universidade federal do cearÁ departamento de …neto e gerardo, pois, com certeza, cada um...
TRANSCRIPT

0
UNIVERSIDADE FEDERAL DO CEARÁ
DEPARTAMENTO DE BIOQUÍMICA E BIOLOGIA MOLECULAR
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
CARACTERIZAÇÃO FÍSICO-QUIMICA E ESTRUTURAL DE POLISSACARÍDEOS
OBTIDOS DE FOLHAS DA PLANTA Aloe barbadensis Miller E AVALIAÇÃO DE
SUAS ATIVIDADES ANTIVIRAL E ANTI-HEMORRÁGICA
YGOR RAPHAEL GOMES ELOY
FORTALEZA / 2012

1
YGOR RAPHAEL GOMES ELOY
CARACTERIZAÇÃO FÍSICO-QUIMICA E ESTRUTURAL DE POLISSACARÍDEOS
OBTIDOS DE FOLHAS DA PLANTA Aloe barbadensis Miller E AVALIAÇÃO DE
SUAS ATIVIDADES ANTIVIRAL E ANTI-HEMORRÁGICA
Tese de Doutorado apresentada
como requisito parcial à obtenção
do grau de doutor pelo Programa de
Pós-Graduação em Bioquímica da
Universidade Federal do Ceará.
Orientadora: Profa. Dra. Norma
Maria Barros Benevides
FORTALEZA
Março/2012



4
Ao grande Designer do Universo, toda honra e toda glória és para ti Senhor.

5
Dedico essa tese com muito amor e carinho à
minha mãe Francisca, a meu pai Carlos, a minha
irmã Patrícia, a minha amada esposa Gardênia, a
minha filha Guadalupe e, em especial, ao casal Ana
Lúcia e Wagner Barbosa, que sempre confiaram em
mim. Dedico, também, aos meus eternos amigos
Catarina e Xavier, que despertaram a minha
curiosidade pela matéria-prima apreciada nesse
trabalho.
Com carinho.

6
AGRADECIMENTOS
Um agradecimento muito especial a profa. Dra. Norma Maria Barros
Benevides, minha orientadora. Agradeço por ter acreditado no projeto que foi
proposto e aceitado a minha inclusão no corpo de pesquisadores do laboratório
de Carboidratos e Lectinas (Carbolec). Obrigado pela boa convivência, pela
amizade e pelas discussões levantadas acerca do projeto de pesquisa. Além
disso, sou grato pela liberdade de ter tomado minhas próprias decisões e
atitudes nas atividades relacionadas ao doutorado.
À profa. Dra. Ana de Fátima Fontenele Urano Carvalho, por ter sido para
mim, desde a graduação, um símbolo de seriedade científica e de competência
profissional.
À profa. Dra. Maria Izabel Florindo Guedes, pela atenção,
disponibilidade e por ter proporcionado o primeiro contato com a área da
virologia.
À profa. Dra. Maria Teresa Villela Romanos, pela disponibilidade e pela
indescritível atenção para comigo. Devo muito mais que um agradecimento,
pois o início de nossa jornada foi marcado por compromisso e um profundo
respeito um pelo outro.
Ao prof. Dr. Vitor Hugo Pomin, pela disponibilidade, seriedade e
compromisso em todas as atividades desenvolvidas durante nossa parceria.
Obrigado mais uma vez, por ter apostado, junto comigo, nas peculiaridades
estruturais dos polissacarídeos de A. barbadensis.
Ao prof. Dr. Paulo Antônio de Souza Mourão, por ter me aceitado em
seu laboratório e ter demostrado um carisma incomparável durante nossas
discussões.
À Profa. Dra. Regina Célia Monteiro de Paula pela receptividade e pela
colaboração.

7
Ao prof. Dr. Francisco José de Abreu Matos, in memoriam, e toda sua
equipe do Horto de Plantas Medicinais da Universidade Federal do Ceará, pela
disponibilidade de coleta das plantas e atenção prestadas.
Aos demais professores do Departamento de Bioquímica e Biologia
Molecular, aos quais reservo grande apreço e consideração, por todos os anos
de convívio e relacionamento amigável.
Aos colegas e amigos do laboratório Carbolec: Ariévilo, Chistiane, Ianna,
Ana Luíza, Bruno, Natássia, Danilo, Érika, Edna, Luana, Trycia, Jane, Gabriela
Almeida, Ticiana Abreu, Ticiana Lima, Willame, Gardênia, Fabiula, Ricardo,
Neto e Gerardo, pois, com certeza, cada um contribuiu um pouco para esse
trabalho. Um agradecimento especial a Ismael e Edfranck, pois foram peças
fundamentais nesse trabalho.
A todos os colegas do Laboratório de Bioquímica Humana (UECE) e, em
especial, Márcia Marques e Raquel.
A todos os colegas e amigos do Laboratório de Tecidos Conjuntivos da
UFRJ e, especialmente, Gustavo Ramalho, Stefan, profa. Dra. Ana Tovar e
Adriana, pela ajuda e disponibilidade a mim prestadas.
Aos colegas e amigos do Laboratório Experimental de Drogas Antivirais
e Citotóxicas (UFRJ) e do Laboratório de Viroses Respiratórias, Entéricas e
Oculares (UFRJ). Sou muito grato pelo apoio prestado por Gabriella da Silva
Mendes, Jéssica Figueiredo Cavalcanti e Soluza. Obrigado pelos ensinamentos
e pela indiscutível disponibilidade.
A Sahra Souza pela atenção no momento de montagem das exsicatas
no Herbário.
Aos meus pais, Carlos e Francisca, pelo amor, orações, confiança, força
e por ter dado todas as ferramentas básicas para que tivesse uma vida digna
com saúde e educação de qualidade.
À minha irmã, pelo carinho e amor concedidos a mim.

8
Ao casal Ana Lúcia e Wagner Barbosa, por ter me apoiado tantos anos
durante minha caminhada no mestrado e doutorado. Obrigado, também, pela
confiança e pelo indiscutível e incondicional amor.
Às minhas princesas Gardênia e Guadalupe, pelo amor, carinho e
confiança. Obrigado por terem marcado, irreversivelmente, minha vida, pois
hoje sou uma pessoa mais feliz, graças a vocês.
A minha sogra, Graça Magalhães, pelo amor e pelas orações.
A todos os demais alunos, professores e funcionários da UFC, UECE e
UFRJ que contribuíram indiretamente para minha formação.

9
Este trabalho foi realizado graças às seguintes Instituições:
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES),
pela concessão da bolsa vinculada ao programa de Pós-graduação em
Bioquímica da Universidade Federal do Ceará.
À Fundação Cearense de Amparo à Pesquisa (FUNCAP), pelos auxílios ao
Programa de Pós-graduação em Bioquímica.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ),
pelos auxílios ao Programa de Pós-graduação em Bioquímica.
Ao Ministério da Ciência e Tecnologia (MCT), pelos auxílios ao Programa de
Pós-Graduação em Bioquímica.
Ao Ministério da Saúde (MS), pelos auxílios ao Programa de Pós-graduação
em Bioquímica.
Ao Programa de Pesquisa para o Sistema Único de Saúde (PPSUS), por ter
fomentado grande parte de minha pesquisa.
Ao Carbolec do Departamento de Bioquímica e Biologia Molecular da
Universidade Federal do Ceará, pelo apoio e suporte para execução da
primeira etapa do trabalho.

10
Ao Laboratório de Tecido Conjuntivo do Hospital Clementino Fraga Filho
(UFRJ) pela execução das análises espectroscópicas de Ressonância
Magnética Nuclear (RMN).
Laboratório de Polímeros do Departamento de Química Orgânica e Inorgânica
da Universidade Federal do Ceará pela execução das análises
espectroscópicas de Infravermelho (IR).
Ao Laboratório de Bioquímica Humana da Universidade Estadual do Ceará,
pela iniciação nas atividades relacionadas à virologia.
Ao Laboratório Experimental de Drogas Antivirais e Citotóxicas (UFRJ) e o
Laboratório de Viroses Respiratórias, Entéricas e Oculares (UFRJ), pela
execução das atividades antivirais e de citotoxicidade.
Ao Laboratório de Bioprospecção de Recursos Regionais (UFC), pela
execução das análises hematológicas.

11
SUMÁRIO
Página
LISTA DE FIGURAS 15
LISTA DE TABELAS
LISTA DE ABREVIATURAS
22
24
RESUMO 25
ABSTRACT 26
CAPÍTULO 1 - INTRODUÇÃO 27
1.1. ASPECTOS SOBRE POLISSACARÍDEOS DE VEGETAIS
SUPERIORES
28
1.1.1. Polissacarídeos de parede celular vegetal 30
1.1.1.1. Celulose 34
1.1.1.2. Polissacarídeos da fase matricial da parede celular 34
1.1.1.2.1. Substâncias pécticas 35
1.1.1.2.2. Hemiceluloses 36
1.1.1.2.3. Polissacarídeos de reserva da parede celular
vegetal
39
1.1.2. Polissacarídeos sulfatados 42
1.1.3. O gênero Aloe 43
1.1.4. Polissacarídeos de A. barbadensis e propriedades
biológicas relacionadas
45
1.2. VIROLOGIA 47
1.2.1. Adenovírus 48
1.2.2. Herpes Simples Vírus (HSV) 50
1.2.3. Metapneumovírus (HMPV) 51
1.2.4. Dengue Vírus (DENV) 55
1.2.5. Infecções Virais Hemorrágicas 57
1.2.6. Hemostasia 58

12
1.3. OBJETIVOS 62
1.3.1. Gerais 62
1.3.2. Específicos 62
CAPÍTULO 2 – CARACTERIZAÇÃO FÍSICO-QUÍMICA E
ESTRUTURAL
64
2.1. MATERIAIS E MÉTODOS 65
2.1.1. Coleta e Identificação 65
2.1.2. Obtenção dos Polissacarídeos Totais (PT) de A.
barbadensis
65
2.1.3. Determinação de Umidade da Polpa 68
2.1.4 Fracionamento dos PT por Cromatografia de Troca
Iônica em Coluna de DEAE-celulose
68
2.1.5. Caracterização Físico-química dos Polissacarídeos 69
2.1.5.1. Determinação de Carboidratos Totais (CT) 69
2.1.5.2. Determinação de Ácidos Urônicos 69
2.1.5.3. Determinação do Teor de Sulfato 69
2.1.5.4. Determinação de Contaminantes Protéicos 70
2.1.5.5. Eletroforese em Gel de Agarose 70
2.1.5.6. Eletroforese em Gel de Poliacrilamida (PAGE) 70
2.1.5.7. Cromatografia de Troca Iônica em Coluna de Mono-
q Acoplada a HPLC
71
2.1.5.8. Determinação da Composição Monossacarídica das
Frações Polissacarídeos
71
2.1.6. Caracterização Estrutural dos Polissacarídeos 72
2.1.6.1. Espectroscopia no Infravermelho Transformada de
Fourier (FTIR)
2.1.6.2. Dessulfatação da Fração Polissacarídica Sulfatada (PII)
72
73
2.1.6.3. Espectroscopia de Ressonância Magnética
Nuclear (RMN)
73
2.2. RESULTADOS E DISCUSSÃO 74
2.2.1. Extração e Rendimento dos Polissacarídeos Totais (PT) de 74

