tyrosine residues at the immunoglobulin-c-type lectin inter-domain boundary of intimin are not...
TRANSCRIPT

Original article
Tyrosine residues at the immunoglobulin-C-type lectin inter-domainboundary of intimin are not involved in Tir-binding but implicated
in colonisation of the host
Stephen Reece a, Cameron P. Simmons a, Robert J. Fitzhenry b, Marjan Ghaem-Maghami a,Rosanna Mundy a, Christine Hale a, Stephen Matthews c, Gordon Dougan a,
Alan D. Phillips b, Gad Frankel a,*a Centre for Molecular Microbiology and Infection, Department of Biological Sciences, Flowers Building,
Imperial College of Science, Technology and Medicine, London SW7 2AZ, UKb Centre for Paediatric Gastroenterology, Royal Free Hospital, London NW3 2QG, UK
c Centre for Structural Biology, Department of Biological Sciences, Wolfson Building, Imperial College of Science,Technology and Medicine, London SW7 2AZ, UK
Received 3 May 2002; accepted 10 September 2002
Abstract
Intimin is an outer membrane adhesion molecule involved in bacterial adhesion to intestinal epithelium by several human and animalenteric pathogens, including enteropathogenic and enterohaemorrhagic Escherichia coli and Citrobacter rodentium. Intimin binds to thetranslocated intimin receptor, Tir, which is delivered to the plasma membrane of the host cell by a type III protein translocation system. Intiminis also implicated in binding to a host cell-encoded intimin receptor (Hir). The receptor-binding activity of intimin resides within the carboxyterminus 280 amino acids (Int280) of the polypeptide. Structural analysis of this region revealed two immunoglobulin-like domains, thesecond of which forms a number of contacts with the distal C-type lectin-like module. Specific orientation differences at this inter-domainboundary, which consists of several tyrosine residues, were detected between the crystal and solution structures. In this study, we determinedthe influence of site-directed mutagenesis of each of four tyrosine residues on intimin–Tir interactions and on intimin-mediated intimateattachment. The mutant intimins were also studied using a variety of in vitro and in vivo infection models. The results show that three of thefour Tyr, although not essential for A/E lesion formation in vitro, are required for efficient colonisation of the mouse host following oralchallenge.
© 2002 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords: Citrobacter rodentium; EPEC; Intimin; Tir
1. Introduction
Intimin is a bacterial adhesion molecule involved in inti-mate attachment of enteropathogenic and enterohaemor-rhagic Escherichia coli (EPEC and EHEC) to mammalianhost cells [1,2]. Intimin targets the translocated intimin re-ceptor (Tir), which is translocated to the plasma membrane ofthe eukaryotic cell by the bacterial type III secretion system[3,4]. In an earlier study, we determined the global fold of the
receptor-binding C-terminal 280 amino acids of intimin(Int280 comprising residues 660–939 in full-length intimin)by a combination of perdueteration, site-specific protonationand multidimensional nuclear magnetic resonance (NMR)[5,6]. More recently, the X-ray crystal structure was pre-sented for this region [7] as well as a higher resolutionsolution structure for the C-terminal 190 residue portion ofintimin (Int190) [8]. The C-terminal 190 amino acids ofintimin (residues 749–939 in full-length intimin) comprisean N-terminal immunoglobulin domain that is coupled to anovel C-type lectin domain (CTLD), which extends an ar-ticulated rod from the bacterial surface and conveys a highlyaccessible ‘adhesive tip’ to the target cell. Int190 contains the
* Corresponding author. Tel.: +44-20-7594-5253;fax: +44-20-7594-3069.
E-mail address: [email protected] (G. Frankel)
Microbes and Infection 4 (2002) 1389–1399
www.elsevier.com/locate/micinf
© 2002 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.PII: S 1 2 8 6 - 4 5 7 9 ( 0 2 ) 0 0 0 2 1 - 7

Tir-binding surface, which resides in the second b-sheet ofthe lectin domain as identified by NMR and X-ray crystal-lography [7,8]. The solution and crystal structures of thisregion are largely identical but some subtle differences arepresent in the relative orientation of the domains and someloop regions [7,8]. It is also noteworthy to point out thatintimin was crystallised in very high salt conditions whichcan induce conformational changes in labile regions, i.e.loops and domain interfaces. Specific differences includeseveral residues near the inter-domain boundary, which showthe highest orientation difference between crystal and solu-tion structures (region Ser870–Ser892). Interestingly, thisregion includes an intriguing group of four tyrosine residues,which are partially exposed and form contacts at the domaininterface [7,8]. The region was also intriguing, as it forms theperiphery of a solvent-exposed cleft in the structure that liesat the domain boundary. Since aromatic residues are oftenimportant in molecular recognition, this region could beenvisaged to be involved in binding a potential ligand. Fur-thermore, this region is more exposed in the solution struc-ture compared to the crystal structure. It was therefore de-cided to mutate these tyrosine residues individually in orderto ascertain whether or not they were involved in an aspect ofintimin function.
2. Materials and methods
2.1. Bacterial strains and plasmids
Bacterial strains used in this study include wild-typeEPEC strain E2348/69 (O127:H6) [9], eae deletion mutantsof E2348/69, strain CVD206 [10], and Citrobacter roden-tium, strain DBS255 [11] and E. coli strains XL1-Blue andBL21. Bacterial strains were grown in L-broth. Media weresupplemented with 50 µg/ml kanamycin, 30 µg/ml chloram-phenicol or 100 µg/ml ampicillin. The plasmids are listed inTable 1.
