thesis - tiho hannover...silvina ribeiro samy, nuno sousa, antonio j. salgado, andreas ratzka,...
TRANSCRIPT

University of Veterinary Medicine Hannover
Hannover Medical School Institute of Neuroanatomy
Peripheral nerve regeneration using hollow and enriched chitosan-based guidance conduits
Thesis
submitted in partial fulfillment of the requirements for the degree of
-Doctor rerum naturalium-
(Dr. rer. nat.)
by
Cora Meyer born in Bremen
Hannover, Germany 2014

Supervisor: Prof. Dr. C. Grothe Supervision group: Prof. Dr. C. Grothe
Prof. Dr. R. Gerardy-Schahn Prof. Dr. K. Schwabe
1st Evaluation: Prof. Dr. C. Grothe Hannover Medical School
Institute of Neuroanatomy Carl-Neuberg-Straße 1 30625 Hannover Prof. Dr. R. Gerardy-Schahn
Hannover Medical School Institute for Cellular Chemistry Carl-Neuberg-Straße 1 30625 Hannover
Prof. Dr. K. Schwabe Hannover Medical School
Department of Neurosurgery Carl-Neuberg-Straße 1 30625 Hannover
2nd Evaluation: Esther Udina i Bonet, MH, PhD Universitat Autònoma Barcelona Department of Cellular Biology, Physiology and Immunology Campus UAB 08193 Bellaterra (Cerdayola del Vallès)
Day of final exam: 10.10.2014

Parts of the thesis have been published previously:
Kirsten Haastert-Talini, Stefano Geuna, Lars Dahlin, Cora Meyer, Lena Stenberg, Thomas Freier, Claudia Heimann, Christina Barwig, Luis F.V. Pinto, Stafania Raimondo, Giovanna Gambarotta, Silvina Ribeiro Samy, Nuno Sousa, Antonio J. Salgado, Andreas Ratzka, Sandra Wrobel, Claudia Grothe (2013).
“Chitosan tubes of varying degrees of acetylation for bridging peripheral nerve defects”. Biomaterials, 2013, 34, 9886-9904.
Parts of this thesis have been submitted previously:
Cora Meyer*, Sandra Wrobel*, Stefania Raimondo, Claudia Heimann, Abraham Shahar, Stefano Geuna, Claudia Grothe, Kirsten Haastert-Talini.
“Engineered Schwann cells delivering fibroblast growth factor-2 increase peripheral nerve regeneration through hydrogel enriched chitosan tubes”. [submitted July 2014] * Both authors contributed equally to the work and share first authorship.
Poster presentations:
Cora Meyer, Kirsten Haastert-Talini, Sandra Worbel, Andreas Ratzka, and Claudia Grothe. “Inflammatory and physiological tissue responses following nerve reconstruction with chitosan tubes of different degrees of acetylation”. 2nd International Symposium on Peripheral Nerve Regeneration, Turin, Italy, January 23-25, 2014. Cora Meyer, Kirsten Haastert-Talini, Sandra Wrobel, and Claudia Grothe. “Addressing the long gap: Chitosan nerve guides supplemented with genetically modified Schwann cells”. 5th Lübeck Regenerative Medicine Symposium, Lübeck, Germany, June 26-27, 2014.
Sponsorship:
This work has received funding from the European Community’s Seventh Framework Programme (FP7-HEALTH-2011) under grant agreement n° 278612 (BIOHYBRID).


Dedicated to my parents


Table of contents
I
Table of Contents Abbreviations ............................................................................................................... IV List of figures .............................................................................................................. VII List of tables ................................................................................................................ IX Summary ..................................................................................................................... XI Zusammenfassung ..................................................................................................... XV 1 Introduction ............................................................................................................ - 1 -
1.1 Clinical relevance of peripheral nerve injuries (PNIs) .................................... - 1 - 1.2 Organization of the peripheral nervous system (PNS) ................................... - 2 - 1.3 Classification of peripheral nerve injuries (PNIs) ........................................... - 4 - 1.4 Degeneration and regeneration processes following injury ........................... - 5 -
1.4.1 Neuronal response ................................................................................... - 6 - 1.4.2 Changes in the distal environment and axonal regeneration .................. - 7 -
1.5 Surgical repair strategies and their limitations ............................................... - 9 - 1.5.1 End-to-end repair ..................................................................................... - 9 - 1.5.2 Autologous nerve graft (ANG) ............................................................... - 10 - 1.5.3 Allograft .................................................................................................. - 10 - 1.5.4 CE and FDA Approved Conduits ........................................................... - 11 -
1.5.4.1. Non-resorbable conduits ................................................................ - 13 - 1.5.4.2 Type I collagen ................................................................................ - 13 - 1.5.4.3 Porcine small intestinal submucosa (SIS) ....................................... - 13 - 1.5.4.4 Polyglycolic acid (PGA) ................................................................... - 14 - 1.5.4.5 Poly D,L lactide-co-ɛ-caprolactone (PCL) ....................................... - 14 -
1.6 Considerations to design the future ideal conduit ........................................ - 14 - 1.6.1 Biocompatible and biodegradable polymers .......................................... - 15 -
1.6.1.1 Chitosan .......................................................................................... - 15 - 1.6.2 Luminal fillers ......................................................................................... - 17 -
1.6.2.1 NVR-Gel-A hyaluronic and laminin-based 3D guidance structure .. - 18 - 1.6.3 Incorporation of support cells ................................................................. - 19 -
1.6.3.1 Schwann Cells (SCs) ...................................................................... - 19 - 1.6.4 Incorporation of neurotrophic factors (NTFs) ......................................... - 20 -
1.6.4.1 Glial cell-derived neurotrophic factor (GDNF) ................................. - 20 - 1.6.4.2 Fibroblast growth factor (FGF)-2 ..................................................... - 21 -
1.7 Tissue responses following implantation of medical devices ....................... - 22 - 1.8 The sciatic nerve model ................................................................................ - 24 - 1.9 Aim of the study ............................................................................................ - 26 -
2. Material ............................................................................................................... - 29 - 3. Methods .............................................................................................................. - 33 -
3.1 Animals ......................................................................................................... - 33 - 3.2 Experimental design and groups .................................................................. - 33 -
3.2.1-a Project 1-Short-term study .................................................................. - 33 - 3.2.1-b Project 1-Long-term study .................................................................. - 34 - 3.2.2 Project 2 ................................................................................................. - 35 -
3.4 Anesthesia and surgical procedures ............................................................ - 36 - 3.5. Functional evaluation of nerve regeneration. .............................................. - 38 -
3.5.1 Static Sciatic Index (SSI) ....................................................................... - 38 - 3.5.2 Von Frey test .......................................................................................... - 39 - 3.5.3 Electrophysiology ................................................................................... - 40 -

Table of contents
II
3.5.3.1 Serial non-invasive electrodiagnostic recordings ............................ - 40 - 3.5.3.2 Final invasive recordings ................................................................. - 42 -
3.5.4 Muscle weight ........................................................................................ - 43 - 3.5.5 Autotomy score ...................................................................................... - 43 -
3.6 (Immuno-) histological evaluation of nerve regeneration ............................. - 44 - 3.6.1 Assessment of nerve tissue ................................................................... - 44 -
3.6.1.1 Project 1-short-term study ............................................................... - 44 - 3.6.1.2 Project 1-long-term study ................................................................ - 46 - 3.6.1.3 Project 2 .......................................................................................... - 47 -
3.6.2 Assessment of fibrous capsule .............................................................. - 48 - 3.6.2.1 Project 1 .......................................................................................... - 48 - 3.6.2.2 Project 2 .......................................................................................... - 50 -
3.7 Molecular and biochemical evaluation of nerve regeneration ...................... - 51 - 3.7.1 Quantitative real-time polymerase chain reaction (qRT-PCR) .............. - 51 - 3.7.2 Western Blot........................................................................................... - 52 -
3.8 Analysis of tube properties after explantation .............................................. - 52 - 3.9 Statistical analysis ........................................................................................ - 53 -
4 Results-Project 1 ................................................................................................. - 55 - 4.1 Gene expression of regeneration-associated proteins ................................ - 55 - 4.2 Matrix formation and presence of brain-derived neurotrophic factor (BDNF)- 56 - 4.2 Static Sciatic Index (SSI) .............................................................................. - 57 - 4.3 Electrophysiological assessment of muscle reinnervation ........................... - 59 - 4.4 Calculation of the lower limb muscle weight-ratio ........................................ - 67 - 4.6 Conduit properties upon explantation and tissue regeneration .................... - 67 - 4.5 Inflammatory tissue response ...................................................................... - 71 -
4.5.1 Connective tissue ................................................................................... - 71 - 4.5.2 Nerve tissue ........................................................................................... - 77 -
5 Results-Project 2 ................................................................................................. - 83 - 5.2 Mechanical pain threshold assessment ....................................................... - 83 - 5.1 Static Sciatic Index (SSI) .............................................................................. - 86 - 5.3 Electrophysiological assessment of muscle reinnervation ........................... - 87 - 5.4 Calculation of the lower limb muscle weight-ratio ........................................ - 95 - 5.3 Autotomy score ............................................................................................. - 95 - 5.5 Macroscopic analysis of the explanted nerve tissue .................................... - 96 - 5.6 Inflammatory tissue response ...................................................................... - 97 -
5.6.1 Connective tissue ................................................................................... - 97 - 5.6.2 Regenerated tissue .............................................................................. - 101 -
6 Discussion ......................................................................................................... - 107 - 6.1 Project 1 ...................................................................................................... - 108 -
6.1.1 Regulation of regeneration associated proteins following nerve lesion and repair ............................................................................................................. - 108 - 6.1.2 Matrix formation and brain-derived neurotrophic factor (BDNF) presence in the tube content ............................................................................................ - 109 - 6.1.3 Reinnervation of distal muscle target................................................... - 111 -
6.1.3.1 Behavior ........................................................................................ - 111 - 6.1.3.2 Electrophysiological recordings ..................................................... - 113 - 6.1.3.3 Morphological evaluation .............................................................. - 115 - 6.1.3.4 Muscle weight ................................................................................ - 116 -
6.1.4 Foreign body response (FBR) upon chitosan implantation degradation- 117 -

Table of contents
III
6.1.5 Concluding remarks-Chances of hollow chitosan conduits on the existing market ........................................................................................................... - 121 -
6.2 Project 2 ...................................................................................................... - 124 - 6.2.1 Schwann cells (SCs) as vehicles for gene delivery ............................. - 124 - 6.2.2 Recovery of mechanical pain sensibility .............................................. - 125 - 6.2.3 Fibroblast growth factor (FGF)-218kDa partly allows reinnervation of distal muscle targets and functional recovery ........................................................ - 128 -
6.2.3.1 Behavior ........................................................................................ - 128 - 6.2.3.2 Electrophysiological recordings ..................................................... - 128 -
6.2.4 The difficulty in finding the right luminal filler for axonal guidance ...... - 131 - 6.2.5 No enhanced inflammatory response elicited by (modified) Schwann cells (SCs) in NVR Gel .......................................................................................... - 133 - 6.2.6 Concluding remarks and considerations regarding future conduits for long nerve defects ................................................................................................. - 133 -
7 References ........................................................................................................ - 136 - 8. Affidavit ............................................................................................................. - 150 - 9. Acknowledgements .......................................................................................... - 152 -

Abbreviations
IV
Abbreviations ADSC Adipose-derived stem cell
ANOVA Analysis of variance
ANG Autologous nerve graft
ANH Adjacent neuropathic hyperalgesia
AUC Area under the curve
AxL Axon loss
BDNF Brain-derived neurotrophic factor
BMSC Bone marrow-derived mesenchymal stem cell
CE Conformité Européenne
CMAP Compound muscle action potential
CNS Central nervous system
CNTF Cilliary neurotrophic factor
DA Degree of acetylation
DAPI 4‘, 6-diamidino-2-phenylindole
DRG Dorsal root ganglion
ECM Extracellular matrix
EG Europäische Gemeinschaft
EV Empty vector
FBGC Foreign body giant cell
FBR Foreign body response
FDA Food and Drug Administration
FGF Fibroblast growth factor
GAP-43 Growth-associated protein-43
Gapdh Glyceraldehyde-3-phosphate dehydrogenase
GDNF Glial cell-derived growth factor
GRAS Generally Recognized As Safe
HA Hyaluronic acid
HE Hematoxylin eosin
IHC Immunohistochemistry
IL Interleukin
i.m. Intramusculary

Abbreviations
V
i.p. Intraperitonealy
ITS Intermediate toe spread
ITSF Intermediate toe spread factor
L Lesion
MAG Myelin-associated glycoprotein
MGC Multinucleated giant cell
MHH Medizinische Hochschule Hannover
mRNA Messenger ribonucleic acid
N Non-lesion
NCAM Neural cell adhesion molecule
NCV Nerve conduction velocity
neo Neonatal
NF Neurofilament
NGF Nerve growth factor
NGS Normal goat serum
nt Non-transfected
NT Neurotrophin
NTF Neurotrophic factor
NVR Neural and vascular reconstruction
OEC Olfactory ensheating cell
P0 Myelin protein zero
PBS Phosphate buffered saline
PCL Poly(D,L lactide-co-ɛ-caprolactone)
PCLF Poly(caprolactone fumarat)
PFA Paraformaldehyde
PGA Polyglycolic acid
PL Plantar
PLGA Poly(lactic-co-glycolic acid)
PLLA Poly(L-lactic acid) PNI Peripheral nerve injury
PNS Peripheral nervous system
Ppia Peptidylpropyl isomerase
PSA Polysialic acid
PVA Polyvinyl alcohol

Abbreviations
VI
qRT-PCR Quantitative real-time polymerase chain reaction
R Receptor
RIPA Radioimmunoprecipitation
RT Room temperature
RT-PCR Real time polymerase chain reaction
s.c. Subcutaneous
SC Schwann cell
SD Sunderland degree
SEM Standard error of means
SFI Sciatic function index
SIS Small intestinal submucosa
SSI Static Sciatic Index
TA Tibialis anterior
UAB Autonomous University of Barcelona
ULUND Lund University
UMINHO University of Minho
UNITO University of Turin
tf Transfected
TrkB Tyrosine kinase receptor B
TS Toe spread
TSF Toe spread factor
US United States

List of figures
VII
List of figures Introduction Figure 1: Anatomical organization of the peripheral nervous system. ..................... - 3 - Figure 2: Degenerative and regenerative events following injuries to the mammalian PNS. ......................................................................................................................... - 7 - Figure 3: Regeneration processes across a 10 mm gap within a hollow nerve guidance conduit. ................................................................................................... - 12 - Figure 4: Chemical structure of chitin and chitosan. .............................................. - 16 - Figure 5: Formation of foreign body giant cells (FBGCs). ..................................... - 24 - Figure 6: The sciatic nerve and its innervation targets. ......................................... - 25 - Methods Figure 7: Experimental design for the long-term study (13 weeks) using hollow chitosan conduits in a 10 mm gap (project 1). ....................................................... - 35 - Figure 8: Experimental design for the long-term study (17 weeks) using enriched tubes in a 15 mm gap (project 2). .......................................................................... - 36 - Figure 9: Chitosan conduit implantation. ................................................................ - 38 - Figure 10: Static Sciatic Index (SSI) ...................................................................... - 39 - Figure 11: Von Frey ................................................................................................ - 40 - Figure 12: Serial non-invasive electrodiagnostic recordings. ................................ - 42 - Figure 13: Autotomy Score. ................................................................................... - 44 - Figure 14: Processing of the nerve tissue gathered 5 and 18 days after surgery (project 1). .............................................................................................................. - 45 - Figure 15: Processing of the regenerated nerve tissue 13 weeks after implantation (project 1). .............................................................................................................. - 46 - Figure 16: Illustration of fibrous capsule analysis. ................................................. - 50 - Figure 17: Regulation of mRNA levels in spinal cord. ........................................... - 55 - Figure 18: Endogenous brain-derived neurotrophic factor (BDNF) detection in tube content. ................................................................................................................... - 56 - Results-Project 1 Figure 19: Static Sciatic Index (SSI). ..................................................................... - 58 - Figure 20: Serial recordings of CMAPs. ................................................................. - 60 - Figure 21: Recovery of the CMAP amplitude. ........................................................ - 62 - Figure 22: Axon Loss. ............................................................................................ - 64 - Figure 23: NCV-ratio. ............................................................................................. - 65 - Figure 24: Analysis of the maximal CMAP. ............................................................ - 66 - Figure 25: Muscle weight determination. ............................................................... - 67 - Figure 26: Macroscopic properties of DAIII tubes. ................................................. - 68 - Figure 27: Regenerated nerve tissue upon explantation. ...................................... - 69 - Figure 28: Fibrous capsule optical density. ............................................................ - 71 - Figure 29: Fibrous capsule analysis. ..................................................................... - 72 - Figure 30: Presence of activated macrophages in the capsule surrounding the conduits. ................................................................................................................. - 73 - Figure 31: Collagen distribution within capsule.. ................................................... - 75 -

List of figures
VIII
Figure 32: Vascularization of the capsule surrounding the conduit. ...................... - 76 - Figure 33: Number of MGCs and presence of chitosan inclusions 18 days after tube implantation within the regenerated nerve tissue.. ................................................ - 78 - Figure 34: Presence of MGCs in the regenerated nerve tissue 13 weeks after surgery. ................................................................................................................... - 79 - Figure 35: Scattered chitosan inclusions in the nerve tissue of the DAIII group. .. - 80 - Figure 36: Activated macrophages in the proximal nerve stump after 18 days. .... - 81 - Results-Project 2 Figure 37: Mechanical pain threshold (lateral) ....................................................... - 84 - Figure 38: Mechanical pain threshold.(medial) ...................................................... - 85 - Figure 39: Static Sciatic Index (SSI). ..................................................................... - 87 - Figure 40: Serial recordings of CMAPs. ................................................................. - 89 - Figure 41: Amplitude-ratio from CMAPs recorded from the tibialis anterior (TA) muscle. ................................................................................................................... - 91 - Figure 42: Amplitude-ratio from CMAPs recorded from the plantar (PL) muscle. . - 92 - Figure 43: Muscle weight-ratio. .............................................................................. - 95 - Figure 44: Autotomy Score. ................................................................................... - 96 - Figure 45: Representative photographs of explanted specimens following a 4 month observation period. ................................................................................................. - 97 - Figure 46: Fibrous capsule analysis. ..................................................................... - 98 - Figure 47: Presence of activated macrophages in the capsule surrounding the conduits. ................................................................................................................. - 99 - Figure 48: Collagen distribution within capsule.................................................... - 100 - Figure 49: Vascularization of the capsule surrounding the conduit. .................... - 101 - Figure 50: Regeneration across the gap. ............................................................. - 103 - Figure 51: Failed regeneration. ............................................................................ - 104 - Figure 52: Cross-section of an enriched chitosan conduit and regenerated nerve tissue 17 weeks following implantation. ............................................................... - 105 -

List of tables
IX
List of tables Methods Table 1: Overview of the collected tissue samples in the short-term study of project 1. ................................................................................................................. - 34 - Results-Project 1 Table 2: Listing of animals per group, which demonstrated evocable CMAPs in the course of the study. ................................................................................................ - 59 - Table 3: Overview of animal numbers showing regenerated nerve tissue 13 weeks after chitosan conduit implantation. ....................................................................... - 70 - Results-Project 2 Table 4: Listing of animals per group, which demonstrated evocable compound muscle action potentials in the course of the study. .............................................. - 88 - Table 5: NCV, AxL, and amplitude-ratio ................................................................ - 94 - Table 6: Tissue regeneration seen upon explantation. .......................................... - 96 - Table 7: Quantity of MGCs and activated macrophages within the (regenerated) nerve tissue. ......................................................................................................... - 102 -

X

Summary
XI
Summary Peripheral nerve regeneration using hollow and enriched chitosan-based guidance conduits Cora Meyer Peripheral nerve injuries (PNIs) represent a global problem, which mainly affects
young men in their work productive years. Following crush injuries, patients will likely
experience a full functional recovery. If the nerve is completely transected, on the
other hand, surgical intervention is needed and the outcome is highly variable.
Cases, which do not allow tensionless end-to-end repair, require a bridging strategy
to overcome the gap. As a clinical standard therapy, surgeons in such situations
usually harvest a negligible sensory nerve and use it to reconnect the two nerve
stumps. This method, though, has disadvantages including the need of a second
surgery, limited amount of available tissue as well as mismatch issues between the
donor nerve and the affected nerve. In addition, only about 50 % of the patients
treated with autologous nerve grafts (ANGs) are reported to have a successful
recovery. Due to these reasons, alternative strategies are needed to bridge nerve
defects. Various synthetic as well as biologically-derived materials have already
been tested and some have been approved by the "Food and Drug Administration"
and "Conformité Européenne" to be marketed as nerve guidance conduits for small
gaps, but none of these conduits is completely accepted in the clinics.
Aim of the present study was to test hollow chitosan-based nerve guidance conduits
of 3 different degrees of acetylation (DAI-III) for their possible application in
treatment of small gaps (e.g. in digital nerves) (project 1). Therefore, short (5 days
and 18 days) and long-term (13 weeks) studies were conducted. In addition, bridging
of long gaps (e.g. in brachial plexus injuries) was addressed by introducing luminal
fillers into the hollow chitosan conduits, which should, amongst others, support
axonal elongation (project 2). Genetically modified neonatal Schwann cells (SCs)
over-expressing either glial cell-derived neurotrophic factor (GDNF) or the low
molecular weight isoform of fibroblast growth factor-2 (FGF-218kDa) were suspended
in a hyaluronic acid and laminin containing gel (NVR-Gel), which was then injected

Summary
XII
into the chitosan scaffold. Empty vector-transfected SCs as well as non-transfected
SCs served as controls besides the reversed ANG in this study of 17 weeks
duration. For both objectives, the nerve guidance conduits were tested in the rat
sciatic nerve model (10 mm or 15 mm gap), which is frequently used to evaluate
peripheral nerve reconstruction. This animal model offers the opportunity to assess
motor and sensory recovery by means of electrophysiological recordings, Static
Sciatic Index, and the von Frey test. Furthermore, the tissue response in presence of
the chitosan implants was analyzed to exclude the possibility of a strong
inflammatory response.
Project 1 led to very encouraging results. Hollow chitosan conduits of all DAs
allowed peripheral nerve regeneration in a similar manner as ANGs across a 10 mm
nerve defect. Motor recovery was observed in almost all animals and the analysis of
the inflammatory response revealed a clear reduction in the number of activated
macrophages and multinucleated giant cells (MGCs) in the fibrous capsule, which
had developed around the conduits over time. In addition, less macrophages were
found after 13 weeks than 18 days in the regenerated nerve tissue, while the number
of MGCs remained comparably equal and low. Noteworthy, no changes in the
regulation of selected regeneration-associated proteins was detected as a response
towards the conduits in dorsal root ganglia or the spinal cord 5 and 18 days after
conduit implantation. Morphometric data, however, revealed increased sprouting of
axons in the presence of DAI conduits, while DAIII conduits demonstrated a fast
degradation rate. Upon explantation, the latter were partly broken and displayed
fissures and cracks. As a consequence of these observations, DAII conduits were
chosen for further experiments, since these conduits were most promising to support
peripheral nerve regeneration.
For project 2, DAII conduits were enriched and tested in a critical 15 mm defect. At
the end of the study, the group that had received ANGs was superior compared to
the experimental groups that had received chitosan conduits containing enriched gel.
Solely, the FGF-218kDa group showed nerve tissue regeneration in 4 out of 7 animals,
which also displayed motor recovery; while it was only one animal from the GDNF
and non-transfected SCs group, respectively. These results indicate that the
introduction of gel as a luminal filler might not be the best choice, since it possibly

Summary
XIII
acts as a physical impediment to peripheral nerve regeneration and only FGF-218kDa
seems to partially compensate for this negative effect.
Reflecting the findings of both studies it can be concluded that DAII chitosan-based
conduits represent an alternative to the marketed conduits for treatment of small
gaps. Besides the promising results obtained in this study, it is especially appealing
for clinical use because of its transparency. The latter property facilitates the
handling procedure for the surgeon and reduces the time needed for correct
positioning of the implant. Regarding treatment of long gaps, however, the search for
an optimal luminal filler / guidance structure continuous and further experiments are
indispensable. Alternatives, which guide the regrowing axons without filling the whole
luminal space, such as highly porous sponges, membranes, and fibers, are probably
better suited for this task than gel.

XIV

Zusammenfassung
XV
Zusammenfassung
Periphere Nervenregeneration mit leeren und angereicherten Chitosan-basierten Leitschienen Cora Meyer Periphere Nervenverletzungen stellen ein globales Problem dar, von welchem
insbesondere junge Männer in ihren arbeitsfähigen Jahren betroffen sind. Im Zuge
von Quetschungen ist eine Wiederherstellung der Funktionalität wahrscheinlich,
während die Heilungschancen nach kompletten Nervendurchtrennungen variieren
und ein chirurgischer Eingriff erforderlich ist. Falls ein spannungsfreies Verbinden
der beiden Nervenstümpfe nicht möglich ist, ist der Einsatz von Nervenleitschienen
unerlässlich. In der Klinik wird in diesen Fällen in der Regel ein autologes
Nerventransplantat verwendet, welches von einem funktionell entbehrlichen
sensiblen Nerven gewonnen wird. Diese Methode hat jedoch Nachteile, da eine
zweite Operation nötig ist, das potenziell zu entnehmende Nervengewebe begrenzt
ist und es zu Inkompatibilitäten zwischen dem Spender- und Empfängernerv
kommen kann. Des Weiteren wird lediglich in der Hälfte der Fälle, welche mit einem
Autotransplantat behandelt werden, eine erfolgreiche Regeneration beobachtet.
Folglich werden alternative Überbrückungsstrategien für Nervenlücken benötigt.
Hierfür wurden bereits verschiedene Synthetik- als auch Biomaterial-basierte
Nervenleitschienen getestet und zum Teil auch von der „Food and Drug
Administration“ (FDA) sowie „Conformité Européenne“, zu Deutsch „Europäische
Gemeinschaft“, zur Behandlung am Menschen zugelassen. Allerdings hat bisher
keine dieser Nervenleitschienen eine breite Akzeptanz in der klinischen Anwendung
gefunden.
Folglich war es ein Ziel dieser Arbeit leere Chitosan-basierte Nervenleitschienen
verschiedener Acetylierungsgrade (DAI-III) für die Überbrückung von kleinen
Nervendefekten, wie sie z.B. bei Handnerven häufig vorkommen, zu testen. Zu
diesem Zwecke wurden Kurzzeit- und Langzeitstudien durchgeführt über Zeiträume
von 5 und 18 Tagen, sowie 13 Wochen (Projekt 1). Des Weiteren wurden Chitosan
Röhrchen mit einem Hyaluronsäure- und Laminin-basiertem Gel (NVR-Gel) befüllt, in

Zusammenfassung
XVI
welchem genetisch veränderte neonatale Schwann Zellen suspendiert waren
(Projekt 2). Letztere wurden dahingehend modifiziert, dass sie entweder eine
erhöhte Expression von GDNF (glial cell-derived neurotrophic factor) oder der 18kDa
FGF (fibroblast growth factor)-2-Isoform aufwiesen. Neben dem reversen autologen
Nerveninterponats wurden physiologische sowie mit einem Leervektor-transfizierte
Schwann Zellen (in Gel) als Kontrollen transplantiert. Mit der Ergänzung von
intraluminalen Leitstrukturen und Schwann Zellen sollte die Behandlung von
größeren Nervendefekten, wie sie z.B. für den Plexus brachialis beschrieben sind,
angesprochen werden, da diese die Regeneration potenziell unterstützen können.
Für beide Projekte wurde das Modell des Nervus ischiadicus bei der Ratte (10 mm
oder 15 mm Lücke) gewählt, welches als Standard-Modell zur Untersuchung
peripherer Nervenregeneration gilt. Es bietet die Möglichkeit, die motorische als
auch sensorische Regeneration u.a. durch elektrophysiologische Messungen, die
Feststellung des SSIs (Static Sciatic Index) als auch durch den von Frey Test zu
beurteilen. Im Rahmen dieser Arbeit wurde ferner die Gewebereaktion auf das
Implantieren der Chitosan-basierten Nervenleitschienen untersucht, um eine starke
Abwehrreaktion auszuschließen.
Die Ergebnisse, die im Projekt 1 erzeugt wurden waren vielversprechend und eine
erfolgreiche, mit dem Autotransplantat vergleichbare, Nervenregeneration konnte für
Chitosan Röhrchen aller Acetylierungsgrade, bei einer Überbrückungsdistanz von
10 mm, festgestellt werden. Bei nahezu allen Tieren konnte die Reinnervation
distaler Muskeln nachgewiesen werden und die Immunantwort zeigte eine deutliche
Reduktion im Zeitverlauf. Letzteres äußerte sich in einer Abnahme aktivierter
Makrophagen und multinukleärer Riesenzellen in der fibrösen Kapsel, welche sich
um die Chitosan Röhrchen gebildet hatte. Zusätzlich konnte eine Verringerung der
Makrophagen im Regenerat beobachtet werde, während die Anzahl der
multinukleären Riesenzellen hier im Zeitraum von 18 Tagen bis 13 Wochen
unverändert gering blieb. Es ist ferner von Bedeutung, dass die Regulation von
beispielhaft ausgewählten Regenerations-assoziierten Proteinen in Spinalganglien
und dem Rückenmark durch die Implantation der Nervenleitschienen nach 5 und 18
Tagen nicht beeinflusst wurde. Eine morphometrische Analyse der distalen
Nervenabschnitte zeigte jedoch ein erhöhtes Maß an axonalen Verzweigungen für
die DAI Gruppe, während die DAIII Röhrchen bereits nach 18 Tagen und verstärkt

Zusammenfassung
XVII
nach 13 Wochen Risse und Bruchstellen aufwiesen und zum Teil gänzlich
auseinandergebrochen waren. Folglich wurden DAII Nervenleitschienen für weitere
Experimente ausgewählt, da sie die periphere Nervenregeneration am besten
unterstützen.
Für Projekt 2 wurden die angereicherten Nervenleitschienen für die Überbrückung
einer Distanz von 15 mm getestet. Bis zum Ende der Studie wurden mit dem
autologen Nerventransplantat konsequent die besten Ergebnisse erzielt. Lediglich
die Gruppe, welche FGF-218kDa überexprimierende Schwann Zellen transplantiert
bekommen hatte, zeigte Regeneration in 4 von 7 Tieren. In diesen konnte auch eine
partielle Wiederherstellung der Funktionalität gezeigt werden, während dies nur für
ein Tier der GDNF sowie physiologischen Schwann Zell Gruppe zutraf. Diese
Ergebnisse implizieren, dass die Nutzung von Gel zur Befüllung von
Nervenleitschienen nicht die ideale Wahl ist. Möglicherweise fungiert es als
physische Barriere und verhindert somit einen erfolgreichen Regenerationsprozess.
FGF-218kDa scheint jedoch einen kompensatorischen Effekt zu haben und das
axonale Wachstum, auch durch das Gel hindurch, zu ermöglichen.
Aus den Resultaten beider Projekte lässt sich folgern, dass DAII Chitosan-basierte
Röhrchen eine Alternative für die bereits erhältlichen Nervenleitschienen darstellen
um kleine Nervendefekte zu überbrücken. Neben den vielversprechenden
Ergebnissen im Rahmen von Projekt 1 besticht dieses Röhrchen durch seine
Transparenz. Diese Eigenschaft macht es für den klinischen Einsatz besonders
attraktiv, da so die Handhabung für den Chirurgen erleichtert wird und eine schnelle
und korrekte Positionierung des Implantates möglich ist. Bezüglich der Behandlung
von großen Nervendefekten geht die Suche nach einer idealen, intraluminalen
Leitstruktur / Füllung weiter und zusätzliche Experimente sind unerlässlich.
Alternativen, welche die Axone leiten ohne den Raum im Röhrchen komplett zu
füllen, wie z.B. sehr poröse Schwämme, Membranen oder Fasern, sind demnach
vermutlich besser für diese Aufgabenstellung geeignet als Gele.

XVIII

Introduction
- 1 -
1 Introduction 1.1 Clinical relevance of peripheral nerve injuries (PNIs) In the last decades, the understanding of biological mechanisms underlying
peripheral nerve injuries (PNIs) and the following regeneration process has
significantly increased. Nevertheless, surgical reconstruction is still challenging and
the clinical results have not substantially improved in the same time frame
(Lundborg, 2000). Despite modern microsurgery techniques that are characterized
by a high level of accuracy, adult patients are likely to suffer from functional
impairment due to incomplete or incorrect target reinnervation (Cobianchi et al.,
2013; Frostick et al., 1998; Navarro et al., 2007; Svennigsen and Dahlin, 2013).
Resulting symptoms include poor sensibility, deficient motor function, cold
intolerance, and neuropathic pain. These are consequences that often add up to a
reduced quality of life for the patient (Lundborg, 2000; Svennigsen and Dahlin,
2013). Children, on the other hand, who e.g. suffer from birth-related brachial plexus
injuries, show a better clinical recovery. Although the mechanisms causing these
differences are not yet understood, it has been proposed that young patients hold a
better cerebral plasticity (cortical reorganization) (Chemnitz et al., 2013; Frykman,
1976; Laurent and Lee, 1994; Svennigsen and Dahlin, 2013; Tajima and Imai, 1989).
According to that, previous existing, but inert connections get activated and new
connections are established via sprouting (Chen et al., 2002). Furthermore, a loss of
myelinated and unmyelinated nerve fibers and a decline in the expression as well as
transport of cytoskeletal proteins has been reported in elderly subjects, thus
explaining the better regeneration seen in children (Verdu et al., 2000).
Worldwide more than one million people are affected by PNIs. In the United States
(US) alone more than 200.000 surgeries on PNIs are performed per year resulting in
five million disability days (Daly et al., 2012; Freier et al., 2005a; Kehoe et al., 2012).
Associated health care costs total approximately $150 billion in the US and therewith
represent a major cost factor (Apel et al., 2008). Additionally, in Europe more than
300.000 PNIs are reported annually (Ciardelli and Chiono, 2006). Causes for PNIs
are manifold and surgical reconstruction may be very challenging. Working in a
military environment for one can result in battlefield wounds, of which approximately
14–22 % include injuries to the upper extremities (e.g. gunshot injuries of the

Introduction
- 2 -
brachial plexus) (Gaudet et al., 2011; Samadian et al., 2009; Waldram, 2003). Due to
the long history of war inflicted injuries, crucial progress in repairing PNIs has been
made e.g. during the American Civil War, World War I, and World War II (Friedman
et al., 2009; Robinson, 2000). Also in daily life situations (during work and leisure
time), however, injuries to the peripheral nervous system (PNS) are common and the
most prominent injuries include those to digital nerves and the brachial plexus
(Ciaramitaro et al., 2010; Fox and Mackinnon, 2011; Svennigsen and Dahlin, 2013;
Thorsén et al., 2012). Mostly young men at productive age suffer from PNIs following
vehicle accidents, but also domestic violence or sport activities represent major
causes for peripheral nerve trauma (Ciaramitaro et al., 2010; Ijkema-Paassen et al.,
2004; Thorsén et al., 2012; Waldram, 2003). Additionally, nerve damage is the result
of conditions like the carpal tunnel syndrome or diabetes (Lekholm et al., 2001;
Moore et al., 2009).
1.2 Organization of the peripheral nervous system (PNS) With multicellular life developing 1 billion years ago, it was about 500 million years
later that the first simple nervous tissue appeared in the phylum Cnidaria
(Grimmelikhuijzen and Westfall, 1995; Selden and Nudds, 2004; Westfall, 1996).
Since then, the complexity has increased and today the nervous system of
vertebrates is organized into the central nervous system (CNS) and PNS.
Approximately 86 billion neurons form the CNS, which is comprised of the brain and
its extension, the spinal cord (Azevedo et al., 2009; Waldram, 2003). The PNS is
composed of cranial nerves, spinal nerves including their roots, peripheral nerves,
and the autonomic nervous system. The soma of motor neurons of the PNS are
located in the anterior horn of the spinal cord and send their axons out to the
periphery to terminate on motor end plates in skeletal muscles. Sensory fibers, with
their cell bodies situated in the dorsal root ganglia (DRGs), convey information from
their pain, thermal, tactile, and stretch receptors, which are found e.g. in skin and
muscles to the CNS (Waldram, 2003).
The functional unit of a peripheral nerve is the axon together with its Schwann cell
(SC). This unit is surrounded by different layers of connective tissue (Fig. 1). The
endoneurium encircles the axons and provides a packaging for nerve fibers, while

Introduction
- 3 -
the perineurium defines fascicles that consist of a bundle of nerve fibers and protects
the nerve from mechanical pressure. Nerve fascicles are then again embedded in a
loose connective tissue, the epineurium (Brushart, 2011a; Waldram, 2003). SCs
spiral around axons and myelinate them in a 1:1 relationship. Approximately, 95 % of
the complete length of an axon is myelinated and only interrupted by nodes of
Ranvier (spaces between adjacent SCs). These sites enable saltatory conduction
due to membrane specializations like a concentrated appearance of voltage-gated
sodium channels (Boiko et al., 2003; Brushart, 2011a).
Figure 1: Anatomical organization of the peripheral nervous system. Somata of sensory neurons are located in DRGs, while those of motor neurons are found in the anterior horn of the spinal cord. Neurons of both modalities send their processes to peripheral targets as part of peripheral nerves. Each peripheral nerve is comprised of various fascicles, which are enclosed by the epineurium, a loose connective tissue layer. Every single fascicle is then again surrounded by another layer of connective tissue, the perineurium, and contains many axons. Each axon is further encircled by SCs, which produce a myelin sheath, as well as by a third connective tissue layer, the endoneurium (the figure was made based on illustrations of (Scholz et al., 2011; Sierpinski Hill, 2009).
The PNS displays a diverse array of axons that vary in their size and end organ and
are characterized by varying nerve conduction velocities (NCV). The latter was used
to subdivide axons into three groups. Group A represents efferent and afferent
myelinated axons of the somatic nervous system that forward information most

Introduction
- 4 -
rapidly. Within this group, axons are further defined and subdivided into alpha
(70-120 m/s), beta (30-70 m/s), gamma (15-30 m/s), and delta (12-30 m/s)
subpopulations according to their NCV (Brushart, 2011a). The largest and fastest of
these neurons, the alpha motoneurons, terminate on extrafusal muscle fibers, while
smaller and slower conducting gamma motoneurons specifically innervate intrafusal
muscle fibers (Manuel and Zytnicki, 2011). Furthermore, neurons of the alpha and
beta subpopulation forward signals from cutaneous and subcutaneous
mechanoreceptors, while gamma neurons take part in sensation of temperature and
pain. A second group is represented by B-fibers, which represent myelinated
preganglionic autonomic fibers. The third group are C-fibers, which are slow
conducting (<2 m/s), unmyelinated axons. Like some A-gamma neurons, they
mediate nociceptive pain and account for 75 % of the total axon number in
cutaneous nerves and their respective DRGs. In general, one single myelinating SC
wraps one large caliber axon (> 2µm) or one non-myelinating SC ensheathes
multiple (up to 100) small unmyelinated axons (C-fibers), thus forming so called
Remak bundles (Berger et al., 2014; Brushart, 2011a; Hall, 2005; Heavner and de
Jong, 1974; Jungnickel et al., 2010; Murinson et al., 2005; Sun et al., 2012).
1.3 Classification of peripheral nerve injuries (PNIs) Mechanisms that cause PNIs are diverse and include crush trauma, laceration,
stretching, and compression. Accordingly, the degree of the resulting PNIs is
variable as is the prognosis for the functional recovery. In 1943, based on the
severity of the lesion, Seddon described three types of injuries: neurapraxia,
axonotmesis and neurotmesis (Seddon, 1942). Sunderland later established a more
detailed classification system (Sunderland, 1951). Neurapraxia, or Sunderland
degree (SD) I, is the mildest type of injury, describing cases, in which the continuity
of the nerve trunk is preserved, but the nerve conduction is blocked at the site of the
injury. In these cases, Wallerian Degeneration does not occur and the segment
distally to the injury continuous to conduct. Full recovery is usually seen within 3 to 4
months with almost the same number of regenerated axons reaching the target
organs (Dubový, 2011; Flores et al., 2000; Isaacs, 2010; Marshall Devor and Ruth
Govrin-Lippmann, 1979; Sunderland, 1990; Waldram, 2003). Axonotmesis (SD II),
on the other hand, is characterized by Wallerian degeneration and loss of

Introduction
- 5 -
spontaneous muscle activity for about three weeks. Axons are disrupted, but the
surrounding connective tissue (epineurium, perineurium, and endoneurium) stays
intact. Therefore, sprouting axons can send their axons through their original
endoneurial tubes and spontaneous recovery is possible. The recovery time
depends on the distance between the injury and distal target organs, with axons
regrowing at a rate of 1-1.5 mm per day (depending on nerve and progress of
reinnervation). If not only the axon is damaged, but also layers of the surrounding
connective tissue, the injury is classified as neurotmesis (Flores et al., 2000; Isaacs,
2010; Seddon et al., 1943; Waldram, 2003). Sunderland further divided injuries of
this level according to the degree of connective tissue disruption (Sunderland, 1990,
1951). SD III indicates that aside from the axons also the endoneurium is affected
and has collapsed. Due to the missing guidance of this layer regenerating axons
have difficulties to find their way and tend to grow together with scar tissue.
Nevertheless, regeneration is possible, but unspecific reinnervation may result in
abnormal motor and sensory recovery. In injuries of SD IV, the epineurium is the
only structure that remains intact and spontaneous recovery does not occur. A
complete transection of the nerve is the highest degree according to Sunderland (SD
V) (Allodi et al., 2012; Flores et al., 2000; Sunderland, 1990, 1951). Surgical
intervention is needed if a patient suffers from an injury that is classified as SD IV or
SD V accordingly, but functional recovery is often incomplete. Furthermore, patients
frequently have to deal with pain and dysthesia due to the formation of neuroma that
are composed of immature axons and connective tissue (Allodi et al., 2012; Isaacs,
2010; Mourad et al., 2001; Navarro et al., 2007; Svennigsen and Dahlin, 2013).
1.4 Degeneration and regeneration processes following injury Unlike the CNS, the PNS has an intrinsic capacity to regenerate and reinnervate
distal target organs following injuries. Neurons perform a switch in function, thus
changing from a transmission state to a growth state (Allodi et al., 2012). Many
axons are able to assemble a new growth cone within hours after nerve crush, while
about 50 % undergo apoptosis (Bradke et al., 2012; Dubový, 2011; Törnqvist and
Aldskogius, 1994). At the same time, the creation of a nerve regeneration friendly
environment (see below) distally to the lesions starts with a process called Wallerian

Introduction
- 6 -
degeneration, if the lesion is at least classified as SD two (Flores et al., 2000;
Liebermann, 1971).
1.4.1 Neuronal response Following nerve injury, morphological changes occur in the cell bodies of the affected
neurons. The localization of the nucleus is altered to a more eccentric position and
its shape appears deformed (Fig. 2). Swelling of the cell body and dissolution of the
Nissl bodies (chromatolysis) are further characteristics that define neurons after
transection (Gaudet et al., 2011; Kreutzberg, 1995; Liebermann, 1971; Navarro et
al., 2007; Pabari et al., 2011).
The axonal regeneration process does not directly start at the level of the lesion, but
rather a few internodal segments more proximal, while the immediately affected
segments undergo degeneration (Raimondo et al., 2011). The neuronal reaction
after transection of the axons is characterized by an increased production of
components relevant for the assembly of growth cones as well as structural
components. Tubulin, actin, and growth-associated protein 43 (GAP-43) are e.g.
reported to be increasingly synthesized and transported, while proteins required for
neurotransmitter generation are down-regulated (Brushart, 2011b; Navarro et al.,
2007; Raimondo et al., 2011). The signals that trigger this shift from a “transmitting”
to a “regenerative” state of the neuron include various mechanisms. For example,
following axonal disruption, and preceding sealing of the axon, an influx of
extracellular calcium and sodium causes a burst of action potentials. Furthermore,
the retrograde transport is interrupted providing the cell body with negative and
positive signals. One negative signal e.g. is the decrease in retrograde transported
nerve growth factor (NGF), while contact with trophic molecules like e.g. cilliary
neurotrophic factor (CNTF) serves as a positive signal (Allodi et al., 2012; Hall, 2005;
Kirsch et al., 2003; Navarro et al., 2007).

