subjects...................................................................................................................................................83 Design.....................................................................................................................................................83 Data analysis..........................................................................................................................................84 RESULTS..................................................................................................................................................86 I. Finger tapping experiment ..................................................................................................................86 II. EEG experiment.................................................................................................................................90 DISCUSSION.............................................................................................................................................91 FIGURES AND TABLES .............................................................................................................................95 DISCUSSION – STABILITY AND BALANCE IN NETWORKS ................................................105
From Schizophrenia to Autism .............................................................................................................108 CONCEPT OF EMERGENCE AND THE IMPORTANCE OF STABILITY AND BALANCE....................................109 The ‘Dynamicist’ view..........................................................................................................................109 Evidence for applicability of dynamical system approach to the brain................................................110 BALANCE WITHIN SENSORY PROCESSING UNDERLIE TEMPERAMENT AND BEHAVIOUR TRAITS ..............111 Occupational Therapy research can provide insight ...........................................................................111 Different sensory processing profiles across people ............................................................................111 A model for sensory processing............................................................................................................112 Sensory processing patterns are different for people with disorders ...................................................113 Relation between sensory processing and temperament and personality traits ...................................114 Evidence from Practice ........................................................................................................................115 WHAT ABOUT A ‘SELF’ WITH A FREEWILL? ...........................................................................................115 Examples of evidence supporting monism............................................................................................116 A PHILOSOPHICAL NOTE ........................................................................................................................118 The possible contribution of the Buddhist tradition to the scientific enterprise. ..................................119 BIBLIOGRAPHY ..............................................................................................................................120
INDEPENDENT COLLABORATION ............................................................................................132
8

List of Abbreviations
TMS – Transcranial Magnetic Stimulation rTMS – repetitive Transcranial Magnetic Stimulation DLPFC - dorsolateral prefrontal cortex L-DLPFC – left dorsolateral prefrontal cortex MT- Motor threshold rMT – resting motor threshold CV – coefficient of variation
9


Introduction and Motivation
Our life is a constant stream of experiences. I see, hear, taste, smell, touch, feel and
think. I can be angry, pleased, tired, self absorbed, self agonising, worried, interested,
and moody. Strong feelings overwhelm me. Praise makes me feel more worthy,
criticism makes me feel unworthy. A loss of a loved one can make me feel destroyed.
I feel I have a unique identity: my own personality, memories, recollections, plans,
my own point of view and anticipations of the world.
What is this self, this ego-centre, that appears and disappears, that seems so constant
and yet so fragile, so familiar and yet so elusive? Moreover, why do pathologies in the
brain, as in the case of Schizophrenia, result in a feeling of a breakdown of the ‘Self’
and of one’s personality?
These questions are at the core of Neuroscience and are the primary reason and
motivation for me to choose this field of study.
In this thesis I would like to contribute to a notion that is steadily growing in the
scientific discourse: that we are nothing more then the result of emergent properties
(Thompson and Varela, 2001) arising from the stability, balance, regulation and
interplay of many interacting networks in our brains. The variability in temperament
and personality between people is due to the variability in mechanisms that balance
inhibition and excitation (Rubenstein and Merzenich, 2003), giving rise to variability
in sensory processing (Dunn, 2001) and to variability in the relative distribution of
processing resources. Abnormalities in the balance between inhibition and excitation
give rise to abnormalities in behavior such as those observed in Schizophrenia and
Autism (Friston, 1998; Rubenstein and Merzenich, 2003).
Wiring patterns in the brain resemble a complex process of tinkering
The wiring patterns in the brain seem to lend support to the view described above. A
closer look at the anatomical wirings provide little proof to the sequential information
processing description of the brain, whereby information enters through the primary
sensory areas and advances via a hierarchy of processing stages, with the primary
10

motor cortex serving as a major gateway for the output of these computations. Rather,
connections from one level of processing to the other are reciprocal and allow higher
synaptic levels to influence earlier levels through feedback connections. In each
cortical area afferent synapses converge and efferent synapses diverge, supporting
both parallel as well as serial processing, allowing each sensory event to initiate
multiple cognitive and behavioral processes (Mesulam, 1998).
An example for this is the first stages of visual processing. Connections come from
the eyes, via the optic nerve, to the thalamus (considered to be the major relay station
in the brain), on to the lateral geniculate nucleus (LGN) and from there to the visual
cortex. Eighty percent of what a LGN cell listens to comes not from the retina but
rather from the dense interconnectedness of other regions in the brain. Moreover,
there are more fibres coming from the cortex down to the LGN than there are going in
the reverse direction (Churchland and Sejnowski, 1988).
This is generally true also in other unimodal sensory areas where the veridical
representation of experience is encoded only in the first few synaptic levels
(Mesulam, 1998). Attentional, motivational and emotional modulations become
increasingly more pronounced from almost the very first stages of processing to the
more downstream levels and help to create a highly edited subjective version of the
world (Mesulam, 1998). These modulations occur via connections coming from the
amygdala, the prefrontal cortex and the limbic areas. Interactions between the external
world and the internal world are allowed via massive connections from the
hypothalamus. Working memory, allowing present events to enter into associative
interactions with past experience, is probably mediated by a complex network of
connections that link the hippocampus to different cortical regions (Mesulam, 1998).
This or similar descriptions of the wiring patterns in the brain, portray a patchwork of
subnetworks, interconnected in a complex process of tinkering rather than in a cleanly
designed unified system. These dense interconnections suggest that everything that is
going on will eventually be a function of what all the components are doing. In this
case it seems that the relative contribution of the different subnetworks to the whole
system arising from the balance, regulation and stability of the local and global
networks should be a key factor in understanding how the rich human experience can
arise from such a biological system.
11

Observations of abnormal human conditions
The concept of stability and balance between excitation and inhibition can be found in
studies of abnormal human conditions that give rise to abnormal and extreme
behaviours, as in the case of Schizophrenia and Autism (Friston, 1998; Rubenstein
and Merzenich, 2003; Stephan et al., 2006). A better understanding of such disorders
should provide insight into the understanding of how the healthy brain gives rise to
what is considered the ‘normal’ human condition.
Schizophrenia is sometimes described as the ‘breaking down of consciousness and
feeling of ‘self’’ (Andreasen, 1997; Churchland, 2002) as the following poem by
Emily Dickenson (who probably suffered from Schizotypal personality disorder
(Winhusen, 2004)) may describe:
I felt a Cleaving in my Mind-
As if my Brain had split-
I tried to match it – Seam by Seam-
But could not make them fit.
The thought behind, I strove to join
Unto the thought before-
But Sequence ravelled out of Sound
Like Balls-Upon a Floor.
-Emily Dickinson "Poem 937" (Taken from (Andreasen, 2001))
Patients with Schizophrenia seem to lose the ability to judge reality, to distinguish
between external and internal events, to understand the other’s intentions and to act
correctly upon them. The disease is characterized by a profound degradation in
cognitive abilities, in motivation and volition.
Autism is defined as a triad of impairments in social interaction, communication and
behavioral flexibility (Rippon et al., 2006). The Autistic spectrum encompasses high
functioning individuals with Asperger’s syndrome as well as those severely impaired
in language and intellectual development.
Interestingly, these pathologies have much in common. Both are described as
heterogeneous disorders, with deficits encompassing practically all brain functions –
12

sensory, emotional, social and cognitive functions (Moldin et al., 2006). In both
disorders, abnormalities in gamma band, considered a main candidate for long
distance synchronization in the brain, are observed (Spencer et al., 2003a; Spencer et
al., 2004; Brown et al., 2005). Both disorders are considered a neurodevelopmental
disorder, suggesting that something is abnormal in the synaptic and plastic processes
during development. In both, abnormalities in gating and regulation of sensory
processing are observed (Dunn, 2001; Brown et al., 2002). In both an interaction of
genetic and environmental factors, each by itself not enough to cause the pathology,
probably underlies the abnormal development (Rubenstein and Merzenich, 2003).
Not surprisingly therefore is the fact that the leading theories today in both
pathologies suggest abnormalities in connectivity – either over connectivity or
dysconnectivity or both (Friston, 1998; Rubenstein and Merzenich, 2003; Rippon et
al., 2006). An imbalance in connectivity is suggested both within subnetworks and
across networks (Rubenstein and Merzenich, 2003; Rippon et al., 2006). This
suggests that in order for the brain to produce behavior that is considered within the
average range, the right balance between connectivity and dysconnectivity needs to be
sustained.
It is of interest that therapists working with Autism and Schizophrenia are
increasingly finding the most efficient interventions to be those that are based on an
integration of perceptual and movement training (Rubenstein and Merzenich, 2003),
together with regulation of sensory processing and emotional and motivational
processing (Greenspan and Wieder, 1999). The fact that these interventions are
reporting relatively high success rates (Case-Smith and Miller, 1999; Baranek, 2002;
Smith et al., 2005), suggests that they may reflect some basic principles of brain
organization.
Research motivation
The motivation for my research arose from an attempt to integrate the above
observations. Together they seem to suggest that understanding the integration of
distributed brain functions into networks and the delicate and subtle interplay between
them may provide important clues to the intriguing question of how the vast spectrum
13

of human behaviors and experiences can arise from the neural architecture in the
brain.
In this thesis I demonstrate the possibility of disturbing the stability of a simple
network underlying the paced finger tapping task and how this disturbance can change
behavior and affect other cognitive processes. We study both the normal brain system
and the presumably dysconnected Schizophrenia brain system. Comparing them
enables us to better understand how the normal brain gives rise to various cognitive
abilities and how the disease process disrupts them.
Background
Networks and Cognition
Two principles of organization
Numerous anatomical, brain lesion, electrophysiology and functional neural imaging
studies have made clear that functional segregation is a principal of organization in
the brain (Friston, 2002a, b; Paus, 2005). However, this cartography by itself fails to
explain how the brain produces its complex cognitive functions. A second principle of
organization arising from the different studies is that of functional integration
(Friston, 2002a). Functional coupling and integration can be achieved via direct
anatomical connections (Mesulam, 1998), via temporal correlation between remote
neurophysiological events (Friston, 2002a) or via mechanisms of synaptic
transmission and plasticity (Stephan et al., 2006).
Anatomical networks
Studies of the wiring patterns of the brain identified at least five anatomically distinct
networks (Mesulam, 1998) (of the many that probably exist): the network underlying
spatial awareness (involving posterior parietal cortex and frontal eye fields); the
language network (involving Wernicke’s and Broca’s areas); the explicit
memory/emotion network (involving hippocampal-entorhinal complex and the
14

amygdale); the face-object recognition network (involving midtemporal and
temporopolar cortices); and the working memory-executive function network (lateral
prefrontal cortex and perhaps the posterior parietal cortex). Individual sensory
processes give rise to streams of processing directed to nodes belonging to each of
these networks and connected to many other brain areas.
Functional connectivity
Functional integration between many different brain areas, underlying complex
cognitive functions, can also arise without direct anatomical connections. Neural
synchrony has been suggested as a candidate for such integration and coordination
through a kind of temporal resonance or "glue" (Engel et al., 2001). This synchrony is
mediated by neuronal groups that oscillate in specific bands and enter into precise
phase-locking over a limited period of time (Varela et al., 2001a). This role of
synchronization of neuronal discharges has been greatly highlighted by results from
microelectrodes in animals (Gray, 1999; Singer, 1999; Engel et al., 2001; Engel and
Singer, 2001). These animal studies have been complemented by studies in humans
using scalp EEG or MEG (Tallon-Baudry et al., 2001), subdural EEG (Le Van Quyen
et al., 1997; Lachaux et al., 2000), fMRI and PET. Recent evidence demonstrated
(Rodriguez et al., 1999) that large-scale integration implicates not only the
establishment of dynamical links but also their dismantling, to give way for a next
cognitive moment. Synchronous patterns of short epochs lasting 50 to 250 ms were
observed with new patterns continually being created, destroyed and subsequently
recreated.
Perturbations as a tool to study brain-behaviour relationships
Brain lesions, electrical stimulation, local cooling, anaesthetic microinjection
Physical perturbations to neural activity have been used by neuroscientists since the
beginning of neuroscience. Irreversible perturbations in the form of brain lesions have
informed us about the essential role of different areas in the brain in various functions
(e.g Broca for language, hippocampus for memory). Reversible perturbation of neural
15

activity with direct electrical stimulation revealed, for example, the somatotopic
organization of the motor cortex (Paus, 2005).
More recently reversible inactivation of specific brain regions has been achieved in
experimental animals using local cooling or local microinjections of anaesthetics and
neurotransmitter agonist/antagonists (Payne and Lomber, 1999).
Intracortical microstimulation has allowed manipulation of the activity of small
groups of neurons with high spatial and temporal precision. These works have been
applied, among others, to the study of attention, visual and somatosensory perception
and studying contextual effects on perception (reviewed by Cohen & Newsome
2004).
Most studies used the various perturbation techniques to assess the local change in
neural activity at the site of perturbations, or to study the effect on some cognitive
function (Cohen and Newsome, 2004). Several investigators studied the effect of
perturbation (e.g. cooling) in one area on adjacent or distal areas (Kotter and Sommer,
2000), or on the rest of the brain (Vanduffel et al., 1997).
Transcranial Magnetic Stimulation
A non invasive tool to stimulate the human brain at a local cortical area is the
technique of Transcranial Magnetic Stimulation (TMS). Coupled with EEG (Paus,
2001b), PET (Chouinard et al., 2003; Paus, 2005) or fMRI (Paus, 2001b) it enables
mapping of the neural connections in the living human brain. Inhibitory or excitatory
effects in a local neuronal population can be induced using the various stimulation
modes of the TMS (see below) and its effect on distributed brain areas can be
simultaneously measured.
Schizophrenia
Schizophrenia is considered the most devastating mental illness. It is estimated to
affect 1% of the world population, striking typically during the late teens and early
twenties (Andreasen, 2001), preventing its victims from fully participating in society
16

and causing great grief to their families. Moreover, Schizophrenia creates an
enormous economic burden, costing society billions of dollars annually.
Defining symptoms
The defining symptoms of Schizophrenia are hallucinations, delusions, thought
disorder, odd behaviour – which were termed “positive” symptoms - and lack of
content of speech, blunted affect, social withdrawal, and the lack of motivation and
goal directed behavior – which were termed “negative” symptoms. Disease onset is
characterized by an episode of psychosis followed by relapses which are usually
treated with antipsychotic medication. Despite the treatment, patients continue to
suffer from a reduction in cognitive abilities and the majority of them can not return
to work or school and have minimal social interactions.
Core cognitive disturbances of this disorder, typically the main cause of suffering to
the patients, include memory and attention deficits, diminished capacity to regulate
willed and stimulus driven action systems (Frith et al., 2000; Torres et al., 2004),
deficient time estimation (Elvevag et al., 2003); and psychomotor disturbances
(Manschreck et al., 1985; Flyckt et al., 1999; Cortese et al., 2005).
Possible causes
Brain abnormalities have been identified in Schizophrenia and implicate multiple
distributed regions in the brain: frontal cortex, temporal cortex, thalamus,
hippocampal complex, basal ganglia and even cerebellum (Andreasen, 2000; Ho et
al., 2004).
Twin studies indicate that there is a genetic factor involved in Schizophrenia. A
monozygotic twin has a 40% probability of developing Schizophrenia if his/her twin
brother or sister has Schizophrenia while a dizygotic twin has only 10% probability.
However genetics alone cannot explain the disease and environmental factors are also
involved.
17

Dopamine supersensitivity occurs in Schizophrenia and has led to the hypothesis that
mal function of dopamine systems may contribute to the symptoms of the disease.
This hypothesis, known as 'The Dopamine Hypothesis of Schizophrenia' suggests that
increases in the level of dopamine in the brain can cause psychosis and that
overactivity in the dopamine systems in the mesolimbic and mesocortical pathways
may contribute to positive and negative symptoms, respectively. Indeed, drugs such as
cocaine and amphetamine, which increases dopamine levels in the brain can cause
psychosis. Additional support to the theory can from the discovery of antipsychotic
drugs that antagonized dopamine binding and reduced positive psychotic symptoms.
A new generation of antipsychotic drugs (called the atypical antipsychotics – see
below) challenged the view that the amount of dopamine blocking was correlated to
clinical benefit. These drugs were just as effective as the dopamine-related
antipsychotics in controlling psychosis, but actually blocked fewer dopamine
receptors. Moreover, they seem to be more effective in the treatment of the negative
symptoms. In addition, the excitatory neurotransmitter Glutamate was also found
effective in treatment of Schizophrenia symptoms. Thus it is clear today that the mal
function of the dopamine system is associated with the initial, acute part of the disease
related to the positive symptoms. It cannot, however, explain by itself how
Schizophrenia or psychosis events are triggered and it is apparent that environmental,
developmental and social factors should also be considered (Seeman et al., 2005). The
drastic deterioration in a wide spectrum of cognitive abilities observed in
Schizophrenia patients suggests that explanation of all aspects of this illness, both
positive but especially the negative ones, requires a more system-view approach (see
below).
Evidence suggesting that Schizophrenia is a neurodevelopmental disorder is
substantial and steadily increasing (Andreasen et al., 1999; Andreasen, 2000;
Andreasen, 2001). Factors affecting multiple stages of brain development are both
genetic and environmental (Rubenstein and Merzenich, 2003). Correlation to head or
birth injuries, viral infections, exposure to toxins and drugs of abuse, hormonal
changes and other factors have been demonstrated (Andreasen, 2001). Since
Schizophrenia typically develops in the late teens and early twenties, a stressful
period in life when a young person must learn to fly out of the parental nest and live
18

his own life, it is possible that the most critical abnormality in brain development
occurs during this final stage of "growing up" (Andreasen, 2001; Peled, 2004).
Conscious integration and Schizophrenia – the 'disconnection hypothesis'
Conscious integration is thought to be disrupted in Schizophrenia. When asked to
describe their suffering, patients with Schizophrenia tend to say: "My thinking is
confused; My ideas don’t seem to connect right; I have trouble filtering out
unimportant information; I feel bombarded by stimuli" (Andreasen, 2001).
Interestingly, both theories of Conscious integration and theories of Schizophrenia
(Shaw et al., 1983; Dolan et al., 1999; Peled and Geva, 2000; Peled et al., 2001) are
associated with the integration of large-scale cortical systems. While conscious
integration is mediated by the establishment of dynamical links and their dismantling,
integrating frontal, parietal, temporal, and occipital areas (Tononi and Edelman,
1998), Schizophrenia is thought to arise from a disturbance in these dynamics
(Friston, 1996; Dolan et al., 1999; Friston, 1999; Peled, 1999b; Tononi and Edelman,
2000; Garcia-Toro et al., 2001; Hoffman and McGlashan, 2001). Disturbance in
dynamics is thought to arise from mal functional effective connectivity and has been
known as “the disconnection hypothesis” (Friston, 1999). This disturbance in
dynamics is supported by dysfunction in synchronous gamma band (~40Hz) activity
(a candidate mechanism for the long-range integration), (Lee et al., 2003; Spencer et
al., 2003b; Spencer et al., 2004), impairment in evoked potentials (Kasai et al., 1999;
Papageorgiou et al., 2001), decreased functional connectivity in the theta-frequency
(4-7Hz) (Koenig et al., 2001), and abnormalities in slow wave activity (Harris et al.,
2001) and in beta activity (14-30Hz) (Itil, 1977). Using PET and fMRI, abnormal
functional connectivity between temporal and frontal regions (Stephan et al., 2006)
and also in cortico-cerebellar-thalamo-cortical circuits was measured (Andreasen et
al., 1996).
The alternative to the disconnection hypothesis (Friston, 1999), is a regionally
specific patho-physiology in one or more neuronal regions in the Schizophrenia brain.
In other words, not pathological interactions between normal regions, but rather a
normal interaction between two or more pathological brain regions. Indeed, the
evidence for abnormal anatomical interruption in connections in Schizophrenia that
19

can support the diconnection hypothesis is not substantial. In addition, many of the
deficits of Schizophrenia can be explained by regionally specific abnormalities of
function or physiology (Friston, 1999). Although these abnormalities could also be
described as abnormalities in connectivity, they do not in themselves provide
evidence for the disconnection hypothesis. Thus, the disconnection hypothesis,
although appealing in many respects, still remains to be proven.
Medication
People with Schizophrenia are treated with antipsychotic medication that affects the
dopamine receptors. Although carrying much hope initially, it was found to be
effective mostly for the treatment of the positive symptoms (delusions and
hallucinations), but does not help patients overcome the negative symptoms and has
many unpleasant side effects. Antipsychotics of a new generation were developed
recently and target other neurotransmitters besides dopamine, and carry more hope as
they work not just on positive psychotic symptoms but also seem to improve negative
symptoms. Although considered first choice treatment today they too have side
effects, such as excessive weight gain, increased tendency to develop diabetes
mellitus, and an effect on the endocrine system (Andreasen, 2001).
Finger Tapping
The synchronized finger taping task has been used since 1886 (Stevens, 1886) to
study aspects of action timing and biological rhythms and to investigate the neural
basis of timing in the brain. In this task, participants are asked to tap in synchrony
with a pacing signal, usually an auditory metronome. Detailed dynamical description
(Kay et al., 1991), analysis of timing errors (Aschersleben and Prinz, 1995; Repp,
2001a), movement trajectories (Balasubramaniam et al., 2004), natural tapping rates
(Collyer et al., 1994), and contribution of tactile reafference (Drewing et al., 2002)
have been conducted.
20

Synchronization task activates distributed areas in the brain
Imaging studies during finger synchronization have identified activation of the
primary motor cortex, premotor cortex, auditory cortex, sensorimotor cortex,
supplementary cortex, basal ganglia, thalamus, the cerebellum, cingulated regions,
prefrontal and temporal association cortices (Lepage et al., 1999; Mayville et al.,
2002; Dhamala et al., 2003; Lewis et al., 2004). These areas can be crudely divided
into lower level components in charge of controlling the finger movement and pace
(e.g. the sensory processing of the auditory stimuli, the timer and the motor
programming and execution components) and higher level components in charge of
attentional, volitional and working memory aspects of the task. These two
subnetworks, together with the peripheral network controlling the muscles comprise
the network underlying the finger tapping task.
Models
Studying the variability in the tapping has led to various models (Vorberg and Wing,
1996; Semjen et al., 2000; Engbert et al., 2002) regarding the source of variability in
tapping and the mechanisms underlying the ability of humans to synchronize to a
rhythmic movement (Aschersleben and Prinz, 1995). The leading model (Wing and
Kristofferson, 1973) suggests two distinct components: a central time keeper that
provides a series of temporal intervals and a peripheral motor component responsible
for execution of the tap. Variability in tapping is due to variability in both central and
peripheral components and limits timing precision.
Cerebellum is a key player in event timing
Patients with cerebellar lesions show increased variability on the rhythmic tapping
task but not on periodic tasks that are smooth and continuous. This is consistent with
the hypothesis that the cerebellum underlies timing of tasks involving an event
(marked by discontinuities) structure. This suggests the cerebellum as a key player in
the time keeping process of tasks such as the finger taping task (Ivry and Spencer,
2004).
21

Perturbations to finger tapping
Perturbations have been applied to the task by varying tapping intervals (Repp,
2001b; Repp and Keller, 2004), varying the contact time (Semjen and Summers,
2002), applying a torque to the finger (Kay et al., 1991) and by transient disturbances
using discrete movements of the contralateral finger (Yoshino et al., 2002). These
works showed a marked stability of movement parameters (Kay et al., 1991) and fast
error correction abilities (Repp, 2001b; Praamstra et al., 2003).
Inhibition of the primary and pre-motor areas prior to the synchronization task using
low frequency rTMS (see below) (Doumas et al., 2005), resulted in an effect on the
error correction component but not on sensorimotor synchronization, demonstrating a
dissociation between them.
Transcranial Magnetic Stimulation
Pulsed magnetic field induces electric field in the brain
Transcranial magnetic stimulation (TMS) is rapidly developing as a powerful, non-
invasive tool for studying the human brain. A pulsed magnetic field creates current
flow in the brain and can temporarily excite or inhibit specific areas. The magnetic
field is generated by driving a current through a coil placed on the scalp. According to
Faraday's law, the changing field B induces an electric field E in the brain tissue. This
field E will, by Ohm's law, drive an electric current in the tissue that causes suitably
oriented neurons to depolarize and thus generate action potentials. As the induced
electric field is of the order of 100mV/mm, the current density in the brain tissue
(conductivity is taken as 0.4 S/m) is 40µA/mm2, which is the same order of
magnitude as during normal neuronal activity (Bailey et al., 2001).
The effects of TMS on neuronal tissue
Knowledge of how different neurons and neural assemblies are activated by TMS is
very limited at the moment. In straight axons, the membrane is depolarized at
22

locations where the electric field along the axon changes, i.e. has a gradient. The
required electric field gradient is effectively also achieved in bent axons (Rotem and
Moses, 2006) and at nerve endings, which are probably the foremost sites of TMS
excitation (Ruohonen and Ilmoniemi, 1999). It is not known which type of cortical
cells, pyramidal (excitatory) or interneurons (with both excitatory and inhibitory
effects) are most sensitive to stimulation. Neither is it known how the stimulus
spreads in the vast neuronal networks in the brain, or how complex cell shapes and
ongoing background neuronal activity affect the outcome of stimulation (Bailey et al.,
2001).
The changes in the local excitability of the neuronal tissue seem to depend strongly on
the TMS parameters. Low frequency trains of TMS at or below 1 Hz reduces the local
excitability as reflected by cerebral metabolic rate or blood flow (Mottaghy et al.,
2000; Paus et al., 2001a). Repetitive TMS (rTMS) at 5-25Hz will usually enhance the
excitability (possibly via local disinhibition). This is true in general, although some
inter individual variability on the effects of different rTMS parameters exits. This
variability may be explained by different levels of background activity in different
brains. On the other hand, there seems to be a fair amount of intra individual stability
of the effects.
Spatial and temporal resolution
Spatial resolution of the TMS is limited. Figure eight coils are more focal than
circular coils because the field under the junction region of the coil is twice the
strength as that under the two loops. Relying on the fact that neurons usually have a
threshold for stimulation, we can usually assume that, over a given range of
intensities, stimulation is limited to sites under the junction region. However, sub
threshold effects undoubtedly occur under all parts of the coil. It is also not clear if
effects of rTMS are confined to the stimulated area. Stimulation in a certain area may
exert effects on distant cortical substrates via multisynaptic pathways. But, since the
stimulus has a peak value at a defined position(s) under the coil and then falls off with
distance, it is still possible to plot the "best point" for stimulation with some accuracy.
In the motor cortex, for example, it is possible to distinguish separate "best points" for
individual muscles, even if only 0.5 cm or so separates them (Jahanshahi and
23

Rothwell, 2000). Thus we can say that the spatial resolution can be as good as 0.5cm
for structures near the surface of the scalp. However, this resolution falls off
considerably for deep structures as in EEG and MEG.
The temporal resolution of the disruptive effect of TMS from a single shock is
roughly of the order of 50-100ms, but this is increased if trains of pulses are used. It is
possible that the effect of trains of stimuli can spread much further than a single
shock, thus reducing the spatial precision (Jahanshahi and Rothwell, 2000).
Orientation of the coil on the scalp can influence which elements of the cortex are
activated by a given intensity. This is because, for physical reasons, currents induced
by TMS can only flow parallel to the surface of the brain. There is no component of
radial flow (Tofts, 1990). Because neurons are excited best by longitudinal rather than
transverse currents, and cortical neurons are arranged in particular orientations with
respect to the surface of the brain, this means that TMS can preferentially activate
specific populations of cortical neurons (Amassian et al., 1992)
Controlling for sensory effects
Although TMS is often said to be a painless way of stimulating the brain through the
scalp, it is not devoid of sensation. A loud click caused by the discharge of the
stimulator through the coil and a definite tactile sensation on the scalp may potentially
interfere with performance, and are hard to mask. This can be overcome by
controlling for these sensory effects by stimulating in a control site or using a control
task. Sham stimulation is also used, where the effects of TMS are simulated by
rotating the magnetic coil relative to the scalp. This sham control method has
limitations since it has been shown that the TMS pulse can still affect the cortex
(Sawaki et al., 1999).
24