2.2. Site-directed mutagenesis
Site-directed mutagenesis was performed using theQuickChange Site-directed mutagenesis kit (Stratagene) fol-lowing the manufacturer’s instructions, employing plasmidpCVD438-encoding intimin a [10] as a template. Comple-mentary mutagenesis oligonucleotide pairs incorporatingsingle amino acid substitutions were as follows:
Sense oligonucleotidesY779/120A – CCACTGTATGGTTGCAAGCTGGTCA-
AGTTAATCTGY887/228A – GGGGCTGCAAATAAAGCTGAATAT-
TATAAGY889/230A – GCAAATAAATATGAAGCTTATAAGTC-
TAGTCY890/231A – GCAAATAAATATGAATATGCTAAGTC-
TAGTCAGAC
Antisense oligonucleotidesY779/120A – CAGATTAACTTGACCAGCTTGCAAC-
CATACAGTGGY/887/228A – GGGGCTGCAAATAAAGCTGAATAT-
TATAAGY889/230A – GACTAGACTTATAAGCTTCATATTTA-
TTTGCY890/231A – GTCTGACTAGACTTAGCATATTCATA-
TTTATTTGC
A deletion mutation consisting of the tyrosine rich region(887YXYY890) within the CTLD of intimin a was con-structed in a similar way using oligonucleotides:
Sense loop – ATGAGCTCTCAGACTATAATTTCAT-GGGTAC
Antisense loop – ATGAGCTCTTATTTGCAGCCCC-CATGCTTTAAAGAC
Mutated plasmid containing staggered nicks was gener-ated by extension of primers annealed to opposite strands ofthe denatured plasmid by temperature cycling (one cycle of95 °C, 30 s, then 16 cycles of 95 °C, 30 s, 55 °C, 1 min, 68 °C,18 min) in the presence of the high-fidelity Pfu DNA poly-merase. Synthesised DNA containing the desired mutationwas selected from the original DNA template by incubationwith Dpn1 at 37 °C for 1 h, on the basis that dam methylatedparental DNA template would be susceptible to digestionwhereas the newly synthesised unmethylated mutated plas-mid would not. Nicks in the plasmid were repaired followingtransformation of 1 µl of the synthesised products into com-petent E. coli XL1-Blue cells. Chloramphenicol-resistanttransformants were randomly selected and inoculated intoovernight L-broths for preparation of plasmid mini-preps
Table 1List of plasmids used
Plasmid Description Reference
pCVD438 pACYC184 encoding intimin a [10]pICC217 pCVD438 encoding intimin aY779/120A TSpICC218 pCVD438 encoding intimin aY887/228A TSpICC219 pCVD438 encoding intimin aY889/230A TSpICC220 pCVD438 encoding intimin aY890/231A TSpICC221 pCVD438 encoding intimin aDY887–Y890A TSpICC18 pET28a encoding Tir-M [3]pICC22 pMal-c2 encoding MBP–Int280a [12]pICC222 pMal-c2 encoding MBP–Int280aY779/120A TSpICC223 pMal-c2 encoding MBP–Int280aY887/228A TSpICC224 pMal-c2 encoding MBP–Int280aY889/230A TSpICC225 pMal-c2 encoding MBP–Int280aY890/231A TSpICC226 pMal-c2 encoding MBP–Int280adY228–Y231 TSpGBT9 A yeast GAL4 DNA-BD ClontechpICC10 PGBT9 expressing Tir [3]pGAD424 A yeast GAL4 DNA-AD ClontechpICC19 pGAD424 expressing Int280a [3]pICC227 pGAD424 expressing Int280aY779/120A TSpICC228 pGAD424 expressing Int280aY887/228A TSpICC229 pGAD424 expressing Int280aY889/230A TSpICC230 pGAD424 expressing Int280aY890/231A TSpICC231 pGAD424 expressing Int280adY228–Y231 TS
1390 S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

(Qiagen). Correct incorporation of each mutation was moni-tored by DNA sequencing using an automated DNA se-quencer. The mutated plasmids were then transformed intoan eae deletion mutant of EPEC strain CVD206 [10].
2.3. Construction of maltose-binding protein(MBP)–Int280 fusions
The mutagenised pCVD438-derivative plasmids werethen used as templates to amplify by PCR the DNA frag-ments encoding the mutated carboxy terminal 280 aminoacid (Int280) regions using one set of primers (forwardprimer 5'-GGAATTCATTACTGAGATTAAGGCT-3'; re-verse primer 5'-CGGGATCCTTATTTTACACAAGTGGC-3'). Following digestion with EcoRI/BamHI, the DNA frag-ments were subcloned into pMAL-c2 for expression asmaltose-binding protein (MBP) fusions (Table 1).
2.3.1. Gel overlays
Purified His-Tir-M (consisting of the intimin-binding re-gion of Tir), expressed from pICC26 in E. coli BL21, waspurified as described [3]. MBP–Int280 derivatives were ex-pressed from recombinants pMal-c2 and purified fromE. coli XL1-Blue as described [12]. His-Tir-M was separatedby SDS-PAGE, blotted onto a nitrocellulose membranewhich was blocked with 10% skimmed milk in PBS, 0.1%Tween-20 overnight. The nitrocellulose membranes werereacted with 5 µg/ml of the purified MBP–Int280 fusions orMBP in PBS, 0.1% Tween-20 for 2 h and washed twice for5 min in PBS, 0.1% Tween-20. MBP–Int280 fusion proteinsbinding to Tir were detected with anti-MBP antiserum(1:2000 for 1 h) and then anti-rabbit antibodies conjugated toalkaline phosphatase (1:2000 for 1 h) as described [3].
2.4. Fluorescent actin stain (FAS) assays, detectionof surface intimin expression and Western blots
Expression of the intimin derivatives on the surface ofCVD206 and their ability to mediate A/E lesion formationwas assessed using rabbit Int280a antiserum [13] and theFAS test developed by Knutton et al. [14], respectively.Briefly, HEp2 cells were grown to approximately 80% con-fluency on coverslips in 24-well plates. Cells were infectedwith 10 µl of static overnight L-broth cultures in mediumlacking antibiotics for 3 h. The monolayers were then washedwith PBS, fixed by the addition of 300 µl 10% formalin for20 min and washed again. For detection of EPEC-associatedintimin, coverslips were incubated with the anti-Int280apolyclonal antiserum (1:50 dilution) for 45 min in DMEMand then with a secondary TRITC-labelled anti-rabbit anti-body (1:250 dilution) for 30 min. Following washes, themonolayers were permeabilised with 0.1% Triton-X 100 for4 min. Filamentous actin was subsequently stained with 5 µlfluorescein isothiocyanate-labelled phalloidin in PBS(0.1 µg/ml) for 20 min (FAS test). Following extensive
washes with PBS, the infected cultures were visualised byfluorescence microscopy.
Expression of the intimin derivatives was also determinedby Western blotting. Briefly, stationary L-broth cultures werediluted 1:100 in DMEM and incubated for 3 h at 37 °C. Anequivalent of an optical density (600) (OD600) of 0.5 wasloaded onto 7.5% SDS-polyacrylamide gel electrophoresis.The electrophoresed polypeptides were transferred to a nitro-cellulose membrane and immunodetection of intimin wasperformed using anti-Int280, diluted 1:500 as described[13,15].
2.5. FACS analysis of surface-expressed intimin by indirectimmunofluorescence
CVD206(pCVD438) derivatives were grown overnight in5 ml LB. Cultures were diluted 1/100 in DMEM and grownto OD600 of 0.6–0.8. Bacteria were washed three times inPBS and fixed for 20 min in 3% formaldehyde in PBS.Bacteria were washed with PBS and incubated with anti-Int280 polyclonal antisera [15] for 30 min at room tempera-ture. Bacteria were then washed with PBS and incubatedwith FITC-conjugated anti-rabbit IgG (Sigma) at room tem-perature for 30 min. Bacteria were washed with PBS and50,000 events were acquired from a Beckton DickinsonFACS analyser using the FL1 channel.
