translation initiation at an acg triplet in mammalian cells* · translation initiation at an acg...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1987 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 262, No. 24, Issue of August 25, pp. 11847-11851, 1987 Printed in U. S. A.
Translation Initiation at an ACG Triplet in Mammalian Cells* (Received for publication, December 19, 1986)
David S. Peabody From the Department of Cell Biology and the Cancer Center, University of New Mexico School of Medicine, Albuquerque, New Mexico 87131
The initiator AUG of the mouse dihydrofolate reduc- tase gene (dhfr) was converted to ACG by site-directed mutagenesis and assayed for expression in cultured monkey cells using an SV40 recombinant called SVGT5dhfr26m2. Synthesis of apparently full-length dihydrofolate reductase (DHFR) protein was signifi- cantly reduced compared to wild-type, but not entirely abolished, suggesting that the ACG triplet was being utilized for translation initiation. In addition, a trun- cated form of DHFR was produced, apparently by ini- tiation at the next in-frame AUG downstream. This result was confirmed in vitro. Transcripts of the dhfr sequence were produced by SP6 RNA polymerase in the presence of m’GpppG and translated in vitro using reticulocyte lysates and wheat germ extracts. The re- sults paralleled those observed in vivo. Synthesis of full-length DHFR was reduced, but not eliminated, and a new species was produced by initiation at an internal site. Amino acid sequence analysis of the products of in vitro translation demonstrated that translation does indeed initiate at the ACG triplet and that it initiates with methionine. Additional mutations were intro- duced which altered the sequence context of the ACG triplet. Mutation of the translation initiation consensus sequence by substitution of the A residue at position -3, or of the G at +4 resulted in a significant decrease in initiation at the ACG and an increase in the level of the internal initiation product. Thus, translation ini- tiation at a non-AUG triplet depends on a favorable sequence context.
The signals in mammalian mRNAs that specify translation initiation sites are still being defined. It is becoming clear, however, that sequences in the vicinity of an AUG triplet are among the factors that influence its ability to serve as a functional initiation codon (1-4). Other things being equal, an AUG serves most efficiently as an initiation codon when it resides within the consensus sequence (A/G)CCAUGG. The relative importance of nucleotides at each of the specific postions in this sequence is reflected in the degree of their conservation. Thus, the -3 position (if the A of the AUG is defined as +1) contains an A residue about 80% of the time and a G residue in most of the remaining cases. Substitution with a pyrimidine at this position causes a sharp reduction in translation initiation at this site. The G at +4 and the C residues at -1 and -2 apparently play a less important role since substitutions here lead to less dramatic reductions in translation efficiency. The translation initiation region of the
*This work was supported by Grant DMB-8510683 from the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
mouse dhfr’ gene contains the sequence AUCAUGG, a near perfect fit to the consensus sequence. Using site-directed mutagenesis techniques we have introduced a number of single nucleotide substititions into this sequence. In a control experiment a mutation was constructed that converted the initiator AUG to a ACG triplet. It was anticipated that this alteration would result in the complete elimination of trans- lation initiation at this site. The mutant dhfr sequence was inserted into the late region of two different SV40 vectors for expression in cultured monkey cells. Recombinant viruses which contain the wild-type dhfr sequence direct the synthesis of large quantities of the mouse DHFR protein. Surprisingly, cells infected with recombinants containing the mutant se- quence still produced a low, but easily detected level of ap- parently authentic DHFR, presumably by translation initia- tion at the ACG triplet. Until recently it was held that AUG was the only initiator triplet in eucaryotic mRNAs (5). How- ever, examples of translation initiation at non-AUG triplets in yeats (6, 7) and at ACG in adeno-associated virus (8) of mammalian cells have now been reported. Moreover, ACG has been shown to be a functional initiation codon in cell- free translation systems from wheat germ and rabbit reticu- locytes programmed with a mutant form of a procaryotic mRNA from the bacteriophage T7 0.3 gene (9). The experi- ments reported here confirm and extend these observations by demonstrating that translation initiates at an ACG triplet of the mutated form of dhfr both in vivo and i n vitro, that the efficiency of initiation at this non-AUG triplet is a function of the surrounding nucleotide sequence, and that translation of the mutant sequence initiates with methionine.