TMS in the study of brain dynamics
Extensive use of TMS
Magnetic stimulation was first achieved by Barker in 1985 and has been used
extensively since then (Cohen, 2000; Gangitano et al., 2001; Leff et al., 2001; Mull
and Seyal, 2001; Aleman et al., 2002; Bohotin et al., 2002; Burt et al., 2002;
Mottaghy et al., 2002; Sack et al., 2002). TMS of motor cortex can produce a muscle
twitch or block movement; TMS of occipital cortex can produce visual phosphenes or
scotomas. Low frequency TMS can produce a local virtual lesion in a particular
cortical areas for a limited time (see review by Jahanshahi & Rothwell, 2000).
Levels of excitation and inhibition in the motor cortex can be studied using single
pulse perturbations in a paired-pulse paradigm in which, depending on the interpulse
interval, suppression or facilitation of the muscle response can be elicited (Paus, 2005;
Tsuji and Rothwell, 2002; Pascual-Leone et al., 1998b). Changes in the onset of this
suppression or facilitation can be used to assess how excitation or inhibition induced
by TMS stimulation of different areas affects the motor cortex.
Studying cortical connectivity and dynamics
As mentioned above, repetitive transcranial magnetic stimulation coupled with an
imaging technique such as PET and EEG can be used to study cortico-cortical
connectivity and dynamics (reviewed by Paus 2005). In EEG (Electroencephalogram)
electrophysiological activity generated by different sources inside the brain is
measured at the scalp. Together with its high temporal resolution, this tool enables the
measurement of global brain activity and dynamics. However, coupling EEG with
TMS requires a compatible EEG system that can measure EEG signals immediately
following TMS stimulation. Consequently special EEG systems have been developed,
using special filters such as gain-control and sample-and-hold circuits, to protect the
EEG amplifiers from the fast-pulsed magnetic field (Ilmoniemi et al., 1997, Thut
2005). The challenge of these pre-amplifier devices is to make the signals return to
the normal DC level after the TMS pulse, with no residual from the large voltage
during the pulse.
25

Massimini et al., 2005a, showed that TMS activation of one cortical area during sleep
does not propagate to other areas as far as it does during wakefulness, thereby
demonstrating a breakdown of effective cortical connectivity during sleep. Jing and
Takigawa, 2000, applied short trains of high frequency (10Hz) rTMS to the left
frontal areas and found they can enhance coherence between frontal areas to parietal
areas for several minutes.
Strens et al., 2002 showed that low frequency, 1Hz rTMS of the motor cortex
increases ipsilateral cotrico-cortical and interhemispheric coherence in the EEG alpha
band (~10Hz). Paus et al, 2001b, found that single pulse TMS induced a brief period
of synchronized activity in the EEG beta range (15-30Hz) in the vicinity of the
stimulation site.
TMS can also be used to study the mechanisms for generation of evoked potentials
components, brain responses time-locked to some "event", (e.g. introduction or
omission of a sensory stimulus or a mental event). For example, Evers et al, 2001,
found that rTMS to the left but not to the right prefrontal cortex significantly
decreased reaction time and latencies of the P300 component, a positive peak in the
potential occurring about 300ms after the occurrence of a low probability task
relevant stimulus. Since P300 is considered a result of updating an internal memory
model, this result indicated the left prefrontal cortex to be involved in this function.
Klimesch et al., 2003, demonstrated the possibility to improve cognitive performance
by influencing brain dynamics. Using rTMS to influence the dynamics of alpha
desynchronization, they showed that they can enhance task performance even 30 s
after stimulation.
TMS in the study of Schizophrenia
The use of TMS in Schizophrenia research has mainly been in the area of measuring
excitability and inhibition of various cortical areas (Geller et al., 1997; Hoffman et al.,
1999; Daskalakis et al., 2001; d'Alfonso et al., 2002; Daskalakis et al., 2002;
Fitzgerald et al., 2002b; Fitzgerald et al., 2002a; Pascual-Leone et al., 2002b).
26

Hoffman et al., 1999, used TMS of left temporoparietal cortex in patients that
reported hallucinated “voices”, demonstrating its possible applications as a future
therapeutic tool. TMS coupled with an imaging tool such as EEG holds the promise to
probe the putative mal connectivity (Massimini et al., 2005b; Litvak et al., 2007)
underlying Schizophrenia deficits. The findings mentioned above about the ability to
change local and distal cortical activation and possibly influence cognitive
performance suggest promising applications in Schizophrenia, since increased or
decreased activity in some cortical areas has been associated with cognitive deficits in
patients with Schizophrenia (Haraldsson et al., 2004).
Main Research Goals
The primary research goal of this work was to use Transcranial Magnetic Stimulation
(TMS) to perturb oscillations invoked in a distributed cognitive network in the brain.
We hypothesized that perturbations to a neural network would illuminate aspects of
connectivity, stability and balance between its components. We compared the effect
of TMS perturbations between the healthy and the Schizophrenia brain which is
believed to be pathologically connected. We expected that comparing TMS
perturbations to the same network but in two different brain systems could reveal
differences in connectivity and stability that may provide important insight to both
human conditions.
The network we chose to study was the neural network that is activated when subjects
are asked to perform a periodic motion of tapping their finger to the beat of an
external auditory metronome, making an effort to keep in pace under TMS
perturbations. Even such a simple task induces the activation of a complex neural
network, involving mechanisms of motor execution, sensory input, sensory feedback,
volition, attention, timing, planning, error correction and awareness of action. TMS
perturbations were applied to this neural network, in synchronization with the
metronome, at the motor command output stage – the area in M1 activating the index
finger - which is easily identified in human subjects. Precise measurements of the
27

finger movement were supplemented by recording a subjective evaluation of the
accuracy.
Manipulating putative components of the network responsible for the task enabled a
further investigation of the connectivity patterns and the stability of the network in
both the healthy and the Schizophrenia brain. This was done by introducing a parallel
mental arithmetic task (to manipulate the attention load) and by inhibition of the left
dorsolateral prefrontal cortex (L-DLPFC), an area which has previously been
associated with high executive functions.
Main Results
TMS perturbations to the network underlying the finger tapping task, at the level of
M1, enabled separation of the timing process involved in controlling the tapping
movement from the motor process in charge of execution of the motor commands.
TMS perturbations profoundly affected the finger's trajectory and kinematics but not,
surprisingly, its timing accuracy. Specifically we observed two abnormal behaviors
that subjects were unaware of; a doubling of the frequency of the tap and a stalling of
the finger for half the period. We speculate that the TMS is causing a release of the
motor plan ahead of time into activation mode. The observed doubles and stalls are
then the result of an indirect interaction in the brain, making use of an existing motor
plan to correct the pre-activation and obtain the temporal goal of keeping the beat.
These results are outlined in Chapter 1.
In patients with Schizophrenia, but not in healthy subjects, these perturbations altered
the ability to correctly judge one's own timing accuracy and invoked high-level
deficiencies related to attention and volition in the form of lapses in movement. These
lapses, defined as little to no movement of the finger during a metronome period,
were observed when these subjects were tapping with the TMS perturbation. These
results imply that TMS can disrupt connectivity between distributed modules of both
high level and low level function in the Schizophrenia but not the healthy network,
supporting theories of disconnection in Schizophrenia (Friston, 1998; Andreasen et
28

al., 1999). Moreover, the lapsing phenomenon, that was much more specific to
Schizophrenia patients, indicate an abnormal link through which an excitation created
in a low level area such as M1 can impact on higher cognitive areas implicated in the
appearance of lapses. These results are further described in Chapter 2.
TMS perturbations to M1 coupled with manipulations to the attention component and
to the L-DLPFC affected timing, motor and high cognitive functions in both healthy
subjects and in subjects with Schizophrenia. In Schizophrenia patients only, lapses
were significantly aggravated by the attention shifting condition, but were
surprisingly almost completely eliminated following the inhibition of the L-DLPFC.
The fact that lesioning it improved performance of subjects with Schizophrenia,
suggests that the L-DLPFC is only a link in a network governing finger tapping,
which is mal-functional in Schizophrenia, and that removal of the L-DLPFC
component alleviates its dysfunction. See Chapter 3 for a further description of these
results.
29

Chapter 1: Transcranial Magnetic Stimulation in a Finger
Tapping Task Separates Motor from Timing Mechanisms
and Induces Frequency Doubling
Introduction
Temporal control of motion requires both a motor program and a timing mechanism,
and debate surrounds where it is located and how it operates (Ivry and Spencer, 2004).
Two forms of timing processes are thought to exist in the brain. One type of
mechanism for timing is a centralized “internal clock”, located e.g. in the cerebellum
or basal ganglia (Rao et al., 1997; Ivry and Spencer, 2004). The second type of timing
mechanism is a cooperative, emergent process that is distributed in the brain and
emerges as a result of a specific task (Ivry and Spencer, 2004; Mauk and Buonomano,
2004). Finger tapping is a simple paradigm for studying such event timing, involving
a repetitive motion that can be precisely monitored. It is considered a concatenation of
discrete movements that is punctuated by events such as surface contact, rather than a
continuous motion (Spencer et al., 2003c; Delignieres et al., 2004; Spencer et al.,
2005). The leading model for finger tapping (Wing and Kristofferson, 1973) proposes
a central timekeeper that provides intervals of the appropriate length, triggering motor
commands at the end of each interval. The model hypothesizes that the clock and
motor functions are distinct, and may involve totally different brain areas.
Prevalent thought is that the motor program in finger tapping involves a chain of
central processes that occur in a given order, resulting in the serial activation of
muscles. Neurophysiological studies in the cerebral cortex have shown that while this
chain model may be oversimplified, a serial order exists in the manner in which cells
in different regions fire during motor planning and execution (Shima et al., 1996;
Crammond and Kalaska, 2000).
External intervention during finger tapping was limited in the past to variations in
timing (Repp, 2001a, b; Praamstra et al., 2003), contact time (Semjen and Summers,
30

2002) and mechanical perturbation (Kay et al., 1991), and mainly served to
investigate the dynamical aspects of motion and of return to stability using error
correction. The advent of TMS has enabled a richer approach and resulted in a
number of insights on timing (Day et al., 1989; Pascual-Leone et al., 1992b; Pascual-
Leone et al., 1992a; Doumas et al., 2005; Verstynen et al., 2006), trajectory control
(Desmurget et al., 1999), stability of motor states (Meyer-Lindenberg et al., 2002),
and motor information processing (Berardelli et al., 1994; Gerloff et al., 1997). The
advantage of using TMS for intervention during finger tapping is its relatively
localized application in the brain, its temporal precision and its wide range of output
powers.
Here, we employ TMS to study the interplay between the primary motor cortex and
the timing machinery in the brain. The TMS is applied to the motor cortex in a
periodic fashion, in synchronization with the metronome governing the finger tapping.
We follow the trajectory of the finger, and investigate the influence of the external
intervention on both the timing of the motion and on the motor execution. We find
that TMS enables the separation of timing and motor functions, and intervenes in a
particularly interesting manner in the motor function.
Materials and Methods
Subjects
The experiment was approved by the Sha’ar Menashe Mental Health Center review
board. Nine healthy right handed subjects took part in this experiment (5 males, age
20-48 mean 29.3±8.7) after screening with the safety questionnaire (Keel et al., 2000)
and an EEG test. Two of the subjects are authors and the remaining seven were paid
for their participation.
Design
Figure 1 presents the experimental setup. Subjects sat comfortably on a chair with
their dominant hand resting on a pad so that the whole hand could rest comfortably as
31

the finger was tapping. A photodiode was fixed into the pad so that the finger allowed
light to enter it in proportion to its height above the pad (see Figure 1 and details
below). The subjects wore headphones and heard clicks with 5ms duration of a
metronome produced via the computer. All subjects were tested at 2.5Hz, and some
(N=7) were tested also at other frequencies, ranging between 2-4Hz. These rates were
chosen since a tapping rate of 2-3.5Hz is considered “natural” for most people
(Stevens, 1886; Collyer et al., 1994).
Subjects were asked to tap to the beat as accurately as they can and to continue doing
so even when perturbations to the finger are produced by the TMS. Subjects were
explicitly told that their finger should be in contact with the target surface when they
heard the metronome signal. They were also explicitly told to try to resist the
perturbations and stay on beat. Subjects were not allowed to watch their tapping
movement and instead were asked to fixate on a convenient point in front of them. At
the end of each trial subjects were asked to rate the perturbations they felt by giving a
score from '1' to '4'. A score of '1' meant they felt no perturbation at all, '2' that they
felt something but it had no interference with the task, '3' that the perturbation
interfered with the task but they overcame the perturbation and '4' that they felt that
the perturbation was so strong that they could not keep to the beat.
Every experimental session began with a pilot test prior to the experiment, to
determine seven TMS intensities that span the range of the '1'-'4' answers (see TMS
parameters). An experimental condition consisted of 21-35 trials, each comprising a
continuous series of ~50 taps. After several taps without TMS (no less than 16), the
next 16 taps were accompanied by TMS. One pulse was given per tap, synchronized
with the metronome beat and at a constant intensity. Subjects were asked at the end of
each trial to give their subjective rating ('1'-'4') and their answer was recorded. The
next trial was carried out ~20 seconds later. At least 3 trials were performed at each of
the seven chosen intensities and the order of the intensities was determined pseudo-
randomly to avoid guessing of the next step by the subjects. In each session a
minimum of two and a maximum of three frequency conditions were tested with a 10-
15 minute break in between. The first frequency used in each session was 2.5Hz. An
interval of at least one week was given between sessions. Since the 2.5Hz condition
32

was always performed together with at least another condition, this condition was
repeated for most subjects, and there were N=20 conditions of 2.5Hz.
In a control experiment, tapping without touching of the pad was executed with two
of the subjects. An additional supporting block placed on the pad elevated the hand,
so that its motion brought it close to the pad at its lowest point but did not touch it. In
this case the photodiode gave only an approximate measure of the trajectory, and we
added a video camera for precise measurement of the finger’s motion. Analysis was
then conducted off-line from a videotape of the motion.
Measurement of finger motion
We designed an efficient yet simple system for determining the finger position with
good accuracy. A photodiode was positioned in the pad on which the finger was
tapping, so that it was viewing a small hole in the pad that was covered by a thin tape.
An incandescent light was placed about half a meter above the photodiode, at a slight
angle to it. When the finger was resting on the hole it blocked the light from reaching
the photodiode and a minimal current was obtained from it. As the finger detached
from the pad and moved higher more light entered the hole, and the photodiode
current increased. Calibration tests (see inset to Figure 1) showed that finger height
was linearly proportional to the photodiode output. Estimate of the degree of accuracy
in timing is 5ms, and height accuracy is 1mm. During the trial the photodiode output
was visualized on a dedicated Labview program controlling the experiment,
effectively giving a visualization of the actual trajectory of the finger.
TMS parameters
Magnetic stimulation was delivered using a Magstim Rapid (Magstim Company Ltd.,
Wales, UK) magnetic stimulator with a 7cm figure-of-eight coil. Motor evoked
potentials (MEP) were recorded from the right first dorsal interosseous (FDI) muscle
using disposable surface electrodes. To determine the optimal site for activation of the
index finger we measured the resting Motor Threshold (rMT), which is defined as the
minimal TMS energy needed to elicit 5 of 10 50µV MEP responses (Pascual-Leone et
al., 1998a). The place of stimulation of the index finger was marked on a swimming
cap that the subjects wore throughout the experiment. Since the resting Motor
33

Threshold (when the finger is completely at rest and not tense) and the working Motor
Threshold (when the finger is moving and the muscles are tense) are different, we
performed a short pilot test in the beginning of each experiment. In this pilot test
subjects were asked to tap to a metronome at a rate of 2.5Hz and a train of 16 single
TMS pulses (each of 200µs duration) time locked to the metronome click was
applied. The intensities were distributed according to a random list around the
predetermined resting MT. The effect of the TMS on the finger was determined by
inspection of the movement displayed online via the dedicated LabView control
program and by the subjective rating '1'-'4' given by the subjects (see above). Based
on this, a set of seven TMS intensities was chosen for each subject, spanning the
range of no visualized or felt (subjective rating '1') perturbation all the way to a clear
visualized and subjective rated '4' perturbation. These intensities varied across
subjects and ranged from as low as 50% rMT to as much as 160% rMT. Intensities
never exceeded the range of comfort for the subject (as reported by the subjects). The
pilot tests were discarded from the subsequent analysis.
After each experimental condition the subjects were given a 10-15 minute break. The
next condition was preceded by a measurement of rMT to determine that no change
occurred in the tissue excitability (Pascual-Leone et al., 1998a) during or after the
experiment and to check that the position that stimulated the index finger did not
change during the break. rMT that varied less than 2% of maximal TMS intensity
above or below the initial rMT was considered unchanged. Similarly the settings were
considered unchanged if the location of activation for the index finger on the cap
moved to a new spot inside a radius of 1cm around the initial spot. In most cases no
change occurred at all.
In order to exclude an auditory effect, sham trials were performed in 4 subjects (one
subject twice) at the end of the last trial of the last condition tested in the session. 4-5
trains of TMS (~16 pulses, intermittent with at least 16 taps without TMS) at the
highest intensity were applied by placing a second, disconnected, coil in the normal
configuration with the connected coil positioned 4cm directly above. In all cases
subjects reported feeling no perturbation to the finger.
34

All TMS parameters were in accordance with the recommended safety guidelines
(Wassermann, 1998).
Recording and Analysis
A National Instruments A/D card (National Instruments Corporation, Austin, TX)
together with National Instruments LabView software were used to digitize and
record the light information arriving from the photodiode. The same program was also
used to create the metronome clicks, send the triggering signals to the TMS and
record the actual TMS stimulation. Data was analyzed using Matlab software (The
MathWorks, Inc, Natick, MA). The points delineating the beginning and end of the
periods where the finger was maximally flexed (“down” and touching the pad) and
maximally extended (“up” position) were visually identified using a special Matlab
program and marked by the experimenters. The tap cycle was divided into 4 parts: 1)
finger at maximum flexion, 2) finger extending and on the way up, 3) finger at
maximum extension and 4) finger flexing on the way down. Each tap was manually
tagged 1-4, depending on where the metronome occurred. Manual analysis was
performed to avoid automatic analysis mistakes that arise due to variability in tapping
between and within subjects.
Taps were considered accurate if the finger was in phase 1 when the metronome
struck. This criterion, although not the standard one for finger tapping tasks, fits best
the requirement given to the subjects during the experiment. Subjects that were "off
beat" more than 70% of the time were excluded from subsequent analysis. We defined
"off beat" when the finger was in positions 2-4. This restriction excluded 2 subjects,
one which was usually at the maximally extended phase (position 3) when the
metronome occurred, and the other usually on the way down. About 3% of the total
taps from accepted subjects were excluded from analysis because they could not be
unambiguously categorized.
A complementary analysis was performed in parallel, using the standard deviation of
the inter-tap interval, which is a more conventional measure of accuracy (Wing and
Kristofferson, 1973). The beginning of the tap was defined as the first contact of the
finger with the pad and the inter-tap interval was defined as the interval between the
35

beginnings of two taps. The synchronization error was defined as the time between
the beginning of each tap and the closest metronome pulse.
The existence of an additional minimum (“dip”) in the finger trajectory in between the
taps that occurred at or near the metronome beat indicated the existence of a “double”.
Both peaks flanking the dip had to be at least 50% of the average height of the regular
peaks in the vicinity of the dip, and its time duration had to be at least 35ms. The
slopes of the dip had to be comparable to the slope of the peaks in its vicinity. The
duration of each half of the double was counted from the two metronome beats to the
minimum of the dip.
If during TMS application the time spent in a tap at the down position was longer than
a given criterion, then that tap was defined as a “stall”. The mean time of the down
phase was first calculated for the no-TMS condition, and to qualify as a stall the time
spent down had to be both more than 150% of this mean and more than two standard
deviations longer than it.
Statistical Analysis
The tests that were used to analyze the data in this study were two-sample one-tailed
t-test for independent samples, paired one-tailed t-test and randomized block design.
Spearman Correlation, which does not assume normal distributions, was used for all
correlation analysis.
Results
TMS was used as a means of perturbing the motor action of the finger while it was
tapping to the beat of a metronome. TMS pulses were given together with the
metronome clicks, in a train that included 16 pulses. Tapping without TMS
perturbations served as a baseline.
Figure 2 shows the height of the finger as a function of time during the experiments at
2.5Hz. Examples of the finger trajectory are displayed. Figure 2a demonstrates the
36

baseline tapping motion of the finger when no TMS was applied and the period of
tapping was 400ms. Figures 2b and 2c demonstrate the two major scenarios for effects
of TMS on finger motion. In the first case the TMS caused the finger to stall at its
lowest position for half a period and only then detach and perform the tap in the
remaining half of the period (we call this kind of movement a “stall”). The mean
duration across all subjects (defined as the time between the first point and last point
of maximum flexion) of the time spent in contact with the pad during the regular,
unperturbed tap was (106±9)ms, while the mean duration across all subjects of time
spent on the pad during a stall was (199±10)ms, very close to 200ms (no statistically
significant difference was found from 200ms, two-tailed t-test P=0.56), which is half
of the tapping period.
In the second case the TMS caused the finger to perform an extra tapping motion,
though not necessarily completing the motion downwards (we call this kind of
movement a “double”). The frequency of the tapping was effectively doubled in this
motion, with each tap extending one half of the metronome period. The mean
durations across all subjects of the first and of the second taps were (212±7)ms and
(186±7)ms respectively, and they were not significantly different from 200ms (two
tailed t-test P=0.16). Figure 2d shows that the doubling motion is unaltered when the
TMS is delivered when the finger is still in motion. The arrows indicate off beat
finger positions (also in Fig. 2c), which were usually on the way down at the time of
the TMS strike (and metronome click). We see no difference in the motion
completing the tap, and subsequent doubles or stalls appear the same as when TMS is
given at the down position (on beat taps).
Overall, the stalls and doubles occurred in 11.2% and 17.3%, respectively, of the taps
where TMS was applied. Table 1 displays the mean occurrence of stalls and doubles
as a function of intensities, grouped into the low, medium and high range. Occurrence
of stalls increased with intensity from 2% to 22% for the lowest and highest
intensities respectively (see Table 1). Occurrence of doubles also increased with
intensity, from ~5% at the low intensities up to ~39% for the highest intensities (Table
1).
37

In both cases, subjects were not aware of the precise nature of the abnormal
movement of their finger. During doubles they reported feeling that the finger was
“lighter” and “flying by itself," and during stalls they felt that the finger was “heavy”
and “hard to move” at certain times. Defining a tap to be accurate if the finger is down
at the time of the metronome sound, we found that the subjective awareness of
accuracy '1'-'4' reported by the subjects after every trial (see Materials and Methods)
correlated well with their actual accuracy on each trial (see Chapter 2). The accuracy
was measured relative to the train of taps without TMS preceding the TMS taps in
each trial.
The structure in both doubles and stalls is interesting because both motions apparently
divide the 400ms time intervals into two parts of equal size. The second half of each
effect consists of a tap which the subjects attempt to complete in 200ms, albeit in the
case of doubles they begin at a higher elevation. Effectively this means when
performing doubles and stalls, the subjects are tapping at 5Hz. One can reasonably
expect the increased difficulty of this task to impair accuracy, but surprisingly this
was not the case. Figure 3 displays the percentage of accurate taps for both the
unperturbed baseline and for the TMS perturbations (two left columns). TMS does not
reduce the accuracy of tapping and on the contrary a slight increase is observed (see
also Doumas et al., 2005), although the difference is not statistically significant (one
tailed t-test, P=0.15). Interestingly, tapping accuracy remained high also when the
finger performed stalls and doubles (see Figure 4).
Although the average tapping accuracy, calculated for all subjects, was high (around
70% of the taps), still there was intra-trial variability in tapping accuracy and off beat
taps also occurred (as displayed for example in figure 2d). Figure 4 displays the
probability for ‘inaccurate taps’, i.e. cases where the metronome click occurred in
other parts of the tapping cycle other then the completely down phase. These
inaccurate taps occurred for taps without and with TMS.
An additional measure of variability in tapping was the variability in finger amplitude
as measured by the coefficient of variation (mean/std) in the maximum height of the
finger. In general finger amplitude variability was larger for taps with TMS, although
it was significant only for the highest TMS intensities (see also figure 11 in Chapter
2).
38

Using the standard measures of accuracy such as the variability in ITI and the
synchronization error gives a complementary view on these results. The ITI
variability increases with TMS intensity (see Figure 5), which is linked to the
variability in the motor execution and the strong fluctuations in the trajectory. On the
other hand, the synchronization error becomes slightly less negative (see Figure 6), so
that the tendency of the finger to touch the pad ahead of the metronome is smaller
with TMS. This is in line with a slight improvement in accuracy during TMS.
Since doubles and stalls result in an effective increase of tap rate to 5Hz, we analyzed
data from the frequency study for accuracy as well. We found that without TMS,
tapping accuracy did fall for all but one subject, when the metronome and tapping
frequency increased from 2.0Hz to 3.0Hz. The one anomalous subject was accurate
16.6% of the time at 2.0Hz, and 46.1% at 3.0Hz, and the average accuracy of the
remaining six subjects decreased from 72.2% at 2.0Hz, to 34% at 3.0Hz. We
therefore find it surprising that the tapping accuracy of 66%, for all subjects, at 2.5Hz
without TMS does not decrease when TMS is given because doubles and stalls
together account for 28.5% of all taps with TMS.
Information on the effect of TMS intensities can be obtained by looking at the
distribution of tapping accuracy according to intensity. The middle and right columns
in Figure 3 display the relative accuracy of tapping with and without TMS, but
separated according to TMS intensities. The right columns show the three trials with
high values of TMS intensities, and the middle ones show the four lower intensities.
While at the highest intensities there is no significant difference between the no-TMS
and TMS conditions (one tailed t-test, P=0.23), at low TMS intensities the subjects
were actually performing better on average (one tailed t-test, P=0.002).
Figure 7 demonstrates the possible irregularities in the trajectory of the finger during
TMS. Figure 7a displays the overall distribution of contact durations for no-TMS and
TMS taps for all subjects. Contact duration was defined as the time the finger was at
the lowest position, i.e. the interval from the time at which maximal flexion was
39

attained to the time at which the finger started moving upward. Shorter contact
durations correspond to doubles, while longer ones correspond to stalls. It is clear that
during TMS the distribution broadens and there are both doubles and stalls. The
distribution of stalls and doubles varied from subject to subject. Some subjects were
more “stallers” while other subjects were more “doublers”. Figure 7b displays the
distribution of stalls and doubles for the individual subjects. No correlation was found
between variables such as motor threshold, gender or age and being a “staller” or a
“doubler”. However, a correlation of -0.58 was found between the contact duration of
the finger with the pad during the no TMS condition and the probability of a subject
to be a “doubler”.
Although on average the subjects’ finger was down in time with the metronome,
inaccurate tapping caused the TMS, which was locked to the metronome, to be
applied also at times when the finger was not completely down. We divided the tap
cycle into 4 parts (see Materials and Methods). Figure 4 compares the probability of a
double or a stall to occur as a result of TMS perturbation at the four different stages.
When calculating the probability for a particular perturbation (stall or double) to occur
out of the total taps during TMS (see Table 2), a number of facts are observed. Stalls
had the highest probability to occur when TMS was applied to the finger when it was
completely up. On the way up, practically no stalls occur, and if an abnormality
occurs there then it is almost sure to result in a double.
It is important to note that no significant effect of the intensity of TMS on the duration
of stalls was observed. For the lowest four intensities it was (179±34)ms and for the
top three intensities it was (194±11)ms, though the difference between these durations
is insignificant (two-tailed t-test, P=0.64). This contrasts with the results of Day et al.
(1989) on the intensity dependence observed in delays of motion as a result of TMS.
The frequency doubling seen during TMS at 2.5Hz, also occurred at 2.0Hz and 3.0Hz.
At 2.0Hz six subjects were tested, and the mean time of the minimum in the double
was at (247±25)ms, which is not statistically significantly different from 250ms (two-
tailed t-test, P=0.9), one half the period at 2.0Hz. For the 3.0Hz condition three
subjects were tested, and the minimum occurred at (200±15)ms, and here too the
difference from one half the period (167ms) is insignificant (two-tailed t-test, P=0.16).
40