Introduction
- 7 -
Figure 2: Degenerative and regenerative events following injuries to the mammalian PNS. (A-B) Following PNIs, degeneration occurs in the directly affected internodal segments of the proximal nerve stump and in the distal nerve stump. Following a brief time period, which allows e.g. influx of extracellular calcium that triggers, amongst others, a burst of action potentials and a subsequent switch to a regenerative state of the neuron, the tip of the proximal nerve stump gets sealed within hours. Myelinating SCs start to release their myelin and the axoplasm undergoes degradation. The rapid inflammatory response includes resident macrophages and invading neutrophils, (C) while recruited macrophages and T-lymphocytes arrive at the lesion site within 2 days and infiltrate the complete distal stump within 3 to 7 days after nerve injury. (E) Dedifferentiated SCs then start to form the bands of Büngner along which axonal sprouts are guided to peripheral target organs. (F) Successful reinnervation ultimately leads to retraction of unsuccessful axon sprouts, reversal of muscle atrophy, chromatolysis as well as the formation of mature SC-axon interactions. The latter are characterized by thinner myelin sheaths and shorter internodal spacing (Deumens et al., 2010; Gaudet et al., 2011; Hall, 2005; Nectow et al., 2011). The figure was made based on illustrations of (Deumens et al., 2010; Gaudet et al., 2011).
1.4.2 Changes in the distal environment and axonal regeneration Distally to the injury site, a degenerative process called Wallerian degeneration
starts initiated by the detachment of axons. Subsequently, axoplasm, axolemma, and
myelin sheaths immediately start to degrade. Calcium dependent proteolytic
enzymes like calpains are mediators causing the breakdown of the cytoskeleton, a
process that starts within 2 days in rodents and within seven days in humans

Introduction
- 8 -
(Dubový, 2011; Gaudet et al., 2011; Hall, 2005). Meanwhile, SCs stop to produce
myelin proteins like myelin basic protein or myelin protein zero (P0), enhance
neurotrophic factor (NTF) and GAP-43 production, and start to proliferate within their
basal lamina tubes. Within 48 h, the myelin sheaths have been segmented into short
pieces. During the first days, SCs play a major role in fragmentation and
phagocytosis of myelin debris. Resident macrophages, which represent about 4-9 %
of the endoneurial cellular population in a normal nerve, then directly begin to
remove axonal as well as myelin debris that contains neurite outgrowth inhibitors like
e.g. myelin-associated glycoprotein (MAG). Production of cytokines and chemokines
by SCs and resident macrophages leads, amongst others, to an accumulation of
blood-derived monocytes within 4 days after injury (Brushart, 2011c; Dubový, 2011;
Freier et al., 2005a; Gaudet et al., 2011; Hall, 2005; Hirata and Kawabuchi, 2002;
Ide, 1996; Stoll et al., 2002). Following differentiation into macrophages, they support
the myelin clearance as well as cytokine production, thereby facilitating SC migration
and axonal growth. Their accumulation reaches a peak 2 weeks after injury with an
150-fold increase in number (Brushart, 2011c). In case of contact with MAG on the
surface of newly formed myelin, recruited macrophages exit from the basal lamina to
re-enter the circulation (Gaudet et al., 2011). Usually, the identification of resident
and invading macrophages is possible, because resident macrophages can be
immunocytochemically recognized with ED-2 antibody and invading macrophages
display immunoreactivity for ED-1 antibody. Following nerve injury, however,
resident macrophages start to proliferate and show ED-1 immunoreactivity making
these two populations indistinguishable (Dubový, 2011). Aside from SCs and
macrophages, neutrophils phagocytize debris in the early phase of Wallerian
degeneration, but do not survive this act and undergo apoptosis. T-lymphocytes, on
the other hand, are mainly crucial during the later phase of axonal regeneration by
producing pro- and anti-inflammatory cytokines (Gaudet et al., 2011; Stoll et al.,
2002).
With the assembly of the growth cone progressing, SCs start to align and form the
bands of Büngner. They serve as a guidance structure for the regrowing axons
towards peripheral reinnervation targets (Gaudet et al., 2011; Pabari et al., 2011;
Svennigsen and Dahlin, 2013). Upon contact, SCs start to remyelinate the
regenerating axons. The morphology of these newly remyelinated axons is

Introduction
- 9 -
characterized by thinner myelin sheaths and shorter internodes compared to the
proximal nerve segment causing a slower nerve NCV (Hall, 2005; Ide, 1996;
Svennigsen and Dahlin, 2013). After weeks to months, the regenerating nerve is
characterized by numerous small nerve bundles called mini-fascicles. In the process
of approaching the periphery, the number of bundles decreases resulting in the
maturation of only the axons that reach distal targets (Freier et al., 2005a).
1.5 Surgical repair strategies and their limitations The severity of peripheral nerve injuries decides if and what kind of surgery is
necessary. Clean nerve transections should be surgically treated as soon as
possible to facilitate an easier approximation and alignment of the two nerve
segments. This, however, is only possible, if the reconnection can be performed
without causing any tension. Short gaps, on the other hand, are usually treated with
commercially available conduits (limited to defects not exceeding 3 cm in humans or
1 cm in rats) or autologous nerve grafts (ANGs), which are also preferentially used
to overcome longer gaps (Bell and Haycock, 2011; Daly et al., 2012; de Ruiter et al.,
2009; Isaacs, 2010; Nectow et al., 2011; Pfister et al., 2011).
1.5.1 End-to-end repair Minor nerve injuries that include no or only very small gaps (not greater than
0.5-2 cm) are treated by reconnecting the two nerve ends, as long as tension-free
repair is possible (Bell and Haycock, 2011; Daly et al., 2012). After removing the
scarred part of the affected nerve tissue, fascicular patterns and longitudinal surface
vessels are consulted as markers to guarantee an accurate alignment. The
reconnection can be achieved with epineurial sutures, but approximation of deep
fascicles is not ensured with this method. Some surgeons therefore align individual
fascicles by suturing their perineurium. This, though, implicates the risk of
misdirecting all axons, if the repair is not performed accurately. Grouped fascicular
alignment, on the other hand, is less problematic. None of these suture methods,
however, has been proven to produce superior results (Isaacs, 2010; Pabari et al.,
2010).

Introduction
- 10 -
1.5.2 Autologous nerve graft (ANG) Defects that cannot be reconnected by end-to-end repair without causing tension
are usually treated with ANGs. This standard therapy includes the harvest of nerve
tissue from the sural nerve or other dispensable sensory nerves like the medial and
lateral antebrachial cutaneous nerves or the superficial radial nerve. The advantage
of these nerves is that they are easily accessible and provide the opportunity to
obtain long nerve grafts (Isaacs, 2010; Pabari et al., 2010). Injuries of small nerves
are usually treated with a single nerve graft, while larger nerves require the use of
cable grafts. In the latter case, multiple grafts are used as interponates to overcome
the gap (Isaacs, 2010). Studies have, however, shown, that motor nerve grafts
support regeneration of motor neurons more effectively, a problem, which probably
occurs due to mismatch issues regarding axonal size, distribution and alignment.
Furthermore, SCs of different nerve modalities express varying phenotypes and
produce modality specific growth factors (Daly et al., 2012; Höke et al., 2006;
Isaacs, 2010; Nichols et al., 2004; Pabari et al., 2010). Additional disadvantages of
this method include donor site morbidity, the need of a second surgery, and the
limited amount of nerve tissue that is available. Furthermore, a prolonged
inflammatory reaction is caused as a result of secondary removal of axonal and
myelin debris. Nevertheless, ANGs have proven to be suitable for nerve gaps of up
to 5 cm length with a success rate of about 50 % (Bell and Haycock, 2011; Daly et
al., 2012; Deumens et al., 2010; Lin and Marra, 2012).
1.5.3 Allograft Since treatment of PNIs with ANGs involves disadvantages like limited amount of
available tissue and donor site morbidity, allografts represent a fitting alternative
strategy to overcome nerve gaps. These grafts can be gained from donor or
cadaveric nerve tissue and provide guidance structures as well as donor SCs. The
required systemic immunosuppression for 18 to 24 months, however, limits the
clinical application of this method, which has so far led to variable results (Bell and
Haycock, 2011; Kehoe et al., 2012; Mackinnon et al., 2001; Pabari et al., 2010; W.
Ray, 2010). Mackinnon and colleagues reported on 10 patients that showed motor
and sensory recovery after nerve repair with cadaveric allografts, while one patient
suffered from rejection of the graft following subtherapeutic immunosuppression
(Mackinnon et al., 2001).

Introduction
- 11 -
To avoid immunosuppression, allografts derived from human nerve tissue have been
decellularized by AxoGen Inc. (Alachua, FL). This process preserves the structure of
the extracellular matrix while clearing cells, cellular debris, and proteins like
chondroitin sulfate proteoglycans that are known to act as neurite outgrowth
inhibitors (axogenic.com, 2014; Kehoe et al., 2012). Decellularized allografts from
AxoGen have received approval from the “Food and Drug Administration” (FDA) and
return of sensation was reported in gaps of up to 3 cm length, while no rejection was
observed (Karabekmez et al., 2009; Kehoe et al., 2012). A multicenter clinical trial
study confirmed these findings with meaningful recovery seen in 87.3 % of the
patients suffering from injuries of up to 5 cm length in sensory, motor, and mixed
nerves (Brooks et al., 2012)
1.5.4 CE and FDA Approved Conduits As an alternative to the standard therapy, short gaps can be treated with nerve
conduits to allow tensionless nerve repair of e.g. digital nerves, since they have been
shown to provide a permissive environment for regrowing axons (Fig. 3). Additionally, this method is time saving, a factor that is important for clinical
application (de Ruiter, 2009; Meek and Coert, 2008; Pabari et al., 2010; Williams et
al., 1983). At first, mostly silicone conduits had been tested, but up to the present
various different conduits have now been experimentally investigated and some
have been cleared for marketing in the US and Europe (de Ruiter, 2009; Meek and
Coert, 2008). To assure that only safe medical devices enter the market, the FDA, a
regulatory system in the US, is responsible to keep the risk minimal for patients (IOM
(Institute of Medicine), 2012). Therefore, manufacturers have to demonstrate safety
and efficacy of high risk devices. The requirements that need be fulfilled, however,
vary depending on the classification of the device. The classification ranges from
low-risk devices in class I (e.g. stethoscopes) to medium-risk class II devices
(computed tomographic scanners and nerve guidance conduits like Neurolac) and
high-risk class III products such as deep-brain stimulators, but also all devices
containing chitin and its derivatives. With the same aim as the FDA, the “Conformité
Européenne” (CE) is responsible for the European market. While low-risk devices
only have to be declared, more complex devices are in need to be cleared by a

Introduction
- 12 -
notified body (specialists in evaluating different products) (Kehoe et al., 2012;
Kramer et al., 2010; Strusycyzk and Struszczyk, 2007).
Figure 3: Regeneration processes across a 10 mm gap within a hollow nerve guidance conduit. Regeneration within a hollow nerve guidance conduit can be divided into five phases: (A) the initial phase is characterized by an influx of fluid that contains NTFs, inflammatory cells, and extracellular matrix precursor molecules. (B) Subsequently, a fibrin cable forms within one week after injury, which connects the two nerve stumps. (C) SCs then start to migrate along the fibrin cable and proliferate to form the bands of Büngner during the second week of regeneration before (D) axonal sprouts can cross the defect in the axonal phase. During this phase, it is likely that the fibrin cable has already vanished. (E) Ultimately, mature SCs start to remyelinate larger regenerated axons (Belkas et al., 2004a; Daly et al., 2012; de Ruiter, 2009; Lundborg et al., 1981; Madison et al., 1987). The figure was made based on illustrations of (Belkas et al., 2004a; Daly et al., 2012).

Introduction
- 13 -
1.5.4.1. Non-resorbable conduits
Salubridge™ and SaluTunnel™ (both Salumedica LLC; Atlanta, GA) are marketed
worldwide and are made of polyvinyl alcohol (PVA) hydrogel. While Salubridge™
serves as a conduit to enable axonal growth across a nerve gap; SaluTunnel™ is
indicated for injuries without substantial loss of nerve tissue. These products are the
only synthetic non-resorbable conduits marketed, but so far no published data from
clinical studies are available (Kehoe et al., 2012).
1.5.4.2 Type I collagen
Advantages of collagen include its adhesive properties for different cell types, its
fibrillar structure, which is preserved during the manufacturing process, and the fact
that it can be easily isolated and purified. There are various conduits based on this
major extracellular matrix (ECM) component. NeuraGen® (Integra Life Sciences
Corporation; Plainsboro, NJ) is reported to be as effective as ANGs and sensory and
motor recovery were equivalent to results with direct suture in a multicenter study
when the nerve gap was 6 mm or less (Archibald et al., 1991; Boeckstyns et al.,
2013; Kehoe et al., 2012). NeuraWrap™ is another product from Integra that has an
identical material composition and is serving as a wrap (Kehoe et al., 2012). Just
recently, Georgiou and colleagues could show robust neuronal regeneration across
a 15 mm gap within 8 weeks when using this wrap in combination with rods of rolled-
up sheets and aligned SCs as fillers (Georgiou et al., 2013). Furthermore,
NeuroMatrix™ and Neuroflex™ (Collagen Matrix Inc., Franklin Lakes, NJ) are also
collagen-based conduits. The latter is highly flexible and has been created to bridge
nerve defects of up to 2.5 cm length. NeuroMend™ is another product of this
company with the same material compositions, but neither of the three devices has
been tested in published animal or clinical studies (Kehoe et al., 2012).
1.5.4.3 Porcine small intestinal submucosa (SIS)
AxoGuard™ nerve connector and AxoGuard™ nerve protector (AxoGen, Alachua,
FL) are based on small intestinal submucosa (SIS), which is a cell-free collagen
matrix that provides a good mechanical support and supports remodeling of the host
tissue by accelerating new vessel growth, cell proliferation, and differentiation
(Kehoe et al., 2012).

Introduction
- 14 -
1.5.4.4 Polyglycolic acid (PGA)
Neurotube® (Synovis Micro Companies Alliance; Birmingham; AL) is a synthetic
porous nerve conduit made of polyglycolic acid (PGA), which allows the infiltration of
oxygen. Results from animal as well as clinical studies vary, but its usage has shown
efficacy in defects up to 3 cm. A disadvantage, however, is the high rate of
degradation and presence of acidic degradation products (Kehoe et al., 2012; Meek
and Coert, 2008).
1.5.4.5 Poly D,L lactide-co-ɛ-caprolactone (PCL)
Neurolac® (Polyganics, Groningen, NL) is the only transparent nerve guidance
conduit, a property that facilitates the correct positioning of the nerve stumps in the
device, and offers the opportunity for the surgeon to remove eventual blood clots that
could hamper nerve regeneration (de Ruiter, 2009; Kehoe et al., 2012;
Polyganics.com, 2014). On the other hand, it is inflexible, highly rigid, thus causing
needle breakage, and swelling of the conduit. Additionally, neuroma formation as
well as foreign body reactions have been described (Kehoe et al., 2012; Meek and
Coert, 2008; Meek and Den Dunnen, 2009).
1.6 Considerations to design the future ideal conduit Approved nerve guidance conduits are so far limited to bridge nerve defects of up to
3 cm length and are associated with insufficient functional recovery. Thus, to
overcome these limitations, new strategies are needed (Bell and Haycock, 2011;
Daly et al., 2012; de Ruiter et al., 2009; Gu et al., 2011). There are various design
possibilities of conduits that can influence axonal regeneration. Future conduits
should not only have mechanical strength, while still being suturable, but should also
be made of biocompatible as well as biodegradable materials. The latter allows
degradation at a reasonable rate, if adjustments are undertaken properly, to avoid
inflammatory reactions. The device should further be flexible without having the
tendency of swelling to avoid compression of regenerated axons. Additionally, the
wall should be semi-permeable to allow exchange of oxygen and nutrients, but at the
same time ingrowth of fibrous tissue should be prevented and NTFs should be kept
inside the conduit. Introduction of 3D guidance structures such as fibers, sponges,
membranes or hydrogels inside the tube could further support axonal outgrowth and

Introduction
- 15 -
SC migration and are thus promising tools to promote regeneration. The same
expectations are held for the incorporation of support cells and NTFs. Also, the
surface structure was found to be important. Smooth surfaces enable the growth of a
free floating nerve cable, while rough surfaces cause a complete covering of the
conduit (Clements et al., 2009; Daly et al., 2012; de Ruiter, 2009; Huang and Huang,
2006; Jiang et al., 2010; Kehoe et al., 2012; Mukhatyar et al., 2014; Pabari et al.,
2011). Also, since the ultimate goal is to market the designed device for clinical
usage, it is important that the end product is easy to produce, to sterilize, and offers
simple handling for the surgeon. The latter includes transparency of the material,
since this characteristic facilitates correct positioning of the conduit (Chalfoun et al.,
2006; Daly et al., 2012; Polyganics.com, 2014)
1.6.1 Biocompatible and biodegradable polymers Biocompatibility and biodegradability are important properties of all materials
included in a nerve conduit. Assessment of biocompatibility is e.g. performed with
regard to blood compatibility (haemolysis, destruction of blood components,
coagulation, and thrombus formation are limiting factors) and histocompatibility
(material should be non-toxic and not elicit immunological rejection responses from
the surrounding tissue). Additionally, degradation in a controlled manner is important
to avoid secondary injuries to the newly grown nerve tissue (Daly et al., 2012).
Otherwise, a second surgery might be necessary to remove the material since its
permanent presence can cause chronic tissue responses or nerve compression.
Moreover, it would be good to match degradation kinetics to the axonal regeneration
rate (Daly et al., 2012; Jiang et al., 2010; Pabari et al., 2011).
Materials that fulfill these requirements include synthetic materials like aliphatic
polyesters (e.g. polycaprolactone) and naturally-derived biopolymers. The latter
includes ECM molecules such as collagen, laminin, and fibronectin, but also silk
fibroin and polysaccharides like alginate or chitin and chitosan (Daly et al., 2012).
1.6.1.1 Chitosan
Chitin is a natural polymer that can be gained from the exoskeleton of arthropods,
cuticles of insects, walls of fungi, and shells of crustaceans (e.g. the deep-water
shrimp Pandalus borealis). It is the second most abundant organic resource on earth
after cellulose (Freier et al., 2005c; Kardas et al., 2012; Singh and Ray, 2000). Intra-

Introduction
- 16 -
and intermolecular hydrogen bonds between the polymer chains lead to a high
insolubility of chitin (Kardas et al., 2012). Full or partly deacetylation of chitin
converts the material into chitosan (Fig. 4), which is insoluble in water and neutral or
basic pH (due to free amino groups), while an acidic pH results in protonation of the
amino functions, thus facilitating solubility in water (Freier et al., 2005a; Kardas et al.,
2012; Singh and Ray, 2000). Chitin, however, degrades faster than chitosan, is more
compatible with blood, and activates fever macrophages (Freier et al., 2005b).
Figure 4: Chemical structure of chitin and chitosan. Chitosan can be gained from chitin through alkaline hydrolysis of the N-acetyl groups. Chitosan degradation occurs by non-specific enzymes such as lysozymes, chitinases, cellulases or hemicellulases, proteases, lipases, ß-1,3-1,4 glucanases as well as chitosanases (Raafat and Sahl, 2009).
Chitosan is comprised of ß(1-4) linked D-glucosamine and N-acetyl-D-glucosamine
subunits with a degree of acetylation (DA) between 0 % and approximately 60 %
(Freier et al., 2005b). Degradation of chitosan occurs, amongst others, by lysozyme,
an enzyme that is present in the human body and has the ability to hydrolyze the
ß(1 -4) linkages between D-glucosamine and N-acetyl-D-glucosamine in chitin and
chitosan. The rate of degradation is thereby related to the distribution of the N-acetyl
groups and to the DA, with higher DAs resulting in a more rapid degradation (Han et
al., 2011; Kean and Thanou, 2010). Chitosan has been investigated for various
biomedical applications due to its biodegradability, but also biocompatibility, low

Introduction
- 17 -
toxicity, and molecular similarity to glycosaminoglycans that are part of the basal
membrane and ECM (Freier et al., 2005a; Siemionow et al., 2010). Furthermore,
chitosan is known to accelerate wound healing and shows antimicrobial activity.
While Chitosan is not cleared as “Generally Recognized As Safe (GRAS)” by the
FDA, it has been approved for some medical applications. Accordingly, several
wound dressings that are based on chitosan are commercially available and it is e.g.
used in food preservation and dentistry as well as being considered as a potential
carrier in drug delivery systems (Itoh et al., 2003; Raafat and Sahl, 2009; Singh and
Ray, 2000). Chitosan has further been studied for peripheral nerve reconstruction
and is thought to be a promising candidate for tissue engineering applications. In
vitro studies revealed that chitosan membranes allow SC adhesion, survival,
migration, and proliferation as well as neuronal survival and differentiation (Simões
et al., 2011; Wang et al., 2005; Yuan et al., 2004). Chitosan has already been tested
alone or in combination with other materials for its possible usage in peripheral nerve
repair. Successful bridging of small gaps (8-12 mm) has repeatedly been reported
with hollow chitosan or chitosan combined conduits (Ao et al., 2011; Han et al.,
2010; Patel et al., 2006; Raisi et al., 2012; Simões et al., 2010; Xie et al., 2008; Xu et
al., 2011; Zheng and Cui, 2012). Treatment of 15 mm gaps with chitosan containing
conduits has also been described and chitosan/PGA and chitosan/poly(lactic-co-
glycolic acid ) (PLGA)-based conduits led to encouraging results bridging 30, 50, or
60 mm defects in dog or rhesus monkey nerve injury models (Ding et al., 2010; Hu et
al., 2013, 2009; Itoh et al., 2003; Wang et al., 2005; Xue et al., 2012).
1.6.2 Luminal fillers Since regeneration following treatment with conduits so far is limited to small gaps,
modifications of hollow tubes are considered that might enhance the success of
target reinnervation in cases of short and long gaps by stabilizing the unformed or
incomplete fibrin cable. Various approaches to incorporate intraluminal guidance
structures have been investigated including multichannel conduits or the introduction
of filaments, sponges, or gels such as collagen- or laminin-containing gels into the
scaffold (Bell and Haycock, 2011; Daly et al., 2012; de Ruiter et al., 2009; Gu et al.,
2011).

Introduction
- 18 -
1.6.2.1 NVR-Gel-A hyaluronic and laminin-based 3D guidance structure
Filling the conduit with ECM components like collagen, fibronectin, laminin, or
hyaluronic acid (HA) is thought to facilitate early ingrowth of axons and support cell
migration (de Ruiter et al., 2009; Jiang et al., 2010; Khaing and Schmidt, 2012; Lin
and Marra, 2012). These components can serve as a basis for hydrogels, which are
injectable and gelling fillers that can also serve as carriers for cells and growth
factors. Typical hydrogels are cross-linked hydrophilic water-soluble polymer
networks, which are used in various medical devices including bone, tissue and
nerve repair (Lin and Marra, 2012).
NVR (Neural and Vascular Reconstruction)-Gel is comprised of HA combined with
laminin (Rochkind et al., 2009; Shahar et al., 2001). HA is a non-sulfated
glycosaminoglycan, a natural repeating disaccharide that is ubiquitous in the human
body (e.g. in the vitreous of the eye and connective tissue). It is e.g. involved in the
regulation of tissue hydration, water transport, inflammation (it has an
anti-inflammatory effect), and lubrication of joints. Accordingly, medical applications
of this hydrophilic molecule have been clinically used for 30 years and include the
supplementation of joint fluid in patients with arthritis and its use as wound dressing.
Moreover, it is biocompatible and non-immunogenic, reduces nerve sensitivity
associated with pain, and regulates cell proliferation, migration, and differentiation.
HA is highly negatively charged and therefore cell adhesive components must be
added such as ECM components like laminin (Burdick and Prestwich, 2011; de
Ruiter et al., 2009; Khaing and Schmidt, 2012; Necas et al., 2008). Laminin, a
trimeric glycoprotein, is part of the basal lamina and promotes neurite outgrowth and
elongation. Like HA, it also has a positive effect on SC proliferation and migration
and additionally provides support for SC adherence. Overall, laminin acts as a cue
that guides regrowing axons and thus has the ability to promote nerve regeneration
(Belkas et al., 2004a; Daly et al., 2012; de Ruiter, 2009; Jiang et al., 2010; Swindle-
Reilly et al., 2012). Importantly, the laminin included in the NVR-Gel is laminin
1,while laminin 5 has been associated with carcinogenesis (Shahar et al., 2001;
Waterman et al., 2007).

Introduction
- 19 -
1.6.3 Incorporation of support cells The addition of support cells to different luminal fillers inside the nerve conduit has
been widely studied and is a favorable tool to enhance peripheral nerve regeneration
across long gaps (de Ruiter, 2009; Pabari et al., 2011). The most promising
candidates to be used as cellular components in an enriched conduit include
adipose-derived stem cells (ADSCs), olfactory ensheating cells (OECs), bone
marrow-derived mesenchymal stem cells (BMSCs), and SCs. (Ao et al., 2011; Bell
and Haycock, 2011; Di Summa et al., 2011; di Summa et al., 2010; Jiang et al.,
2010; Radtke et al., 2011; Wei et al., 2011; Xue et al., 2012; ; Yuan, 2004 et al.,
2004; Zheng and Cui, 2012).
1.6.3.1 Schwann Cells (SCs)
SCs are the main glial cells of the vertebrate PNS and are known for their essential
role during peripheral nerve regeneration. Aside from expressing a myelinating or
unmyelinating phenotype, sensory and motor phenotypes have been described for
SCs. They express different molecular markers, thereby creating modality specific
pathways and facilitating correct target reinnervation. The most prominent example
is the L2/HNK1 carbohydrate epitope, which is only expressed by SCs encircling
motor axons, while neural cell adhesion molecule (NCAM)-positive cells are
associated with sensory axons. L2/HNK1, however, is down-regulated following
injury and reemerges upon contact with regenerating axons (Allodi et al., 2012; Höke
et al., 2006; Nichols et al., 2004).
Following PNIs, SCs switch their phenotype to a regenerative state and start to
proliferate (reaching up to 4-17 times higher numbers compared to intact nerve) and
migrate (Daly et al., 2012; Gu et al., 2011; Haastert et al., 2006b; Madduri and
Gander, 2010). Previously, it has been shown that incorporation of SCs improves
peripheral nerve regeneration across a 2 cm gap in the rat median nerve and across
a distance of 18 mm in the sciatic nerve (Ansselin et al., 1997; Brushart, 2011a;
Sinis et al., 2005). Autologous SCs were further reported to allow regeneration over
a 6 cm gap in the peroneal nerve of rabbits, if applied in combination with Matrigel
within a gluteal vein (Strauch et al., 2001). SCs provide a substratum consisting of
structural and adhesive ECM molecules for axonal outgrowth and secrete a cocktail
of NTFs, thus creating the ideal environment for regeneration (Huang and Huang,

Introduction
- 20 -
2006). Depending on the modality of the affected nerve and the phenotype of the
SCs, the composition of produced NTFs varies. While SCs of the sensory system
produce high levels of NGF, brain derived neurotrophic factor (BDNF), insulin like
growth factor, and erythropoietin, SCs of the motor system secrete glial cell-derived
growth factor (GDNF) (Gu et al., 2011; Madduri and Gander, 2010). Low and high
molecular isoforms of fibroblast growth factor (FGF)-2 are also up-regulated by SCs
at the lesion site and differentially promote peripheral nerve regeneration (Grothe et
al., 2001; Haastert et al., 2006a). SC seeded chitosan conduits have already been
tested and were successful in improving regeneration (Ao et al., 2011).
1.6.4 Incorporation of neurotrophic factors (NTFs) Since the Nobel prize-awarded discovery of NGF and its role as a target-derived
factor for the survival and maintenance of neurons during development by Rita
Levi-Montalcini and Viktor Hamburger, half a century ago, multiple families of NTFs
that are part of various biological activities have been identified. NGF, BDNF,
neurotrophin (NT)-3, NT-4/5, and NT-6 are part of the NT-family, CNTF belongs to a
neuropoietic family, and GDNF and FGF are eponyms for their respective family. In
the last decades, other sources of trophic support have been identified such as SCs
and the nerve regenerating promoting effects of these factors have been
investigated (Jiang et al., 2010; Levi-Montalcini and Angeletti, 1968; Levi-Montalcini,
1987; Oppenheim and Bartheld, 2008). For example, null mutants of NGF or its
receptor trkA suffered from loss of sympathetic and some small sensory neurons,
while NT-3 and trkC absence induced loss of large diameter neurons (Oppenheim
and von Bartheld, 2008; Snider, 1994). Furthermore, controlled release (e.g. via
genetically engineered SCs) of NTFs promotes outgrowth and survival of regrowing
axons and is therefore an option to improve nerve regeneration across long gaps
(Daly et al., 2012; Pfister et al., 2007; Timmer et al., 2003).
1.6.4.1 Glial cell-derived neurotrophic factor (GDNF)
GDNF, a member of the tumor necrosis factor-ß superfamily, has been widely
investigated and is mainly associated with being a survival factor for mesencephalic
dopaminergic cells and motor neurons as well as for some sensory and autonomic
neurons (Boyd and Gordon, 2003a; Brushart, 2011a; Höke et al., 2002; Jiang et al.,
2010; Oppenheim et al., 1995; Yan et al., 1995). It is retrogradely transported to

Introduction
- 21 -
motor and sensory neurons and is up-regulated in DRGs as well as ventral roots
following injury (Höke et al., 2006, 2002). Signaling occurs, amongst others, via
receptor complexes that consist of a GFR-α1 and ret subunit, both of which are
up-regulated in axotomized motoneurons, while GDNF messenger ribonucleic acid
(mRNA) has not been described in regular or lesioned motoneurons. The latter,
however, as well as the protein, have been found to be increased in SCs of the
sciatic nerve following injury peaking at 7 days (Boyd and Gordon, 2003a; Höke et
al., 2002). Synthetic polymer conduits sustainingly releasing GDNF have further
been shown to allow regeneration of motor and sensory neurons over major gaps of
15 mm length (Fine et al., 2002; Kokai et al., 2011). Moreover, blending of chitosan
conduits with laminin and GDNF as well as delivery of the latter via transfected SCs
in a silicone tube has led to an enhanced functional recovery over a 10 mm gap (Li
et al., 2006; Patel et al., 2007). The number of myelinated axons has further been
reported to be greater with GDNF released from a rod than with NGF or NT-3 and
axonal elongation promoting effects have been reported following local
administration (Barras et al., 2002; Fine et al., 2002; Madduri et al., 2009).
Additionally, impaired regeneration following chronic axotomy has been associated
with a decline of GDNF expression by chronically denervated SCs (Höke et al.,
2002). In correspondence, GDNF administration via microspheres or pumps has
been shown in some studies to only be effective following nerve repair after a
2 month period of chronic axotomy, but fails to support regeneration in cases of
immediate nerve repair (Boyd and Gordon, 2003a, 2003b; Wood et al., 2013b).
Furthermore, intraperitoneal (i.p.) injection of GDNF affected SCs by inducing an
increased proliferation of unmyelinated SCs as well as myelination of small
unmyelinated SCs (Höke et al., 2003).
1.6.4.2 Fibroblast growth factor (FGF)-2
The vertebrate FGF family consists of 22 members that have been grouped into 7
subfamilies. These factors take part in regulating cell proliferation, migration, and
differentiation during development and play a role in tissue repair following injury.
Signaling of these NTFs takes place via binding to high-affinity tyrosine
transmembrane receptors (R) 1-4 and heparin or heparin sulfate proteoglycans
(Dorey and Amaya, 2010; Grothe and Nikkhah, 2001; Ornitz and Itoh, 2001;
Woodbury and Ikezu, 2014).

Introduction
- 22 -
FGF-2 has been detected in DRGs during development and adulthood and has been
reported to be up-regulated following nerve injuries. Also, SCs as well macrophages
express FGF-2 and its receptors at the lesion site (Grothe and Nikkhah, 2001;
Grothe et al., 2001, 1996). Furthermore, FGF-2 as well as the FGFR1-3 are usually
expressed at low levels in adult sciatic nerve, but are subjected to an up-regulation
after nerve crush (Grothe et al., 2001). Administration of exogenous FGF-2 to the
lesion site has been found to prevent cell death of sensory neurons and promote
neurite outgrowth and vascularization of regrowing axons (Aebischer et al., 1989;
Danielsen et al., 1989; Grothe and Nikkhah, 2001). Different isoforms of FGF-2
result from the alternate start codons with the additional amino-terminal sequence of
the high-molecular-weight isoforms containing a nuclear-localization signal (Grothe
and Nikkhah, 2001; Ornitz and Itoh, 2001; Woodbury and Ikezu, 2014). Following
nerve transection, these isoforms are differently regulated with an overall increase of
all three isoforms in the proximal nerve stump (higher up-regulation of 18-and 21-
kDa FGF-2 isoform than 23-kDa isoform), while a decrease has been described in
the distal stump after 5 days using RT-PCR (real-time polymerase chain reaction),
and no FGF-2 protein has been found with Western Blot analysis after 7 days. In the
case of a nerve crush, however, an up-regulation of all three isoforms is seen in the
proximal and distal nerve segment (Brushart, 2011a; Meisinger and Grothe, 2002).
In accordance with the differing patterns of up-regulation of these isoforms, it was
reported that they have different effects on peripheral nerve regeneration. While
FGF-218kDa inhibits myelination, but allows functional motor recovery, FGF-221/23kDa
has been associated with promotion of early sensory target reinnervation over long
gaps of 15 mm (Haastert et al., 2006a). Furthermore, it was recently shown that
FGF-218kselectively promotes neurite outgrowth of spinal motor neurons in vitro
(Allodi et al., 2013).
1.7 Tissue responses following implantation of medical devices Implantation of biomaterial devices causes a series of tissue responses within 2 to 4
weeks after surgery including blood-material interactions, provisional matrix
formation, acute (less than a week) and chronic inflammation (usually of short
duration) as well as fibrous capsule development. The foreign body response (FBR)
is part of the chronic inflammation and includes monocytes, macrophages, and the

Introduction
- 23 -
formation of multinucleated giant cells (MGCs) at tissue/material interfaces. The
latter separate the FBR from granulation tissue, which is characterized by the
presence of macrophages, fibroblasts, and capillaries, and which represents a
precursor to fibrous capsule formation. Two types of MGCs have been described
and are known as Langhans-type cells and foreign body giant cells (FBGCs). Cells
of the Langhans-type are thought to be characteristic for sarcoidosis and
tuberculosis and feature up to 20 nuclei in their periphery, which are arranged in a
circular or semi-circular “horse-shoe” pattern. FBGCs, on the other hand, have been
shown to form in the presence of foreign bodies like medical devices and can contain
tens to hundreds nuclei, which are irregularly distributed. Both types, however, are
formed from blood monocytes. While it has been suggested that these variants of
MGCs are identical except for their nuclei arrangement, it has more recently been
stated that they fulfill different functions. Furthermore, it was demonstrated that their
formation is induced by differential cytokine signals (McNally and Anderson, 2011;
Okamoto et al., 2003).
Following medical device implantation, an infection is likely, if the acute and chronic
inflammation persists for more than three weeks. Depending on the surface
topography of the implanted material, the degree of the FBR varies, with rough
surfaces provoking a broader reaction. Regarding formation of FBGCs it is thought
that they originate from biomaterial surface adherent macrophages that undergo
fusion (Fig.5). Although they might continue to surround the device for the duration
of implantation, it is not known whether FBGCs remain activated or not. Fusion,
however, is reported to be, amongst others, mediated via mannose receptors
following an up-regulation that is initiated by Interleukin (IL)-4 and IL-13. Usually
macrophages are able to phagocytize small particles (< 5 µm) while larger particles
result in the formation of MGCs (Anderson, 2001; Anderson et al., 2008; Brodbeck
and Anderson, 2009; McNally and Anderson, 2011).

Introduction
- 24 -
Figure 5: Formation of foreign body giants cells (FBGCs). Blood-derived monocytes transform to biomaterial adherent monocytes/macrophages and ultimately to FBGCs at the tissue/biomaterial interface. A group of adhesion receptors (integrins) enables the binding to certain blood proteins that have been adsorbed to biomaterials as well as to ECM molecules such as laminin. Adhesion success is thought to be necessary for FBGC formation (Anderson, 2001). The figure was made based on illustrations of (Anderson, 2001; McNally and Anderson, 2011).
1.8 The sciatic nerve model The rat sciatic nerve model is frequently used to determine the experimental
outcome of peripheral nerve reconstruction (Angius et al., 2012; Swett et al., 1991).
This mixed nerve is formed by spinal nerves L3-L6 and contains sensory and motor
axons. Accordingly, it is involved in movement, proprioreception, skin sensation as
well as smooth muscle functions (Swett et al., 1986; Wang et al., 2005). The ~2000
motoneurons and ~11,000 sensory neurons that form the sciatic nerve mainly reside
in L4 and L5 DRGs (only some are found in L3 and even less in L6). The main
branches of the sciatic nerve include the tibial nerve, common peroneal nerve, and
caudal sural cutaneous nerve (Fig. 6). 96 % of the axons of the sural nerve originate
in DRGs, while only 4 % belong to motoneurons. The common peroneal nerve is the
second largest branch of the sciatic nerve containing a third of all motoneurons
found in the sciatic nerve as well as mainly large diameter sensory neurons. The
largest branch, however, is the tibial nerve, which holds about half of the
motoneurons of the sciatic nerve and also most sensory neurons (~ 4700) (Maciel et
al., 2013; Pardo et al., 2012; Rigaud et al., 2008; Swett et al., 1991, 1986; Ueda,
2006; Wang et al., 2005).

Introduction
- 25 -
Figure 6: The sciatic nerve and its innervation targets. The sciatic nerve contains axons from motoneurons and sensory neurons and is formed mainly by spinal nerves L3-L6. It runs parallel to the femur and splits into three main branches: the tibial nerve, common peroneal nerve, and caudal sural cutaneous nerve. The deep peroneal nerve, a branch from the common peroneal nerve, innervates, amongst other, the tibialis anterior (TA) muscle, while the lateral plantar branch of the tibial nerve supplies the short flexor muscles of digits (plantar (PL) muscles). Innervation of the lateral plantar surface occurs via the caudal sural cutaneous nerve and lateral plantar nerve, while the medial plantar surface is innervated via a branch from the saphenous nerve and a branch from the medial plantar nerve (Gelderd and Chopin, 1977; Pardo et al., 2012; Popesko et al., 1990; Swett et al., 1986).
Evaluation of muscle reinnervation following nerve repair in this model can be
evaluated with regard to motor recovery via electrophysiological recordings of
compound muscle action potentials (CMAPs). This method allows, amongst other,

Introduction
- 26 -
the determination of the NCV. Recordings are performed from target muscles of the
sciatic nerve like the gastrocnemius muscle, tibialis anterior (TA) (synonym in
animals: cranial tibial) muscle, and short flexor muscles of digits, which is here
referred to as plantar muscle (PL). The branch that innervates the TA muscle is the
deep peroneal nerve, which arises from the common peroneal nerve, while
innervation of the PL muscle occurs via the muscular branches of the tibial nerve
(Nickel et al., 2004; Popesko et al., 1990). Another widely used method to assess
muscle reinnervation is the Static Sciatic Index (SSI), which is calculated by
measuring the distances between toes. Following sciatic nerve injuries, the toe
positions are altered due to the missing innervation of the muscles (Bervar, 2000).
The sciatic nerve model is further suitable to evaluate sensory recovery using
mechanical or thermal algesimeter tests (Cobianchi et al., 2013; Popesko et al.,
1990).
1.9 Aim of the study Although, ANGs continue to be the clinical standard therapy to bridge peripheral
nerve defects their usage comes with disadvantages (donor site morbidity, second
surgery, time consuming, and mismatch issues). Various nerve guidance conduits
have already been approved to be officially marketed, but none of the products fulfils
the requirements to prevail on the market. Therefore, alternative materials are
needed that facilitate peripheral nerve regeneration and keep the FBR minimal.
Addressing this issue, hollow chitosan conduits of 3 different DAs were tested in a
10 mm rat sciatic nerve model regarding their possible usage to reconstruct small
nerve defects (e.g. digital nerve defects). Short-term studies over 5 and 18 days
were performed as well as a long-term study (13 weeks) and analyzed with regard to
various parameters. Following an implantation period of 18 days and 13 week, the
inflammatory response was assessed through an (immuno-) histological analysis of
the fibrous capsule that had formed around the conduits as well as of nerve tissue.
Moreover, the physiological regulation of regeneration-associated proteins in DRGs,
nerve stumps, and the lumbar spinal cord region was analyzed by quantitative
(q)RT-PCR as part of the short-term studies. Furthermore, the functional recovery
was investigated via electrophysiological recordings and determination of the SSI in

Introduction
- 27 -
course of the long-term study. Mechanical tube properties were further noted for all
studies to ensure stability of the conduits.
Since treatment of long nerve defects (medial-, and ulnar nerves, brachial plexus
injuries) is thought to require a 3D guidance structure within conduits that provides
support for migrating SCs and axonal sprouts a second study was conducted.
Chitosan conduits filled with enriched NVR-Gel containing genetically modified SCs
either over-expressing GDNF or FGF-218kDa, were tested in a 15 mm rat sciatic nerve
model for a time period of 17 weeks. The functional outcome was determined
through electrophysiological recordings and SSI, while sensory recovery was further
assessed by determination of the mechanical pain threshold. Additionally, the
inflammatory response after an implantation time of 17 weeks was analyzed.