MATERIALS AND METHODS
Construction of Viral Recombinants and Analysis of Their Prod- ucts-The mutations were produced by the method of Zoller and Smith (10) using synthetic oligodeoxyribonucleotides. produced on the Applied Biosystems model 380A automated DNA synthesizer to mutagenize a 200-base pair fragment of dhfr cloned in M13mp8. The mutants (see Fig. 1) were identified by hybridization to the mutagenic oligonucleotide and their structures were confirmed by DNA sequence analysis (11, 12). After introduction of the nucleotide substitutions, the complete dhfr sequence was reconstructed and inserted into a lytic viral vector called SVGT5. The construction and properties of the SVGT5dhfr26 recombinant have been described elsewhere (13). A derivative containing the ACG mutation was constructed by similar methods and was called SVGT5dhfr26m2. Recombinant viral stocks containing the SV40 early mutant tsA58 as helper, were used to infect CVl cells at multiplicities of 1-10 plaque-forming units/cell. Protein was extracted about 48 h postinfection when approximately 70% of cells exhibited cytopathic effect. They were analyzed for the presence of mouse DHFR by the protein blotting method of Burnette (14) using antibody to DHFR and ‘251-labeled protein A.
Transcription and Translation in Vitro-The wild-type and mutant
The abbreviations used are: dhfr, mouse dihydrofolate reductase cDNA sequence; DHFR, dihydrofolate reductase; SDS, sodium do- decyl sulfate; HPLC, high performance liquid chromatography; PTH- Met, phenylthiohydantoin derivative of methionine.
11847
11848 Initiation at an ACG Triplet in Mammalian Cells forms of the dhjr sequence were placed under control of the bacteri- ophage SP6 promoter by inserting them as HindIII-BglII fragments between the Hind111 and BamHI sites of pSP64 (15) to produce the plasmids known as pSPD26, pSPD26m2, pSPD26m3, pSPD26m15, and pSPD26m16. After purification the plasmids were linearized by digestion at the unique EcoRI site within the polylinker of pSP64 and near the 3' end of the dhfr sequence. Synthesis and capping of RNAs was accomplished by transcription in the presence of m7GpppG as described by Pelletier and Sonenberg (16). Transcription was monitored by incorporation of [32P]CTP into trichloroacetic acid- precipitable material and transcripts were shown to be full-length by electrophoresis in denaturing agarose gels (17). Translation was per- formed by adding about 0.1-0.2 pg of RNA to reticulocyte lysates (purchased from Bethesda Research Laboratories) or to wheat germ extracts prepared as described by Anderson et al. (18). The ["SI methionine-labeled products were analyzed by electrophoresis in pol- yacrylamide gels containing SDS (19) followed by autoradiography (20).
Amino Acid Sequence Analysis-Identities of the products of trans- lation in reticulocyte lysates were confirmed by sequence analysis of the polypeptides labeled with [35S]methionine. Acetylation of the amino terminus was prevented by performing the translation reac- tions in the presence of oxalacetic acid and citrate synthase as described by Palmiter (21). After separation by electrophoresis in polyacrylamide gels containing SDS (19), the radiolabeled proteins were electrophoretically transferred to glass fiber filters by the method of Aebersold et al. (22) and subjected to amino acid sequence analysis on the Applied Biosystems model 470A protein sequenator. Half of the material collected after each cycle was counted in a liquid scintillation counter. The labeled material released in a given cycle was analyzed by HPLC for coelution with a PTH-methionine stan- dard.