Four subjects were tested 3.5Hz, in which the minimum occurred at (130±6)ms,
insignificantly different from half the period (142.9ms).
We did find a clear effect of frequency on the occurrence of stalls and doubles. Figure
8a displays the ratio between stalls and doubles at different frequencies for all
subjects. A shift from doubles to stalls as frequency increases is observed in all
subjects but one. The change can be so dramatic that a log-linear plot is required.
Figure 8b displays the distribution of tap duration as a measure for stalls and doubles,
for all subjects, at the lowest (2Hz) and highest (3.5Hz) tapping rates. While accuracy
drops with frequency, it does so both for the no-TMS and the TMS conditions. There
was no significant difference for the relative change in accuracy (defined as the ratio
between accuracy at TMS and accuracy at no TMS) across frequencies (P=0.2 for
difference between 2Hz and 2.5Hz conditions, P=0.4 for difference between 2.5Hz
and 3-3.5Hz conditions). Although experiments always started with the 2.5Hz rate, an
order effect can be ruled out because the second and third frequency conditions were
randomly ordered and still the same frequency dependence was obtained across
subjects. We monitored motor excitability after each condition and checked that no
significant change occurred in the rMT or in the stimulation point (Pascual-Leone et
al., 1998a).
It was previously shown that asymmetry of flexion and extension exists in
synchronized tapping and that this asymmetry may assist movement timing,
especially in more difficult tapping conditions (Balasubramaniam et al., 2004). We
checked whether TMS perturbations affected the relative timing between the
extension and flexion phases by measuring flexion and extension average velocities
(measured as ratio between the amplitude of each phase and the time). Figure 9a
demonstrates the distribution of up velocities and down velocities for the no-TMS and
TMS conditions for all subjects. A clear increase in the velocities of the upward
moving finger during TMS is observed (Figure 9a, left panel) and is greater for the
higher intensities (Figure 9b). There is no effect on the flexion phase (downward
movement) (Figure 9a, right panel). This increase in the velocity of the upward
movement was also observed when we analyzed separately taps that were stalls and
taps that were doubles (data not shown). A randomized block design with subjects as
blocks and the extension and flexion phase velocities during no TMS or TMS as four
41

treatments revealed that the velocities at the extension and flexion phase were
significantly different from each other in both TMS and no TMS tapping (P<0.0001).
A possible explanation for the existence of stalls is suggested by the report of subjects
that their finger is “stuck” to the pad during stalls. To check whether the pad has a
role in causing stalls, we performed a preliminary test of tapping with no pad. The
data is less precise, and the absence of the sensation of touching the pad seems to
detract from the ability to tap precisely. However, it soon became evident that stalls
no longer occurred. In their place, we often observed an “inverse double” where the
finger performed an additional half tap at the lowest part of the trajectory.
TMS causes a loud click when activated, which could cause sensory facilitation and
could conceivably induce some of the effects we attribute to the magnetic field.
Nikouline et al., 1999, showed, for example, that the click elicits auditory-evoked
potentials. We therefore performed a control experiment under sham conditions,
where we put a coil that was not connected to the stimulator on the head of the subject
in the regular position and activated another coil that was placed just above the sham
coil. This coil was activated at the same times as in the regular conditions. In this way
the subject felt the pressure of the coil on the head and heard the click coming from a
very similar direction. None of the effects reported here were observed for the sham
condition. This control also serves to check for the possibility of an order confound.
Discussion
Our main observation is that TMS applied synchronously with the metronome
induced complex finger trajectories but did not affect the timing of the tapping events.
The tapping accuracy, as judged by the position of the finger at the time of the
metronome click, was slightly improved by TMS at low intensities. While this is
unexpected, it is in line with recent observations of Doumas et al., (2005) that motor
cortex inhibition decreased the tap-tone asynchrony (tapping inaccuracy).
The trajectory of the finger is profoundly affected by the TMS. At the high intensities
large deviations from normal finger tapping occur, while the finger still manages to
42

arrive at the pad, remarkably on time with the metronome. The deviations can be
categorized into two abnormal behaviors, stalls and doubles. A consistent additional
effect is the enhanced velocity of the finger on its way up.
Our measure of accuracy is different from the conventional, standard definition using
the inter-tap interval. The spatial resolution of the finger trajectory supplied by our
measurement apparatus enables the separation of contact times into the different
phases of the finger motion and we found that TMS does not reduce the probability
for the finger to be in the down phase with the metronome. The more standard
measure of synchronization error also showed a slight improvement in accuracy that
resulted in a slight shift of the reported negative error (Aschersleben and Prinz, 1995)
towards zero.
These two observations lead us to conclude that the timekeeping and motor functions
in the finger tapping task are controlled by distinct neural systems, and that the TMS
applied to the primary motor cortex affects only the motor function directly. There
does, however, seem to be an indirect interaction between the motor and timing
circuitry (see below). Abnormal motion in response to TMS occurred randomly
within a trial, indicating that the effect of TMS was limited to individual taps. This
supports the assumption that continuous tapping movement is composed of multiple
discrete events. This probabilistic effect of TMS on the movement trajectory is not
particular for our protocol and is probably due to fluctuations in the excitability levels
in the cortex.
The observed changes in trajectory cannot be attributed simply to muscular effects,
since deviations in the movement occur as much as 100ms after application of the
TMS. In comparison, the direct muscular response measured by EMG when TMS is
applied at rest occurs within about 50ms. The effect of the TMS was never apparent
before the ongoing motion – either flexion or extension – was completed. This is in
line with previous measurements (Day et al., 1989; Verstynen et al., 2006) and
strengthens our conclusion that we are affecting directly the motor plan rather than the
motor execution.
43

The dependence of the number of stalls on frequency may be related to the asymmetry
between the upward and downward phases of the trajectory. The probability for
getting a stall was highest if the TMS was delivered when the finger was fully
extended or on the way down, while it rarely occurred if the finger was on the way up.
Coupled with the decrease we measure in relative time spent on the way up as the
frequency increases, this may explain the change in relative occurrences of doubles
and stalls.
While stalls and doubles look very different, we attribute them to the same basic
mechanism. We understand the stall to be caused by the presence of the pad, and in its
absence would expect an exaggerated flexion, perhaps coupled to an inverse double.
Explaining the precise motion created by the TMS in conjunction with tapping, using
mathematical models and simulations, is beyond the scope of the current thesis and
poses an interesting direction for future research.
A recent paper (Verstynen et al., 2006) employed a complementary approach to
evaluate the relative contributions of the primary motor cortex in response timing and
execution. Their protocol involved an unpaced tapping paradigm, during which the
TMS pulses were applied at random times, independent of the subjects’ responses.
The measurement apparatus employed in their study was a telegraph-style response
key. They find a similar intensity dependant global increase in ITI variability,
regardless of its timing with respect to the TMS pulse. This is attributed to noise that
is added on to the implementation, and this is similar to our findings on the higher
variability in ITI under TMS, which we attribute to deviations in timing caused by
complex finger trajectories under TMS.
Verstynen et al., 2006, report a delay in the tap defining the end of the interval that
occurs when the TMS is applied during a critical time window of 100 ms just prior to
the flexion onset. This corresponds to TMS occurring during what we term phase 3 of
the trajectory (finger completely up). In this case stalls are most probable to occur. It
would be interesting to see whether in the full spatial trajectories of the Verstynen
experiment, stall like behavior occurs. Note that in our experiments the stalls do not
cause deviations of the mean ITI. This is probably because of our metronome - paced
tapping paradigm.
44

The paced versus unpaced paradigms also impact on long-term effects. In our case,
the next metronome beat and its associated TMS pulse reset the timing and cut off any
long term correlations and any changes in ITI caused by delays of the motion. This is
very different in the unpaced results reported by Verstynen et al., 2006.
Delay of execution of voluntary motion by TMS, reminiscent to the stalls we observe,
has been previously described (Day et al., 1989; Pascual-Leone et al., 1992b; Pascual-
Leone et al., 1992a; Berardelli et al., 1994; Haggard and Magno, 1999; Verstynen et
al., 2006). We view the stalls and observed delays as different. First, in the absence of
a pad we observed no obvious stalls. Second, the delay interval in previous
experiments is intensity dependent, while we observe constant stall intervals. Third,
the duration of the motion following the delay was unchanged in previous
experiments, while in ours it was speeded up. It is plausible that the absence of delays
in our experiment is related to the ongoing nature of the task, without stop-and-go
triggering.
Day et al., 1989, showed that it is possible to selectively delay the two independent
phases of the motion (agonist and antagonist). It is possible that in our case, the stalls
are a kind of mistiming between the two phases, which in the absence of a pad would
lead to different motions.
The main candidate in the brain for the timekeeping function during tapping is the
cerebellum (Ivry et al., 2002; Spencer et al., 2003c). A primary pathway from the
cerebellum to the motor cortex is known to exist (Strick et al., 1993; Middleton and
Strick, 1994). However, our attempts to target the cerebellum with TMS, following
the protocol of Theoret et al., 2001, and thereby to cause change in the timekeeping,
were not successful. While this could indicate that the timing circuitry is located
elsewhere, due to the depth of the cerebellum we are unable to exclude the possibility
that TMS was not effective enough (Jancke et al., 2004), and in practice a virtual
lesion was not created there.
Spencer et al., 2005, showed that lesions in the cerebellum lead to impaired timing
control of tapping. Interestingly, the appearance of doubles is also reported in that
45

paper, where both normal subjects and cerebellar patients occasionally executed an
additional tap within the time frame of a single tap. The probability of lesioned patient
to perform a double was greater by about a factor of three. These authors suggest that
the doubles may reflect a release of inhibition of planned movements. Doubles also
appear in the data of Meyer and Voss, 2000, for tapping with TMS, although their
existence is not addressed directly.
We therefore conclude that the appearance of doubles in our case indicates that we do
influence the timekeeper, albeit by an indirect interaction with the motor function.
The involvement of the timekeeper is strengthened by the precise halving of the
period that we observe in both stalls and doubles.
The observed inter-subject variation in the distribution of stalls and doubles may be
related to the different tapping patterns employed by the different subjects. Semjen
and Summers, 2002, found that varying the contact duration of movement affected
trajectories but not timing or accuracy. We similarly find a correlation between
duration of contact with the pad and the probability for a subject to be a “staller” or a
“doubler”. It may also be that the cognitive strategy adopted by the subjects to resist
the TMS induced perturbations (Bonnard et al., 2003) accounts for this inter-subject
variability, although it is probably not the only explanation. It is also possible that
different accentuation patterns (Sternad and Corcos, 2001) may affect the distribution
of stalls and doubles across subjects. This remains to be investigated in future
experiments.
A persistent result we obtained is the speeding up of the finger on its motion upward
for taps occurring during TMS, both in the presence of stalls and doubles and in their
absence. If the TMS causes delayed antagonist activation then the agonist motion
would be unhindered and hence naturally faster. Mechanical perturbation can also
lead to speeding up of the finger (Kay et al., 1991), but the major effect there was a
change in tapping phase.
The downward motion, on the other hand, is towards the temporal goal controlled by
the metronome beat (Balasubramaniam et al., 2004), and therefore under control of
the timekeeping mechanism. That explains why the downward motion is naturally
46

faster (Balasubramaniam et al., 2004). This can also explain why the downward
motion is not affected by the TMS.
It is possible that a generalized motor plan exists for the tapping motion where a
representation of the sequence of events in a tap and the relative timing between them
is kept (Vorberg and Wing, 1996). Upon transformation from intention to action an
explicit temporal goal is set by a timer (e.g the cerebellum (Spencer et al., 2003c)) and
the exact performance is then controlled by parameters that satisfy the various
temporal, mechanical and dynamical constraints of the motion (Hogan et al., 1987).
The TMS perturbation can interfere in this process by releasing the plan into
activation mode earlier than intended. This would require an adjustment of the
temporal parameters in order to carry out the planned sequence of extension-flexion
motions and obtain the initial goal, resulting in a multiple activation of this sequence,
in our case a double tap.
Since the representation of the motor plan needs to be continuous to account for the
repetitive nature of the motion, it may be that the order of sequence events (i.e. first
extension and then flexion) is flexible and can be inverted. In this case, when the plan
is released earlier than intended by the TMS perturbation, an inverted double tap can
also occur, accounting for the inverted doubling we presume is happening in case of
the stalls.
Subjects were generally unaware of the precise change in the trajectory, even when
performing an additional tapping step. This suggests that fast error corrections to the
motor function are automatic, occur outside of M1 (Doumas et al., 2005) and do not
enter into awareness. On the other hand, the subjects were well aware of the precision
with which they were tapping, i.e. of the timekeeper function. It follows that
timekeeping is a higher -level function then the motor program.
The possibility that TMS causes an external activation of the motor command prior to
the intended time is further supported by previous observations of an effect of a startle
on the speeding up of movement execution (Sanegre et al., 2004). When a startling
auditory stimulus is applied at the same time as the imperative signal in a simple
reaction time task experiment, movement execution is speeded up (Valls-Sole et al.,
47

1999). In this case it is hypothesized that the effect of a startle on reaction time is due
to external activation of the whole set of motor commands prepared for the intended
movement, either via subcortical pathways or by intersensory facilitation (Pascual-
Leone et al., 1992b; Pascual-Leone et al., 1992a).
An intriguing fact is that both abnormal trajectories occur in precisely one half of the
period set by the timekeeper, effectively doubling the frequency. It is interesting to
note in this context that the phenomenon of frequency doubling is known to occur in
certain nonlinear dynamical systems. The motor and timekeeping functions must
interact, since the timekeeper must adapt to correct for the changes set by the motor
dysfunction, and the resultant motion combines input from both these neural systems.
The appearance of higher harmonics, as in frequency doubling, may be an indication
of an interaction involving nonlinear terms between the oscillator governing the finger
tapping and the periodic forcing of the TMS.
The question remains: what is the precise form of the interaction that causes trajectory
deviations at exactly double the requested frequency and not other combinations.
Frequency doubling phenomenon can appear in oscillating systems when perturbed
mechanically by an external force. Thus we suggest that the explanation lies in the
nonlinear interaction of the muscular system involved in the oscillatory motion with
the external TMS perturbation. However, the exact formulation of this interaction is
beyond the scope of this work.
48


Figures and Tables
Figure 1. Finger measurement setup. Subjects sat comfortably on a chair with their dominant hand
resting on a foam pad. A photodiode was positioned inside the pad, and a thin semi-transparent tape
covered the aperture of the photodiode box. An incandescent light was placed about half a meter above
the photodiode, at a slight angle to it. When the finger was resting on the hole it blocked the light from
reaching the photodiode and a minimal current was obtained from it. As the finger detached from the
pad and moved higher more light entered the hole (left inset), and the photodiode current increased.
Right inset shows a calibration test for one subject. The finger height was linearly proportional to the
photodiode output.
49

Figure 2. Tapping Profiles. Finger height versus time for the three types of observed motion: a)
Normal tapping without TMS, b) the stall effect with TMS, in which the finger remains down much
longer, and c) the double motion with TMS, in which the finger taps at twice the required rate. d)
shows examples of doubles occurring when subjects were off beat (indicated by arrows). There is no
sharp deviation in motion as a result of TMS though the finger is moving, and subsequent taps are
similar to those in part c). The dashed vertical lines are separated by 400 ms, and indicate the times of
metronome signals (in a) as well as TMS pulses in b), c) and d).
50

Figure 3. Tapping Accuracy. Ratio of accurate taps to total taps performed without TMS (blue
columns) and with TMS pulses locked with the metronome signals (red columns), for all subjects. A
tap is considered accurate if the finger is down and in contact with the pad when the metronome
signals. The left most columns show the tapping accuracy for all intensities. Middle columns show
tapping accuracy for the lowest four intensities and the right most columns show the tapping accuracy
for the highest three intensities.
51

Figure 4. Probability of stalls and doubles to occur within the four phases of the tap cycle. If the
subjects’ finger was not down in time with the metronome, then their tapping was inaccurate and the
metronome click occurred in other parts of the tapping cycle. This caused the TMS, which was locked
to the metronome, also to be applied at times when the finger was not completely down. Shown is the
division of the taps, for all subjects, according to where the metronome and TMS (when TMS was
applied) occurred in the four different parts of the tap cycle (see Materials and Methods). Blue
columns: all the taps during no TMS, red columns: all the taps during TMS, yellow columns: taps
where doubles occurred and green columns: taps where stalls occurred.
52

Figure 5. Variability of the inter-tap interval. The variability of the inter-tap interval (ITI), measured
as the mean standard deviation of the ITI, as a function of the seven intensities for taps with TMS (red)
and taps without TMS (blue). Inset: the mean standard deviation of the ITI for TMS and no TMS taps
collapsed across all intensities.
53

Figure 6. Distribution of synchronization error. The distribution of the synchronization error,
defined as the time between the occurrence of the metronome and the first touch of the pad for taps
with no TMS (blue curve) and taps with TMS (red curve). Distribution curves were normalized with
the absolute of the mean error in the no TMS taps. A clear shift of the synchronization error
distribution peak from negative values towards zero is observed for the taps with TMS.
54

Figure 7. The possible irregularities of the finger trajectory. a, Average distribution of the time
the finger stayed down in contact with the pad, upon completion of a tap for all taps without TMS (blue
curve) and all taps with TMS (red curve). The longer times indicate the existence of stalls while the
shorter times the existence of doubles. Times are normalized by the average contact duration in the no-
TMS condition.
b, Frequency of stalls (red column) and doubles (blue column) for all subjects. While subjects do
execute both anomalous motions, they tend to favor one of the two motions. All the subjects were right
handed. No clear and statistically significant correlations were found between the favored motion
(double or stall) and physical characteristics. Subjects performed between N=2 and N=5 trials, except
NH for whom N=1.
55

Figure 8. Frequency dependence of stalls and doubles. a, The ratio of stalls to doubles for all the
subjects in the different frequency conditions they were tested for. The increase of stalls relative to
doubles was so dramatic in some cases that a log-linear axis was used. b, Transition from stalls to
doubles as measured by the average distribution of the contact duration (see figure 7a), for the two
extreme frequencies (2 Hz – right plot, 3.5 Hz – left plot). The blue line is the distribution of time the
finger stayed down for all the taps without TMS, the red line is the distribution of time for all taps with
TMS. The green line is the difference between the blue and the red lines. A clear shift from left to right
of the dip in the difference is observed between the 2Hz and the 3.5Hz conditions, indicating less
doubles and more stalls.
56

Figure 9. Effect of TMS on finger velocity. a, Distribution of the velocities during the up, extension
motion (left) and the down, flexion motion (right) for all the taps occurring without TMS (blue) and
with TMS (red). b, The mean normalized up velocity as a function of the seven TMS intensity levels
(see Materials and Methods). Although the actual TMS intensities varied between subjects, the seven
intensity levels are a measure of the reactivity of the cortex to the TMS stimulation and are distributed
around each individual's rMT. In blue is the baseline velocity level when no TMS was applied (and
therefore no dependence on intensity) and in red are the velocities when TMS was applied.
57

Table 1. Distribution of doubles and stalls across intensities. The first three columns display the
average occurrence of doubles and stalls (mean and standard error) for the first two, middle three and
top two intensities respectively. The last column displays the average occurrence of doubles and stalls
for all intensities collapsed together.
Table 2. Division of taps according to where the metronome occurred during the tap cycle. The
first and second rows display the distribution of taps according to where the I. metronome click (in the
case of no TMS application) or II. the metronome click together with a TMS pulse, occurred during
one of four phases of the tap cycle. The last three rows display the probability that in case II (during
TMS) the taps were stalls, doubles or regular taps (not stalls or doubles). In bold are the largest
probabilities for stalls and doubles.
58

Chapter 2: Transcranial Magnetic Stimulation at M1
Disrupts Cognitive Networks in Patients of Schizophrenia.
Introduction
Leading theories in Schizophrenia posit that different symptoms of the disease are
caused by both a breakdown of connectivity between distributed brain circuits that
underlie the fundamental cognitive and psychomotor functions, and the opposite
extreme of over-connectivity (Friston, 1998). In particular, the 'Cognitive Dysmetria'
model points to abnormal inter-regional connectivity within the cortico-cerebellar-
thalamo-cortical circuit due to cerebellar dysfunction (Andreasen et al., 1996; Ho et
al., 2004; Daskalakis et al., 2005). Dysfunction in this circuit is thought to disrupt
synchrony, coordination, and timing of mental processes (Andreasen et al., 1999).
It is particularly instructive to study such disruptions using the network underlying
externally paced finger tapping task, since it combines sensorimotor coordination and
synchrony with aspects of timing and time perception. Although a simple task, the
neural network invokes sensory input, motor control, event timing, error correction
and higher executive functions that interact in a timed, repetitive fashion. Surprisingly
few studies have checked the performance of Schizophrenia patients in this
synchronization task, and those that did, found that medicated Schizophrenia patients
were deficient in motor synchrony in this task (Manschreck et al., 1981; Manschreck
et al., 1982).
In this chapter we studied the network’s function by creating a periodic, precisely
controlled external perturbation to the internal periodic process. We used Transcranial
Magnetic Stimulation (TMS) to intervene in the oscillatory motion created by the
internal circuitry activated by the finger-tapping task. TMS pulses were synchronized
with a metronome and applied to the network at the level of the primary motor cortex,
while the subject tried to tap to the beat. We hypothesized that differences between
normal and Schizophrenia patients in neural network connectivity would manifest
59

themselves as differences in the effects of the TMS perturbations on activity governed
by the network.
We have shown in Chapter 1 that TMS pulses to M1 during paced finger tapping had
a great impact on the finger trajectory, without hindering the accuracy in timing in
healthy subjects. The results of that study revealed a dissociation of timing and motor
components for healthy subjects. Since timing deficits are known to exist in
Schizophrenia patients (Davalos et al., 2003; Elvevag et al., 2003) we expected in the
present work, to find some difference to appear already at the level of the
sensorimotor synchrony. Our first surprise is that TMS perturbations left the tapping
accuracy intact in Schizophrenia patients, even when profound trajectory deviations
occurred.
Our second observation is that the perturbations at the level of M1 were able to induce
an alteration in timing perception of Schizophrenia patients, and to trigger lapses in
the performance of tapping, i.e. higher-level deficiencies related to attention and
volition. We believe this provides evidence for the altered connectivity of the
Schizophrenia network as compared to the normal network. It furthermore suggests
the potential use of TMS perturbations as a means to probe connectivity between the
components of other distributed networks in the brain of Schizophrenia patients.
Materials and Methods
Design, measurement of finger motion, TMS parameters, recording and analysis were
as described in Chapter 1.
Subjects
The experiment was approved by the local Helsinki committee. Nine healthy subjects
(5 males and 4 females, age 20-48 mean 29.3±8.7, 9 right handed), including two of
the authors, and nineteen Schizophrenia patients (17 males, age 23-47 mean 39.5±5.6,
15 right handed, 2 left handed, and 2 females, ages 27 and 57, both right handed) took
part in this experiment, and were paid for their participation. After a complete
60

description of the study was given, written informed consent was obtained. Patients
were diagnosed according to the DSM-IV (First et al., 1995) criteria and rated for
symptom severity with the positive and negative symptom scale (PANSS) (Kay et al.,
1989). Schizophrenia patients with a history of neurological disorders, comorbidity
and drug abuse were excluded from the study. All Schizophrenia patients received
their regular medication during the time of the experiment, three of which were on
atypical antipsychotic medication in all mean dose equivalent of 412mg
chlorpromazine.
Atypical medication involved therapeutic doses of Ziprasadon, typical medications
included Haloperidol, Perphenazine and Fluphnazine. Only two patients who were
medicated with Perphenazine, needed continuous medications with Biperidon to avoid
developing Akatisia, and they did not have extrapyramidal symptoms while on
medication with Biperidon, thus allowing them to participate in the study. Two
patients received Carbamazepine treatment but with low doses for symptomatic relief
only. Patients had on average, a nine year illness duration and a mixture of positive
and negative symptoms prevailing, thus were diagnosed at times as residual
Schizophrenia and paranoid Schizophrenia depending on aggravated and remission
periods. Due to institutional withdrawal, none of the patients used alcohol or other
psychoactive drugs during the three months before the experiments. Of the 19
subjects with Schizophrenia 14 were smokers and 5 were not, while only one of the 9
healthy subjects smoked.
Some Schizophrenia patients found it hard to use the '1'-'4' scale to judge the affect of
TMS on movement. In this case the exact description of their subjective experience
and the experimenter's observation were recorded and translated into a scale of '1'-'4'.
We defined a tap to be "on beat" if the finger was down (maximal flexion) when the
metronome occurred, and excluded 2 healthy subjects that were not on beat more than
70% of the time. For most of the subjects the experiment was repeated more than once
(on different days), in total there were N=20 and N=31 repeats of this experiment for
the normal subjects and Schizophrenia patients respectively. About 3% and 4% of the
total taps, in the case of the normal subjects and Schizophrenia patients respectively,
61

were excluded from the analysis because they could not be unambiguously
categorized.
Our study concentrated on phenomena particular to Schizophrenia, and therefore
included more people with Schizophrenia than healthy controls. While some results
presented here apply to both groups, of particular interest were the momentary lapses
of attention observed in a majority of the patients tested, but in none of the controls.
For clarity and for greater statistics, we expanded our study to include more patients.
Statistical analysis
The tests that were used to analyze the data in this study were two-sample one-tailed
t-test for independent samples, and paired one-tailed t-test. When normality was
violated Wilcoxon signed rank was used instead of the paired t-test. When equality of
variance was violated the Satterthwaite t-test was used. Spearman Correlation, which
does not assume normal distributions, was used for all correlation analysis. The
analysis of the subjective rating was performed using a 1-way ANOVA with the
subjective response as the independent variable (factor) at 4 levels, and the change in
accuracy as the dependent variable.
Results
Figure 10 shows graphs of the finger height as a function of time during the
experiment. Figure 10a demonstrates the baseline tapping motion of the finger
without TMS for Schizophrenia patients. While the motion is similar in both groups,
the Schizophrenia patients spend ~26% more time at the maximally flexed phase than
the healthy subjects ('Down Time' in Table 3). Figures 10b and 10c demonstrate the
two major effects of TMS perturbations on finger movement in Schizophrenia
patients as was shown for healthy subjects in Chapter 1. A "double" is shown in figure
10b, and in 10c a "stall" (see also Chapter 1). In the case of a stall, TMS caused the
finger to stay at its lowest position for half or more of the period and then perform a
tap in the rest of the period. In a double, TMS caused the finger to perform an extra
62

tapping motion (though not necessarily completing the motion downwards),
effectively doubling the frequency of tapping.
A summary of results on tapping with and without TMS is given in Table 3.
Schizophrenia patients had 46% more stalls than normal subjects and 28% fewer
doubles than normal subjects (both not statistically significant). As was the case for
normal subjects (see Chapter 1), the distribution of doubles and stalls in the
Schizophrenia group varied from subject to subject and some were more "stallers" (11
of 19 subjects) while others more "doublers" (7 of 19 subjects), and one patient
executed stalls and doubles equally. Table 3 gives the duration of stalling before
resuming a tap ("Stall time"), and the duration of the first and second taps in a double
("Double time"). Note that for both groups, both stalls and doubles divide the
metronome period in half.
The major observation made in Chapter 1 was that TMS perturbations in normal
subjects did not result in a greater difficulty to stay synchronized with the metronome.
In fact, a slight increase was seen (Doumas et al., 2005) for the lowest TMS
intensities (see Chapter 1). A number of factors led us to expect that Schizophrenia
patients would be less accurate in tapping even without TMS (Manschreck et al.,
1981; Manschreck et al., 1985) and that TMS perturbations would affect their
accuracy (either increasing or reducing it) in a greater manner. Among these are the
hypothesis regarding timing deficits in Schizophrenia (Davalos et al., 2003; Elvevag
et al., 2003) together with reports of motor deficits (Vrtunski et al., 1989; Wolff and
O'Driscoll, 1999; Doumas et al., 2005), and side-effects of medication (Leucht et al.,
1999; Tandon and Fleischhacker, 2005).
However, when analyzing across all intensities we found that the probability for
controls and patients to be down with the metronome was the same when tapping with
TMS and without TMS and no statistically significant difference was found between
the two conditions (see Table 3, "Tapping accuracy"). While for normal subjects we
observed a significant improvement in accuracy at the lower intensities (see Chapter
1), no such increase was observed for the Schizophrenia patients (data not shown).
63