- 28 -

Materials
- 29 -
2. Material Materials Manufacturer 1 ml single-use syringe Injekt® Braun, Germany 5 ml single-use syringe Injekt® Braun, Germany Adson forceps World Precision Instruments, Germany Anatomical forceps World Precision Instruments, Germany AxioVision Zeiss, Germany Bipolar steel hook electrode Medizintechnik MHH, Germany
Chitosan hollow tubes DAI-III Medovent GmbH, Germany & Altakitin S.A, Portugal
Cover slip (Superfrost Plus®) Thermo Scientific, Gerhard Menzel GmbH, Germany
Dumont forceps #5 World Precision Instruments, Germany Dumont forceps #55 World Precision Instruments, Germany Ethilon 4-0 Ethicon®, Belgium Ethilon 9-0 Ethicon®, USA
Glass slide (Menzel-Gläser) Thermo Scientific, Gerhard Menzel GmbH, Germany
Iris scissors World Precision Instruments, Germany iScript Kit BioRad, Germany Micro scissor World Precision Instruments, Germany Mowiol® Calbiochem, Germany Needle electrodes Natus Europe GmbH, Germany Nitrocellulose membrane (Amersham Hybond ECL) GE Healthcare, UK NVR Gel N.V.R. LABS LTD., Israel Raucodrape® Lohmann & Rauscher, Germany Photoshop® CS2 Adobe® Systems Inc., USA Polysorb™ 3-0 Syneture™ Tyco Healthcare, USA Rats Janvier, France Razor Kai Co., LTD, Japan Rneasy Plus Micro Kit 50 Quiagen®, Germany Scalpel No 21 Feather® Safety Razor Co. LTD., Japan Thermostatic blanket (Bosotherm 1500) Bosch & Sohn GmbH, Germany Canon MV 20i, Digital video camcorder Canon, Japan
Instruments & Software Manufacturer Electronic von Frey anesthesiometer IITC Life Sciences, USA CellP software Olympus, Germany CellSense dimensions software Olympus, Germany CellSense entry software Olympus, Germany Citadel tissue processor 2000 Thermo Scientific, USA GraphPad PRISM® 6.0 GraphPad Software, Inc., USA ImageJ software National Institutes of Health, USA Microscope (Olympus IX70) Olympus, Germany Microscope (Olympus BX51) Olympus, Germany

Materials
- 30 -
Microscope (Olympus BX53) Olympus, Germany Microtome RM 2155 Leica, Germany StepOnePlus instrument Applied Biosystems®, Germany StepOne™ software Applied Biosystems®, Germany Toshiba Tecra 8200 Toshiba, Japan
Portable electrodiagnostic device Medtronic functional diagnostics A/S, Denmark
Antibodies Manufacturer Dilution Western Blot α-BDNF Santa Cruz, Germany (sc-546) 1:1000 Primary antibodies - Immunhistochemistry
α-NF200 Sigma-Aldrich®, Germany (N4142) 1:500
α-ED1 Serotec, UK (MCA275R) 1:1000 α-GAP43 Millipore™, Germany (AB5220) 1:200 Secondary antibodies - Immunhistochemistry ALEXA 488 α-rabbit Invitrogen, Germany (A11034) 1:1000 ALEXA 555 α-mouse Invitrogen, Germany (A21422) 1:1000
Chemical Manufacturer 4', 6-diamidin-2-phenylindol (DAPI) Sigma-Aldrich®, Germany (1:2000) Acetic acid Merck, Germany Acetic fuchsin Merck, Germany Amitripyline hydrochloride (Amitriptylin®) Neuraxpharm, Germany Bovine serum albumin (BSA) PAA, Germany Bepanthen® Bayer Vital GmbH, Germany Brain derived nuerotrophic factor (BDNF) PeproTech Inc, USA Braunol® B. Braun Meisungen AG, Germany Buprenorphine (Buprenovet®) Bayer Vital GmbH, Germany Butorphanol tartrate (Torbugesic®) Pfitzer GmbH, Germany Chloral hydrate Sigma-Aldrich®, Germany Sodium citrate tribasic dihydrate (0.01M, pH 6.0) Sigma-Aldrich®, Germany Corbit-balsam Hecht, Germany Distilled water (H2O) Millipore™, Germany Eosin Roth®, Germany Ethanol Mallinckrodt Baker, Germany Hematoxylin Roth®, Germany
Light green Chroma Gesellschaft, Schmidt & Co., Germany
Liquid nitrogen Linde Gas Therapeutics, Germany Milk powder Heirler, Germany Normal goat serum (NGS) Gibco, Germany

Materials
- 31 -
Orcein Merck, Germany Paraformaldehyd (PFA) Sigma-Aldrich®, Germany Phosphate buffered saline (PBS) Biochrom, Germany Rabbit serum Sigma-Aldrich®, Germany Sodium chloride Braun, Germany Sodium dodecyl sulfate (SDS) Roth®, Germany SYBR-Green PCR Master Mix Applied Biosystems®, Germany Triton-X-100 Roche, Germany Wolframato phosphoric acid Merck, Germany

- 32 -

Methods
- 33 -
3. Methods 3.1 Animals All animal experiments followed the German Animal Protection Act and were
approved by the local animal protection committee (Bezirksregierung LAVES,
Hannover, Germany, AZ 33.12-4202-04-12/0816). Adult female Wistar rats
(220-250 g) were obtained from Janvier (France) and housed in groups of 4 under
standard conditions (room temperature (RT) 22 ± 2 °C; humidity 55 ± 5 %; 14 h
light/10 h dark cycle). Food and water was available ad libitum for the complete
duration of the experiments.
3.2 Experimental design and groups Hollow chitosan conduits (inner diameter: 2.1 mm, wall thickness: 0.3 mm) of
different DAs were tested in short and long-term studies (project 1 / 10 mm gap).
Furthermore, a second long-term study was performed with chitosan conduits
containing enriched NVR-Gel. This gel is comprised of extracellular matrix
components such as HA (high molecular weight, 1 %) and laminin 1 (10 ng/ml) and
was used as a matrix to deliver genetically modified SCs, either overexpressing
GDNF or FGF-218kDa, to the injury site (Rochkind et al., 2009; Shahar et al., 2001)
(project 2 / 15 mm gap). The experimental procedures for both projects are
separately described in the following text.
3.2.1-a Project 1-Short-term study A total of 36 animals out of 44 were subjected to sciatic nerve surgery and a 10 mm
gap was introduced. Subsequently, nerve reconstruction was performed with hollow
chitosan conduits of a low (2 %), medium (5 %), or high (20 %) DA (n = 12,
respectively). The remaining eight animals served as controls. Following an
implantation time of 18 days, the fibrous capsule that had developed around the
tubes as well as the proximal tissue were harvested and embedded in Paraffin for
(immuno-) histological analyses. Furthermore, conspicuous features of the conduits
upon explantation were noted regarding their mechanical properties. Additionally,
DRGs, nerve stumps, and the lumbar spinal cord region were harvested at both time
points and snap frozen for qRT-PCR, while the tube contents were collected and
used for Western Blot analysis (Tab. 1).

Methods
- 34 -
Table 1: Overview of the collected tissue samples in the short-term study of project 1. n.a. = non analyzed; n.av. = not available
3.2.1-b Project 1-Long-term study Sixty-eight rats were included in this study and a 10 mm gap was introduced into the
sciatic nerve of all animals. Overall 51 of these defects were treated with hollow
chitosan conduits of the three different DAs (see 3.2.1-a), while ANGs were used to
reconnect the two nerve stumps in the remaining cases (n = 17, respectively) (Fig. 7). Tests for functional evaluation of the nerve reconstruction were performed
4, 9, 12, and 13 weeks after surgery (SSI, electrodiagnostic recordings). On the final
day of observation, animals were sacrificed and the regenerated nerve tissue and
fibrous capsules were harvested for (immuno-) histological and morphometric
analyses. Additionally, the muscle weight of the TA muscle and PL muscle was
determined as were the mechanical tube properties upon explantation.

Methods
- 35 -
Figure 7: Experimental design for the long-term study (13 weeks) using hollow chitosan conduits in a 10 mm gap (project 1). (A) Illustration of nerve reconstruction with hollow chitosan conduits in the 10 mm rat sciatic nerve model. (B-D) Schematic illustration of the three experimental groups that were treated with chitosan conduits of different DAs (DAI = 2%, DAII = 5%, DAIII = 20%) after introduction of 10 mm gap into the sciatic nerve as well as of (E) the control group that received reversed ANGs.
3.2.2 Project 2 For the second project a 15 mm, gap was introduced into the sciatic nerve of 40
female Wistar rats and the nerve was reconstructed using chitosan conduits
enriched with NVR-Gel containing 1) non-transfected neonatal SCs (nt-neoSCs) 2)
SCs that were transfected with 3) an empty vector (tf-SCs-EV), 3) GDNF (tf-neoSCs-
GDNF), or 4) FGF-218kDa (tf-neoSCs-FGF-218kDa) (n = 8, respectively). The
preparation of the conduits was performed by Sandra Wrobel (MSc, Medizinisiche
Hochschule Hannover (MHH) Neuroanatomy), and is described in detail in her PhD-
thesis. In short, three days prior to transplantation 2.5 x 106 neoSCs (neoSCs; 8.-9
passage; purity: 75-90 %) were genetically modified by AMAXA nucleofection
(transfection rate: 73-80 %). Afterwards, the cells were allowed to recover in culture
before a total of 1 x 106 t-SCs as well as nt-SCs were mixed with 0.5 % NVR-Gel
(110 µl 1st series and 130 µl 2nd series) and placed inside each chitosan conduit.
24 h following conduit preparation, surgery was performed and the conduits were
used to bridge the nerve defects. Over-expression of GDNF and FGF-218kDa was
confirmed by Western Blot analyses by (see Sandra Wrobel, PhD-thesis). In control
animals the nerve defect was bridged with 5) ANGs (n = 8) (Fig. 8). Tests regarding

Methods
- 36 -
sensory and motor recovery were performed 4, 9, 12, and 16 weeks after surgery by
means of SSI and mechanical pain threshold assessment. Furthermore,
electrodiagnostic measurements were performed following an implantation time of 9,
12, and 16 weeks. On the final day of observation, animals were sacrificed and the
regenerated nerve tissues (if regeneration was successful) as well as the fibrous
capsule that had formed around the conduits were harvested for (immuno-)
histological analysis. In cases of failed or thin tissue regeneration proximal nerve
stumps or total samples (nerve stumps + conduit (+ fibrous capsule)) were
embedded. Additionally, the muscle weight of the TA muscle and gastrocnemius
muscle was determined as were the mechanical tube properties upon explantation.
Schematic illustration of the four experimental groups that were treated with chitosan conduits (DA = 5 %) enriched with NVR-Gel containing either (A) nt-neoSCs, SCs that were transfected with (B) an EV, (C) GDNF, or (D) FGF-218kDa. (E) ANGs served as controls.
3.4 Anesthesia and surgical procedures During surgery and electrodiagnostic recordings, the rats were in general
anesthetized by an i.p. injection of chloral hydrate (370 mg/kg) and intramuscularly
(i.m.) applied buprenorphine (0.045 mg/kg, Buprenovet®) (project 1 & 2). On the two
days following the initial surgery, the animals also received butorphanol tartrate
(0.5 mg/kg, Torbugesic ®) via subcutaneous injection (s.c.) as pain relief (project
Figure 8: Experimental design for the long-term study (17 weeks) using enriched tubes in a 15 mmgap (project 2).

Methods
- 37 -
1-short-term study and project 2). Additionally, amitriptyline hydrochloride
(9 mg/kg/day, Amitriptylin®) was administered through the drinking water, starting
two weeks prior to the surgery and was given for the time period of the experiments
to minimize autotomy events (project 2) (Navarro et al., 1994).
Animals were prepared for surgery by shaving the left hind leg and disinfecting the
skin. During surgery, to keep the body temperature stable, the rats were placed on a
thermostatic blanket. Aseptic techniques were further used to ensure sterility. A skin
incision was made with a surgical blade along the femur, and the gluteus and biceps
femoris muscles were separated by sharp and blunt dissection to expose the sciatic
nerve. The wound was kept open with the help of small retractors. The sciatic nerve,
exposed at the mid-thigh level, was transected with a single micro scissors cut at a
constant point (6 mm from the exit of the gluteus muscle). For tube transplantation,
5 mm of the sciatic nerve end were removed and 14 (Fig. 9) or 19 mm long conduits
were used to bridge the nerve defects leaving a 10 mm (project 1) or 15 mm gap
(project 2) between the two nerve stumps, respectively. Control animals of the
long-term studies received reversed ANGs of 10 (projects 1) or 15 mm (project 2)
length. Therefore, a sciatic nerve piece of the desired length was cut out, reversed
(distal-proximal), and flipped 90° before 3 sutures (9-0) were placed 120° apart from
each other. In control animals of the 5 and 18 days short-term studies (project 1), the
nerve defect was not bridged, but the proximal and distal nerve ends were folded
over and sutured to themselves after removal of the 5 mm nerve piece (Haastert-
Talini et al., 2013). The critical steps of the surgeries were conducted by Prof.
Haastert-Talini (MHH Neuroanatomy), while it was my responsibility to expose the
sciatic nerve and close the wound. Jennifer Metzen (biology lab assistant, MHH
Neuroanatomy), was further in charge of all aspects regarding anesthesia.

Methods
- 38 -
Figure 9: Chitosan conduit implantation. Representative photograph showing a hollow chitosan conduit bridging a 10 mm sciatic nerve defect.
3.5. Functional evaluation of nerve regeneration. During the observation periods of both long-term studies (project 1 and 2), the
progress of functional recovery was monitored by means of SSI, electrophysiological
recordings (serial non-invasive and final invasive), and calculation of the lower limb
muscle weight ratio. Additionally, in the scope of project 2, the von Frey test was
performed to assess sensory recovery, while cases of autotomy were noted using an
autotomy score. The investigators were blinded to the conditions of sciatic nerve
reconstruction applied to the animals.
3.5.1 Static Sciatic Index (SSI) To assess motor recovery after nerve repair the SSI was determined. This index is
based on changes in the hind paw posture that occur following sciatic nerve injury
due to missing innervation. In case of successful regeneration changes of the
posture gradually decrease. For project 1 SSI analysis was conducted before
surgery and during weeks 1, 4, 9, and 12 following surgery. In the course of project
2, however, this test was additionally performed 16 after nerve repair.
For this test, a plastic box was placed on a glass table and rats were transferred
inside the box (Fig. 10). A webcam (Canon MV 20i) placed under the glass table and
a notebook (Toshiba Tecra 8200) allowed the image acquisition of the paw positions.
These images were then exported to a freely available image-editing program
(AxioVision, Zeiss, Jena, Germany). The distance between toes 1 and 5 (toe spread,
TS) as well as toes 2 and 4 (intermediate toe spread, ITS) was measured for the
lesioned (L) and non-lesioned (N) hind paws. The SSI was then calculated using the

Methods
- 39 -
following formulas, which were introduced by Bervar (Bervar, 2005, 2000) (TSF = toe
spread factor; ITSF = intermediate toe spread factor):
TSF= (L-TS - N-TS) / N-TS
ITSF= (L-ITS - N-ITS) / N-ITS
SSI= (108.44 * TSF) + (31.85 * ITSF) - 5.49
The setup for the SSI includes (A) a webcam placed under a glass table, on which the animals are positioned so that (B) images can be taken of the paw positions. For calculation of the SSI, the toe spread (TS, marked in pink) and intermediate toe spread (ITS, marked in green) are measured. Animals that displayed autotomy were not included in the test and account for changes in numbers of evaluated animals.
3.5.2 Von Frey test An electronic von Frey algesimeter was used to assess the mechanical pain
threshold to determine sensory recovery following nerve reconstruction. Therefore,
plastic compartments were positioned on a metallic mesh stand and animals were
placed inside the boxes and allowed to acclimate to the surrounding (15 minutes)
(Fig. 11). Following habituation, a blunted needle connected to a force sensor in a
hand-held probe was used to stimulate the medial and lateral site in the intact and
injured paw. Pressure was increased gradually and in a slow manner until the animal
withdrew the tested paw. The applied force needed to induce paw lifting was
Figure 10: Static SciaticIndex (SSI).

Methods
- 40 -
displayed as the weight-force (in grams). A cut-off value was set to 40 g, when it was
observed that the paw was lifted simply by the hand-held probe and not as a result
from an active response from the animal or in cases where no paw lifting at all was
seen. The interval between the stimuli was at least 3 min. The paw withdrawal
pressure was recorded five times for each site and the three steadiest values were
used for calculation of a mean that was then used for statistical analysis. The test
was performed three times before surgery for habituation and then in weeks 1, 4, 9,
13, and 16 following surgery.
Figure 11: Von Frey. Sensory recovery was assessed under usage of the von Frey algesimeter. (A) The plastic chambers, in which the animals were placed and allowed to habituate for 15-30 minutes. Following habituation, a B) hand held probe that consists of a force sensor and a blunted needle was used to stimulate the lateral and medial side of the paws, which can be seen through the (C) holey metal plate.
3.5.3 Electrophysiology 3.5.3.1 Serial non-invasive electrodiagnostic recordings
For evaluation of ongoing muscle reinnervation in the long-term studies, a portable
electrodiagnostic device (Keypoint Portable) was used and recordings were
performed in weeks 4, 9, and 13 for project 1 and in weeks 9, 12, and 16 for project
2. Therefore, animals were anesthetized as described above and fixated with the
hind limbs extended on a metal plate on top of a thermostatic blanket. Only during
recordings, the thermostatic blanket was turned off to avoid interfering signals. The
sciatic nerve was stimulated with single electrical pulses (100 μs duration and
supramaximal intensity) delivered by monopolar needles (30 G, diameter 0.3 mm,
length 10 mm), which were percutaneously placed either at the sciatic notch, thus

Methods
- 41 -
proximal to the injury (proximal stimulation: cathode directly at sciatic notch, anode
1 cm more proximal) or in the popliteal fossa (distal stimulation) (Fig. 12). The
CMAPs of the TA and PL muscles were recorded by means of another pair of
monopolar needles inserted across the skin on the muscle belly. To ensure
reproducibility, the recording needles were placed with the help of anatomical
landmarks to secure the same placement on all animals. The active recording
electrode was placed subcutaneously at the first third of the distance between knee
and ankle on the TA muscle, or at the third metatarsal space for PL muscle
recordings. The reference needle electrode was inserted at the distal phalange of the
second or fourth toe. A ground needle electrode was inserted into the skin. Taking
into account the latency difference of the two stimulation points and the distance
between them, the electrodiagnostic device calculated automatically the NCV
(Haastert-Talini et al., 2013). For each animal, four recordings per time point and
recording site (TA, PL muscles of both body sides) were performed and means were
used for further calculations. A NCV-ratio was calculated according to the following
equation (L = recorded values at one time point on the lesioned site, NM = mean of all
recorded values from the non-lesioned site obtained during the complete follow-up
period):
NCV-ratio = L/NM
Furthermore, the amplitude and the area under the curve (AUC) of the negative
CMAP peaks were analyzed. For evaluation of the amplitude, a ratio was calculated
according to the concept described for the NCV. AUC-values of the CMAPs were
used to calculate the percentage of axon loss (AxL) according to the following
equation:
AxL [%] = NM-L/NM x 100

Methods
- 42 -
Figure 12: Serial non-invasive electrodiagnostic recordings. To evaluate the functional outcome electrophysiological recordings from the TA and PL muscle were conducted. (A) For non-invasive recordings, grounding, reference, and stimulation electrodes were placed at defined landmarks. Furthermore, recording electrodes were positioned either on the muscle belly of the (B) TA or (C) PL muscle.
Regarding recordings from the TA muscle, it is important to note that potential
artifacts from neighboring muscles were seen and early regeneration signals partly
fused together with these artifacts. If the correct signal was not clearly identifiable, it
was not analyzed. The recordings were performed together with Prof. Haastert-Talini
(MHH Neuroanatomy).
3.5.3.2 Final invasive recordings
On the last day of observation, animals of the long-term studies were once more
anesthetized and the sciatic nerve was exposed consecutively on the lesioned and
non-lesioned side. The recording electrodes were placed as described above. Using
a bipolar steel hook electrode the sciatic nerves were directly stimulated proximally
and distally to the transplants. Single electrical pulses (100 μs duration) with

Methods
- 43 -
gradually increased intensity (not exceeding 8 mA) were applied in order to evaluate
the maximal CMAP. The current required to elicit the maximal CMAP was also
determined as was the latency of these signals. Furthermore, the NCV as well as
AUC were calculated by stimulating the sciatic nerve, proximal and distal to the
injury, with a current intensity of 1 mA (Haastert-Talini et al., 2013). The NCV-ratio,
AxL, and amplitude-ratio were then calculated using the equations noted under
3.5.3.1, but only values obtained during the invasive recordings were taken into
account for calculations.
3.5.4 Muscle weight After completing the long-term experiments, the animals were sacrificed and the
gastrocnemius and TA muscle of both hind limbs were excised and weighted in order
to determine the muscle weight ratio ([g]ipsilateral/[g]contralateral) (Haastert-Talini et
al., 2013).
3.5.5 Autotomy score Events of autotomy were reported using an autotomy score based on a point-
system. As part of this system one point per automutilation site (nails, distal
phalanges, proximal phalanges, and plantar pads) was counted (Fig. 13).

Methods
- 44 -
Figure 13: Autotomy Score. (A-B) Nails, the proximal and distal phalanges from the animals toes (U1-U5) as well as the plantar pads (indicated by asterisks) were taken into account for the determination of the autotomy score.
3.6 (Immuno-) histological evaluation of nerve regeneration 3.6.1 Assessment of nerve tissue 3.6.1.1 Project 1-short-term study
After an implantation time of 18 days, the transplants were harvested together with
the matrix that had formed inside the tube. While the tube content was used for
Western Blot analysis (see 3.7.2), the proximal nerve stumps were fixated in 4 %
paraformaldehyde (PFA) overnight at 4°C. Afterwards, samples were
paraffin-embedded with the help of a Citadel tissue-embedding-automatic unit and
serial 7 µm cross-sections were prepared (Fig. 14). Blind-coded sample sections
(from 3 randomly chosen animals per conduit group as well as from 2 random control
animals), taken from the distal end and approximately 390 µm more proximal, were
subjected to hematoxylin-eosin (HE)-staining in order to determine the number of
MGCs per mm2. Therefore, sections were examined manually under light microscopy
(Olympus BX53) and overview photomicrographs were used for measurements of
the area.

Methods
- 45 -
Figure 14: Processing of the nerve tissue gathered 5 and 18 days after surgery (project 1). After 5 and 18 days, a matrix had formed inside the tube, but no regenerated nerve cable was found. Therefore, the proximal nerve stump was paraffin-embedded and 7 µm sections were prepared for (immuno-) histological analyses.
Additional blind coded sample sections (from the same animals), taken at defined
distances in distal-proximal direction (0 µm, 72 µm, 152 µm, 320 µm, 392 µm and
472 µm), were stained for activated macrophages (ED1) and GAP-43. Following a
pre-incubation step in citrate buffer (heating until boiling then 9 min at 95°C and
30 min at RT), the sections were incubated in phosphate buffered saline (PBS) +
5 % rabbit serum as a blocking step prior to incubation with primary mouse α-ED1
antibody (1:1000, in blocking) at 4°C overnight. The next day, sections were washed
with PBS (3x) before being incubated with ALEXA 555-conjugated secondary goat
α-mouse antibody (1:1000, in blocking) for 1h at RT. A second blocking step in 0.3 %
Triton + 3 % normal goat serum (NGS) followed before rabbit α-GAP43 (1:200; in
blocking) was applied overnight at 4°C. As a next step, the sections were washed in
PBS (3x) and incubated with ALEXA 488-conjugated goat α-rabbit antibody (1:1000,
in blocking) for 1h at RT. The sections were finally counterstained with the nuclear
dye 4’, 6’-diamino-2-phenylindole (DAPI, 1:2000 in PBS) in PBS for 2 min at RT and
then mounted with Mowiol. For quantification of the number of activated ED1+
macrophages/mm2, seven photomicrographs per section were randomly taken (20x
magnification) almost covering the whole cross section and counting was performed
using ImageJ software.

Methods
- 46 -
3.6.1.2 Project 1-long-term study
On the last day of observation, the transplants with the adjacent nerve tissue were
harvested and the regenerated nerve tissue was extracted from the chitosan tubes.
Afterwards, the regenerated nerve tissue was separated into a proximal and a distal
part by a cut 1 mm distal to the distal suture (Fig. 15). Five mm of the distal segment
were then prepared for the stereological analysis and shipped to our partner
laboratory at the University of Turin (UNITO) (for methods and results see (Haastert-
Talini et al., 2013). The proximal regenerated nerve tissue was subjected to fixation
in 4 % PFA overnight at 4°C. Next, paraffin-embedding was performed in a Citadel
tissue-embedding-automatic unit and serial 7 µm cross-sections were prepared.
Two blind-coded sample sections (from randomly chosen animals; n = 5,
respectively) taken from the distal end as well as 320 µm more proximal were
subjected to HE-staining in order to examine the number of MGCs per mm2.
Sections were examined manually under light microscopy (Olympus BX53).
Overview photomicrographs were used for measurements of the area.
Figure 15: Processing of the regenerated nerve tissue 13 weeks after implantation (project 1). 13 weeks after surgery, the regenerated nerve tissue was exposed and prepared for further analysis. Therefore, a cut was made 1 mm behind the distal suture and the proximal part was embedded in paraffin for (immuno-) histological analysis, while the distal part was sent to our partner laboratory (UNITO, Italy) for morphometric evaluation. From the proximal nerve tissue serial 7 µm sections were prepared (in distal to proximal direction, covering a distance of approximately 1000 µm) and subjected to different stainings.
Additional blind-coded sample sections (from randomly chosen animals; n = 8,
respectively), taken at defined distances in distal-proximal direction, were subjected

Methods
- 47 -
to double-immunohistochemistry (IHC) for activated macrophages (ED1) and
neurofilament (NF) 200. Quantification of the number of activated macrophages/mm2
was performed by Sandra Wrobel (MSc, MHH Neuroanatomy) (for methods and
results see (Haastert-Talini et al., 2013)).
3.6.1.3 Project 2
Following an implantation time of 17 weeks, the transplants were exposed and the
nerve tissue was cut 2 mm distal to the distal suture site, thus leaving a proximal as
well as distal segment. In accordance to project 1, 5 mm of the distal segment were
prepared for morphometric analysis and sent to our partner laboratory (UNITO). In
case of clear tissue regeneration, the chitosan scaffold was carefully removed and
the proximal tissue sample was subjected to fixation in 4% PFA (overnight at 4°) for
subsequent paraffin-embedding and (immuno-) histological analysis. Furthermore, in
case of failed regeneration single proximal nerve stumps were prepared for
histological analysis. Samples displaying thin regeneration as well as one sample of
successful regeneration, and representative examples of failed regeneration were
embedded together with the chitosan conduit, distal nerve stump, and, if applicable,
together with the surrounding connective tissue.
Seven µm cross-sections were prepared from samples showing clear tissue
regeneration as well as of representative proximal nerve stumps gained from cases
of failed regeneration. The sectioning was undertaken in distal-proximal direction as
described for project 1 and covered a distance of approximately 600 µm. After
discarding a 1.5 mm piece of the sample a second series of 600 µm was cut and
gathered for histological analysis. Two sections of each series were HE-stained and
evaluated with regard to the number of MGCs, tissue thickness, and area as
described for project 1 (long-term study). Adjacent sections were subjected to
ED1-staining, to enable counting of activated macrophages, and NF200 staining.
ED1-staining staining, however, was performed according to a new staining protocol
compared to project 1. Therefore, paraffin cross-sections were incubated in PBS with
3 % milk powder and 0.5 % Triton X 100 for blocking before incubation with primary
mouse α-ED1 antibody (1:1000, in blocking solution) at 4°C overnight. Following
three washing steps in PBS, incubation with Alexa 555-conjugated secondary goat
α-mouse antibody (1:1000, in blocking) for 1h at RT was performed. Afterwards, a

Methods
- 48 -
second blocking step with PBS containing 3 % milk powder and 0.5 % triton X-100
and subsequent overnight incubation with primary rabbit α-NF200 antibody was
conducted. Next, three washing steps followed, before incubation with Alexa
488-conjugated secondary goat α-rabbit antibody (1:1000, in blocking) for 1h at RT
and counterstaining with DAPI (1:2000, in PBS) was performed. Finally, sections
were mounted using Mowiol. Five images (20x magnification) were acquired per
section or less if the whole section was already covered and counting of activated
macrophages was performed with the help of ImageJ software by Natalie Schecker
(internship, MHH Neuroanatomy).
Exemplary samples of thin as well as failed tissue regeneration were cut in the
middle of the tube and sections were gathered covering a distance of about 300 µm.
Staining for activated macrophages and axons was performed as described above.
3.6.2 Assessment of fibrous capsule Eighteen days and 13 weeks (project 1) as well as 17 weeks (project 2) after
surgery, a fibrous capsule / connective tissue had formed around the conduit, which
was macroscopically and histologically analyzed. For the macroscopic evaluation,
the tissue was dissected, spread on a scale paper glass plate, and photographs
were taken of every sample. From the photographs, three classes of macroscopic
connective tissue optical density were defined as translucent, translucent-opaque,
and opaque (only project 1). Afterwards, samples of the connective tissue (n = 4-6)
were folded and sutured at the proximal end so that the chitosan conduit contact
surfaces were facing each other. The tissue was then fixed overnight in 4 % PFA
and paraffin-embedded. For histological analysis, blind-coded 7 µm paraffin sections
were cut from the distal end of the connective tissue and subjected to HE-, Trichrom-
, or Orcein-staining as well as to IHC for activated macrophages.
3.6.2.1 Project 1
In 4-6 HE-stained sections per sample, covering a distal-proximal distance of about
400 µm, the connective tissue area and thickness (at 4 randomly chosen points)
were determined. Light microscopic evaluation (Olympus BX53 and Olympus BX51)
was aided by the programs CellSense Dimension and CellSense Entry and overview
photomicrographs were used for measurements (Fig. 16). Furthermore, the number

Methods
- 49 -
of MGCs per mm2 within the former contact area with the chitosan conduits (18 days)
or in the whole section (13 weeks) was manually quantified. Trichrom- and
Orcein-stainings were performed to visualize the distribution of collagen and elastic
fibers, which are amongst others found in the wall of vessels (Ushiki, 2002). For the
Trichrom-staining, sections were subjected to hematoxylin, before being washed-
offed under running water. Afterwards, 20 ml solution containing acetic acid (1 ml in
99 ml aqua dest) and acetic fuchsin (0.5 g, in 100 ml 1 % acetic acid) were mixed
with 10 ml light green (1 g in 100 ml 1% acetic acid) and 10 ml wolframato
phosphoric acid (1 g in 100 ml H2O) and the slides subjected to this solution.
Following a washing step in aqua dest, the slides were subjected to 1 % acetic acid,
before being pressed with filter paper, dehydrated, and mounted with corbit-balsam.
For orcein-staining, slides were exposed to a staining solution (for 45 min) containing
orcein (1 g in 99 ml 70 % ethanol) mixed with 25 % hydrochloric acid. Following a
washing step in aqua dest the slides were placed in 96 %, 100 % ethanol, and xylol,
consecutively, before being mounted with corbit-balsam. Representative
photomicropgraphs were taken using the programs CellSense Dimension and
CellSense Entry
IHC was performed to estimate the number of ED1 stained cells (n = 4-5 / group).
Therefore, two blind-coded 7 µm paraffin cross sections, approximately 320 µm
apart from each other, were stained for ED1 as described above (see 3.6.1.1) and
after incubation with goat α-mouse antibody the sections were counterstained
directly with DAPI and mounted with Mowiol. Two photomicrographs each, were
taken from an area with minimal as well as maximal ED1 signal per section (20x
magnification) using an IX70 microscope and CellP software. The number of ED1+
cells was counted as number/mm2 with the help of ImageJ software. The nuclear
DAPI counterstain was used to clearly identify ED1+ cells (Haastert-Talini et al.,
2013).

Methods
- 50 -
Figure 16: Illustration of fibrous capsule analysis. 18 days and 13 weeks after implantation, connective tissue had developed around the implanted scaffolds. (A) The area of the connective tissue was determined in HE-stained 7 µm paraffin cross-sections. In these sections, the inner surfaces of the connective tissue (former contact area with chitosan conduits) are facing each other. (B) Furthermore, the thickness of the connective tissue was measured at 4 randomly chosen locations. (C, D) After 18 days, MGCs were further quantified within the contact area with the chitosan conduits, while for the long-term studies the whole tissue area was taken into account. Scale bars: 200 µm, 200 µm, 10 µm, a 5 µm.
3.6.2.2 Project 2
For the second project, 7 µm thick sections were cut from the distal end of the
samples and four sections within a distance of 2 cm were subjected to HE-staining
and analyzed with regard to number of MGCs, tissue thickness and area as
described above. Also, Trichrom- and Orcein-stainings were conducted according
the procedures noted for project 1.
Additional two sections obtained from the distal end, covering a distance of
approximately 300 µm, were stained for activated macrophages using staining
protocol stated under 3.6.1.3. Following the incubation step with Alexa 555
conjugated secondary goat α-mouse antibody (1:1000, in blocking) and subsequent
counterstaining for DAPI, however, samples were mounted with Mowiol. Four
photomicrographs per section were acquired (Olympus IX70) and cell counting was
performed as described before using ImageJ software as described for project 1.

Methods
- 51 -
3.7 Molecular and biochemical evaluation of nerve regeneration 3.7.1 Quantitative real-time polymerase chain reaction (qRT-PCR) The ipsi- and contralateral lumbar region of the spinal cord of animals subjected to
the 5 and 18 days observation period were dissected and snap frozen in liquid
nitrogen (5 days: DAI, DAII, DAIII n = 3 each, control n=4; 18 days: DAI + DAIII n = 3
each, DAII + control n=4 each). Tissue samples of each animal were homogenized
and total RNA was extracted according to the manufacturer’s instructions (RNeasy
Plus Micro Kit 50). The eluate of 14 μl was used completely for cDNA synthesis
using the iScript Kit (BioRad, Germany). Primer sequences were as previously
stated for BDNF, GAP-43, and NGF (Ratzka et al., 2012). Further primer sequences
were as follows: FGF-2-F: 5’-GAACCGGTACCTGGCTATGA-3’; FGF-2-R: 5’-
CCAGGCGTTCAAAGAAGAAA-3’; IL-6-F: 5’- CGTTTCTACCTGGAGTTTGTGAAG-
3’; IL-6-R: 5’- GGAAGTTGGGGTAGGAAGGAC-3’; tyrosine kinase receptor B
(TrkB)-F: 5’- CCCAATTGTGGTCTGCCG-3’; TrkB-R: 5’-
CTTCCCTTCCTCCACCGTG-3‘. Equal PCR efficiency was certified by serial cDNA
dilutions and estimated to be 100 %. qRT-PCR was performed with Power SYBR-
Green PCR Master Mix on a StepOnePlus instrument as described before (Ratzka et
al., 2012). Fold changes in mRNA levels are stated in comparison to the levels
measured on the respective day at the contralateral side of the control animals.
Calculation was done by using the 2(-∆∆Ct) method and normalized to the
housekeeping genes glyceraldehyde-3-phosphate dehydrogenase (Gapdh)-F: 5′-
GAACATCATCCCTGCATCCA-3′, Gapdha-R: 5′-CCAGTGAGCTTCCCGTTCA-3′;
and peptidylprolyl isomerase (Ppia)-F: 5’-TGTGCCAGGGTGGTGACTT-3’; Ppia-R:
5’- TCAAATTTCTCTCCGTAGATGGACTT-3’) (Haastert-Talini et al., 2013; Rumpel
et al., 2013).
Furthermore, DRGs (L3-L5) were gathered for qRT-PCR to investigate expression
pattern of the noted factors as well as of IL-6. This work was performed by Dr.
Andreas Ratzka (MHH Neuroanatomy) and methods as well as results can be
looked up in (Haastert-Talini et al., 2013). Also, proximal and distal nerve stumps
were collected, but processing of the samples was not successful likely due to an
insufficient material amount.

Methods
- 52 -
3.7.2 Western Blot Conduit contents were collected 5 and 18 days after nerve reconstruction and pooled
from 5-6 animals per DA, respectively, to verify the presence of BDNF. Therefore,
samples were homogenized in 40 μl Radioimmunoprecipitation assay (RIPA) buffer
according to (Ratzka et al., 2012). Samples were then boiled in Laemli buffer (5 min)
and separated by sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis
(PAGE) (12 % gel), before being transferred (50 μg) electrophoretically to
nitrocellulose membranes with 4 ng recombinant BDNF serving as positive control.
Membranes were then probed with α-BDNF antibody (1:1000) and analyzed
according to (Ratzka et al., 2012) (Haastert-Talini et al., 2013). This work was
performed by Kerstin Kuhlemann (VMTA, MHH Germany).
3.8 Analysis of tube properties after explantation On day 5 and 18 after nerve reconstruction (project 1 short-term study), the chitosan
tubes were explanted. Within 18 days, a fibrous capsule had developed, which was
macroscopically classified and processed for histological analysis (see 3.6.2). On
day 5 and day 18, representative conduits and their contents were stored for
Western Blot analysis of the content (see 3.7.2). Therefore, the middle parts of the
tubes were harvested by cutting of the proximal and distal end of the tubes at the
suture sites. Also, upon explantation in week 13 (long-term studies) representative
conduits were explanted, dissected from surrounding tissue, and stored in 0.9 %
NaCl solution for further analysis of biomaterial properties. The assets of the
conduits, the connective tissue as well as the macroscopic nerve cable peculiarities
were then noted and connective tissue samples were processed for histological
evaluation. Furthermore, tube samples (before and after nerve reconstruction) were
analyzed by Scanning electron microscopic or biochemical tests (n = 3 / per group)
in our partner laboratory at the University of Minho (UMINHO) (for methods and
results see (Haastert-Talini et al., 2013)). For project 2, again the conduit and
connective tissue properties were noted in the scope of the explantation procedure,
before being prepared for further analysis.

Methods
- 53 -
3.9 Statistical analysis The data obtained from the various functional and molecular tests as well as the
(immuno-) histological analysis were subjected to One-way or Two-way ANOVA
(Analysis of Variance) followed by Dunn’s or Tukey’s multiple comparison test
(GraphPad Prism 6). In general non-parametric statistical approaches were preferred
due to the significant deviation from normality. Furthermore, a Chi-square test was
performed, if applicable. Results are presented as means ± standard error of the
mean (SEM) and p-values were set to < 0.05, < 0.01, and < 0.001 as level of
significance.

- 54 -

Results-Project 1
- 55 -
4 Results-Project 1 4.1 Gene expression of regeneration-associated proteins qRT-PCR was performed to assure that chitosan does not influence the
physiological regulation of NTFs (NGF, FGF-2, BDNF), the cytokine IL-6, BDNF-
receptor TrkB, and GAP-43 following nerve lesion. Therefore, the lumbar region of
the spinal cord was explanted from representative animals of all experimental groups
5 and 18 days after nerve injury and mRNA was extracted. All data were normalized
to the values measured on the respective day for the contralateral nerve of the
control animals.
NGF and IL-6 expression was below detection limit and could not be quantified. No
significant up- or down-regulations of the additional analyzed factors were seen in
the ipsilateral spinal cord of all chitosan conduit grafted animals as well as the
animals of the control group, when compared to the expression levels seen in the
contralateral spinal cord of the control animals 5 and 18 days after surgery,
respectively (Fig. 17).
Figure 17: Regulation of mRNA levels in spinal cord. 5 and 18 days after conduit implantation or control treatment, the lumbar region of spinal cords were harvested and total mRNA was extracted. qRT-PCR was conducted to determine gene expression of representative NTFs and regeneration-associated proteins but did not reveal any significant differences between all experimental groups with regard to their regulation following nerve injury. Values were normalized to the non-lesioned site of the control group. DAI, DAII, DAIII n= 3 each; control n=4. Results were tested for significance (p < 0.05) using Two-way ANOVA.

Results-Project 1
- 56 -
In addition, there were no significant changes in the expression patterns of the
analyzed factors between day 5 and 18. Thus, it can be concluded that implanted
chitosan conduits do not influence the physiological regulation of NTFs following
nerve injury.
4.2 Matrix formation and presence of brain-derived neurotrophic factor (BDNF) Five days after implantation, the content of the tube appeared jellylike, while matrix
formation was seen after an observation period of 18 days (Fig. 18 A-C). This matrix
was rather translucent and partly displayed revascularization (small vessels traveling
along). Furthermore, blood accumulations could be seen accompanying the matrix.
To detect the relative amounts of secreted BDNF into the chitosan conduit the
content with the formed matrix was collected and Western Blot analysis performed.
For both time points, 5 as well as 18 days after nerve reconstruction, pre-mature and
mature BDNF could be detected in the pooled conduit content with no obvious
differences between the groups of different DAs (Fig. 18 D).
Figure 18: Endogenous brain-derived neurotrophic factor (BDNF) detection in tube content. (A-C) Eighteen days after nerve reconstruction a matrix had formed inside the implanted tubes with a jellylike surrounding. The complete tube content was collected and (D) endogenous BDNF was detected to be present in the direct environment of the regenerating nerve tissue (Haastert-Talini et al., 2013).