RESULTS
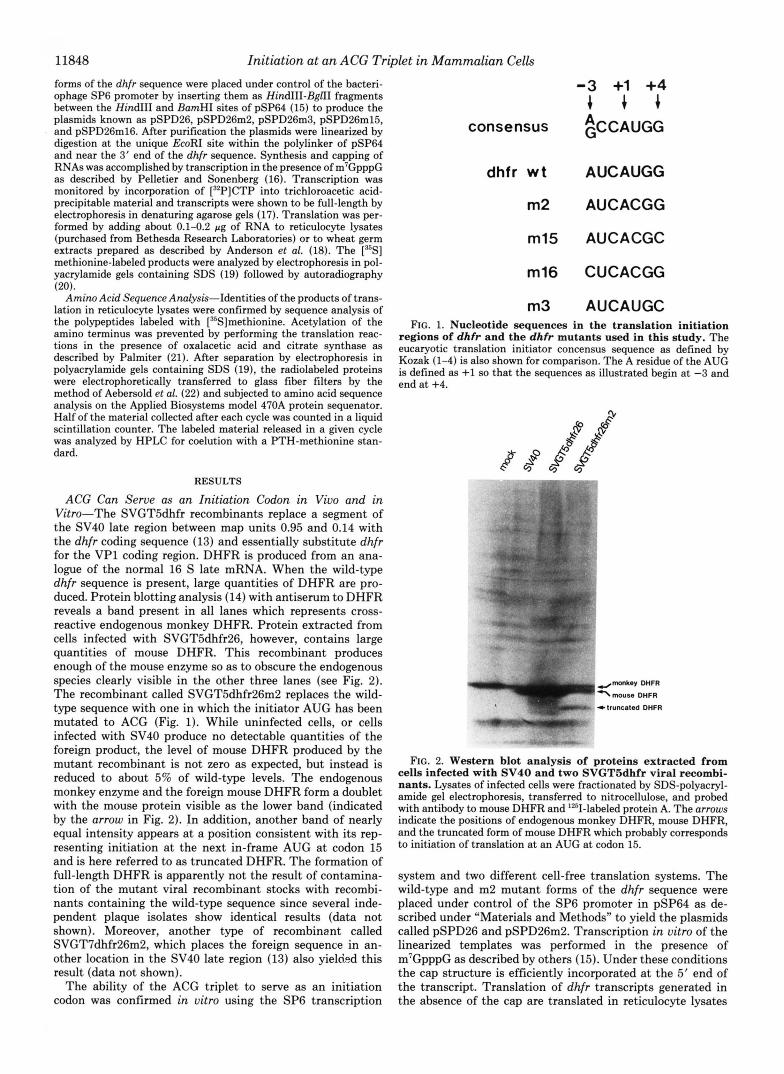
ACG Can Serve as an Initiation Codon in Vivo and in Vitro-The SVGT5dhfr recombinants replace a segment of the SV40 late region between map units 0.95 and 0.14 with the dhfr coding sequence (13) and essentially substitute dhfr for the VP1 coding region. DHFR is produced from an ana- logue of the normal 16 S late mRNA. When the wild-type dhfr sequence is present, large quantities of DHFR are pro- duced. Protein blotting analysis (14) with antiserum to DHFR reveals a band present in all lanes which represents cross- reactive endogenous monkey DHFR. Protein extracted from cells infected with SVGT5dhfr26, however, contains large quantities of mouse DHFR. This recombinant produces enough of the mouse enzyme so as to obscure the endogenous species clearly visible in the other three lanes (see Fig. 2). The recombinant called SVGTSdhfr26m2 replaces the wild- type sequence with one in which the initiator AUG has been mutated to ACG (Fig. 1). While uninfected cells, or cells infected with SV40 produce no detectable quantities of the foreign product, the level of mouse DHFR produced by the mutant recombinant is not zero as expected, but instead is reduced to about 5% of wild-type levels. The endogenous monkey enzyme and the foreign mouse DHFR form a doublet with the mouse protein visible as the lower band (indicated by the arrow in Fig. 2). In addition, another band of nearly equal intensity appears at a position consistent with its rep- resenting initiation at the next in-frame AUG at codon 15 and is here referred to as truncated DHFR. The formation of full-length DHFR is apparently not the result of contamina- tion of the mutant viral recombinant stocks with recombi- nants containing the wild-type sequence since several inde- pendent plaque isolates show identical results (data not shown). Moreover, another type of recombinent called SVGT7dhfr26m2, which places the foreign sequence in an- other location in the SV40 late region (13) also yielded this result (data not shown).
The ability of the ACG triplet to serve as an initiation codon was confirmed in vitro using the SP6 transcription
consensus
dhfr w t
m2
m15
m16
m3
-3 +1 +4 + + + ~CCAUGG
AUCAUGG
AUCACGG
AUCACGC
CUCACGG
AUCAUGC FIG. 1. Nucleotide sequences in the translation initiation
regions of dhfr and the dhfr mutants used in this study. The eucaryotic translation initiator concensus sequence as defined by Kozak (1-4) is also shown for comparison. The A residue of the AUG is defined as +1 so that the sequences as illustrated begin at -3 and end at +4.