13
A. barbadensis
2.2.2. Cromatografia de Troca Iônica em Matriz de DEAE-celulose 75
2.2.3. Caracterização Físico-química 79
2.2.3.1. Análises Químicas Colorimétricas 79
2.2.3.2. Eletroforese em Gel de Poliacrilamida (PAGE) 82
2.2.3.3. Cromatografia de Troca Iônica em Coluna de
Mono-q Acoplada a HPLC
84
2.2.3.4. Eletroforese em Gel de Agarose 86
2.2.3.5. Análise da Composição Monossacarídica das
Frações PI e PII por GC-MS
88
2.2.4. Análises Estruturais 95
2.2.4.1. Espectroscopia de Absorção na Região do
Infravermelho
95
2.2.4.2. Ressonância Magnética Nuclear (RMN) 101
2.3. CONCLUSÕES 107
CAPÍTULO 3 - Avaliação das Atividades Antiviral, Prócoagulante
e Anti-hemorrágica e Toxicidade
109
3.1. MATERIAIS E MÉTODOS 110
3.1.1. CITOTOXICIDADE 110
3.1.1.1. Culturas de Células 110
3.1.1.2. Materiais Testados 111
3.1.1.3. Toxicidade das Frações para Cultura de Células 111
3.1.1.3.1. Verificação da Alteração da Morfologia Celular 111
3.1.1.3.2. Verificação da Viabilidade Celular 112
3.1.2. ATIVIDADE INIBITÓRIA SOBRE OS ADENOVÍRUS (Ad) E
OS VÍRUS HERPES SIMPLES (HSV)
112
3.1.2.1. Amostras Celulares e Virais 112
3.1.2.2. Titulação dos Vírus 112
3.1.2.3. Avaliação da Atividade Antiviral por Redução do
Título Viral
113
3.1.3. ATIVIDADE INIBITÓRIA SOBRE O METAPNEUMOVÍRUS
(HMPV) E O VÍRUS DENGUE TIPO 1 (DENV-1)
114

14
3.1.2.1. Amostras Celulares e Virais 114
3.1.2.2. Atividade antiviral 114
3.1.2.3. Extração do RNA viral 115
3.1.2.4. qRT-PCR em tempo real 115
3.1.4. HEMOSTASIA 116
3.1.4.1. Ensaios de Coagulação in vitro 116
3.1.4.1. Tempo de tromboplastina parcial ativada (TTPa) 116
3.1.4.2. Tempo de Protrombina (TP) 116
3.1.4.2. Ensaios de Coagulação ex vivo 117
3.1.4.2.1. Animais 117
3.1.4.2.2. Avaliação do Efeito Hemorrágico 117
3.1.4.2.3. Tempo de tromboplastina parcial ativada (TTPa)
ex vivo
119
3.1.4.3. Avaliação Sistêmica dos PT de A. barbadensis na
toxicidade subcrônica em camundongos.
119
3.1.4.4. Parâmetros Hematológicos e Bioquímicos 120
3.1.4.5. Análises Histológicas 120
3.1.4.6. Análises Estatísticas 121
3.1. RESULTADOS E DISCUSSÕES 122
3.2.1. Citotoxicidade e Viabilidade Celular 122
3.2.2. Atividade Inibitória Sobre os Vírus Adenovírus (Ad) e os
Vírus Herpes Simples (HSV)
126
3.2.3. Atividade Inibitória Sobre o HMPV e o DENV-1 133
3.2.4. Hemostasia 138
3.2.4.1. Atividades Prócoagulante e Anti-Hemorrágica das
Frações PI e PII
138
3.2.4.2. Avaliação Sistêmica dos PT de A. barbadensis na
toxicidade subcrônica em camundongos.
141
3.3. CONCLUSÕES 145
REFERÊNCIAS BIBLIOGRÁFICAS 146

15
LISTA DE FIGURAS
Capítulo 1 - Introdução
Figura Página
1.1 Representação esquemática da formação de ligação O-glicosídica
(condensação) proveniente da reação da hidroxila (álcool) de um
monossacarídeo com o carbono anomérico (hemiacetal) de outra
molécula de açúcar, formando um grupo acetal e liberação de uma
molécula de água. O inverso da reação é a hidrólise.
29
1.2 Fórmula de Haworth dos monossacarídeos que constituem os
polissacarídeos de parede celular. A) ácido á-D-galacturônico (á-
GalA); B) á-L-ramnose (á-Rha); C) â-D-galactose (â-Gal); D) á-L-
arabinofuranose (á-Ara); E) â-D-glucose (â-Glc); F) â-D-xilose (â-
Xil); G) â-D-manose (â-Man); H) ácido á-D-glucurônico (á-GlcA); I)
á-L-galactose (á-L-Gal); J) á-L-fucose (á-Fuc); K) á-L-
arabinopiranose (á-Ara p); L) ácido urônico D-lyxo-5-hexosulose
(Hua); M) â-D-apiose (â-Api); N) ácido á-L-acético (á-AceA); O)
ácido [2-ceto]-3-deoxi-á-D-manno-octulosônico (á-Kdo); P) ácido [2-
ceto]-3-deoxi-â-D-lyxo-heptulosárico (â-Dha); Q) Metil á-D-
galacturonato [éster] (MeGalA); R) ácido 4-O-Metil-á-D- glucurônico
[éter] (MeGlcA); S) ácido 3-O-acetil-á-D-galacturônico [éster] (3
AcGalA); T) 5-O-Feruloil-á-L-arabinose [éster] (Fer-Ara). Linha
superior, principais componentes das pectinas; 2ª linha, principais
componentes das hemiceluloses; 3ª linha, açúcares de origem
variada; 4ª linha, ramnogalactorunanos tipo II; última linha, açúcares
com substituintes não-glicídicos.
31
1.3 Modelo tridimensional de parede celular primária do tipo I mostrando
as interações moleculares entre celulose, xiloglucanas e pectinas.
33

16
Nessa figura, não está sendo mostrado gluco- e galactomananos. B)
Modelo tridimensional da parede celular do tipo II, mostrando a
interação molecular entre celulose, glucuronoarabinoxilanas,
pectinas e substâncias aromáticas.
1.4 Estrutura e nomenclatura dos principais oligossacarídeos obtidos a
partir de tratamento enzimático de xiloglucanas. Nas cadeias
principais e ramificações as setas indicam: , ligações â-(1 4); ,
ligação á-(1 6); ⇑, ligação á- (1 2).
38
1.5 Estruturas químicas das principais mananas de reserva encontradas
em tecidos de vegetais. (A) – galactomanana, sendo que só pode
ser nomeado como tal quando possuir ramificações com resíduos de
galactose superior a 10%. Setas indicam os monossacarídeos
principais, manose e galactose; (B) – glucomanana. A seta superior
indica grupo O-acetil na posição C-6. As setas inferiores indicam as
principais unidades monossacarídicas, manose e glucose. No
entanto, há substituições com resíduos de galactose (não mostrado).
41
1.6 Representação esquemática da estrutura de adenovírus (A) e
herpes simples vírus (B).
49
1.7 Representação esquemática da estrutura do HMPV e seu ciclo de
replicação viral. (1) Glicosaminoglicanos (GAGs), presentes na
membrana plasmática da célula hospedeira são reconhecidos pela
proteína de ligação G e, as integrinas ávâ1, pela proteína de fusão
F; (2) Inclusão do nucleocapsídeo viral no citoplasma celular; (3)
Síntese de RNA mensageiro viral; (4) Replicação do RNA viral; (5)
Tradução das proteínas virais [fusão (F), de ligação (G e HN), de
baixa hidrofobicidade (SH), nucleoproteína (N), fosfoproteína (P),
proteína da matriz (M) e polimerase (L)] e migração para o Retículo
Endoplasmático (RE) e Complexo de Golgi; (6) Brotamento das
53

17
partículas virais.
1.8 Vírus da Dengue. (a) Partícula viral imatura. Círculo correspondendo
a proteínas que sobressaem da superfície do capsídeo; (b)
Proteínas do envelope (E) de partícula viral madura. Cores azul,
amarelo e vermelho indicam os diferentes domínios das proteínas E.
Verde indica as junções dos domínios.
56
1.9 Representação do modelo da coagulação baseado em superfícies
celulares, compreendendo as fases de iniciação, amplificação e
propagação.
60
Capítulo 2 – Caracterização Físico-química e Estrutural
Figura Página
2.1 Fotografia do plantio de espécimes de Aloe barbadensis em canteiro
irrigado no Horto de Plantas Medicinais da UFC; (B) corte
transversal de uma folha, demonstrando a casca e a polpa interna;
(C) retirada da casca; (D) sistema de filtração a vácuo; (E)
Classificação taxonômica da espécie A. barbadensis.
66
2.2 Fluxograma de obtenção dos polissacarídeos totais (PT) de
Aloe barbadensis.
67
2.3 Cromatografia de troca iônica em coluna de DEAE-celulose dos PT
de Aloe barbadensis. As frações PI e PII foram eluídas com tampão
acetato de sódio 50 mM, pH 6,0, sem NaCl e com o mesmo tampão
contendo 0,5 M de NaCl, respectivamente. A cromatografia foi
monitorada pelo método fenol-ácido sulfúrico (DUBOIS et al., 1956).
(-♦ -) Dosagem de carboidratos totais; (-o-) concentração de NaCl
(M).
76

18
2.4 Eletroforese em gel de poliacrilamida (PAGE) dos PT e das frações
PI e PII. (A) Coloração por azul de toluidina; (B) Coloração por
stainsall. M – Padrões dos GAGs dextram sulfato de baixa massa
molecular (~8 kDa) (M1), condroitim-4-sulfato (M2), condroitim-6-
sulfato (M3) e dextram sulfato de alta massa molecular (~500 kDa)
(M4).
83
2.5 Purificação dos polissacarídeos sulfatados de Aloe barbadensis por
cromatografia de troca iônica em matriz Mono-q, acoplada a sistema
de (HPLC), em gradiente de concentração de 0,5 – 3 M de NaCl
(linha pontilhada).
85
2.6 Eletroforese em gel de agarose 0,5% da fração de polissacarídeos
eluída com Tampão acetato de sódio 50 mM, pH 6,0, contendo 0,5M
de NaCl (PII). (A) Coloração com stainsall (início do processo de
descoramento). (B) Coloração com stainsall (final do processo de
descoramento). M – Padrões dos glicosaminoglicanos (GAGs)
heparam sulfato (HS), dermatam sulfato (DS) e condroitim sulfato
(CS).
87
2.7 Análise das unidades monossacarídicas por cromatografia gasosa
acoplada a espectrometria de massa (GC-MS). (A) Derivados
alditois-acetato (D-manose, D-glucose e D-galactose) obtidos do
hidrolisado da fração PI de Aloe barbadensis; (B) espectro de massa
da D-manose; (C) Espectro de massa da D-glucose e (D) espectro
de massa da D-galactose.
90
2.8 Análise das unidades monossacarídicas por cromatografia gasosa
acoplada a espectrometria de massa (GC-MS). (A) Derivados
alditois-acetato de L-fucose obtidos do hidrolisado da fração PI de
Aloe barbadensis; (B) espectro de massa da L-fucose.
91