TMS had two further effects on the finger motion of people with Schizophrenia that
were more pronounced than in healthy subjects. Large deviations were seen in the
height the finger achieves in each tap. The distribution of these heights was more
irregular for the Schizophrenia patients than for the healthy subjects. This is shown in
Figure 11, which displays the coefficient of variation (CV) of this distribution as a
function of TMS intensity for healthy people and people with Schizophrenia. CV is
defined as the ratio of the standard deviation of the distribution to its mean, and is a
measure of amplitude variation. While the amplitude was more variable in patients
even during unperturbed tapping, the application of TMS caused a larger increase in
variability for people with Schizophrenia, and this increase was even larger at the
higher intensities (the increase was significant for intensity levels 2 and 4-7 P<0.03,
DF=48).
Figure 12 displays the average ratio of the up velocity (velocity during extension) to
the down velocity (velocity during flexion) when tapping with and without TMS, for
the healthy (blue bars) and Schizophrenia patients (red bars). This ratio is closer to
one for more symmetric tapping profiles. While the tapping profile of normal subjects
tends to be asymmetric, with a slow up movement and a faster movement toward the
target (Balasubramaniam et al., 2004), the tapping profile of the Schizophrenia
patients is more symmetric (pooled t-test, P=0.008, DF=49). Application of TMS
perturbations causes an increase in symmetry for both groups (for normal P=0.0008,
for Schizophrenia patients P<0.0001); however this increase is significantly more
pronounced for the Schizophrenia patients (pooled t-test, P=0.05, DF=49). As with
healthy subjects (see Chapter 1), the increase in up velocity grows with increasing
TMS intensity (Figure 12 inset), though at the highest intensities (where subjects
report a tired feeling – see below), there is a decrease.
Similar to healthy subjects (see Chapter 1), Schizophrenia patients were not aware of
the abnormal movement of their finger. They only reported a feeling of "losing
control" or of the finger "feeling tired". In order to measure the ability of both healthy
subjects and Schizophrenia patients to judge their accuracy we calculated the actual
change in accuracy between tapping without TMS and tapping with TMS for each
trial, and compared it to the subjective rating ‘1’-‘4’ (see Chapter 1, Materials and
Methods) given for this trial. As in the case of Figure 3 in Chapter 1, accuracy was
64

defined as the number of times the finger was on the pad when the metronome struck
divided by the total number of taps. Defining AT to be the accuracy with TMS, AnT
without TMS, the change in accuracy is given by 100.(AT-AnT)/AnT. This
normalization of the tapping accuracy during TMS taps by the tapping accuracy
during no TMS taps was assential in order to account for trials where tapping
accuracy was relatively low even without TMS perturbations (see Chapter 1 for
discussion of tapping accuracy variability). Table 4 shows what the mean change in
accuracy actually was for the different subjective ratings ‘1’–‘4’ (see Materials and
Methods in Chapter 1). The first two rows show this relation for all trials at all
intensities for the healthy and Schizophrenia subjects respectively. A 1-way ANOVA
analysis with the subjective rating as factor produced a significant effect for
subjective response only for healthy subjects (P=0.01, DF=3) and not for the
Schizophrenia patients (P=0.7, DF=3). We attribute the fact that healthy subjects rated
'2' when they were (on average) more accurate, to the slight increase in accuracy in
the range of the low TMS intensities (see Chapter 1 Figure 3).
As described in the Materials and Methods section in Chapter 1, the subjective ratings
‘2’-‘4’ related to cases where the TMS perturbations were felt by the subjects. We
thus correlated these three scores with the change in accuracy related to them (see
Table 4, first row) and found a high negative correlation of -0.99 for the healthy
subjects, indicating their high ability to judge the effect of the TMS perturbations on
their accuracy (producing the expected negative slope). In contrast, no correlation
(0.36) between accuracy and subjective rating ‘2’-‘4’ was found for the Schizophrenia
patients (Table 4, second row), suggesting a reduced ability in judging the affect of
TMS on their accuracy.
It is possible that reduced judgment of accuracy could be a general deficit in
Schizophrenia and not a consequence of the TMS perturbations. We therefore
checked the judgment of accuracy of the patients at the lowest intensity, where TMS
perturbations are considered negligible. In this case the distribution of ‘1’s in patient’s
reports was ~80% while the distribution of ‘2’s and ‘3’s was ~10-20%, and no ‘4’s
were reported. This indicated that indeed patients felt little or no disturbance from
TMS at the lowest intensity, and that this intensity can serve to control for the effect
of the Schizophrenia pathology itself on subjective judgment. The last row in Table 4
65

displays Schizophrenia patients' actual change in tapping accuracy correlated with
their subjective rating at the lowest intensity. When significant change in accuracy
occurred patients reported ‘1’, while when almost no change occurred they reported
‘2’ and ‘3’. Thus, with weak TMS perturbations, Schizophrenia patients had no
problem in judgment of accuracy. The implication is that the impaired ability to judge
tapping accuracy at the higher intensities (see second row in Table 4) is not inherent
to schizophrenia, but an effect of the TMS perturbation to M1.
In addition to the doubling and stalling (observed for all Schizophrenia patients), a
subset of patients showed an additional effect. Figure 10d displays the phenomenon of
lapses that occurred. These lapses, defined as little to no movement of the finger
during a metronome period, were observed when these subjects were tapping with the
TMS perturbation. In all, lapses were seen in 12 of the 19 patients, but most severely
in 7 patients, whom we term strongly lapsing. Lapses occurred with a probability of
9.5-29% in these 7 patients during TMS, while in all 7 normal controls we observed
lapses in just 3 subjects with probability of less than 2-3.6% (see Table 5). When
correlating the occurrence of lapses for the strongly lapsing with the PANSS (Positive
and Negative Symptoms Scale) of each subject we found a correlation of 0.55
(Spearman correlation, P=0.04) with the average score of positive symptoms while no
significant correlation with average score of negative symptoms or with the average
score of general symptoms. Spearman correlation analysis was also preformed with
each PANSS symptom separately. Significant correlation or nearly significant
correlation was found between the occurrence of lapses and Delusions (0.68,
P=0.007), Hallucinatory Behavior (0.58, P=0.03), Poor Rapport (0.47, P=0.09), and
Unusual Thought Content (0.6, P=0.02). Lapses were also found to be correlated with
the occurrence of stalls (0.63, P=0.04) and anti-correlated with the occurrence of
doubles (-0.56, P=0.07). Lapses were not correlated to TMS intensity and could occur
for intensities that are both beneath and above the individual motor threshold.
All our Schizophrenia patients were medicated, some taking typical antipsychotic
medication known to have motor side effects (Leucht et al., 1999; Tandon and
Fleischhacker, 2005). We found that atypical medication was correlated to the
occurrence of stalls (0.58, P=0.03), and no correlation between the type of drugs
(typical or atypical), with the occurrence of lapses or doubles. Moreover, our results
66

regarding the motor abnormalities and the timing effects were robust, whereas our
Schizophrenia patient population was heterogonous with respect to the types of
medication they were taking. Taken together with past results that no significant
difference was found between medicated and non medicated patients when subjects
were asked to tap as fast as they could (Flyckt et al., 1999), we believe that the major
effects of TMS perturbations in this study are not related to medication.
Although at least fourteen of the nineteen Schizophrenia patients tested in this
experiment were heavy smokers (more than a pack of cigarettes per day) we did not
find a correlation between being (or not being) a smoker and having (or not having)
doubles, stalls or lapses.
Discussion
In comparison to normal subjects, TMS perturbations caused greater responses and
variability in the finger movements of Schizophrenia patients, especially at the high
intensities, but they did not elicit a new pattern of behavior. The enhanced sensitivity
and severity of motor anomalies in Schizophrenia patients is not very surprising
(Vrtunski et al., 1989; Hoy et al., 2004) and is usually attributed to deficits in motor
cortical inhibition (Pascual-Leone et al., 2002a) and to dysfunction of the dopamine
system (Yang et al., 2003; Yang et al., 2004). It is noteworthy that no correlation with
this type of medication was found for most observed abnormalities. We therefore
believe that the differences in motor function between healthy subjects and subjects
with Schizophrenia can be attributed to such deficiencies, e.g. in the abundance of
dopamine receptors or to local deficits of level of inhibition and excitation of the
motor circuit. In contrast to our expectations, TMS perturbations did not affect the
timing component, and Schizophrenia patients were as good as normal subjects in
keeping pace with the metronome.
On the other hand, perturbations applied to M1 did affect the higher brain function
related to the judgment of one’s own timing, indicating that this higher level
component may be tampered with via its interaction with other components in the
network. Since this does not occur in normal subjects, we can conclude that the
67

connectivity of the network is in some way deficient and that the connections are
more sensitive to perturbations than in the healthy brain. Abnormalities in the
connections between components associated with subjective timing judgment and the
other components in the brain relate to previous reports of timing deficiencies
(Davalos et al., 2003; Elvevag et al., 2003) and support the "Cognitive Dysmetria"
model (Andreasen et al., 1999). They also relate to models suggesting impairment in
the mechanism underlying the awareness to an action and the ability to compare its
outcome with the intended internal motor plan (Frith et al., 2000; Alain et al., 2002).
We find the occurrence of lapses in patients during TMS perturbations of M1 to be
particularly fascinating. Lapses in cognition are known to occur in Schizophrenia
patients during speech, for example in an interview or during a discussion. A-priori
this would seem to be an unrelated, high level deficiency.
Recently, Weismann et al., 2006, described the neural correlates of momentary lapses
in attention resulting in increased reaction times to visual stimuli. They provided a
system-wide description of this phenomenon, showing how lapses in attention
correlated with reduced activation in right frontal areas implicated with control of
attention, reduced deactivation of a widespread ‘default-mode’ network, reduced
stimulus-triggered activity in sensory cortices and increased activity in frontal and
parietal brain regions involved in the recovery from these lapses (Weissman et al.,
2006). The lapses we observe may therefore be associated to the lapses in attention
described by Weismann et al., 2006, may originate in similar brain regions, and have
similar effects on brain activity, i.e reduction of processing resources of the
behaviorally related areas.
Interestingly, we found correlations between appearance of lapses and the existence of
symptoms that underlie deficits in high brain functions such as judgment of reality
(delusions, hallucinations) and thought content (unusual thought content). A possible
explanation for the appearance of lapsing in patients with Schizophrenia is the
existence of an inherent weakness or instability in the regions responsible for attention
(Lewis and Gonzalez-Burgos, 2006). Such a weakness would also explain the
phenomenon of “blocking” in patients, where they seem to lose attention momentarily
during an interview. Perturbations to M1, similar to internal disturbances in the
68

patient (hallucinations etc.) could perturb the centers of attention sufficiently to cause
a lapse. This explanation, however, does not address the fact that lapsing occur during
one or two precise metronome periods.
A potentially more interesting hypothesis is that intervention in one part of the
network causes action in another part, but only in patients of Schizophrenia. Using the
system-wide observation of (Friston, 1998; Peled, 1999a; Weissman et al., 2006), we
can frame an explanation for the appearance of lapses that relies on the dysconnection
hypothesis (Friston, 1998; Andreasen et al., 1999). We attribute the lapses to the
existence of mal-connectivity in the cortical circuitry that regulates the motor control,
timing, attention and volition involved in finger tapping. Components of this network
should include the prefrontal cortex (Callicott et al., 2003), the anterior cingulate
cortex (Paus, 2001a), the auditory cortex, the primary and pre-motor cortices (Lewis
et al., 2004), the cerebellum (Ivry et al., 2002), the basal ganglia and the sensorimotor
cortex (Praamstra et al., 2003). If we model this network as a block diagram with
nodes and links, we can view the occurrence of lapses as evidence of an anomalous
effective over-connectivity in the neural circuitry that is specific to people with
Schizophrenia. Lapsing in response to TMS indicates that a link exists, through which
an excitation created in a low level area such as M1 can impact on higher, cognitive
areas, perhaps the ACC or other areas implicated in the appearance of lapses
(Weissman et al., 2006). The connection may not be a direct link, and may be relayed
through a number of other nodes, but it is not made in the healthy brain. This effective
or idealized link may actually be strengthened in practice only by weakening of other,
inhibitory links.
While Schizophrenia patients exhibit similar effects of TMS as healthy controls, in
general they were more easily perturbed, and to greater extent. The lapses, however,
are a much more specific to Schizophrenia. We postulate that the motor cortex is
erroneously connected to a node in the network which is deficient in people with
Schizophrenia, possibly mediated by components regulating attention and working
memory (Weissman et al., 2006). This suggests that TMS perturbations can be used to
alter and possibly to modify connectivity in distributed components of a network
(Haraldsson et al., 2004). Ongoing work to determine the nature of these connections,
69

as well as the components involved and methods of correction, holds the promise to
demonstrate the utility of TMS in understanding and treating Schizophrenia.
70

Figures and Tables
Figure 10. Finger Tapping Profiles of Schizophrenia Patients (N=19). Measured finger trajectories
shown as height versus time. Green dashed lines indicate the time of the metronome click and for b),
c), and d) the time of the Transcranial Magnetic Stimulation (TMS) pulse as well. Shown in a) is
typical finger motion without the TMS pulses. b) shows the doubles wherein an extra flexion is
executed halfway between metronome beats. c) shows the stalls which include finger inactivity lasting
half the time interval of beats. In part d) there is a beat in which no finger motion is observed, preceded
by doubles and followed by normal taping. This effect is particular to Schizophrenia patients and not
seen in healthy subjects. We termed this effect 'Lapses' (see text).
71

Figure 11. Height of Finger Variation for Healthy Subjects (Controls, N=9) and Schizophrenia
Patients (N=19). Coefficient of variation (mean/std) in the maximum height of the finger, plotted
versus Transcranial Magnetic Stimulation (TMS) intensity. Red curves are data from Schizophrenia
patients and blue are from healthy subjects. Dashed curves show data from finger tapping without
TMS, while solid curves indicate finger tapping with TMS pulses. Schizophrenia patients evidently
displayed greater deviations in trajectory than healthy subjects. Without TMS these deviations were
roughly constant (dashed curves), but were greater with TMS (solid curves), and showed a clear
dependence on the magnitude of the applied magnetic field (TMS intensity). Furthermore, the deviation
in Schizophrenia patients follows the same dependence on intensity as in healthy subjects, with an
initial increase followed by a constant plateau, and ending with greater deviation at the highest
intensities.
72

Figure 12. Asymmetry of Finger Velocity for Healthy Subjects (Controls, N=9) and
Schizophrenia Patients (N=19). Top panel: Ratio of finger velocity going up to velocity going down,
for Schizophrenia patients and healthy subjects. The columns on the left show this ratio during tapping
without Transcranial Magnetic Stimulation (TMS), and a clear increase with TMS is seen in the
columns on the right. Both groups experience an increase of up velocity relative to down velocity,
though patients had a significantly greater increase with the application of TMS. Bottom Panel: shows
the dependence of this increase with TMS intensity; here the up velocity with TMS is normalized by
the up velocity without TMS, and the increase implies this is a direct result of TMS.
73

Table 3. Average percentages and times for finger tapping. Average percentage of stalls and
doubles out of all taps performed with TMS. 'Down time' refers to the duration the finger stayed on the
pad when tapping without TMS. 'Stall time' is the duration the finger stayed down during a stall.
'Double time 1' is the time from a metronome click to the nadir of the first flexion in a double, and
'double time 2' is the time from the nadir to the next metronome click. Tapping accuracy is the average
percentage of taps in which the metronome occurred while the finger was on the pad (see Materials and
Methods). Tapping accuracy is displayed for taps without TMS ('no TMS') and for taps with TMS
('TMS'). All values are presented as 'mean' ± 'standard deviation'.
74
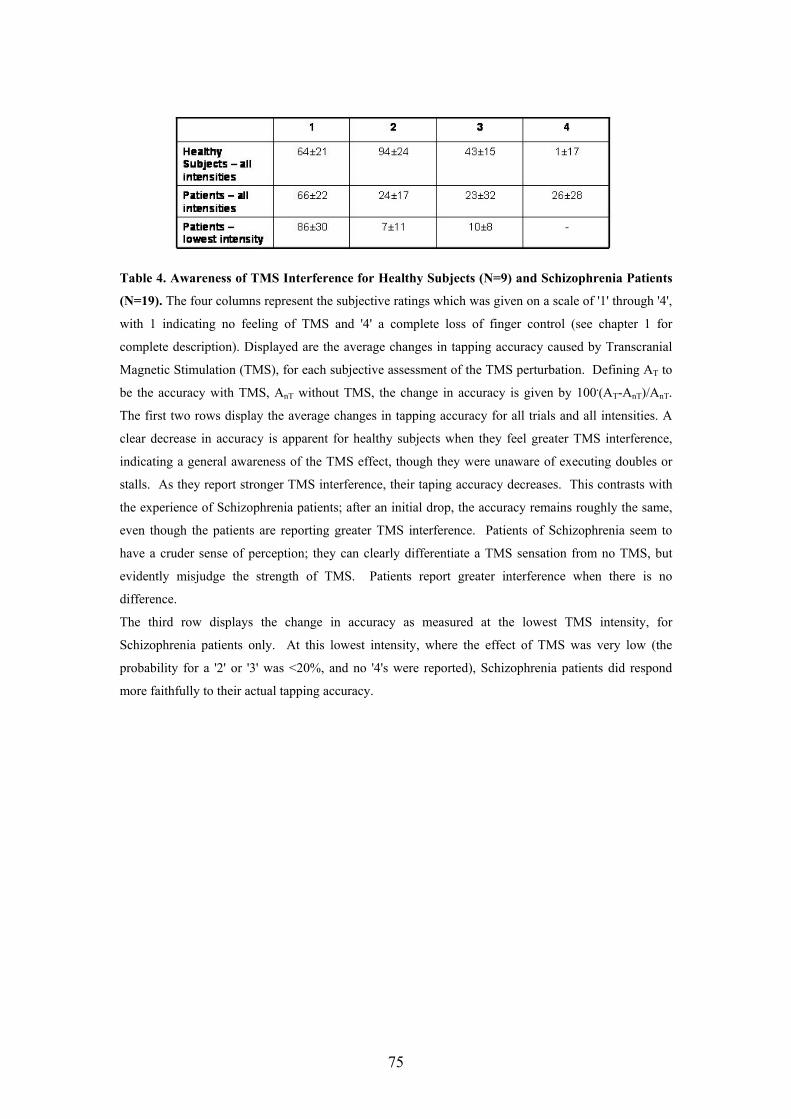
Table 4. Awareness of TMS Interference for Healthy Subjects (N=9) and Schizophrenia Patients
(N=19). The four columns represent the subjective ratings which was given on a scale of '1' through '4',
with 1 indicating no feeling of TMS and '4' a complete loss of finger control (see chapter 1 for
complete description). Displayed are the average changes in tapping accuracy caused by Transcranial
Magnetic Stimulation (TMS), for each subjective assessment of the TMS perturbation. Defining AT to
be the accuracy with TMS, AnT without TMS, the change in accuracy is given by 100.(AT-AnT)/AnT.
The first two rows display the average changes in tapping accuracy for all trials and all intensities. A
clear decrease in accuracy is apparent for healthy subjects when they feel greater TMS interference,
indicating a general awareness of the TMS effect, though they were unaware of executing doubles or
stalls. As they report stronger TMS interference, their taping accuracy decreases. This contrasts with
the experience of Schizophrenia patients; after an initial drop, the accuracy remains roughly the same,
even though the patients are reporting greater TMS interference. Patients of Schizophrenia seem to
have a cruder sense of perception; they can clearly differentiate a TMS sensation from no TMS, but
evidently misjudge the strength of TMS. Patients report greater interference when there is no
difference.
The third row displays the change in accuracy as measured at the lowest TMS intensity, for
Schizophrenia patients only. At this lowest intensity, where the effect of TMS was very low (the
probability for a '2' or '3' was <20%, and no '4's were reported), Schizophrenia patients did respond
more faithfully to their actual tapping accuracy.
75

Table 5. Rate of occurrence of lapses in Schizophrenia. The percentage of finger tapping trials in
which a subject with Schizophrenia experienced a lapse of attention. All subjects are listed; the four
left columns for people with Schizophrenia, and the rightmost two for healthy control subjects. The
last two healthy subjects (DK and IH) were excluded for poor tapping accuracy, but their lapsing
information is nonetheless given for completeness. The first seven subjects with Schizophrenia showed
the strongest propensity to commit a lapse, and are termed 'strongly lapsing' in the text.
76

Chapter 3: Probing Connectivity and Stability in the
Schizophrenia Brain using TMS Perturbations
Introduction
The interactions between various distributed neuronal modules that underlie cognitive
function and the dynamical nature of their connections has been of primary interest in
the past decade (Tononi et al., 1994; Varela et al., 2001b; Friston, 2002a; Meyer-
Lindenberg et al., 2002). Moreover, a pathological state such as Schizophrenia,
characterized by cognitive and emotional abnormalities has been hypothesized to be
the result of dysfunction in the connectivity of such distributed networks (Friston,
1998; Andreasen et al., 1999).
In this chapter we study the connectivity and stability of the Schizophrenia brain
using two techniques. In the first part we consider the externally paced finger tapping
task (discussed in the two previous chapters) and study the effect of TMS
perturbations to the network underlying it while manipulating its components. In the
second part we study EEG responses evoked by TMS as a means to study functional
connectivity in Schizophrenia.
The finger tapping task is considered a simple voluntary motor task, yet it activates a
distributed network of frontal, sensorimotor, cerebellar and deep brain areas that
underlie brain functions of volition, awareness of action, attention, timing, motor
planning, motor execution, sensorifeedback and error monitoring (Ivry and
Richardson, 2002; Praamstra et al., 2003; Taylor, 2003). In Chapter 1 and 2 we have
demonstrated that Transcranial Magnetic Stimulation (TMS) can be used to dissociate
components of such a network and study the interaction between them.
In chapter two we described our observations that TMS perturbations to the
Schizophrenia brain affected components of the finger tapping system that are usually
assumed to reside in high level brain areas, distant from the perturbed site (M1).
77

Perturbations to the motor component induced lapses in the movement of the finger of
subjects with Schizophrenia but only rarely in healthy controls (see Figure 14 for an
example of lapses). These lapses seemed related to the “blocking” observed
occasionally in patients during an interview and to the lapses in attention described by
Weissman et al., 2006. Patients who had many lapses also displayed strong positive
symptoms of the disease (see Chapter 2). Specifically, significant correlation was
found between the occurrence of lapses and Delusions, Hallucinatory Behavior and
Unusual Thought Content. TMS perturbations also greatly affected the ability of
subjects with Schizophrenia to judge their timing accuracy while in healthy subjects
this ability remained intact (see Chapter 2).
This ability to reach remote high level brain components via perturbations to a low
level motor component is in itself interesting and points at the potential of the
technique. However, the fact that it was observed in the Schizophrenia system but not
in the parallel healthy system suggests differences in connectivity and possibly
stability of these two networks and supports the disconnection theories of
Schizophrenia (Friston, 1998; Andreasen et al., 1999).
In principle, differences in connectivity of a neural network can be induced artificially
either by completely eliminating a component from the network or by changing its
relative contribution. In this paper we utilize both these approaches, and study
connectivity between other modules of the paced finger tapping network in both the
healthy brain and the brain with Schizophrenia. In the first case we manipulated the
putative contribution of the attention component to the network (Zanone et al., 2001;
Monno et al., 2002; Lau et al., 2004), by deviating attention to a cognitive task
introduced in parallel to the finger tapping task (Troyer et al., 1999) In the second
case we temporary eliminated the putative contribution of the left dorsolateral
prefrontal cortex to the network (Paus, 2001a; Callicott et al., 2003; Dalley et al.,
2004), by applying low-threshold, low frequency TMS stimulation to create a local
inhibition of this area prior to the finger tapping task (Pascual-Leone et al., 1998a).
In subjects with Schizophrenia only, we found that deviating attention from the finger
tapping task increased the occurrence of lapses while inhibition of the L-DLPFC
decreased finger abnormalities (trajectory and variability of motion) during TMS. We
78

interpret these results as supporting the notion of a mal connected network in the brain
with Schizophrenia. The L-DLPFC, known to be mal functional in Schizophrenia
(Lewis and Gonzalez-Burgos, 2006), seems to play a role in the creation of abnormal
connectivity and applying stress on one of the nodes of the network (by changing
attention load) seems to increase vulnerability to perturbations. Our results
demonstrate that manipulation of different components of a network can change
connectivity patterns of the network and its overall stability, and this can be probed
using TMS.
Another powerful tool to study functional connectivity is the use of EEG
(Electroencephalogram) together with TMS. EEG measures the superposition of
electrical signals generated by distributed multiple sources inside the brain, thus
providing a tool to measure global brain connectivity. In healthy controls TMS
stimulation at the vertex evoked a sequence of three clearly circumscribed peaks after
the stimulus. In patients no clear response pattern was observed. Some activation
peaks seen in controls were completely absent in patients while others were reduced
in amplitude and differed in their scalp potential distribution.
These findings demonstrate that there is a difference between healthy subjects and
patients with respect to the EEG responses evoked by TMS and suggest that these
responses can be used to study changes in brain connectivity and responsiveness in
Schizophrenia.
Materials and Methods
I. Finger Tapping Experiment
Basic design, measurement of finger motion and TMS parameters were as described
in Chapter 1.
79

