tissue site-specific enhancer function of the upstream regulatory
TRANSCRIPT

Vol. 63, No. 2JOURNAL OF VIROLOGY, Feb. 1989, p. 957-9600022-538X/89/020957-04$02.00/0Copyright © 1989, American Society for Microbiology
Tissue Site-Specific Enhancer Function of the Upstream RegulatoryRegion of Human Papillomavirus Type 11 in Cultured Keratinocytes
BETTIE M. STEINBERG,'* KAREN J. AUBORN,1 JANET L. BRANDSMA,1 AND LORNE B. TAICHMAN2
Department of Otolaryngology and Communicative Disorders, Long Island Jewish Medical Center, New Hyde Park, NewYork 11042,1 and Department of Oral Biology and Pathology, School ofDental Medicine, State University ofNew York
at Stony Brook, Stony Brook, New York 117942
Received 18 July 1988/Accepted 24 October 1988
The human papillomavirus type 11 regulatory region was cloned upstream of a reporter complex andmicroinjected into nuclei of individual primary human keratinocytes. Genital and laryngeal keratinocytes,normal host cells for this virus, exhibited higher levels of expression than cutaneous keratinocytes. We concludethat a papillomavirus enhancer(s) shows preferences among epithelial cells from different tissue sites.
Human papillomaviruses (HPVs) cause benign papillomasor warts (for a review, see reference 13) and are involved ingenital tract malignancies (20). The HPVs are epitheliotropicand have some tissue site specificity (12). For example, HPVtype 11 (HPV-11) is found predominantly in lesions of genitalepithelium and mucosal surfaces, not epidermis. Kreider etal. (9) found that epithelial tissues from these same sitesgrow as papillomas in nude mice after infection with HPV-11virions. The basis for this tissue site preference is unknown.Many cellular and viral enhancers show cell type speci-
ficity and are responsible for tissue-specific expression (10,11). HPVs contain a number of constitutive and inducibleenhancer sequences in the upstream regulatory region(URR) (1, 2, 7, 8), including one that appears to be kerati-nocyte specific (1). To our knowledge, no one has describedenhancers in the URR that show a preference for differentepithelial tissue sites.
In this study, we asked if the tropism of HPV-11 forcertain epithelial tissues is regulated in part by geneticelements in the URR. To do this, we linked the URR ofHPV-11 to the enhancer-minus segment of the simian virus40 (SV40) early region and introduced the construct intohuman keratinocyte cultures from a variety of sites. Toensure that all cells received similar amounts ofDNA and togauge the response of individual cells, the construct wasmicroinjected into the nuclei of the cells. Transfection, themost commonly used method of gene transfer in these typesof studies, is especially inefficient in primary cells, making itdifficult to compare different cell isolates, and does notprovide information about individual cell variability. There-fore, we used microinjection.Two recombinant plasmids were constructed for this
study. The enhancer-minus pPT-1 was generated by cloningthe SV40 promoter and T-antigen (Tag) gene, nucleotides(nt) 200 to 2533, into pGEM3. pPT-11R was cloned byinserting the URR of HPV-11 (nt 7178 to 33) upstream of theSV40 promoter in pPT-1, in the same orientation relative tothe promoter as exists in HPV-11. The positive control DNAwas pPP1, with the SV40 early region (nt 294 to 2533) clonedinto pfx3 (J. Sambrook, personal communication). The con-structs were linearized in plasmid sequences prior to micro-injection. Chloramphenicol acetyl transferase constructsnormally used for enhancer assays were not used because
* Corresponding author.