FIG. 2. Western blot analysis of proteins extracted from cells infected with SV40 and two SVGT5dhfr viral recombi- nants. Lysates of infected cells were fractionated by SDS-polyacryl- amide gel electrophoresis, transferred to nitrocellulose, and probed with antibody to mouse DHFR and '251-labeled protein A. The arrows indicate the positions of endogenous monkey DHFR, mouse DHFR, and the truncated form of mouse DHFR which probably corresponds to initiation of translation at an AUG at codon 15.
system and two different cell-free translation systems. The wild-type and m2 mutant forms of the dhfr sequence were placed under control of the SP6 promoter in pSP64 as de- scribed under "Materials and Methods" to yield the plasmids called pSPD26 and pSPD26m2. Transcription in vitro of the linearized templates was performed in the presence of m7GpppG as described by others (15). Under these conditions the cap structure is efficiently incorporated at the 5' end of the transcript. Translation of dhfr transcripts generated in the absence of the cap are translated in reticulocyte lysates

Initiation at an ACG Triplet in Mammalian Cells 11849
at least 50-fold less efficiently than those produced in reac- tions that contain the cap (data not shown).
The results of translation of the wild-type and m2 mutant transcripts in the reticulocyte lysate system are shown in Fig. 3A. The unmodified dhfr sequence directs the synthesis of full-length DHFR but no substantial quantity of any other species. The m2 mutant yields a different result. The amount of full-length DHFR now represents about 80% of the total translation products and an additional band is seen whose mobility indicates it is probably the result of translation initiation at the next AUG in the sequence at codon 15. The mobility of this species is identical with that of the shortened form of DHFR seen in vivo (Fig. 2).
The results of translation in the wheat germ extract are shown in Fig. 3B. Translation of the m2 mutant RNA results in reduced, but still substantial levels of full-length DHFR and larger amounts of product apparently corresponding to initiation at the next AUG at codon 15. Densitometer scans indicate that about 35% of the product is full-length. In addition, small amounts of two other species can also be observed (especially in the wild-type lane). While these could be the result of degradation of the full-length protein, their molecular sizes are consistent with the idea that they repre- sent translation initiation at AUG triplets at codons 38 and 53. Interestingly, the first four AUGs of the dhfr coding region are in the same reading frame and each lies within a sequence that is a good fit to the initiator consensus sequence defined by Kozak (Fig. 1 and Refs. 1-4).
The Sequence Context of the ACG Influences Its Ability to Serve as an Initiation Codon-Since ACG does not usually function as an initiation codon, its ability to do so might be a function of surrounding nucleotide sequence in the same way that the efficiency of ACG initiators depends on sequence context. Kozak has shown that translation initiation is opti- mal when the initiator AUG is present in the sequence (A/ G)CCAUGG (1-4). The dhfr initiation site, AUCAUGG, is a very close fit to this consensus sequence. To test the impor- tance of the consensus sequence for initiation at ACG, two additional mutants (m15 and m16) were constructed, cloned in pSP64, and analyzed by in vitro transcription and trans- lation as described above and under “Materials and Methods.” The sequence alterations in the initiation region of each mutant are shown in Fig. 1 and the results of translation in reticulocyte lysates are shown in Fig. 4. The mutation called m15 is substituted at position +4 and changes the sequence
A B -RNA wt m2 -RNP, wt m2
FIG. 3. Translation in vitro of dhfr and dhfrm2-specific RNAs generated by transcription with SP6 RNA polymerase (described under YMaterials and Methods”). The products of translation in reticulocyte lysates are shown in A while those synthe- sized in wheat germ extracts are shown in B.
wt m2 1n15 m16
FIG. 4. Translation in reticulocyte lysates of the wild-type and the three mutant forms of dhfr RNA generated in vitro using SP6 RNA polymerase. The position of full-length DHFR is indicated by the arrow.
of the initiation region of m2 from AUCACGG to AUCACGC. When m15 RNA is translated in vitro about 50% of the total product is full-length DHFR, a modest reduction compared to wild-type. The mutation called m16 changes the most conserved element of the consensus sequence (apart from the AUG itself) by changing the A residue at position -3 to a C to give the sequence CUCACGG. This results in a substantial reduction in the level of full-length DHFR and an elevation in the level of initiation at the internal site. In fact, the full- length protein now represents only about 5% of the protein synthesized. In addition, small amounts of a third product are observed. The identity of this species is unclear, but may represent initiation at a GUG triplet at codon 11. Preliminary experiments indicate that GUG can also serve as an initiation codon when present in a favorable context, as it is in this case (data not shown). Amino acid sequence analysis will be nec- essary to fully define the nature of these additional products.