2.6. In vitro organ culture of human intestinal mucosa
Human tissue was obtained, with fully informed parentalconsent and Local Ethical Committee approval, during rou-tine investigation of patients for potential intestinal disor-ders. Mucosal biopsies of proximal small intestinal mucosa(fourth part duodenum) and terminal ileum were taken usinggrasp biopsy forceps during routine endoscopy (OlympusPCF paediatric endoscope). All endoscopic biopsies weretaken from areas showing no obvious pathology or otherabnormality, and all intestinal histology was subsequentlyreported to be normal in the material used in this study.
In vitro organ culture (IVOC) was performed as describedpreviously for 8 h [16]. Each bacterial strain was examined inhuman IVOC on at least three occasions using tissue fromdifferent children. In all cases, an uninoculated specimen wasincluded with each experimental culture to exclude the pos-sibility of in vivo bacterial colonisation; a non-adhesive con-trol (CVD206) was included to exclude non-specific adhe-sion; and a positive control (CVD206(pCVD438)) wasincluded to demonstrate that the sample was susceptible toA/E lesion formation. After incubation with bacteria or ap-propriate control solutions, IVOC specimens were washedthoroughly three times to remove any non-adherent bacteriaand then prepared for scanning electron microscopy (SEM)as described previously [16]. Samples were fixed with 2.5%glutaraldehyde in 0.1 M phosphate buffer, post-fixed in 1%aqueous osmium tetroxide and dehydrated in 2, 2 dimethoxy-propane. Specimens were transferred to absolute ethanol,
1391S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

critically point dried using liquid carbon dioxide in anEmitech K850 apparatus, mounted on aluminium stubs, sput-ter coated with gold-palladium using a Polaron E5100 sput-ter coater, and viewed in a JEOL 5300 SEM at an accelerat-ing voltage of 30 kV.
2.7. Yeast two-hybrid system
The mutagenised pCVD438-derivative plasmids werethen used as templates to amplify by PCR the DNA frag-ments encoding the mutated Int280 regions using one set ofprimers (forward primer 5'-GGAATTCATTACTGAG-ATTAAGGCT-3', reverse primer 5'-CGGGATCCTTATT-TTACACAAGTG GC-3'). After digestion with EcoRI-BamHI, the DNA fragments were subcloned into the yeasttwo-hybrid vector pGAD424.
Yeast two-hybrid system pGAD424-encoding Int280-derivative plasmids (carrying the GAL4 activation domain,AD) were transformed with pICC10 (pGBT9-encoding Tirand carrying the GAL4-binding domain, BD), into the yeasttwo-hybrid host PJ69-4A (MAT a trp1-901 leu2-3112ura3-52 his3-200 gal4D gal80D LYS2::GAL1-HIS3 GAL2-ADE1 met2::GAL7-lacZ), which confers the advantage ofthree independent reporter genes under the control of threedifferent GAL promoters. They were initially selected for theplasmid-encoded TRP1 and LEU2 genes. The resultingtransformants were then replica plated onto 3-aminotriazolecontaining medium to select for the HIS3 reporter, and ontoSC minus Trp, Leu, Ade to select for the ADE2 reporter. Thefunction of LacZ reporter was quantified in cell extracts byassaying for b-galactosidase activity using o-nitrophenylb-D-galactopyranoside as a substrate [17].
2.8. Challenge of mice with C. rodentium
Female, specific pathogen-free, C3H/Hej mice(6–8 weeks old) were purchased from Harlan Olac (Biches-ter, United Kingdom). All mice were housed in individualventilated cages with free access to food and water. Bacterialinoculums were prepared by culturing bacteria overnight at37 °C in 10 ml of L-broth containing nalidixic acid(100 µg/ml) plus chloramphenicol (50 µg/ml). After incuba-tion, bacteria were harvested by centrifugation and resus-pended in a 1-ml volume of PBS. Unanaesthetised mice werethen orally inoculated with 200 µl of the bacterial suspensionusing a gavage needle. The viable count of the inoculum wasdetermined by retrospective plating on L-agar containingappropriate antibiotics.
2.9. Measurement of pathogen burden
Mice were killed 14 d post-infection by cervical disloca-tion. The distal 6 cm of colon was removed and weighed afterremoval of faecal pellets. Colons were then homogenisedmechanically using a Seward 80 stomacher (London, En-gland) and the number of viable bacteria in colonic homoge-
nates determined by viable count on L-agar containing ap-propriate antibiotics.
2.10. Statistical analysis
All statistical analysis was carried out using a two-tailedStudent’s t-test of type two sample equal variance (homosce-dastic).
3. Results
3.1. Site-directed mutagenesis of tyrosines Tyr779/120,Tyr887/288, Ty889/230 and Tyr890/231
Tyr779/120 is present in the Ig domain and makes contactwith Tyr889/230 and Tyr890/231 (positions numbered ac-cording to Int280/whole intimin) in the CTLD (Fig. 1),
Fig. 1. Schematic representation of the structure of Int190 and the locationof the tyrosine mutation sites at the domain interface. The backbone isrepresented by a worm and the mutated side chains are space filled. Numbe-ring is according to full-length intimin.
1392 S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

whereas Tyr887/288 does not contact either Tyr779/120,Ty889/230 or Tyr890/231.