19
2.9 Análise das unidades monossacarídicas por cromatografia gasosa
acoplada a espectrometria de massa (GC-MS). (A) Derivados
alditois-acetato (D-manose, D-glucose e D-galactose) obtidos do
hidrolisado da fração PII de Aloe barbadensis; (B) espectro de
massa da D-manose; (C) Espectro de massa da D-glucose e (D)
espectro de massa da D-galactose.
92
2.10 Análise das unidades monossacarídicas por cromatografia gasosa
acoplada a espectrometria de massa (GC-MS). Derivados alditois-
acetato de L-arabinose (A) e D-xilose (C) obtidos do hidrolisado da
fração PII de Aloe barbadensis; (B) espectro de massa da L-
arabinose; (D) Espectro de massa da D-xilose.
93
2.11 Espectro de absorção na região do infravermelho (4000-2000 cm-1)
dos PT (linha preta) e das frações PI (linha vermelha) e PII (linha
azul) de Aloe barbadensis.
96
2.12 Espectro de absorção na região do infravermelho (2000-500 cm-1)
dos PT (A) e da fração PI (B) de Aloe barbadensis.
97
2.13 Espectro de absorção na região do infravermelho (2000-500 cm-1)
da fração PII de Aloe barbadensis.
99
2.14 Comparação dos espectros de absorção dos PT (linha preta) e das
frações PI (linha vermelha) e PII (linha azul) de Aloe barbadensis na
região do infravermelho (2000-500 cm-1).
100
2.15 Espectros 1D de 1H-RMN em 800 MHz da fração PII nativa (A) e
dessulfatada (B) de Aloe barbadensis.
102
2.16 Espectros 1D de 1H-RMN em 800 MHz dos sinais correspondentes
aos prótons anoméricos de polissacarídeos da fração PII nativa (A) e
dessulfatada (B) de Aloe barbadensis.
103

20
2.17 Espectros 2D heteronuclear 1H/13C DEPT-HSQC da fração PII nativa
de Aloe barbadensis.
105
2.18 Espectros 2D heteronuclear 1H/13C DEPT-HSQC da fração PII
dessulfatada de Aloe barbadensis.
106
Capítulo 3 – Avaliação das Atividades Antiviral, Prócoagulante e Anti-
hemorrágica e Toxicidade
Figura Página
3.1 Representação esquemática do modelo de análise do efeito
hemorrágico por tempo de sangramento. Nesse modelo, o volume
de sangue perdido é calculado a partir da concentração de
hemoglobina dissolvida na água destilada contida na proveta.
118
3.2 Fotomicrografia das células de: A – rim de macaco rhesus [Macaca
Mulatta (LLC-MK2)]; B – carcinoma de laringe humana (HEP-2); C –
carcinoma cervical uterino HeLa; D – rim de macaco verde africano
[Cercopithecus aethiops (Vero)]; E – larva de mosquito Aedes
albopictus (C6/36). (Aumento 100x)
123
3.3 Efeito citopático em células de rim de macaco verde africano
Cercopithecus aethiops (Vero) infectadas com HSV-1. (a) Células
arredondadas desprendidas da monocamada (seta); (b) Foco de
infecção em monocamada de células (círculo); (c) Célula
arredondada com tamanho superior à células normais (seta). Pode
ser observado prolongamentos citoplasmáticos; (d) Células sinciciais
gigantes (asterisco).
128
3.4 Efeito citopático de HMPV em células de rim de macaco rhesus
[Macaca Mulatta (LLC-MK2)]. Flechas indicam aglomeração celular.
136

21
3.5 Avaliação do efeito anti-hemorrágico (A) e do tempo de
tromboplastina parcial ativada (TTPa) (B) em ensaios in vivo em
ratos, após tratamento com as frações PI e PII. * p<0,05 indica
diferença significativa de PI (1,5 mg/Kg) comparada ao grupo
controle.
139
3.6 Fotomicrografia de cortes histológicos, corados com hematoxilina e
eosina, dos órgãos: coração (A e B), fígado (C e D), rim (E e F),
baço (G e H), timo (I e J) e linfonodo (K e L) de camundongos
machos Swiss tratados com salina (figuras da esquerda) e 10 mg/kg
dos PT de A.barbadensis (figuras da direita). Os resultados
referentes às fêmeas se mostraram idênticos aos machos. Aumento
400x.
144

22
LISTA DE TABELAS
Capítulo 2 – Caracterização Físico-química e Estrutural
Tabela Página
2.1 Composição química de PT e das frações PI e PII obtidos de
folhas de Aloe barbadensis.
80
2.2 Composição química de frações de espécies do gênero Aloe. 81
2.3 Composição de monossacarídeos das frações PI e PII de Aloe
barbadensis comparadas com as obtidas para as espécies A.
barbadensis e A. arborescens por outros autores.
94
Capítulo 3 – Avaliação das Atividades Antiviral, Prócoagulante e Anti-
hemorrágica e Toxicidade
Tabela Página
3.1 Efeito dos polissacaríeos totais (PT) e das frações
polissacarídicas PI e PII de Aloe barbadensis na viabilidade de
células de rim de macaco rhesus [Macaca Mulatta (LLC-MK2)],
carcinoma de laringe humana (HEP-2), carcinoma cervical
uterino (HeLa), rim de macaco verde africano [Cercopithecus
aethiops (Vero)] e de larva de mosquito Aedes albopictus
(C6/36).
124
3.2 Percentagem de inibição dos polissacarídeos totais (PT) e das
frações PI e PII de Aloe barbadensis contra os vírus HSV-1 e
129

23
HSV-2 em células de rim de macaco verde africano
Cercopithecus aethiops (Vero).
3.3 Atividade inibitória dos polissacarídeos totais (PT) e das
frações PI e PII de Aloe barbadensis sobre os vírus HSV-1 e
HSV-2 em células de rim de macaco verde africano
Cercopithecus aethiops (Vero).
132
3.4 Efeito inibitório dos polissacarídeos totais (PT) e das frações PI
e PII de Aloe barbadensis sobre o HMPV em células de rim de
macaco rhesus [Macaca Mulatta (LLC-MK2)].
134
3.5 Atividade inibitória dos polissacarídeos totais (PT) e das
frações PI e PII de Aloe barbadensis sobre o HMPV em células
de rim de macaco rhesus [Macaca Mulatta (LLC-MK2)].
135
3.6 Análise de toxicidade por dose repetida dos PT de Aloe
barbadensis em camundongos machos e fêmeas Swiss
142
3.7 Análises hematológicas de sangue de camundongos Swiss
machos e fêmeas tratados com os PT de Aloe barbadensis.
143

24
LISTA DE ABREVIATURAS
1D e 2D – uni e bidimensional
Ad – adenovírus
C6/36 – células de larva de mosquito
Aedes albopictus
CC50 – concentração citotóxica para 50%
das células em cultura
CMNT – concentração máxima não tóxica
D2O – água deuterada
DEAE – dietilaminoetil
DENV – vírus da dengue
DEPT – Distortionless Enhancement by
Polarization Transfer
DMB – azul de 1,9-dimetilmetileno
DMSO – Dimetil sulfóxido
DO – Densidade Óptica
ED50 – dose efetiva capaz de inibir 50%
do efeito citopático viral
FT – fator tissular
GAG – glicosaminoglicano
GC-MS – cromatografia gasosa acoplada
a espectrômetro de massa
H1N1 – Vírus influenza
HeLa – células de carcinoma cervical
humano
HEp-2 – células de carcinoma de laringe
humana
HIV – Vírus da imunodeficiência humana
HMPV – Metapneumovírus
HPLC – Cromatografia líquida de alto
desempenho
HSQC – Heteronuclear Single Quantum
Coherence
HSV – Vírus herpes simples
I% – percentagem de inibição
IIV - índice de inibição viral
IR – infravermelho
IS – índice de seletividade
KOH – Hidróxido de Potássio
LLC-MK2 – células de rim de macaco
rhesus (Macaca mulata)
MEM – Meio mínimo essencial
NaCl – Cloreto de Sódio
NMSO3 – lipídeo sialil sulfatado
PAGE – Eletroforese em gel de
poliacrilamida
PI – Fração PI de Aloe barbadensis não
retida em cromatografia de DEAE
PII – Fração PII de Aloe barbadensis
eluída com 0,5 M de NaCl em
cromatografia de DEAE
PNC – polissacarídeos negativamente
carregados
PT – Polissacarídeos totais de Aloe
barbadensis
qRT-PCR – Reação em cadeia da
polimerase em tempo real
RMN – Ressonância Magnética Nuclear
RSV – Vírus sincicial respiratório
SFB – Soro fetal bovino
TCA – Ácido tricloracético
TCID50 – dose infectante para 50% dos
cultivos celulares
TP - Tempo da trombina
TTPa - Teste de tromboplastina parcial
ativada
Vero - Células de rim de Cercopithecus
aethiops

25
RESUMO
Este trabalho teve como objetivos caracterizar físico-química e estruturalmentepolissacarídeos obtidos de folhas de Aloe barbadensis e avaliar suas atividadesantiviral, anti-hemorrágica e pró-coagulante e possíveis sinais de toxicidade. Foirealizada extração aquosa de polissacarídeos totais (PT) de A. barbadensis,seguido de precipitação por etanol e remoção dos contaminantes proteicos comTCA. A cromatografia em DEAE-celulose foi eficiente no fracionamento dos PT,onde foram obtidas as frações PI e PII. A caracterização físico-química mostrouque a fração PI é composta por manose (78,4%), glucose (7,3%), galactose(2,1%), fucose (2,8%) e ácidos urônicos (10,0%), e isenta de grupos éster sulfato.Enquanto, a fração PII é constituída por manose (39,2%), glucose (22,2%),galactose (26,3%), arabinose (3,8%), xilose (1,1%), ácidos urônicos (8,0%) egrupos éster sulfato (12,0%). Na revelação das bandas polissacarídicas da fraçãoPII, obtidas por PAGE e gel de agarose corados com stainsall foi constatado apresença de duas bandas que apresentaram diferentes colorações, roxa e ciana,correspondentes a presença de grupos sulfato e carboxilados, respectivamente.Na análise estrutural das frações PI e PII, por espectroscopia no IR, foidemonstrado que a fração PI apresenta unidades monossacarídicas de â-manoseO-acetiladas (812,2 e 960 onda.cm-1) e ácidos urônicos neutros (1738 onda.cm-1)em sua estrutura. Diferentemente, a fração PII, mostrou-se ser constituída porunidades monossacarídicas de manose (1014,7 onda.cm-1), galactose (1078,2onda.cm-1), ácidos urônicos carregados negativamente (1635 onda.cm-1) e éstersulfato (1329,5 e 1260,9 onda.cm-1). Na avaliação estrutural da fração PII por RMNfoi comprovado à presença do grupo éster sulfato. Em relação às atividadesbiológicas, o teste de citotoxicidade mostrou que os PT e as frações PI e PII nãoapresentaram toxicidade para a maioria das células testadas e não forameficientes na inibição de vírus não envelopados Ad-19 e Ad-41. No entanto, os PTe a fração PI foram capazes de inibir a infecção causada por HSV-1 e HSV-2. Emensaios com metapneumovírus (HMPV), a fração PII apresentou atividade antiviralsuperior à ribavirina. Embora os PT e as frações PI e PII não terem apresentadoatividade contra dengue vírus sorotipo 1 (DENV-1), os PT puderam inibir ahemorragia em ratos, diminuindo o tempo de sangramento e o tempo deprotrombina. Os PT não apresentaram toxicidade em camundongos, masaumentaram o tamanho do baço e o número de plaquetas sanguíneas. Pode serconcluído que extratos foliares de A. barbadensis podem apresentar polissacarídeosneutros ou carregados negativamente por grupos carboxilados/sulfatados. Em adição,além de apresentar atividade inibitória contra os vírus HSV-1, HSV-2 e HMPV, podemtambém apresentar efeito anti-hemorrágico e prócoagulante, propriedades essas,importantes, visto que a complicação de muitas viroses leva a quadros hemorrágicos.Além disso, os PT de A. barbadensis não apresentaram toxicidade expressiva,podendo ser utilizada como agente terapêutico seguro e eficaz.
Palavras-chave: Aloe barbadensis, polissacarídeos, vírus herpes simples,metapneumovírus e atividade anti-hemorrágica.