Amino Acid Sequence Analysis of DHFR Produced from AUG and ACG Containing RNAs-The [35S]methionine-la- beled products of translation in reticulocyte lysates were subjected to amino acid sequence analysis in order to deter- mine whether methionine was the initiating amino acid, and whether translation was truly initiating at ACG and not at some other nearby non-AUG triplet. Normally the amino- terminal methionine of DHFR is cleaved from the molecule so that valine (codon 2) is present at the mature amino terminus (23). Since the methionine aminopeptidase activity responsible for the cleavage is usually present in reticulocyte lysates, sequence analyses were performed on the products of translation of two additional mutant RNAs. One of them, m3, retains the normal AUG initiation codon, but by changing the G residue normally present at +4 to C, introduces a substi- tution of isoleucine for valine at position 2 of the amino acid sequence. The other mutant, m15, was described in the pre- vious section. To reiterate, m15 is a double mutant which converts the initiator AUG to ACG, but, like m3, also changes the nucleotide at +4 in the RNA to C. According to the rules developed by Sherman and Stewart (24), proteins with isoleu- cine at position 2 are not generally susceptible to the action of methionine aminopeptidase. Thus, the amino-terminal me- thionine should be retained permitting its identification by sequence analysis. As a preliminary indication that this is the

11850 Initiation at an ACG Triplet in Mammalian Cells
B
PTH-MET
i residue 1
10 20 Iractuon no. n
0 1 5 10 15 20
CYCLE NO.
PTH-MET
1 residue 1 t
10 20 lractton no.
0 pmnm4hm 5 1 0 15 2 0
CYCLE NO. FIG. 5. Amino acid sequence analysis of DHFR m3 (panel
A ) and m15 (panel B ) . Protein labeled in uitro with [35S]methio- nine (about 22,000 and 17,000 cpm, respectively) was subjected to automated sequential Edman degradations in an Applied Biosystems model 470A protein sequenator. One-half of the material from each cycle was counted in a liquid scintillation counter and the remaining half of material from cycles which contained radioactivity was ana- lyzed by HPLC against a PTH-methionine standard. The results of HPLC analysis of the material released in cycle 1 are shown in the insets.
case, it was observed that the m3 and m15 DHFR proteins had electrophoretic mobilities in SDS-polyacrylamide gels which were slightly reduced compared with the wild-type and m2 mutant proteins (data not shown).
The full-length products of translation of the m3 and m15 RNAs in reticulocyte lysates containing [35S]methionine were subjected to automated sequential Edman degradation as described under “Materials and Methods.” The results are shown in Fig. 5. It is clear that the amino-terminal residue is methionine in both cases. This is indicated not only by the release of radiolabeled material in cycle 1, but also by the fact that this material coelutes with a PTH-methionine standard when analyzed by HPLC (see inset to Fig. 5). Moreover, [35S] methionine is recovered in the first cycle in yields of about 70% of the theoretical maximum considering that the DHFR molecule contains seven methionines. The near identity of the results for molecules initiating at AUG and ACG indicates that methionine is apparently the only amino acid present in any significant quantity at the amino terminus when ACG is the initiator triplet. It is also apparent that translation initi- ation occurs at the correct sites since the next release of [35S] methionine is at residue 15 in both cases as predicted from the known sequence. The material released at cycle 15 was also shown to coelute with PTH-methionine (data not shown). Thus, in the case of the m15 mutant translation ix:itiates at ACG and it initiates with methionine.