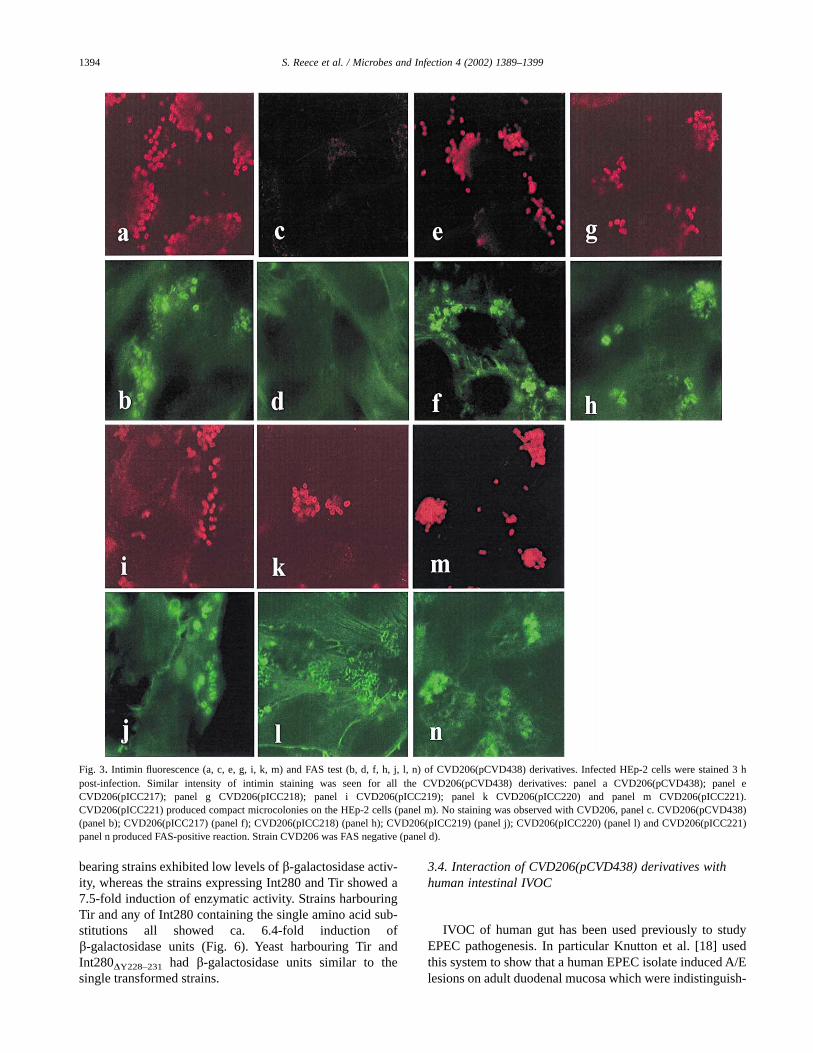
In order to determine the importance of the Tyr779/120,Tyr887/288, Ty889/230 and Tyr890/231 residues for intimin-mediated A/E lesion formation on HEp-2 cells, Tyr to Alasubstitutions were introduced, individually, into pCVD438harbouring the eae gene encoding intimin a [10], generatingplasmids pICC217 (intiminY779/120A), pICC218(intiminY887/288A), pICC219 (intiminY889/230A) and pICC220(intiminY890/231A) (Table 1). The mutant pCVD438 deriva-tives were introduced into CVD206 [10], which harbours anull deletion in eae, and the CVD206 derivatives were sub-jected to a number of biological assays designed to determinethe influence of the mutations on intimin function in vitro.Western blot analysis of whole cell lysates prepared from thedifferent CVD206(pCVD438) derivatives reacted similarlywith anti-Int280a antiserum (Fig. 2). No reactivity was de-tected with CVD206 (Fig. 2). Indirect immunofluorescencestaining showed that after a 3-h incubation with HEp-2 cellsall the adhering CVD206(pCVD438) derivatives, but notCVD206, expressed intimin (Fig. 3). Surface intimin expres-sion was quantified using FACS analysis. AlthoughCVD206(pICC220) presented somewhat lower fluorescentintensity, overall similar levels of surface intimin expressionwere observed with all the other CVD206(pCVD438) deriva-tives, while no signal was observed using CVD206 (Fig. 4).Since expression of intimin was not markedly affected byintroduction of the site-directed mutations, the ability of thedifferent CVD206(pCVD438) derivatives to induce actin po-lymerisation in infected HEp-2 cells (FAS test; [14]), amarker for A/E lesion formation, was determined. This assayshowed that none of the single Tyr substitutions detectablyaffected the ability of the strains to produce a positive FASreaction while CVD206 was FAS negative (Fig. 3).
3.2. Deleting the tyrosine-rich region—effect on biologicalactivity of intimin
Since mutagenesis of each of the four Tyr residues indi-vidually did not detectably affect A/E lesion formation onHEp-2 cells, we produced a derivative of pCVD438 in whichthe entire Tyr-rich region within the CTLD was deleted(Y887/228-Y890/231) (pICC221). Western blot of wholecell lysate of CVD206(pICC221) revealed a similar level of
intimin expression, although more degradation productswere detected (Fig. 2). However, FACS analysis revealed alevel of surface intimin expression similar toCVD206(pCVD438) (Fig. 4). Indeed, CVD206(pICC221)was capable of producing a FAS-positive reaction (Fig. 3),although the adherent bacteria produced more compact mi-crocolonies compared with the other strains (Fig. 2). Theseresults suggest that the Tyr-rich region is not essential for A/Elesion formation in vitro.
3.3. Binding of the modified intimins to Tir
The fact that all strains produced a FAS-positive reactionsuggested that all the intimin derivatives could bind Tir. Inorder to determine this experimentally and compare the dif-ferent intimin derivatives, the Int280 domains harbouring themutation were amplified by PCR and cloned in pMALc-2plasmid, allowing expression of the Int280 domains as trans-lational fusions with MBP. The MBP–Int280 fusion proteinswere purified by affinity chromatography [12] and equalamounts (data not shown) were used together with purifiedTir-M (consisting of the intimin-binding region of Tir [3]) ina gel overlay-binding assay. This revealed that none of thesingle amino acid substitutions detectably affected Tir-binding. In contrast, despite producing a FAS-positive reac-tion, MBP–Int280DY228–Y231, exhibited only a weak Tir-binding activity (Fig. 5).
The yeast two-hybrid system is designed to identify pro-tein–protein interactions through the functional restorationof the yeast GAL4 transcriptional activator in vivo [17]. Inthis study, we expressed the mutations, individually, from arecombinant pGAD424 yeast two-hybrid system vector en-coding Int280, plasmids pICC227 (Int280Y120A), pICC228(Int280Y228A), pIC229 (Int280Y230/A), pICC230(Int280Y231/A) and pICC231 (Int280DY228–231) (Table 1). Wepreviously reported that in the yeast two-hybrid system, theinteraction of Int280 (pICC19) with the whole Tir polypep-tide (pICC10) is more efficient than the interaction of Int280with Tir-M [3]. Accordingly, the DNA fragment encoding thewhole Tir polypeptide, expressed from the second yeasttwo-hybrid system vector, pGBT9 (pICC10) [3], was used todetermine the effect of the mutagenesis on Int280–Tir proteininteraction.
Plasmid pICC10 was co-transformed with each of thedifferent pGAD424-based plasmids into a derivative of theyeast stain PJ69-4A selected previously as a reporter forintimin–Tir interaction [3]. Replica plating these coloniesonto selective media yielded vigorously growing colonies,and hence a positive two-hybrid phenotype, in yeast strainsexpressing both Tir and Int280 (pICC10 and pICC19) and Tirand all the single amino acid substitutions (data not shown).In contrast, yeast harbouring pICC10 and pICC231 did notgrow on the selective medium.
The function of the non-selective reporter, lacZ, was alsoassessed in these strains by measuring b-galactosidase activi-ties (Fig. 6). The host (data not shown) or single plasmid-
Fig. 2. Western blot of CVD206(pCVD438)-derivative whole cell lysates.The nitrocellulose membrane was probed with the Int280a antiserum. Full-length intimin was detected only in strains harbouring the pCVD438 deriva-tives. Lane 1, CVD206(pCVD438); lane 3, CVD206(pICC217); lane 4,CVD206(pICC218); lane 5, CVD206(pICC219); lane 6, CVD206(pICC220); lane 7, CVD206(pICC221). No intimin was detected inCVD206 (lane2).