Subjects
The experiment was approved by the local Helsinki committee. 8 healthy subjects (5
males and 3 females, age 23-48 mean 33.3±10.3, all right handed) and 11 subjects
with Schizophrenia (10 males, 1 female, age 28-47 mean 37.8±9.5, 10 right handed, 1
left handed) took part in this experiment, and were paid for their participation. After
complete description of the study to the subjects, written informed consent was
obtained. All subjects with Schizophrenia were on medication during the time of the
experiment, 3 were on atypical antipsychotic medication. All healthy subjects were
screened with the safety questionnaire (Keel et al., 2000) and an EEG testing prior to
their participation.
Design
Basic design was as described in Chapter 1. To manipulate the contribution of the left
dorsolateral prefrontal cortex and the contribution of attention load to the network
underlying the paced finger tapping task we ran two different experiments. In both
experiments subjects performed an externally paced finger tapping task (described
below). In the first experiment (COG condition) a secondary mental arithmetic task
was introduced in parallel to the tapping task. This task required adding a random
series of numbers, each of which were either '1' or '2'. The digits were displayed on
the center of a screen which was situated ~ 40cm in front of the subjects, at a visual
angle of 0.1 rad. Each digit was displayed for 600 ms and the next digit appeared after
1400 ms in the same place.
In the second experiment (DLPFC condition), low threshold 1Hz rTMS was applied
to the left dorsolateral prefrontal cortex to create a virtual lesion in this area (see TMS
Parameters) prior to the initiation of the finger tapping task. This caused a
deactivation of the L-DLPFC for the period of time long enough to perform the whole
finger tapping task. The metronome was activated immediately after the induction of
the virtual lesion in the L-DLPFC, and the finger tapping task begun.
The basic finger tapping task setup is described in detail in see Chapter 1. Some
subjects with Schizophrenia found it hard to use the '1'-'4' scale describing the
subjective judgment of the perturbations to the finger (see Chapter 1). In this case the
80

exact description of their subjective experience and the experimenter's observation
were recorded and translated later into a scale of '1'-'4'. When tested in the COG
condition, participants had to remember the calculated sum as well as there subjective
report and report both in the end of each trial. Three subjects with Schizophrenia (OS,
VP and AR) could not perform the tapping task while trying to add the randomly
appearing numbers. In their case they were asked to count the number of '2's that
occurred in the displayed set and report that at the end of each trial. Our general
impression was that reporting both the subjective report and the calculated sum was
too hard for most patients and thus the subjective rating was not used as reliable data
for subsequent analysis.
The basic finger tapping test (see Chapter 1) served also as a baseline control
condition ("baseline condition") and was performed in addition to the main condition
(COG or DLPFC) in both tests.
Every experimental session began with a pilot test, to determine seven TMS
intensities that span the range of the ‘1’-‘4’ answers (see Chapter 1). An experiment
comprised of one of the test conditions (COG or DLPFC) and a baseline condition.
Every condition within an experiment consisted of 21 trials, each comprising a
continuous series of ~50 taps. After several taps without TMS (no less than 16), the
next 16 taps were accompanied by TMS. One pulse was given per tap, synchronized
with the metronome signal that lasted 5 ms and at a constant intensity. The next trial
was carried out after subjects gave their report of their subjective experience '1'-'4'
(~20 seconds later). Three trials were performed at each of the seven chosen
intensities and the order of the intensities was determined pseudo-randomly to avoid
guessing of the next step by the subjects. The pilot tests were discarded from the
subsequent analysis.
In each experimental session the subjects were usually first tested with the baseline
condition and after a 10-15min break they were tested for one of the main (COG or
DLPFC) conditions. 7 healthy subjects and 7 subjects with Schizophrenia performed
the COG Experiment. 8 healthy subjects and 9 subjects with Schizophrenia performed
the DLPFC Experiment. Seven healthy subjects and 5 subjects with Schizophrenia
81

performed both experiments, in which case the experiments were run on different
days, at least 48 hours apart.
Creating a virtual lesion with TMS
To create a virtual lesion in the DLPFC we applied 1 Hz rTMS in a sub threshold
intensity (90% of the rMT) for 25 minutes. This was twice the time it takes for a
baseline experiment, and ensured that the virtual lesion effect remains during the
entire tapping test. The L-DLPFC site was determined by measuring 5 cm anterior to
the index finger site in M1. The coil was oriented in 45 degrees to the midline.
All TMS parameters were in accordance with the recommended safety guidelines
(Wassermann, 1998).
Recording and Analysis
Recording and analysis was as described in Chapter 1.
Based on our definition of being “on beat”, we excluded 2 healthy subjects, one male
and one female, one being constantly at the maximally extended phase when the
metronome occurred and the other usually on the way down. We did however include
these subjects in the analysis of lapses (see below).
About 6% of the total taps for healthy subjects and 27% for subjects with
Schizophrenia were excluded from the analysis because they could not be
unambiguously categorized. All taps however were used for the analysis of lapses as
this analysis was done manually.
Statistical analysis
The statistical tests we performed here were in most cases the two-sample t-test for
independent samples, and the paired t-test. When normality was violated Wilcoxon
signed rank was used instead of the paired t-test. When equality of variance was
violated the Satterthwaite t-test was used. Spearman Correlation, which does not
assume normal distributions, was used for all correlation analysis. The analysis of the
subjective rating was performed using a 1-way ANOVA with the subjective response
82

as the independent variable (factor, at 4 levels) and the probability as the dependent
variable
II. EEG Experiment.
Subjects 6 healthy subjects and 6 subjects with schizophrenia took part in this experiment.
Design A 64 channel ActiveTwo Biosemi EEG device (Biosemi Instrumentations,
Amsterdam, Netherlands) was used to record TMS-evoked potentials over the entire
scalp. Magnetic stimulation was delivered using a Magstim Rapid (Magstim
Company Ltd., Wales, UK) magnetic stimulator with a 7 cm figure-of-eight coil. The
precise timing of the TMS pulse was recorded in the specialized status (trigger input)
channel of the EEG.
Subjects sat comfortably in a chair. They were connected to the EEG system with 64
scalp electrodes (except for one healthy subject for whom a 30 channel setup was
used) positioned at extended 10-20 system locations. The TMS coil was placed over
the Cz electrode. For the healthy subjects, white noise, was generated by Matlab
software (The MathWorks, Inc), and was applied to the subject's ears to reduce the
effect of the TMS click. To reduce the electrical artifact, we used a stethoscope which
had its chest piece removed and replaced by an earphone that generated the white
noise directly to the subject's ears via the air filled tubes. The hospital environment
made it difficult to generate the white noise, thus for subjects with schizophrenia, ear
plugs were used instead.
The EEG signal was displayed by the Biosemi ActiView acquisition program during
the experiment. In experiments where subject’s alertness was monitored a module
added to the open source Actiview software was used for 8-12 Hz (alpha) band
filtering of the signal coming from a single channel (usually Pz) and determination of
its amplitude. When the amplitude signal was greater than a predetermined threshold
(individually determined for each subjects prior to the initiation of the experimental
setup), a trigger signal was sent to the magnetic stimulator activating the coil (given
83

that at last 3 seconds have passed since the previous TMS pulse). The EEG response
to this stimulation was immediately recorded with no saturation of the EEG
amplifiers.
For 5 healthy subjects and 5 patients two TMS conditions were tested. In the first
condition subjects were asked to relax and close their eyes, a state that elicited alpha
waves in their EEG. Between 30 and 50 single pulses of TMS at 70% of the maximal
stimulator output were applied over Cz whenever the amplitude threshold was
reached with an inter pulse interval of at least 3 sec. In the second condition the
subjects were asked to open their eyes so alpha waves were significantly reduced, and
TMS was applied manually by the experimenter (inter pulse interval of at least 3 sec).
Two sham conditions were additionally tested to control for the effect of the auditory
click. In both, the experimental setup was identical to the first condition described
above except for the placement of the coil. In the first case, the TMS coil was placed
2 cm in the air above the Cz electrode. In the second case the TMS coil was placed
~40 cm away from the subject. In some subjects an additional sham conditions where
the coil was placed 2cm above the Fz or Pz electrodes were used for analysis.
Data analysis The basic protocol for data analysis in the present study was previously described in
detail in Litvak et al. (in preparation). It is based on the multiple source approach
originally suggested by Berg and Scherg, 1994, for correction of eye artifacts. Brain
Electrical Source Analysis (BESA) software (MEGIS Software GmbH, Gräfelfing,
Germany) was used for data preprocessing and artifact correction. Each raw data file
was examined manually prior to the analysis. Channels heavily contaminated by noise
and epochs contaminated by artifacts other than the stimulation artifact were excluded
from the analysis. Epochs from -100 to +300 ms relative to the magnetic stimulus
were baseline corrected (baseline -100 to 0 ms).
In a previous study of Litvak et al. (in preparation) the data analyzed was recorded
using a specially adapted EEG system where the TMS artifact was blocked with a
sample-and-hold circuit of Virtanen et al., 1999. This led to complete loss of data
segment between 0 and 7 ms relative to the stimulus. The time period of several tens
84

of ms following the reconnection of the recording system at 7ms post stimulus was
contaminated with residual artifacts that could be corrected with the algorithm of Berg
and Scherg, 1994. BioSemi recording system used in the present study did not block
the recording at the time of the magnetic stimulus. Due to the wide dynamic range of
the system the stimulus artifact was completely recorded without saturation. However,
we found that the part of the artifact close to the stimulus onset had a complex scalp
pattern and could not be completely removed in software. Therefore, the time interval
between -2 and 10ms in each epoch was removed and replaced with spline
interpolation based on the time segments preceding and following the removed
interval. The interpolation did not remove very large artifacts found after 10ms post-
stimulus. These residual artifacts were corrected after computing the average of all
trials in a single raw data file (between 30 and 50 trials) using the algorithm of Berg
and Scherg, 1994, implemented in BESA. Artifact topographies were determined
based on principal component analysis (PCA) of a manually set time segment
contaminated by the artifact. The final result as in the study of Litvak et al. (in
preparation) was data clean of artifacts where the physiological responses could be
examined starting from about 10ms after the stimulus.
As described above conditions differed within and between subjects with respect to
the subject’s level of alertness (TMS pulses triggered while the alpha power was
increased vs. TMS pulses triggered with no relation to alpha power) and the kind of
measures used to block the coil click. In the case of sham stimulation there were
several kinds of coil positions as described above. In order to increase the signal to
noise ratio and enable comparison between subjects all the corrected averages of
responses to TMS at Cz under different conditions within a subject were averaged
weighted by the number of trials. The final number of trials used for averaging per
subject was 60±28 (min 29, max 107) for controls and 87±37 (min 32, max 129) for
patients.
Artifacts were also present in sham trials, though their amplitude and spatial extent
were usually reduced relative to real stimulation. Therefore sham data was analyzed
in exactly the same way as the stimulation data including artifact correction.
Following the artifact correction sham signals were averaged between different sham
conditions within subject weighted by the number of trials. The final number of sham
85

trials used for averaging per subject was 58±19 (min 27, max 84) for controls and
109±72 (min 35, max 201) for patients.
For comparison between subjects the missing channels were interpolated using spline
interpolation.
Results
I. Finger tapping experiment
We used TMS to perturb the voluntary motion of tapping in pace with a metronome,
while manipulating the contribution of different components of the network. Subjects
were asked to tap as accurately as they could to an external cue and try to resist the
TMS perturbations when they occurred. In one experiment we tested the effect of
manipulating the subjects' attentional resources by introducing an additional cognitive
task in parallel to the tapping task. In the second we tested the effect of inducing a
temporal inhibition of the left dorsolateral prefrontal cortex on subjects' performance.
In Chapters 1 and 2 we have reported that TMS perturbations applied while subjects
were performing the finger tapping task (similar to the baseline condition described
here) did not affect the timing accuracy but had a profound effect on the movement of
the finger. Specifically we observed two patterns of behavior of the finger. In the first,
the TMS caused the finger to stall for approximately half a period before performing
the tap in the remaining half of the period (see example in Figure 14). In the second
case the TMS caused the finger to perform an extra tap (see example in Figure 14),
effectively causing a doubling of the frequency (see Chapter 1).
The performance of healthy subjects and subjects with Schizophrenia in the cognitive
task is displayed in table 6. Mean performance was (84.8±8.0)% for healthy subjects
and (81.2±14.8)% for subjects with Schizophrenia (without OS, VP, AR who
performed a simpler task (see "Materials and Methods"), mean performance was
(71.9±11.2)% ).
86

Adding a cognitive task in parallel to the tapping task (COG condition) caused an
overall effect of decrease in timing accuracy in both healthy subjects and subjects
with Schizophrenia, when compared to the baseline condition (table 7a and 7b, left
column). In the case of healthy subjects the overall effect of decrease in accuracy was
statistically significant (P=0.02) while in subjects with Schizophrenia it was not
(P=0.18). The average absolute change in accuracy in healthy subjects and in subjects
with Schizophrenia, when tapping without and with TMS, is displayed in table 8.
Manipulating attention load changed the response of the finger to TMS perturbations
when compared with the baseline condition. The effect of adding a cognitive task on
the occurrence of doubles and stalls is summarized in table 9. A significant reduction
occurred only for doubles in healthy subjects and for stalls in patients. The overall
variability of movement can be measured using the coefficient of variation of the
distribution of the finger's height (see Chapter 2). In healthy subjects the COG
condition did not cause a significant change in variability while for subjects with
Schizophrenia a significant increase of 77% (P=0.02) in variability occurred in both
TMS and no TMS tapping.
In addition we found that the parallel cognitive task caused Schizophrenia subjects'
finger to spend more time down on the pad relative to the baseline condition when
tapping with TMS (from (246±10) ms to (339±42) ms (P=0.05)). No such effect was
observed in healthy subjects.
In healthy subjects we reported (see Chapter 1) that their awareness of what their
finger was doing was partial. They were generally unaware of the abnormal
trajectories (stalls and doubles) their finger was performing and only reported a
feeling of "losing control" or of the finger "feeling tired". In contrast, their perception
of timing accuracy remained intact under TMS perturbations (see Chapter 2).
Perception of timing in subjects with Schizophrenia, on the other hand, was greatly
affected by TMS (see Chapter 2).
Figure 15a displays the distributions of subjective ratings of accuracy '1'-'4' reported
by the healthy subjects (See Materials and Methods) for the baseline condition and for
the COG condition collapsed over all intensities. Healthy subjects (patients were not
analyzed for subjective rating, see Materials and Methods) rated more '1's and '2's
87

(feeling "nothing", or feeling "something" but not a perturbation to the movement)
and less '3's and '4's (feeling perturbations to the finger) in the COG condition relative
to the baseline condition. In a 1-way ANOVA with the subjective response as the
independent variable we found a main affect of subjective response for the COG
condition (P=0.009). Figure 15b displays the subjective ratings for the two highest
intensities, when TMS perturbations are assumed to be greatest. While in the baseline
condition subjects reported only feeling perturbed ('3's and '4's), with the parallel
cognitive task subjects reported that sometimes they did not feel any perturbation ('1's
and '2's) even in such high intensities. The inset demonstrates the relation between the
actual change in accuracy and the subjective rating for the two lowest intensities.
Assuming that at these low intensities TMS perturbations were close to zero, this inset
serves as control to isolate the effect of the cognitive task only on the ability of
subjects to accurately monitor their timing accuracy. The negative slope of the graph
indicates that the cognitive task, without TMS perturbations, did not affect subjects'
ability to correctly judge their timing accuracy. Thus, any effect on the subject’s
judgment, as seen in Figure 15a, was due to the TMS perturbations being applied
during the parallel cognitive task.
In Chapter 2 we reported the relatively high occurrence of lapses (defined as little to
no movement of the finger for at least one whole metronome period) in tapping of
some subjects with Schizophrenia during TMS perturbations (see examples of lapses
in Figure 14). These lapses hardly ever occurred in healthy subjects' tapping (see
Chapter 2). Adding a parallel cognitive task caused a significant increase in lapses for
subjects with Schizophrenia (see Table 10) (mean increase (8.6±3.5)%, P=0.035) and
even induced lapses in two subjects that previously did not have them (BM and SG).
No lapses were found in healthy subjects' tapping while they were performing the
parallel cognitive task.
In Chapter 2 we found a correlation between the occurrence of lapses and positive
symptoms such as delusions and hallucinations. Correlating the occurrence of lapses
in the COG condition with the PANSS scores of the individual subjects with
Schizophrenia who performed this condition, we found high correlations with Poor
impulse control (C=-0.87 P=0.01), Unusual Thought Content (C=0.83 P=0.02),
88

Hostility (C=0.7 P=0.08) and Stereotypical Thinking (C=0.8 P=0.04). No correlation
was found between the occurrence of lapses and medication.
Inhibition of the left dorsolateral prefrontal cortex (DLPFC condition) prior to the
finger tapping task caused an overall effect of decrease in accuracy in both healthy
subjects and subjects with Schizophrenia (see Table 7a and 7b, right columns),
although this effect was nearly statistically significant only for the subjects with
Schizophrenia (P=0.16 for healthy subjects and P=0.07 for subjects with
Schizophrenia). The change in absolute accuracy for both healthy subjects and
subjects with Schizophrenia, during tapping with TMS and without TMS, is displayed
in table 8a and 8b, right column.
L-DLPFC inhibition had an effect on the motor component of the network. The affect
of L-DLPFC inhibition on occurrence of stalls and doubles is summarized in table 9,
right column. For subjects with Schizophrenia DLPFC caused a significant decrease
in occurrence of doubles and stalls while for healthy subjects no significant change
was observed. Like in the COG condition, no change in variability (as defined by the
coefficient of variation of the distribution of the finger's maximal height) was
observed for healthy subjects. Interestingly however, for subjects with Schizophrenia,
while L-DLPFC inhibition increased variability for no TMS tapping (by 58%,
P=0.02), adding TMS reversed this effect to no change in variability similar to the
healthy subjects case.
Only two of the nine Schizophrenia subjects that preformed the DLPFC condition had
lapses in the baseline condition preformed at the same day and only one of them
exhibited a single lapse after L-DLPFC inhibition. Given all the lapsing data for all
Schizophrenia subjects (N=19) that preformed a baseline condition (see chapter 2),
the probability to have trials with lapses was 8.3% and 37% of the patients had above
10% trials with lapses. The probability to have trials with lapses taking all the
baseline conditions of the subjects that participated in the DLPFC condition (N=9)
was 3.8%. The fact that almost zero trials with lapses (0.5%) were observed in the
DLPFC condition suggests that L-DLPFC inhibition may have affected lapse
probability. No lapses were observed in healthy subjects.
89

We did not find an effect of L-DLPFC inhibition on the subjective perception of
healthy subjects.
II. EEG experiment
Figure 16 shows the TMS evoked potential waveforms before and after artifact
correction. The CPz electrode was chosen because it is close to the stimulation site
and picked up the physiological response to TMS, but was less affected by noise than
the Cz electrode directly touched by the coil. Even after removing the initial part of
the artifact by interpolation the residual artifact amplitude exceeded the physiological
signal amplitude by three orders of magnitude. Nevertheless, the residual artifact
could be corrected using the method of Berg and Scherg, 1994. After artifact
correction clear activity peaks could be seen in most subjects.
Figure 17 shows the grand averages of artifact corrected data for effective stimulation
and for sham. For both healthy controls (Fig 17A) and patients (Fig. 17B) a series of
clear peaks can be seen in response to effective stimulation that are absent in the
response to sham stimulation. However, the earliest peak clearly seen in healthy
controls in frontal channels around 29 ms after the stimulus is completely absent in
patients. The amplitude of other peaks in patients is also reduced.
These findings are summarized in Figure 18 which shows a butterfly plot of the grand
averages of responses to effective stimulation separately for healthy controls and
patients and scalp maps of potential distribution at the peaks of the responses seen in
healthy controls. It can be seen that the clearly circumscribed peaks seen in healthy
controls around 29, 43 and 59 ms after the stimulus are reduced in amplitude and
smeared in time in patients. In healthy controls at 29ms after the stimulus clear frontal
negativity and parietal positivity are observed. In patients the frontal negativity is
absent (as can be also seen in Fig. 17B) and the parietal positivity is greatly reduced
in amplitude. In addition, the negativity around 43ms after the stimulus is shifted
frontally and the centro-parietal positivity around 59 ms is greatly reduced in
amplitude.
90

Discussion
In this work we studied the effect of TMS perturbations on the stability and
connectivity in the Schizophrenia brain relative to the healthy brain. First, we studied
TMS perturbations to the network underlying the finger tapping task, under the
manipulation of the contribution of different components that was conducted in
parallel. We assumed that manipulating the contribution of different components will
have a general effect on the network and will cause a change in connectivity patterns
and possibly in stability. The left dorsolateral prefrontal cortex and the attention
component were both chosen because of their reported role in functions required in
the finger tapping task, such as timing (Mangels et al., 1998; Grondin and
Rammsayer, 2003; Dalley et al., 2004), performance (Troyer et al., 1999; Dalley et
al., 2004), voluntary action (Paus, 2001a; Monno et al., 2002; Lau et al., 2004) and
awareness (Stephan et al., 2002; Callicott et al., 2003; Taylor, 2003). Indeed, tapping
was affected in both the COG and DLPFC conditions even without the additional
TMS perturbations.
Attention has long been considered a central mediator that manipulates the amount of
processing activity allocated by the central nervous system to a certain task (Troyer et
al., 1999; Monno et al., 2002). Previous works have reported the effect of attention
load on coordination of motion (Zanone et al., 2001; Monno et al., 2002) and have
shown that intention and attention modify the stability of spontaneous coordination
dynamics between oscillating limbs. We therefore expected that changing the
attention load of the finger tapping task via a dual-task paradigm would affect
stability of the dynamical system associated with this task. We indeed found an
interaction at various levels of the network activated by the finger tapping task: the
timing accuracy, the subjective feeling and the motor execution.
First we found an effect of TMS perturbations on the timing component. Whereas
TMS perturbations to M1 without the manipulation of the attention load did not affect
timing accuracy and even caused a slight improvement (see Chapter 1 and 2),
introducing these manipulations caused a decrease in timing accuracy during TMS.
91

This is in line with previous reports of the connection between timing and attention
(Grondin and Rammsayer, 2003; Jantzen et al., 2004).
Second, manipulation of the attention component caused a disruption in the subjective
feeling of the TMS perturbation in the finger. Although healthy subjects' perception of
their accuracy remained intact, they reported feeling less perturbed by the TMS even
at high intensities of perturbation. These results support our previous observations for
the existence of connections linking low and high level functions (see Chapter 2).
Third, an effect of TMS on the motor component was found in the tapping of patients
with Schizophrenia, as observed by variability in amplitude and prolongation of
timing contact with the pad. While the latter is understood in terms of the general
deficiency in motor function in these patients (Vrtunski et al., 1989; Wolff and
O'Driscoll, 1999) which is exacerbated and amplified by the COG conditions, the
former may be due to a connection linking attention and movement error monitoring
processes.
As to the lapses described in Chapter 2, reducing attention in the COG condition
caused a significant increase in the lapses induced by TMS while no lapses occurred
in healthy subjects. This lends further support to our hypothesis that lapses are due to
higher cognitive functions related to attention mechanisms (see discussion Chapter 2)
(Weissman et al., 2006). The addition of an attention load as created by the COG
condition naturally reduces the ability to follow the beat and enhances the probability
of a lapse.
In Chapter 2 we suggested two alternative explanations to the occurrence of these
lapses. In the first, lapses were attributed to a weakness or instability in the attention
bearing areas, which was exacerbated by TMS. The second explanation implicated a
network of links between (among others) timing, motor, attention and volition. Lapses
occur due to mal-connectivity through which M1 can affect higher brain functions in
patients but not in healthy subjects. The aggravation of lapses during the attention
load is then understood as a result of putting additional stress on an already stressed
node in the network creating a change in balance that may make the network less
stable to perturbation. Patients of Schizophrenia are particularly liable to this stress
92

(Friston and Frith, 1995; Yang et al., 2004; Lewis and Gonzalez-Burgos, 2006) since
some of the attention nodes are known to be deficient.
In principle, the L-DLPFC condition should have provided an experimental setup to
examine the creation of attentional lapses and help to resolve between these two
explanations. The L-DLPFC was previously reported to be involved in high executive
functions such as volition and attention (MacDonald et al., 2000; Paus, 2001a).
Moreover, disturbed activation of the DLPFC appears to be specific to the disease
process in Schizophrenia (Lewis and Gonzalez-Burgos, 2006). We therefore expected
that inhibiting this area will profoundly affect volitional,attentional and other high
executive aspects of the finger tapping task (such as lapses) and decrease subjects’
ability to resist the TMS perturbations. Furthermore, we expected that subjects with
Schizophrenia would react more to this manipulation than healthy subjects.
Unfortunately, this issue could not be resolved in our experiments, since only two
subjects had a low number of lapses in the baseline condition prior to the inhibition of
the D-LPFC. The effect of D-LPFC inhibition caused the elimination of lapses in one
subjects while in the other one lapse was observed in all his trials. However, it is
important to note, that although an important area was inhibited, no lapses appeared in
all other subjects. This suggests that also in strong lapsing subjects, D-LPFC would
alleviate lapses. In this case, this will serve to support the explanation involving a
network. If weakening the L-DLPFC will avoid lapsing (i.e. improve attention) this
may indicate that a (perhaps indirect) link connecting motor to attention areas is no
longer active returning the network to more normal function. This remains to be
studied in the future and is beyond the scope of the present work. However, it
demonstrates this setup and similar setups as a means to use TMS for manipulation
and beneficial intervention (Haraldsson et al., 2004) in a network of the brain.
Intestingly, however, only in the patients with Schizophreina, did the DLPFC
condition cause a decrease in stalls and doubles and alleviat the effect on finger
variability during TMS, bringing it closer to healthy performance. If the mal
functional D-LPFC causes deficiencies in error correction mechanisms, possibly by
affecting links to other nodes in the network (e.g. the parietal cortex and the pre-
SMA, (Fiehler et al., 2004; Praamstra et al., 2003)), its inhibition may enhance error
93

correction abilities and reduce abnormalities in finger trajectories during TMS
perturbations.
In the EEG experiment we recorded fluctuations in electrical activity in response to
the presentation of a single TMS pulse. By measuring the time it takes for the
different peaks in activity to occur after presentation of the stimulus, it is possible to
determine the sequence and timing of the specific areas activated within the brain.
Significant differences were found in TMS evoked responses between the group of
healthy controls and the group of patients with schizophrenia. While in healthy
subjects TMS evoked a consistent response in all subjects, characterized by well
defined and timed peaks, in the Schizophrenia subjects the responses were smeared in
time and amplitude.
One possibility could be that these differences were due to different effects of the coil
click – in healthy subjects the click was masked by white noise while for patients it
was masked by ear plugs. However, the differences observed in the potentials were
consistent with previous findings from a similar study where recordings were
performed using a sophisticated artifact-resistant EEG system and a similar masking
method of the click for both groups (Massimini et al, 2005b), suggesting they reflect
neural processing rather then coil click effects.
In principle the networks of areas generating EGG responses evoked by TMS should
have similar sequence and time courses in all people, except when abnormalities and
pathologies in brain processing exist. It has been previously shown that changes in
long term connectivity result in changes in the form of the EEG potentials evoked by
TMS (Massimini et al., 2005a). Thus the fact that in the Schizophrenia brain
responses to TMS were not consistent serves as a further support to the existence of
deficits in processing and connectivity between areas. These preliminary findings
suggest that EEG responses to TMS can be used to study changes in brain
connectivity and responsiveness in Schizophrenia.
94
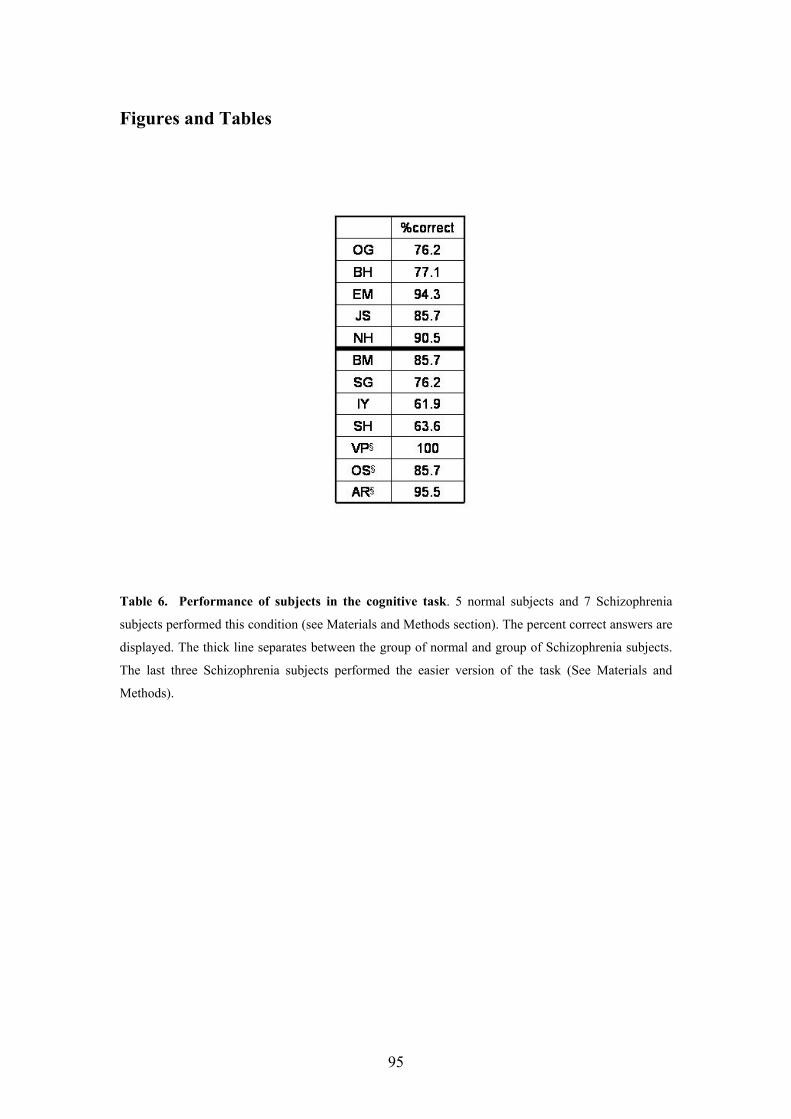
Figures and Tables
Table 6. Performance of subjects in the cognitive task. 5 normal subjects and 7 Schizophrenia
subjects performed this condition (see Materials and Methods section). The percent correct answers are
displayed. The thick line separates between the group of normal and group of Schizophrenia subjects.
The last three Schizophrenia subjects performed the easier version of the task (See Materials and
Methods).
95

a.
b.
Table 7. Affect of condition on tapping accuracy. Accuracy is defined as the percent of taps for
which the metronome occurred while the finger was down on the pad (tagged ‘1’ in the manual
analysis, see Materials and Methods section). The left column is the effect of the COG condition on
tapping accuracy and the right column is the effect of the LPFC condition on tapping accuracy. Cases
where the accuracy was reduced relative to the baseline condition were given a value of “one”,
otherwise they were given a “zero”. a. Normal subjects. b. Schizophrenia subjects.
96