DISCUSSION
Work from several laboratories has begun to define the nature of eucaryotic translation initiation sites. Much of the
conceptual basis for this work has been derived from Kozak’s (25) scanning hypothesis which in its original form suggested that a translation initiation site was defined entirely as the AUG nearest the 5‘ end of the mRNA. However, as the amount of sequence information for a large number of mRNAs increased, exceptions to the first AUG rule made it clear that this was not a complete description of translation initiation signals. Evidence has accumulated that clearly demonstrates the importance of the nucleotide sequence immediately sur- rounding an AUG in determining its ability to function as an initiation codon (1-4). While it is now recognized that an AUG triplet is not by itself sufficient to specify an efficient initiation site, it has been widely believed that an AUG triplet was at least a necessary component of the initiation site. This view was supported by an analysis of a large number of mutations in the CYCl gene of yeast which was interpreted to show that AUG was the only initiation codon in eucaryotes (5). This situation was contrasted with procaryotes where GUG and UUG are known to serve as functional initiation codons, albeit at lower efficiencies than AUG (26). However, the information presented in this paper and experiments recently reported by others now indicate that necleotide tri- plets other than AUG are competent to direct translation initiation under certain circumstances. Thus, in yeast, AUA, UUG, and several other non-AUG triplets have been shown to be capable of inefficiently initiating translation of mutant forms of CYCl mRNA (6,7). Adeno-associated virus of mam- malian cells initiates translation of one of its coat proteins at a naturally occurring ACG triplet (8). Moreover, a mutant of the bacteriophage T7 0.3 gene, in which the initiator AUG has been altered to ACG has been shown to be functional in cell-free translation systems from reticulocytes and wheat germ (9).
The results presented here offer another example of initi- ation at a non-AUG triplet when the initiator AUG of mouse dhfr is converted to ACG by site-directed mutagenesis. More- over, just as the ability of AUG triplets to serve as initiation codons depends on surrounding sequences (1-4), the effi- ciency with which the ACG is utilized is also modulated by sequence context. The effects of altering the sequence context of the ACG roughly parallel those recently reported for an AUG initiator by Kozak where substitution at the +4 position had a relatively small, but significant effect on translation efficiency and substitutions at -3 were major determinants of initiator strength (4). In the present study initiation at the ACG triplet was substantially reduced when the A residue at -3 was converted to C, while the substitution at +4 was less effective in reducing initiation efficiency. The importance of the upstream A residue for initiation at another non-AUG triplet was also observed in the yeast cyc-1 system where a double mutant which changed the conserved A residue at -3 to U reduced the efficiency of initiation at UUG an additional 10-fold (6). In the two other known examples of initiation at ACG, the initiator triplet is found within a favorable sequence environment according to the rules of Kozak (1-4). In the cases of both the adeno-associated virus and the mutant T7 0.3 gene (9) the initiation site has the sequence AAGACGG.
It is unclear why the efficiency of utilization of the ACG triplet for translation initiation should be higher in vitro than it is in vivo. Several unpublished observations,2 however, are relevant to this question. When dhfr mutants altered in the sequence context of the normal initiator AUG are translated in reticulocyte lysates or in wheat germ extracts, no obvious difference in translation efficiency is observed relative to wild- type. For example, when a pyrimidine residue is substituted
D. S. Peabody, unpublished observations.

Initiation at an ACG Triplet in Mammalian Cells 11851
for the A normally present at position -4, as much DHFR is produced as with the normal sequence.* Moreover, no evidence of internal initiation events is seen. These cell-free systems, therefore, seem to be less sensitive to subtle alterations of mRNA structure than is the translation machinery within the living cell. In addition, the conditions chosen for translation in vitro can affect the fidelity of the initiation codon recog- nition process. For instance, the efficienciy of utilization of the ACG triplet as compared to initiation at the internal site on the dhfr m2 mutant RNA is substantially affected by changing the concentration of magnesium ions in wheat germ extracts. More efficient initiation at ACG occurs at a higher concentration of magnesium than that used in this study and which was adjusted for optimal incorporation of the radioac- tive amino acid (not shown).