1393S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399
bearing strains exhibited low levels of b-galactosidase activ-ity, whereas the strains expressing Int280 and Tir showed a7.5-fold induction of enzymatic activity. Strains harbouringTir and any of Int280 containing the single amino acid sub-stitutions all showed ca. 6.4-fold induction ofb-galactosidase units (Fig. 6). Yeast harbouring Tir andInt280DY228–231 had b-galactosidase units similar to thesingle transformed strains.
3.4. Interaction of CVD206(pCVD438) derivatives withhuman intestinal IVOC
IVOC of human gut has been used previously to studyEPEC pathogenesis. In particular Knutton et al. [18] usedthis system to show that a human EPEC isolate induced A/Elesions on adult duodenal mucosa which were indistinguish-
Fig. 3. Intimin fluorescence (a, c, e, g, i, k, m) and FAS test (b, d, f, h, j, l, n) of CVD206(pCVD438) derivatives. Infected HEp-2 cells were stained 3 hpost-infection. Similar intensity of intimin staining was seen for all the CVD206(pCVD438) derivatives: panel a CVD206(pCVD438); panel eCVD206(pICC217); panel g CVD206(pICC218); panel i CVD206(pICC219); panel k CVD206(pICC220) and panel m CVD206(pICC221).CVD206(pICC221) produced compact microcolonies on the HEp-2 cells (panel m). No staining was observed with CVD206, panel c. CVD206(pCVD438)(panel b); CVD206(pICC217) (panel f); CVD206(pICC218) (panel h); CVD206(pICC219) (panel j); CVD206(pICC220) (panel l) and CVD206(pICC221)panel n produced FAS-positive reaction. Strain CVD206 was FAS negative (panel d).
1394 S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

able from those seen in vivo. Hicks et al. [16] showed thatA/E formation by EPEC in IVOC is an intimin-dependentevent.
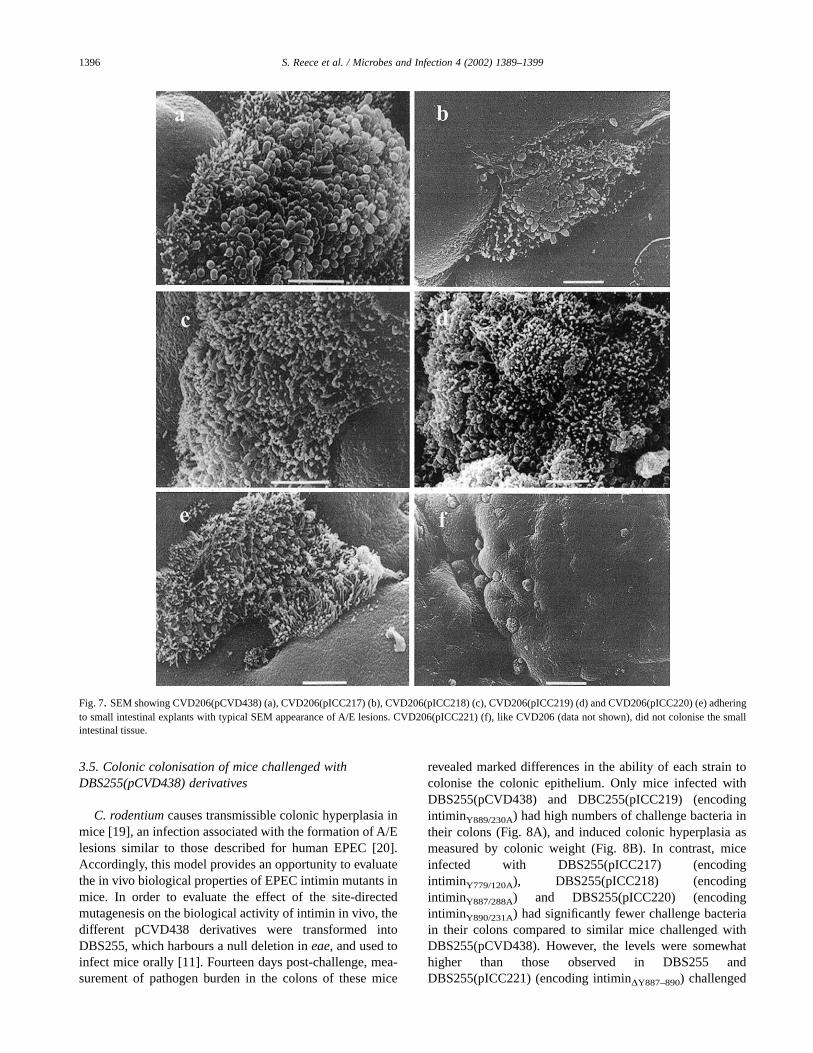
Histologically normal tissue obtained from the distalduodenum and terminal ileum was examined after infectionwith the different CVD206(pCVD438) derivatives to inves-tigate the effect of the site-directed mutagenesis on intimin-mediated mucosal attachment and A/E lesion formation us-ing an ex vivo infection model. Examination of the infectedbiopsies by SEM revealed that the single amino acid substi-tutions did not affect adhesion to and A/E lesion formation onthe IVOC by the different CVD206(pCVD438) derivatives(Fig. 7). In contrast, deletion of the Tyr-rich region abrogatedthe ability of CVD206(pICC221) to bind and induce A/Elesions on the human gut specimens (Fig. 7).
Fig. 4. Detection of surface intimin expression by FACS. Surface intimin expression was not affected by the mutagenesis; a comparable level of fluorescentintensity was seen on CVD206(pCVD438) (panel b) or the mutated intimin polypeptides: CVD206(pICC217) (panel c); CVD206(pICC218) (panel d);CVD206(pICC219) (panel e); and CVD206(pICC221) (panel g). CVD206(pICC220) (panel f) presented somewhat lower fluorescent intensity. Intimin was notdetected on CVD206 (panel a) and on a small proportion of CVD206(pCVD438) (panel b) and CVD206(pICC218) (panel d).
Fig. 5. Detection of Int280–Tir interaction using gel overlays. ImmobilisedTir-M was overlayed with the different MBP–Int280 derivatives. Bindingwas detected using rabbit MBP antiserum. MBP–Int280 (lane 2),MBP–Int280Y120A (lane 3), MBP–Int280Y1288A (lane 4), MBP–Int280Y230A
(lane 5) MBP–Int280Y231A (lane 6) interacted similarly with Tir-M.MBP–Int280DY228–Y231 bound weakly to the immobilised Tir-M (lane 7),while MBP only (lane 1) did not bind Tir-M.
Fig. 6. Detection of Int280–Tir interaction using the yeast two-hybrid sys-tem. b-Galactosidase assay showing 6.4–7.5 increase in enzymatic activityin strains co-expressing the whole Tir polypeptide and Int280 or any of thesingle Tyr substitutions compared with yeast expressing either Int280 or Tironly. Int280Y779/120A, Int280Y887/228A, Int280Y889/230A and Int280Y890/231A,analysed by the yeast two-hybrid system, produced levels of reporter geneactivation that were significantly lower than the same experiment usingwild-type Int280 (P < 0.05), but significantly higher than the same experi-ment using negative controls (P < 0.05). Int280Dl228–231 produced levels ofreporter gene activation that were not significantly different from those inthe same experiment using negative controls (P > 0.05). Error bars representstandard deviation of data for at least three separate experiments.