Results-Project 1
- 57 -
4.2 Static Sciatic Index (SSI) Motor recovery was assessed through determination of the SSI before as wells as 1,
4, 9, and 12 weeks after nerve transection and subsequent nerve reconstruction.
Prior to surgery, healthy animals displayed a SSI around ± 0 (ANG -2.07 ± 1.75; DAI
-7.30 ± 1.71; DAII -4.62 ± 1.72; DAIII -7.96 ± 1.65) indicating that there was no
impairment in the position of the animals paws. One week following nerve lesion and
nerve repair with ANGs or chitosan conduits, the distance between the toes was
altered resulting in a significant decrease of the index for all groups (ANG -73.94 ±
3.21; DAI -74.35 ± 2.84; DAII -76.14 ± 3.31; DAIII -70.72 ± 5.89; (Fig. 19). For the
duration of the complete observation period there was no significant improvement of
the SSI. For the ANG group the SSI mean value was still -54.07 ± 9.07, for the DAI
group -66.12 ± 6.19, for the DAII group -77.92 ± 4.76, and for the DAIII group it was -
75.70 ± 5.38. In addition to the maintained impairment over time, we did not detect
any significant SSI-value differences between the experimental groups.
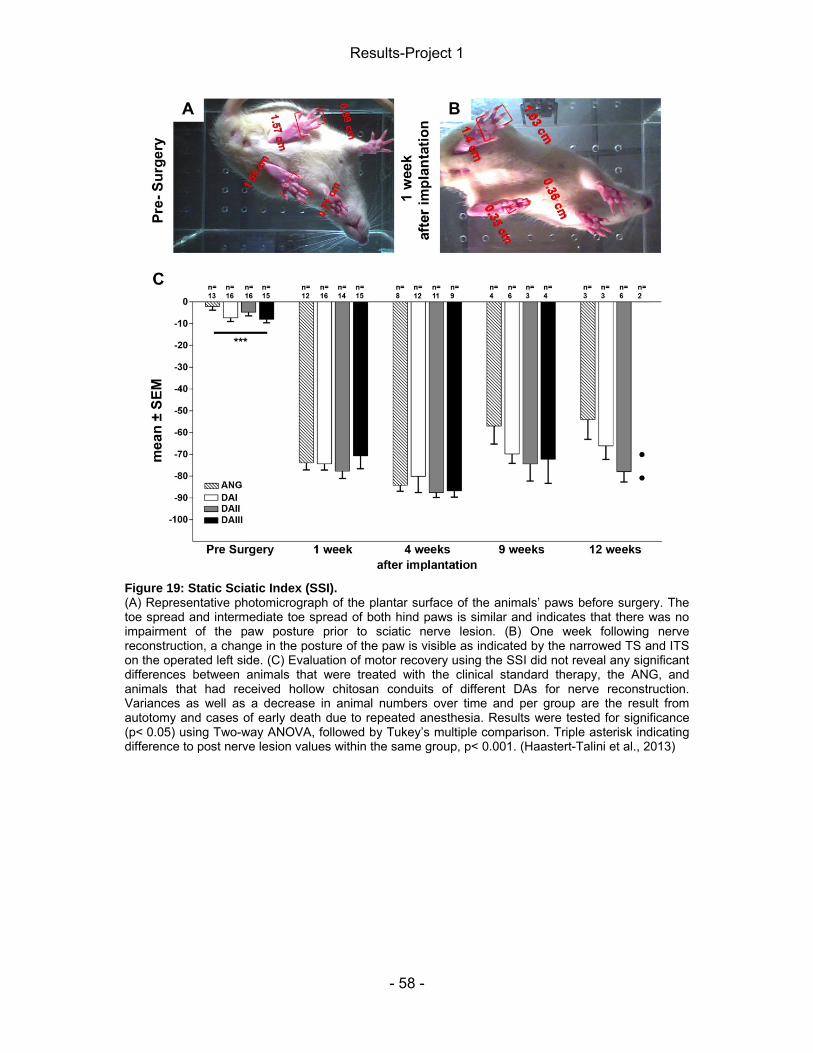
Results-Project 1
- 58 -
Figure 19: Static Sciatic Index (SSI). (A) Representative photomicrograph of the plantar surface of the animals’ paws before surgery. The toe spread and intermediate toe spread of both hind paws is similar and indicates that there was no impairment of the paw posture prior to sciatic nerve lesion. (B) One week following nerve reconstruction, a change in the posture of the paw is visible as indicated by the narrowed TS and ITS on the operated left side. (C) Evaluation of motor recovery using the SSI did not reveal any significant differences between animals that were treated with the clinical standard therapy, the ANG, and animals that had received hollow chitosan conduits of different DAs for nerve reconstruction. Variances as well as a decrease in animal numbers over time and per group are the result from autotomy and cases of early death due to repeated anesthesia. Results were tested for significance (p< 0.05) using Two-way ANOVA, followed by Tukey’s multiple comparison. Triple asterisk indicating difference to post nerve lesion values within the same group, p< 0.001. (Haastert-Talini et al., 2013)

Results-Project 1
- 59 -
4.3 Electrophysiological assessment of muscle reinnervation Reinnervation of the distal muscle targets was investigated by means of non-
invasive electrodiagnostic recordings 4, 9, and 12 weeks after nerve reconstruction
from the TA and PL muscles. Additionally, a final invasive-recording was performed
prior to tissue harvest in week 13.
First non-invasive recordings after 4 weeks did not show any evocable CMAPs. After
9 weeks, 8 out of 13 animals that had received ANGs displayed TA muscle
reinnervation, while this was only the case for single animals of the experimental
groups that had received chitosan conduits of different DAs for nerve reconstruction.
With a delay of additional three weeks, however, an increased number of animals of
conduit repair groups also demonstrated successful functional regeneration, which
was then confirmed with the results obtained during invasive recordings (Tab. 2). Measurements performed on the PL muscle yielded similar results, with the number
of chitosan conduit treated animals showing recordable CMAPs, reaching a level
comparable to that of ANG animals 13 weeks after implantation.
Table 2: Listing of animals per group, which demonstrated evocable CMAPs in the course of the study.
The shape and amplitude of the recorded CMAPs changed in the course of the
study. Four weeks following surgery, either no signals were detected or artifacts
were displayed when recordings were performed from the TA muscle (Fig. 20). These multiphasic artifacts could have been caused through interference from
neighboring muscles or they represent early reinnervation, which is characterized by
asynchronous firing of only a few motor units (Fugleholm et al., 2000; Navarro and
Udina, 2009; Rupp et al., 2007). Solely recordings with amplitudes clearly above
baseline were evaluated to exclude the chance of false positive results. During next
measurements, however, CMAPs were evident from both muscles, which were then

Results-Project 1
- 60 -
analyzed with regard to amplitude (indicating recruited axons / muscle fibers), AUC
(indicating AxL) and NCV.
Figure 20: Serial recordings of CMAPs. Recovery of CMAPs recorded from (A) TA and (B) PL muscles. With progressing reinnervation and a growing number of available muscle units as well as improving synchronization the amplitudes of the signal gradually increased. Note the changing scale for recordings from the lesioned and healthy side at the left margin.

Results-Project 1
- 61 -
For amplitude analysis the maximal amplitude of the M wave was measured and
results are expressed as a ratio (0 = no signal, no recruited axons; 1 = as many
recruited axons as on the non-lesioned side). In case of inclusion of “0” values
resulting from unsuccessful CMAP recordings from the TA muscle, an increase in
amplitude was seen for all experimental groups from week 9 (TA: ANG 0.18 ± 0.04;
DAI 0.05 ± 0.03; DAII 0.03 ± 0.02; DAIII 0.08 ± 0.03) to 12 (TA: ANG 0.36 ± 0.02;
DAI 0.29 ± 0.04; DAII 0.25 ± 0.05; DAIII 0.31 ± 0.04) that was statistically significant
(p< 0.05 for ANG; P< 0.001 for chitosan reconstructed groups). When disregarding
these values, on the other hand, the increase from week 9 to 12 is not significant
(Fig. 21 A). For the CMAPs obtained from the PL muscle no significant increase in
amplitude size could be found with or without inclusion of “0” values (Fig. 21 C). Additionally, no significant differences were seen between all experimental groups,
including the ANG group.

Results-Project 1
- 62 -
Figure 21: Recovery of the CMAP amplitude. Calculated amplitude-ratio from CMAPs recorded from (A, B) TA and (C, D) PL muscle in the course of (A, C) non-invasive recordings performed 9 as well as 12 weeks after conduit implantation and during (B, D) final invasive recordings. No significant differences between the experimental groups or between the results obtained during the different test intervals were found. Note that “0” values are not part of this scheme. Results were tested for significance (p < 0.05) using (A, C) Two- or (B, D) One-way ANOVA.
Invasive recordings performed directly before explantation, confirmed the
functionality of the regenerated nerve tissue and the amplitude-ratio was again
calculated. With regard to results obtained from TA muscle recordings the chitosan
conduit treated animals displayed amplitudes similar to the ANG group (Fig. 21 B), which had a size of about 30 % of the amplitude recorded from fully innervated
contralateral muscles. PL muscle CMAP amplitudes only reached around 20 % of
healthy control values when grafting was undertaken with chitosan conduits, while
the percentage was 39 % for ANGs (Fig. 21 D). This comparatively high percentage,
however, results from varying values within the ANG group, since one animal

Results-Project 1
- 63 -
responded with an almost complete amplitude recovery during stimulation and a
second animal even displayed a higher amplitude size on the lesioned than the
non-lesioned side. Excluding these two animals reduces the average amplitude-ratio
to 0.2 ± 0.04 (~20 %) and consequently to the same level seen with chitosan conduit
grafting.
Results obtained from evaluation of the AUC (AxL) of the CMAP are in line with the
findings of the amplitude analysis. A significant increase in axon numbers is seen for
ANG (P< 0.05), DAI (P< 0.001) and DAII animals (P< 0.01) in case of “100 %” value
inclusion (which indicates complete AxL) from week 9 (TA: ANG 84.48 ± 3.94; DAI
98.10 ± 1.90; DAII 95.76 ± 2.96) to 12 (TA: ANG 63.24 ± 2.34; DAI 69.76 ± 4.04;
DAII 72.59 ± 5.79). Due to a high SEM, however, this is not the case for the DAIII
group. Statistical analysis only of values from positive recordings, on the other hand,
did not reveal a significant increase in axon number over time for any of the groups
(Fig. 22 A). For PL muscle recordings, no significant change in AxL was seen over
time regardless of “100 %” value inclusion or exclusion (Fig. 22 C). Recordings from
direct nerve stimulation confirm the comparable functional recovery of all
experimental groups in terms of TA and PL muscle reinnervation (Fig. 22 B, D). Additional exclusion of the values from the two animals of the ANG group, which
showed a gain in axon number compared to the healthy control nerve when
recordings were undertaken from the PL muscle, shifts the average AxL in this group
to 56.84 ± 5.07, thus approximating the AxL seen with chitosan conduit treatment.
Differences between the experimental groups were not found, regardless of the
investigated time point.

Results-Project 1
- 64 -
Figure 22: Axon Loss. AxL in the sciatic nerve of the injured hind limb was calculated for (A, C) non-invasive and (C, D) invasive measurements from the (A, B) TA and (C, D) PL muscle. No significant differences between the experimental groups or different time intervals were seen. Note that “100 %” values are not part of this scheme. Results were tested for significance (p< 0.05) using (A, C) Two- or (B, D) One-way ANOVA.
The NCV was automatically calculated by the recording device based on the
different latencies obtained from proximal and distal stimulation of the sciatic nerve.
Recordings from the TA muscle revealed a significant increase of NCV from week 9
(TA: ANG 0.2 ± 0.05; DAI 0.02 ± 0.02; DAII 0.04 ± 0.03; DAIII 0.11 ± 0.04) to 12 (TA:
ANG 0.4 ± 0.03; DAI 0.3 ±0.03; DAII 0.26 ± 0.05; DAIII 0.31 ± 0.04) when “0” values
are taken into account for analysis. This, however, is not the case if these values are
excluded. As previously described for the amplitude and the AxL, no change in NCV
is seen for recordings from the PL muscle, in the course of non-invasive
measurements. Also, no differences among the groups were found during
non-invasive as well as invasive recordings for both recording sites (Fig. 23).
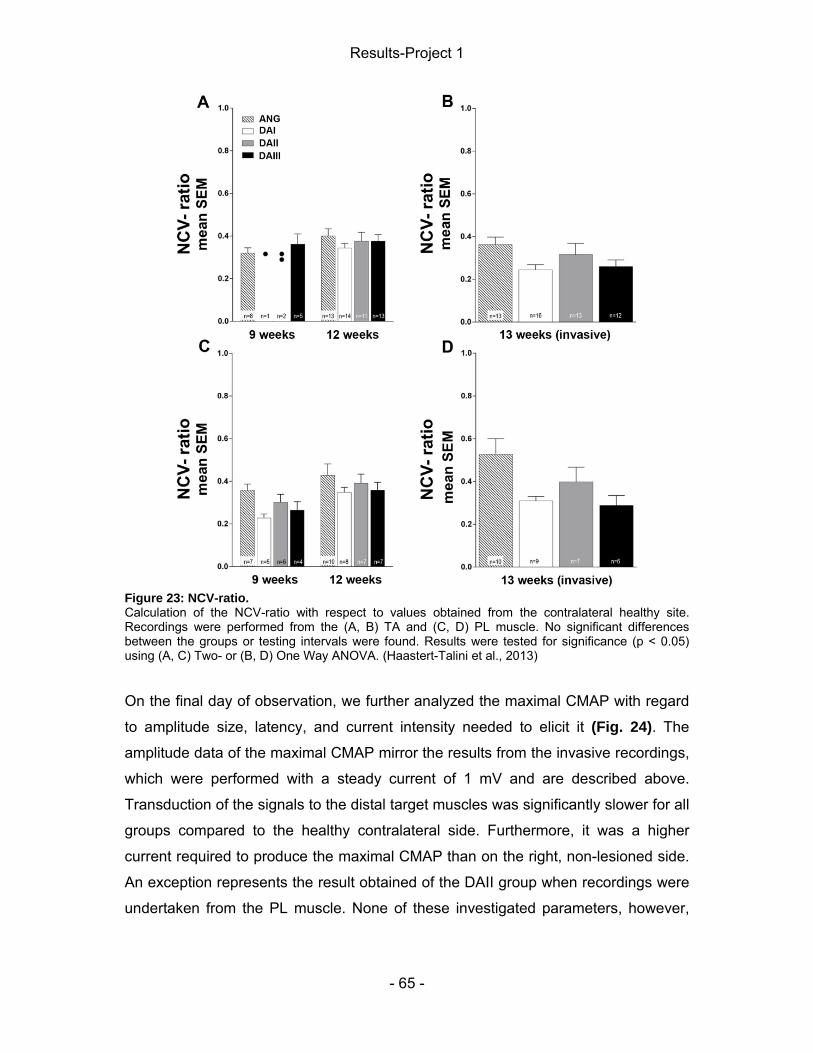
Results-Project 1
- 65 -
Figure 23: NCV-ratio. Calculation of the NCV-ratio with respect to values obtained from the contralateral healthy site. Recordings were performed from the (A, B) TA and (C, D) PL muscle. No significant differences between the groups or testing intervals were found. Results were tested for significance (p < 0.05) using (A, C) Two- or (B, D) One Way ANOVA. (Haastert-Talini et al., 2013)
On the final day of observation, we further analyzed the maximal CMAP with regard
to amplitude size, latency, and current intensity needed to elicit it (Fig. 24). The
amplitude data of the maximal CMAP mirror the results from the invasive recordings,
which were performed with a steady current of 1 mV and are described above.
Transduction of the signals to the distal target muscles was significantly slower for all
groups compared to the healthy contralateral side. Furthermore, it was a higher
current required to produce the maximal CMAP than on the right, non-lesioned side.
An exception represents the result obtained of the DAII group when recordings were
undertaken from the PL muscle. None of these investigated parameters, however,

Results-Project 1
- 66 -
revealed significant differences between the experimental groups including the ANG
group.
Figure 24: Analysis of the maximal CMAP. The maximal CMAP recorded during final invasive recordings from (A, C, E) TA and (B, D, F) PL muscle was analyzed with regard to its (A, B) amplitude, (C, D) latency and (E, F) required current intensity. Significant differences to the healthy contralateral side are marked by asterisk (* =p< 0.05; ** = p< =.01; *** = P < 0.001). Results were tested for significance using One-way ANOVA followed by Tukey’s multiple comparison.

Results-Project 1
- 67 -
4.4 Calculation of the lower limb muscle weight-ratio At the end of the observation period, the TA and gastrocnemius muscles were
harvested to assess muscle atrophy. The harvested tissue from the lesioned side
was macroscopically smaller compared to the non-lesioned side, but the extent of
visible atrophy differed for single animals (Fig. 25 A-C). Nevertheless, there were no
significant differences detected for either muscle between the experimental groups
including the ANG group (Fig. 25 D, C).
Figure 25: Muscle weight determination. Representative photographs of harvested muscle tissue from animals of the (A) ANG, (B) DAII, and (C) DAIII group. No statistical differences (P< 0.05) between the calculated muscle weight ratios could be found for the (D) TA or (E) gastrocnemius muscle using One-way ANOVA (P < 0.05). (Haastert-Talini et al., 2013)
4.6 Conduit properties upon explantation and tissue regeneration As part of the short- as well as long-term studies the regenerated nerve tissue was
cut out as a final step. In case of nerve reconstruction using chitosan conduits, this

Results-Project 1
- 68 -
procedure was conducted with the conduit still around the bridged distance. Five
days after conduit implantation, no macroscopic differences between individual
conduits were visible, while after 18 days, DAIII chitosan conduits already showed
signs of beginning degradation. In 2 out of 6 animals within this group small
longitudinal fissures were seen and another sample displayed a rough surface. The
degradation of DAIII conduits was further in progress after 13 weeks, when 4 out of
15 samples demonstrated longitudinal fissures, 8 out of 15 samples broke apart
upon nerve tissue harvest, and the other three samples were longitudinally broken
and compressed (Fig. 26).
Figure 26: Macroscopic properties of DAIII tubes. Representative photographs showing chitosan conduits of a high DA and regenerated nerve tissue within the bridging material. 13 weeks after surgery, the implanted conduits displayed fissures, crack lines and the tendency to break apart upon explantation (A-D). (A) As a result, the regenerated nerve tissue was partly secondary injured and appeared cleaved.
DAI and DAII conduits, on the other hand, featured a regular shape and firmness
(13/16 and 11/15 respectively) and only in few cases a slightly bent shape (3 within
each group) or a minor compression (DAII: 1/15) was observed, both observations
were accompanied by regular firmness of the sample (Haastert-Talini et al., 2013).

Results-Project 1
- 69 -
Regenerated nerve tissue (Fig. 27) was found in all grafted animals. Only 1 of 15
DAII animals displayed a thin cable (Tab. 3) and the regenerated tissue in 2 out of
16 DAIII animals was, most likely, secondary injured due the early degradation
process of these conduits.
Figure 27: Regenerated nerve tissue upon explantation. Representative photographs showing the regenerated nerve tissue upon explantation from animals of the (A) ANG, (B) DAI, (C) DAII, and (D) DAIII group.

Results-Project 1
- 70 -
Table 3: Overview of animal numbers showing regenerated nerve tissue 13 weeks after chitosan conduit implantation.
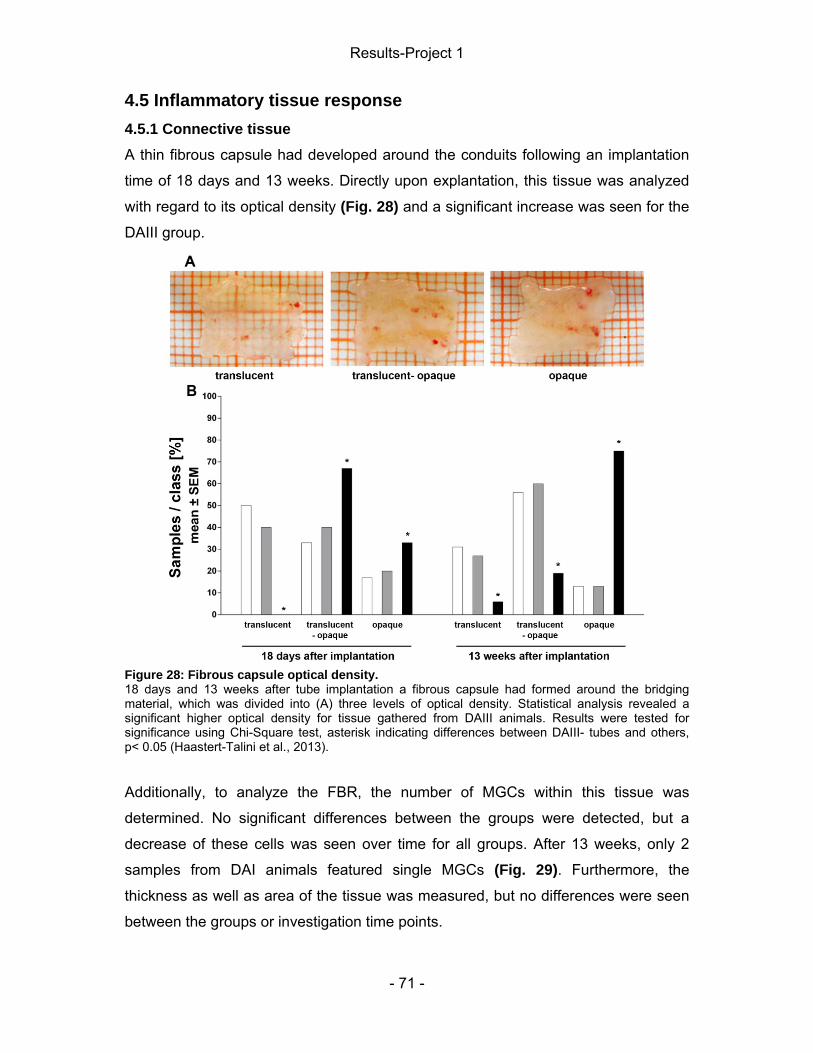
Results-Project 1
- 71 -
4.5 Inflammatory tissue response 4.5.1 Connective tissue A thin fibrous capsule had developed around the conduits following an implantation
time of 18 days and 13 weeks. Directly upon explantation, this tissue was analyzed
with regard to its optical density (Fig. 28) and a significant increase was seen for the
DAIII group.
Figure 28: Fibrous capsule optical density. 18 days and 13 weeks after tube implantation a fibrous capsule had formed around the bridging material, which was divided into (A) three levels of optical density. Statistical analysis revealed a significant higher optical density for tissue gathered from DAIII animals. Results were tested for significance using Chi-Square test, asterisk indicating differences between DAIII- tubes and others, p< 0.05 (Haastert-Talini et al., 2013).
Additionally, to analyze the FBR, the number of MGCs within this tissue was
determined. No significant differences between the groups were detected, but a
decrease of these cells was seen over time for all groups. After 13 weeks, only 2
samples from DAI animals featured single MGCs (Fig. 29). Furthermore, the
thickness as well as area of the tissue was measured, but no differences were seen
between the groups or investigation time points.
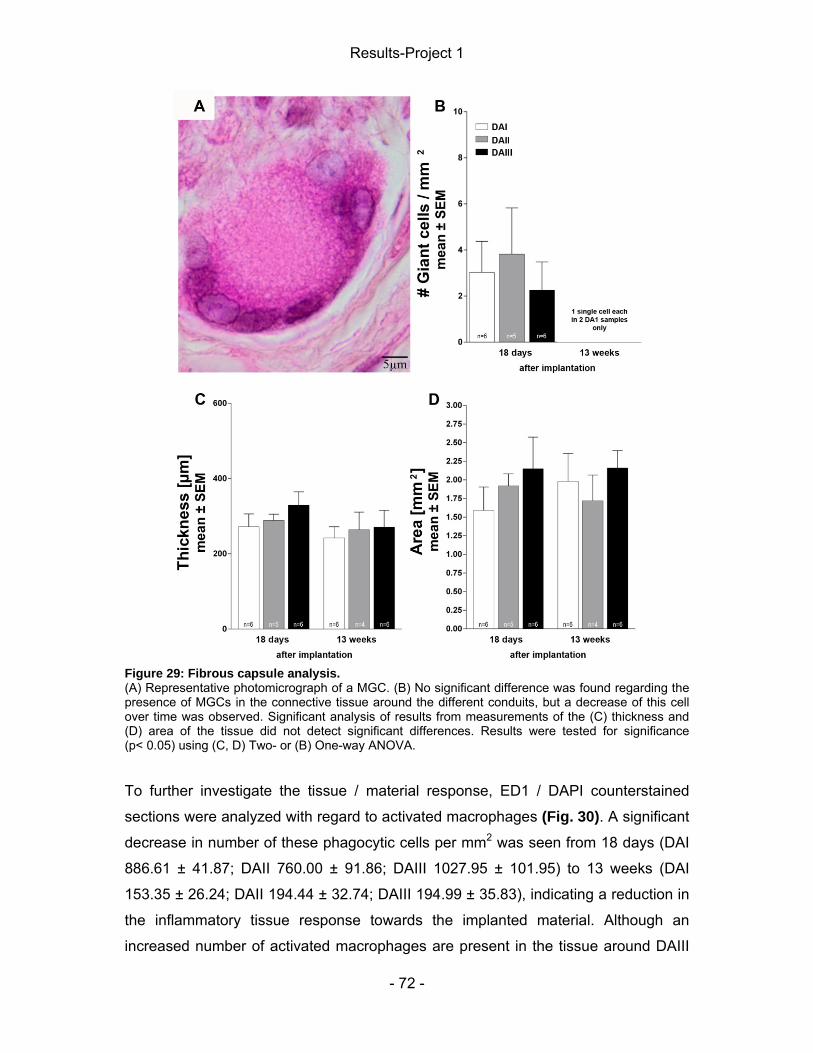
Results-Project 1
- 72 -
Figure 29: Fibrous capsule analysis. (A) Representative photomicrograph of a MGC. (B) No significant difference was found regarding the presence of MGCs in the connective tissue around the different conduits, but a decrease of this cell over time was observed. Significant analysis of results from measurements of the (C) thickness and (D) area of the tissue did not detect significant differences. Results were tested for significance (p< 0.05) using (C, D) Two- or (B) One-way ANOVA.
To further investigate the tissue / material response, ED1 / DAPI counterstained
sections were analyzed with regard to activated macrophages (Fig. 30). A significant
decrease in number of these phagocytic cells per mm2 was seen from 18 days (DAI
886.61 ± 41.87; DAII 760.00 ± 91.86; DAIII 1027.95 ± 101.95) to 13 weeks (DAI
153.35 ± 26.24; DAII 194.44 ± 32.74; DAIII 194.99 ± 35.83), indicating a reduction in
the inflammatory tissue response towards the implanted material. Although an
increased number of activated macrophages are present in the tissue around DAIII

Results-Project 1
- 73 -
tubes 18 days following nerve reconstruction, the differences between the groups
are not significantly different, independently from the investigated time point.
Figure 30: Presence of activated macrophages in the capsule surrounding the conduits. (A) Connective tissue formed around the implanted nerve guidance devices and was (B) cut longitudinally and folded with the areas formerly lying against the conduit facing each other. (C) Numbers of activated macrophages within the tissue. No significant differences were found between the experimental groups, but a decrease was seen over time. Results were tested for significance (p< 0.05) using Two-way ANOVA, Tukey’s multiple comparisons, triple asterisk indicating difference to 18 day values, p< 0.001. (D) Representative photographs of an ED1- immunopositive/DAPI counterstained section 18 days after conduit implantation and (E, F) activated macrophages in detail.
Furthermore, additional sections were investigated for elastic fibers (Orcein-staining),
and collagen (Trichrom-staining). After 18 days, a granulation tissue was seen with
many cells, but only a limited share of collagen and vessels. By 13 weeks, this
granulation tissue had been replaced by a vascular and collagenous fibrous capsule.
Thereby, collagen fibers were facing the conduit and the blood supply was located in

Results-Project 1
- 74 -
the layer fronting the surrounding tissue (Fig. 31, 32). Also, only few cells were seen
compared to 18 days and elastic fibers were confined to the blood vessel walls. The
portion of dense connective tissue, characterized by collagen, and loose connective
tissue, including the passing vessels, varied between samples, but certain patterns
could not be assigned to specific groups.

Results-Project 1
- 75 -
Figure 31: Collagen distribution within capsule. (A, D) 18 days after nerve reconstruction, granulation tissue had formed around the chitosan conduits, which did not feature vessels, only little collagen, but many cells. (B, C, E, F) After 13 weeks, however the collagen portion in the tissue increased (marked by light green color) as revealed by Trichrom-staining. Collagen fibers were mainly traveling in close proximity to the chitosan conduit. * = collagen. Scale bars: 200 µm, 100 µm, 200 µm, 20 µm¸20 µm, and 20 µm.
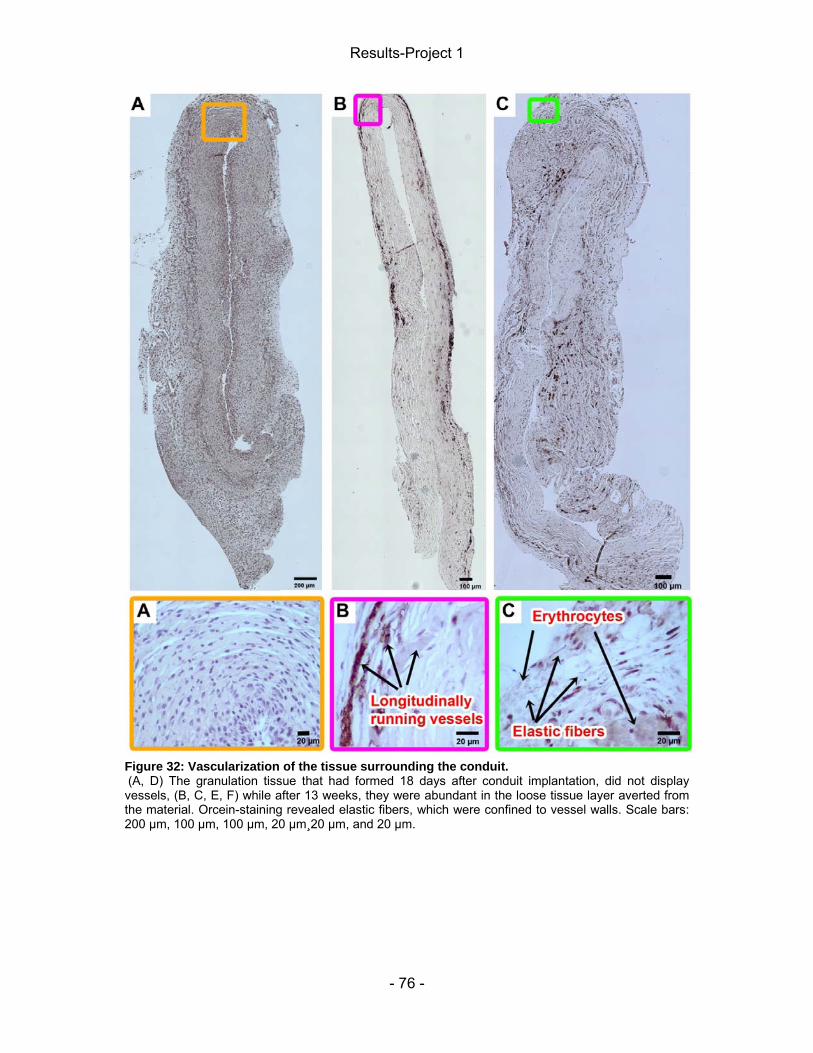
Results-Project 1
- 76 -
Figure 32: Vascularization of the tissue surrounding the conduit. (A, D) The granulation tissue that had formed 18 days after conduit implantation, did not display vessels, (B, C, E, F) while after 13 weeks, they were abundant in the loose tissue layer averted from the material. Orcein-staining revealed elastic fibers, which were confined to vessel walls. Scale bars: 200 µm, 100 µm, 100 µm, 20 µm¸20 µm, and 20 µm.

Results-Project 1
- 77 -
4.5.2 Nerve tissue The nerve tissue harvested in the final day of observation was analyzed with regard
to the FBR. Therefore, the presence of MGCs within the nerve tissue was
investigated (Fig. 33). After 18 days, an unstable matrix had formed inside the tube,
consequently only the proximal nerve stump without regenerated nerve tissue was
prepared for histological analysis (Fig 33 A, B). Only few MGCs were found in the
tissue and no differences between the groups were detected (Fig. 33 C). In line with
the conduit properties observed for the DAIII group upon explantation scattered
chitosan inclusions were discovered in the sections of two out of three animals of this
group (Fig. 33 D, E, G, H), while this was not the case for the DAI group, and only
one single chitosan piece was seen in the periphery of an explanted sample from a
DAII animal (Fig 33 I).

Results-Project 1
- 78 -
Figure 33: Number of MGCs and presence of chitosan inclusions 18 days after tube implantation within the regenerated nerve tissue. The proximal nerve stump of explanted samples was subjected to histological analysis with regard to the presence of MGCs. (C) No statistical differences in the number of MGCs between the experimental groups were revealed 18 days after surgery. Results were tested for significance using One-way ANOVA (p< 0.05). Scattered chitosan inclusions were found in two samples of the (D, E, G, H) DAIII groups, while only one single piece was present in the tissue surrounding a DAII conduit. Abbreviations: C= chitosan; MGC= multinucleated giant cell.
After 13 weeks, regenerated nerve tissue was present inside the tube and sections
were prepared starting 1 mm behind the distal suture (Fig. 34 A, B). In line with
results from the 18 day short-term study no significant differences between the
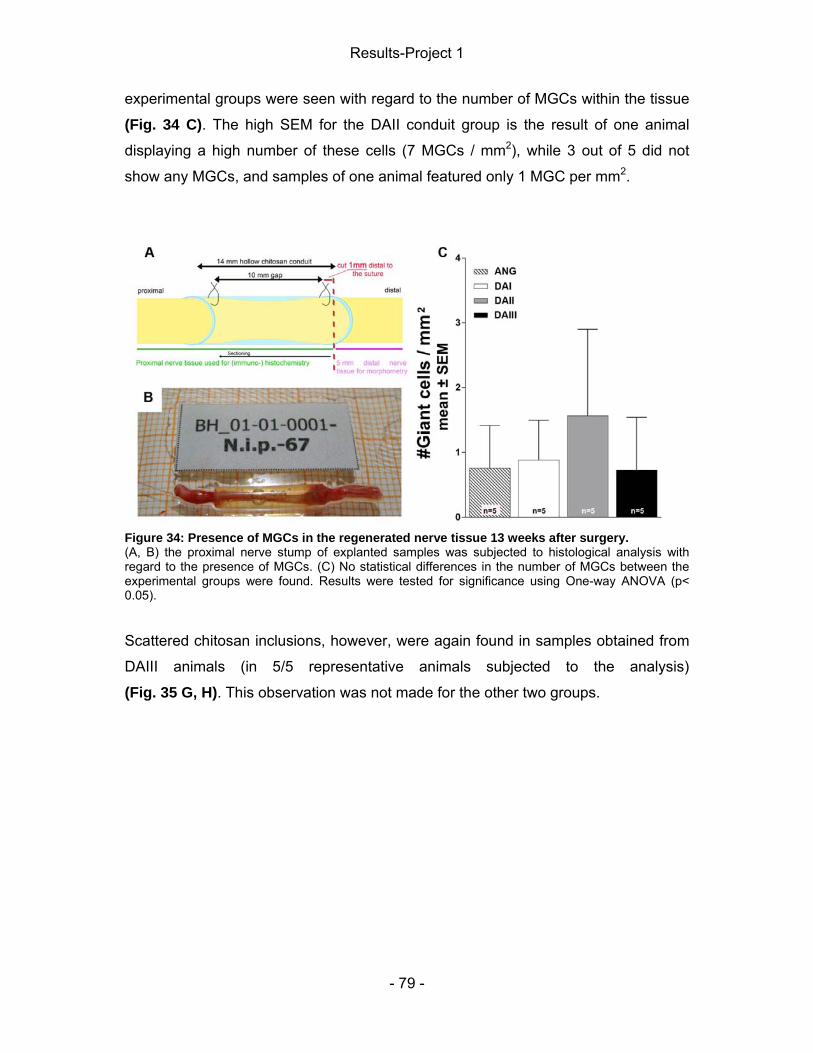
Results-Project 1
- 79 -
experimental groups were seen with regard to the number of MGCs within the tissue
(Fig. 34 C). The high SEM for the DAII conduit group is the result of one animal
displaying a high number of these cells (7 MGCs / mm2), while 3 out of 5 did not
show any MGCs, and samples of one animal featured only 1 MGC per mm2.
Figure 34: Presence of MGCs in the regenerated nerve tissue 13 weeks after surgery. (A, B) the proximal nerve stump of explanted samples was subjected to histological analysis with regard to the presence of MGCs. (C) No statistical differences in the number of MGCs between the experimental groups were found. Results were tested for significance using One-way ANOVA (p< 0.05).
Scattered chitosan inclusions, however, were again found in samples obtained from
DAIII animals (in 5/5 representative animals subjected to the analysis)
(Fig. 35 G, H). This observation was not made for the other two groups.

Results-Project 1
- 80 -
Figure 35: Scattered chitosan inclusions in the nerve tissue of the DAIII group. (A-I) Representative photomicrographs of sections obtained from the regenerated nerve tissue of the (A, B) ANG group and newly grown tissue found inside (C, D) DAI, (E, F) DAII, or (G, H) DAIII conduits. Sections obtained from DAIII animals displayed chitosan inclusions. Furthermore, the thickness of the connective tissue surrounding the axons was increased compared to the other groups. Results regarding the latter were obtained by Sandra Wrobel (M.Sc., MHH Neuroanatomy) and can be looked up in (Haastert-Talini et al., 2013). Abbreviation: C= chitosan.
Consecutive sections from the 18 day study were counterstained for ED1
immunopositive cells and DAPI to assess the number of activated macrophages in

Results-Project 1
- 81 -
the regenerated nerve tissue (Fig 36). Between the experimental groups no
significant differences were found.
Figure 36: Activated macrophages in the proximal nerve stump after 18 days. (A) Sections were prepared form the proximal nerve stump and staining was performed for (B) activated macrophages. (C) Between the grafted groups no significant difference in the number of these cells was found. Results were tested for significance using One Way ANOVA, not including the ANG group (P < 0.05).
Also, sections of regenerated nerve tissue obtained 13 weeks after surgery, were
investigated for activated macrophages. This analysis was performed by Sandra
Wrobel (M.Sc., MHH Neuroanatomy) and can be looked up in (Haastert-Talini et al.,
2013).