Table 8. Change in tapping accuracy relative to the baseline condition. Accuracy is defined as the
percent of taps for which the metronome occurred while the finger was down on the pad (see Materials
and Methods section). The difference between tapping accuracy in the COG or LPFC condition minus
the tapping accuracy at the baseline condition is displayed (negative values mean that the baseline
condition accuracy was higher). The statistical significance of this difference is displayed as the P value
of a two-sided t-test. Asterisks signify cases where the difference was statistically significant (P<0.05).
97

Table 9. Change in occurrence of “doubles” and “stalls” relative to baseline condition. “Doubles”
are taps in which the finger performed an extra tap during the metronome period and “stalls” are taps in
which the finger stalled for approximately half the period before performing the tap (see Chapter 1 and
2). The occurrence, in percent (out of the total number of taps), of these variations in tapping for the
COG/LPFC minus the baseline condition are displayed (negative values mean the baseline occurrence
was higher). The statistical significance of this difference is displayed as the P value of a two-sided t-
test. Asterisks signify cases where the difference was statistically significant (P<0.05).
98

Table 10. . Rate of occurrence of lapses in Schizophrenia in the COG condition The percentage of
finger tapping trials in which a subject with Schizophrenia experienced a lapse of attention during the
condition in which attention was manipulated (right column) compared to the baseline condition
performed on the same day (left column). .
99

Figure 14. Example of lapses, “doubles” and “stalls” in the tapping. Displayed are three tapping
profiles of Schizophrenia patients. Highlighted in yellow are examples of lapses in the tapping. Lapses
are defined as little to no movement of the finger for at least one whole metronome period. Highlighted
in blue are examples of taps where the finger performed and extra tap. These taps were tagged as
“doubles”. Highlighted in pink is an example of a tap where the finger stalled for approximately half a
period before performing the tap in the remaining half of the period. Such tagged were tagged as
“stalls”. See Chapter 1 and 2 for a more detailed description of these categories of taps.
100

Figure 15. Distribution of subjective rating of accuracy. ‘1’-‘4’ are the subjective ratings of
accuracy as described in the Materials and Methods section. Blue is for the baseline condition and red
is for the COG condition. Top panel: The distribution of the subjective rating collapsed over all
intensities. Bottom panel: The distribution of the subjective rating for the two highest intensities only.
The inset is the distribution of the subjective ratings for the two lowest intensities which serve as a
control to isolate the effect of the cognitive task on the subjective rating (assuming at these low
intensities TMS perturbations were close to zero).
101

Figure 16. TMS evoked potential waveforms in the CPz electrode. Left panels are for controls
while right panels are for patients with Schizophrenia. Upper panels: Before artifact correction. Lower
panels: after artifact correction. Even after removing the initial part of the artifact by interpolation the
residual artifact amplitude exceeded the physiological signal amplitude by three orders of magnitude.
Nevertheless, the residual artifact could be corrected using the method of Berg and Scherg (1994).
After artifact correction clear activity peaks could be seen in most subjects.
102

A
B
A
B
Figure 17. The grand averages of artifact corrected data. Response to effective stimulation is in
black and to sham is in gray. For both healthy controls (upper panel) and patients (lower panel) a series
of clear peaks can be seen in response to effective stimulation that are absent in the response to sham
stimulation. However, the earliest peak clearly seen in healthy controls in frontal channels around 29
ms after the stimulus is completely absent in patients. The amplitude of other peaks in patients is also
reduced.
103

Figure 18 Grand averages of responses. Displayed are the grand averages of responses to effective
stimulation separately for healthy controls and patients and scalp maps of potential distribution at the
peaks of the responses seen in healthy controls. It can be seen that the clearly circumscribed peaks seen
in healthy controls around 29, 43 and 59 ms after the stimulus are reduced in amplitude and smeared in
time in patients. In healthy controls at 29ms after the stimulus clear frontal negativity and parietal
negativity are observed. In patients the frontal negativity is absent (as can be also seen in Fig. 17B) and
the parietal positivity is greatly reduced in amplitude. In addition, the negativity around 43ms after the
stimulus is shifted frontally and the centro-parietal positivity around 59 ms is greatly reduced in
amplitude.
104

Discussion – Stability and Balance in Networks
The experiments described in the previous chapters detail the effect of TMS
perturbations on the finger tapping network in the healthy and Schizophrenia brain. In
general, it seems that TMS perturbations affected this network at all its levels – at the
peripheral level controlling the finger muscles, at the ‘lower’ level components in
charge of motor and timing aspects, and at the ‘higher’ level components in charge of
volition, attention and monitoring aspects of the task.
In the first chapter we have demonstrated the ability to apply TMS to the network
underlying the finger tapping task in the healthy brain at one of its nodes (M1), and to
affect its components selectively. TMS pulses to M1 during paced finger tapping had
a great impact on the finger trajectory, without hindering the accuracy in timing in
healthy subjects. Thus, TMS enabled the separation of timing and motor functions and
demonstrated that in the healthy network even profound perturbations to one node do
not permeate to other nodes.
It is tempting to speculate whether the interesting phenomenon of effective frequency
doubling induced by TMS is related to the phenomenon of frequency doubling that is
known to occur in certain nonlinear dynamical systems (for example, when an
oscillating system is perturbed by an external force). Non linear interactions may arise
between the network of muscles and peripheral connections involved in the
oscillatory motion of the finger and the external TMS perturbations. Alternatively,
non linearity may arise from the complex interaction between the timing and motor
components in the attempt to keep with the goal of the finger tapping task despite the
TMS perturbations. The exact formulation of these interactions may provide insight to
their nature and present a natural extension and the next step beyond this work.
In the second chapter we compared between healthy and Schizophrenia subjects.
Unlike the healthy network that was quite resistant to perturbations, in Schizophrenia
patients we could affect higher cognitive functions related to attention and volition.
Perturbations at the level of M1 were able to induce an alteration in timing perception
of Schizophrenia patients, and to trigger lapses in the performance of tapping.
105

In the third chapter we have demonstrated how manipulations to different components
can change connectivity patterns of the network and its overall stability and how this
can be probed using TMS. Deviating attention from the finger tapping task affected
the subjective awareness of the healthy subjects to their finger during TMS
perturbations to M1. In patients with Schizophrenia it increased the occurrence of
lapses in Schizophrenia patients’ tapping. Inhibition of the L-DLPFC affected the
occurrence of finger abnormalities and improved its variability to the level of baseline
behavior as observed in the healthy subjects. Interestingly, in the subjects that
participated in the L-DLPFC inhibition condition, only one lapse was observed
although the probability for occurrence of trials with lapses was much higher (8.3%
for N=19 subjects). These results were interpreted as supporting the notion of a mal
connected network in the Schizophrenia brain, in which the L-DLPFC, known to be
mal functional in Schizophrenia (Lewis and Gonzalez-Burgos, 2006), plays a role in
the creation of abnormal connectivity.
These results demonstrate the use of TMS to study connectivity and stability of a
distributed cognitive network in the brain. In the healthy network, perturbations to M1
did not diffuse to other components. Moreover the existence of the effective
frequency doubling by doubles and stalls suggested that the timing component was
not only unperturbed, but was even able to compensate for perturbation-induced
deviations in the trajectory, probably via feedback connections.
The lapses that were found in Schizophrenia tapping when TMS perturbations were
applied, were seen to be correlated with Schizophrenia deficits that are related to high
cognitive functions and were suggested to be related to the lapses in attention
described by Weismann et al., 2006. The fact that perturbations to a low level
component (M1) induced them, led us to suggest that in the Schizophrenia network,
but not in the healthy network, there exists an effective link from lower to higher
areas which enables excitation in low levels to impact on higher levels. This link is an
‘effective link’ in the sense that it does not need to be a direct anatomical link but
rather the result of changes in connectivity in other links of the network, such as the
weakening of inhibitory links that normally prevent this direct influence of lower
levels on higher ones.
106

Manipulating attention load produced a change in the relative stability of the network
in the healthy brain, so that perturbations to M1 affected awareness of action, which
probably resides in higher brain areas. The fact that timing accuracy and occurrence
of doubles and stalls was reduced provides evidence for an effect of the perturbations
also on the timing component. This causal relation between a motor component, the
awareness of action component and the timing component provides evidence that
indeed areas activated by the finger tapping task constitute a network which is
connected via effective connections.
Unlike the healthy network, the Schizophrenia network seemed less resistant to
perturbations. TMS to M1 affected components underlying awareness of action and
increased the occurrence of lapses, supporting our hypothesis that these lapses are
related to high brain functions. Additional manipulation of the attention components
increases this effect profoundly and suggests a further decrease of stability to the
already less stable network. The decrease in finger variability and the preliminary
result of almost complete absence of lapses in the DLPFC condition suggests that
inhibition of the L-DLPFC may increase the stability of the network. If this is indeed
the case this supports the notion that interplay between inhibitory and excitatory
influences underlie network stability. The effective link between lower to higher
areas, created by this abnormal connectivity, and allowing TMS perturbations to the
low level area M1 to induce lapses and affect error correction mechanisms, would be
weakened when a mal functional node in the network (the L-DLPFC) is removed.
Further experiments are needed with subjects with Schizophrenia that have lapses
during the baseline condition in order to establish the result that L-DLPFC inhibition
can alleviate lapses.
The inconsistent EEG response pattern evoked by TMS stimulation that was observed
in Schizophrenia further suggests a deficit in network processing probably arising
from mal-connectivity.
It can be interesting to check in future experiments how these TMS evoked responses
change as a function of different manipulations to networks in the healthy and
Schizophrenia brain and if increasing stability (for example by inhibiting a mal-
functional area) produces a clearer response pattern.
107

The results described above support the hypothesis that Schizophrenia is a disorder of
effective connectivity (Friston, 1998; Andreasen et al., 1999). It suggests that when
violation of stability and balance within and probably between networks occurs,
abnormal effective connectivity arises leading to abnormal behaviour. This violation
of stability and balance seems to be caused when the relative contribution of different
nodes of the network is manipulated. This may cause a change in the overall
dynamics, probably due to changes in neuronal activity, changing the relative
influence that a neural system exerts over the others. In our case, this seemed to occur
when a pathology in a certain brain area existed (e.g. the mal-functional D-LPFC), or
due to an external manipulation (e.g changing the attention load, or inhibiting a
certain region). It is more difficult to interpret our results in the framework of the
alternative hypothesis that suggests Schizophrenia to be a deficit of impaired
functional specialization (see Background). The direct contribution of the impairment
in the function of the DLPFC to the Schizophrenia disease seems to be secondary to
its contribution to the impairment in the effective connectivity of the neural network.
This is supported by the fact that perturbations to a lower, seemingly unrelated area
(M1) can impact on high cognitive functions such as attention and induce lapses.
The fact that Schizophrenia is considered a developmental disorder by most
neurologists (Andreasen, 2001), can go together with the notion of stability and
balance. It may reflect a system in which genetic and environmental factors affected
stability of the developing brain network, causing it to be vulnerable to relatively
extreme stress conditions. Interestingly, the onset of the disease parallels the stage in
life when young adult leave the parental nest, need to support themselves and build
their own nest, a relatively stressful condition (Andreasen, 2001; Peled, 2004). These
(or other) extreme stress conditions may cause a breakdown in a network prone to
instabilities, at different levels, resulting in a ‘breakdown in consciousness’ often used
to describe the illness (Andreasen, 1997).
From Schizophrenia to Autism As mentioned in the introduction, some common features can be found between
Schizophrenia and Autism, two heterogeneous deficits involving many aspects of
108

cognitive functions. As in Schizophrenia, the importance of a delicate balance
between processes of specialization and integration for the healthy neural and
cognitive development and its disruption in Autism has recently been emphasized,
(Brock et al., 2002) and supported by animal models (Rubenstein and Merzenich,
2003). In Autism this balance is disrupted in a way that specialized units are
effectively disconnected from one another resulting in deficits in functions that
require high integration such as one’s ability to adjust to an ever changing
environment (inflexibility and rigidity), or one’s social and communication skills.
The importance of balance and stability within and between networks seems to be a
key factor in understanding how the vast spectrum of human conditions arises from
the complex network of the brain. In the following paragraphs I present additional
correlative evidence to further emphasize the role of stability and balance.
Concept of Emergence and the importance of stability and balance
The ‘Dynamicist’ view
Recently, the stage by stage information processing metaphor of the brain is being
abandoned (Engel et al., 2001; Thompson and Varela, 2001). Instead, it has become
increasingly clear to neuroscientists that neurons should be studied as members of
large ensembles that are constantly arising and disappearing in a context dependent
manner. The dense and complex interconnectivity inside the brain suggests that all the
components will eventually have an influence on everything that is going on.
In the ‘dynamicist’ view (Engel et al., 2001), afferent signals are subject to local
computations operating on the input but also to modulation by specific synchronizing
and desynchronizing influences that are mediated by long-range interactions from
distributed areas. These modulatory influences carry contextual predictions that
compete for stable, resonant states of the involved cell populations. A stable state
reflects a successful match of the input to the predictions and result in amplification of
a specific pattern of coherent neural discharges, which supports the stabilization of
large-scale patterns of temporal coherence. Any area can, in principle, modulate any
other area to which it is connected, and what counts is not the superiority in
109

anatomical hierarchy but rather a ‘democratic’ ‘majority votes’ organization
(Churchland and Sejnowski, 1988; Engel et al., 2001). These modulatory effects or
priming of neuronal assemblies do not necessarily require an external stimulus but can
also be triggered by internal processes and interactions, such as input coming from the
internal milieu (homeostatic, autonomic regulation), memories, emotions etc.
Ongoing activity is thus not merely noise but rather a continuous process of
predictions and of priming signals permitting rapid selection among multiple events
or inputs (Kenet et al., 2003).
In this view, the balance between excitation and inhibition (or between
synchronization and desynchronization) is critical to achieve an output signal that is
salient enough to influence other areas and participate in the ongoing processing
(Rippon et al., 2006). It is also important to allow different assemblies to become
dominant at different moments and allow rapid transitions between different states.
This description of the brain as a self organizing system is very appealing. In these
systems the concept of stability is central as the selection of a particular dynamic
pattern from the vast array of possible combinations depends on the relative stability.
An emergent state of self organization depends on the occurrence of sudden
macroscopic transitions between states of differing stability. These systems exhibit
pronounced susceptibility to small disturbances and large-scale alterations can be
caused by minute changes to the system’s state.
Consequently, different theories of emergence and self organization have been used to
describe the brain’s dynamics and recently researchers have tried to find evidence for
this description in the brain. Tools and methods of nonlinear dynamics are
progressively becoming more and more popular in analysis of brain signals such as
EEG and MEG (Stam, 2005).
Evidence for applicability of dynamical system approach to the brain
In humans, the dynamical systems approach to study the brain was widely used with a
large degree of success in the field of movement science (see review by Sternad,
2000). Corroborative evidence for its applicability comes from the observation of
110

electrophysiological changes as a consequence of alterations of stability in motor
behaviour (Bressler and Kelso, 2001; Stam, 2005). Recently, Meyer-Lindenberg et al.,
2002, demonstrated more directly the applicability of nonlinear dynamics to the
brain. In a combined TMS and neuroimaging study, they showed the ability to elicit
transitions between states of movement (fingers tapping in phase or out of phase) that
have differing stability with TMS. TMS caused sustained and macroscopic behavioral
transitions from the unstable out-of-phase movement attractor to the stable in-phase
movement attractor, whereas the stable pattern could not be affected. Moreover the
strength of the TMS perturbation required (considered a measure of neural stability)
was linked to the degree of behavioral stability.
Balance within sensory processing underlie temperament and
behaviour traits
Occupational Therapy research can provide insight
The relation between how the working of the brain can account for the vast spectrum
of human behavior is still an open and mysterious question. A field that can give
important insight and help bridge the gap is the field of Occupational Therapy.
Occupational therapists have accumulated much experience with a wide spectrum of
human conditions and their observations and intervention strategies can be insightful.
Interestingly, occupational therapists use concepts such as “suppression” (the ability
to screen certain stimuli so that other stimuli receive more careful attention), “balance
of power” (i.e. complementary function of the various parts of the brain), “balance of
excitation and inhibition” and “intersensory integration” when they come to
understand the different human conditions they come across in their clinic and design
the best intervention to support performance (Dunn, 1997).
Different sensory processing profiles across people
In studies of populations of children and adults, consistent patterns of temperament,
personality and sensory processing emerge (Dunn and Brown, 1997; Brown et al.,
2001; Dunn, 2001). This suggests that one’s temperament and personality may be the
111

behavioral manifestation of one’s sensory processing patterns and nervous system
functions.
Indeed, when asked to characterize their experiences, people usually describe it from
a sensory point of view, using sensory words to share their experiences. A person is
usually described in terms of his interests in, tolerance for and pleasure for sensations.
People have different thresholds for noticing, responding to, and becoming irritated
with sensations. Internal (sensation of the body, i.e touch, visceral) and external
(sensations of the environment, i.e auditory and visual) information, together with
attentional, memory, motivation and emotional processes modulate sensory
information and affects the way sensory information is processed, causing low
thresholds (higher sensitivity) for some types of sensory channels and higher
thresholds (lower sensitivity) for other sensory inputs. This neural regulation occurs
through mechanisms that balance excitation and inhibition, and reflect the natural
tension that exists between the different modulatory factors (e.g. internal versus
external information) (Dunn, 2001). The interplay between excitation and inhibition is
determined by a combination of genetic and environmental factors.
The combination of threshold levels for the different sensory channels affect people’s
daily choices and are reflected in their mood, temperament, attention levels, problem
solving strategies and ways of organising their life (Baranek et al., 1997; Dunn and
Brown, 1997; Dunn, 2001). Cognitive processing is optimal when the internal and
external information processing both enable task performance. These combinations of
thresholds are not rigid for each individual and can change during the day and
throughout life (Dunn, 2001).
A model for sensory processing
Recently, a model for sensory processing that characterizes patterns of responding to
task and environmental demands, was proposed (Dunn, 2001). These patterns are
based on a person’s neurological thresholds and self regulation strategies. The
neurological threshold continuum (the amount of input the nervous system requires
before responding) moves from low to high thresholds. The behavioral response
112

continuum moves from self-regulation strategies that are passive (tendency to let
things happen) to those that are active (attempting to control the incoming input).
When these continua intersect, four basic patterns of sensory processing emerge,
representing the anchor points of the continua (Dunn et al., 2002).
Persons that have high neurological thresholds with passive self regulation strategies
(low registration), do not notice sensory events in daily life that others notice readily.
They may not notice when other people come into the room or food or dirt on their
face and hands. Others may have to call the person’s name several times or use
additional cues to catch the person’s attention (Dunn et al., 2002).
Persons who have high neurological thresholds but an active self regulation strategy
are sensation seekers. They enjoy sensory experiences and find ways to enhance and
extend sensory events in daily life. They like physical movements such as climbing,
twirling, swinging, and bouncing. They search for additional sensory experiences for
themselves, such as humming and other mouth noises, touching objects, feeling
vibrations in stereo speakers and appliances, wearing perfume, and smelling flowers
(Dunn, 2001; Dunn et al., 2002).
Persons who have low neurological thresholds experience more sensory events then
others in daily life. They are easily distracted by various sensory cues such as
movement, sound or smell. They are more sensitive to temperature, food textures, and
certain fabric textures. Those with an active self-regulation strategy are sensory
avoiders, finding ways to limit sensory input throughout daily life. They stay away
from distracting settings, leave the room when noise levels are too high, or people are
bumping into them. They adhere to daily routines in order to generate only familiar,
predictable sensory patterns and become unhappy when these rituals are disrupted
perhaps because of increasing unpredictability (Dunn, 2001; Dunn et al., 2002).
Sensory processing patterns are different for people with disorders
Studies indicate that sensory processing is significantly different for persons with
disorders and various disabilities, such as depression, ADHD, Autism, Asperger
syndrome, Schizophrenia and sensory modulation disorder (Brown et al., 2002; Dunn
and Bennett, 2002; Myles et al., 2004).
113

Interestingly, people with Autism and Schizophrenia have significant sensory
processing disturbances (Brown et al., 2002). In Autism, children with Asperger
syndrome and Autism have significantly different sensory processing patterns than
normal children (Dunn et al., 2002; Myles et al., 2004). Rigidity and inflexibility,
found for example in people with Autism, reflect coping with low thresholds that
quickly overwhelm some person’s nervous systems (Baranek et al., 1997). Moreover,
in some cases severe communication and relationship disorders may be secondary to
various sensory and motor processing deficits which can be conceptualized as
Multisystem Developmental Disorder (MSDD) rather than Autistic disorders (Wieder,
1996).
In Schizophrenia, McGhie and Chapman, 1961, identified disturbances of attention,
perception, and changes in motility and body awareness as key features of the onset of
the disease. They categorized people’s description of their early experiences with
Schizophrenia. The description which emerged was that of having to face an unstable
and newly fluctuating relationship between perception of self (body sensations) and
the environment. During this early period, persons seem to have increasing awareness
of all stimuli and cannot organize the myriad of sensations, making it more and more
difficult to function. Recently it had been proposed that delusions emerge from
attempts to make meaning out of this increasingly undifferentiated sensory input
(Frith et al., 2000). Sensory processing challenges were linked to the cognitive
impairments characteristic of the disorder (Frith et al., 2000). Brown et al., 2001,
described persons with Schizophrenia as having simultaneously low registration
(reflecting high threshold and low sensitivity) and sensory avoidance (caused by very
high sensitivity and an attempt to avoid being overwhelmed) (Brown et al., 2002).
Dunn hypothesized that this pattern of poor noticing on the one hand, and
hyperresponsivenss leading to withdrawal (sensory avoidance), on the other hand,
may reflect the small range within which these persons can receive sensory input and
use it to participate successfully in daily activities(Dunn, 2001).
Relation between sensory processing and temperament and personality traits
Researchers who have studied temperament and personality traits found factor
structures that seem to reflect levels of reactivity suggesting the possible impact that
sensory processing may have on behavioral and personality traits (Dunn, 2001;
114

Rothbart et al., 2003; Nigg, 2006). For example, sensory avoidance is associated with
negative affect and neuroticism; sensory seeking with extraversion, agreeableness and
openness; low registration with persistence, effortful attention and conscientiousness;
and sensory sensitivity to irritability/anger (Dunn, 2001).
Evidence from Practice
If indeed irregularities in the balance of excitation and inhibition underlie the core
deficits in disorders such as Autism, then efficient interventions should directly
address these irregularities. Recently, sensory intervention programs that try to
regulate abnormalities in sensory processing, have been introduced and have been
found to improve behavior, such as a decrease in the frequency of non engaged
behavior and an increase in the frequency of goal directed behavior (Case-Smith and
Bryan, 1999; Baranek, 2002; Schaaf and Miller, 2005). Dynamic developmental
programs that consider the child's functional emotional developmental capacities,
individual differences in sensory processing and modulation in motor planning and
sequencing, as well as child/caregiver and family interaction (Greenspan and Wieder,
1999) are gaining popularity over the traditional behavioral intervention therapies that
put emphasis on teaching outward behavior and concrete skills.
The picture outlined above suggests that we are the result of the coherent patterns
emerging from the complex dynamics of our brain, which in turn is the result of the
balance and stability between the densely connected sub networks (underlying
sensory, emotional, vestibular, homeostasis processing) that constitute the global
network of the brain. At any one time, the activity of any cortical area and its outputs
to other areas will reflect and influence virtually all the activity in the entire brain.
Variations in this balance accounts for the variability between people and
abnormalities in this balance and instability of networks account for abnormal human
conditions. Is it really necessary that there be anything else?
What about a ‘Self’ with a freewill?
“You enter the brain through the eye, march up the optic nerve and round the cortex,
looking behind every neuron, and then before you know it, you emerge into daylight
115

on the spike of a motor nerve impulse, scratching your head and wondering where the
self is”. (Dennett, Elbow Room 74-75). Taken from (Varela et al., 1993)
Is this indeed all there it to it? Am I simply the imprint of the interactions between my
sensory systems and other systems in the brain and the resultant emergent self-
organisational dynamics? But what about ‘I’, ‘Me’, the strong feeling of ‘Self’ that
has awareness and a free will to chose to make a movement?
The Dualistic (mind and brain are separate) versus the Monistic (mind is a product of
brain) view are great questions at the heart of neuroscience and Philosophy and this
thesis cannot pretend to give a full and closed answer. However, some evidence does
exist to support monism, that there may really be no such thing as free will, an inner
causal agent or a thing that is not identical with one’s body or with anything physical.
Examples of evidence supporting monism Libet et al., 1983, presented results that suggested that freewill is a perception. They
asked subjects to view a clock with a moving spot and say at what time they first had
the subjective experience of intending to act (termed W) and when the time of
awareness of actually moving (termed M). To control the ability of correctly judging
such times, random stimulations to the skin were applied and subjects were also asked
to time these events (termed S). Movement related cortical potentials were assessed
using EEG recorded simultaneously from subjects’ scalps. The movement related
cortical potentials have a number of components. Two components precede
movement and are called the readiness potentials (termed BP1 and BP2) and only the
third component is the motor potential. Depending on whether the movement was
preplanned or spontaneous, BP1 began 1050 ms and 575 ms prior to EMG onset,
respectively. W and M occurred about 300 ms and 90 ms, respectively, prior to EMG
onset. BP1 occurred 800 ms and 350 ms prior to W in the case of the preplanned
movements and spontaneous movement respectively. This provides evidence that the
brain initiates a movement before awareness of volition actually occurs. The
subjective sense of having moved preceded the actual onset of movement suggesting
that the brain assumes that if it issues a motor command, the movement will be
generated. This is supported by an experiment (Haggard and Magno, 1999) where
116