chloramphenicol acetyl transferase was not detectable withantibodies.Primary laryngeal and epidermal keratinocytes were cul-
tured as previously described (4, 15) and were plated onmarked glass cover slips in MCDB 153 (19; Clonetics, Inc.).Nearly confluent monolayers were fed Ham F12 mediumcontaining 10% fetal calf serum and 10 mM hydrocortisoneto enhance flattening of the cells. Fibroblasts derived fromlarynx and foreskin tissues were cultured in Dulbecco mod-ified Eagle medium with 10% fetal calf serum. A minimum of100 cells were microinjected with each DNA construct asdescribed (3). Cells were fixed with methanol-acetone (1:1,vol/vol) 20 to 22 h after injection with monoclonal antibodypAB-416 and were stained against Tag (6), avidin-biotin-peroxidase complex (Vector Laboratories) and diaminoben-zidine. This early detection of transient Tag expression wasused to minimize autoregulation of the SV40 promoter (5, 14)or heterologous regulation of the type seen with HPV-18after 48 h (18).To determine if the frequency of Tag-positive cells was
related to the amount of DNA injected, laryngeal kerati-nocytes were injected with 2, 20, and 50 copies of the variousconstructs (Fig. 1). At 50 copies per cell, the enhancer-minuspPT-1 yielded a significant number of Tag-positive nuclei.Injections of 2 and 20 copies per cell gave nearly equalpercentages of positive cells with all constructs. The HPV-11URR was able to efficiently substitute for the SV40 enhancerin these cells. All further experiments were done with 2 ,ug ofDNA (two to four copies per cell) per ml, to keep the pPT-1background as low as possible. It has previously been shownthat the variation in the injected volume ranges from 1 x10-11 to 2 x 10-11 ml per cell (3). This variation is inherentin the.technique.To confirm that we were, in fact, detecting an enhancer-
like function, the HPV-11 URR was also tested in theopposite orientation, resulting in 62% Tag-positive cells. Asimilar increase in enhancer function with an HPV-16 con-struct has been reported by Cripe et al. (1). We chose to testfor tissue specificity with the URR orientation found in thevirus itself.When the constructs were tested in different types of cells,
both the percentage of Tag-positive cells and the intensity ofstaining within the nuclei varied. The differences in stainingintensity can be seen in Fig. 2. To quantitate this, the percentlight absorption was measured for each stained nucleus witha photometer attached to a Leitz microscope. Results were
957

958 NOTESJ.VO.
80 1
701-cn I
C-,5Q-
0 0
10
M2 COPIES/CELL=120 COPIES/CELL=50 COPIES/CELL.1 IHK 1K~~~~~~5
pPT- 1
FIG. 1. Titration of DNA constructs (enhancer-minus pPT-1[pPT-1], pPl, and pPT-11R) used for microinjection experiments.DNAs were adjusted to 2, 20, or 50 jig/ml in phosphate-bufferedsaline, and approximately 10-11 ml was microinjected per cell (3)into cultured laryngeal epithelial cells to give 2, 20, or 50 copies ofDNA per cell. Cells were stai'ned for Tag with monoclonal antibodypAB-416 (6) 20 to 22 h later, and the percentage of injected cells thatwere Tag positive was determined. Note the high background levelsseen with the enhancer-minus pPT-1 when 50 copies per cell wereinjected.