26
ABSTRACT
The aim of this study was to investigate the physicochemical and structuralparameters of polysaccharides obtained from Aloe barbadensis leaves and toevaluate the antiviral and cytotoxic activities and procoagulant, anti-bleedingeffects. In addition, toxicological analysis was carried out. The pulp was submittedto aqueous extraction (70 °C) and the total polysaccharides (PT) obtained byethanol precipitation, TCA was used to remove the protein contamination. Thefractionation of PT with DEAE-celulose resulted in two fractions (PI and PII). Thephysicochemical characterization showed that PI fraction presents mannose(78,4%), glucose (7,3%), galactose (2,1%), fucose (2,8%) and uronic acid (10,0%).Sulfate esters were not detected in PI fraction. On the other hand, PII fractionpresents the monosaccharides mannose (39,2%), glucose (22,2%), galactose(26,3%), arabinose (3,8%), xylose (1,1%), uronic acids (8,0%) and sulfate estersgroups (12,0%). The polysaccharidics of PII obtained by PAGE and agarose gelelectrophoresis were revealed with toluidine blue and stainsall dye showing thepresence of two different bands, one purple (indicative of sulfate) and another cyan(indicative of carboxilated groups). Structural analysis of PI and PII fractions by IRspectroscopy demonstrated that PI fraction is composed of residues of â-mannoseO-acetylated (812.2 and 960 cm-1) and uronic acids (1738 cm-1). In contrast, the PIIfraction is composed of mannose (1014.7 cm-1), galactose (1078.2 cm-1), negativelycharged uronic acids (1635 cm-1) and sulfate ester (1329.5 and 1260.9 cm-1).These results corroborate with NMR analyses that suggest the presence of sulfategroups in PII structure. The cytotoxicity evaluation showed that PT and the fractionsPI and PII did not show toxicity against most of tested cells and the same fractionswere not effective against non-enveloped virus (Ad 19 and Ad 41) inhibition.However, the PT and PI fraction were able to inhibit the infection caused by HSV-1and HSV-2. In addition, PII fraction presents antiviral activity againstmetapneumovirus. Although the PT and the fractions PI and PII did not showactivity against dengue virus serotype 1 (DENV-1), PT could inhibit the bleedingeffects in rats, reducing the bleeding and prothrombin time. The PT showed notoxicity in mice, but increased spleen size and number of blood platelets. Inconclusion, A. barbadensis leaves contain neutral or negatively chargedcarboxylated/sulfated polysaccharides. In addition, besides having inhibitory activityagainst HSV-1, HSV-2 and HMPV, they can also anti-bleeding and procoagulanteffect. Moreover, the PT of A. barbadensis showed no significant toxicity and canbe used as a safe and effective therapeutic agent.
Keywords: Aloe barbadensis, polysaccharides, herpes simplex virus,metapneumovirus and anti-bleeding effect.


28
1.1. ASPECTOS SOBRE POLISSACARÍDEOS DE VEGETAIS
SUPERIORES
Um dos grupos de macromoléculas mais abundantes no reino vegetal é
o dos carboidratos, que existem sob a forma de monossacarídeos,
oligossacarídeos, polissacarídeos e seus derivados (ROBYT, 2001).
Teoricamente, cada monossacarídeo, cetona ou aldeído polihidroxilado, pode
ocorrer sob duas formas isoméricas distintas (enantiômeros, D- ou L-). Já os
oligo- e polissacarídeos são formados por várias unidades desses monômeros
unidos por ligações O-glicosídicas. Essas ligações são formadas quando uma
hidroxila (OH–) de um monossacarídeo reage com o átomo de carbono
anomérico de outra molécula de açúcar (hemiacetal) formando um acetal e
liberando H2O (Figura 1.1) (NELSON; COX, 2008). As ligações entre resíduos
ao longo da cadeia polissacarídica apresentam duas formas adicionais de
estereoisômeros (anômeros, á ou â) definidos pela orientação da ligação entre
o átomo de carbono anomérico e o átomo de oxigênio que liga os dois
açúcares (FRY, 2011).
Dessa forma, existe uma grande variedade de polissacarídeos que são
encontrados em diferentes plantas e em diferentes tecidos/órgãos, como
folhas, sementes, raízes, tubérculos etc. Esses polímeros são muito mais
complexos que as proteínas, lipídeos e ácidos nucléicos, já que são as únicas
macromoléculas capazes de se ligar com outra através de diferentes ligações
(1 2, 1 3, 1 4 e 1 6) em configurações diferentes, á ou â. Em adição, a
cadeia principal do polímero pode apresentar ramificações, aumentando mais
ainda o grau de complexidade (NI; YATES; TIZARD, 2004). De acordo com sua
função, os polissacarídeos podem ser classificados em dois grandes grupos, os
estruturais e os de reserva. Apesar de que essa definição ainda é muito
discutida, os polissacarídeos de reserva estão relacionados no provimento de
monossacarídeos que serão utilizados durante o crescimento e
desenvolvimento da planta, enquanto os estruturais, com raras exceções, são
encontrados nas paredes celulares, que envolvem as células vegetais
(AVIGAD; DEY, 1997).

29
Figura 1.1: Representação esquemática da formação de ligação O-glicosídica(condensação) proveniente da reação da hidroxila (álcool) de ummonossacarídeo com o carbono anomérico (hemiacetal) de outra molécula deaçúcar, formando um grupo acetal e liberação de uma molécula de água. Oinverso da reação é a hidrólise. Fonte: Nelson e Cox (2010).

30
1.1.1. Polissacarídeos de parede celular vegetal
Enquanto que as células animais apresentam uma matriz extracelular de
carboidratos na superfície de sua membrana plasmática (glicocálice), as
células vegetais apresentam uma estrutura adicional, a parede celular, que
confere não só resistência mecânica, mas também está envolvida no
movimento da água, defesa contra patógenos, crescimento, diferenciação e
comunicação celular (COSGROVE, 2005). Os vegetais apresentam,
aproximadamente, 35 tipos celulares que contêm esse envoltório, com exceção
dos gametas (MELLINGER, 2006). Ele está presente em todos os estágios de
desenvolvimento da célula, se comportando como estrutura dinâmica que se
altera no decorrer da vida celular (CARPITA; Mc CANN, 2000).
Em Angiospermas (plantas que apresentam sementes envolvidas por
frutos) podem ser observados dois tipos distintos de parede celular, a primária
e a secundária. Segundo o modelo clássico, a parede celular primária (PCP) é
encontrada em células jovens sendo constituída por duas fases, uma
microfibrilar e outra matricial (FRY, 2011). A fase microfibrilar é composta por
celulose, um polímero altamente insolúvel em água constituído por resíduos de
glucose â(1 4) ligados. Quando observada através de microscopia eletrônica,
é percebido que essa fase é bastante homogênea sendo distinguida da fase
matricial pelo alto grau de cristalinidade. A fase matricial é constituída por
outros polissacarídeos, pectinas e hemiceluloses, glicoproteínas e compostos
fenólicos (O’NEILL; YORK, 2003).
Como a fase matricial é bastante heterogênea, Carpita e Gibeaut (1993)
propuseram dois modelos estruturais para PCP de angiospermas com base
nas diferenças entre as pectinas e hemiceluloses. Atualmente, os modelos
ainda são utilizados, subdividindo as PCP em tipo I e tipo II (FRY, 2011). Em
adição, os monossacarídeos que constituem os polissacarídeos presentes na
fase matricial da PCP estão resumidos na Figura 1.2.

31
Figura 1.2: Fórmula de Haworth dos monossacarídeos que constituem ospolissacarídeos de parede celular. A) ácido á-D-galacturônico (á-GalA); B) á-L-ramnose (á-Rha); C) â-D-galactose (â-Gal); D) á-L-arabinofuranose (á-Ara); E)â-D-glucose (â-Glc); F) â-D-xilose (â-Xil); G) â-D-manose (â-Man); H) ácido á-D-glucurônico (á-GlcA); I) á-L-galactose (á-L-Gal); J) á-L-fucose (á-Fuc); K) á-L-arabinopiranose (á-Ara p); L) ácido urônico D-lyxo-5-hexosulose (Hua); M) â-D-apiose (â-Api); N) ácido á-L-acético (á-AceA); O) ácido [2-ceto]-3-deoxi-á-D-manno-octulosônico (á-Kdo); P) ácido [2-ceto]-3-deoxi-â-D-lyxo-heptulosárico(â-Dha); Q) Metil á-D-galacturonato [éster] (MeGalA); R) ácido 4-O-Metil-á-D-glucurônico [éter] (MeGlcA); S) ácido 3-O-acetil-á-D-galacturônico [éster] (3AcGalA); T) 5-O-Feruloil-á-L-arabinose [éster] (Fer-Ara). Linha superior,principais componentes das pectinas; 2ª linha, principais componentes dashemiceluloses; 3ª linha, açúcares de origem variada; 4ª linha,ramnogalactorunanos tipo II; última linha, açúcares com substituintes não-glicídicos. Fonte: Fry (2011)