TMS over the motor cortex caused a delay in the RT for movement, but less in the
judgment of when the movement occurred. This suggests that the motor cortex may
be down stream from the site of movement awareness.
Taylor and McColsky, 1990, used “backward masking” (i.e a rapid sequential
presentation of two visual stimuli in a way that the first stimulus is not perceived) and
showed that reaction time to the masked stimulus were the same as those for
responses to the easily perceived, nonmasked stimulus. Thus voluntary movements
can be triggered with stimuli that are not perceived.
Brasil-Neto et al., 1992, showed that TMS can bias motor choice. Subjects were asked
to move their right or left hand upon hearing the click of the magnetic coil. TMS
affected hand preference only when it was delivered to the motor area and only if
movement was initiated within the first 200 ms after stimulation. Thus freely chosen
movements can be externally biased without perception of external influence and
without disrupting the conscious perception of volition.
In the context of all that was said above, it is interesting to note that Schizophrenia
and Autism may contribute also to the understanding of ‘self’ as it presents a case of
self-dysfunction. During a psychotic episode, Schizophrenia subjects may have good
autobiographical memory, but suffer deep confusion about self / noself boundaries.
An example is the phantom hand experiments where patients respond to a tactile
stimulus but claim that the sensation belongs to someone else (Peled et al., 2003).
Another example are the auditory hallucinations that appear to be the patients’ own
thoughts or inner speech but are not recognized as such (Churchland, 2002). In
Autism, the sense of self seems also to be disrupted. For example, in patients with
Autism, a typical reduction in activity in several cortical areas known as the 'default
state' network (Greicius et al., 2003) implicated in self-referential processing, was not
observed (Iacoboni, 2006). Impairment of self-face recognition has also been reported
in these patients (Irani et al.).
117

A philosophical note
“For my part, when I enter most intimately into what I call myself. I always stumble
on some particular perception or other, of heat or cold, light or shade, love or hatred,
pain or pleasure. I never catch myself at any time without a perception, and never can
observe anything but the perception.” David Hume
I would like to end this thesis with a very personal and perhaps philosophical
perspective. This view is based upon recent attempts of western neuroscientists to
expand and open the understanding of the different human experiences to other
conditions which are less familiar to the Western mind but are a daily experience to
many in the East.
Earlier, the fragmented experience of people with Schizophrenia was described,
reporting confused thinking, difficulties in connecting ideas, trouble filtering
unimportant information etc (Andreasen, 2001). This feeling of fragmentation, at
times, may also arise in what is considered a 'normal and healthy' experience.
An example for such fragmentation may be found in the simple situation when people
are asked to sit quietly and observe one’s mind. They often report upon feeling their
mind wander and finding it hard to stay connected to their momentary experience for
long intervals. In addition there is often a feeling of a hard to control turmoil activity
in the mind, where perceptions, thoughts, feelings, desires, fears, and every other kind
of mental content pursue each other endlessly.
This difficulty of being mindful of one’s experiences without the mind wandering
uncontrollably, being at times aware and at other times unaware of one’s state gives
rise to a feeling of fragmentation that indeed may give rise to a dualistic point of view
in which mind and brain are separate.
Eastern meditation practices give rise to a somewhat different experience. The
firsthand account of serious practitioners in Eastern meditation, who attain a high
degree of mindfulness, is that with training, the relative time during which they can be
mindful of their daily experience grows progressively. They often report that when
118

this is achieved, their experience becomes nothing more than a constant stream of
perceptions and the feeling of fragmentation is much less frequent.
A recent surge in research comparing highly trained meditators to untrained
individuals indeed has revealed differences in EEG patterns, in cortical structure and
in responses to the perceptual rivalry task (Lutz et al., 2004; Carter et al., 2005;
Cysarz and Bussing, 2005; Lazar et al., 2005). This suggests that understanding this
specific human experience may provide further insight to the understanding of the
wide spectrum of human experiences mentioned above.
The possible contribution of the Buddhist tradition to the scientific enterprise. To my mind, of the eastern traditions Buddhism seems particularly well suited to
interact with science because of its development of techniques and systematic
methodologies to investigate the human experience (Varela et al., 1993). Doctrines of
no self and of nondualism grew out of this method and may have a significant
contribution to make to the discussion of dualism versus monism, through a dialogue
with cognitive science (Varela et al., 1993).
In the context of all that was said in this Thesis, and if indeed the reports by highly
trained mediators of a less fragmented experience is correct, then these practitioners
may represent a highly stable brain condition that stands at the other end of the
spectrum from the Schizophrenia and Autism condition. Comparing stability and
connectivity in brain networks across these two sides of the spectrum may provide
further insight to how human behaviour arises and may suggest therapeutic
interventions to increase stability in presumably less stable brains.
119


Bibliography
Alain C, McNeely HE, He Y, Christensen BK, West R (2002) Neurophysiological evidence of error-monitoring deficits in patients with schizophrenia. Cerebral Cortex 12:840-846.
Aleman A, Schutter DJ, Ramsey NF, van Honk J, Kessels RP, Hoogduin JM, Postma A, Kahn RS, de Haan EH (2002) Functional anatomy of top-down visuospatial processing in the human brain: evidence from rTMS. Brain Res Cogn Brain Res 14:300-302.
Amassian VE, Cracco RQ, Maccabee PJ, Cracco JB (1992) Cerebellofrontal Cortical Projections in Humans Studied with the Magnetic Coil. Electroencephalography and Clinical Neurophysiology 85:265-272.
Andreasen NC (1997) Linking Mind and Brain in the Study of Mental Illnesses: A Project for a Scientific Psychopathology. Science 275:1586-1593.
Andreasen NC (2000) Schizophrenia: the fundamental questions. Brain Research Reviews 31:106-112.
Andreasen NC (2001) Brave New Brain: Conquering Mental Illness in the Era of the Genome. New York: Oxford University Press.
Andreasen NC, Nopoulos P, O'Leary DS, Miller DD, Wassink T, Flaum M (1999) Defining the phenotype of schizophrenia: cognitive dysmetria and its neural mechanisms. Biol Psychiatry 46:908-920.
Andreasen NC, O'Leary DS, Cizadlo T, Arndt S, Rezai K, Ponto LL, Watkins GL, Hichwa RD (1996) Schizophrenia and cognitive dysmetria: a positron-emission tomography study of dysfunctional prefrontal-thalamic-cerebellar circuitry. Proc Natl Acad Sci U S A 93:9985-9990.
Aschersleben G, Prinz W (1995) Synchronizing Actions with Events - the Role of Sensory Information. Perception & Psychophysics 57:305-317.
Bailey CJ, Karhu J, Ilmoniemi RJ (2001) Transcranial magnetic stimulation as a tool for cognitive studies. Scandinavian Journal of Psychology 42:297-305.
Balasubramaniam R, Wing AM, Daffertshofer A (2004) Keeping with the beat: movement trajectories contribute to movement timing. Experimental Brain Research 159:129-134.
Baranek GT (2002) Efficacy of sensory and motor interventions for children with autism. Journal of Autism and Developmental Disorders 32:397-422.
Baranek GT, Foster LG, Berkson G (1997) Tactile defensiveness and stereotyped behaviors. American Journal of Occupational Therapy 51:91-95.
Berardelli A, Inghilleri M, Polidori L, Priori A, Mercuri B, Manfredi M (1994) Effects of transcranial magnetic stimulation on single and sequential arm movements. Exp Brain Res 98:501-506.
Bohotin V, Fumal A, Vandenheede M, Gerard P, Bohotin C, Maertens de Noordhout A, Schoenen J (2002) Effects of repetitive transcranial magnetic stimulation on visual evoked potentials in migraine. Brain 125:912-922.
Bonnard M, Camus M, de Graaf J, Pailhous J (2003) Direct evidence for a binding between cognitive and motor functions in humans: A TMS study. Journal of Cognitive Neuroscience 15:1207-1216.
Brasil-Neto JP, Pascual-Leone A, Valls-Sole J, Cohen LG, Hallett M (1992) Focal transcranial magnetic stimulation and response bias in a forced-choice task. J Neurol Neurosurg Psychiatry 55:964-966.
120

Berg, P and M. Scherg (1994) A multiple source approach to the correction of eye artifacts. Electroencephalogr Clin Neurophysiol 90(3): 229-41.
Bressler SL, Kelso JAS (2001) Cortical coordination dynamics and cognition. Trends in Cognitive Sciences 5:26-36.
Brock J, Brown CC, Boucher J, Rippon G (2002) The temporal binding deficit hypothesis of autism. Development and Psychopathology 14:209-224.
Brown C, Tollefson N, Dunn W, Cromwell R, Filion D (2001) The adult sensory profile: Measuring patterns of sensory processing. American Journal of Occupational Therapy 55:75-82.
Brown C, Cromwell RL, Filion D, Dunn W, Tollefson N (2002) Sensory processing in schizophrenia: missing and avoiding information. Schizophrenia Research 55:187-195.
Brown C, Gruber T, Boucher J, Rippon G, Brock J (2005) Gamma abnormalities during perception of illusory figures in autism. Cortex 41(3): 364-376.
Burt T, Lisanby SH, Sackeim HA (2002) Neuropsychiatric applications of transcranial magnetic stimulation: a meta analysis. Int J Neuropsychopharmacol 5:73-103.
Callicott JH, Mattay VS, Verchinski BA, Marenco S, Egan MF, Weinberger DR (2003) Complexity of prefrontal cortical dysfunction in schizophrenia: more than up or down. Am J Psychiatry 160.
Carter OL, Presti DE, Callistemon C, Ungerer Y, Liu GB, Pettigrew JD (2005) Meditation alters perceptual rivalry in Tibetan Buddhist monks. Current Biology 15:R412-R413.
Case-Smith J, Bryan T (1999) The effects of occupational therapy with sensory integration emphasis on preschool-age children with autism. American Journal of Occupational Therapy 53:489-497.
Case-Smith J, Miller H (1999) Occupational therapy with children with pervasive developmental disorders. American Journal of Occupational Therapy 53:506-513.
Chouinard PA, Van der Werf YD, Leonard G, Paus T (2003) Modulating neural networks with transcranial magnetic stimulation applied over the dorsal premotor and primary motor cortices. Journal of Neurophysiology 90:1071-1083.
Churchland PS (2002) Self-representation in nervous systems. Science 296. Churchland PS, Sejnowski TJ (1988) Perspectives on cognitive neuroscience. Science 242. Cohen LG (2000) A window into the role of inhibitory and excitatory mechanisms of
perception? J Physiol 529 Pt 2:283. Cohen MR, Newsome WT (2004) What electrical microstimulation has revealed about the
neural basis of cognition. Curr Opin Neurobiol 14. Collyer CE, Broadbent HA, Church RM (1994) Preferred rates of repetitive tapping and
categorical time production. Percept Psychophys 55:443-453. Cortese L, Caligiuri MP, Malla AK, Manchanda R, Takhar J, Haricharan R (2005)
Relationship of neuromotor disturbances to psychosis symptoms in first-episode neuroleptic-naive schizophrenia patients. Schizophr Res 75.
Crammond DJ, Kalaska JF (2000) Prior information in motor and premotor cortex: activity during the delay period and effect on pre-movement activity. J Neurophysiol 84:986-1005.
Cysarz D, Bussing A (2005) Cardiorespiratory synchronization during Zen meditation. European Journal of Applied Physiology 95:88-95.
d'Alfonso AA, Aleman A, Kessels RP, Schouten EA, Postma A, van Der Linden JA, Cahn W, Greene Y, de Haan EH, Kahn RS (2002) Transcranial magnetic stimulation of left auditory cortex in patients with schizophrenia: effects on hallucinations and neurocognition. J Neuropsychiatry Clin Neurosci 14:77-79.
121

Dalley JW, Cardinal RN, Robbins TW (2004) Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neurosci Biobehav Rev 28:771-784.
Daskalakis ZJ, Christensen BK, Fitzgerald PB, Fountain SI, Chen R (2005) Reduced cerebellar inhibition in schizophrenia: A preliminary study. American Journal of Psychiatry 162:1203-1205.
Daskalakis ZJ, Kapur S, Christensen BK, Chen R, Fitzgerald PB, Zipursky RB (2001) Direct demonstration of impaired cortical inhibition in schizophrenia using TMS. Schizophrenia Research 49:200-201.
Daskalakis ZJ, Christensen BK, Chen R, Fitzgerald PB, Zipursky RB, Kapur S (2002) Evidence for impaired cortical inhibition in schizophrenia using transcranial magnetic stimulation. Arch Gen Psychiatry 59:347-354.
Davalos DB, Kisley MA, Ross RG (2003) Effects of interval duration on temporal processing in schizophrenia. Brain and Cognition 52:295-301.
Day BL, Rothwell JC, Thompson PD, Maertens de Noordhout A, Nakashima K, Shannon K, Marsden CD (1989) Delay in the execution of voluntary movement by electrical or magnetic brain stimulation in intact man. Evidence for the storage of motor programs in the brain. Brain 112 ( Pt 3):649-663.
Delignieres D, Lemoine L, Torre K (2004) Time intervals production in tapping and oscillatory motion. Hum Mov Sci 23:87-103.
Desmurget M, Epstein CM, Turner RS, Prablanc C, Alexander GE, Grafton ST (1999) Role of the posterior parietal cortex in updating reaching movements to a visual target. Nat Neurosci 2:563-567.
Dhamala M, Pagnoni G, Wiesenfeld K, Zink CF, Martin M, Berns GS (2003) Neural correlates of the complexity of rhythmic finger tapping. Neuroimage 20:918-926.
Dolan RJ, Fletcher PC, McKenna P, Friston KJ, Frith CD (1999) Abnormal neural integration related to cognition in schizophrenia. Acta Psychiatr Scand Suppl 395:58-67.
Doumas M, Praamstra P, Wing AM (2005) Low frequency rTMS effects on sensorimotor synchronization. Experimental Brain Research 167:238-245.
Drewing K, Hennings M, Aschersleben G (2002) The contribution of tactile reafference to temporal regularity during bimanual finger tapping. Psychological Research-Psychologische Forschung 66:60-70.
Dunn W (1997) Implementing neuroscience principles to support habilitation and recovery. In: Occupational Therapy: Enabling Function and Well-Being. (Christiansen C, Baum C, eds), pp 186-232. Thorofare NJ: Slack Incorporated.
Dunn W (2001) The sensations of everyday life: Empirical, theoretical, and pragmatic considerations. American Journal of Occupational Therapy 55:608-620.
Dunn W, Brown C (1997) Factor analysis on the sensory profile from a national sample of children without disabilities. American Journal of Occupational Therapy 51:490-495.
Dunn W, Bennett D (2002) Patterns of sensory processing in children with Attention Deficit Hyperactivity Disorder. Otjr-Occupation Participation and Health 22:4-15.
Dunn W, Myles BS, Orr S (2002) Sensory processing issues associated with Asperger syndrome: a preliminary investigation. American Journal of Occupational Therapy 56:97-102.
Elvevag B, McCormack T, Gilbert A, Brown GDA, Weinberger DR, Goldberg TE (2003) Duration judgements in patients with schizophrenia. Psychological Medicine 33:1249-1261.
Engbert R, Krampe RT, Kurths J, Kliegl R (2002) Synchronizing movements with the metronome: Nonlinear error correction and unstable periodic orbits. Brain and Cognition 48:107-116.
122

Engel AK, Singer W (2001) Temporal binding and the neural correlates of sensory awareness. Trends in Cognitive Sciences 5:16-25.
Engel AK, Fries P, Singer W (2001) Dynamic predictions: Oscillations and synchrony in top-down processing. Nature Reviews Neuroscience 2:704-716.
Evers S, Bockermann I, Nyhuis PW (2001) The impact of transcranial magnetic stimulation on cognitive processing: an event-related potential study. Neuroreport 12:2915-2918.
First MB, Spitzer RL, Gibbon M, Williams JBW (1995) SCID (DSM-IV) Structured clinical interview for Axis I DSM-IV Disorders - Patients Edition (SCID-I/P). New York: Biometrics Research Department, New York State Psychiatric Institute.
Fitzgerald PB, Brown TL, Daskalakis ZJ, Kulkarni J (2002a) Tms reveals abnormal plasticity in schizophrenia. Schizophrenia Research 53:218-219.
Fitzgerald PB, Brown TL, Daskalakis ZJ, deCastella A, Kulkarni J (2002b) A study of transcallosal inhibition in schizophrenia using transcranial magnetic stimulation. Schizophr Res 56:199-209.
Flyckt L, Sydow O, Bjerkenstedt L, Edman G, Rydin E, Wiesel FA (1999) Neurological signs and psychomotor performance in patients with schizophrenia, their relatives and healthy controls. Psychiatry Research 86:113-129.
Friston K (2002a) Functional integration and inference in the brain. Progress in Neurobiology 68:113-143.
Friston K (2002b) Beyond phrenology: What can neuroimaging tell us about distributed circuitry? Annual Review of Neuroscience 25:221-250.
Friston KJ (1996) Theoretical neurobiology and schizophrenia. Br Med Bull 52:644-655. Friston KJ (1998) The disconnection hypothesis. Schizophr Res 30:115-125. Friston KJ (1999) Schizophrenia and the disconnection hypothesis. Acta Psychiatr Scand
Suppl 395:68-79. Friston KJ, Frith CD (1995) Schizophrenia - a Disconnection Syndrome. Clinical
Neuroscience 3:89-97. Frith CD, Blakemore SJ, Wolpert DM (2000) Explaining the symptoms of schizophrenia:
Abnormalities in the awareness of action. Brain Research Reviews 31:357-363. Gangitano M, Mottaghy FM, Pascual-Leone A (2001) Phase-specific modulation of cortical
motor output during movement observation. Neuroreport 12:1489-1492. Garcia-Toro M, Blanco C, Gonzalez A, Salva J (2001) Psychopathology and the binding
problem. Med Hypotheses 57:718-723. Geller V, Grisaru N, Abarbanel JM, Lemberg T, Belmaker RH (1997) Slow magnetic
stimulation of prefrontal cortex in depression and schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry 21:105-110.
Gerloff C, Corwell B, Chen R, Hallett M, Cohen LG (1997) Stimulation over the human supplementary motor area interferes with the organization of future elements in complex motor sequences. Brain 120 ( Pt 9):1587-1602.
Gray CM (1999) The temporal correlation hypothesis of visual feature integration: Still alive and well. Neuron 24:31-47.
Greenspan SI, Wieder S (1999) A functional developmental approach to autism spectrum disorders. Journal of the Association for Persons with Severe Handicaps 24:147-161.
Greicius MD, Krasnow B, Reiss AL, Menon V (2003) Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. PNAS 100:253-258.
Grondin S, Rammsayer T (2003) Variable foreperiods and temporal discrimination. Q J Exp Psychol A 56.
Haggard P, Magno E (1999) Localising awareness of action with transcranial magnetic stimulation. Exp Brain Res 127:102-107.
123

Haraldsson HM, Ferrarelli F, Kalin NH, Tononi G (2004) Transcranial Magnetic Stimulation in the investigation and treatment of schizophrenia: a review. Schizophrenia Research 71:1-16.
Harris AWF, Bahramali H, Slewa-Younan S, Gordon E, Williams L, Li WM (2001) The topography of quantified electroencephalography in three syndromes of schizophrenia. International Journal of Neuroscience 107:265-278.
Ho BC, Mola C, Andreasen NC (2004) Cerebellar dysfunction in neuroleptic naive schizophrenia patients: Clinical, cognitive, and neuroanatomic correlates of cerebellar neurologic signs. Biological Psychiatry 55:1146-1153.
Hoffman RE, McGlashan TH (2001) Neural network models of schizophrenia. Neuroscientist 7:441-454.
Hoffman RE, Boutros NN, Berman RM, Roessler E, Belger A, Krystal JH, Charney DS (1999) Transcranial magnetic stimulation of left temporoparietal cortex in three patients reporting hallucinated "voices". Biol Psychiatry 46:130-132.
Hogan N, Bizzi E, Mussa-Ivaldi FA, Flash T (1987) Controlling multijoint motor behavior. Exerc Sport Sci Rev 15:153-190.
Hoy KE, Fitzgerald PB, Bradshaw JL, Farrow M, Brown TL, Armatas CA, Georgiou-Karistianis N (2004) Motor overflow in schizophrenia. Psychiatry Research 125:129-137.
Iacoboni M (2006) Failure to deactivate in autism: the co-constitution of self and other. Trends in Cognitive Sciences 10:431-433.
Ilmoniemi RJ, Virtanen J, Ruohonen J, Karhu J, Aronen HJ, Naatanen R, Katila T (1997). Neuronal responses to magnetic stimulation reveal cortical reactivity and connectivity. NeuroReport 8: 3537-3540.
Irani F, Platek SM, Panyavin IS, Calkins ME, Kohler C, Siegel SJ, Schachter M, Gur RE, Gur RC Self-face recognition and theory of mind in patients with schizophrenia and first-degree relatives. Schizophrenia Research In Press, Corrected Proof.
Itil TM (1977) Qualitative and Quantitative Eeg Findings in Schizophrenia. Schizophrenia Bulletin 3:61-79.
Ivry RB, Richardson TC (2002) Temporal control and coordination: The multiple timer model. Brain and Cognition 48:117-132.
Ivry RB, Spencer RMC (2004) The neural representation of time. Current Opinion in Neurobiology 14:225-232.
Ivry RB, Spencer RM, Zelaznik HN, Diedrichsen J (2002) The cerebellum and event timing. In: Crebellum: Recent Developments in Cerebellar Research, pp 302-317.
Jahanshahi M, Rothwell J (2000) Transcranial magnetic stimulation studies of cognition: an emerging field. Experimental Brain Research 131:1-9.
Jancke L, Steinmetz H, Benilow S, Ziemann U (2004) Slowing fastest finger movements of the dominant hand with low-frequency rTMS of the hand area of the primary motor cortex. Exp Brain Res 155:196-203.
Jantzen KJ, Steinberg FL, Kelso JAS (2004) Brain networks underlying human timing behavior are influenced by prior context. Proceedings of the National Academy of Sciences of the United States of America 101:6815-6820.
Jing HK, Takigawa M (2000) Observation of EEG coherence after repetitive transcranial magnetic stimulation. Clinical Neurophysiology 111:1620-1631.
Kasai K, Okazawa K, Nakagome K, Hiramatsu KI, Hata A, Fukuda M, Honda M, Miyauchi M, Matsushita M (1999) Mismatch negativity and N2b attenuation as an indicator for dysfunction of the preattentive and controlled processing for deviance detection in schizophrenia: a topographic event-related potential study. Schizophrenia Research 35:141-156.
124

Kay BA, Saltzman EL, Kelso JAS (1991) Steady-State and Perturbed Rhythmical Movements - a Dynamic Analysis. Journal of Experimental Psychology-Human Perception and Performance 17:183-197.
Kay S, Fisbein A, Opler L (1989) The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophrenia Bulletin 13:261-276.
Keel JC, M.J. S, Wassermann EM (2000) A safety screening questionnaire for transcranial magnetic stimulation. Clinical Neurophysiology 112:720.
Kenet T, Bibitchkov D, Tsodyks M, Grinvald A, Arieli A (2003) Spontaneously emerging cortical representations of visual attributes. Nature 425:954-956.
Klimesch W, Sauseng P, Gerloff C (2003) Enhancing cognitive performance with repetitive transcranial magnetic stimulation at human individual alpha frequency. European Journal of Neuroscience 17:1129-1133.
Koenig T, Lehmann D, Saito N, Kuginuki T, Kinoshita T, Koukkou M (2001) Decreased functional connectivity of EEG theta-frequency activity in first-episode, neuroleptic-naive patients with schizophrenia: preliminary results. Schizophrenia Research 50:55-60.
Kotter R, Sommer FT (2000) Global relationship between anatomical connectivity and activity propagation in the cerebral cortex. Philos Trans R Soc Lond B Biol Sci 355.
Lachaux JP, Rodriguez E, Martinerie J, Adam C, Hasboun D, Varela FJ (2000) A quantitative study of gamma-band activity in human intracranial recordings triggered by visual stimuli. Eur J Neurosci 12:2608-2622.
Lau HC, Rogers RD, Ramnani N, Passingham RE (2004) Willed action and attention to the selection of action. Neuroimage 21.
Lazar SW, Kerr CE, Wasserman RH, Gray JR, Greve DN, Treadway MT, McGarvey M, Quinn BT, Dusek JA, Benson H, Rauch SL, Moore CI, Fischl B (2005) Meditation experience is associated with increased cortical thickness. Neuroreport 16:1893-1897.
Le Van Quyen M, Adam C, Lachaux JP, Martinerie J, Baulac M, Renault B, Varela FJ (1997) Temporal patterns in human epileptic activity are modulated by perceptual discriminations. Neuroreport 8:1703-1710.
Lee KH, Williams LM, Breakspear M, Gordon E (2003) Synchronous Gamma activity: a review and contribution to an integrative neuroscience model of schizophrenia. Brain Research Reviews 41:57-78.
Leff AP, Scott SK, Rothwell JC, Wise RJ (2001) The planning and guiding of reading saccades: a repetitive transcranial magnetic stimulation study. Cereb Cortex 11:918-923.
Lepage R, Beaudoin G, Boulet C, O'Brien I, Marcantoni W, Bourgouin P, Richer F (1999) Frontal cortex and the programming of repetitive tapping movements in man: lesion effects and functional neuroimaging. Cognitive Brain Research 8:17-25.
Leucht S, Pitschel-Walz G, Abraham D, Kissling W (1999) Efficacy and extrapyramidal side-effects of the new antipsychotics olanzapine, quetiapine, risperidone, and sertindole compared to conventional antipsychotics and placebo. A meta-analysis of randomized controlled trials. Schizophrenia Research 35:51-68.
Lewis DA, Gonzalez-Burgos G (2006) Pathophysiologically based treatment interventions in schizophrenia. 12:1016-1022.
Lewis PA, Wing AM, Pope PA, Praamstra P, Miall RC (2004) Brain activity correlates differentially with increasing temporal complexity of rhythms during initialisation, synchronisation, and continuation phases of paced finger tapping. Neuropsychologia 42:1301-1312.
125