1395S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399
3.5. Colonic colonisation of mice challenged withDBS255(pCVD438) derivatives
C. rodentium causes transmissible colonic hyperplasia inmice [19], an infection associated with the formation of A/Elesions similar to those described for human EPEC [20].Accordingly, this model provides an opportunity to evaluatethe in vivo biological properties of EPEC intimin mutants inmice. In order to evaluate the effect of the site-directedmutagenesis on the biological activity of intimin in vivo, thedifferent pCVD438 derivatives were transformed intoDBS255, which harbours a null deletion in eae, and used toinfect mice orally [11]. Fourteen days post-challenge, mea-surement of pathogen burden in the colons of these mice
revealed marked differences in the ability of each strain tocolonise the colonic epithelium. Only mice infected withDBS255(pCVD438) and DBC255(pICC219) (encodingintiminY889/230A) had high numbers of challenge bacteria intheir colons (Fig. 8A), and induced colonic hyperplasia asmeasured by colonic weight (Fig. 8B). In contrast, miceinfected with DBS255(pICC217) (encodingintiminY779/120A), DBS255(pICC218) (encodingintiminY887/288A) and DBS255(pICC220) (encodingintiminY890/231A) had significantly fewer challenge bacteriain their colons compared to similar mice challenged withDBS255(pCVD438). However, the levels were somewhathigher than those observed in DBS255 andDBS255(pICC221) (encoding intiminDY887–890) challenged
Fig. 7. SEM showing CVD206(pCVD438) (a), CVD206(pICC217) (b), CVD206(pICC218) (c), CVD206(pICC219) (d) and CVD206(pICC220) (e) adheringto small intestinal explants with typical SEM appearance of A/E lesions. CVD206(pICC221) (f), like CVD206 (data not shown), did not colonise the smallintestinal tissue.
1396 S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

animals (Fig. 8A). Nevertheless, infection with these bacteriadid not induce colonic hyperplasia, as measured by colonicweight (Fig. 8B).
4. Discussion
EPEC, an established aetiological agent of human infan-tile diarrhoea, is a bacterial pathogen that subverts intestinalepithelial cell function to produce distinctive ‘attaching &effacing’ (A/E) lesions. These lesions are characterised bylocalised destruction (effacement) of brush border microvilli,intimate bacterial attachment to the host cell membrane andformation of an actin-rich cytoskeletal structure beneath inti-mately attached bacteria. All the genes required for A/Elesion formation are encoded by the locus of enterocyte
effacement (LEE) pathogenicity island [21]. This encodes atype III secretion system [22], type III secreted proteinsEspA, EspB, and EspD [23], an intimate EPEC adhesin,intimin [1,2] and an intimin receptor, Tir [4] that is translo-cated and inserted into the host cell membrane by the EPECtype III secretion system, where it binds to intimin. Intim-in–Tir interaction leads to cytoskeletal accumulation beneathintimately attached bacteria and A/E lesion formation [23].Two further LEE-encoded proteins, EspF [24] and Map [25],are translocated into the host cell cytosol by the type IIIsecretion system, although these EPEC effector proteins donot appear to be involved in A/E lesion formation.
Intimin is a colonisation factor required for full virulencein human volunteers and animal models [11,26–28]. More-over, antigenic variation within the receptor-binding regionof intimin allows the definition of five subclasses of intimintypes (a, b, c, d and e) [15,29], variation which contributes tohost and tissue specificity [30]. Indeed, we have recentlyreported that changing a single amino acid (Val897Ala)within the solvent-exposed, Tir-binding interface, of intimina, resulted in a phenotype characteristic of intimin c [31].However, despite the reported variation in this region ofintimin, several conserved motifs are recognisable. Theseinclude the 880WAGGANKYX(Y/H)Y890 motif, compris-ing the Tyr-rich region (Fig. 1), previously hypothesised to bepart of the binding site.
In this study, we systematically mutagenised each of thefour Tyr residues at the Int280 D2/D3 interface and testedhow this affected intimin activity in vitro and in vivo. In theHEp-2 infection model, we did not detect any differencesbetween the ability of CVD206 expressing the mutated in-timins to mediate A/E lesions compared with CVD206 ex-pressing the wild-type intimin. Consistent with this, surfaceexpression of the intimin mutants, determined by FACS andimmunofluorescence, was similar to the wild-type intimin aswas the level of total intimin seen by western blots. Inaddition, the ability of the mutated intimin to bind recombi-nant Tir was also not compromised. Similarly, CVD206expressing the wild-type and mutated intimins were able tobind to and induce A/E lesions on human intestinal mucosalsurfaces in vitro. Due to the lack of a clear phenotype for eachof the four individual mutations, we decided to delete theTyr-rich region altogether. This drastic step did not affectsecretion of intimin to the membrane compartment of thebacterial cell, or, when expressed in CVD206, A/E lesionformation on HEp-2 cells. However, in the absence of theTyr-rich region, intimin seems to be less stable on westernblots, and MBP–Int280DY288–Y231 bound Tir very ineffi-ciently. Consistent with this, CVD206(pICC221) was unableto colonise or induce A/E lesion on human IVOC. Collec-tively, these results show that despite affecting intimin activi-ties, the Tyr-rich region is not essential for A/E lesion forma-tion on HEp-2 cells. The reason for the compactmicrocolonies that CVD206(pICC221) bacteria produced isnot clear.
Fig. 8. Virulence of C. rodentium strains expressing mutated intimin mole-cules. (A) The data depict the number of C. rodentium recovered fromcolonic tissue of individual mice orally infected 14 d previously. Miceinfected with DBS255(pCVD438) and DBS255(pICC219) had high patho-gen burdens. In contrast, mice infected with DBS255(pICC217),DBS255(pICC218), DBS255(pICC220) had lower bacterial loads, althoughstill higher than DBS255 and DBC255(pICC221) from which no bacteriawere recovered. (B) The distal 6 cm of the colon was weighed 14 dpost-challenge. Mice infected with DBS255(pICC219) produced colon wei-ghts that were not significantly different from colon weights of mice infectedwith DBS255(pCVD438) (P > 0.05). There was no significant differencebetween the colon weights of mice infected with avirulent DBS255 and thecolon weights of mice infected with DBS255(pICC217),DBS255(pICC218), DBS255(pICC220) or DBS255(pICC221) (P > 0.05).