32
As PCP do tipo I estão presentes nas dicotiledôneas e nas
monocotiledôneas não comelinóides. As comelinídeas compõem as
monocotiledôneas pertencentes às ordens Arecales, Commelinales, Poales e
Zingiberales. Assim, as PCP do tipo I apresentam fase matricial constituída,
principalmente, por xiloglucanas, polímeros bastantes extensos formados, em
sua maioria, por resíduos de xilose, que se ligam fortemente às microfibrilas de
celulose (Figura 1.3 A). Os componentes dessa interação compreendem cerca
de 50% da parede celular e estão embebidos por uma matriz de pectinas e
compostos pécticos (20-35 %). Em adição, outras hemiceluloses,
glicoproteínas e compostos fenólicos são encontrados nas PCP de tipo I, em
menor quantidade (HOFFMAN et al., 2005).
As PCP de tipo II são caracterizadas por conter baixa quantidade de
pectina e xiloglucanos, e as microfibrilas de celulose estão interligadas,
principalmente, por hemiceluloses do tipo glucuronoarabinoxilanas. Esse tipo
de PCP está ausente nas dicotiledôneas e presentes nas monocotiledôneas
comelinóides. A quantidade de proteínas estruturais é menor que nas PCP do
tipo I e os polissacarídeos formam ligações cruzadas com compostos fenólicos.
As hemiceluloses presentes nas PCP tipo II são formadas por uma mistura de
â-D-glucanas ligados tanto por ligações (1 3) quanto por (1 4) (GIBEAUT et
al., 2005) (Figura 1.3 B).
Em algumas células especializadas, que requerem alta força mecânica e
reforço estrutural, a parede celular é apresentada muito mais espessa devido à
deposição de celulose, hemicelulose e lignina. Essas células entram em um
processo de lenhificação que se inicia desde a região da PCP se estendendo
para o interior da célula formando a parede celular secundária. A deposição de
lignina e formação de parede celular secundária interrompe o crescimento
celular e desencadeia o amadurecimento e diferenciação terminal celular
(ALBERSHEIM et al., 1996; COSGROVE, 2005). A constituição química dessa
parede é de, aproximadamente, 40-45 % de celulose, 15-35 % de
hemicelulose, 15-30 % de lignina e traços de pectina (MELLINGER, 2006).
Considerando a relevância com a qual os polissacarídeos da PCP apresentam
nesse trabalho, os próximos subitens trazem uma breve revisão sobre os
mesmos.

33
Figura 1.3: A) Modelo tridimensional de parede celular primária do tipo I
mostrando as interações moleculares entre celulose, xiloglucanas e pectinas.
Nessa figura, não está sendo mostrado gluco- e galactomananos. B) Modelo
tridimensional da parede celular do tipo II, mostrando a interação molecular
entre celulose, glucuronoarabinoxilanas, pectinas e substâncias aromáticas.Fonte: Carpita e Mc Cann, 2000.

34
1.1.1.1. Celulose
A celulose é sintetizada por um complexo enzimático chamado celulose
sintase (CESA) que está inserido na membrana plasmática em unidades
hexaméricas chamadas de rosetas. Cada microfibrila de celulose é formada do
agrupamento espontâneo e da cristalização de dúzias de cadeias de â-D-
glucanas (1,4)-ligadas. É possível que hemiceluloses, como as xiloglucanas,
possam estar anexadas às microfibrilas de celulose como observado na Figura
1.3 (COSGROVE, 2005; HARRIS; SMITH, 2006).
De acordo com O’Looney e Fry (2005) a celulose é relativamente
resistente a hidrólises com ácido trifluoroacético (TFA) 2M a 120 °C por 1h,
embora uma pequena fração possa ser liberada como glucose, principalmente
se houver pré-tratamento com álcalis fortes. No entanto, hidrólise ácidas
utilizando H2SO4 a 120 °C com subsequente neutralização com BaCO3, pode
atingir uma taxa de conversão de celulose à glucose superior ao procedimento
anterior.
1.1.1.2. Polissacarídeos da fase matricial da parede celular
Diferente da fase microfibrilar, que é composta por celulose, os
polissacarídeos da matriz apresentam grande variedade de resíduos de açúcar
e ligações glicosídicas. Os genes que codificam para CESA pertencem a uma
superfamília chamada de Cellulose synthase-like (CSL). Oito famílias de genes
CSL (CSLA até CSLH) foram caracterizadas e apresentam sequências
características de â-glicosiltransferases. As enzimas codificadas por esses
genes são as melhores candidatas para a síntese dos polissacarídeos da
matriz, que ocorre no complexo de Golgi e, não, na membrana celular
(RICHMOND; SOMERVILLE, 2000; SCHEIBLE; PAULY, 2004). De acordo com
o processo de extração, os polissacarídeos da matriz podem ser classificados
como substâncias pécticas ou hemiceluloses.

35
1.1.1.2.1. Substâncias pécticas
As substâncias pécticas fazem parte de um grupo heterogênico de
polissacarídeos de estrutura acídica que são extraídos com agentes quelantes,
água quente ou ácidos diluídos. Suas propriedades ácidas estão relacionadas
à presença de resíduos de ácido galacturônico (GalA) ligados na cadeia por
á(1 4) (POPPER, 2008; WILLATS; KNOX; MIKKELSEN, 2006). Os resíduos
de GalA podem ser encontrados na sua forma metil-esterificada, formando
polímeros de pectinas, ou na forma desesterificada, ácidos pécticos. Ambos
podem apresentar associações com polissacarídeos neutros formando
polímeros que variam entre 50 a 150 KDa (COSGROVE, 2005).
Segundo Fry (2011), os polissacarídeos pécticos podem ser
classificados em quatro classes: homogalacturonanas, ramnogalacturonanas
do tipo I (RG I), ramnogalacturonanas do tipo II (RG II) e xilogalacturonanas.
As homogalacturonanas apresentam uma cadeia linear constituída por
resíduos de GalA não-esterificados e metil-esterificados (MeGalA). Os resíduos
de MeGalA formam polímeros neutros ao longo da cadeia de
homogalacturonanas que, provavelmente, são formadas a partir da ação
progressiva de metilesterases. Pode também ser observado a presença de
grupamentos O-acetil substituindo os C-2 ou C-3, que influenciam na
solubilidade da molécula (ISHII, 1997). Comercialmente, esses polissacarídeos
são extraídos usando solução aquosa de ácido mineral, geralmente o ácido
nítrico, a pH de 1-3, temperatura de 50-90 °C por 3-12 horas. Após a solução
ter sido concentrada, os polissacarídeos são precipitados por etanol ou 2-
propanol (HARRIS; SMITH, 2006).
As RG I são heteropolissacarídeos com cadeia principal composta por
repetições do dissacarídeo [ 4)-á-D-GalA-(1 2)-á-L-Rha-(1 ]. Juntamente
com as homogalacturonanas, formam os dois principais constituintes da matriz
péctica de dicotiledôneas e monocotiledôneas não-comelinóides (CARPITA;
GIBEAUT, 1993; MAY, 2000; WILLATS; KNOX; MIKKELSEN, 2006). Ao longo
da cadeia, os resíduos de á-L-Rha são substituídos no C-4 por, geralmente,

36
unidades de arabinose e/ou galactose. Dessa forma, as RG I são classificadas
em arabinanas, galactanas e arabinogalactanas (CIPRIANI, 2004). Além
desses substituintes, podem ser encontrado resíduos de á-Fuc, á-GluA,
MeGluA e Fer-Ara (ver Figura 1.2) (MELLINGER, 2006). As RG II diferem das
RG I por possuírem em suas cadeias principais monossacarídeos raros como
apiose, 2-O-metil-fucose, 2-O-metil-xilose, ácido acérico, entre outros (JÚNIOR,
2006).
As xilogalacturonanas são polímeros pouco estudados, encontrados em
monocotiledôneas e constituídos por resíduos de á-GalA ligados por (1 4).
Esses resíduos são substituídos majoritariamente por unidades de xilose e, em
menor quantidade por fucose e apiose (FRY, 2011).
1.1.1.2.2. Hemiceluloses
Apesar da dificuldade conceitual do termo hemicelulose, o mesmo é
utilizado para designar polissacarídeos presentes em tecidos vegetais que
ocorrem juntamente com as microfibrilas de celulose, através de pontes de
hidrogênio, e que podem ser isolados através de extrações aquosas ou
soluções alcalinas de base forte, como KOH ou NaOH (GABRIELLI et al.,
2000). Apesar da semelhança com a celulose, as ramificações em sua
estrutura previnem a formação de microfibrilas de origem hemicelulósica
(JARVIS, 2009). A composição química, abundância e distribuição desses
polímeros dependem da espécie, tecido, condições de crescimento, idade da
planta e protocolo de extração utilizado (KITAYAMA et al., 2000; MELLINGER,
2006).
Em adição aos monossacarídeos hemicelulósicos descritos na Figura
1.2 E-H, as hemiceluloses também podem apresentar resíduos de D-galactose,
L-arabinose, ácido 4-O-metilglucurônico e ácido D-galacturônico. As principais
hemiceluloses encontradas em paredes celulares da maioria das plantas são
as xiloglucanas, xilanas, glucuronomananas e mananas (CUI & WANG, 2009;
FRY, 2011; MOLLER et al., 2007; ).

37
As xiloglucanas são as hemiceluloses mais abundantes e, assim como a
celulose, sua cadeia principal é constituída por unidades de (1 4)-â-D-Glcp.
Grande parte desses resíduos é ramificada em C-6 por resíduos de á-D-Xylp,
sendo que as unidades de xilose são substituídas por resíduos de â-D-Galp ou
á-L-Fucp no C-2 (SILVA, 1993). As xiloglucanas podem ser hidrolisadas por
endo-(1 4)-â-D-glucanase (celulase) ou por endoglucanases xiloglucana-
específicas (XEG). Para ambas as enzimas, a clivagem se dá no mesmo sítio,
ou seja, nos resíduos de glucose não substituídos (FRY, 2011; HARRIS;
SMITH, 2006). Na Figura 1.4 pode ser observada a estrutura simplificada e
nomenclatura dos principais oligossacarídeos obtidos a partir de tratamento
enzimático de xiloglucanos.
O segundo tipo de hemicelulose mais abundante é o das xilanas, que
tem uma cadeia principal composta por resíduos de (1 4)-â-Xil. As unidades
de xilose são substituídas, frequentemente, por á-Ara, á-GlcA e á-MeGlcA no
C-2. Dependendo da proporção dos seus substituintes as xilanas podem
receber nomenclaturas variadas como arabinoxilanas, glucuronoxilanas etc.
(COSGROVE, 2005) Esses polissacarídeos são encontrados, principalmente,
na parede celular secundária dos tecidos formando pontes de hidrogênio com a
celulose, ligações covalentes com as ligninas e ligações ésteres com
grupamento acetil e ácidos fenólicos (DINAND; VIGNON, 2001). Os grupos O-
acetil também estão presentes, especialmente nos resíduos de xilose (FRY,
2011).
As glucuronomananas geralmente são classificadas como mucilagens e
são completamente distintas das mananas que serão descritas a seguir. Esses
polissacarídeos apresentam em sua cadeia principal repetições do
dissacarídeo [-4)-â-GlcA-(1 2)-á-Man-(1 ], sendo que, alternadamente, são
encontrados resíduos de â-Man ao invés de á-Man (FRY, 2011). Na posição C-
3 das unidades de â-GlcA e á-Man, geralmente, estão anexadas cadeias
laterais ricas em á-Ara, â-Gal, á-Fuc e, em algumas vezes, â-Xil (SILVA, 1993).
Devido à importância das mananas para o presente trabalho, esses
polissacarídeos foram descritos no item subsequente.