- 82 -

Results-Project 2
- 83 -
5 Results-Project 2 5.2 Mechanical pain threshold assessment The second study was performed with the aim to address bridging of long gaps by
introducing luminal fillers into the conduit. Therefore, chitosan conduits were
enriched with NVR-Gel containing genetically modified SCs over-expressing either
GDNF or FGF-218kDa. Investigation parameters were broadened compared to project
1, thus sensory recovery was assessed through determination of the nociceptive
pain threshold in the lateral and medial side of the hind paws. One and 4 weeks
following nerve reconstruction, no withdrawal responses were seen when the lateral
side was stimulated. Five weeks later, all animals of the ANG group showed some
recovery (12.64 ± 1.26 g), but the pressure required to elicit a response was
significantly decreased compared to the values obtained from stimulation of the
contralateral side (20.16 ± 0.30 g) (Fig. 37). An observation, which has, amongst
others, already been described in previous studies by our partner laboratory at the
autonomous University of Barcelona (UAB) (Cobianchi et al., 2014, 2013). On the
other hand, only single animals of the nt-neoSCs (18.93 g), tf-neoSCs-GDNF (20.93
g), and tf-neoSCs-FGF-218kDa (15.60 g) groups demonstrated signs of reinnervation.
In the following weeks, a higher pain threshold was measured for animals of the
ANG group (week 12: 21.44 ± 0.54; week 16: 23.10 ± 0.62) that did not differ
significantly from healthy control values of the contralateral side (week 12: 15.15 ±
1.59; week 16: 17.76 ± 1.28). Until the end of the observation period, only one more
animals of the tf-neoSCs-FGF-218kDa group responded to stimulation of the lateral
side of the paw, while most animals of the groups that had received enriched
chitosan conduits for nerve repair did not show withdrawal responses.
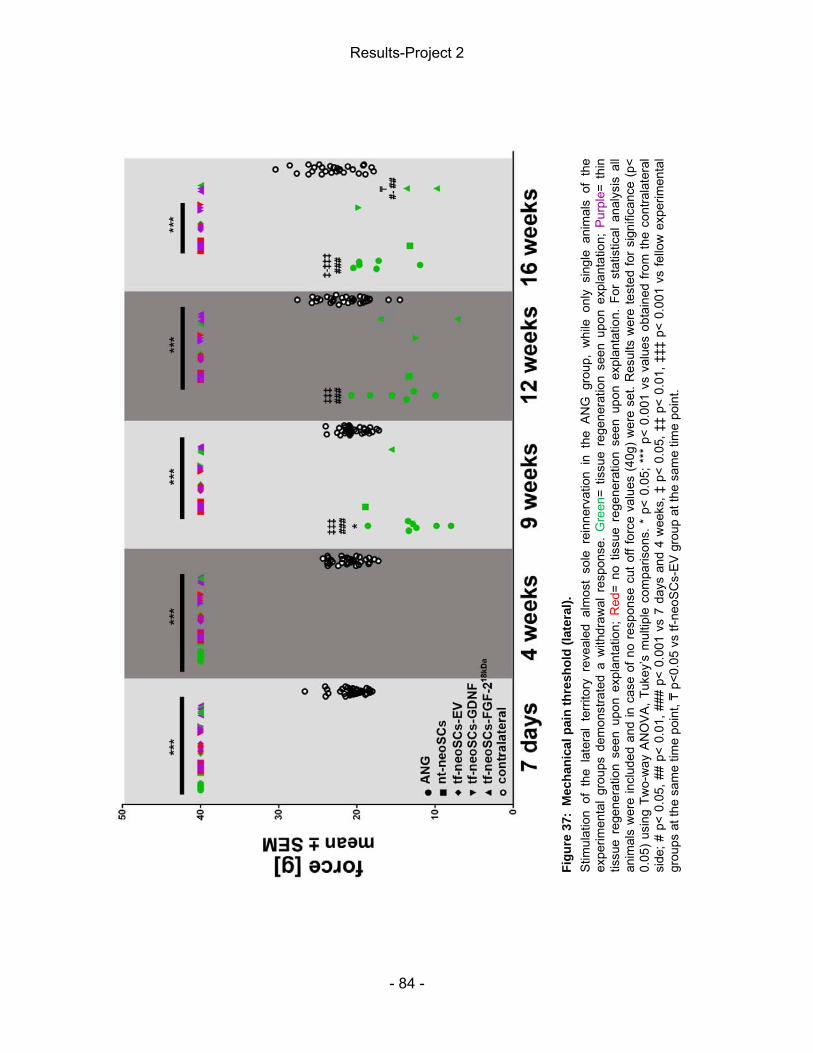
Results-Project 2
- 84 -
Stim
ulat
ion
of t
he l
ater
al t
errit
ory
reve
aled
alm
ost
sole
rei
nner
vatio
n in
the
AN
G g
roup
, w
hile
onl
y si
ngle
ani
mal
s of
the
ex
perim
enta
l gr
oups
dem
onst
rate
d a
with
draw
al r
espo
nse.
Gre
en=
tissu
e re
gene
ratio
n se
en u
pon
expl
anta
tion;
Pur
ple=
thi
n tis
sue
rege
nera
tion
seen
upo
n ex
plan
tatio
n; R
ed=
no t
issu
e re
gene
ratio
n se
en u
pon
expl
anta
tion.
For
sta
tistic
al a
naly
sis
all
anim
als
wer
e in
clud
ed a
nd in
cas
e of
no
resp
onse
cut
off
forc
e va
lues
(40
g) w
ere
set.
Res
ults
wer
e te
sted
for
sign
ifica
nce
(p<
0.05
) us
ing
Two-
way
AN
OVA
, Tuk
ey’s
mul
tiple
com
paris
ons.
* p
< 0.
05; *
** p
< 0.
001
vs v
alue
s ob
tain
ed fr
om th
e co
ntra
late
ral
side
; # p
< 0.
05, #
# p<
0.0
1, #
## p
< 0.
001
vs 7
day
s an
d 4
wee
ks, ‡
p<
0.05
, ‡‡
p< 0
.01,
‡‡‡
p<
0.00
1 vs
fello
w e
xper
imen
tal
grou
ps a
t the
sam
e tim
e po
int, ₸
p<0.
05 v
s tf-
neoS
Cs-
EV
gro
up a
t the
sam
e tim
e po
int.
Fi
gure
37:
Mec
hani
cal p
ain
thre
shol
d (la
tera
l).
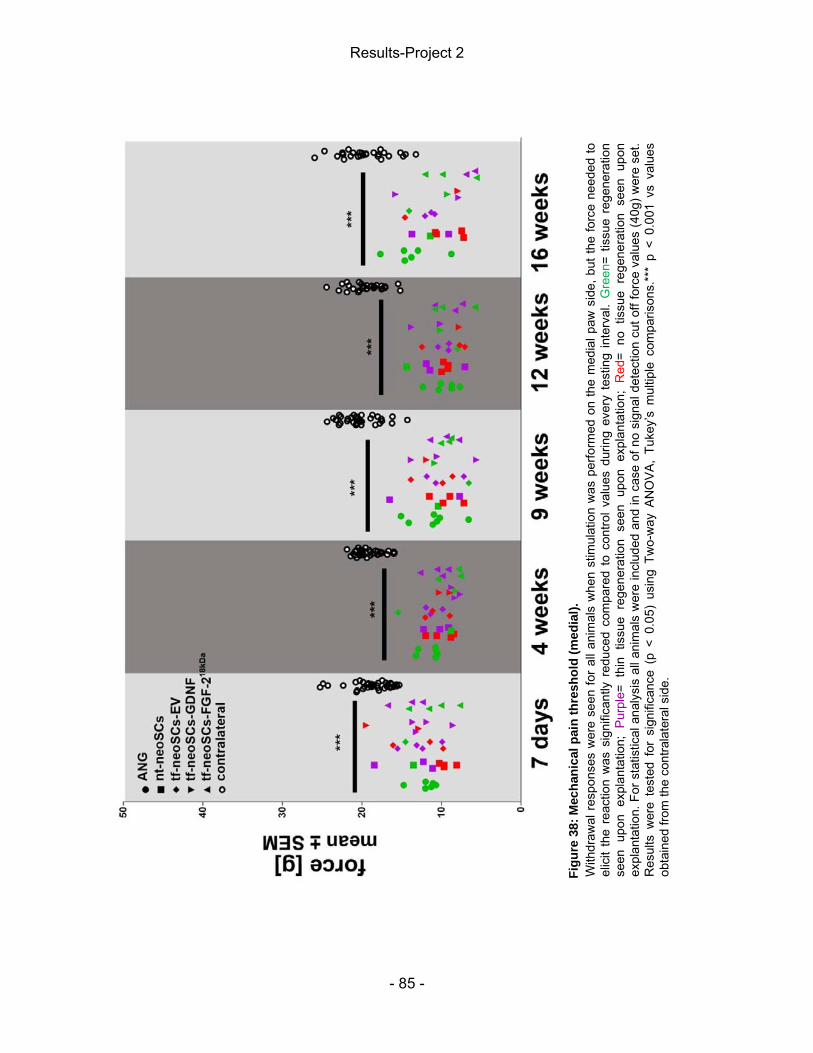
Results-Project 2
- 85 -
Figu
re 3
8: M
echa
nica
l pai
n th
resh
old
(med
ial).
W
ithdr
awal
res
pons
es w
ere
seen
for
all a
nim
als
whe
n st
imul
atio
n w
as p
erfo
rmed
on
the
med
ial p
aw s
ide,
but
the
forc
e ne
eded
to
elic
it th
e re
actio
n w
as s
igni
fican
tly r
educ
ed c
ompa
red
to c
ontro
l val
ues
durin
g ev
ery
test
ing
inte
rval
. G
reen
= tis
sue
rege
nera
tion
seen
upo
n ex
plan
tatio
n; P
urpl
e= t
hin
tissu
e re
gene
ratio
n se
en u
pon
expl
anta
tion;
Red
= no
tis
sue
rege
nera
tion
seen
upo
n ex
plan
tatio
n. F
or s
tatis
tical
ana
lysi
s al
l ani
mal
s w
ere
incl
uded
and
in c
ase
of n
o si
gnal
det
ectio
n cu
t off
forc
e va
lues
(40g
) wer
e se
t. R
esul
ts w
ere
test
ed f
or s
igni
fican
ce (
p <
0.05
) us
ing
Two-
way
AN
OVA
, Tu
key’
s m
ultip
le c
ompa
rison
s.**
* p
< 0.
001
vs v
alue
s ob
tain
ed fr
om th
e co
ntra
late
ral s
ide.

Results-Project 2
- 86 -
The pain threshold measured in response to stimulation of the medial paw side, a
territory that is also partly innervated by the saphenous nerve, was significantly
decreased following nerve repair (ANG: 11.98 ± 0.60 g; nt-neoSCs 11.65 ± 1.14 g; tf-
neoSCs-EV 12.86 ± 0.85 g; tf-neoSCs-GDNF 13.27 ± 1.45 g; tf-neoSCs-FGF218kDa
12.35 ± 1.11 g) compared to values obtained from the contralateral side (18.54 ±
0.42) (Fig. 38). For the complete duration of the study, however, the animals
continued to show signs of hyperalgesia on the lesioned side and the measured pain
threshold remained significantly decreased. ₸
5.1 Static Sciatic Index (SSI) Following nerve reconstruction, using enriched chitosan conduits or ANGs, motor
recovery was analyzed at various time points via SSI as seen for project 1.
Pre-surgery values confirmed that there was no paw position impairment before
implantation (ANG -1.90 ± 2.93; nt-neoSCs -3.15 ± 2.83; tf-neoSCs-EV -0.64 ± 3.11;
tf-neoSCs-GDNF -2.25 ± 2.30; tf-neoSCs-FGF-218kDa -3.73 ± 2.50) (Fig. 39). Shortly
after lesion and subsequent reconstruction, however, the SSI dropped to around
-100 % (ANG -95.69 ± 1.97; nt-neoSCs -96.91 ± 2.32; tf-neoSCs-EV -92.70 ± 1.45;
tf-neoSCs-GDNF -100.52 ± 3.64; tf-neoSCs-FGF-218kDa -94.37 ± 1.58) due to the
missing innervation from the sciatic nerve branches.

Results-Project 2
- 87 -
Figure 39: Results of the calculation of the Static Sciatic Index (SSI). Evaluation of distal target reinnervation using SSI did not reveal any significant difference between animals that were treated with ANGs, and animals that received enriched chitosan conduits for nerve reconstruction. Results were tested for significance (p < 0.05) using Two-way ANOVA, Tukey’s multiple comparisons, ***= difference to post nerve lesion values, p < 0.001, and #= difference to 4 weeks post lesion values of tf-neoSCs-EV, p < 0.05.
Four month later, there was no significant improvement regarding distal muscle
target reinnervation assessed by SSI for all experimental groups when compared to
values obtained 1 week after lesion. For the ANG group the SSI mean value was still
-83.64 ± 11.03, for nt-neoSCs -80.08 ± 5.69, for tf-neoSCs-EV -74.40 ± 2.71, for
tf-neoSCs-GDNF -80.32 ± 5.01 and for tf-neoSCs-FGF218kDa -92.03 ± 5.02.
5.3 Electrophysiological assessment of muscle reinnervation Motor recovery was further evaluated by means of non-invasive nerve conduction
tests 9, 13, and, 17 weeks after sciatic nerve lesion and subsequent repair, as well
as by additional final invasive recordings. Measurements performed at the earliest
time point revealed reinnervation of the TA and PL muscle for almost all animals of
the ANG group, while this was only seen for single animals of the nt-neoSCs and
tf-neoSCs-EV group. Four weeks later, all animals of the ANG group showed
evocable CMAPs, while the signal detected for the animal of the tf-neoSCs-EV group
after 9 weeks was not confirmed. Instead one animal of the tf-neoSCs-FGF-218kDa
group showed signs of reinnervation. On the last day of observation, however,

Results-Project 2
- 88 -
signals were recorded from 4 out of 7 animals of the tf-neoSCs-FGF-218kDa group
when recordings were undertaken from the TA muscle (Tab. 4). Table 4: Listing of animals per group, which demonstrated evocable CMAPs in the course of the study.
These findings were confirmed during final invasive recordings. One of these
animals further displayed PL muscle reinnervation. Exemplary CMAPs recorded from
the TA and PL muscle during the different recording sessions are depicted in Fig. 40.

Results-Project 2
- 89 -
Figure 40: Serial recordings of CMAPs. Recovery of CMAPs recorded from (A) TA and (B) PL muscle following nerve repair of a 15 mm gap with either an ANG or enriched chitosan conduit containing FGF-218kDa over-expressing SCs. Note the changing scale for recordings from the lesioned and healthy side at the left margin.

Results-Project 2
- 90 -
The data gained from the recordings were used to calculate the NCV, AUC (AxL),
and amplitude-ratio, with the latter two providing information with regard to axon loss
or recruited axons, respectively.
At the end of the observation period, solely tf-neoSCs-FGF-218kDa treatment allowed
TA muscle reinnervation that was comparable to results seen following ANG
treatment (Fig. 41). The latter, however, enabled recovery of the amplitude size to
almost 50 % of contralateral control values, while the FGF-218kDa group reached
13 %. Regarding PL muscle reinnervation ANG treatment was superior to all groups,
since only one animal of the tf-neoSCs-FGF-218kDa group showed CMAPs during the
final invasive recording (Fig. 42). Also, single animals of the nt-neoSCs and
tf-neoSCs-GDNF groups displayed PL muscle CMAPS during non-invasive
recordings, but these signals were not stable and could not be confirmed during
invasive recordings.

Results-Project 2
- 91 -
Non
-inva
sive
reco
rdin
gs fr
om T
A m
uscl
es re
veal
ed s
igni
fican
t diff
eren
ces
betw
een
the
AN
G g
roup
and
all
grou
ps th
at h
ad
been
tre
ated
with
eith
er o
f th
e en
riche
d ch
itosa
n co
ndui
ts.
Add
ition
ally
, so
lely
tre
atm
ent
with
the
gol
d st
anda
rd le
d to
a
sign
ifica
nt im
prov
emen
t of m
uscl
e re
inne
rvat
ion
over
tim
e (T
wo-
way
AN
OV
A fo
llow
ed b
y Tu
key’
s m
ultip
le c
ompa
rison
, # <
0.
05 v
s A
NG
13
and
17 w
eeks
; *
p< 0
.05
vs A
NG
gro
up o
f th
e sa
me
wee
k).
Fina
l inv
asiv
e re
cord
ings
17
wee
ks a
fter
surg
ery,
how
ever
, re
veal
ed s
igni
fican
t di
ffere
nces
bet
wee
n th
e pe
rform
ance
of
the
ANG
gro
up a
nd t
he a
nim
als
treat
ed
with
the
diff
eren
t va
riant
s of
enr
iche
d co
ndui
ts,
exce
pt f
or t
he g
roup
tha
t ha
d re
ceiv
ed t
f-neo
SC
s-FG
F-218
kDa t
reat
men
t (K
rusk
al-W
allis
test
follo
wed
by
Dun
n’s
mul
tiple
com
paris
on, *
p<
0.05
vs
AN
G g
roup
of t
he s
ame
wee
k. G
reen
= tis
sue
rege
nera
tion
seen
up
on
expl
anta
tion;
P
urpl
e=
thin
tis
sue
rege
nera
tion
seen
up
on
expl
anta
tion;
R
ed=
no
tissu
e re
gene
ratio
n se
en u
pon
expl
anta
tion.
For
sta
tistic
al a
naly
ses
all a
nim
als
wer
e in
clud
ed a
nd in
cas
e of
no
sign
al d
etec
tion
“0” v
alue
s w
ere
set.
Figu
re 4
1: A
mpl
itude
-rat
io fr
om C
MA
Ps re
cord
ed fr
om th
e tib
ialis
ant
erio
r (TA
) mus
cle.

Results-Project 2
- 92 -
PL
mus
cle
rein
nerv
atio
n fo
llow
ing
treat
men
t w
ith A
NG
s w
as s
uper
ior
to a
ll ot
her
treat
men
t st
rate
gies
at
ever
y re
cord
ing
time
poin
t. (N
on-in
vasi
ve: T
wo-
way
AN
OV
A fo
llow
ed b
y Tu
key’
s m
ultip
le c
ompa
rison
, # <
0.0
5 vs
AN
G 1
3 an
d 17
wee
ks; #
# <
0.05
vs
AN
G 1
3 an
d 17
wee
ks, *
p<
0.05
vs
ANG
gro
up o
f the
sam
e w
eek;
Inva
sive
: Kru
skal
-W
allis
test
follo
wed
by
Dun
n’s
mul
tiple
com
paris
on, *
< 0
.05
vs A
NG
gro
up o
f th
e sa
me
wee
k). (
B)
Gre
en=
tissu
e re
gene
ratio
n se
en u
pon
expl
anta
tion;
Pur
ple=
thi
n tis
sue
rege
nera
tion
seen
upo
n ex
plan
tatio
n; R
ed=
no t
issu
e re
gene
ratio
n se
en u
pon
expl
anta
tion.
For
sta
tistic
al a
naly
ses
all
anim
als
wer
e in
clud
ed a
nd i
n ca
se o
f no
sig
nal
dete
ctio
n “0
” val
ues
wer
e se
t.
Figu
re 4
2: A
mpl
itude
-rat
io fr
om C
MA
Ps re
cord
ed fr
om th
e pl
anta
r (PL
) mus
cle.

Results-Project 2
- 93 -
Results of NCV- and AxL-calculation support the findings of the amplitude analysis.
During non-invasive TA muscle recordings, animals of the ANG group showed
significant faster NCVs and more available axons compared to the other
experimental groups (Tab. 5). In the course of invasive tests, however, no significant
differences were found between the NVC and AxL calculated for ANG and tf-
neoSCs-FGF-218kDa treated animals. An increase in NCV over time was seen solely
for the ANG group when recordings were undertaken from the PL muscle.
Furthermore, a significant decrease of AxL over time was seen for the ANG (TA and
PL) and FGF-218kDa (TA) groups.

Results-Project 2
- 94 -
Ta
ble
5: N
CV,
AxL
, and
am
plitu
de-r
atio
Te
stin
g fo
r sig
nific
ance
(p<
0.05
) was
per
form
ed u
sing
Kru
skal
-Wal
lis te
st o
r Tw
o-w
ay A
NO
VA, f
ollo
wed
by
Tuke
y’s
or D
unn’
s m
ultip
le c
ompa
rison
. *p
< 0.
05 v
s fe
llow
exp
erim
enta
l gro
ups,
**p
< 0.
01 v
s fe
llow
exp
erim
enta
l gro
ups,
***
p< 0
.001
vs
fello
w e
xper
imen
tal g
roup
s, #
# p<
0.0
1 vs
AN
G 1
7 w
eeks
, ##
# p<
0.0
01 v
s A
NG
17
wee
ks,
§ p<
0.0
5 vs
AN
G 1
3 w
eeks
, §§
p<
0.01
vs
ANG
13
wee
ks,
$$ p
<0.0
1 vs
nt-n
eoS
Cs,
tf-n
eoS
Cs-
EV
, tf-
neoS
Cs-
GD
NF,
$$$
p<
0.00
1 vs
nt-n
eoSC
s, tf
-neo
SC
s-EV
, tf-n
eoSC
s-G
DN
F

Results-Project 2
- 95 -
5.4 Calculation of the lower limb muscle weight-ratio The muscle weight-ratio was calculated subsequently to harvest of the TA and
gastrocnemius muscles. Solely nerve reconstruction using enriched chitosan
conduits featuring SCs that over-expressed FGF-218kDa led to a muscle weight that
did not differ significantly from that seen after ANG treatment although an optical
difference was still visible (Fig. 43).
Figure 43: Muscle weight-ratio. The TA as well as gastrocnemius muscle weights were determined upon explantation of the nerve tissue specimens. Representative photomicrographs showing cut muscles from animals that demonstrated successful tissue regeneration: (A) ANG group, (B) FGF-218kDa over-expressing group. (C) Harvested muscles from an animal without tissue regeneration that had been treated with a conduit containing non-transfected SCs. (D, E) Significant differences were found comparing the ANG group with all experimental groups, except for the group that had received conduits with SCs over-expressing FGF-218kDa, although an optical difference was seen (Kruskal-Wallis test, followed by Dunn’s multiple comparison, *< 0.05 vs ANG group). Green= tissue regeneration seen upon explantation; Purple= thin tissue regeneration seen upon explantation; Red= no tissue regeneration seen upon explantation.
5.3 Autotomy score The degree of automutilation was regularly assessed using an autotomy score.
Predominantly animals that had received enriched chitosan conduits for peripheral

Results-Project 2
- 96 -
nerve reconstruction displayed automutilation, while this was only case for one
animal of the ANG group. Thereby, animals that showed tissue regeneration as well
as animal with failed regeneration after 17 weeks were affected. Animals of the
tf-neoSCs-GDNF group, however, demonstrated highest ratings on the score
(Fig. 44).
Figure 44: Autotomy Score. Animals predominantly displayed no automutilation or mild toe nail biting (1 point), but some cases of higher degrees of automutilation were also seen. For the duration of the experiment, animals of the tf-neoSCs-GDNF group demonstrated the highest score. Green= tissue regeneration seen upon explantation; Purple= thin tissue regeneration seen upon explantation; Red= no tissue regeneration seen upon explantation. Results were tested for significance (p< 0.05) using Two Way ANOVA.
5.5 Macroscopic analysis of the explanted nerve tissue Following final functional tests, regenerated nerve tissue was collected if regeneration had occurred. All animals of the ANG group showed regeneration (Tab. 6, Fig. 45A), while this was only the case for single animals of the experimental groups. Solely the tf-neoSCs-FGF-218kDa treatment group stood out within the conduit groups with 4 out of 7 animals showing clear tissue regeneration (Fig. 45B), and the remaining animals of the group displaying thin tissue regeneration. Table 6: Tissue regeneration seen upon explantation.

Results-Project 2
- 97 -
Figure 45: Representative photographs of explanted specimens following a 4 month observation period. (A) Successful tissue regeneration following ANG treatment. (B) Tissue regeneration through a chitosan conduit filled with NVR-Gel and SCs overexpressing FGF-218kDa. (C) Thin tissue regeneration (arrows) through a chitosan conduit filled with NVR-Gel and empty vector transfected SCs. (D) Failed regeneration through a chitosan conduit filled with NVR-Gel and SCs over-expressing GDNF.
5.6 Inflammatory tissue response 5.6.1 Connective tissue The fibrous capsule that had developed around the conduits was harvested and
analyzed with regard to MGCs, its thickness, and area. Only single MGCs were
found in one out of four animals of the tf-neoSCs-EV group and in two out of six
animals of the tf-neoSCs-FGF-218kDa group. Furthermore, no significant differences
were seen in the thickness of the connective tissue as well as its area when
comparing samples collected from the different experimental groups (Fig. 46).

Results-Project 2
- 98 -
Figure 46: Fibrous capsule analysis. Connective tissue that had developed around the chitosan tubes was analyzed with regard to (A, B) presence of MGCs (arrows), its (C) thickness, and (D) area. No significant differences between the experimental groups were detected (One way ANOVA; p < 0.05).
To further analyze the inflammatory response staining for activated macrophages
(ED1+) was performed on consecutive sections, but counting of these cells also did
not reveal significant differences between the groups (Fig. 47).

Results-Project 2
- 99 -
Figure 47: Presence of activated macrophages in the capsule surrounding the conduits. (A) Sections of connective tissue surrounding the enriched chitosan conduits were stained for activated macrophages. Some vessels also appear in red as indicated by arrows. (B) Detail of activated macrophages. (C) No significant differences were found between the experimental groups. Results were tested for significance (p< 0.05) using Kruskal-Wallis test.
Further sections were examined with regard to collagen formation and elastic fiber
presence. As seen with hollow chitosan conduits (project 1), collagen formed directly
at the material / tissue interface, while vascularization of the tissue occurred in the
outer, more loose layers of the capsule. The samples displayed variations regarding
the content of collagen and loose connective tissue (FIG. 48, 49), but these
observations could not be assigned to specific groups. Also, single samples of the
experimental groups (nf-neoSCs, tf-neoSCs-EV, and tf-neoSCs-GDNF each one
sample) were characterized by the presence of numerous cells in the direct contact
area with the material as depicted in Fig. 49D and H.

Results-Project 2
- 100 -
Figure 48: Collagen distribution within capsule. (A-C) Collagen bands (light green) are seen in the direct contact area with the chitosan as revealed by TriChrom-staining. (E-G) Details of the contact areas. (D) Representative overview photomicrograph of a sample displaying numerous cells compared to other capsules. (H) Detail of the same sample, which also featured a high number of activated macrophages when stained for ED1.

Results-Project 2
- 101 -
Figure 49: Vascularization of the capsule surrounding the conduit. (A-D) Orcein-staining revealed the presence of elastic fibers in connection with vessels in the outer layers of the connective tissue that had formed around the implant. (E-H) Details of vessels showing elastic fibers as part of the vessel wall.
5.6.2 Regenerated tissue Samples of successful tissue regeneration were subjected to HE- and ED1-staining
and analyzed towards the presence of MGCs and activated macrophages,
respectively. Only single MGCs were detected in the samples, except for one animal
of the FGF-218kDa group. In this case a higher number of these cells were found in
two out of four analyzed sections, while no MGCs were seen in the other two
sections. The same animal also displayed the highest number of macrophages when
compared to other samples of successful regeneration (Tab. 7).
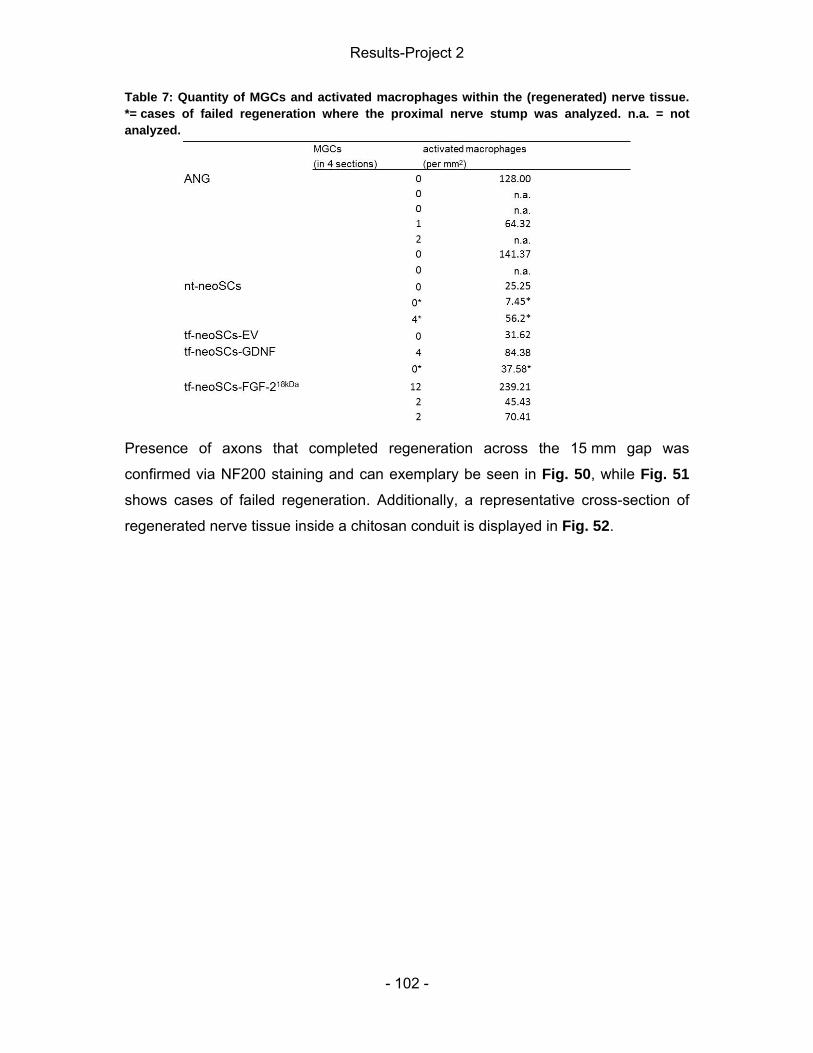
Results-Project 2
- 102 -
Table 7: Quantity of MGCs and activated macrophages within the (regenerated) nerve tissue. *= cases of failed regeneration where the proximal nerve stump was analyzed. n.a. = not analyzed.
Presence of axons that completed regeneration across the 15 mm gap was
confirmed via NF200 staining and can exemplary be seen in Fig. 50, while Fig. 51
shows cases of failed regeneration. Additionally, a representative cross-section of
regenerated nerve tissue inside a chitosan conduit is displayed in Fig. 52.

Results-Project 2
- 103 -
Figure 50: Regeneration across the gap. (A) Schematic illustration of the approximate localization of the stained sections from regenerated samples obtained from animals of the (C, D) ANG and (E, F) FGF-218kDa group. (C, E) Sections within the bridged distance (orange box) and (D, F) close to the distal suture site (pink box). (B) Representative staining of a healthy control nerve.

Results-Project 2
- 104 -
Figure 51: Failed regeneration. Schematic illustration of the approximate localization of the stained sections from samples obtained from animals of the (B, C) nt-neoSCs and (D, E) tf-neoSCs-GDNF groups. (C, E) Sections close to the suture site (pink box) and (B, D) 1.5 mm more proximal (orange box) were stained for activated macrophages and neurofilament. (C, F,). No or only very few axons were seen directly at the entering site into the chitosan conduits, however, sections of a more proximal region stained positive for neurofilament.

Results-Project 2
- 105 -
Figure 52: Cross-section of an enriched chitosan conduit and regenerated nerve tissue 17 weeks following implantation. Representative photomicrograph of a cross-section obtained 17 weeks after surgery from an FGF-218kDa animal, which also demonstrated functional recovery by means of electrophysiological recordings.

- 106 -

Discussion
- 107 -
6 Discussion PNIs have been a challenging research field in the last century and represent a
global clinical problem. Although, there has been great progress in development of
nerve guidance conduits based on synthetic and non-synthetic materials, the
functional outcome for patients is often unsatisfactory. Consequently, the ANG
remains the clinical standard therapy. Despite refinement of this technique, however,
bridging nerve defects with dispensable sensory nerves does also not guarantee a
full recovery. Furthermore, this method is associated with disadvantages like a
limited availability of autologous nerve tissue and mismatch issues. Consequently,
alternatives have to be created for the currently available bridging strategies that
allow improvement of peripheral nerve regeneration across small gaps, but also
address critical gaps.
As part of this thesis, we tested hollow chitosan conduits of three different DAs
(DAI= 1%, DAII= 5%, DAIII= 20%) for peripheral nerve repair of small gaps
(Project 1). Therefore, we introduced a 10 mm defect into the sciatic nerve of rats
and evaluated the performance of the chitosan conduits with regard to functional
outcome and foreign body response elicited through the implantation of the material.
Both issues were addressed in a long-term study (13 weeks) with ANGs serving as
positive control. Secondary, short-term-studies were conducted to look at the
regulation of regeneration associated proteins as well as at the tissue response
shortly upon nerve lesion in the presence of chitosan (5 days and 18 days).
In addition, as a second project of my thesis, we focused on NVR-Gel as a matrix
inside the tube, which contained genetically modified SCs over-expressing GDNF or
FGF-218kDa to improve peripheral nerve regeneration across critical defects
(Project 2). Therefore, a 15 mm gap was bridged with enriched chitosan conduits
and the functional recovery and device / tissue reaction was analyzed in a long-term
study of 17 weeks duration.

Discussion-Project 1
- 108 -
6.1 Project 1 6.1.1 Regulation of regeneration associated proteins following nerve lesion and repair In response to nerve damage, various NTFs and cytokines as well as their receptors
are subjected to putative expression changes in a time and spatial manner. Neurons
and SCs up-regulate these survival and axonal outgrowth promoting factors and
thereby provide the basis for successful nerve regeneration (Boyd and Gordon,
2003a; Gordon, 2009). It is therefore of importance that the physiological regulation
is not disturbed as a result of material implantation. Proximal and distal nerve
stumps, the lumbar region of the spinal cord as well as DRGs (L3-L5) were collected
5 and 18 days after nerve lesion and repair with chitosan conduits. Using qRT-PCR,
the expression level of NTFs (NGF, FGF-2, BDNF), the cytokine IL-6, the BDNF-
receptor TrkB, and GAP-43 were determined and normalized to values obtained
from the contralateral side of control animals. Analysis of the nerve stumps,
however, was not successful, most likely because the distance (5 mm) that was
harvested was insufficient.
NGF as well as IL-6 expression was below detection limit in the spinal cord. The
former is in accordance with reports that NGF is neither expressed by motoneurons
in a detectable amount nor up-regulated upon injury (Boyd and Gordon, 2003a;
Josephson et al., 2001). Another study, on the other hand, has demonstrated NGF
expression in ventral spinal cords, but no regulation in any direction was revealed 1
or 2 days after injury (Cobianchi et al., 2013). IL-6 transcripts have previously been
reported to be immediately up-regulated by axotomized motoneurons, peaking at
24 h before returning to undetectable baseline expression within 21 days after nerve
injury (Kiefer et al., 1993). Also, an increase of IL-6 mRNA levels has been reported
by SCs of ventral roots after 4 and 7 days using an in vitro model that mimics
degeneration (Allodi et al., 2013). BDNF, FGF-2, GAP-43, and trkB were all neither
significantly up- nor down-regulated in the spinal cord and also no differences
between the experimental groups including the control group were seen. Previous
results regarding BDNF expression in the ventral horn have revealed that there is no
up-regulation 1 or 3 days following nerve injury (Cobianchi et al., 2013). But also an
up-regulation in the spinal cord 2 days after injury has been reported before

Discussion-Project 1
- 109 -
subsequent down-regulation to below baseline levels by 2 weeks. Additionally,
GAP-43 has already been shown to not be significantly up- or down-regulated
(Haastert-Talini et al., 2011). Allodi et al. (2013) further described an up-regulation of
FGF-2 by SCs of ventral roots in the same study set-up as noted for IL-6.
Results addressing expression levels of the same factors in DRGs following nerve
repair with chitosan conduits were obtained by Dr. Ratzka (MHH, Institute of
Neuroanatomy) (Haastert-Talini et al., 2013). The data show that regardless of the
time point (5 days or 18 days) NGF, FGF-2, IL-6, BDNF. and GAP-43 were all
up-regulated in the control group as well as in the conduit treated groups, while no
change in expression was seen for TrkB. BDNF has been previously described to be
up-regulated 1 to 3 days after injury, before returning to baseline levels by 14 days
(Cobianchi et al., 2013; Haastert-Talini et al., 2011). An in vitro model mimicking
degeneration after injury has revealed IL-6 up-regulation in dorsal roots to a similar
extent as seen in this study after 4 days before decreasing again following a 7 days
culture period (Allodi et al., 2013). An increase of GAP-43 transcripts has already
been described 2 days after injury, while baseline levels were reported after 2 weeks
and only in case of electrical stimulation did the levels remain significantly higher
(Haastert-Talini et al., 2011). Furthermore, up-regulation of mRNA levels of NGF and
FGF-2 in DRGs have already been stated (Cobianchi et al., 2013; Grothe et al.,
1996; Meisinger and Grothe, 2002).
Since the regulation patterns of the analyzed growth factors are comparable in
conduit treated animals as well as control animals, it can be concluded that chitosan
does not affect these molecular processes.
6.1.2 Matrix formation and brain-derived neurotrophic factor (BDNF) presence in the tube content Following an implantation time of 18 days, a matrix formation was seen in our
short-term study, which was not further analyzed, since we collected the entire tube
content for Western Blot analysis. Our partner laboratory at the Lund University
(ULUND), however, examined the matrix in a separate study (see (Haastert-Talini et
al., 2013)). They evaluated the formed matrix after 21 days with regard to its
thickness, axonal outgrowth, and presence of SCs. The latter were seen within the

Discussion-Project 1
- 110 -
matrix along the complete conduits of different DAs, but DAI as well as DAII tubes
contained less apoptotic SCs in the distal region than DAIII conduits. Analyses of
matrix thickness and axonal outgrowth did not show significant differences between
the tubes, but in all experimental groups axons had almost completed the growth
across the 10 mm defect. This kind of matrix formation has previously been
described as an important phase on the way to successful regeneration. One week
after conduit implantation, formation of a fibrin cable has been reported, along which,
amongst others, SCs begin to migrate within the second week from the proximal and
distal nerve stumps, thereby aligning to form the bands of Büngner. Subsequently,
axonal sprouts enter the conduit 7-14 days after repair and use the matrix as a
guidance structure while crossing the defect. The fibrin cable itself, however, is
believed to vanish after fulfilling its duty during cell migration (Belkas et al., 2004a;
Dahlin et al., 1995; Daly et al., 2012; Lundborg et al., 1981; Madison et al., 1987;
Seckel, 1990; Williams et al., 1983).
From the matrix formation seen regardless of the DA it can be concluded that the
early regenerative processes described in hollow conduits are allowed in the
presence of chitosan as well. The reason for the higher number of apoptotic SCs in
the presence of chitosan of a high DA (about 20 %) is not clear. A good
biocompatibility of chitosan membranes and SCs has been shown by our laboratory
together with a partner laboratory (UMINHO, Portugal) in vitro, however, the highest
DA tested was 5 % (Wrobel et al., 2014). Other studies undertaking SC seeding on
chitosan membranes and fibers could further demonstrate viability and migration if
the DA was around 8 % (Yang et al., 2009; Yuan et al., 2004). Interestingly, Wenling
et al. have reported that chitosan films with a DA lower than 10 % are a more
suitable substrate for spreading and proliferation of SCs (Wenling et al., 2005). While
a DA of 20 % resulted in a spherical shape of the SCs, they displayed a polar
morphology on lower DAs of e.g. 5 %. Additionally, more SCs attached on films of a
low DA. This is in agreement with observations that cells in general prefer hydrophilic
surfaces with positively charged amine groups, which are properties that both
account for chitosan with low DAs, while chitin is highly hydrophobic and displays
less amine groups (Aranaz et al., 2009; Dutta et al., 2004; Ma et al., 2002; Tomihata
and Ikada, 1997). The cause for the change in morphology, however, is not known

Discussion-Project 1
- 111 -
as well as what kind of physiological changes cause an increase in apoptosis in the
presence of chitosan of a high DA.
Furthermore, BDNF was chosen as an exemplary NTF to verify its secretion into the
conduit after nerve repair. Five and 18 days following surgery, BDNF was detected in
the pooled tube content with no difference between the DAs. mRNA level of BDNF
have been reported to increase 3 days after sciatic nerve transection before reaching
maximal levels 3-4 weeks later (Boyd and Gordon, 2003a; Meyer et al., 1992). With
regard to protein levels in the tube content, however, no such increase could be
seen from day 5 to 18 in our study, but an increase in proBDNF levels. The presence
of BDNF in the tube matrix is of importance since it has e.g. been reported to be
anterogradely transported in sensory neurons, thus accumulating in the tip of the
proximal nerve stump following nerve injury, before probably being released to
support prevention of axotomy induced death (Tonra et al., 1998). Moreover, BDNF
has been shown to enhance motoneuron outgrowth in vitro using an outgrowth
assay of spinal cord slices in a collagen matrix (Allodi et al., 2011).
6.1.3 Reinnervation of distal muscle target Following PNIs, functional recovery occurs if injured neurons successfully send their
axons out and connect to distal targets subsequently recreating motor endplates and
reestablishing appropriate reinnervations. Improvement of functionality is the most
important factor for the patient (Fugleholm et al., 2000; Valero-Cabré et al., 2001).
Therefore, to evaluate the progress regarding this parameter, motor recovery was
assessed by means of SSI and electrophysiological recordings.
6.1.3.1 Behavior
Since the introduction of the SSI by Bervar, when it was described as a time-saving
alternative to the sciatic function index (SFI), this method has been widely used in
our and other laboratories (Bervar, 2000; Bozkurt et al., 2011, 2008; Dornseifer et
al., 2011; Korte et al., 2011; Luís et al., 2008). In the present study, however, we
could not detect significant improvement over time regarding motor recovery using
the SSI, while it was demonstrated and reflected in the electrophysiological results.
Such inconsistency of results, obtained by both parameters, has been described
before. In a previous study, it has been shown that SSI and electrophysiological
results only correlate in cases of nerve crush, while following gap introduction and

Discussion-Project 1
- 112 -
subsequent regeneration, false negative results are reported (Korte et al., 2011).
This phenomenon has already been stated for the SFI when results did not correlate
with histological findings (Shenaq et al., 1989). Accordingly, the usefulness of this
evaluation method has to be questioned with regard to future experiments that
investigate treatment strategies in connection with nerve transection.
Importantly, however, it is known that the functional outcome following nerve
transection using nerve conduits or ANG treatment is often unsatisfactory for the
patient (Isaacs, 2010). This can be attributed to the loss of endoneurial tubes as
guidance structures upon nerve transection and the consequence that axons need to
find their way to and through the distal nerve stump to ultimately reestablish
connection with correct target organs (Bozkurt et al., 2008; Fugleholm et al., 2000;
Ransom, 2012; Thomas, 1989; Valero-Cabré et al., 2004). Unfortunately,
electrophysiological as well as histological data do not provide information regarding
accurateness of target reinnervation. Accordingly, quantitative target reinnervation
and remyelination represent no guarantee of a successful complex functional
recovery. Therefore, the inconsistency of results obtained from SSI and
electrophysiological as well as histological investigations can be explained by
quantitative, but not appropriate, reinnervation of distal muscle targets (Fugleholm et
al., 2000; Navarro and Udina, 2009; Sarikcioglu et al., 2009; Valero-Cabré et al.,
2001).
To allow a better functional evaluation of peripheral nerve regeneration, various
studies have tested alternatives to the SSI and SFI in the last decades, thereby
focusing on assessment of locomotor function by video gait analysis (Costa et al.,
2009). This method allows the determination of the angle formed by the knee, ankle,
and feet. Therefore, animals are trained to walk on a treadmill. Following sciatic
nerve injury, a decrease of the angles is observed, which then gradually increases
again with proceeding regeneration. Also, different gait cycle phases (e.g. initial
contact, toe off, mid swing) can be evaluated. Importantly, this method has been
suggested to be more sensitive than SFI in detecting motor recovery (De Boer et al.,
2012; Kokai et al., 2011; Lin et al., 1996; Patel et al., 2007). Furthermore, Bozkurt et
al. described an automatic gait analysis system (“CatWalk”), which allows the
quantification of dynamic parameters such as intensity of paw prints. In a similar