Of the hundreds of naturally occurring eucaryotic mRNAs whose sequences are now known, only one so far has been reported to initiate at a non-AUG triplet. Non-AUG initiation codons are relatively rare even in bacteria, however, where their existence has been recognized for some time. A recent study used site-directed mutagenesis to show that GUG and UUG are less efficient than AUG in directing translation initiation in Escherichia coli (26). In the few cases examined so far, non-AUG triplets are much less efficient initiators in eucaryotes as well. This may by itself explain their rarity. Translation efficiency may usually be modulated by more subtle alterations of sequence or secondary structure sur- rounding an AUG. However, the utilization of a non-AUG initiator could provide an additional means of reducing the levels of a protein product that may be required only in small amounts. Moreover, as the results of these experiments indi- cate, it can also provide the means of producing more than one protein from a single mRNA by permitting the utilization of internal initiation sites.
These observations raise the possibility that other non- AUG triplets may be recognized as translation initiation sites when surrounded by the appropriate nucleotide sequences. Indeed, a preliminary analsyis of all possible single nucleotide substitutions in the dhfr AUG indicates that most of them are utilized to some extent by the translation initiation ap- paratus. Further experiments are required to define the range of non-AUG triplets that can function as initiation codons and to fully characterize the influence of nearby sequences.
Acknowledgments-Gratitude is expressed to Latif Kazim for per- forming the amino acid sequence analysis, to Gavin Pickett for technical assistance, and to John Atkins for criticisms.
REFERENCES
1. Kozak, M. (1981) Nucleic Acids Res. 9,5233-5252 2. Kozak, M. (1984) Nucleic Acids Res. 12, 857-872 3. Kozak, M. (1984) Nature 308,241-246 4. Kozak, M. (1986) Cell 44,283-292 5. Sherman, F., McKnight, G. & Stewart, J. W. (1980) Biochim.
6. Zitomer, R. S., Walthall, D. A., Raymond, B. C. & Hollenberg, C. P. (1984) Mol. Cell. Biol. 4 , 1191-1197
7. Baim, S. B., Goodhue, C. T., Pietras, D. F., Eustice, D. C., Labhard, M., Friedman, L. R., Hampsey, D. M., Stiles, J. I. & Sherman, F. (1985) in Sequence Specificity in Transcription and Translation (Calendar, R., and Gold, L., eds) pp. 351-362, Allen R. Liss Inc., New York
8. Becerra, S. P., Rose, J. A., Hardy, M., Baroudy, B. M. & Ander- son, C. W. (1985) Proc. Natl. Acud. Sci. U. S. A. 82,7919-7923
9. Anderson, C. W. & Buzash-Pollert, E. (1985) Mol. Cell. Biol. 5 ,
10. Zoller, M. J. & Smith, M. (1983) Methods Enzymol. 100, 468-
11. Sanger, F., Coulson, A. R., Barrell, B. G., Smith, A. J. H. & Roe,
12. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl. Acud.
13. Subramani, S., Mulligan, R. C. & Berg, P. (1981) Mol. Cell. Biol.
14. Burnette, W. N. (1981) Anal. Biochem. 112,195-203 15. Melton, D. A., Krieg, P. & Green, M. R. (1984) Nucleic Acids Res.
16. Pelletier, J. & Sonenberg, N. (1985) Cell 40, 515-526 17. Bailey, J. M. & Davidson, N. (1976) Anal. Biochem. 7 0 , 75-85 18. Anderson, C. W., Stews, J. W. & Dudock, B. S. (1983) Methods
19. Laemmli, U. K. (1970) Nature 227, 680-685 20. Chamberlain, J. P. (1979) Anal. Biochem. 98, 132-135 21. Palmiter, R. D. (1977) J. Biol. Chem. 252,8781-8783 22. Aebersold, R. H., Teplow, D. B., Hood, L. E. & Kent, S. B. H.
23. Stone, D. & Phillips, A. W. (1977) FEBS Lett. 74 , 85-87 24. Sherman, F. & Stewart, J. W. (1982) in The Molecular Biology of
the Yeast Saccharomyces (Jones, E. W., and Broach, J. R., eds) pp. 301-333, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Biophys. Acta 609,343-346
3621-3624
500
B. A. (1981) J. Mol. Biol. 143 , 161-178
Sci. U. S. A. 74,5463-5467
1,854-864
12,7035-7056
Enzymol. 101,635-644
(1986) J. Biol. Chem. 261 , 4229-4238
25. Kozak, M. (1978) Cell 15 , 1109-1123 26. Reddy, P., Peterkofsky, A. & McKenney, K. (1985) Proc. Natl.
Acud. Sci. U. S. A. 82,5656-5660