38
Figura 1.4: Estrutura e nomenclatura dos principais oligossacarídeos obtidos a
partir de tratamento enzimático de xiloglucanas. Nas cadeias principais e
ramificações as setas indicam: , ligações â-(1 4); , ligação á-(1 6); ⇑,
ligação á- (1 2). Fonte: Fry et al., 1993.

39
1.1.1.2.3. Polissacarídeos de reserva da parede celular vegetal
No mesmo tecido vegetal, uma população de polissacarídeos pode ser
classificada hora como de “reserva” e hora como “estrutural”. Essa definição
ainda não é bem estabelecida, visto que a maioria dos polissacarídeos
estruturais é submetida a processos de hidrólise e despolimerização para o
crescimento celular (AVIGRAD; DEY, 1997). Uma vez despolimerizados, os
polissacarídeos remanescentes do processo de hidrólise podem ser utilizados,
transitoriamente, como materiais de reserva em estágios fisiológicos
específicos. Dentre outros polissacarídeos que apresentam essa dupla
funcionalidade é destacada as mananas, que apresentam alta importância
econômica para a indústria química, farmacêutica e alimentícia (NISHINARI et
al., 2007).
Além disso, as mananas são consideradas hemiceluloses
mucilaginosas. Como descrito anteriormente, as hemiceluloses são formadas
por polissacarídeos extraíveis por soluções aquosas ou álcalis. A mucilagem é
definida como qualquer substância de alta massa molar que confira aspecto
coloidal quando esteja em solução. Essas substâncias podem ser encontradas
em todos os tecidos da planta como produtos pré-formados, ou seja, são
sintetizadas pelo metabolismo normal do vegetal, não necessitando que a
mesma sofra injúria ou ataque de patógenos para serem produzidas (JANI et
al., 2009). As mananas de parede celular vegetal podem ser subdivididas em:
mananas puras, glucomananas, galactomananas e galactoglucomananas
(AVIGRAD; DEY, 1997; BUCKERIDGE; SANTOS; TINÉ, 2000; MATHUR;
MATHUR, 2005; NISHINARI et al., 2007).
As mananas puras são encontradas tanto em dicotiledôneas como em
monocotiledôneas. Esses polissacarídeos são mais abundantes em sementes,
sendo uma importante fonte de reserva durante o processo de germinação.
Além disso, podem ser encontradas em parênquimas de reserva foliar de
plantas suculentas (NI; YATES; TIZARD, 2004). São constituídas, em mais de
90%, por resíduos de â(1 4)-D-mananopiranose e, em minoria, por
ramificações formadas por unidades de á(1 6)-galactose (AVIGRAD; DEY,

40
1997). Ou seja, são estruturalmente relacionados às galactomananas, sendo
estas mais ramificadas, chegando apresentar mais de 50% de resíduos de
galactose em sua estrutura (Figura 1.5 A). Diferente da celulose, esses dois
tipos de polissacarídeos apresentam certo grau de solubilidade variando de
acordo com o seu grau de ramificação.
As galactomananas são amplamente distribuídas na natureza e
apresentam considerável atenção no meio acadêmico e industrial, visto que
são consideradas importantes agentes espessantes e geleificantes em meio
aquoso (LIMA, 2000). Os genes que codificam as enzimas que adicionam
resíduos de açúcar na cadeia de polimanose estão sendo amplamente
estudados. É sabido que a síntese de galactomananas é realizada pela
combinação de dois eventos: primeiro, a ação de uma manana sintase, que
adiciona resíduos de manose â(1 4)-ligados e, em segundo lugar, a atuação
de uma á-galactosiltransferase, que adiciona unidades de galactose na cadeia
principal de manana (NISHINARI et al., 2007). Em contraste, a mobilização de
galactomananos no tecido vegetal só é possível devido à atuação de enzimas
do tipo á-galactosidase, endo â-mananases e â-manosidases (BUCKERIDGE;
SANTOS; TINÉ, 2000; MATHUR; MATHUR, 2005).
As glucomananas (Figura 1.5 B) apresentam, em sua estrutura, resíduos
â-glicosil alternados com â-manosil. Essa cadeia principal pode apresentar
resíduos substituídos por unidades de á-galactosil (1 6)-ligados e,
eventualmente, O-acetilados no C-6 (NISHINARI et al., 2007; SINGH;
SHELLEY, 2007). A enzimologia de degradação envolve a ação de esterases
(que removem o grupo O-acetil), endo-â-manosidases, exo-â-manosidases e â-
glucosidases. Ao passo que as glucomananas se tornam mais ramificadas, são
chamadas de galactoglucomananas.

41
Figura 1.5: Estruturas químicas das principais mananas de reserva
encontradas em tecidos de vegetais. (A) – galactomanana, sendo que só pode
ser nomeado como tal quando possuir ramificações com resíduos de galactose
superior a 10%. Setas indicam os monossacarídeos principais, manose e
galactose; (B) – glucomanana. A seta superior indica grupo O-acetil na posição
C-6. As setas inferiores indicam as principais unidades monossacarídicas,
manose e glucose. No entanto, há substituições com resíduos de galactose
(não mostrado). Fonte: Lima, 2000.

42
1.1.2. Polissacarídeos sulfatados
Os polissacarídeos sulfatados compreendem um grupo complexo e
heterogêneo formado por unidades monossacarídicas carregadas
negativamente devido à presença de radicais sulfatos. Muitos estudos mostram
que esses polímeros aniônicos podem ser encontrados em vertebrados (SENNI
et al., 2011), invertebrados (VILELA-SILVA et al., 2002) e, principalmente, em
algas marinhas (JIAO et al., 2011). Contudo, salvo raros relatos, esses
polissacarídeos não têm sido encontrados em plantas vasculares terrestres
(AQUINO; GRATIVOL; MOURÃO, 2011; FRY, 2011). Dentre os
polissacarídeos de algas, organismos evolutivamente próximos às plantas
vasculares, podem ser destacados as galactanas sulfatadas, fucanas e
arabino-galactanas.
As galactanas sulfatadas estão presentes, principalmente, em algas
vermelhas (Rodophyta) e compreendem polissacarídeos formados por
unidades dissacarídicas repetitivas de â(1 3)-D-galactopiranose (unidade A) e
á(1 4)-galactopiranose (unidade B) que se alternam ao longo da cadeia
principal (JIAO et al., 2011). Essas unidades podem apresentar diversas
variações originadas de substituições das hidroxilas. As substituições mais
frequentes são de grupos O-sulfato, metil-ésteres, acetal de ácido pirúvico e
glicosilações (VAN DE VELDE et al., 2002). As galactanas são classificadas de
acordo com a variação esterioquímica da unidade B, podendo a mesma
apresentar configuração D, classificadas como carragenanas, ou L,
classificadas como agaranas (ZIBETTI, 2005). Esses polissacarídeos podem
ser classificados de acordo com outras características estruturais como padrão
e grau de sulfatação e, além disso, presença ou ausência de 3,6-anidro-á-
galactopiranose na unidade B (CAMPO et al., 2009).
As fucanas sulfatadas, ou fucoidanas, são polissacarídeos ricos em
resíduos de fucose e alginato e estão presentes em algas pardas (Phaeophyta)
e equinodermos marinhos (SENNI et al., 2011). Além disso, apresentam em
sua estrutura os monossacarídeos xilose, galactose, manose, glucose e ácidos
urônicos. As arabino-galactanas são encontradas em algas verdes

43
(Chlorophyta) e se apresentam como heteropolissacarídeos bastante
ramificados, sendo compostos, principalmente, por unidades de galactose e
arabinose (RODRIGUES, 2011).
As Angiospermas apresentam mais características em comum com
algas da ordem das Charophytas (Charales e Coleochaetales) que são
endêmicas de águas fluviais e de ambientes terrestres. De acordo com Fry
(2011), atualmente, esse grupo é classificado como táxon único que
compreende organismos que possuem polissacarídeos não sulfatados e ricos
em ácidos urônicos. Embora não sejam observados polissacarídeos sulfatados
em plantas terrestres e de água doce, Aquino et al. (2005; 2011) mostraram
que há correlação entre salinidade e biossíntese de polissacarídeos
negativamente carregados. Em plantas que são intolerantes a alta salinidade
(glicófitas), há uma produção de polissacarídeos ricos em ácidos urônicos
quando submetidas a essa condição. No entanto, para plantas tolerantes
(halófitas), os polissacarídeos sintetizados apresentaram, além dos ácidos
urônicos, grupos sulfato em suas estruturas.
1.1.3. O gênero Aloe
O gênero Aloe é constituído por plantas pertencentes à família Aloaceae
e compreende mais de 300 espécies amplamente adaptadas a condições
áridas. São plantas xerófitas e suculentas que apresentam tecidos/órgãos
capazes de acumular água e mantê-la na forma de mucilagem. Isso é possível
já que suas folhas apresentam cutícula espessa recoberta por cera, estômatos
localizados na base da cavidade supraestomatal e realizam o metabolismo do
ácido das crassuláceas (CAM), evitando assim a perda de água em seus
tecidos (NEWTON, 2004; NI et al., 2004).
A Aloe barbadensis Miller [sinonímia científica: Aloe vera (L.) Burm. F.,
A. indica Royle, A. chinensis Bak., A. elongata Murray, A. officinalis Forsk., A.
perfoliata L.; nomenclatura vernácula: babosa (Brasil), erva-babosa, sábila

44
(América Latina), aloés de Curaçao (Alemanha/Inglaterra), aloés (França),
ghrita kumari (India), Iu hui ye (China)] é a espécie mais estudada da família
Aloaceae (CAMPESTRINI, 2007; SUNG, 2006). É uma planta arbustiva, dióica,
rizomatosa, entouceirada, acaulescente, perene, com altura que varia de 0,50 a
1,20 m. As folhas apresentam comprimento médio de 55 cm, largura entre 6 a
9 cm e espessura de 3 cm, arranjadas em roseta apresentando espinhos
triangulares marginais (CAMPESTRINI, 2007). Quando submetidas a corte
transversal, pode ser observada uma camada de células epidérmicas, que
compõem a casca, e a polpa, que corresponde ao parênquima de reserva do
mesófilo foliar (NI et al., 2004). Segundo Femenia et al. (1999), a polpa,
também chamada de gel mucilaginoso, possui aspecto vítreo, e é constituído
majoritariamente por água (98,5 – 99,5 % de matéria fresca). Em adição, é
observada a presença de células associadas aos feixes vasculares, que
liberam um exsudato amarelado, rico em compostos fenólicos, quando
rompidos por alguma injúria (NEWTON, 2004).
A. barbadensis é mais conhecida pela sua sinonímia A. vera, na qual a
maioria das pessoas associa a planta a determinados produtos cosméticos
para pele e cabelos. Dentre as espécies do gênero Aloe, a A. barbadensis é a
mais explorada pela indústria da cosmetria e farmacêutica (REYNOLDS;
DWECK, 1999). Essa planta já foi citada como planta da imortalidade pelos
egípcios a 1000 A.C., mas só realmente foi documentada como medicinal, pela
primeira vez, no “Herbário Grego”, escrito por Dioscorides no primeiro século
D.C. Dessa forma, A. barbadensis é tida como milenar por já ter sido utilizada
por várias civilizações: egípcia, fenícia, suméria, romana, grega, indiana,
chinesa e alemã (PEUSER, 2003; SUNG, 2006).
Apesar de ter sido esquecida durante a primeira guerra mundial, devido
ao bloqueio continental do comércio, ela ressurgiu depois da segunda grande
guerra, sendo alvo principal de grandes grupos de pesquisa (PEUSER, 2003).
Dentre estes, pode ser destacado o grupo coreano da Seoul National
University, no qual possuem um projeto bem elaborado – Creation of Aloe
Pharmaceuticals Project (CAP Project) – e a companhia Aloecorp (Aloe
Corporation), que é sediada nos Estados Unidos, que possui suas plantações