1397S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

The mouse pathogen C. rodentium was used to assess theaffect of mutagenesis on the function of intimin in vivo. Theexpression of intimin a in C. rodentium DBS255 restores theability of the strain to colonise the mouse colon and inducecolonic hyperplasia [32,33]. In addition, intimin a can bindTir from C. rodentium [34]. Evaluating intimin mutants inthe C. rodentium in vivo infection model, where the bacteriaalso have to compete with the normal intestinal flora, differ-entiated between the biological activities of the individualsingle Tyr amino acid substations. Substitution of Tyr889 didnot notably affect the level of DBS255(pICC219) colonisa-tion or colonic hyperplasia (as measured by colon weight),whereas substitution of the adjacent residue, Tyr890Ala,resulted in decreased DBS255(pICC220) colonisation withno hyperplastic reaction. Phenotypes similar to that exhibitedby DBS255(pICC220) were observed with the other twoDBS255 derivatives expressing intimin Tyr779Ala(DBS255(pICC217)) and Tyr887Ala (DBS255(pICC218)).Importantly, the levels of infecting bacteria recovered fromthe colons were somewhat higher than those observed inDBS255-challenged animals. In contrast, following infec-tion with DBS255(pICC221) (encoding intiminDY887–890),no challenged bacteria were recovered and no hyperplasiawas recorded. This might reflect the increased sensitivity ofintiminDY887–890 to proteolysis or to a complete loss of func-tion in vivo.
Three of the tyrosine mutants are significantly attenuatedfor colonisation of the mouse, yet the mutated intimin mol-ecule is able to bind to Tir and induce A/E lesions in vitro.This implies that Tir-binding site is unaffected by mutation.However, the mutants that affect the colonisation of themouse support the hypothesis of a second function for in-timin in pathogenesis [35]. This role most likely involves theformation or the stabilisation of a structure that is involved incolonisation, which could include the binding of a host cellreceptor in the early stages of infection. Perhaps the regionformed by the tyrosine residues, particularly Tyr779 andTyr890, is part of a binding site within intimin that is distinctfrom the well-characterised intimin–Tir interaction surface.This is not unreasonable, as this group of tyrosine residues isdistant from the Tir-binding site and lies in a partially ex-posed cleft situated at the domain interface (Fig. 1). Indeed,the fact that Tyr 779 and Tyr 890 are conserved, within thevariable Int190 region, in all the different intimin types,implies that these residues are maintained through a force ofnatural selection. More specifically, Tyr779 is present in thefirst domain and makes contacts with Tyr889 and Tyr890,whereas Tyr887 does not contact either Tyr779, Tyr889 orTyr890. Furthermore, this region also shows some distinctdifferences between the solution and crystal states, whichcan often be a pointer for functionally significant residues.The identity of the putative host intimin receptor within themouse colon remains elusive, although two potential host-cell-encoded intimin receptors (Hirs) have been reported, i.e.b1 integrins [36] and nucleolin [37]. Further studies arerequired to determine their relevance in vivo and the exist-
ence of yet other unidentified Hirs. Our laboratory togetherwith others is involved in unravelling the identity and biologyof potential Hirs.
Acknowledgements
We are grateful to Rob Heuschkel, Simon Murch andMike Thomson for their endoscopic skills in the provision ofendoscopic biopsies. This investigation was supported bygrants from Action Research and the BBSRC.
References
[1] A.E. Jerse, J.Yu, B.D. Tall, J.B. Kaper,A genetic locus of enteropatho-genic Escherichia coli necessary for the production of attaching andeffacing lesions on tissue culture cells, Proc. Natl. Acad. Sci. USA 7(1990) 7839–7843.
[2] A.E. Jerse, J.B. Kaper, The eae gene of enteropathogenic Escherichiacoli encodes a 94-kilodalton membrane protein, the expression ofwhich is influenced by the EAF plasmid, Infect. Immun. 59 (1991)4302–4309.
[3] E.L. Hartland, M. Batchelor, R.M. Delahay, C. Hale, S. Matthews,G. Dougan, S. Knutton, I. Connerton, G. Frankel, Binding of intiminfrom enteropathogenic Escherichia coli to Tir and to host cells, Mol.Microbiol. 32 (1999) 151–158.
[4] B. Kenny, R. DeVinney, M. Stein, D.J. Reinscheid, E.A. Frey,B.B. Finlay, Enteropathogenic E. coli (EPEC) transfers its receptorfor intimate adherence into mammalian cells, Cell 91 (1997) 511–520.
[5] G. Kelly, S. Prasannan, S. Daniell, K. Fleming, G. Frankel, G. Dou-gan, I. Connerton, S. Matthews, Structure of the cell-adhesion frag-ment of intimin from enteropathogenic Escherichia coli, Nat. Struct.Biol. 6 (1999) 313–318.
[6] G. Kelly, S. Prasannan, S. Daniel, G. Frankel, G. Dougan, I. Conner-ton, S. Mathews, Sequential assignment of the triple-labelled 30.1kDa cell adhesion domain of intimin from enteropathogenic E. coli, J.Biomol. NMR. 12 (1998) 189–191.
[7] Y. Luo, E.A. Frey, R.A. Pfuetzner, A.L. Creagh, D.G. Knoechel,C.A. Haynes, B.B. Finlay, N.C. Strynadka, Crystal structure of en-teropathogenic Escherichia coli intimin-receptor complex, Nature405 (2000) 1073–1077.
[8] M. Batchelor, S. Prasannan, S. Daniell, S. Reece, I. Connerton,G. Bloomberg, G. Dougan, G. Frankel, S. Matthews, Structural basisfor recognition of the translocated intimin receptor (Tir) by intiminfrom enteropathogenic Escherichia coli, EMBO J. 19 (2000)2452–2464.
[9] M.M. Levine, E.J. Berquist, D.R. Nalin, D.H. Waterman, R.B. Hor-nick, C.R. Young, S. Stoman, B. Rowe, Escherichia coli that causediarrhoea but do not produce heat-labile or heat-stable enterotoxinsand are non-invasive, Lancet i (1978) 119–122.
[10] M.S. Donnenberg, J.B. Kaper, Construction of an eae deletion mutantof enteropathogenic Escherichia coli by using a positive-selectionsuicide vector, Infect. Immun. 59 (1991) 4310–4317.
[11] D.B. Schauer, S. Falkow, The eae gene of Citrobacter freundii biotype4280 is necessary for colonization in transmissible murine colonichyperplasia, Infect. Immun. 61 (1993) 4654–4661.
[12] G. Frankel, D.C. Candy, P. Everest, G. Dougan, Characterization ofthe C-terminal domains of intimin-like proteins of enteropathogenicand enterohemorrhagic Escherichia coli, Citrobacter freundii, andHafnia alvei, Infect. Immun. 62 (1994) 1835–1842.
1398 S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399

[13] G. Frankel, A.D. Philips, M. Novakova, M. Batchelor, S. Hicks,G. Dougan, Generation of Escherichia coli intimin-derivatives withdiffering biological activities using site-directed mutagenesis of theintimin C-terminus domain, Mol. Microbiol. 29 (1998) 559–570.