LARYNX
AV
NECK A
pPl pPT- 1 1R
standardized using the SV40 transformed cell line (16),which was stained with each experiment to correct for smallvariations in backgro'und staining.The ability of the constructs to function in different cells is
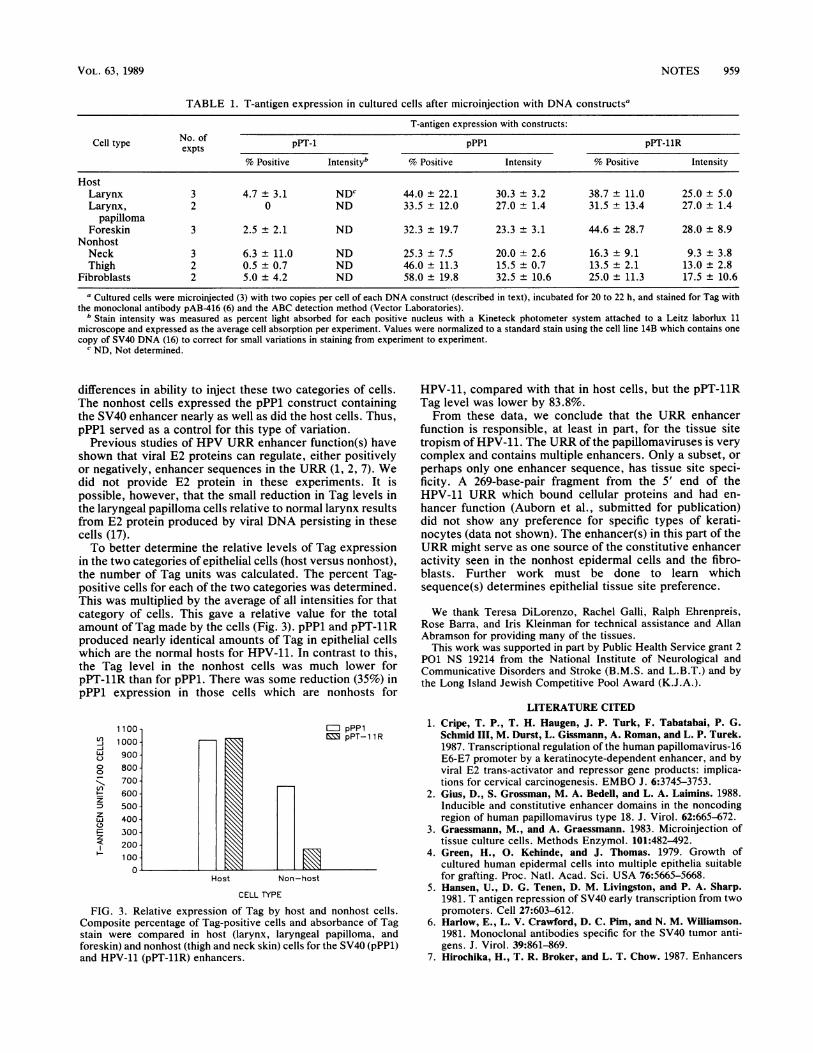
shown in Table 1. Epithelial cells from the normal hosttissues for HPV-11 (larynx and foreskin), as well as thosefrom the laryngeal papillomas, showed similar percentagesof Tag-po'sitive cells and staining intensities with both pPP1and pPT-11R. In contrast, both the percent Tag-positivecells and the intensities for pPT-11R 'in the thigh and neckskin cells (nonhost for HPV-11) were clearly lower than theresults for pPP1. In fact, the values for these cells werelower than for fibroblasts. Fibroblasts are nonpermissive forHPV replication and do not function as iargets for theseviruses in vivo; thus, they served as controls to evaluate thelevel of non-site-specific enhancer function.The variation in the percentage of Tag-positive cells from
experiment to experiment could reflect differences in theability to inject the cells, differences in the cells fromdifferent patients, or both. It is difficult to microinjectepithelial cells, since the penetration of the needle is affectedby the state of differentiation of the cell. However, once thecells were satisfactorily injected, the variabil'ity in intensityamong experiments with the same cell type was low. Thedifferences between host and nonhost cells were not due to
F 0
*A
#K
pPl 11RFIG. 2. Variation in staining intensity seen when DNA constructs were microinjected into different cultured epithelial cells. Cells were
injected with 2 pLg (2 copies per cell) of pPl or pPT-11R (11R) DNA and stained 20 to 22 h later for Tag. Only a portion of the injected areais shown. The difference in distribution of stained nuclei in larynx and foreskin reflects the number of injected cells within the specific fields,not the total number of positive cells.
J. VIROL.
0 ''
I.. ..,N
':--:.
40
NOTES 959
TABLE 1. T-antigen expression in cultured cells after microinjection with DNA constructsa
T-antigen expression with constructs:
Cell type No. of pPT-1 pPPl pPT-11R
% Positive Intensityb % Positive Intensity % Positive Intensity
HostLarynx 3 4.7 ± 3.1 NDc 44.0 ± 22.1 30.3 + 3.2 38.7 ± 11.0 25.0 ± 5.0Larynx, 2 0 ND 33.5 ± 12.0 27.0 + 1.4 31.5 ± 13.4 27.0 ± 1.4
papillomaForeskin 3 2.5 ± 2.1 ND 32.3 ± 19.7 23.3 ± 3.1 44.6 ± 28.7 28.0 ± 8.9
NonhostNeck 3 6.3 ± 11.0 ND 25.3 ± 7.5 20.0 ± 2.6 16.3 ± 9.1 9.3 ± 3.8Thigh 2 0.5 ± 0.7 ND 46.0 ± 11.3 15.5 ± 0.7 13.5 ± 2.1 13.0 ± 2.8
Fibroblasts 2 5.0 ± 4.2 ND 58.0 ± 19.8 32.5 ± 10.6 25.0 ± 11.3 17.5 ± 10.6
a Cultured cells were microinjected (3) with two copies per cell of each DNA construct (described in text), incubated for 20 to 22 h, and stained for Tag withthe monoclonal antibody pAB416 (6) and the ABC detection method (Vector Laboratories).