45
no Texas e Tampico (México) (LEE, 2006). Estes grupos são responsáveis pela
maior parte da pesquisa feita com A. barbadensis mundialmente.
1.1.4. Polissacarídeos de A. barbadensis e propriedades biológicas
relacionadas
A. barbadensis é uma planta conhecida milenarmente por apresentar
inúmeras atividades biológicas/farmacológicas (HABEEB et al., 2007;
SERRANO et al., 2006). Dentre estas podemos destacar sua atividade anti-
inflamatória, analgésica, hipoalergênica, cicatrizante, antitumoral, bactericida,
antileishmaniose, imunomoduladora e, dentre outras, antiviral (HAMMAN, 2008;
REYNOLDS;DWECK, 1999).
O gel de A. barbadensis é proveniente do extrato das células
parequimáticas do mesófilo foliar. Esse gel possui mais de 300 substâncias
bioativas e é constituído, principalmente, por água e polissacarídeos (celulose,
pectinas, hemiceluloses, glucomananas, glucomananas acetiladas,
galactogalacturanas, glucogalactomananas, galactoglucoarabinomananas e
mananas puras acetiladas). Além disso, compostos fenólicos, terpenóides,
antraquinona, aminoácidos, vitaminas e sais minerais estão presentes no gel
(FEMENIA et al., 1999; HAMMAN, 2008; NI; YATES; TIZARD, 2004; PEUSER,
2003).
Muitos relatos mostram que as glucomananas são os polissacarídeos
mais abundantes em todo o gênero Aloe. No entanto, estudos provam que
outros polissacarídeos tais como pectinas, glucanas, galactanas, arabinanas,
galactoglucorunanas, podem ser detectados, compondo muitas vezes a fração
majoritária (ESUA; RAUWALD, 2006; McCONAUGHY et al., 2008; NI; YATES;
TIZARD, 2004; PUGH et al., 2001; STRICKLAND; PELLEY, 2004). De acordo
com Ni, Yates e Tizard (2004), a razão para tal discrepância se deve a
mudanças climáticas e/ou diferenças na localização geográfica.

46
O maior volume de artigos publicados está relacionado à acemanana,
glucogalactomanana linear composto por resíduos manosil β(1à 4)-ligados,
com o carbono 2 ou 3 acetilados e com algumas cadeias possuindo resíduos
de galactose ligados ao carbono 6 (CHOW et al., 2005; FEMENIA et al., 2003;
HAMMAN, 2008; IM et al., 2005; LEUNG et al., 2004). Esse polissacarídeo é
relacionado à maioria das respostas biológicas conhecidas do gel mucilaginoso
de A. barbadensis.
Acemanana é definido como um imunoestimulante demonstrando
atividade adjuvante na produção de anticorpos específicos. Quando
administrado intraperitonealmente em camundongos com câncer, esse
polissacarídeo é capaz de curar ou reduzir significativamente os tumores. Além
disso, o mesmo tem sido efetivo no tratamento de fibrossarcoma em felinos e
caninos (REYNOLDS; DWECK, 1999). A atividade antitumoral da acemanana é
mediada via ativação de mecanismos de defesa do hospedeiro e não por
apresentar efeito citotóxico nas células tumorais. Algumas das atividades
imunoestimulantes da acemanana têm sido mediadas através da ativação de
monócitos/macrófagos (LEE et al., 2001). Na presença de INFγ, o
polissacarídeo induz a ativação destas células, aumenta sua atividade
fagocítica e, subsequentemente, eleva a produção de óxido nítrico (NO) pelos
macrófagos induzindo-os a desencadear um processo de apoptose. Estas
atividades têm sido relacionadas à presença de receptores de manose
presentes nos macrófagos (CHOW et al., 2005; LEUNG et al., 2006,
REYNOLDS; DWECK, 1999).
Em adição, a manana acetilada de A. barbadensis apresenta efeito
antiviral. É mostrado na literatura que esse polímero quando administrado com
aciclovir tem efeito sinergístico no combate à infecção desencadeada pelo vírus
herpes simples (HSV). Em ensaios similares este polissacarídeo tem sido
utilizado com sucesso como adjuvante contra poliomavírus. Além disso,
apresenta efeitos benéficos significantes em felinos contaminados com o vírus
da imunodeficiência felina (FIV) e inibe a replicação do vírus causador da
síndrome da imunodeficiência humana (HIV). É sabido que acemanana
aumenta a viabilidade de linfócitos infectados com HIV-1 e reduz a carga viral.
É também relatado o efeito sinergístico que a mesma molécula tem com o

47
azidotimidina (AZT) e seu efeito tóxico em citomegalovírus (CHINNAH et al.,
1992; FRIEDMAN; SPECTER; BENDINELLI, 2006; HAMMAN, 2008;
MONTANER et al., 1996; PULSE; UHLIG, 1990; SAOO et al., 1996; SHARMA;
KARACA; PERTILE, 1994; SYED et al., 1997).
Existem muitas evidências in vitro e in vivo que polissacarídeos
carregados negativamente exercem atividade antiviral (CAMPO et al., 2009;
JIAO et al., 2011; KIM et al., 2000; MIAO et al., 2004; SENNI et al., 2011).
Essas atividades têm sido relacionadas às suas características aniônicas.
Moléculas com maior grau de sulfatação, por exemplo, podem inibir os
primeiros estágios de multiplicação viral, como a adsorção e penetração (EO et
al., 1999; JIAO et al., 2011). Muitos relatos mostram a eficiência que estes
polissacarídeos possuem contra o vírus herpes simples (HSV), vírus da
imunodeficiência humana (HIV), vírus influenza (H1N1) e vírus da dengue
(DENV) (ARENA et al., 2006; CAMPO et al., 2009; EO et al., 1999; LIU et al.,
2004; MIAO et al., 2004; SENNI et al., 2011; TALARICO; DAMONTE, 2007;
ZHU et al., 2006).
1.2. VIROLOGIA
Os vírus são parasitas intracelulares obrigatórios que usam a maquinaria
celular para síntese dos componentes virais. Sua estrutura é organizada de
forma simples: material genético consistindo, basicamente, por DNA ou RNA,
de fita simples ou dupla, linear ou circular; cobertura proteica (capsídeo) e, em
muitos grupos, a presença de envelope, camada lipídica que envolve o
capsídeo (WHITE; FENNER, 1994). Ou seja, algumas famílias virais são
envelopadas e outras apresentam apenas o capsídeo (não envelopadas). Em
adição, vírus de RNA podem apresentar o material genético orientado de forma
que podem ser traduzidos diretamente pelos ribossomos celulares (RNA +), ou
orientado de forma inversa (RNA -). Apesar dos vírus não realizarem processos
metabólicos, muitas partículas virais podem albergar enzimas, que são
importantes durante o processo de infecção (RÁCZ; MENCK, 2005).

48
1.2.1. Adenovírus
Os vírus causadores das adenoviroses humanas são divididos em sete
espécies, seguindo nomenclatura de A a G, baseada nas propriedades
imunológicas, biológicas e bioquímicas, totalizando 52 sorotipos (JONES et al.,
2007). São vírus não envelopados, têm 70 a 90 nm de diâmetro e simetria
icosaédrica. O genoma é constituído por uma única molécula de DNA de fita
dupla (dsDNA) que codifica para, aproximadamente, 40 polipeptídeos
diferentes, dos quais 30% representam proteínas estruturais (Figura 1.6 A)
(ROBINSON et al., 2009).
As manifestações clínicas, dependendo do hospedeiro e do sorotipo,
incluem faringite, febre faringoconjuntival, conjuntivite, ceratoconjuntivite,
bronquite, pneumonia, cistite hemorrágica, gastroenterite, miocardite e hepatite
(GAINOTTI et al., 2010). Alguns adenovírus podem persistir por anos, como
infecção latente, nas adenoides e tonsilas, e são eliminados pelas fezes meses
após a infecção inicial (RÁCZ, 2005). Dentre as espécies estudadas, D e F têm
tido grande importância por estarem associados nas infecções
ceratoconjuntivite epidêmica e gastroenterite, respectivamente (LENAERTS;
NAESENS, 2006).
Três espécies do sorotipo D, adenovírus 8 (Ad8), Ad19 e Ad37, causam
doenças oculares graves que são, geralmente, espalhadas pelo contato direto.
Além da ceratite e conjuntivite, as infecções causadas por esses sorotipos são
também caracterizadas por dor, edema, lacrimação, fotofobia, hemorragias e
formações pseudomembranares (JOHANSSON et al., 2007). Os adenovírus
entéricos (espécie F; sorotipos principais Ad40 e Ad41), depois dos rotavírus,
são os principais causadores de diarreia não bacteriana em crianças. Esses
sorotipos apresentam tropismo para o trato intestinal e, tipicamente, não
causam doenças em outras regiões do corpo (LEUNG; BROWN, 2011). Em
adição, pacientes portadores de HIV são, corriqueiramente, acometidos por
infecções intensas, causadas por Ad 40 e 41. Em muitos casos, infecções
cruzadas com Citomegalovírus intensificam o quadro diarreico (UCKUN et al.,
2004).

49
Figura 1.6: Representação esquemática da estrutura de adenovírus (A) e vírus
herpes simples (B). FONTE: (A) Marinheiro, 2009; (B) Medicaltrue, 2012.