[14] S. Knutton, T. Baldwin, P.H. Williams, A.S. McNeish, Actin accumu-lation at sites of bacterial adhesion to tissue culture cells: basis of anew diagnostic test for enteropathogenic and enterohemorrhagic Es-cherichia coli, Infect. Immun. 57 (1989) 1290–1298.
[15] J. Adu-Bobie, G. Frankel, C. Bain, A.G. Goncaleves, L.R. Trabulsi,G. Douce, S. Knutton, G. Dougan, Detection of intimin a, b, c, and d,four intimin derivatives expressed by attaching and effacing microbialpathogens, J. Clin. Microbiol. 36 (1998) 662–668.
[16] S. Hicks, G. Frankel, J.B. Kaper, G. Dougan, A.D. Phillips, Role ofintimin and bundle forming pili in enteropathogenic Escherichia coliadhesion to paediatric intestine in vitro, Infect. Immun. 66 (1998)1570–1578.
[17] P. James, J. Halladay, E.A. Craig, Genomic libraries and a host straindesigned for highly efficient two-hybrid selection in yeast, Genetics144 (1996) 1425–1436.
[18] S. Knutton, D.R. Lloyd, A.S. McNeish, Adhesion of enteropathogenicEscherichia coli to human intestinal enterocytes and cultured humanintestinal mucosa, Infect. Immun. 55 (1987) 69–77.
[19] S.W. Barthold, G.L. Coleman, R.O. Jacoby, E.M. Livstone, A.M. Jo-nas, Transmissible murine colonic hyperplasia, Vet. Pathol. 15 (1976)223–236.
[20] D.B. Schauer, S. Falkow, Attaching and effacing locus of a Citro-bacter freundii biotype that causes transmissible murine colonic hy-perplasia, Infect. Immun. 61 (1993) 2486–2492.
[21] T.K. McDaniel, K.G. Jarvis, M.S. Donnenberg, J.B. Kaper, A geneticlocus of enterocyte effacement conserved among diverse enterobacte-rial pathogens, Proc. Natl. Acad. Sci. USA 92 (1995) 1664–1668.
[22] K.G. Jarvis, J.A. Giron, A.E. Jerse, T.K. McDaniel, M.S. Donnenberg,J.B. Kaper, Enteropathogenic Escherichia coli contains a putativetype III secretion system necessary for the export of proteins involvedin attaching and effacing lesion formation, Proc. Natl. Acad. Sci. USA92 (1995) 7996–8000.
[23] G. Frankel, A.D. Phillips, I. Rosenshine, G. Dougan, J.B. Kaper,S. Knutton, Enteropathogenic and enterohaemorrhagic Escherichiacoli: more subversive elements, Mol. Microbiol. 30 (1998) 911–921.
[24] B.P. McNamara, M.S. Donnenberg, A novel proline-rich protein,EspF, is secreted from enteropathogenic Escherichia coli via the typeIII export pathway, FEMS Microbiol. Lett. 166 (1998) 71–78.
[25] B. Kenny, M. Jepson, Targeting of an enteropathogenic Escherichiacoli (EPEC) effector protein to host mitochondria, Cell. Microbiol. 2(2000) 579–590.
[26] M.S. Donnenberg, S. Tzipori, M.L. McKee, A.D. O’Brien, J. Alroy,J.B. Kaper, The role of the eae gene of enterohemorrhagic Escheri-chia coli in intimate attachment in vitro and in a porcine model, J.Clin. Invest. 92 (1993) 1418–1424.
[27] M.S. Donnenberg, C.O. Tacket, S.P. James, G. Losonsky, J.P. Nataro,S.S. Wasserman, J.B. Kaper, M.M. Levine, Role of the eaeA gene inexperimental enteropathogenic Escherichia coli infection, J. Clin.Invest. 92 (1993) 1412–1417.
[28] M.L. McKee, A.R. Melton-Celsa, R.A. Moxley, D.H. Francis,A.D. O’Brien, Enterohemorrhagic Escherichia coli O157:H7 requiresintimin to colonize the gnotobiotic pig intestine and to adhere toHEp-2 cells, Infect. Immun. 63 (1995) 3739–3744.
[29] E. Oswald, H. Schmidt, S. Morabito, H. Karch, O. Marches, A. Cap-rioli, Typing of intimin genes in human and animal enterohemorrhagicand enteropathogenic Escherichia coli: characterization of a newintimin variant, Infect. Immun. 68 (2000) 64–71.
[30] A.D. Phillips, G. Frankel, Intimin-mediated tissue specificity in en-teropathogenic Escherichia coli interaction with human intestinalorgan cultures, J. Infect. Dis. 181 (2000) 1496–1500.
[31] S. Reece, C.P. Simmons, R.J. Fitzhenry, S. Matthews, A.D. Phillips,G. Dougan, G. Frankel, Site-directed mutagenesis of intimin alphamodulates intimin-mediated tissue tropism and host specificity, Mol.Microbiol. 40 (2001) 86–98.
[32] G. Frankel, A.D. Phillips, M. Novakova, H. Field, D.C. Candy,D.B. Schauer, G. Douce, G. Dougan, Intimin from enteropathogenicEscherichia coli restores murine virulence to a Citrobacter rodentiumeaeA mutant: induction of an immunoglobulin A response to intiminand EspB, Infect. Immun. 64 (1996) 5315–5325.
[33] L.M. Higgins, G. Frankel, I. Connerton, N.S. Goncalves, G. Dougan,T.T. MacDonald, Role of bacterial intimin in colonic hyperplasia andinflammation, Science 285 (1999) 588–591.
[34] E.L. Hartland, V. Huter, L.M. Higgins, N.S. Goncalves, G. Dougan,A.D. Phillips, T.T. MacDonald, G. Frankel, Expression of intimingamma from enterohemorrhagic Escherichia coli in Citrobacter ro-dentium, Infect. Immun. 68 (2000) 4637–4646.
[35] G. Frankel, A.D. Phillips, L.R. Trabulsi, S. Knutton, G. Dougan,S. Matthews, Intimin and the host cell – is it bound to end in Tir(s)?,Trends Microbiol. 9 (2001) 214–218.
[36] G. Frankel, O. Lider, R. Hershkoviz, A.P. Mould, S.G. Kachalsky,D.C.A. Candy, L. Cahalon, M.J. Humphries, G. Dougan, The cell-binding domain of intimin from enteropathogenic Escherichia colibinds to beta1 integrins, J. Biol. Chem. 271 (1996) 20359–20364.
[37] J.F. Sinclair, A.D. O’Brien, Cell-surface localized nucleolin is a eu-caryotic receptor for the adhesin intimin-gamma of enterohemor-rhagic Escherichia coli O157: H7, J. Biol. Chem. 277 (2002)2876–2885.
1399S. Reece et al. / Microbes and Infection 4 (2002) 1389–1399