Discussion-Project 1
- 113 -
manner, another study used a Tescan pressure sensor grid to determine timing and
contact variables (Boyd et al., 2007; Bozkurt et al., 2008). Thereby, they could show
that animals decrease contact pressure on the injured hind limb following sciatic
nerve injury, while the contact force on the contralateral side increases. Still, finding
the appropriate evaluation method for functional recovery remains challenging.
Another possibility is the use of a forelimb rather than the sciatic nerve rat model
(Costa et al., 2009). The median nerve model e.g. offers the opportunity to perform
electrophysiological recordings, but also grip strength tests can be carried out. For
this test the rat / mouse can be held by the tail, while grasping a bar and the maximal
force generated when pulling the tail is recorded (Park and Höke, 2014).
Furthermore, a skilled walking test can be executed, during which animals have to
walk across a horizontal ladder with variable spacing of the rugs (Costa et al., 2009;
Metz and Whishaw, 2009) Wang et al. additionally reported the possibility to assess
wrist kinetics using a 2D motion analysis (Wang et al., 2008).
6.1.3.2 Electrophysiological recordings
Nevertheless, electrophysiological recordings and histological evaluation are
indispensable and provide useful information regarding the progress of regeneration
as well as regarding the amount of axons reaching distal muscle. Thereby, repeated
measurements can be performed since non-invasive recordings are not harmful to
the regeneration process and animals only have to be anesthetized (Navarro and
Udina, 2009). Four weeks after surgery, however, we did not see any CMAPs. This
observation is in line with reports that axons reach the distal end of a conduit (10 mm
gap) approximately after 3 weeks, a time frame that was also reported by our partner
laboratory (ULUND), who described axonal outgrowth into the chitosan tube for
about 7.9-8.6 mm after 3 weeks (Haastert-Talini et al., 2013; Williams et al., 1983).
Axons regrow with a rate of 1-3 mm per day. But due to retrograde axonal
degeneration as well as staggered growth at the suture site, a process that is
associated with progressive reinnervation of appropriate muscle pathways by motor
axons, the distance of the defect is not overcome earlier (Al-Majed et al., 2000; Daly
et al., 2012; Gordon, 2009; Gordon et al., 2003; Huelsenbeck et al., 2012; Kimura,
1984). Second measurements after 9 weeks, on the other hand, revealed
reinnervation for most of the animals that had received an ANG (TA: 62 %, PL:
80 %), while this was only the case for 6–29 % (TA) or 33-50 % (PL) of the

Discussion-Project 1
- 114 -
experimental groups. Last non-invasive as well as final invasive recordings
demonstrated successful reinnervation also in a higher number of animals that had
received chitosan conduits as treatment and which approached the level of
reinnervation seen with ANG treatment. The delay seen in nerve repair using hollow
chitosan conduits might be due to missing guidance structures for the distance of the
defect. Axonal elongation following ANG repair is eventually faster since the tissue
serves as a substratum for the growth cones and the SCs of this denervating
segment take part in the creation of a regeneration promoting microenvironment
(Wang et al., 2001). CMAPs selectively give information regarding function and
regeneration of α-motoneurons (Navarro and Udina, 2009). Using chitosan conduits
of different DAs, nerve reconstruction was comparably successful with respect to
skeletal muscle reinnervation by these motoneurons as seen with ANGs. No
differences were found within the treatment groups with regard to NCV, AxL (AUC),
or amplitude, but the level of healthy control values was not reached. This
observation is in line with previous results for treatment of nerve defects of 10-
14 mm length using hollow guidance conduits based on materials like PLGA / PCL,
PLGA / Pluronic F127, silk fibroin, collagen, but also chitin or chitosan (Dornseifer et
al., 2011; Oh et al., 2008; Panseri et al., 2008; Yang et al., 2007). The amplitude of
the measured CMAP provides information about the number of regrown motor axons
as well as regarding the size of the motor unit. But, a full amplitude recovery does
not necessarily mean that regeneration of all motoneurons was successful. Some
axons fail at reaching distal muscle targets and subsequently allow regenerating
axons to branch, thereby increasing the degree of reinnervation (Archibald et al.,
1991; Fugleholm et al., 2000; Navarro and Udina, 2009; Pfister et al., 2011; Valero-
Cabré et al., 2001).
The observation that measurable PL innervation occurred earlier than TA innervation
is hard to explain since CMAPs are expected to occur first in more proximal target
muscles (Navarro and Udina, 2009; Valero-Cabré et al., 2001). TA and PL muscles
are innervated by different branches of the sciatic nerve, but still it would be more
reasonable to find a first detectable signal in the more proximal muscle. A stronger
recovery of the TA amplitude compared to the PL amplitude as seen in this study
has already been reported before and can likely be explained by less axons reaching
the more distal muscle (Valero-Cabré et al., 2004, 2001). The latency, with which the

Discussion-Project 1
- 115 -
signals arrived at the muscles, was as expected higher for recordings from the more
distal target (PL) and reflects the conduction time of the largest recruited motor fibers
and therefore the degree of myelination and the size of the axons (Archibald et al.,
1991; Navarro and Udina, 2009; Valero-Cabré et al., 2001). Additionally, no
significant differences between the experimental groups including the control group
(ANG) were found. Contralateral intact control values, though, were superior to all
results obtained by the different bridging strategies like seen before in similar
experimental settings (Mancuso et al., 2011; Valero-Cabré et al., 2001). It has
already been previously noted that the latency in general is prolonged for a long time
after nerve lesion and differences are rarely detected between treatment groups.
This is an observation that also accounts for NCVs (Navarro and Udina, 2009;
Valero-Cabré et al., 2001). Furthermore, the current needed to elicit the maximal
CMAP when recordings were undertaken from the TA muscle was not different
between the treatment groups but differed to contralateral healthy control values.
Interestingly, only for PL recordings and only with DAII conduits, the current needed
to elicit the maximal CMAP was comparable to contralateral control values.
6.1.3.3 Morphological evaluation
Distal nerve stumps harvested upon explantation were sent to our partner laboratory
(UNITO, Italy) and subjected to morphometric analysis. Results are published in
Haastert-Talini et al. (2013) and show that in general the total number of myelinated
nerve fibers in all groups is higher than on the contralateral intact control side, but
only DAI treatment resulted in significantly more fibers in comparison to the ANG
group. The total numbers of myelinated fibers in healthy and ANG samples reported
are in line with morphometric results described before (Daly et al., 2013; de Ruiter et
al., 2008; Haastert-Talini et al., 2013; Luis et al., 2007). For hollow tube treatment,
however, lower numbers have been reported in former studies using e.g. PLGA or
poly (caprolactone fumarat) (PCLF) conduits (Daly et al., 2013; de Ruiter et al.,
2008), while other studies have stated similar numbers using PLGA or PCL conduits
(Luis et al., 2007; Varejão et al., 2003) as bridging material for 10 mm gaps.
Nevertheless, excessive sprouting as seen with DAI conduits is undesirable. Shortly
after lesion, lateral budding starts close to the injury site resulting in up to 25
branches per axon. While branches are pruned off with time, this is eventually not
the case for all of them. Consequently, single axons are often misguided and

Discussion-Project 1
- 116 -
reinnervate several muscles via their branches, thereby often eliciting antagonizing
effects. Also, muscles (hyperinnervation) as well as motor endplates
(polyinnervation) are targeted by two or more motoneurons. Altogether, these
processes increase the chance of disturbed motor coordination (Ijkema-Paassen et
al., 2002; Klimaschewski et al., 2013; Valero-Cabré et al., 2004, 2001). The reason
for increased sprouting in the presence of chitosan with a low DA, however, is not
clear. Investigation of axon and fiber diameters as well as myelin thickness revealed
differences to healthy values but no differences were seen among the treatment
groups, while no differences at all were detected regarding the g-ratio (axon
diameter / fiber diameter). Comparable results regarding these parameters have
been already reported bridging 10-14 mm gaps amongst others with chitosan
conduits, but also collagen, silk fibroin, PCL, PCLF and PLGA (Bini et al., 2004; Daly
et al., 2013; de Ruiter et al., 2008; Dornseifer et al., 2011; Varejão et al., 2003; Wang
et al., 2010, 2001; Yang et al., 2007).
6.1.3.4 Muscle weight
To further assess the reinnervation of distal muscle targets, the TA and
gastrocnemius muscles were harvested and their weight determined. No differences
were found between the experimental groups indicating comparable reinnervation
following ANG and chitosan conduit implantation. Nevertheless, none of the groups
reached healthy control values. The latter is in accordance with previous studies
testing PGA, PLGA, PCLF, collagen, but also chitosan conduits in 10-14 mm gap
models, while in some cases the conduits could not reach ANG levels (Alluin et al.,
2009; Daly et al., 2013; Dornseifer et al., 2011; Patel et al., 2006; Raisi et al., 2012;
Waitayawinyu et al., 2007).
Altogether, the functional, morphometric, and muscle weight data point out that
chitosan conduits of all DAs allow comparable results as the standard therapy
(ANG). But, an important finding is that DAI conduits seem to encourage excessive
sprouting and are therefore not convenient for further experiments. Also, healthy
control values have not been reached and a recovery time of 13 weeks was not
enough for nerve maturation to pre-surgery levels. This discrepancy, however, has
frequently been reported for hollow conduits of varying materials including those that
have received FDA and/or CE approval and is thought to only be overcome with the

Discussion-Project 1
- 117 -
right intraluminal guidance structure or filling materials that feature appropriate
textures and cues (Daly et al., 2013; Oh et al., 2008; Panseri et al., 2008;
Waitayawinyu et al., 2007; Yang et al., 2009).
6.1.4 Foreign body response (FBR) upon chitosan implantation degradation Chitosan has already been widely investigated for its use in tissue engineering, but
limitations were encountered due to its poor mechanical properties (Freier et al.,
2005c; Madihally and Matthew, 1999; Siemionow et al., 2010; Xie et al., 2008; Yang
et al., 2004). Notably techniques, however, have been adjusted by our partner
Medovent GmbH that make it possible to control the mechanical properties (Freier et
al., 2005c). 18 days, but especially 13 weeks after implantation, however, fissures
and cracks were displayed by the chitosan conduits with the highest DA (20%).
Additionally, it was partly observed that the conduits had completely collapsed and
thereby partly damaged newly grown nerve tissue. An accelerated degradation rate
of these conduits was expected, since lysozymes, the enzymes responsible for the
degradation of chitosan in our body, hydrolyze the ß(1-4) linkages between N-acetyl
glucosamine and glucosamine in chitosan as well as chitin and the degradation rate
is consequently in proportion to the DA (Han et al., 2011; Kean and Thanou, 2010;
Wang et al., 2010; Zhang and Neau, 2001). Previously, Tomihata and Ikada have
reported that about 20 % of chitosan with a DA of 16 % was degraded 12 weeks
upon implantation into a pouch in sub dermal tissue, while almost no weight loss was
seen for chitosan with a DA 10 % (Tomihata and Ikada, 1997). In line with this in our
study, conduits with a middle or low DA (5 and 1 %) did not show any macroscopic
signs of degradation. Interestingly, Wang and colleagues, who also used chitosan
(DA= 6.5 %) as a bridging material, have stated that the conduit had barely degraded
after 12 weeks (Wang et al., 2010). They further reported on a thin fibrous tissue
capsule that had developed around the conduit like we have seen it. Others,
however, have reported complete degradation 3 or 6 months after implanting
chitosan conduits (DA= 17 % or 7.7 %, respectively) with PLGA or PGA filaments
inside the lumen in the dog sciatic nerve model (Shen et al., 2010; Wang et al.,
2005). It has, however, been taken into account that the DA was higher, the
observation time partly longer, and that they used a different and bigger animal
model, which was proposed to maybe influence the degradation rate (Xie et al.,
2008). Xie and colleagues have worked with the same DA, mixed the chitosan with

Discussion-Project 1
- 118 -
polylactic acid, and tested conduits in a rat sciatic nerve model (Xie et al., 2008).
They detected many cracks and described the conduit as coarse 12 weeks after
surgery.
The thin fibrous capsule around the conduits was further analyzed to look at the
inflammatory response towards the chitosan conduits. In general, it is described that
upon implantation of a medical device, plasma proteins (e.g. fibrinogen, fibronectin)
are adsorbed to the material and likely undergo conformational changes. This results
in the exposition of binding sites for immune cells, thus initiating the FBR, which
serves as protection of the body and restructuring of the affected, injured area. Acute
inflammation, which is marked by infiltration of neutrophils within hours to days after
injury, is followed by chronic inflammation. The latter is characterized by the
formation of granulation tissue, a stage of repair and hallmark of healing, which
features macrophages, monocytes, lymphocytes, and collagen as well as
proliferating blood vessels. Additionally, MGCs partly surround the foreign material
through fusion of macrophages with the amount of present cells depending on the
form and topography of the material. In case of successful tissue reconstruction, the
last stage of the FBR is the formation of a collagenous and vascular capsule; if,
however, the immune reaction towards the material is not fading, foreign body
granuloma can occur, which are comprised of modified macrophages (epithelioid
macrophages) (Anderson, 2001; Anderson et al., 2008; Hu et al., 2001; Kisseleva
and Brenner, 2008; Morais et al., 2010; Thevenot et al., 2008). Eighteen days after
implantation of hollow chitosan conduits, the inner side of the formed capsule, which
faced the material surface, featured macrophages as well as few MGCs, a clear sign
of chronic inflammation. Furthermore, collagen forming cells were seen indicating
that tissue remodeling was in process. Interestingly, the MGCs did not contain many
nuclei (less than 20), which were mostly located in the periphery. These
characteristics are known for giant cells of the Langhans-type and are usually
associated with e.g. sarcoidosis and tuberculosis rather than the FBR. The latter
usually includes formation of FBGCs, which feature more and randomly distributed
nuclei. The possibility that Langhans-type cells are precursors of FBGCs has,
however, been discussed (Rhee et al., 1978). Also, following subcutaneous
implantation of cellophane foil, formation of both Langhans-type and foreign body
MGCs has been reported (Smetana, 1987). Moreover, McNally and Anderson noted

Discussion-Project 1
- 119 -
that a gray area exists between these two variants of MGCs and that further studies
are mandatory to elucidate different phenotypes and functions of different types of
MGCs (McNally and Anderson, 2011). Interestingly, following an implantation time of
13 weeks, the number of activated macrophages was significantly decreased, thus
pointing towards a reduction of the inflammatory response. Instead, dense regular
collagen fibers were visible in the contact area with the material, while vessels were
found in the area averted from it. Additionally, only single MGCs were found after 13
weeks in two animals that had received DAI conduits for nerve reconstruction. It was,
however, rather expected that these cells would be present in increased numbers in
the DAIII group, since degradation in this group was already in progress, but this was
not the case. Furthermore, although we could see macroscopic differences in the
optical density of the tissue, no significant differences were seen when it was
measured. The capsule displayed a thickness of 200-300 µm, which is slightly
thicker than known from literature (50-200 µm) following e.g. chitosan implantation in
a subcutaneous air pouch model or nerve reconstruction with poly(2-hydroxyethyl
methacrylate-co-methyl methacrylate) nerve conduits (Barbosa et al., 2010; Belkas
et al., 2005; Morais et al., 2010). On the other hand, it has been also reported that
following subcutaneous implantation of poly(L-lactide acid) (PLLA) the thickness of
the surrounding capsule is thicker during the first 6 month and then decreases to 100
to 200 µm (Mainil-Varlet et al., 1996). Previously, various studies have reported the
lack of a visible inflammation response towards chitosan or chitosan in combination
with other materials like PGA, PLGA (Raisi et al., 2012; Shen et al., 2010, 2009;
Wang et al., 2005). Others, conversely, have described the formation of fibrous
capsules (Simões et al., 2011; Tomihata and Ikada, 1997; Wang et al., 2010).
Simões et al. (2011) subcutaneusly implanted chitosan membranes, which had been
subjected to different drying methods (37°C vs. freeze drying) and ultimately
displayed variances in their porous microstructure. Eight weeks afterwards, they
detected a mild chronic inflammation in form of a collagenous fibrous capsule (37°C)
or a strong granulomatous reaction (freeze drying) depending on the production
procedure. In a similar experimental setting, Tomihata and Ikada described a mild
tissue reaction towards chitosan membranes with varying DAs (DA: 0, 20, 16 or
27 %), while there was strong reaction towards chitin and chitosan with a DA of 31 %
due to rapid degradation (Tomihata and Ikada, 1997). Analogous results were
obtained by Hidaka et al., who saw a mild tissue reaction following chitosan

Discussion-Project 1
- 120 -
membrane implantation with a DA around 0 % or 6 %, while the initial inflammatory
response was stronger with a DA between 20 % and 35 % (Hidaka et al., 1999).
Furthermore, they speculated that the more negatively charged surface of chitosan
with a higher DA lead to an increased plasma protein adhesion, thus triggering the
inflammtion by higher acetylated chitosan derivatives. Another study, in which
chitosan with a DA of 4 % and 15 % was implanted subcutaneously, supported these
findings. Higher recruitment numbers of inflammatory cells to the implant site and
also increased adherance of these cells to the material was reported in the presence
of a higher DA. Furthermore, they also stated that the capsule around 15 % DA
chitosan was thicker (Barbosa et al., 2010). Also, in the presence of other
biodegradable materials, the formation of a fibrous capsule has been reported as
well as formation of MGCs. A study testing PLLA / plasticizer triethylcitrate
membranes reported that MGCs were present in the developed capsule after 7 days
and increased in number up to the 180th day of observation (Maluf-Meiken et al.,
2006). This is contrast to our results, where we saw a decrease or rather none of
these cells at all except for samples of two DAI animals. Other studies implanting
PLLA materials reported the presence of MGCs for weeks to months and also the
eventual formation of a fibrous capsule (Mainil-Varlet et al., 1996; Pistner et al.,
1993).
Although we did not see differences in the inflammatory response between the
experimental groups with regard to analysis of the fibrous capsule that had
developed around the conduits, other observations were made when analyzing the
regenerated nerve tissue. Counting of MGCs also did not reveal differences between
the conduits and also activated macrophages were equally present in the nerve
tissue after 18 days, but 13 weeks after implantation most ED1-positive cells were
found in samples of the ANG and DAIII groups. The latter results were obtained by
Sandra Wrobel ( MSc, Institute of Neuroanatomy, MHH) and can be looked up in
((Haastert-Talini et al., 2013)). This observation can probably be assigned to the
presence of chitosan fragments that were detected in 5 out of 5 analyzed animals in
the DAIII group after 13 weeks. Interestingly, already after 18 days, fragments were
found in this group, supporting the finding of very fast degradation of these conduits
and their risk for newly formed nerve tissue, although those fragments were mainly
localized in the outer tissue layer. Activated macrophages, however, were present in

Discussion-Project 1
- 121 -
higher numbers 18 days after implantation than after 13 weeks indicating a reduction
of the inflammatory response as it was already described for the fibrous capsule.
The great number of macrophages after 18 days, though, is in line with the
understanding that after nerve lesion, nerve stumps are highly invaded by
hematogenous macrophages, which also infiltrate the fibrin matrix from both sides.
They secrete growth factors, pro- as well as anti-inflammatory cytokines, and are
thus of importance for debris clearing, but also for promotion of SC proliferation and
axonal regeneration in early phases of regeneration (Díaz-Flores et al., 1995; Kiefer
et al., 2001; Zochodne and Cheng, 2000).
Altogether, we saw a mild tissue reaction towards chitosan conduits in the formation
of a thin fibrous capsule. This kind of FBR, however, has been frequently described
in studies testing various materials and does not endanger successful nerve tissue
regeneration. The observation of fragments of DAIII conduits in the nerve tissue
already after 18 days as well as after 13 weeks, on the other hand, bears a risk for
the newly formed nerve tissue and is a criterion of exclusion for conduits with a high
DA. Together with the finding of increased sprouting with DAI conduits, which is
undesired since it provokes hyper- and polyinnervation as well as innervation of
wrong targets, it can be concluded that hollow DAII chitosan conduits are the eligible
candidate for bridging small defects.
6.1.5 Concluding remarks-Chances of hollow chitosan conduits on the existing market The market of conduits for peripheral nerve repair is growing and different materials
have already gained approval by the FDA or CE to be used in clinical applications
with NeuraGen® (collagen), Neurotube® (PGA), and Neurolac® (PCL) representing
the mostly described and applied products. Nevertheless, none of the marketed
conduits so far has encountered unlimited acceptance and although it is often
suggested that these conduits can be used for gaps of up to 3 cm length, it was also
proposed that the realistic bridgeable distance is probably less (Isaacs, 2010). Just
recently, Liodaki et al. described insufficient nerve regeneration in four patients using
NeuraGen® (Liodaki et al., 2013). In agreement, successful sensory and motor
recovery was only seen when the defect was 6 mm or less in a multicenter clinical
trial using NeuraGen® (Boeckstyns et al., 2013). Furthermore, case studies revealed

Discussion-Project 1
- 122 -
neuroma formation using this type of conduit (Moore et al., 2009). The same authors
also described a failed case of regeneration through a Neurotube® conduit. In this
case, the conduit was fully degraded after 9 months and a gap defect between the
proximal and distal stump was left. In general, variable results were obtained so far
with this conduit, in total emphasizing that successful regeneration is only seen in
small gaps (Berger et al., 1994; Isaacs, 2010; Kehoe et al., 2012; Meek and Coert,
2008; Navissano et al., 2005). With regard to Neurolac®, complications have been
reported. Meek and den Dunnen e.g. used porous Neurolac® conduits for nerve
reconstruction in rats and described fragmentation as well as severe swelling and
inflammation (Meek and Den Dunnen, 2009). Implantation of less porous conduits
resulted in not as much swelling, but fragments were still found after 16 as well as 24
months, although the material was no longer macroscopically visible at the latter time
point. These fragments, which were accompanied by MGCs and macrophages,
however, where only found in the epineurium of the regenerated nerve and did not
influence the nerve regeneration (Jansen et al., 2004; Meek and Jansen, 2009).
Nevertheless, it was also noted that the conduit might be too stiff, thus causing
needle breakage and problems in case of positioning over joints. Also, in a clinical
case, neuroma formation was described upon collapse of the conduit (Chiriac et al.,
2012; Hernández-Cortés et al., 2010; Kehoe et al., 2012; Meek and Coert, 2008).
Given the limitations of already approved conduits, the here tested hollow chitosan
conduits of a DAII can most likely compete on the market and a first clinical trial is in
planning for bridging of small defects. Advantages of hollow DAII conduits include
their transparency, which so far is only displayed by the Neurolac® conduit, as well
as a mild tissue reaction. Nevertheless, so far we do not know when and in which
time frame the DAII conduits will degrade. Therefore, we currently perform long-term
studies in the rat sciatic nerve model addressing this issue. Furthermore, it is unclear
how the conduit would perform if the nerve defect was localized over a joint, where
kinking could cause damage to the nerve tissue. To conclude, the hollow DAII
chitosan conduit with a DA of 5 % has the best requirements to get a place on the
market of peripheral nerve reconstruction of small defects, but some questions
remain to be answered in the future.

Discussion-Project 1
- 123 -

Discussion-Project 2
- 124 -
6.2 Project 2 6.2.1 Schwann cells (SCs) as vehicles for gene delivery Due to their multifaceted and significant role during peripheral nerve regeneration,
the delivery of NTFs to the site of injury has been extensively studied and different
methods have been applied to make them available in a sustained manner. This
includes e.g. NTF-loaded nerve guidance conduits as well as application via
introduction of microspheres, osmotic minipumps, or matrices such as gels,
sponges, and filaments, but also transplantation of physiological or genetically
modified cells has widely been investigated (Belkas et al., 2004b; Haastert et al.,
2006a; Hadlock et al., 2000; T. Ikeda et al., 2014; Jungnickel et al., 2011; Kokai et
al., 2011, 2010; Madduri et al., 2010; Pabari et al., 2011; Pfister et al., 2007; Timmer
et al., 2003; Wood et al., 2013a). SCs, the glial cells of the PNS, proliferate, loose
their myelin, migrate to the injury site, phagocytize myelin debris, guide regrowing
axons, and represent a major source of NTFs. Accordingly, their transplantation has
been suggested to have a beneficial effect on peripheral nerve regeneration.
Consequently, autologous naïve as well as genetically modified and NTF
over-expressing SCs are candidates for artificial biohybrid nerve devices (Madduri
and Gander, 2010; Pfister et al., 2007; Reichert et al., 1994). Advantages of this
delivery method include long-term availability at the site the injury, the lack of
non-physiological degradation products, and the ability of these cells to respond to
changes in the environment. A drawback regarding SC transplantation, however, is
the ex vivo proliferation and purification procedure of these cells (Di Summa et al.,
2011; Dove, 2002; Madduri and Gander, 2010; Pfister et al., 2007). Therefore,
alternative cell sources including e.g. OECs, BMSCs, and ADSCs have also been
investigated in connection with peripheral nerve repair and revealed promising
results (Ao et al., 2011; Di Summa et al., 2011; di Summa et al., 2010; Gu et al.,
2014; Radtke et al., 2011, 2009; Wei et al., 2011). For example, BMSC-derived SCs
have been reported to allow similar nerve regeneration as seen with sciatic nerve
derived SCs when seeded into a chitosan conduit (Ao et al., 2011). Also, chitosan /
silk fibroin-based nerve grafts enhanced regeneration when ADSCs were included
for treatment of a 10 mm gap in the rat, but ANGs were not included as a positive
control (Wei et al., 2011). In another study, however, results obtained with fibrin
conduits containing ADSCs differentiated into a SC-like phenotype were comparable

Discussion-Project 2
- 125 -
to those with ANGs (Di Summa et al., 2011). Interestingly, in the scope of a short-
term study, it was demonstrated that SCs hold a higher regenerative potential
compared to ADSCs as well as BMSCs (di Summa et al., 2010). Due to their multi-
functional role in the PNS and in response to PNIs, SCs therefore remain ideal
candidates for transplantation experiments (Madduri and Gander, 2010). Collagen
nerve guidance conduits seeded with SCs have already been shown to enhance
nerve regeneration in a 18 mm rat sciatic nerve rat model and autogenous veins
containing SCs performed better in a 40 mm tibial nerve rabbit model than empty
veins, although they could not compete with ANGs (Ansselin et al., 1997; Pfister et
al., 2007; Zhang et al., 2002). Autogenous venous nerve conduits were further
reported to successfully bridge a 6 cm gap in rabbits when SCs in Matrigel were
included, but no functional tests were conducted in this case (Strauch et al., 2001).
Also, Hadlock et al. could show an improved performance of small intestinal
submucosa-based nerve guides by adding SCs, although the loaded conduits did not
reach ANG levels in a 7 mm gap with regard to SFI and extensor postural thrust
testing (Hadlock et al., 2001). In another study, bridging of a 6 mm gap was most
successful with SC-seeded collagen conduits and combined administration of an
immunosuppressant (FK506) (Udina et al., 2004). Also, genetically modified SCs
have already been successfully transplanted and demonstrated to have regeneration
promoting effects (Haastert et al., 2006a; Timmer et al., 2003).
6.2.2 Recovery of mechanical pain sensibility In the scope of the second study, the battery of tests was extended to allow a more
widespread evaluation of the performance of the tested conduits. Therefore, the Von
Frey test was conducted to assess the mechanical pain sensibility since
hyperalgesia (a decrease in pain threshold), has been reported in patients following
nerve lesions (Bennett and Xie, 1988; Bridges et al., 2001; Casals-Díaz et al., 2009;
Cobianchi et al., 2014; Kingery et al., 1994; Sandkühler, 2009). For this test, animals
were placed in plastic compartments on a metallic mesh and medial as well as
lateral paw sites was stimulated with a non-noxious mechanical stimulus to
determine return of sensibility. Basis for this test is that innervation of muscles
proximal to the knee is preserved upon sciatic nerve injury, thus allowing a
withdrawal response in case of touch sensation (Cobianchi et al., 2014). Under
healthy conditions, tibial and sural branches of the sciatic nerve as well as the

Discussion-Project 2
- 126 -
saphenous nerve provide sensory innervation to the medial paw site, while the
lateral area is exclusively innervated by the sciatic nerve (Cobianchi et al., 2013).
Due to these innervation patterns, it is of importance to differentiate between
responses obtained from different paw regions or interpretation of results can be
misleading (Cobianchi et al., 2014).
Following lesion of the sciatic nerve and subsequent repair using either ANGs or
chitosan conduits containing NVR-Gel, which was supplemented with naïve or
genetically modified SCs, respectively, hyperalgesia was seen when stimulating the
medial paw territory on the lesioned side compared to the respective healthy
contralateral side. This observation is in line with results obtained by Cobianchi et al.
(2013), who reported increased sensitivity in the same tests following end-to-end
repair in the rat sciatic nerve model. Also, as a result of nerve transection and
excision of 2 mm from the distal stump mechanical hyperalgesia was observed for
weeks (Casals-Díaz et al., 2009). The reason for the mechanical hyperalgesia after
sciatic nerve injury can be found in collateral sprouting from the intact neighboring
saphenous nerve into the area that was formerly innervated by the sciatic nerve.
This process is known as adjacent neuropathic hyperalgesia (ANH) and is promoted
by NGF secretion by the denervated tissue. In case of successful target-organ
reinnervation through the sciatic nerve, the increased sensitivity might be reversed or
attenuated (Casals-Díaz et al., 2009; Cobianchi et al., 2014, 2013; Diamond et al.,
1987; Kingery and Vallin, 1989; Kingery et al., 1994). With the results obtained in this
study, however, we do not see a reduction of ANH including the ANG group. This is
in agreement with reports that, as a consequence of nerve transection with distal
segments exclusion, ANH is described for at least 12 weeks and normal sensitivity is
not restored for about 2 months after suture repair (Cobianchi et al., 2014, 2013;
Kingery et al., 1994). The persisting hypersensitivity also in case of reinnervation has
further been proposed to be the result of chronic sensitization of the saphenous
nerve pathway including spinal cord circuits (e.g. via unmasking of pre-existing
connections or changes involving inhibitory interneurons) (Casals-Díaz et al., 2009;
Cobianchi et al., 2014; Navarro et al., 2007; Sandkühler, 2009; Sotgiu and Biella,
1995).

Discussion-Project 2
- 127 -
Stimulation of the lateral paw site did not provoke withdrawal response in any of the
tested animals 7 days as well as 4 weeks after nerve lesion indicating complete
denervation of this territory as expected. In all animals of the ANG group, however,
return of sensation was seen after a time period of 9 weeks, but the response could
already be detected upon light stimulation representing hyperalgesia. In similar
manner, single animals of the experimental groups responded to stimulation. One
animal each of the nt-neoSCs and tf-neoSCs-GDNF group as well as a total of 3
animals of the tf-neoSCs-FGF-218kDa group showed signs of sensory recovery. The
general observation that after nerve injury no response is seen is in agreement with
prior findings and can be attributed to the sole innervation of this paw site through
the sciatic nerve in healthy animals. Recent testing of hollow chitosan conduits in a
15 mm gap model by our partner laboratory revealed similar withdrawal responses
(Talk: “Tubulization with chitosan guides for the repair of long gap peripheral nerve
injury in the rat” by Francisco Gonzalez-Perez from UAB at the 2nd International
Symposium on Peripheral Nerve Regeneration in Turin, Italy). Based on experiments
that included cutting of the saphenous nerve, the hypersensitivity in the lateral paw
site after 4 weeks was attributed to the arrival of regenerating axons to this territory
and not ANH (Cobianchi et al., 2014).
Interestingly, Cobianchi et al. could show that after nerve transection, the number of
peptidergic intra-epidermal nerve fibers was reduced in lateral and medial paw
territories (Cobianchi et al., 2014). First regenerating axons were seen reaching the
epidermis 4 weeks following surgery, while massive reinnervation was observed
after 2 months with numbers of peptidergic fibers, which have been described to be
involved in the signaling of noxious pain and maintenance of hypersensitivity,
exceeding healthy control values. Furthermore, they could ascribe the early
hyperalgesia in the medial paw region to saphenous peptidergic fiber sprout growth
in the subepidermal plexus, while the decreased pain threshold seen for stimulation
of the lateral paw region correlated with skin reinnervation.
Altogether, the obtained results show reinnervation of distal target-organs in the
ANG group as indicated by withdrawal responses seen earliest 9 weeks after nerve
injury upon stimulation of the lateral paw territory on the lesioned side. Regarding the
experimental groups, however, only single animals displayed sensory recovery,

Discussion-Project 2
- 128 -
which mainly belonged to the FGF-218kDa group. The animals that responded to
stimulation of the plantar paw surface could later be correlated to show regenerated
nerve tissue, thus supporting the findings in this test.
6.2.3 Fibroblast growth factor (FGF)-218kDa partly allows reinnervation of distal muscle targets and functional recovery 6.2.3.1 Behavior
As already seen with hollow chitosan conduits to bridge short nerve defects in
project 1, we could not detect any motor recovery in the differently treated groups
using the SSI test. Since only single animals of the experimental groups revealed
nerve tissue regeneration upon explantation, this result is not surprising. Aside from
the fact that this test only provides minimal information in case of studies of complete
nerve transection (see 6.1.3.1), another problem is that sometimes it is difficult to
gain data. It is frequently observed that after nerve transection, rats and mice start to
bite off their nails and phalanges, a behavior that is referred to as autotomy (Costa et
al., 2009; Sarikcioglu et al., 2009). Discussions are ongoing if this self-harming
procedure is the response to painful perception in the injured nerve (neuropathic
pain) or to the feeling of insensitiveness and the need to shed off some kind of
foreign body part. During our study, autotomy was first observed 3 weeks after
surgery, a time point, which has previously been noted as a usual starting point
(Sarikcioglu et al., 2009). With advancing observation period the number of animals
that showed signs of autotomy increased. Thereby, it was noticeable that animals of
the ANG, except for one animal, did not display autotomy, while it was frequently
observed across the experimental groups with most severe cases among the GDNF
group. Also, another study reported severe cases of autotomy when fibrin sealant
containing GDNF was locally applied to a transected sciatic nerve (Jubran and
Widenfalk, 2003). This enhanced toe biting within the GDNF group could support the
theory that autotomy is indeed the reaction towards insensitiveness in this body part
since analgetic, neuropathic pain reducing effects of this factor have been reported
(Boucher, 2001; Boucher et al., 2000).
6.2.3.2 Electrophysiological recordings
Also, in project 2, electrophysiological tests were conducted to elucidate functional
recovery. Reinnervation of distal muscle targets and subsequent motor recovery was

Discussion-Project 2
- 129 -
seen for all animals, which had received ANGs for nerve reconstruction. On the other
hand, CMAPs were only detected in single animals of the experimental groups and
here mainly the FGF-218kDa treatment lead to positive recordings. These results are
in line with reports indicating that FGF-218kDa has a promoting effect on neurite
outgrowth of spinal motoneurons in vitro (Allodi et al., 2013). The authors of this
study further suggested that FGF-218kDa likely exhibits its effect during early stages of
nerve repair by encouraging interactions between polysialic acid (PSA)-NCAM and
FGFR-1 with the latter being highly expressed by SCs accompanying motor neurites.
Furthermore, our laboratory could previously show that FGF-2 in general plays an
important role after PNI and different effects can be attributed to the isoforms;
something that is also supported through the dissimilar regulation of these isoforms
with regard to temporal as well as spatial parameters. While the high molecular
isoform, which has already enhanced nerve regeneration across a 15 mm gap, has
mainly been associated with sensory regeneration and has been found to be mostly
up-regulated in DRGs, the low molecular isoform has been demonstrated to allow
motor recovery and is rather prominently seen close to the injury site (Grothe et al.,
2000, 1996; Haastert et al., 2006a; Meisinger and Grothe, 2002; Timmer et al.,
2003). Moreover, Jungnickel et al. could demonstrate that transgenic mice
over-expressing FGF-2 display an increased number of proliferating SCs as well as
more axons 1 week after nerve crush (Jungnickel et al., 2006). An important function
of FGF-2 was suggested during early phases of regeneration by regulation of SCs
differentiation as well as by directly influencing axons (Fujimoto et al., 1997;
Jungnickel et al., 2004). In addition, FGF-2 was shown to be involved, amongst
others, in remyelination, protection of DRGs from lesion-induced death, and injury-
induced neuronal apoptosis. These different observations emphasize the crucial role
of this factor during peripheral nerve regeneration (Grothe et al., 2006; Haastert et
al., 2006a; Jungnickel et al., 2010, 2006, 2004; Otto et al., 1987). Interestingly, Han
et al. (2010) have reported improved functional outcome when chitosan conduits
(DA ~ 16 %) were filled with heparin-incorporated fibrin-fibronectin gel, which served
as a delivery system for FGF-2 compared to control, PBS filled, conduits in a 10 mm
gap (Han et al., 2010). Using the latter, however, they could only report successful
reinnervation in 5 out of 9 animals, while we recorded CMAPs in 13 out of 14
animals that were treated with hollow DAII chitosan conduits during project 1. The
differences regarding the success rate can possibly be attributed to the different DA

Discussion-Project 2
- 130 -
as well as varying production processes / fine tuning methods. Furthermore, FGF-2
has previously promoted nerve tissue regeneration across a 15 mm gap within
4 weeks when it was released from a BSA / ethylene-vinyl acetate based conduit
(Aebischer et al., 1989). Also, Wang et al. have reported enhanced nerve
regeneration across a 15 mm gap when delivering FGF-2 with regard to functional
and morphometric parameters, but the comparison to the ANG was missing (Wang
et al., 2003). Recently, Ikeda et al. (2014) could show that double layered nerve
conduits, consisting of PLA and PCL, in combination with induced pluripotent stem
cell-derived neurospheres and a FGF-drug delivery system accelerated nerve
regeneration in the mice sciatic nerve model compared to conduits alone (M. Ikeda
et al., 2014).
The missing reinnervation seen with GDNF supports former findings that this factor
might possibly be only effective in case of delayed repair and fails to promote
regeneration when administered immediately after nerve transection (Boyd and
Gordon, 2003a, 2003b; Höke et al., 2002; Wood et al., 2013a, 2013b). Moreover, it
has been reported that sustained over-expression of GDNF by SCs trapped axons in
acellular nerve grafts ultimately preventing functional regeneration across a 14 mm
gap in the rat sciatic nerve model (Santosa et al., 2013). On the other hand, it has
been described that GDNF promotes motor as well as sensory neurite outgrowth in
vitro and serves as a protective factor for these neurons (Allodi et al., 2013;
Henderson et al., 1994; Li et al., 1995; Madduri and Gander, 2010; Madduri et al.,
2009). Furthermore, Patel et al. could show that GDNF and laminin blended chitosan
conduits enhanced functional and sensory recovery as assessed by gait analysis,
sensitivity testing, and muscle weight determination in a 10 mm gap (Patel et al.,
2007). Similarly, 7 weeks after nerve repair, Fine et al. have described the formation
of a regeneration cable across a 15 mm gap when GDNF was slowly released (Fine
et al., 2002). That GDNF is an important factor for motoneuron regrowth was
confirmed by the finding that it promotes facial nerve regeneration (Barras et al.,
2002). Li and colleagues could further show enhanced recovery (10 mm gap) by
means of morphometric and electromyography examination compared to hollow
silicone tube (Li et al., 2006). In addition, Kokai and colleagues have reported
functional recovery across a long gap (15 mm) when delivering GDNF to the lesion
site via microspheres (Kokai et al., 2011). After 16 weeks, they saw higher muscle

Discussion-Project 2
- 131 -
twitch force in GDNF treated animals than in negative controls (empty
microspheres), while results were comparable with isografts (nerve graft from rats of
the same strain). Regarding muscle weight, however, the isograft was superior.
Interestingly, all animals that had received GDNF showed nerve tissue regeneration
while this was only the case in 70% of the negative control. Noteworthy is, however,
that they saw a high amount of collagen content within the lumen in both conduit
treated groups and fibers only appeared organized in the GDNF group. This is in
contrast to the tissue regeneration seen in this study. Mainly in the presence of FGF-
218kDa, some regeneration was seen and axons were organized, while collagen
contributed less to the complete nerve section. Furthermore, Chew et al. have
reported functional recovery in 44% of rats that had received fibers releasing GDNF
when bridging a 15 mm gap in the rat, while it was less without the growth factor
(Chew et al., 2007). In this study, however, it is not clear if the failed regeneration
can be explained by the missing effect of GDNF following immediate repair or if the
used luminal filler maybe had a negative effect on the regeneration and only FGF-
218kDa partly enabled regeneration against this obstacle. The idea that the hydrogel
represented some kind of barrier for the regrowing axons is supported by a study
from our partner laboratory that revealed nerve regeneration and functional recovery
across a 15 mm gap in 3 out of seven rats using hollow chitosan conduits (Talk:
“Tubulization with chitosan guides for the repair of long gap peripheral nerve injury in
the rat” by Francisco Gonzalez-Perez from UAB at the 2nd International Symposium
on Peripheral Nerve Regeneration in Turin, Italy).
6.2.4 The difficulty in finding the right luminal filler for axonal guidance In general, it is thought that structures inside the nerve guidance conduits could
facilitate SC migration as well as axonal outgrowth by replacing or supporting the
initially formed fibrin cable (Daly et al., 2012). In this study, however, the impression
is given that a hydrogel as SC carrier may have hampered nerve regeneration.
Possibly, the gel used was too dense, thus preventing the formation of the initial
fibrin matrix as well as subsequent migration of SCs, growth of axonal sprouts, and
diffusion of molecules. Usage of less dense gel, however, is difficult, since it would
leak from the conduit immediately after nerve reconstruction and the transplanted
cells would no longer be present at the site of need. The results of the in vivo study
regarding NVR-Gel do not coincide with in vitro results obtained by Sandra Wrobel

Discussion-Project 2
- 132 -
during her PhD-thesis (Institute of Neuroanatomy, MHH). She could show that
transfected SCs display their bipolar shape within the gel and neurite outgrowth
assays led to promising results. Recently, another in vitro study suggested great
potential of this hydrogel for in vivo applications (Ziv-Polat et al., 2014). Thus,
modifications of the NVR-Gel could potentially enhance its application in vivo, but so
far in vitro and in vivo results are diverging.
While some have used different hydrogels and were able to enhance nerve
regeneration, others have reported impeded regeneration as a result of the
introduction of matrixes (Apel et al., 2008; Hill et al., 2011; Karabekmez et al., 2009;
Pfister et al., 2007; Valentini et al., 1987; Wang et al., 1998; Zheng et al., 2010). It
has e.g. been attempted to bridge a 4 mm rat sciatic nerve gap using collagen- and
laminin-containing gels. Acellular remnants of the gel could still be identified after 12
weeks and less myelinated axons were found compared to saline filled conduits
(Valentini et al., 1987). Consequently, the authors suggested that a faster absorbed
delivery matrix might enable regeneration. Also, a SC-containing
chitosan/glycerol-ß-phosphate disodium salt hydrogel injection into conduits led to
complete failure of nerve regeneration across a 10 mm gap in the rat sciatic nerve
model, while SCs in suspension or culture medium alone allowed regeneration
(Zheng et al., 2010). Interestingly, Madison et al. have demonstrated that
laminin-containing gel initially enhanced nerve regeneration at 2 weeks, but acted
inhibitory at 6 weeks when bridging a 4 mm defect (Madison et al., 1987). The
authors suggested that the observed reversal occurred due to swelling and
decreased luminal space with the laminin gel, something that might have also
happened in this study. Moreover, the implantation of collagen tubes containing
SC-seeded polyglactin filaments in addition to SCs in Matrigel allowed only partial
regeneration, but failed completely regarding motor recovery (Lohmeyer et al.,
2007). It was concluded that the amount of inserted materials possibly acted as a
physical impediment to nerve regeneration or hampered cell function. Noteworthy,
Pfister and colleagues (2007) summarized in a review that growth factor free
matrixes result in poor regeneration, while the addition of these factors generally
mediated a better result than empty tubes. This holds not completely true for the
here tested chitosan conduits, since GDNF did not enhance nerve regeneration
compared to physiological SCs or empty vector transfected SCs in NVR-Gel. The
FGF-218kDa treated animals are the only group showing successful regeneration in

Discussion-Project 2
- 133 -
about 57% giving the impression that this growth factor partly enabled regeneration
even in a non-permissive environment created through the presence of the probably
too dense gel. Interestingly, we found NF200-positive fibers proximal to the proximal
suture in cases of failed regeneration, while this was not the case directly at the
suture site. This observation further supports the idea that axons started to regrow
but did not cross the suture line to grow inside the hydrogel filled conduit.
6.2.5 No enhanced inflammatory response elicited by (modified) Schwann cells (SCs) in NVR Gel The inflammatory response was comparably assessed as described for project 1 by
analyzing the fibrous capsule that had formed around the conduits as well as the
(regenerated) nerve tissue. The determination of the capsules thickness and area as
well as counting of activated macrophages and MGCs resulted in similar results as
seen for hollow chitosan conduits indicating that there was no increased
inflammatory response due to the presence of the NVR Gel or SCs, regardless if the
latter were genetically modified or not. The same observation was made for analysis
of the regenerated nerve tissue or nerve stumps in cases of failed regeneration.
6.2.6 Concluding remarks and considerations regarding future conduits for long nerve defects Studies that address the treatment of long peripheral nerve defects have to take into
account numerous variables that decide whether a conduit will have success in
promoting peripheral nerve regeneration or not. Although the PNS has an intrinsic
capacity to regenerate, problems like misdirection of axons, the deprivation of
regeneration promoting factors (e.g. due to SC denervation), and muscular atrophy
deteriorate the chance of accurate reinnervation and functional recovery limit the
success rate especially across long gaps and in cases of delayed repair (Allodi et al.,
2012; de Ruiter et al., 2008; Gordon, 2009; Gu et al., 2014; Klimaschewski et al.,
2013; Lin et al., 2011; Saito and Dahlin, 2008; Scheib and Höke, 2013)
Intraluminal guidance structures / fillers are thought to be favorable, since they
potentially support SC migration and axonal outgrowth, but as seen in this study,
their introduction can also disturb nerve regeneration (Daly et al., 2012). We
observed complete failure of regeneration in most animals of the enriched conduits

Discussion-Project 2
- 134 -
treated animals except for the FGF-218kDa group and partly only detected very thin
tissue connections between the two nerve stumps, which only occasionally
contained axons. Successful regeneration including functional recovery was only
seen when SCs over-expressing FGF-218kDa were suspended in the gel. Compared
to hollow chitosan conduits, however, which were previously tested in a long gap by
our partner laboratory, the gene delivery did not enhance the rate of successful
regeneration (Talk: “Tubulization with chitosan guides for the repair of long gap
peripheral nerve injury in the rat” by Francisco Gonzalez-Perez from UAB at the 2nd
International Symposium on Peripheral Nerve Regeneration in Turin, Italy). It can be
assumed that the limited success of the enriched nerve conduit is based on the
density of the used gel and that only FGF-218kDa partly promoted axonal outgrowth in
this unfavorable environment. For future experiments, it is therefore of importance to
identify a more suitable delivery method for these genetically modified cells, which
finds a balance in providing a guidance structure without impeding axonal outgrowth
due to unintended barrier functions of the filler (Lin and Marra, 2012). Alternatives
could be very porous sponges and fibers as well as membranes that only cover a
limited area of the conduits cross-section, thus leaving space for regrowing axons
but still offer a guidance surface for growth cones. Furthermore, growth factor
delivery methods are in need of modification in a way to allow a spatial and temporal
dependent release that adjusts to different phases of nerve regeneration. This issue
is, however, very complex, since it has to be determined which factors are needed at
what time point of regeneration as well as in what dose and at what localization, and
most importantly a sophisticated delivery method has to be found that allows this
kind of regulation. Also, different regulation profiles have to be established for nerves
of different modalities, since it was shown that sensory and motor nerves prefer
varying environments and are usually accompanied by SCs of differing phenotypes
(Höke et al., 2006). Furthermore, synergistic effects of different factors have to be
elucidated to increase the regeneration capacity.