b Stain intensity was measured as percent light absorbed for each positive nucleus with a Kineteck photometer system attached to a Leitz laborlux 11microscope and expressed as the average cell absorption per experiment. Values were normalized to a standard stain using the cell line 14B which contains onecopy of SV40 DNA (16) to correct for small variations in staining from experiment to experiment.
c ND, Not determined.
differences in ability to inject these two categories of cells.The nonhost cells expressed the pPP1 construct containingthe SV40 enhancer nearly as well as did the host cells. Thus,pPP1 served as a control for this type of variation.
Previous studies of HPV URR enhancer function(s) haveshown that viral E2 proteins can regulate, either positivelyor negatively, enhancer sequences in the URR (1, 2, 7). Wedid not provide E2 protein in these experiments. It ispossible, however, that the small reduction in Tag levels inthe laryngeal papilloma cells relative to normal larynx resultsfrom E2 protein produced by viral DNA persisting in thesecells (17).To better determine the relative levels of Tag expression
in the two categories of epithelial cells (host versus nonhost),the number of Tag units was calculated. The percent Tag-positive cells for each of the two categories was determined.This was multiplied by the average of all intensities for thatcategory of cells. This gave a relative value for the totalamount of Tag made by the cells (Fig. 3). pPP1 and pPT-11Rproduced nearly identical amounts of Tag in epithelial cellswhich are the normal hosts for HPV-11. In contrast to this,the Tag level in the nonhost cells was much lower forpPT-11R than for pPP1. There was some reduction (35%) inpPP1 expression in those cells which are nonhosts for
1100 1=J pPp1V) GM pPTD D R_i 1000 pP-1Lu 900o 8000
Z-. 700U,
600zD3 500zLu 400
300z200
Host Non-host
CELL TYPE
FIG. 3. Relative expression of Tag by host and nonhost cells.Composite percentage of Tag-positive cells and absorbance of Tagstain were compared in host (larynx, laryngeal papilloma, andforeskin) and nonhost (thigh and neck skin) cells for the SV40 (pPP1)and HPV-11 (pPT-11R) enhancers.
HPV-11, compared with that in host cells, but the pPT-11RTag level was lower by 83.8%.From these data, we conclude that the URR enhancer
function is responsible, at least in part, for the tissue sitetropism of HPV-11. The URR of the papillomaviruses is verycomplex and contains multiple enhancers. Only a subset, orperhaps only one enhancer sequence, has tissue site speci-ficity. A 269-base-pair fragment from the 5' end of theHPV-11 URR which bound cellular proteins and had en-hancer function (Auborn et al., submitted for publication)did not show any preference for specific types of kerati-nocytes (data not shown). The enhancer(s) in this part of theURR might serve as one source of the constitutive enhanceractivity seen in the nonhost epidermal cells and the fibro-blasts. Further work must be done to learn whichsequence(s) determines epithelial tissue site preference.
We thank Teresa DiLorenzo, Rachel Galli, Ralph Ehrenpreis,Rose Barra, and Iris Kleinman for technical assistance and AllanAbramson for providing many of the tissues.
This work was supported in part by Public Health Service grant 2P01 NS 19214 from the National Institute of Neurological andCommunicative Disorders and Stroke (B.M.S. and L.B.T.) and bythe Long Island Jewish Competitive Pool Award (K.J.A.).
LITERATURE CITED1. Cripe, T. P., T. H. Haugen, J. P. Turk, F. Tabatabai, P. G.