Libet B, Gleason CA, Wright EW, Pearl DK (1983) Time of Conscious Intention to Act in Relation to Onset of Cerebral-Activity (Readiness-Potential) - the Unconscious Initiation of a Freely Voluntary Act. Brain 106:623-642.
Litvak V, Levit-Binnun N, Handzy N, Moses E, Pratt H, Zaaroor M, Peled A (2007) Differences in the TMS evoked EEG responses between Schizophrenia patients and normal controls. In preparation.
Litvak V, Komssi S, Schreg M, Hoechstetter K, Classen J, Zaaroor M, Pratt H, Kahkonen S (2007). Artifact correction and source analysis of early electroencephalographic responses evoked by transcranial magnetic stimulation over primary motor cortex. Neuroimage May 18 2007.
Lutz A, Greischar LL, Rawlings NB, Ricard M, Davidson RJ (2004) Long-term meditators self-induce high-amplitude gamma synchrony during mental practice. Proceedings of the National Academy of Sciences of the United States of America 101:16369-16373.
MacDonald AW, 3rd, Cohen JD, Stenger VA, Carter CS (2000) Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 288.
Mangels JA, Ivry RB, Shimizu N (1998) Dissociable contributions of the prefrontal and neocerebellar cortex to time perception. Cognitive Brain Research 7:15-39.
Manschreck TC, Maher BA, Rucklos ME, Vereen DR (1982) Disturbed Voluntary Motor-Activity in Schizophrenic Disorder. Psychological Medicine 12:73-84.
Manschreck TC, Maher BA, Rucklos ME, Vereen DR, Ader DN (1981) Deficient Motor Synchrony in Schizophrenia. Journal of Abnormal Psychology 90:321-328.
Manschreck TC, Maher BA, Waller NG, Ames D, Latham CA (1985) Deficient Motor Synchrony in Schizophrenic Disorders - Clinical Correlates. Biological Psychiatry 20:990-1002.
Massimini M, Ferrarelli F, Huber R, Esser SK, Singh H, Tononi G (2005a) Breakdown of Cortical Effective Connectivity During Sleep. Science 309:2228-2232.
Massimini M, Ferrarelli F, Peterson MJ, Esser SK, Huber R, Lazar M, Alexander A, Tononi G (2005b) Cortical EEG response to TMS in schizophrenia patients and normal controls. In: Society for Neuroscience, p Program No. 443.445. Washington DC.
Mauk MD, Buonomano DV (2004) The neural basis of temporal processing. Annual Review of Neuroscience 27:307-340.
Mayville JM, Jantzen KJ, Fuchs A, Steinberg FL, Kelso JAS (2002) Cortical and subcortical networks underlying syncopated and synchronized coordination revealed using fMRI. Human Brain Mapping 17:214-229.
McGhie A, Chapman J (1961) Disorders of attention and perception in early schizophrenia. British Journal of Medical Psychology 34:103-116.
Mesulam M (1998) From sensation to cognition. Brain 121:1013-1052. Meyer BU, Voss M (2000) Delay of the execution of rapid finger movement by magnetic
stimulation of the ipsilateral hand-associated motor cortex. Experimental Brain Research 134:477-482.
Meyer-Lindenberg A, Ziemann U, Hajak G, Cohen L, Berman KF (2002) Transitions between dynamical states of differing stability in the human brain. Proc Natl Acad Sci U S A 99:10948-10953.
Middleton FA, Strick PL (1994) Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function. Science 266:458-461.
Moldin SO, Rubenstein JLR, Hyman SE (2006) Can autism speak to neuroscience? Journal of Neuroscience 26:6893-6896.
Monno A, Temprado J-J, Zanone P-G, Laurent M (2002) The interplay of attention and bimanual coordination dynamics. Acta Psychol (Amst) 110.
126

Mottaghy FM, Gangitano M, Sparing R, Krause BJ, Pascual-Leone A (2002) Segregation of areas related to visual working memory in the prefrontal cortex revealed by rTMS. Cereb Cortex 12:369-375.
Mottaghy FM, Krause BJ, Kemna LJ, Topper R, Tellmann L, Beu M, Pascual-Leone A, Muller-Gartner HW (2000) Modulation of the neuronal circuitry subserving working memory in healthy human subjects by repetitive transcranial magnetic stimulation. Neuroscience Letters 280:167-170.
Mull BR, Seyal M (2001) Transcranial magnetic stimulation of left prefrontal cortex impairs working memory. Clin Neurophysiol 112:1672-1675.
Myles BS, Hagiwara T, Dunn W, Rinner L, Reese M, Huggins A, Becker S (2004) Sensory issues in children with Asperger syndrome and autism. Education and Training in Developmental Disabilities 39:283-290.
Nigg JT (2006) Temperament and developmental psychopathology. Journal of Child Psychology and Psychiatry 47:395-422.
Nikouline V, Ruohonen J, Ilmoniemi RJ (1999) The role of the coil click in TMS assessed with simultaneous EEG. Clinical Neurophysiology 110:1325-1328.
Papageorgiou C, Kontaxakis VP, Havaki-Kontaxaki BJ, Stamouli S, Vasios C, Asvestas P, Matsopoulos GK, Kontopantelis E, Rabavilas A, Uzunoglu N, Christodoulou GN (2001) Impaired P600 in neuroleptic naive patients with first-episode schizophrenia. Neuroreport 12:2801-2806.
Pascual-Leone A, Houser CM, Grafman J, Hallett M (1992a) Reaction time and transcranial magnetic stimulation. Lancet 339:1420.
Pascual-Leone A, Manoach DS, Birnbaum R, Goff DC (2002a) Motor cortical excitability in schizophrenia. Biological Psychiatry 52:24-31.
Pascual-Leone A, Brasil-Neto JP, Valls-Sole J, Cohen LG, Hallett M (1992b) Simple reaction time to focal transcranial magnetic stimulation. Comparison with reaction time to acoustic, visual and somatosensory stimuli. Brain 115 Pt 1:109-122.
Pascual-Leone A, Tormos JM, Keenan J, Tarazona F, Canete C, Catala MD (1998b) Study and modulation of human cortical excitability with transcranial magnetic stimulation. Journal of Clinical Neurophysiology 15:333-343.
Paus T (2001a) Primate anterior cingulate cortex: Where motor control, drive and cognition interface. Nature Reviews Neuroscience 2:417-424.
Paus T (2001b) Integration of transcranial magnetic stimulation and brain imaging. Biological Psychiatry 49:6S-6S.
Paus T (2005) Inferring causality in brain images: a perturbation approach. Philosophical Transactions of the Royal Society B-Biological Sciences 360:1109-1114.
Paus T, Castro-Alamancos MA, Petrides M (2001a) Cortico-cortical connectivity of the human mid-dorsolateral frontal cortex and its modulation by repetitive transcranial magnetic stimulation. Eur J Neurosci 14:1405-1411.
Paus T, Sipila PK, Strafella AP (2001b) Synchronization of neuronal activity in the human primary motor cortex by transcranial magnetic stimulation: an EEG study. J Neurophysiol 86:1983-1990.
Payne BR, Lomber SG (1999) A method to assess the functional impact of cerebral connections on target populations of neurons. J Neurosci Methods 86.
Peled A (1999b) Multiple constraint organization in the brain: A theory for schizophrenia. Brain Research Bulletin 49:245-250.
Peled A (2004) Brain dynamics and mental disorders, a project for a scientific psychiatry: Yozmot Heiliger.
Peled A, Geva AB (2000) The perception of Rorschach inkblots in schizophrenia: A neural network model. International Journal of Neuroscience 104:49-61.
127

Peled A, Pressman A, Geva AB, Modai I (2003) Somatosensory evoked potentials during a rubber-hand illusion in schizophrenia. Schizophrenia Research 64:157-163.
Peled A, Geva AB, Kremen WS, Blankfeld HM, Esfandiarfard R, Nordahl TE (2001) Functional connectivity and working memory in schizophrenia: An EEG study. International Journal of Neuroscience 106:47-61.
Praamstra P, Turgeon M, Hesse CW, Wing AM, Perryer L (2003) Neurophysiological correlates of error correction in sensorimotor-synchronization. Neuroimage 20:1283-1297.
Rao SM, Harrington DL, Haaland KY, Bobholz JA, Cox RW, Binder JR (1997) Distributed neural systems underlying the timing of movements. Journal of Neuroscience 17:5528-5535.
Repp BH (2001a) Processes underlying adaptation to tempo changes in sensorimotor synchronization. Hum Mov Sci 20:277-312.
Repp BH (2001b) Phase correction, phase resetting, and phase shifts after subliminal timing perturbations in sensorimotor synchronization. J Exp Psychol Hum Percept Perform 27:600-621.
Repp BH, Keller PE (2004) Adaptation to tempo changes in sensorimotor synchronization: Effects of intention, attention, and awareness. Quarterly Journal of Experimental Psychology Section a-Human Experimental Psychology 57:499-521.
Rippon G, Brock J, Brown C, Boucher J (2006) Disordered connectivity in the autistic brain: Challenges for the `new psychophysiology'. International Journal of Psychophysiology In Press, Corrected Proof.
Rodriguez E, George N, Lachaux JP, Martinerie J, Renault B, Varela FJ (1999) Perception's shadow: long-distance synchronization of human brain activity. Nature 397:430-433.
Rotem A, Moses E (2006) Magnetic stimulation of curved nerves. IEEE Trans Biomed Eng 53:414-420.
Rothbart MK, Ellis LK, Rueda MR, Posner MI (2003) Developing mechanisms of temperamental effortful control. Journal of Personality 71:1113-1143.
Rubenstein JLR, Merzenich MM (2003) Model of autism: increased ratio of excitation/inhibition in key neural systems. Genes Brain and Behavior 2:255-267.
Ruohonen J, Ilmoniemi RJ (1999) Modeling of the stimulating field generation in TMS. Electroencephalogr Clin Neurophysiol Suppl 51:30-40.
Sack AT, Hubl D, Prvulovic D, Formisano E, Jandl M, Zanella FE, Maurer K, Goebel R, Dierks T, Linden DE (2002) The experimental combination of rTMS and fMRI reveals the functional relevance of parietal cortex for visuospatial functions. Brain Res Cogn Brain Res 13:85-93.
Sanegre MT, Castellote JM, Haggard P, Valls-Sole J (2004) The effects of a startle on awareness of action. Experimental Brain Research 155:527-531.
Sawaki L, Okita T, Fujiwara M, Mizuno K (1999) Specific and non-specific effects of transcranial magnetic stimulation on simple and go/no-go reaction time. Experimental Brain Research 127:402-408.
Schaaf RC, Miller LJ (2005) Occupational therapy using a sensory integrative approach for children with developmental disabilities. Mental Retardation and Developmental Disabilities Research Reviews 11:143-148.
Seeman P, Weinshenker D, Quirion R, Srivastava LK, Bhardwaj SK, Grandy DK, Premont RT, Sotnikova TD, Boksa P, El-Ghundi M, O'Dowd BF, George SR, Perreault ML, Männistö PT, Robinson S, Palmiter RD, and Tallerico T (2005) Dopamine sensitivity correlates with D2high states, implying many paths to psychosis. Natl Acad Sci U S A. 2005 March 1; 102(9): 3513–3518.
128

Semjen A, Summers JJ (2002) Timing goals in bimanual coordination. Quarterly Journal of Experimental Psychology Section a-Human Experimental Psychology 55:155-171.
Semjen A, Schulze HH, Vorberg D (2000) Timing precision in continuation and synchronization tapping. Psychological Research-Psychologische Forschung 63:137-147.
Shaw JC, Colter N, Resek G (1983) EEG coherence, lateral preference and schizophrenia. Psychol Med 13:299-306.
Shima K, Mushiake H, Saito N, Tanji J (1996) Role for cells in the presupplementary motor area in updating motor plans. Proc Natl Acad Sci U S A 93:8694-8698.
Singer W (1999) Neuronal synchrony: A versatile code for the definition of relations? Neuron 24:49-65.
Smith SA, Press B, Koenig KP, Kinnealey M (2005) Effects of sensory integration intervention on self-stimulating and self-injurious behaviors. American Journal of Occupational Therapy 59:418-425.
Spencer KM, Nestor PG, Niznikiewicz MA, Salisbury DF, Shenton ME, McCarley RW (2003a) Abnormal neural synchrony in schizophrenia. J Neurosci 23.
Spencer KM, Nestor PG, Niznikiewicz MA, Salisbury DF, Shenton ME, McCarley RW (2003b) Abnormal neural synchrony in schizophrenia. Journal of Neuroscience 23:7407-7411.
Spencer KM, Nestor PG, Perlmutter R, Niznikiewicz MA, Klump MC, Frumin M, Shenton ME, McCarley RW (2004) Neural synchrony indexes disordered perception and cognition in schizophrenia. Proc Natl Acad Sci U S A 101.
Spencer RMC, Ivry RB, Zelaznik HN (2005) Role of the cerebellum in movements: control of timing or movement transitions? Experimental Brain Research 161:383-396.
Spencer RMC, Zelaznik HN, Diedrichsen J, Ivry RB (2003c) Disrupted timing of discontinuous but not continuous movements by cerebellar lesions. Science 300:1437-1439.
Stam CJ (2005) Nonlinear dynamical analysis of EEG and MEG: Review of an emerging field. Clinical Neurophysiology 116:2266-2301.
Stephan KE, Baldeweg T, Friston KJ (2006) Synaptic plasticity and dysconnection in schizophrenia. Biol Psychiatry 59.
Stephan KM, Thaut MH, Wunderlich G, Schicks W, Tian B, Tellmann L, Schmitz T, Herzog H, McIntosh GC, Seitz RJ, Homberg V (2002) Conscious and subconscious sensorimotor synchronization--prefrontal cortex and the influence of awareness. Neuroimage 15.
Sternad D (2000) Debates in dynamics: A dynamical systems perspective on action and perception. Human Movement Science 19:407-423.
Sternad D, Corcos D (2001) Effect of task and instruction on patterns of muscle activation: Wachholder and beyond. Motor Control 5:307-336.
Stevens LT (1886) On The Time Sense. Mind 11:393-404. Strens LHA, Oliviero A, Bloem BR, Gerschlager W, Rothwell JC, Brown P (2002) The
effects of subthreshold 1 Hz repetitive TMS on cortico- cortical and interhemispheric coherence. Clinical Neurophysiology 113:1279-1285.
Strick PL, Hoover JE, Mushiake H (1993) Evidence for "output channels" in the basal ganglia and cerebellum. In: Role of the cerebellum and basal ganglia in voluntary movement (Mano N, Hamada I, DeLong MR, eds), pp 171-180. Amsterdam: Elsevier Science.
Tallon-Baudry C, Bertrand O, Fischer C (2001) Oscillatory synchrony between human extrastriate areas during visual short-term memory maintenance. Journal of Neuroscience 21:art. no.-RC177.
129

Tandon R, Fleischhacker W (2005) Comparative efficacy of antipsychotics in the treatment of schizophrenia: A critical assessment. Schizophrenia Research 79:145-155.
Taylor JG (2003) Paying attention to consciousness. Prog Neurobiol 71:305-335. Taylor JL, McCloskey DI (1990) Triggering of preprogrammed movements as reactions to
masked stimuli. J Neurophysiol 63:439-446. Theoret H, Haque J, Pascual-Leone A (2001) Increased variability of paced finger tapping
accuracy following repetitive magnetic stimulation of the cerebellum in humans. Neuroscience Letters 306:29-32.
Thompson E, Varela FJ (2001) Radical embodiment: neural dynamics and consciousness. Trends in Cognitive Sciences 4:418-425.
Thut G, Ives JR, Kampmann F, Pastor MA, Pascual-Leone A (2005) A new device and protocol for combining TMS and online recordings of EEG and evoked potentials. J Neurosci Methods. 141(2): 207-17.
Tofts PS (1990) The distribution of induced currents in magnetic stimulation of the nervous system. Phys Med Biol 35:1119-1128.
Tononi G, Edelman GM (1998) Consciousness and the integration of information in the brain. Adv Neurol 77:245-279; discussion 279-280.
Tononi G, Edelman GM (2000) Schizophrenia and the mechanisms of conscious integration. Brain Res Brain Res Rev 31:391-400.
Tononi G, Sporns O, Edelman GM (1994) A Measure for Brain Complexity - Relating Functional Segregation and Integration in the Nervous-System. Proceedings of the National Academy of Sciences of the United States of America 91:5033-5037.
Torres IJ, O'Leary DS, Andreasen NC (2004) Symptoms and interference from memory in schizophrenia: evaluation of Frith's model of willed action. Schizophr Res 69.
Troyer AK, Winocur G, Craik FIM, Moscovitch M (1999) Source memory and divided attention: Reciprocal costs to primary and secondary tasks. Neuropsychology 13:467-474.
Tsuji T, Rothwell JC (2002) Long lasting effects of rTMS and associated peripheral sensory input on MEPs, SEPs and transcortical reflex excitability in humans. J Physiol 540:367-376.
Valls-Sole J, Rothwell JC, Goulart F, Cossu G, Munoz E (1999) Patterned ballistic movements triggered by a startle in healthy humans. Journal of Physiology-London 516:931-938.
Vanduffel W, Payne BR, Lomber SG, Orban GA (1997) Functional impact of cerebral connections. Proc Natl Acad Sci U S A 94.
Varela F, Thompson E, Rosch E (1993) The Embodied Mind: Cognitive Science and Human Experience: MIT Press.
Varela F, Lachaux JP, Rodriguez E, Martinerie J (2001a) The brainweb: phase synchronization and large-scale integration. Nat Rev Neurosci 2:229-239.
Varela F, Lachaux JP, Rodriguez E, Martinerie J (2001b) The brainweb: Phase synchronization and large-scale integration. Nature Reviews Neuroscience 2:229-239.
Verstynen T, Konkle T, Ivry RB (2006) Two types of TMS-induced movement variability after stimulation of the primary motor cortex. Journal of Neurophysiology 96:1018-1029.
Virtanen J, Ruohonen J, Naatanen R, Illmoniemi RJ (1999) Instrumentation for the measurement of electric brain responses to transcranial magnetic stimulation. Med Biol Eng Comput. 37: 322-326.
Vorberg D, Wing A (1996) Modeling variabilty and dependence in timing. In: Handbook of perception and action (Heuer H, Keele SW, eds), pp 181-262: London: Academic Press.
130

Vrtunski PB, Simpson DM, Meltzer HY (1989) Voluntary Movement Dysfunction in Schizophrenics. Biological Psychiatry 25:529-539.
Wassermann EM (1998) Risk and safety of repetitive transcranial magnetic stimulation: report and suggested guidelines from the international workshop on the safety of repetitive transcranial magnetic stimulation, June 5-7, 1996. Evoked Potentials-Electroencephalography and Clinical Neurophysiology 108:1-16.
Weissman DH, Roberts KC, Visscher KM, Woldorff MG (2006) The neural bases of momentary lapses in attention. Nat Neurosci 9.
Wieder S (1996) Integrated treatment approaches for young children with multisystem developmental disorder. Infants and Young Children 8:24-34.
Wing AM, Kristofferson AB (1973) Response delays and the timing of discrete motor responses. Perception and Psychophysics 14:5-12.
Winhusen S (2004) Emily Dickinson and Schizotypy. The Emily Dickinson Journal 13:77-96.
Wolff AL, O'Driscoll GA (1999) Motor deficits and schizophrenia: the evidence from neuroleptic-naive patients and populations at risk. Journal of Psychiatry & Neuroscience 24:304-314.
Yang YK, Chiu NT, Chen CC, Chen M, Yeh TL, Lee IH (2003) Correlation between fine motor activity and striatal dopamine D-2 receptor density in patients with schizophrenia and healthy controls. Psychiatry Research-Neuroimaging 123:191-197.
Yang YK, Yeh TL, Chiu NT, Lee IH, Chen PS, Lee LC, Jefffies KJ (2004) Association between cognitive performance and striatal dopamine binding is higher in timing and motor tasks in patients with schizophrenia. Psychiatry Research-Neuroimaging 131:209-216.
Yoshino K, Takagi K, Nomura T, Sato S, Tonoike M (2002) MEG responses during rhythmic finger tapping in humans to phasic stimulation and their interpretation based on neural mechanisms. Biological Cybernetics 86:483-496.
Zanone PG, Monno A, Temprado JJ, Laurent M (2001) Shared dynamics of attentional cost and pattern stability. Hum Mov Sci 20.
131

Independent Collaboration
The idea for the experiments, the design of the experimental structure, procedures and protocols and the design and construction of the fingertapping and EEG setup were done by Nava Levit-Binnun. The recruitment of subjects and the connection to Sha'ar Menashe hospital, which was essential for the ethical approval and medical supervision of the experiments, was done by Nava Levit-Binnun. Experiments were conducted by Nava Levit-Binnun with the assistance of Dr. Nestor Handzy and Dr. Avi Peled (who provided the medical supervision). Supplementary control experiments, in response to the request of referees while reviewing our manuscript were conducted by Dr. Nestor Handzy. Data analysis of the finger tapping experiments was conducted by Nava Levit-Binnun and Nestor Handzy with the technical assistance of Assaf Weiss. Data analysis of the EEG experiment was conducted by Dr. Vladimir Litvak using analysis tools developed by him in his PhD thesis.
132


לגלוש TMSעל כך שייתכן שהמניפולציות שביצענו שינו את יציבות הרשת וגרמו לערעורי ה .גם למרכיבים אחרים
ל להגברה של השפעת עירורי ה "גם אצל נבדקים עם סכיזופרניה גרמו המניפולציות הנ
TMSההשפעה המעניינת ביותר הייתה על העצירות התקופתיות . על מרכיבי הרשת שוניםהוגברו , שקודם לכן הופיעו אצל נבדקים עם סכיזופרניה, התקופתיותעצירות ה. בעשל האצ
גרמה L-DLPFCאינהיביציה של , .בצורה משמעותית כאשר הקשב הוסט למשימת החישובונראה , TMSשיפור משמעותי בביצועים של האצבע כאשר תופפה יחד עם עירעורי ה ל
לפונקציות ביצוע DLPFC בעבר יוחס ה .התקופתיותירות שגרמה לצמצום תופעת העצ. ופגיעה בפעילותו תוארה כאפיון ספציפי של תהליך המחלה בסכיזופרניה, גבוהות במוח
והשפיעה , העובדה שהשתקתו של אזור זה שיפרה את התפקוד אצל נבדקים עם סכיזופרניה הינו L-DLPFCמצביעה על כך שאכן ה, תזמון וקשב, על מנגנונים אפשריים של תיקון טעות
החלשת ה . ושהוא אינו תקין בסכיזופרניה, חוליה בתוך הרשת השולטת על משימת התיפוף– L-DLPFCמקטין את השפעת אי תפקודו, באמצעות פעולת האינהיביציה.
על מנת למדוד את תגובת הפעילות ) EEG) ElectroEncephaloGramכאשר השתמשנו ב מצאנו הבדלים , דקים בריאים ונבדקים עם סכיזופרניה אצל נבTMSהגלובלית במוח לעירור
הבדלים אלו עולים בקנה אחד עם . TMS ל EEGהן באמפליטודה והן בזמנים של תגובות ה (Massimini et al., 2005)תוצאות שהתקבלו לאחרונה באמצעות כלים מתוחכמים יותר
ככלי TMSור המתקבלות בעקבות עירEEGותומך בטענה שניתן להשתמש בתגובות .למדידת שינויים בתגובתיות ובקישוריות במוח
תומכות בהיפותזת הקישוריות הלא תקינה בסכיזופרניה שהתקבלו בשני הניסויים תוצאות ה
כיבים שונים של רשת קוגניטיבית נרחבת באמצעות רומדגימות את היכולת לתמרן ולחקור מTMS .
להבנת הוויסות והיציבות של האינטראקציות התוצאות נידונות במונחים של התרומה שלהם
של רשתות במוח וכיצד שינויים בהם יכולים להסביר את הספקטרום הנרחב של התנהגויות . אנושיות
133

134
תמצית
סכיזופרניה היא מחלה הרסנית הפוגעת באחוז אחד מאוכלוסיית העולם בתחילת חייהם תיאוריות מובילות . שפחה הקרובהומהווה נטל כלכלי על החברה וכאב רב למ, הבוגרים
הן של יתר קישוריות והן של תת -בחקר המחלה מצביעות על בעיות בקישוריות במוח שבירה של "סכיזופרניה לעיתים מתוארת כ. העומדות בבסיס מחלה זאת-קישוריות, לכן. בהיותה משפיעה כמעט בכל תחום הקשור בחוויה המורכבת של להיות אדם, "התודעה" נורמלית" הפתולוגיה של הסכיזופרניה עשויה לאפשר תובנה כיצד החוויה האנושית ההבנת
.מתהווה מהארכיטקטורה הביולוגית של המוח
המטרה העיקרית של מחקר זה הייתה להשתמש בעירורים הנוצרים מסטימולציה מגנטית על מנת לחקור ) (TMS – Transcranial Magnetic Stimulation(טראנס גולגולתית
הן אצל אנשים בריאים והן אצל , קישוריות ויציבות של רשתות קוגנטיביות נרחבות במוח .אנשים עם סכיזופרניה
TMS הינו שיטת עירור לא חודרנית שבה פולס של שדה מגנטי חזק הניתן על הקרקפת מאפשר השדה המגנטי דועך במהירות עם המרחק ולכן. משרה זרמים חשמליים בתוך המוח
.עירור יעיל רק באיזורים הקרובים יחסית לפני השטח של הקורטקס
TMSככלי לעורר את , ניתן בעת משימה של תיפוף האצבע לפי תזמון של מטרונום חיצונילמרות שמשימה זאת הינה פשוטה . התנודות הפנימיות שנוצרות בעקבות משימה זו במוח
, סנסומוטוריים, של איזורים פרונטליםהיא מפעילה רשת נוירונלית נרחבת, יחסיתמודעות , האחראים לפונקציות מוח כגון הנעה פנימית, צברלאריים ואיזורי מוח עמוקים
היו TMSערעורי ה. משוב חושי וניטור טעויות, תכנון וביצוע מוטורי, תזמון, קשב, לתנועה ).M1(י מסונכרנים למטרונום וניתנו לרשת זו ברמת האזור המוטורי הראשונ
בשלב ראשון הצלחנו להראות הן אצל נבדקים בריאים והן אצל נבדקים עם סכיזופרניה
השפיעו TMSערעורי . מאפשר הפרדה של מרכיבים שונים ברשת הנוירונליתTMSשעירור בצורה ניכרת על תנועת האצבע וגרמו לשתי צורות התנהגות לא רגילות אליהן הנבדקים לא
למרבה . והשהיה של האצבע למשך חצי זמן מחזור, ה של תדירות התיפוףהכפל: היו מודעיםדבר המצביע על כך שתהליכי התזמון , ההפתעה התזמון המדויק של התיפוף לא הושפע
המעורבים בבקרה של תנועת התיפוף הינם נפרדים מהתהליכים המוטוריים האחראים .לביצוע הפקודות המוטוריות
למרכיבים אחרים של הרשת ופגעו ביכולת TMSלשו עירורי ה בנבדקים עם סכיזופרניה גבנוסף חשפו העירורים קשיי תפקוד גבוהים הקשורים לשימת לב . להעריך את דיוק התזמון
עצירות תקופתיות ). Lapses(ולהנעה פנימית בצורת עצירות תקופתיות של תנועת האצבע צבע למשך זמן מחזור של שהוגדרו כעצירה מוחלטת או כמעט מוחלטת של הא, אלו
.TMSנצפו בזמנים שבו ניתנו עירורי ה , המטרונום
מתוארים לעיל תוך כדי מניפולציה של מרכיבים TMSביצענו את עירורי ה , בהמשךהקשב הוסט ממשימת , בתנאי הראשון. משוערים של הרשת הנוירונאלית שתוארה לעיל
גרמנו לאינהיביציה , תנאי השניב. התיפוף אל משימה מקבילה של חישוב אריתמטי L-DLPFC: left(של הקורטקס השמאלי הפרה פרונטלי דורסולטרלי ) השתקה(
dorsolateral prefrontal cortex ( על ידי עירור חוזר שלTMS . צורת עירור זו ניתנה לפניתחילת משימת התיפוף באופן שההשפעה האינהיביטורית מתאפשרת למשך של כל זמן
.הניסוי
השפיעו כעת ,1M ברמת TMSל לכך שעירורי ה "אצל נבדקים בריאים גרמו המניפולציות הנ תוצאה זאת מצביעה. לא רק על המרכיב המוטורי אלא גם על התזמון ועל המודעות לאצבע