- 135 -

References
- 136 -
7 References Aebischer, P., Salessiotis, A.N., Winn, S.R., 1989. Basic fibroblast growth factor released from
synthetic guidance channels facilitates peripheral nerve regeneration across long nerve gaps. Journal of Neuroscience Research 23, 282–89.
Allodi, I., Casals-Díaz, L., Santos-Nogueira, E., Gonzalez-Perez, F., Navarro, X., Udina, E., 2013. Fgf-2 low molecular weight selectively promotes neuritogenesis of motor neurons in vitro. Molecular Neurobiology 47, 770–81.
Allodi, I., Guzmán-Lenis, M.-S., Hernàndez, J., Navarro, X., Udina, E., 2011. In vitro comparison of motor and sensory neuron outgrowth in a 3D collagen matrix. Journal of Neuroscience Methods 198, 53–61.
Allodi, I., Udina, E., Navarro, X., 2012. Specificity of peripheral nerve regeneration: Interactions at the axon level. Progress in Neurobiology 98, 16–37.
Alluin, O., Wittmann, C., Marqueste, T., Chabas, J.-F., Garcia, S., Lavaut, M.-N., Guinard, D., Feron, F., Decherchi, P., 2009. Functional recovery after peripheral nerve injury and implantation of a collagen guide. Biomaterials 30, 363–73.
Al-Majed, A.A., Neumann, C.M., Brushart, T.M., Gordon, T., 2000. Brief electrical stimulation promotes the speed and accuracy of motor axonal regeneration. The Journal of Neuroscience 20, 2602–8.
Anderson, J.M., 2001. Biological responses to materials. Annual Review of Materials Research 31, 81–110.
Anderson, J.M., Rodriguez, A., Chang, D.T., 2008. Foreign body reaction to biomaterials. Seminars in Immunology 20, 86–100.
Angius, D., Wang, H., Spinner, R.J., Gutierrez-Cotto, Y., Yaszemski, M.J., Windebank, A.J., 2012. A systematic review of animal models used to study nerve regeneration in tissue-engineered scaffolds. Biomaterials 33, 8034–9.
Ansselin, A.D., Fink, T., Davey, D.F., 1997. Peripheral nerve regeneration through nerve guides seeded with adult Schwann cells. Neuropathology and Applied Neurobiology 23, 387–98.
Ao, Q., Fung, C.-K., Tsui, A.Y.-P., Cai, S., Zuo, H.-C., Chan, Y.-S., Shum, D.K.-Y., 2011. The regeneration of transected sciatic nerves of adult rats using chitosan nerve conduits seeded with bone marrow stromal cell-derived Schwann cells. Biomaterials 32, 787–96.
Apel, P.J., Garrett, J.P., Sierpinski, P., Ma, J., Atala, A., Smith, T.L., Koman, L.A., Van Dyke, M.E., 2008. Peripheral nerve regeneration using a keratin-based scaffold: Long-term functional and histological outcomes in a mouse model. Journal of Hand Surgery 33, 1541–7.
Aranaz, I., Mengibar, M., Harris, R., Panos, I., Miralles, B., Acosta, N., Galed, G., Heras, A., 2009. Functional characterization of chitin and chitosan. Current Chemical Biology 3, 203–30.
Archibald, S.J., Krarup, C., Shefner, J., Li, S.T., Madison, R.D., 1991. A collagen-based nerve guide conduit for peripheral nerve repair: An electrophysiological study of nerve regeneration in rodents and nonhuman primates. The Journal of Comparative Neurology 306, 685–96.
axogenic.com, 2014. AxoGen [WWW Document]. Azevedo, F.A.C., Carvalho, L.R.B., Grinberg, L.T., Farfel, J.M., Ferretti, R.E.L., Leite, R.E.P., Jacob
Filho, W., Lent, R., Herculano-Houzel, S., 2009. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of Comparative Neurology 513, 532–41.
Barbosa, J.N., Amaral, I.F., Aguas, A.P., Barbosa, M.A., 2010. Evaluation of the effect of the degree of acetylation on the inflammatory response to 3D porous chitosan scaffolds. Journal of Biomedical Materials Research Part A 93, 20–8.
Barras, F.M., Pasche, P., Bouche, N., Aebischer, P., Zurn, A.D., 2002. Glial cell line-derived neurotrophic factor released by synthetic guidance channels promotes facial nerve regeneration in the rat. Journal of Neuroscience Research 70, 746–55.
Belkas, J.S., Munro, C.A., Shoichet, M.S., Johnston, M., Midha, R., 2005. Long-term in vivo biomechanical properties and biocompatibility of poly(2-hydroxyethyl methacrylate-co-methyl methacrylate) nerve conduits. Biomaterials 26, 1741–9.
Belkas, J.S., Shoichet, M.S., Midha, R., 2004a. Peripheral nerve regeneration through guidance tubes. Neurological Research 26, 151–60.
Belkas, J.S., Shoichet, M.S., Midha, R., 2004b. Axonal guidance channels in peripheral nerve regeneration. Operative Techniques in Orthopaedics 14, 190–8.

References
- 137 -
Bell, J., Haycock, J.W., 2011. Next generation gerve Guides - materials, fabrication, growth factors and cell delivery. Tissue engineering Part B Reviews 44, 1–38.
Bennett, G.J., Xie, Y.-K., 1988. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 33, 87–107.
Berger, A., Lassner, F., Schaller, E., 1994. The Dellon tube in injuries of peripheral nerves. Handchirurgie, Mikrochirurgie, plastische Chirurgie 26, 44–7.
Berger, M.J., Hubli, M., Krassioukov, A. V, 2014. Sympathetic skin responses and autonomic dysfunction in spinal cord injury. Journal of neurotrauma Online ahe.
Bervar, M., 2000. Video analysis of standing-an alternative footprint analysis to assess functional loss following injury to the rat sciatic nerve. Journal of Neuroscience Methods 102, 109–16.
Bervar, M., 2005. Effect of weak, interrupted sinusoidal low frequency magnetic field on neural regeneration in rats: functional evaluation. Bioelectromagnetics 26, 351–6.
Bini, T.B., Gao, S., Xu, X., Wang, S., Ramakrishna, S., Leong, K.W., 2004. Peripheral nerve regeneration by microbraided poly(L-lactide-co-glycolide) biodegradable polymer fibers. Journal of Biomedical Materials Research. Part A 68, 286–95.
Boeckstyns, M.E.H., Sørensen, A.I., Viñeta, J.F., Rosén, B., Navarro, X., Archibald, S.J., Valss-Solé, J., Moldovan, M., Krarup, C., 2013. Collagen conduit versus microsurgical neurorrhaphy: 2-year follow-up of a prospective, blinded clinical and electrophysiological multicenter randomized, controlled trial. The Journal of Hand Surgery 38, 2405–11.
Boiko, T., Van Wart, A., Caldwell, J.H., Levinson, S.R., Trimmer, J.S., Matthews, G., 2003. Functional specialization of the axon initial segment by isoform-specific sodium channel targeting. The Journal of Neuroscience 23, 2306–13.
Boucher, T., 2001. Neurotrophic factors and neuropathic pain. Current Opinion in Pharmacology 1, 66–72.
Boucher, T.J., Okuse, K., Bennett, D.L., Munson, J.B., Wood, J.N., McMahon, S.B., 2000. Potent analgesic effects of GDNF in neuropathic pain states. Science 290, 124–7.
Boyd, B.S., Puttlitz, C., Noble-Haeusslein, L.J., John, C.M., Trivedi, A., Topp, K.S., 2007. Deviations in gait pattern in experimental models of hindlimb paresis shown by a novel pressure mapping system. Journal of Neuroscience Research 85, 2272–83.
Boyd, J.G., Gordon, T., 2003a. Neurotrophic factors and their receptors in axonal regeneration and functional recovery after peripheral nerve injury. Molecular Neurobiology 27, 277–323.
Boyd, J.G., Gordon, T., 2003b. Glial cell line-derived neurotrophic factor and brain-derived neurotrophic factor sustain the axonal regeneration of chronically axotomized motoneurons in vivo. Experimental Neurology 183, 610–9.
Bozkurt, A., Deumens, R., Scheffel, J., O’Dey, D.M., Weis, J., Joosten, E.A., Führmann, T., Brook, G.A., Pallua, N., 2008. CatWalk gait analysis in assessment of functional recovery after sciatic nerve injury. Journal of Neuroscience Methods 173, 91–8.
Bozkurt, A., Scheffel, J., Brook, G.A., Joosten, E.A., Suschek, C. V, O’Dey, D.M., Pallua, N., Deumens, R., 2011. Aspects of static and dynamic motor function in peripheral nerve regeneration: SSI and CatWalk gait analysis. Behavioural Brain Research 219, 55–62.
Bradke, F., Fawcett, J.W., Spira, M.E., 2012. Assembly of a new growth cone after axotomy: the precursor to axon regeneration. Nature Reviews. Neuroscience 13, 183–93.
Bridges, D., Ahmad, K., Rice, A.S., 2001. The synthetic cannabinoid WIN55,212-2 attenuates hyperalgesia and allodynia in a rat model of neuropathic pain. British Journal of Pharmacology 133, 586–94.
Brodbeck, W.G., Anderson, J.M., 2009. Giant cell formation and function. Current Opinion in Hematology 16, 53–7.
Brooks, D.N., Weber, R. V, Chao, J.D., Rinker, B.D., Zoldos, J., Robichaux, M.R., Ruggeri, S.B., Anderson, K.A., Bonatz, E.E., Wisotsky, S.M., Cho, M.S., Wilson, C., Cooper, E.O., Ingari, J. V, Safa, B., Parrett, B.M., Buncke, G.M., 2012. Processed nerve allografts for peripheral nerve reconstruction: a multicenter study of utilization and outcomes in sensory, mixed, and motor nerve reconstructions. Microsurgery 32, 1–14.
Brushart, T.M., 2011a. Chapther 1: Peripheral nerve anatomy, in: Nerve Repair. Oxford University Press, pp. 3–18.
Brushart, T.M., 2011b. Chapter 9: Nerve regeneration, in: Nerve Repair. p. 253. Brushart, T.M., 2011c. Chapter 9: Nerve Regeneration, in: Nerve Repair. pp. 283–289. Burdick, J.A., Prestwich, G.D., 2011. Hyaluronic acid hydrogels for biomedical applications. Advanced
Materials 23, H41–56.

References
- 138 -
Casals-Díaz, L., Vivó, M., Navarro, X., 2009. Nociceptive responses and spinal plastic changes of afferent C-fibers in three neuropathic pain models induced by sciatic nerve injury in the rat. Experimental Neurology 217, 84–95.
Chalfoun, C.T., Wirth, G.A., Evans, G.R.D., 2006. Tissue engineered nerve constructs: where do we stand? Journal of Cellular and Molecular Medicine 10, 309–17.
Chemnitz, A., Andersson, G., Rosén, B., Dahlin, L.B., Björkman, A., 2013. Poor electroneurography but excellent hand function 31 years after nerve repair in childhood. Neuroreport 24, 6–9.
Chen, R., Cohen, L.G., Hallett, M., 2002. Nervous system reorganization following injury. Neuroscience 111, 761–73.
Chew, S.Y., Mi, R., Hoke, A., Leong, K.W., 2007. Aligned protein-polymer composite fibers enhance nerve regeneration: A potential tissue-engineering platform. Advanced Functional Materials 17, 1288–96.
Chiriac, S., Facca, S., Diaconu, M., Gouzou, S., Liverneaux, P., 2012. Experience of using the bioresorbable copolyester poly(DL-lactide-ε-caprolactone) nerve conduit guide NeurolacTM for nerve repair in peripheral nerve defects: report on a series of 28 lesions. The Journal of Hand Surgery, European Volume 37, 342–9.
Ciaramitaro, P., Mondelli, M., Logullo, F., Grimaldi, S., Battiston, B., Sard, A., Scarinzi, C., Migliaretti, G., Faccani, G., Cocito, D., 2010. Traumatic peripheral nerve injuries: epidemiological findings, neuropathic pain and quality of life in 158 patients. Journal of the Peripheral Nervous System 15, 120–7.
Ciardelli, G., Chiono, V., 2006. Materials for peripheral nerve regeneration. Macromolecular Bioscience 6, 13–26.
Clements, I.P., Kim, Y., English, A.W., Lu, X., Chung, A., Bellamkonda, R. V, 2009. Thin-film enhanced nerve guidance channels for peripheral nerve repair. Biomaterials 30, 3834–46.
Cobianchi, S., Casals-Diaz, L., Jaramillo, J., Navarro, X., 2013. Differential effects of activity dependent treatments on axonal regeneration and neuropathic pain after peripheral nerve injury. Experimental Neurology 240, 157–67.
Cobianchi, S., de Cruz, J., Navarro, X., 2014. Assessment of sensory thresholds and nociceptive fiber growth after sciatic nerve injury reveals the differential contribution of collateral reinnervation and nerve regeneration to neuropathic pain. Experimental Neurology 255, 1–11.
Costa, L.M., Simões, M.J., Mauricio, A.C., Varejão, S.P., 2009. Methods and protocols in peripheral nerv eregeneration experimental research: Part IV-Kinematic gait analysis to quantify peripheral nerve regeneration in the rat, in: International Review of Neurobiology. pp. 127–39.
Dahlin, L.B., Zhao, Q., Bjursten, L.M., 1995. Nerve regeneration in silicone tubes: distribution of macrophages and interleukin-1β in the formed fibrin matrix. Restorative Neurology and Neuroscience 8, 199–203.
Daly, W., Yao, L., Zeugolis, D., Windebank, A., Pandit, A., 2012. A biomaterials approach to peripheral nerve regeneration: bridging the peripheral nerve gap and enhancing functional recovery. Journal of the Royal Society Interface 9, 202–21.
Daly, W.T., Knight, A.M., Wang, H., de Boer, R., Giusti, G., Dadsetan, M., Spinner, R.J., Yaszemski, M.J., Windebank, A.J., 2013. Comparison and characterization of multiple biomaterial conduits for peripheral nerve repair. Biomaterials 34, 8630–9.
Danielsen, N., Pettmann, B., Vahlsing, H.L., Manthorpe, M., Varon, S., 1989. Fibroblast growth factor effects on peripheral nerve regeneration in a silicone chamber model. Journal of Neuroscience Research 20, 282–89.
De Boer, R., Borntraeger, A., Knight, A.M., Hébert-Blouin, M.-N., Spinner, R.J., Malessy, M.J.A., Yaszemski, M.J., Windebank, A.J., 2012. Short- and long-term peripheral nerve regeneration using a poly-lactic-co-glycolic-acid scaffold containing nerve growth factor and glial cell line-derived neurotrophic factor releasing microspheres. Journal of Biomedical Materials Research Part A 100, 2139–46.
De Ruiter, G.C., Spinner, R.J., Malessy, M.J.A., Moore, M.J., Sorenson, E.J., Currier, B.L., Yaszemski, M.J., Windebank, A.J., 2008. Accuracy of motor axon regeneration across autograft, single-lumen, and multichannel poly(lactic-co-glycolic acid) nerve tubes. Neurosurgery 63, 144–53; discussion 153–5.
De Ruiter, G.C.W., 2009. Misdirection and guidance of regenerating motor axons after experimental nerve injury and repair: Chapter 5: Review of the experimental and clinical literature on nerve tubes for peripheral nerve repair.
De Ruiter, G.C.W., Malessy, M.J.A., Yaszemski, M.J., Windebank, A.J., Spinner, R.J., 2009. Designing ideal conduits for peripheral nerve repair. Neurosurgical Focus 26, E5.

References
- 139 -
Deumens, R., Bozkurt, A., Meek, M.F., Marcus, M.A.E., Joosten, E.A.J., Weis, J., Brook, G.A., 2010. Repairing injured peripheral nerves: Bridging the gap. Progress in Neurobiology 92, 245–76.
Di Summa, P.G., Kalbermatten, D.F., Pralong, E., Raffoul, W., Kingham, P.J., Terenghi, G., 2011. Long-term in vivo regeneration of peripheral nerves through bioengineered nerve grafts. Neuroscience 181, 278–91.
Di Summa, P.G., Kingham, P.J., Raffoul, W., Wiberg, M., Terenghi, G., Kalbermatten, D.F., 2010. Adipose-derived stem cells enhance peripheral nerve regeneration. Journal of Plastic, Reconstructive & Aesthetic Surgery 63, 1544–52.
Diamond, J., Coughlin, M., Macintyre, L., Holmes, M., Visheau, B., 1987. Evidence that endogenous beta nerve growth factor is responsible for the collateral sprouting, but not the regeneration, of nociceptive axons in adult rats. Proceedings of the National Academy of Sciences of the United States of America 84, 6596–600.
Díaz-Flores, L., Gutiérrez, R., Varela, H., Evora, P., Valladares, F., Rodríguez, M., Rancel, N., Alvarez-Argüelles, H., 1995. Contribution of the proximal and distal nerve stumps to peripheral nerve regeneration in silicone chambers. Histology and Histopathology 10, 937–46.
Ding, F., Wu, J., Yang, Y., Hu, W., Zhu, Q., Tang, X., Liu, J., Gu, X., 2010. Use of tissue-engineered nerve grafts consisting of a chitosan/poly(lactic-co-glycolic acid)-based scaffold included with bone marrow mesenchymal cells for bridging 50-mm dog sciatic nerve gaps. Tissue Engineering. Part A 16, 3779–90.
Dorey, K., Amaya, E., 2010. FGF signalling: diverse roles during early vertebrate embryogenesis. Development 137, 3731–42.
Dornseifer, U., Fichter, A.M., Leichtle, S., Wilson, A., Rupp, A., Rodenacker, K., Ninkovic, M., Biemer, E., Machens, H.-G., Matiasek, K., Papadopulos, N.A., 2011. Peripheral nerve reconstruction with collagen tubes filled with denatured autologous muscle tissue in the rat model. Microsurgery 31, 632–41.
Dove, A., 2002. Cell-based therapies go live. Nature Biotechnology 20, 339–43. Dubový, P., 2011. Wallerian degeneration and peripheral nerve conditions for both axonal
regeneration and neuropathic pain induction. Annals of Anatomy 193, 267–75. Dutta, P.K., Dutta, J., Tripathi, V.S., 2004. Chitin and chitosan: Chemistry, properties and
applications. Journal of Scientific & Industrial Research 63, 20–31. Fine, E.G., Decosterd, I., Papaloïzos, M., Zurn, A.D., Aebischer, P., 2002. GDNF and NGF released
by synthetic guidance channels support sciatic nerve regeneration across a long gap. European Journal of Neuroscience 15, 589–601.
Flores, A.J., Lavernia, C.J., Owens, P.W., 2000. Anatomy and physiology of peripheral nerve injury and repair. The Journal Of The American Academy Of Orthopaedic Surgeons 150, 537–41, 551, 550.
Fox, I.K., Mackinnon, S.E., 2011. Adult peripheral nerve disorders: nerve entrapment, repair, transfer, and brachial plexus disorders. Plastic and Reconstructive Surgery 127, 105e–118e.
Freier, T., Jiminez Hamann, M., Katayana, Y., Musoke-Zawedde, P., Piotrowicz, A., Yuan, Y., Shoichet, M.S., 2005a. Chapter 9: Biodegradable polymers in neural tissue engineering, in: Handbook of Biodegradable Polymeric Materials and Their Applications. pp. 1–49.
Freier, T., Koh, H.S., Kazazian, K., Shoichet, M.S., 2005b. Controlling cell adhesion and degradation of chitosan films by N-acetylation. Biomaterials 26, 5872–8.
Freier, T., Montenegro, R., Shan Koh, H., Shoichet, M.S., 2005c. Chitin-based tubes for tissue engineering in the nervous system. Biomaterials 26, 4624–32.
Friedman, A.H., Elias, W.J., Midha, R., 2009. Introduction: peripheral nerve surgery-biology, entrapment, and injuries. Neurosurgical Focus 26, E1.
Frostick, S.P., Yin, Q., Kemp, G.J., 1998. Schwann cells, neurotrophic factors, and peripheral nerve regeneration. Microsurgery 18, 397–405.
Frykman, G.K., 1976. Peripheral nerve injuries in children. The Orthopedic Clinics of North America 7, 701–16.
Fugleholm, K., Schmalbruch, H., Krarup, C., 2000. Post reinnervation maturation of myelinated nerve fibers in the cat tibial nerve: chronic electrophysiological and morphometric studies. Journal of the Peripheral Nervous System 5, 82–95.
Fujimoto, E., Mizoguchi, A., Hanada, K., Yahima, M., Ide, C., 1997. Basic fibroblast growth factor promotes extension of regenerating axons of peripheral nerve. In vivo experiments using a Schwann cell basal lamina tube model. Journal of Neurocytology 26, 511–28.
Gaudet, A.D., Popovich, P.G., Ramer, M.S., 2011. Wallerian degeneration: gaining perspective on inflammatory events after peripheral nerve injury. Journal of Neuroinflammation 8, 110.

References
- 140 -
Gelderd, J.B., Chopin, S.F., 1977. The vertebral level of origin of spinal nerves in the rat. The Anatomical Record 188, 45–7.
Georgiou, M., Bunting, S.C.J., Davies, H.A., Loughlin, A.J., Golding, J.P., Phillips, J.B., 2013. Engineered neural tissue for peripheral nerve repair. Biomaterials 34, 7335–43.
Gonzalez-Perez, F., 2014. Talk (2nd international symposium on peripheral nerve regeneration): “Tubulization with chitosan guides for the repair of long gap peripheral nerve injury in the rat”.
Gordon, T., 2009. The role of neurotrophic factors in nerve regeneration. Neurosurgical Focus 26, E3. Gordon, T., Sulaiman, O., Boyd, J.G., 2003. Experimental strategies to promote functional recovery
after peripheral nerve injuries. Journal of the Peripheral Nervous System 8, 236–50. Grimmelikhuijzen, C.J., Westfall, J.A., 1995. The nervous systems of cnidarians. EXS 72, 7–24. Grothe, C., Haastert, K., Jungnickel, J., 2006. Physiological function and putative therapeutic impact
of the FGF-2 system in peripheral nerve regeneration-lessons from in vivo studies in mice and rats. Brain Research Reviews 51, 293–9.
Grothe, C., Heese, K., Meisinger, C., Wewetzer, K., Kunz, D., Cattini, P., Otten, U., 2000. Expression of interleukin-6 and its receptor in the sciatic nerve and cultured Schwann cells: Relation to 18-kD fibroblast growth factor-2. Brain Research 885, 172–81.
Grothe, C., Meisinger, C., Claus, P., 2001. In vivo expression and localization of the fibroblast growth factor system in the intact and lesioned rat peripheral nerve and spinal ganglia. The Journal of Comparative Neurology 434, 342–57.
Grothe, C., Meisinger, C., Hertenstein, A., Kurz, H., Wewetzer, K., 1996. Expression of fibroblast growth factor-2 and fibroblast growth factor receptor 1 messenger RNAs in spinal ganglia and sciatic nerve: regulation after peripheral nerve lesion. Neuroscience 76, 123–35.
Grothe, C., Nikkhah, G., 2001. The role of basic fibroblast growth factor in peripheral nerve regeneration. Anatomy and Embryology 204, 171–77.
Gu, X., Ding, F., Williams, D.F., 2014. Neural tissue engineering options for peripheral nerve regeneration. Biomaterials 35, 6143–56.
Gu, X., Ding, F., Yang, Y., Liu, J., 2011. Construction of tissue engineered nerve grafts and their application in peripheral nerve regeneration. Progress in Neurobiology 93, 204–30.
Haastert, K., Lipokatic, E., Fischer, M., Timmer, M., Grothe, C., 2006a. Differentially promoted peripheral nerve regeneration by grafted Schwann cells over-expressing different FGF-2 isoforms. Neurobiology of Disease 21, 138–53.
Haastert, K., Mauritz, C., Matthies, C., Grothe, C., 2006b. Autologous adult human Schwann cells genetically modified to provide alternative cellular transplants in peripheral nerve regeneration. Journal of Neurosurgery 104, 778–86.
Haastert-Talini, K., Geuna, S., Dahlin, L.B., Meyer, C., Stenberg, L., Freier, T., Heimann, C., Barwig, C., Pinto, L.F.V., Raimondo, S., Gambarotta, G., Samy, S.R., Sousa, N., Salgado, A.J., Ratzka, A., Wrobel, S., Grothe, C., 2013. Chitosan tubes of varying degrees of acetylation for bridging peripheral nerve defects. Biomaterials 34, 9886–904.
Haastert-Talini, K., Schmitte, R., Korte, N., Klode, D., Ratzka, A., Grothe, C., 2011. Electrical stimulation accelerates axonal and functional peripheral nerve regeneration across long gaps. Journal of Neurotrauma 28, 661–74.
Hadlock, T., Sundback, C., Hunter, D., Cheney, M., Vacanti, J.P., 2000. A polymer foam conduit seeded with Schwann cells promotes guided peripheral nerve regeneration. Tissue Engineering 6, 119–27.
Hadlock, T.A., Sundback, C.A., Hunter, D.A., Vacanti, J.P., Cheney, M.L., 2001. A new artificial nerve graft containing rolled Schwann cell monolayers. Microsurgery 21, 96–101.
Hall, S., 2005. The response to injury in the peripheral nervous system. The Journal of Bone & Joint Surgery 87-B, 1309–19.
Han, H., Ao, Q., Chen, G., Wang, S., Zuo, H., 2010. A novel basic fibroblast growth factor delivery system fabricated with heparin-incorporated fibrin-fibronectin matrices for repairing rat sciatic nerve disruptions. Biotechnology Letters 32, 585–91.
Han, T., New, N., Furuike, T., Tokura, S., Tamura, H., 2011. Methods of N-acetylated chitosan scaffolds and its in vitro biodegradation by lysozyme. Biomedical Science and Engineering 5, 15–23.
Heavner, J.E., de Jong, R.H., 1974. Lidocaine blocking concentrations for B- and C-nerve fibers. Anesthesiology 40, 228–33.
Henderson, C., Phillips, H., Pollock, R., Davies, A., Lemeulle, C., Armanini, M., Simmons, L., Moffet, B., Vandlen, R., Simpson LC Corrected to Simmons, L., Et, A., 1994. GDNF: a potent survival factor for motoneurons present in peripheral nerve and muscle. Science 266, 1062–4.

References
- 141 -
Hernández-Cortés, P., Garrido, J., Cámara, M., Ravassa, F.O., 2010. Failed digital nerve reconstruction by foreign body reaction to Neurolac nerve conduit. Microsurgery 30, 414–6.
Hidaka, Y., Ito, M., Mori, K., Yagasaki, H., Kafrawy, A.H., 1999. Histopathological and immunohistochemical studies of membranes of deacetylated chitin derivatives implanted over rat calvaria. Journal of Biomedical Materials Research 46, 418–23.
Hill, P.S., Apel, P.J., Barnwell, J., Smith, T., Koman, L.A., Atala, A., Van Dyke, M., 2011. Repair of peripheral nerve defects in rabbits using keratin hydrogel scaffolds. Tissue engineering. Part A 17, 1499–505.
Hirata, K., Kawabuchi, M., 2002. Myelin phagocytosis by macrophages and nonmacrophages during Wallerian degeneration. Microscopy research and technique 57, 541–7.
Höke, A., Gordon, T., Zochodne, D.W., Sulaiman, O.A.R., 2002. A decline in glial cell-line-derived neurotrophic factor expression is associated with impaired regeneration after long-term Schwann cell denervation. Experimental Neurology 173, 77–85.
Höke, A., Ho, T., Crawford, T.O., LeBel, C., Hilt, D., Griffin, J.W., 2003. Glial cell line-derived neurotrophic factor alters axon schwann cell units and promotes myelination in unmyelinated nerve fibers. The Journal of Neuroscience 23, 561–7.
Höke, A., Redett, R., Hameed, H., Jari, R., Zhou, C., Li, Z.B., Griffin, J.W., Brushart, T.M., 2006. Schwann cells express motor and sensory phenotypes that regulate axon regeneration. Journal of Neuroscience 26, 9646–55.
Hu, N., Wu, H., Xue, C., Gong, Y., Wu, J., Xiao, Z., Yang, Y., Ding, F., Gu, X., 2013. Long-term outcome of the repair of 50 mm long median nerve defects in rhesus monkeys with marrow mesenchymal stem cells-containing, chitosan-based tissue engineered nerve grafts. Biomaterials 34, 100–11.
Hu, W.J., Eaton, J.W., Ugarova, T.P., Tang, L., 2001. Molecular basis of biomaterial-mediated foreign body reactions. Blood 98, 1231–8.
Hu, X., Huang, J., Ye, Z., Xia, L., Li, M., Lv, B., Shen, X., Luo, Z., 2009. A novel scaffold with longitudinally oriented microchannels promotes peripheral nerve regeneration. Tissue Engineering. Part A 15, 3297–308.
Huang, Y.-C., Huang, Y.-Y., 2006. Biomaterials and strategies for nerve regeneration. Artificial Organs 30, 514–22.
Huelsenbeck, S.C., Rohrbeck, A., Handreck, A., Hellmich, G., Kiaei, E., Roettinger, I., Grothe, C., Just, I., Haastert-Talini, K., 2012. C3 peptide promotes axonal regeneration and functional motor recovery after peripheral nerve injury. Neurotherapeutics 9, 185–98.
Ide, C., 1996. Peripheral nerve regeneration. Neuroscience Research 25, 101–21. Ijkema-Paassen, J., Jansen, K., Gramsbergen, A., Meek, M.F., 2004. Transection of peripheral
nerves, bridging strategies and effect evaluation. Biomaterials 25, 1583–92. Ijkema-Paassen, J., Meek, M.F., Gramsbergen, A., 2002. Reinnervation of muscles after transection
of the sciatic nerve in adult rats. Muscle & Nerve 25, 891–7. Ikeda, M., Uemura, T., Takamatsu, K., Kazuki, K., Tabata, Y., Ikada, Y., Nakamura, H., 2014.
Acceleration of peripheral nerve regeneration using nerve conduits in combination with induced pluripotent stem cell technology and a basic fibroblast growth factor drug delivery system. Journal of Biomedical Materials Research. Part A 102, 1370–8.
Ikeda, T., Ikeda, K., Yamamoto, K., Ishizaki, H., Yoshizawa, Y., Yanagiguchi, K., Yamada, S., Hayashi, Y., 2014. Fabrication and characteristics of chitosan sponge as a tissue engineering scaffold. BioMed Research International 2014. Article ID 786892, 8 pages.
IOM (Institute of Medicine), 2012. Ensuring safe food and medical products through stronger regulatory systems abroad.
Isaacs, J., 2010. Treatment of acute peripheral nerve injuries: current concepts. The Journal of Hand Surgery 35, 491–7.
Itoh, S., Suzuki, M., Yamaguchi, I., Takakuda, K., Kobayashi, H., Shinomiya, K., Tanaka, J., 2003. Development of a nerve scaffold using a tendon chitosan tube. Artificial Organs 27, 1079–88.
Jansen, K., Meek, M.F., van der Werff, J.F.A., van Wachem, P.B., van Luyn, M.J.A., 2004. Long-term regeneration of the rat sciatic nerve through a biodegradable poly(DL-lactide-epsilon-caprolactone) nerve guide: tissue reactions with focus on collagen III/IV reformation. Journal of Biomedical Materials Research. Part A 69, 334–41.
Jiang, X., Lim, S.H., Mao, H.-Q., Chew, S.Y., 2010. Current applications and future perspectives of artificial nerve conduits. Experimental Neurology 223, 86–101.
Josephson, A., Widenfalk, J., Trifunovski, A., Widmer, H.R., Olson, L., Spenger, C., 2001. GDNF and NGF family members and receptors in human fetal and adult spinal cord and dorsal root ganglia. Journal of Comparative Neurology 440, 204–17.

References
- 142 -
Jubran, M., Widenfalk, J., 2003. Repair of peripheral nerve transections with fibrin sealant containing neurotrophic factors. Experimental Neurology 181, 204–12.
Jungnickel, J., Claus, P., Gransalke, K., Timmer, M., Grothe, C., 2004. Targeted disruption of the FGF-2 gene affects the response to peripheral nerve injury. Molecular and Cellular Neurosciences 25, 444–52.
Jungnickel, J., Eckhardt, M., Haastert-Talini, K., Claus, P., Bronzlik, P., Lipokatic-Takacs, E., Maier, H., Gieselmann, V., Grothe, C., 2011. Polysialyltransferase-overexpression in Schwann cells mediates different effects during peripheral nerve regeneration. Glycobiology 22, 107–15.
Jungnickel, J., Haase, K., Konitzer, J., Timmer, M., Grothe, C., 2006. Faster nerve regeneration after sciatic nerve injury in mice over-expressing basic fibroblast growth factor. Journal of Neurobiology 66, 940–8.
Jungnickel, J., Haastert, K., Grzybek, M., Thau, N., Lipokatic-Takacs, E., Ratzka, A., Nölle, A., Claus, P., Grothe, C., 2010. Mice lacking basic fibroblast growth factor showed faster sensory recovery. Experimental Neurology 223, 166–72.
Karabekmez, F.E., Duymaz, A., Moran, S.L., 2009. Early clinical outcomes with the use of decellularized nerve allograft for repair of sensory defects within the hand. Hand 4, 245–9.
Kardas, I., Struszcyk, M.H., Kucharska, M., van den Broek, L.A.M., van Dam, J.E.G., Ciechanska, D., 2012. Chapter 11: Chitin and chitosan as functional biopolymers for industrial applications, in: The European Polysacchride Network of Excellence (EPNOE). pp. 329–73.
Kean, T., Thanou, M., 2010. Biodegradation, biodistribution and toxicity of chitosan. Advanced Drug Delivery Reviews 62, 3–11.
Kehoe, S., Zhang, X.F., Boyd, D., 2012. FDA approved guidance conduits and wraps for peripheral nerve injury: A review of materials and efficacy. Injury 43, 553–72.
Khaing, Z.Z., Schmidt, C.E., 2012. Advances in natural biomaterials for nerve tissue repair. Neuroscience Letters 519, 103–14.
Kiefer, R., Kieseier, B.C., Stoll, G., Hartung, H.P., 2001. The role of macrophages in immune-mediated damage to the peripheral nervous system. Progress in Neurobiology 64, 109–27.
Kiefer, R., Lindholm, D., Kreutzberg, G.W., 1993. Interleukin-6 and transforming growth factor-beta 1 mRNAs are induced in rat facial nucleus following motoneuron axotomy. The European Journal of Neuroscience 5, 775–81.
Kimura, J., 1984. Principles and pitfalls of nerve conduction studies. Annals of neurology 16, 415–29. Kingery, W.S., Lu, J.D., Roffers, J.A., Kell, D.R., 1994. The resolution of neuropathic hyperalgesia
following motor and sensory functional recovery in sciatic axonotmetic mononeuropathies. Pain 58, 157–68.
Kingery, W.S., Vallin, J.A., 1989. The development of chronic mechanical hyperalgesia, autotomy and collateral sprouting following sciatic nerve section in rat. Pain 38, 321–32.
Kirsch, M., Terheggen, U., Hofmann, H.D., 2003. Ciliary neurotrophic factor is an early lesion-induced retrograde signal for axotomized facial motoneurons. Molecular and Cellular Neuroscience 24, 130–8.
Kisseleva, T., Brenner, D.A., 2008. Mechanisms of fibrogenesis. Experimental Biology and Medicine 233, 109–22.
Klimaschewski, L., Hausott, B., Angelov, D.N., 2013. The pros and cons of growth factors and cytokines in peripheral nerve regeneration, in: International Review of Neurobiology. pp. 137– 171.
Kokai, L.E., Bourbeau, D., Weber, D., McAtee, J., Marra, K.G., 2011. Sustained growth factor delivery promotes axonal regeneration in long gap peripheral nerve repair. Tissue Engineering. Part A 17, 1263–75.
Kokai, L.E., Ghaznavi, A.M., Marra, K.G., 2010. Incorporation of double-walled microspheres into polymer nerve guides for the sustained delivery of glial cell line-derived neurotrophic factor. Biomaterials 31, 2313–22.
Korte, N., Schenk, H.C., Grothe, C., Tipold, A., Haastert-Talini, K., 2011. Evaluation of periodic electrodiagnostic measurements to monitor motor recovery after different peripheral nerve lesions in the rat. Muscle & Nerve 44, 63–73.
Kramer, D.B., Xu, S., Kesselheim, A.S., 2010. Regulation of medical devices in the United States and European Union. The New England Jounal of Medicine 1–8.
Kreutzberg, G.W., 1995. Reaction of the neuronal cell body to axonal damage., in: TThe Axon: Structure, Function and Pathophysiology. Oxford University Press, Oxford, pp. 355–374.
Laurent, J.P., Lee, R.T., 1994. Topical Review: Birth-elated upper brachial plexus injuries in infants: operative and nonoperative approaches. Journal of Child Neurology 9, 111–7.