Schmid III, M. Durst, L. Gissmann, A. Roman, and L. P. Turek.1987. Transcriptional regulation of the human papillomavirus-16E6-E7 promoter by a keratinocyte-dependent enhancer, and byviral E2 trans-activator and repressor gene products: implica-tions for cervical carcinogenesis. EMBO J. 6:3745-3753.
2. Gius, D., S. Grossman, M. A. Bedell, and L. A. Laimins. 1988.Inducible and constitutive enhancer domains in the noncodingregion of human papillomavirus type 18. J. Virol. 62:665-672.
3. Graessmann, M., and A. Graessmann. 1983. Microinjection oftissue culture cells. Methods Enzymol. 101:482-492.
4. Green, H., 0. Kehinde, and J. Thomas. 1979. Growth ofcultured human epidermal cells into multiple epithelia suitablefor grafting. Proc. Natl. Acad. Sci. USA 76:5665-5668.
5. Hansen, U., D. G. Tenen, D. M. Livingston, and P. A. Sharp.1981. T antigen repression of SV40 early transcription from twopromoters. Cell 27:603-612.
6. Harlow, E., L. V. Crawford, D. C. Pim, and N. M. Williamson.1981. Monoclonal antibodies specific for the SV40 tumor anti-gens. J. Virol. 39:861-869.
7. Hirochika, H., T. R. Broker, and L. T. Chow. 1987. Enhancers
VOL. 63, 1989

960 NOTES
and trans-acting E2 transcriptional factors of papillomaviruses.J. Virol. 61:2599-2606.
8. Hirochika, H., R. Hirochika, T. R. Broker, and L. T. Chow.1988. Functional mapping of the human papillomavirus type 11transcriptional enhancer and its interaction with the trans-actingE2 proteins. Genes Dev. 2:54-67.
9. Kreider, J. W., M. K. Howett, M. H. Stoler, R. J. Zaino, and P.Welsh. 1987. Susceptibility of various human tissues to trans-formation in vivo with human papillomavirus type 11. Int. J.Cancer 39:459-465.
10. Maniatis, T., S. Goodbourn, and J. A. Fischer. 1987. Regulationof inducible and tissue-specific gene expression. Science 236:1237-1244.
11. McKnight, S., and R. Tjian. 1986. Transcriptional selectivity ofviral genes in mammalian cells. Cell 46:795-805.
12. Pfister, H. 1984. Biology and biochemistry of papillomaviruses.Rev. Physiol. Biochem. Pharmacol. 99:111-181.
13. Pfister, H., J. Kribke, W. Dietrich, T. Ifner, and P. G. Fuchs.1986. Classification of the papillomaviruses-mapping the ge-nome. CIBA Found. Symp. 120:3-22.
14. Rio, D. C., A. Robbins, R. Meyers, and R. Tjian. 1980. Regula-tion of simian virus 40 early transcription in vitro by purified
tumor antigen. Proc. Natl. Acad. Sci. USA 77:5706-5710.15. Steinberg, B. M., A. L. Abramson, and R. P. Meade. 1982.
Culture of human laryngeal papilloma cells in vitro. Otolaryn-gol. Head Neck Surg. 90:728-735.
16. Steinberg, B., R. Pollack, W. Topp, and M. Botchan. 1978.Isolation and characterization of T-antigen-negative revertantsfrom a line of transformed rat cells containing one copy of theSV40 genome. Cell 13:19-32.
17. Steinberg, B. M., W. C. Topp, P. S. Schneider, and A. L.Abramson. 1983. Laryngeal papillomavirus infection duringclinical remission. N. Engl. J. Med. 308:1261-1264.
18. Thierry, F., J. M. Heard, K. Dartmann, and M. Yaniv. 1987.Characterization of a transcriptional promoter of human papil-lomavirus and modulation of its expression by simian virus 40and adenovirus early antigens. J. Virol. 61:134-142.
19. Tsao, M. C., B. J. Walthall, and R. G. Ham. 1982. Clonal growthof normal epidermal keratinocytes in a defined medium. J. CellPhysiol. 110:219-229.
20. Zur Hausen, H., L. Gissmann, and J. R. Schlehofer. 1985.Viruses in the etiology of human genital cancer. Prog. Med.Virol. 30:170-188.
J. VIROL.