References
- 143 -
Lekholm, C., Sundkvist, G., Lundborg, G., Dahlin, L., 2001. The diabetic hand-complications of diabetes. Lakartidningen 98, 306–12.
Levi-Montalcini, R., 1987. The nerve growth factor: thirty-five years later. The EMBO Journal 6, 1145–54.
Levi-Montalcini, R., Angeletti, P.U., 1968. Nerve growth factor. Physiological Reviews 48, 534–69. Li, L., Wu, W., Lin, L.F., Lei, M., Oppenheim, R.W., Houenou, L.J., 1995. Rescue of adult mouse
motoneurons from injury-induced cell death by glial cell line-derived neurotrophic factor. Proceedings of the National Academy of Sciences of the United States of America 92, 9771–5.
Li, Q., Ping, P., Jiang, H., Liu, K., 2006. Nerve conduit filled with GDNF gene-modified Schwann cells enhances regeneration of the peripheral nerve. Microsurgery 26, 116–21.
Liebermann, A.R., 1971. The axon reaction: A review of the principal features of perikaryal repsonses to axon injury. International Review of Neurobiology 14, 49–124.
Lin, F.M., Pan, Y.C., Hom, C., Sabbahi, M., Shenaq, S., 1996. Ankle stance angle: a functional index for the evaluation of sciatic nerve recovery after complete transection. Journal of Reconstructive Microsurgery 12, 173–7.
Lin, S., Xu, J., Hu, S., Xu, L., Zhang, C., Wang, Y., Gu, Y., 2011. Combined application of neutrophin-3 gene and neural stem cells is ameliorative to delay of denervated skeletal muscular atrophy after tibial nerve transection in rats. Cell Transplantation 20, 381–90.
Lin, Y.-C., Marra, K.G., 2012. Injectable systems and implantable conduits for peripheral nerve repair. Biomedical Materials 7, 9 pages.
Liodaki, E., Bos, I., Lohmeyer, J.A., Senyaman, O., Mauss, K.L., Siemers, F., Mailaender, P., Stang, F., 2013. Removal of collagen nerve conduits (NeuraGen) after unsuccessful implantation: focus on histological findings. Journal of Reconstructive mMcrosurgery 29, 517–22.
Lohmeyer, J.A., Shen, Z.-L., Walter, G.F., Berger, A., 2007. Bridging extended nerve defects with an artifcial nerve graft containing Schwann cells pre-seeded on polyglactin filaments. The International Journal of Artificial Organs 30, 64–74.
Luis, A.L., Rodrigues, J.M., Amado, S., Veloso, A.P., Armada-Da-Silva, P.A.S., Raimondo, S., Fregnan, F., Ferreira, A.J., Lopes, M.A., Santos, J.D., Geuna, S., Varejão, A.S.P., Maurício, A.C., 2007. PLGA 90/10 and caprolactone biodegradable nerve guides for the reconstruction of the rat sciatic nerve. Microsurgery 27, 125–37.
Luís, A.L., Rodrigues, J.M., Geuna, S., Amado, S., Simões, M.J., Fregnan, F., Ferreira, A.J., Veloso, A.P., Armada-da-Silva, P.A.S., Varejão, A.S.P., Maurício, A.C., 2008. Neural cell transplantation effects on sciatic nerve regeneration after a standardized crush injury in the rat. Microsurgery 28, 458–70.
Lundborg, G., 2000. A 25-year perspective of peripheral nerve surgery: Evolving neuroscientific concepts and clinical significance. Journal of Hand Surgery 25, 391–414.
Lundborg, G., Dahlin, L.B., Danielsen, N.P., Hansson, H.A., Larsson, K., 1981. Reorganization and orientation of regenerating nerve fibres, perineurium, and epineurium in preformed mesothelial tubes - an experimental study on the sciatic nerve of rats. Journal of Neuroscience Research 6, 265–81.
Ma, Z., Gao, C., Yuan, J., Ji, J., Gong, Y., Shen, J., 2002. Surface modification of poly-L-lactide by photografting of hydrophilic polymers towards improving its hydrophilicity. Journal of Applied Polymer Science 85, 2163–71.
Maciel, F.O., Viterbo, F., Chinaque, L.D.F.C., Souza, B.M., 2013. Effect of electrical stimulation of the cranial tibial muscle after end-to-side neurorrhaphy of the peroneal nerve in rats. Acta Cirúrgica Brasileira 28, 39–47.
Mackinnon, S.E., Doolabh, V.B., Novak, C.B., Trulock, E.P., 2001. Clinical outcome following nerve allograft transplantation. Plastic and Reconstructive Surgery 107, 1419–29.
Madduri, S., Gander, B., 2010. Schwann cell delivery of neurotrophic factors for peripheral nerve regeneration. Journal of the Peripheral Nervous System 15, 93–103.
Madduri, S., Papaloïzos, M., Gander, B., 2009. Synergistic effect of GDNF and NGF on axonal branching and elongation in vitro. Neuroscience Research 65, 88–97.
Madduri, S., Papaloïzos, M., Gander, B., 2010. Trophically and topographically functionalized silk fibroin nerve conduits for guided peripheral nerve regeneration. Biomaterials 31, 2323–34.
Madihally, S. V., Matthew, H.W.T., 1999. Porous chitosan scaffolds for tissue engineering. Biomaterials 20, 1133–42.
Madison, R.D., da Silva, C., Dikkes, P., Sidman, R.L., Chiu, T.-H., 1987. Peripheral nerve regeneration with entubulation repair: Comparison of biodegradeable nerve guides versus polyethylene tubes and the effects of a laminin-containing gel. Experimental Neurology 95, 378–90.

References
- 144 -
Mainil-Varlet, P., Gogolewski, S., Nieuwenhuis, P., 1996. Long-term soft tissue reaction to various polylactides and their in vivo degradation. Journal of Materials Science: Materials in Medicine 7, 713–21.
Maluf-Meiken, L.C. V, Silva, D.R.M., Duek, E.A.R., Alberto-Rincon, M.C., 2006. Morphometrical analysis of multinucleated giant cells in subdermal implants of poly-lactic acid in rats. Journal of Materials Science: Materials in Medicine 17, 481–5.
Mancuso, R., Santos-Nogueira, E., Osta, R., Navarro, X., 2011. Electrophysiological analysis of a murine model of motoneuron disease. Clinical Neurophysiology 122, 1660–70.
Manuel, M., Zytnicki, D., 2011. Alpha, beta and gamma motoneurons: functional diversity in the motor system’s final pathway. Journal of Integrative Neuroscience 10, 243–76.
Marshall Devor and Ruth Govrin-Lippmann, 1979. Selective regeneration of sensory fibers following nerve crush injury. Experimental Neurology 65, 243–54.
McNally, A.K., Anderson, J.M., 2011. Macrophage fusion and multinucleated giant cells of inflammation. Advances in Experimental Medicine and Biology 713, 97–111.
McNally, A.K., Anderson, J.M., 2011. Foreign body-type multinucleated giant cells induced by interleukin-4 express select lymphocyte co-stimulatory molecules and are phenotypically distinct from osteoclasts and dendritic cells. Experimental and Molecular Pathology 91, 673–81.
Meek, M.F., Coert, J.H., 2008. US Food and Drug Administration/Conformit Europe-approved absorbable nerve conduits for clinical repair of peripheral and cranial nerves. Annals of Plastic Surgery 62, 110–6.
Meek, M.F., Den Dunnen, W.F.A., 2009. Porosity of the wall of a Neurolac nerve conduit hampers nerve regeneration. Microsurgery 29, 473–8.
Meek, M.F., Jansen, K., 2009. Two years after in vivo implantation of poly(DL-lactide-epsilon-caprolactone) nerve guides: has the material finally resorbed? Journal of Biomedical Materials Research Part A 89, 734–8.
Meisinger, C., Grothe, C., 2002. Differential regulation of fibroblast growth factor (FGF)-2 and FGF receptor 1 mRNAs and FGF-2 isoforms in spinal ganglia and sciatic nerve after peripheral nerve lesion. Journal of Neurochemistry 68, 1150–8.
Metz, G.A., Whishaw, I.Q., 2009. The ladder rung walking task: a scoring system and its practical application. Journal of Visualized Experiments 28, 1204.
Meyer, M., Matsuoka, I., Wetmore, C., Olson, L., Thoenen, H., 1992. Enhanced synthesis of brain-derived neurotrophic factor in the lesioned peripheral nerve: different mechanisms are responsible for the regulation of BDNF and NGF mRNA. The Journal of Cell Biology 119, 45–54.
Moore, A.M., Kasukurthi, R., Magill, C.K., Farhadi, H.F., Borschel, G.H., Mackinnon, S.E., 2009. Limitations of conduits in peripheral nerve repairs. Hand 4, 180–6.
Morais, J.M., Papadimitrakopoulos, F., Burgess, D.J., 2010. Biomaterials/tissue interactions: possible solutions to overcome foreign body response. The AAPS Journal 12, 188–96.
Mourad, P.D., Lazar, D.A., Curra, F.P., Mohr, B.C., Andrus, K.C., Avellino, A.M., McNutt, L.D., Crum, L.A., Kliot, M., 2001. Ultrasound accelerates functional recovery after peripheral nerve damage. Neurosurgery 48, 1136–1141.
Mukhatyar, V., Pai, B., Clements, I., Srinivasan, A., Huber, R., Mehta, A., Mukhopadaya, S., Rudra, S., Patel, G., Karumbaiah, L., Bellamkonda, R., 2014. Molecular sequelae of topographically guided peripheral nerve repair. Annals of Biomedical Engineering 42, 1436–55.
Murinson, B.B., Hoffman, P.N., Banihashemi, M.R., Meyer, R.A., Griffin, J.W., 2005. C-fiber (Remak) bundles contain both isolectin B4-binding and calcitonin gene-related peptide-positive axons. The Journal of Comparative Neurology 484, 392–402.
Navarro, X., Butí, M., Verdú, E., 1994. Autotomy prevention by amitriptyline after peripheral nerve section in different strains of mice. Restorative Neurology and Neuroscience 6, 151–7.
Navarro, X., Udina, E., 2009. Chapter 6: Methods and protocols in peripheral nerve regeneration experimental research: part III-electrophysiological evaluation., in: International Review of Neurobiology. pp. 105–26.
Navarro, X., Vivó, M., Valero-Cabré, A., 2007. Neural plasticity after peripheral nerve injury and regeneration. Progress in Neurobiology 82, 163–201.
Navissano, M., Malan, F., Carnino, R., Battiston, B., 2005. Neurotube for facial nerve repair. Microsurgery 25, 268–71.
Necas, J., Bartosikova, L., Brauner, P., Kolar, J., 2008. Hyaluronic acid (hyaluronan): a review. Veterinarni Medicina 53, 397–411.
Nectow, A.R., Marra, K.G., Kaplan, D.L., 2011. Biomaterials for the development of peripheral nerve guidance conduits. Tissue Engineering Part B Reviews 18, 40–50.

References
- 145 -
Nichols, C.M., Brenner, M.J., Fox, I.K., Tung, T.H., Hunter, D.A., Rickman, S.R., Mackinnon, S.E., 2004. Effects of motor versus sensory nerve grafts on peripheral nerve regeneration. Experimental Neurology 190, 347–55.
Nickel, P., Schummer, A., Seiferle, E., 2004. Peripheres Nervensystem, in: Lehrbuch Der Anatomie Der Haustiere Band IV. pp. 228–384.
Oh, S.H., Kim, J.H., Song, K.S., Jeon, B.H., Yoon, J.H., Seo, T.B., Namgung, U., Lee, I.W., Lee, J.H., 2008. Peripheral nerve regeneration within an asymmetrically porous PLGA/Pluronic F127 nerve guide conduit. Biomaterials 29, 1601–9.
Okamoto, H., Mizuno, K., Horio, T., 2003. Langhans-type and foreign-body-type multinucleated giant cells in cutaneous lesions of sarcoidosis. Acta Dermato-Venereologica 83, 171–4.
Oppenheim, R., Bartheld, C. von, 2008. Chapter 19: Programmed cell death and neurotrophic factors, in: Fundamental Neuroscience. pp. 438–444.
Oppenheim, R.W., Houenou, L.J., Johnson, J.E., Lin, L.F., Li, L., Lo, A.C., Newsome, A.L., Prevette, D.M., Wang, S., 1995. Developing motor neurons rescued from programmed and axotomy-induced cell death by GDNF. Nature 373, 344–6.
Oppenheim, R.W., von Bartheld, C.S., 2008. Chapter 19: Programmed cell death and neurotrophic factors, in: Fundamental Neuroscience. pp. 450–452.
Ornitz, D.M., Itoh, N., 2001. Fibroblast growth factors. Genome Biology 2, REVIEW3005. Otto, D., Unsicker, K., Grothe, C., 1987. Pharmacological effects of nerve growth factor and fibroblast
growth factor applied to the transectioned sciatic nerve on neuron death in adult rat dorsal root ganglia. Neuroscience Letters 83, 156–60.
Pabari, A., Yang, S., Mosahebi, A., Seifalian, A., 2011. Recent advances in artificial nerve conduit design: strategies for the delivery of luminal fillers. Journal of Controlled Release 156, 2–10.
Pabari, A., Yang, S., Seifalian, A., Mosahebi, A., 2010. Modern surgical management of peripheral nerve gap. Clinics in Plastic Surgery 63, 1941–8.
Panseri, S., Cunha, C., Lowery, J., Del Carro, U., Taraballi, F., Amadio, S., Vescovi, A., Gelain, F., 2008. Electrospun micro- and nanofiber tubes for functional nervous regeneration in sciatic nerve transections. BMC Biotechnology 8, 39.
Pardo, I.D., Garman, R.H., Weber, K., Bobrowski, W.F., Hardisty, J.F., Morton, D., 2012. Technical guide for nervous system sampling of the cynomolgus monkey for general toxicity studies. Toxicologic Pathology 40, 624–36.
Park, J.-S., Höke, A., 2014. Treadmill exercise induced functional recovery after peripheral nerve repair is associated with increased levels of neurotrophic factors. PloS One 9, e90245.
Patel, M., Mao, L., Wu, B., Vandevord, P.J., 2007. GDNF-chitosan blended nerve guides: a functional study. Journal of Tissue Engineering and Regenerative Medicine 1, 360–7.
Patel, M., Vandevord, P.J., Matthew, H., Wu, B., DeSilva, S., Wooley, P.H., 2006. Video-gait analysis of functional recovery of nerve repaired with chitosan nerve guides. Tissue Engineering 12, 3189–99.
Pfister, B.J., Gordon, T., Loverde, J.R., Kochar, A.S., Mackinnon, S.E., Cullen, D.K., 2011. Biomedical engineering strategies for peripheral nerve repair: Surgical applications, state of the art, and future challenges. Critical Reviews in Biomedical Engineering 39, 81–124.
Pfister, L.A., Papaloïzos, M., Merkle, H.P., Gander, B., 2007. Nerve conduits and growth factor delivery in peripheral nerve repair. Journal of the Peripheral Nervous System 12, 65–82.
Pistner, H., Gutwald, R., Ordung, R., Reuther, J., Mühling, J., 1993. Poly(L-lactide): a long-term degradation study in vivo. I. Biological results. Biomaterials 14, 671–7.
Polyganics.com, 2014. Polyganics-Bioresorbable medical devices [WWW Document]. Popesko, P., Rajtová, V., Horák, J., 1990. A color atlas of anatomy of small laboratory animals.
Volume two: rat, mouse, hamster. Raafat, D., Sahl, H.-G., 2009. Chitosan and its antimicrobial potential-a critical literature survey.
Microbial Biotechnology 2, 186–201. Radtke, C., Aizer, A.A., Agulian, S.K., Lankford, K.L., Vogt, P.M., Kocsis, J.D., 2009. Transplantation
of olfactory ensheathing cells enhances peripheral nerve regeneration after microsurgical nerve repair. Brain Research 1254, 10–7.
Radtke, C., Wewetzer, K., Reimers, K., Vogt, P.M., 2011. Transplantation of olfactory ensheathing cells as adjunct cell therapy for peripheral nerve injury. Cell Transplantation 20, 145–52.
Raimondo, S., Fornaro, M., Tos, P., Battiston, B., Giacobini-Robecchi, M.G., Geuna, S., 2011. Perspectives in regeneration and tissue engineering of peripheral nerves. Annals of Anatomy 193, 334–40.

References
- 146 -
Raisi, A., Azizi, S., Delirezh, N., Heshmatian, B., Amini, K., 2012. Use of chitosan conduit for bridging small-gap peripheral nerve defect in sciatic nerve transection model of rat. Iranian Journal of Veterinary Surgery 5, 89–100.
Ransom, B.R., 2012. Neuroglia (Google eBook). Oxford University Press. Ratzka, A., Kalve, I., Ozer, M., Nobre, A., Wesemann, M., Jungnickel, J., Koster-Patzlaff, C., Baron,
O., Grothe, C., 2012. The co-layer method as an efficient way to genetically modify mesencephalic progenitor cells transplanted into 6-OHDA rat model of Parkinson’s disease. Cell Transplantion 21, 749–62.
Ray, W. Z., Mackinnon, S. E., 2010. Management of nerve gaps: Autografts, allografts, nerve transfers, and end-to-side neurorrhaphy. Experimental Neurology 223, 77–85.
Reichert, F., Saada, A., Rotshenker, S., 1994. Peripheral nerve injury induces Schwann cells to express two macrophage phenotypes: phagocytosis and the galactose-specific lectin MAC-2. The Journal of Neuroscience 14, 3231–45.
Rhee, H.J., Hillebrands, W., Daems, W.T., 1978. Are Langhans giant cells precursors of foreign-body giant cells? Archives of Dermatological Research 263, 13–21.
Rigaud, M., Gemes, G., Barabas, M.-E., Chernoff, D.I., Abram, S.E., Stucky, C.L., Hogan, Q.H., 2008. Species and strain differences in rodent sciatic nerve anatomy: implications for studies of neuropathic pain. Pain 136, 188–201.
Robinson, L.R., 2000. Traumatic injury to peripheral nerves. Muscle & Nerve 23, 863–73. Rochkind, S., El-Ani, D., Nevo, Z., Shahar, A., 2009. Increase of neuronal sprouting and migration
using 780 nm laser phototherapy as procedure for cell therapy. Lasers in Surgery and Medicine 41, 277–81.
Rumpel, R., Alam, M., Klein, A., Oezer, M., Wesemann, M., Jin, X., Krauss, J.K., Schwabe, K., Ratzka, A., Grothe, C., 2013. Neuronal firing activity and gene expression changes in the subthalamic nucleus after transplantation of dopamine neurons in hemiparkinsonian rats. Neurobiology of Disease 59, 230–243.
Rupp, A., Dornseifer, U., Fischer, A., Schmahl, W., Rodenacker, K., Jütting, U., Gais, P., Biemer, E., Papadopulos, N., Matiasek, K., 2007. Electrophysiologic assessment of sciatic nerve regeneration in the rat: surrounding limb muscles feature strongly in recordings from the gastrocnemius muscle. Journal of Neuroscience Methods 166, 266–77.
Saito, H., Dahlin, L.B., 2008. Expression of ATF3 and axonal outgrowth are impaired after delayed nerve repair. BMC Neuroscience 9, 88.
Samadian, M., Rezaee, O., Haddadian, K., Sharifi, G., Abtahi, H., Hamidian, M., Khormaee, F., Sodagari, F., 2009. Gunshot injuries to the brachial plexus during wartime. British Journal of Neurosurgery 23, 165–9.
Sandkühler, J., 2009. Models and mechanisms of hyperalgesia and allodynia. Physiological Reviews 89, 707–58.
Santosa, K.B., Jesuraj, N.J., Viader, A., MacEwan, M., Newton, P., Hunter, D.A., Mackinnon, S.E., Johnson, P.J., 2013. Nerve allografts supplemented with Schwann cells overexpressing glial-cell-line-derived neurotrophic factor. Muscle & Nerve 47, 213–23.
Sarikcioglu, L., Demirel, B.M., Utuk, A., 2009. Walking track analysis: an assessment method for functional recovery after sciatic nerve injury in the rat. Folia Morphologica 68, 1–7.
Scheib, J., Höke, A., 2013. Advances in peripheral nerve regeneration. Nature Reviews. Neurology. 9, 668–76.
Scholz, T., Sumarto, A., Krichevsky, A., Evans, G.R.D., 2011. Neuronal differentiation of human adipose tissue-derived stem cells for peripheral nerve regeneration in vivo. Archives of Surgery 146, 666–74.
Seckel, B.R., 1990. Enhancement of peripheral nerve regeneration. Muscle & Nerve 13, 785–800. Seddon, H.J., 1942. A Classification of Nerve Injuries. British Medical Journal 2, 237–9. Seddon, H.J., Medawar, P.B., Smith, H., 1943. Rate of regeneration of peripheral nerves in man. The
Journal of Physiology 102, 191–215. Selden, P.A., Nudds, J.R., 2004. Chapter 1: Ediacara, in: Evolution of Fossil Ecosystems. p. 10. Shahar, A., Nevo, Z., Rochkind, S., 2001. Cross-linked hyaluronic acid-laminin gels and use thereof in
cell culture and medical implants. Shen, H., Shen, Z.-L., Zhang, P.-H., Chen, N.-L., Wang, Y.-C., Zhang, Z.-F., Jin, Y.-Q., 2010. Ciliary
neurotrophic factor-coated polylactic-polyglycolic acid chitosan nerve conduit promotes peripheral nerve regeneration in canine tibial nerve defect repair. Journal of Biomedical Materials Research. Part B, Applied biomaterials 95, 161–70.
Shen, X., Ruan, J., Zhou, Z., Zeng, Z., Xie, L., 2009. Evaluation of PLGA/chitosan/HA conduits for nerve tissue reconstruction. Journal of Wuhan University of Technology 24, 566–70.

References
- 147 -
Shenaq, J.M., Shenaq, S.M., Spira, M., 1989. Reliability of sciatic function index in assessing nerve regeneration across a 1 cm gap. Microsurgery 10, 214–9.
Siemionow, M., Bozkurt, M., Zor, F., 2010. Regeneration and repair of peripheral nerves with different biomaterials: review. Microsurgery 30, 574–88.
Sierpinski Hill, P., 2009. Regeneration of peripheral nerves using neuroinductive biomaterial scaffolds. Simões, M., Gärtner, A., Shirosaki, Y., Costa, R.G. da, Cortez, P., Gartnër, F., Santos, J., Lopes, M.,
Geuna, S., Varejão, A., Maurício, A., 2011. In vitro and in vivo chitosan for peripheral nerve reconstruction. Acta Medica Portuguesa 24, 43–52.
Simões, M.J., Amado, S., Gärtner, A., Armada-Da-Silva, P.A.S., Raimondo, S., Vieira, M., Luís, A.L., Shirosaki, Y., Veloso, A.P., Santos, J.D., Varejão, A.S.P., Geuna, S., Maurício, A.C., 2010. Use of chitosan scaffolds for repairing rat sciatic nerve defects. Italian Journal of Anatomy and Embryology 115, 190–210.
Singh, D.K., Ray, A.R., 2000. Biomedical applications of chitin, chitosan, and their derivatives. Journal of Macromolecular Science, Part C: Polymer Reviews 40, 69–83.
Sinis, N., Schaller, H.-E., Schulte-Eversum, C., Schlosshauer, B., Doser, M., Dietz, K., Rösner, H., Müller, H.-W., Haerle, M., 2005. Nerve regeneration across a 2-cm gap in the rat median nerve using a resorbable nerve conduit filled with Schwann cells. Journal of Neurosurgery 103, 1067–76.
Smetana, K., 1987. Multinucleate foreign-body giant cell formation. Experimental and Molecular Pathology 46, 258–65.
Snider, W.D., 1994. Functions of the neurotrophins during nervous system development: what the knockouts are teaching us. Cell 77, 627–38.
Sotgiu, M.L., Biella, G., 1995. Spinal expansion of saphenous afferents after sciatic nerve constriction in rats. Neuroreport 6, 2305–8.
Stoll, G., Jander, S., Myers, R.R., 2002. Degeneration and regeneration of the peripheral nervous system: from Augustus Waller’s observations to neuroinflammation. Journal of the peripheral nervous system 7, 13–27.
Strauch, B., Rodriguez, D.M., Diaz, J., Yu, H.L., Kaplan, G., Weinstein, D.E., 2001. Autologous Schwann cells drive regeneration through a 6-cm autogenous venous nerve conduit. Journal of Reconstructive Microsurgery 17, 589–95; discussion 596–7.
Strusycyzk, M.H., Struszczyk, K.J., 2007. Medical applications of chitin and its derivatives, in: Polish Chitin Society. pp. 139– 147.
Sun, W., Miao, B., Wang, X.-C., Duan, J.-H., Wang, W.-T., Kuang, F., Xie, R.-G., Xing, J.-L., Xu, H., Song, X.-J., Luo, C., Hu, S.-J., 2012. Reduced conduction failure of the main axon of polymodal nociceptive C-fibres contributes to painful diabetic neuropathy in rats. Brain 135, 359–75.
Sunderland, S., 1951. A classification of peripheral nerve injuries producing loss of function. Brain 74, 491–516.
Sunderland, S., 1990. The anatomy and physiology of nerve injury. Muscle & Nerve 13, 771–84. Svennigsen, Å., Dahlin, L., 2013. Repair of the Peripheral Nerve-Remyelination that Works. Brain
Sciences 3, 1182–97. Swett, J.E., Torigoe, Y., Elie, V.R., Bourassa, C.M., Miller, P.G., 1991. Sensory neurons of the rat
sciatic nerve. Experimental Neurology 114, 82–103. Swett, J.E., Wikholm, R.P., Blanks, R.H., Swett, A.L., Conley, L.C., 1986. Motoneurons of the rat
sciatic nerve. Experimental Neurology 93, 227–52. Swindle-Reilly, K.E., Papke, J.B., Kutosky, H.P., Throm, A., Hammer, J.A., Harkins, A.B., Willits, R.K.,
2012. The impact of laminin on 3D neurite extension in collagen gels. Journal of Neural Engineering 9, 046007.
Tajima, T., Imai, H., 1989. Results of median nerve repair in children. Microsurgery 10, 145–6. Thevenot, P., Hu, W., Tang, L., 2008. Surface chemistry influences implant biocompatibility. Current
Topics in Medicinal Chemistry 8, 270–80. Thomas, P.K., 1989. Invited review: focal nerve injury: guidance factors during axonal regeneration.
Muscle & Nerve 12, 796–802. Thorsén, F., Rosberg, H.-E., Steen Carlsson, K., Dahlin, L.B., 2012. Digital nerve injuries:
epidemiology, results, costs, and impact on daily life. Journal of Plastic Surgery and Hand Surgery 46, 184–90.
Timmer, M., Robben, S., Müller-Ostermeyer, F., Nikkhah, G., Grothe, C., 2003. Axonal regeneration across long gaps in silicone chambers filled with Schwann cells overexpressing high molecular weight FGF-2. Cell Transplantation 12, 265–77.
Tomihata, K., Ikada, Y., 1997. In vitro and in vivo degradation of films of chitin and its deacetylated derivatives. Biomaterials 18, 567–75.

References
- 148 -
Tonra, J.R., Curtis, R., Wong, V., Cliffer, K.D., Park, J.S., Timmes, A., Nguyen, T., Lindsay, R.M.,
Acheson, A., DiStefano, P.S., 1998. Axotomy upregulates the anterograde transport and expression of brain-derived neurotrophic factor by sensory neurons. The Journal of Neuroscience 18, 4374–83.
Törnqvist, E., Aldskogius, H., 1994. Motoneuron survival is not affected by the proximo-distal level of axotomy but by the possibility of regenerating axons to gain access to the distal nerve stump. Journal of Neuroscience Research 39, 159–65.
Udina, E., Rodríguez, F.J., Verdú, E., Espejo, M., Gold, B.G., Navarro, X., 2004. FK506 enhances regeneration of axons across long peripheral nerve gaps repaired with collagen guides seeded with allogeneic Schwann cells. Glia 47, 120–9.
Ueda, H., 2006. Molecular mechanisms of neuropathic pain-phenotypic switch and initiation mechanisms. Pharmacology and Therapeutics.
Ushiki, T., 2002. Collagen fibers, reticular fibers and elastic fibers. A comprehensive understanding from a morphological viewpoint. Archives of Histology and Cytology 65, 109–26.
Valentini, R.F., Aebischer, P., Winn, S.R., Galletti, P.M., 1987. Collagen- and laminin-containing gels impede peripheral nerve regeneration through semipermeable nerve guidance channels. Experimental Neurology 98, 350–6.
Valero-Cabré, A., Tsironis, K., Skouras, E., Navarro, X., Neiss, W.F., 2004. Peripheral and spinal motor reorganization after nerve injury and repair. Journal of Neurotrauma 21, 95–108.
Valero-Cabré, A., Tsironis, K., Skouras, E., Perego, G., Navarro, X., Neiss, W.F., 2001. Superior muscle reinnervation after autologous nerve graft or poly-L-lactide-epsilon-caprolactone (PLC) tube implantation in comparison to silicone tube repair. Journal of Neuroscience Research 63, 214–23.
Varejão, A.S.P., Cabrita, A.M., Meek, M.F., Fornaro, M., Geuna, S., Giacobini-Robecchi, M.G., 2003. Morphology of nerve fiber regeneration along a biodegradable poly (DLLA-epsilon-CL) nerve guide filled with fresh skeletal muscle. Microsurgery 23, 338–45.
Verdu, E., Ceballos, D., Vilches, J.J., Navarro, X., 2000. Influence of aging on peripheral nerve function and regeneration. Journal of the Peripheral Nervous System 5, 191–208.
Waitayawinyu, T., Parisi, D.M., Miller, B., Luria, S., Morton, H.J., Chin, S.H., Trumble, T.E., 2007. A comparison of polyglycolic acid versus type 1 collagen bioabsorbable nerve conduits in a rat model: an alternative to autografting. The Journal of Hand Surgery 32, 1521–9.
Waldram, M., 2003. Peripheral nerve injuries. Trauma 5, 79–96. Wang, G., Lu, G., Ao, Q., Gong, Y., Zhang, X., 2010. Preparation of cross-linked carboxymethyl
chitosan for repairing sciatic nerve injury in rats. Biotechnology Letters 32, 59–66. Wang, H., Spinner, R.J., Sorenson, E.J., Windebank, A.J., 2008. Measurement of forelimb function by
digital video motion analysis in rat nerve transection models. Journal of the peripheral nervous system : JPNS 13, 92–102.
Wang, K.K., Nemeth, I.R., Seckel, B.R., Chakalis-Haley, D.P., Swann, D.A., Kuo, J.W., Bryan, D.J., Cetrulo, C.L., 1998. Hyaluronic acid enhances peripheral nerve regeneration in vivo. Microsurgery 18, 270–5.
Wang, S., Cai, Q., Hou, J., Bei, J., Zhang, T., Yang, J., Wan, Y., 2003. Acceleration effect of basic fibroblast growth factor on the regeneration of peripheral nerve through a 15-mm gap. Journal of Biomedical Materials Research. Part A 66, 522–31.
Wang, S., Wan, A.C., Xu, X., Gao, S., Mao, H.Q., Leong, K.W., Yu, H., 2001. A new nerve guide conduit material composed of a biodegradable poly(phosphoester). Biomaterials 22, 1157–69.
Wang, X., Hu, W., Cao, Y., Yao, J., Wu, J., Gu, X., 2005. Dog sciatic nerve regeneration across a 30-mm defect bridged by a chitosan/PGA artificial nerve graft. Brain 128, 1897–910.
Waterman, E.A., Sakai, N., Nguyen, N.T., Horst, B.A.J., Veitch, D.P., Dey, C.N., Ortiz-Urda, S., Khavari, P.A., Marinkovich, M.P., 2007. A laminin-collagen complex drives human epidermal carcinogenesis through phosphoinositol-3-kinase activation. Cancer Research 67, 4264–70.
Wei, Y., Gong, K., Zheng, Z., Wang, A., Ao, Q., Gong, Y., Zhang, X., 2011. Chitosan/silk fibroin-based tissue-engineered graft seeded with adipose-derived stem cells enhances nerve regeneration in a rat model. Journal of Materials Science. Materials in Medicine 22, 1947–64.
Wenling, C., Duohui, J., Jiamou, L., Yandao, G., Nanming, Z., Xiufang, Z., 2005. Effects of the degree of deacetylation on the physicochemical properties and Schwann cell affinity of chitosan films. Journal of Biomaterials Applications 20, 157–177.
Westfall, I.A., 1996. Ultrastructure of synapses in the first-evolved nervous systems. Journal of Neurocytology 25, 735–46.

References
- 149 -
Williams, L.R., Longo, F.M., Powell, H.C., Lundborg, G., Varon, S., 1983. Spatial-temporal progress of peripheral nerve regeneration within a silicone chamber: parameters for a bioassay. The Journal of Comparative Neurology 218, 460–70.
Wood, M.D., Gordon, T., Kemp, S.W.P., Liu, E.H., Kim, H., Shoichet, M.S., Borschel, G.H., 2013a. Functional motor recovery is improved due to local placement of GDNF microspheres after delayed nerve repair. Biotechnology and Bioengineering 5, 1272–81.
Wood, M.D., Gordon, T., Kim, H., Szynkaruk, M., Phua, P., Lafontaine, C., Kemp, S.W., Shoichet, M.S., Borschel, G.H., 2013b. Fibrin gels containing GDNF microspheres increase axonal regeneration after delayed peripheral nerve repair. Regenerative Medicine 8, 27–37.
Woodbury, M.E., Ikezu, T., 2014. Fibroblast growth factor-2 signaling in neurogenesis and neurodegeneration. Journal of Neuroimmune Pharmacology 9, 92–101.
Wrobel, S., Serra, S.C., Ribeiro-Samy, S., Sousa, N., Heimann, C., Barwig, C., Grothe, C., Salgado, A.J., Haastert-Talini, K., 2014. In vitro evaluation of cell-seeded chitosan films for peripheral nerve tissue engineering. Tissue Engineering. Part A Epub ahead.
Xie, F., Li, Q.F., Gu, B., Liu, K., Shen, G.X., 2008. In vitro and in vivo evaluation of a biodegradable chitosan-PLA composite peripheral nerve guide conduit material. Microsurgery 28, 471–479.
Xu, H., Yan, Y., Li, S., 2011. PDLLA/chondroitin sulfate/chitosan/NGF conduits for peripheral nerve regeneration. Biomaterials 32, 4506–16.
Xue, C., Hu, N., Gu, Y., Yang, Y., Liu, Y., Liu, J., Ding, F., Gu, X., 2012. Joint use of a chitosan/PLGA scaffold and MSCs to bridge an extra large gap in dog sciatic nerve. Neurorehabilitation and Neural Repair 26, 96–106.
Yan, Q., Matheson, C., Lopez, O.T., 1995. In vivo neurotrophic effects of GDNF on neonatal and adult facial motor neurons. Nature 373, 341–4.
Yang, Y., Ding, F., Wu, J., Hu, W., Liu, W., Liu, J., Gu, X., 2007. Development and evaluation of silk fibroin-based nerve grafts used for peripheral nerve regeneration. Biomaterials 28, 5526–35.
Yang, Y., Gu, X., Tan, R., Hu, W., Wang, X., Zhang, P., Zhang, T., 2004. Fabrication and properties of a porous chitin/chitosan conduit for nerve regeneration. Biotechnology Letters 26, 1793–7.
Yang, Y., Wu, J., Wang, X., Liu, J., Ding, F., Gu, X., 2009. Fabrication and evaluation of chitin-based nerve guidance conduits used to promote peripheral nerve regeneration. Advanced Engineering Materials 11, B209–B218.
Yuan, Y., Zhang, P., Yang, Y., Wang, X., Gu, X., 2004. The interaction of Schwann cells with chitosan membranes and fibers in vitro. Biomaterials 25, 4273–8.
Zhang, F., Blain, B., Beck, J., Zhang, J., Chen, Z., Chen, Z.-W., Lineaweaver, W.C., 2002. Autogenous venous graft with one-stage prepared Schwann cells as a conduit for repair of long segmental nerve defects. Journal of Reconstructive Microsurgery 18, 295–300.
Zhang, H., Neau, S.H., 2001. In vitro degradation of chitosan by a commercial enzyme preparation: effect of molecular weight and degree of deacetylation. Biomaterials 22, 1653–8.
Zheng, L., Ao, Q., Han, H., Zhang, X., Gong, Y., 2010. Evaluation of the chitosan/glycerol-beta-phosphate disodium salt hydrogel application in peripheral nerve regeneration. Biomedical Materials 5, 35003.
Zheng, L., Cui, H.-F., 2012. Enhancement of nerve regeneration along a chitosan conduit combined with bone marrow mesenchymal stem cells. Journal of Materials Science: Materials in Medicine.
Ziv-Polat, O., Shahar, A., Levy, I., Skaat, H., Neumann, S., Fregnan, F., Geuna, S., Grothe, C., Haastert-Talini, K., Margel, S., 2014. The role of neurotrophic factors conjugated to iron oxide nanoparticles in peripheral nerve regeneration: In vitro studies. BioMed Research International. Article ID 267808, 10 Pages.
Zochodne, D.W., Cheng, C., 2000. Neurotrophins and other growth factors in the regenerative milieu of proximal nerve stump tips. Journal of Anatomy 196, 279–83.

Affidavit
- 150 -
8. Affidavit
I herewith declare that I autonomously carried out the PhD-thesis entitled “Peripheral nerve regeneration using hollow and enriched chitosan-based guidance conduits”.
No third party assistance has been used.
I did not receive any assistance in return for payment by consulting agencies or any other person. No one received any kind of payment for direct or indirect assistance in correlation to the content of the submitted thesis.
I conducted the project at the following institution: Institute of Neuroanatomy Hannover Medical School The thesis has not been submitted elsewhere for an exam, as a thesis or for evaluation in a similar context. I hereby affirm the above statements to be complete and true to the best of my knowledge. date, signature

- 151 -

Acknowledgements
- 152 -
9. Acknowledgements The last three years have been a unique journey, which would not have been the
same without the people accompanying me during this time. Therefore, I would like
to thank them in the next lines.
My first thank you belongs to my main supervisor Prof. Dr. Claudia Grothe for
providing me with the opportunity to be a part of this great EU-project and to work in
the research field of peripheral nerve regeneration. Thank you for your scientific
advice and support especially during the final months of this work.
Thank you to my co-supervisors Prof. Dr. Gerardy-Schahn and Prof. Dr. Schwabe for
your support and helpful input during these years. And thank you to Esther Udina,
MH, PhD, for agreeing to be my external referee for this work.
A special thank you to Prof. Dr. Kirsten Haastert-Talini for all the work you put into
the projects, the uncountable hours spend together doing electrophysiological
recordings and surgeries, laughs we shared, and for always making time to discuss
and answer questions.
Thank you to the whole Medovent-Team for allowing me to gain an insight into the
manufacturing of chitosan-based products, introducing me to carnival (Fasching),
Kräppel and for having a great time in Mainz.
Thank you to the whole BIOHYBRID-consortium that allowed me to meet so many
excellent researchers from all over Europe.
I also would like to thank Maike Wesemann, Silvana Taubeler-Gerling and Hildegard
Streich for all the work you did for the BIOHYBRID-project.
A huge thank you to Natascha Heidrich and Silke Fischer for every single section of
the approximately thousands you did in the course of my studies. I think we can fill a
whole room with all the boxes of sections you cut and stained. No kidding!

Acknowledgements
- 153 -
Thank you to Kerstin Kuhlemann for taking me in to your aisle during my long
journey to find my final desk, all the encouraging words, and helpful comments.
Thank, thank, thank you to Jenny Metzen! We spend so much time together looking
at rat feet and I could not imagine another person I wanted to do this with. Also thank
you for transportation help during rain, snow, and wind and bringing a smile to my
face in so many situations. And let’s not forget the great laughs we had over a glass
of wine.
Thank you to my current and past office-co-owners of the last 1 ½ years. Dr. Regina
Rumpel, Dr. Anna Effenberg, Michela Morano, Dr. Alex Klein, and Dr. Andreas
Ratzka. I’m glad I somehow managed to snatch a seat in your office (ok, I kind of
creped in), because I enjoyed so many moments with all of you. A special thank you
for tolerating the looks of my desk(s) during the last months, but I guess this is how it
has to be shortly before thesis-submission. Thank you to Dr. Andreas Ratzka for
helping me with all things regarding PCR and giving me my very own shrimp. Thank
you Dr. Regina Rumpel for sharing a sparkling weekend with me in Berlin and the
love for certain entertainment shows and books. Thank you Dr. Anna Effenberg for
great movie nights and sometimes talking weird, which always causes laughs. Thank
you to Dr. Alex Klein for all the encouragements regarding work and life. Thank you
Michela for being a part of our team for some months and for always having a smile
on your face when entering the office early in the morning.
Dr. Meltem Hohmann, Inga Stockbrügger, Verena Lübben, Michela Morano, Dr.
Benjamin Förthmann, Dr. Niko Hensel, Dr. Regina Rumpel, Sebastian Rademacher,
Sandra Wrobel, Jenny Metzen, Timo Konen, Maria Stößel thank you all for the so
much needed coffee breaks, candy breaks, cake breaks, pizza breaks as well as all
the evenings we spend together. I would never want to exchange this team for
another.
Thank you to Sandra Wrobel and Natalie Schecker for counting macrophages for
hours, days, and months!

Acknowledgements
- 154 -
A huge thank you to Sandra Wrobel for being my BIOHYBRID-partner of three years.
We shared so many moments and experiences in the office, during travels, and
evenings that I will cherish forever. Thank you for a special boat trip in Barcelona,
loving the Sushi-Groupon as much as I do, and watching almost all matches from the
German soccer team on their road to win the World Cup with me.
Thank you to Mariana Behrmann, Jana Wohltmann, and Nora Schaffarczyk for being
close even when there is an ocean between us. Thank you Mariana for the laughs
we shared on Skype and doing a US trip with me that will be a once in a lifetime
experience. Thank you Jana for all the talks we had, being the best friend one can
ask for and for just being who you are. Thank you Nora for all the videos and photos
that keep me up-to-date when you are again enjoying the sun in Florida, they really
have brightened my days.
Thank you to all my friends in Bremen, Jessica Wöhlke, Susanne Adrich, Jule
Hägermann, Mareike Hinze, Sarah Schreck, Julia Bauer, Svenja Ludwig, Tanja
Behnke, Andreas Kerzel, and Till Sonnenberg for being a part of my life since my
childhood or teenage years.
Thank you to Julia Budde for sharing my addiction to concerts and loving as many
music styles as I do. I hope we will attend hundreds more and continue to have great
talks on our ways to the venues.
Last but not least, I would like to thank my parents, Dietlinde and Gerd Meyer, for
their continuous support and being such a big part in my life. Thank you for always
being there, being patient, and sometimes reminding me of all the possibilities life
has to offer. I love you.