50
1.2.2. Vírus Herpes Simples (HSV)
Os herpesvírus pertencem à família Herpesviridae, a qual apresenta
mais de 100 espécies que estão distribuídas em três subfamílias:
Alphaherpesvirinae, que ocasionam lesões na pele e mucosas, e Beta e
Gammaherpesvirinae, que estão relacionadas a manifestações sistêmicas
(KNIPE; HOWLEY, 2001). Um vírion dessa família é composto por um core
interno, contendo molécula de DNA linear de dupla fita, circundado por um
capsídeo icosaédrico, formado por 162 capsômeros, 150 hexâmeros e 12
pentâmeros e, adicionalmente, um envelope lipoproteico com espículas
glicoproteicas (Figura 1.6 B) (MEHNERT; CANDEIAS, 2005; WHITE; FENNER,
1994). Até então, nove diferentes herpesvírus foram isolados de humanos,
sendo eles: vírus herpes simples 1 (HSV-1), vírus herpes simples 2 (HSV-2),
vírus varicela-Zoster (HHV-3, VZV), vírus Epstein-Barr (HHV-4, EBV),
citomegalovírus humano (HHV-5, HCMV), vírus herpes 6A, 6B, 7 e 8 (HHV-6A,
HHV-6B, HHV-7, HHV-8) (FATAHZADEH; SCHWARTZ, 2007; GEMIN, 2008).
Doenças associadas aos HSV afetam, aproximadamente, 60 a 95 % dos
humanos adultos (FATAHZADEH; SCHWARTZ, 2007). A latência e a
reativação viral são características comuns nos HSV (DONG et al, 2012;
KLEYMANN, 2003; LEFLORE; ANDERSON; FLETCHER, 2000; VILLARREAL,
2003). As manifestações clínicas são variadas e influenciadas pela porta de
entrada viral e pela imunidade do hospedeiro (BRADY, BERNSTEIN, 2004). O
primeiro contato com o HSV-1 ocorre na faixa de seis meses a três anos de
idade, sendo transmitido, principalmente, por contato com saliva. O HSV-2 é
adquirido, geralmente, na fase da adolescência, coincidindo com o início das
atividades sexuais (EO et al., 1999; KHAN et al., 2005; ZHU et al., 2006).
A exposição inicial por HSV leva à invasão viral nas células epiteliais e
consequente replicação intracelular (FATAHZADEH; SCHWARTZ, 2007). Após
infecção primária, as partículas virais migram para as células do sistema
nervoso, principalmente, para os gânglios trigeminal e sacral, onde
permanecem latentes (ASHLEY; WALD, 1999; MILLER; DANAHER; JACOB,
1998). A latência crônica predispõe o hospedeiro a recorrentes ataques através

51
da reativação viral, aumentando o potencial de transmissão (WHITLEY;
ROIZMAN, 2001). Uma vez reativado, os vírus migram ao longo de neurônios
sensoriais para células inervadas da mucosa, onde sofre replicação e conduz à
formação de um aglomerado de vesículas na região do local inicial de
exposição (FATAHZADEH; SCHWARTZ, 2007). Essas infecções são
acompanhadas pela formação de sincícios (células gigantes plurinucleadas) no
local infectado (MANDAL et al., 2008).
Dentro do espectro de doenças causadas por HSV, pode ser destacada
uma das formas mais graves, a encefalite, que apresenta mortalidade
significativa (20-30% dos casos), apesar do uso de antivirais (TONOMURA et
al., 2010). A encefalite devasta o sistema nervoso central causando necrose
hemorrágica nos tecidos. A doença pode evoluir para uma leucoencefalite
hemorrágica aguda, causando demielinação das células nervosas (KABAKUS
et al., 2005). Dentre os pacientes mais afetados, podem ser destacadas
pessoas imunossuprimidas, incluindo HIV positivos (KURT-JONES et al.,
2004).
Fatores importantes são considerados para selecionar o tratamento de
pacientes acometidos de infecções por HSV. Dentre eles podemos destacar o
estado imune do hospedeiro, o sítio de infecção e se a infecção é primária ou
recorrente. Atualmente, os agentes antivirais como aciclovir, valaciclovir,
foscarnet, cidofovir, penciclovir, famciclovir, docosanol, trifluridina e vidarabina
são bastante utilizados e, em adição, pesquisas são realizadas para o
desenvolvimento de uma vacina (BRADY; BERNSTEIN, 2004; CUNNINGHAM
et al., 2012).
1.2.3. Metapneumovírus (HMPV)
As infecções do trato respiratório são as formas de infecção mais
comuns que afetam o homem e, dentre essas, predominam as de causa viral.
Uma das consequências da infecção é a pneumonia, que é definida como
qualquer anormalidade nas trocas gasosas, nos alvéolos, acompanhada por

52
inflamação do parênquima pulmonar (FIGUEIREDO, 2006). No entanto, devido
a grande variedade de sintomas relacionados à pneumonia viral, existem
dificuldades no diagnóstico etiológico.
Dentre os agentes causadores de doenças respiratórias virais, pode ser
destacado o metapneumovirus humano (HMPV). Esse vírus foi relatado pela
primeira vez por Van den Hoogen et al. (2001), a partir da análise de secreções
do trato respiratório de uma criança holandesa acometida por bronquite
(MENDES et al., 2010). Este vírus pertence à família Paramixoviridae,
juntamente com seus principais integrantes, parainfluenza vírus (1, 2, 3, 4a e
4b) e vírus sincicial respiratório (RSV) (DE CLERCQ, 2004). Como mostrado na
Figura 1.7, o HMPV é envelopado, apresenta RNA de fita simples, com
orientação negativa (RNA–), contendo três glicoproteínas presentes no
envelope: de fusão (F), de adesão (G) e uma de hidrofobicidade baixa (SH)
(YU et al., 2012). A proteína F é bastante conservada e a proteína G revela um
alto nível de diversidade, sendo esta classificada em dois grandes grupos, A e
B, e, em adição, subdividida em subgrupos 1 e 2 (FEUILLET et al., 2012). Huck
et al. (2006) sugerem uma nova divisão para os subgrupos A2, em A2a e A2b.
Tanto F quanto G, são alvos em potencial para o processo de neutralização,
realizado pelo sistema imunológico. Em especial, a proteína G é classificada
como uma proteína de membrana altamente glicosilada, através de ligações O-
e N-, com domínios extracelulares, transmembranares e intracelulares
(THAMMAWAT et al., 2008).
Estudos epidemiológicos mostram que o HMPV é um patógeno
respiratório distribuído em todos os continentes (WYDE et al., 2003) e que
pode infectar indivíduos de todas as idades, isso inclui recém-nascidos
(XEPAPADAKI et al., 2004), crianças e adultos (TOLLEFSON; COX;
WILLIAMS, 2010). Além disso, infecções causadas em indivíduos
imunossuprimidos (VAN DEN HOOGEN et al., 2003) e infecções nosocomiais
(TOLLEFSON; COX; WILLIAMS, 2010) têm sido relatadas. Em casos mais
delicados, indivíduos podem apresentar co-infecções com RSV, que apresenta
padrão de infecção semelhante ao HMPV (RUOHOLA et al., 2009). No entanto,
diferente do RSV, HMPV possui dois genes que codificam para duas proteínas

53
não estruturais (NS1 e NS2) que induz resposta imune diferente no hospedeiro
(LING; TRAN; TENG, 2009).
Figura 1.7:Representação esquemática da estrutura do HMPV e seu ciclo dereplicação viral. (1) Glicosaminoglicanos (GAGs), presentes na membranaplasmática da célula hospedeira são reconhecidos pela proteína de ligação Ge, as integrinas ávâ1, pela proteína de fusão F; (2) Inclusão do nucleocapsídeoviral no citoplasma celular; (3) Síntese de RNA mensageiro viral; (4) Replicaçãodo RNA viral; (5) Tradução das proteínas virais [fusão (F), de ligação (G e HN),de baixa hidrofobicidade (SH), nucleoproteína (N), fosfoproteína (P), proteínada matriz (M) e polimerase (L)] e migração para o Retículo Endoplasmático(RE) e Complexo de Golgi; (6) Brotamento das partículas virais.
FONTE:Feuillet et al., 2012

54
HMPV, preferencialmente, infecta as células epiteliais ciliadas do trato
respiratório. O reconhecimento dessas células é feito através de proteínas de
ligação, o que inclui a proteína G e outras duas, H e HN. GAG´s, presentes na
membrana plasmática da célula hospedeira (Figura 1.7), são reconhecidos
pelas proteínas de ligação e, em adição, integrinas ávâ1, pela proteína de
fusão F (CSEKE et al., 2009; THAMMAWAT et al., 2008). A incubação ocorre
por um período entre 4 e 6 dias, sendo que as partículas virais começam a ser
eliminadas pela célula em um período de 5 dias a 2 semanas (FEUILLET et al.,
2012). Durante o processo de tradução do RNA viral, é realizada produção de
nucleoproteína (N), fosfoproteína (P), proteína da matriz (M) e polimerase (L).
Em seguida, os componentes proteicos virais e uma molécula de RNA com
orientação negativa são montadas formando, assim, uma partícula infecciosa
(Figura 1.7).
As infecções causadas pelo HMPV podem se localizar na região do trato
respiratório superior (ITRS) ou inferior (ITRI). Dentre os sintomas mais comuns,
são destacadas a bronquite, pneumonia, exacerbação asmática e inflamação
na garganta. Em pacientes com ITRS, é muito comum febre, coriza, tosse,
rinite, faringite, otalgia e anormalidades na membrana do tímpano. Em adição,
indivíduos que apresentam ITRI, não só podem apresentar ITRS como,
também, estridor pulmonar, dispneia, taquipneia, hipóxia e rouquidão
(HERMOS; VARGAS; McADAM, 2010). Esse quadro sintomático pode ser
intensificado se o paciente é uma criança, ou transplantado ou com o sistema
imunológico suprimido. O progresso da doença para pneumonia hemorrágica
falência respiratória e choque séptico tem sido relatado na literatura (DONOSO
et al., 2008; RUUSKANEN et al., 2011).
Embora muito similar com RSV, não existe tratamento antiviral
específico para HMPV. Tem sido relatado o uso de ribavirina, heparina e
lipídeo sialil sulfatado (NMSO3) como drogas capazes de inibir a replicação do
HMPV (ABED; BOIVIN, 2006).

55
1.2.4. Vírus da Dengue (DENV)
A Dengue é uma arbovirose causada por um Flavivirus, pertencente à
família Flaviviridae, e transmitida pelos mosquitos Aedes aegyptii e Aedes
albopictus, apresentando quatro sorotipos conhecidos (DENV-1, DENV-2,
DENV-3 e DENV-4) (WHITEHORN; SIMMONS, 2011). As partículas virais são
formadas por um nucleocapsídeo envelopado contendo genoma constituído por
uma fita simples de RNA com orientação positiva (RNA+) (ACOSTA;
CASTILLA; DAMONTE, 2011). O genoma de, aproximadamente, 11 kilo pares
de bases (kpb), organizado em uma única fase de leitura aberta e que codifica
para uma poliproteína que, por sua vez, é clivada em 3 proteínas estruturais (C,
PrM e E) e 7 não estruturais (NS1, NS2a, NS2b, NS3, NS4a, SN4b e NS5),
através de mudanças póstraducionais (Figura 1.8) (KATO et al., 2010).
Clinicamente pode se manifestar como infecção inaparente, dengue
clássica (DC) ou dengue grave [febre hemorrágica da dengue (FHD)/síndrome
de choque da dengue (SCD)] (MURRELL; WU; BUTLER, 2011). O próprio
nome da doença vem do dialeto africano suaíle onde Ki denga pepo quer dizer
“possuído subitamente por um espírito”. A expressão era utilizada por africanos
do leste para descrever uma doença transmitida por mosquitos que,
abruptamente, dominava seres humanos, causando horríveis cefaléias, dores
oculares e dores e inchaço das articulações, que são sintomas que
caracterizam a DC (SAVALIA; SINDHI, 2008). Entretanto, a dengue pode
evoluir para um quadro mais agressor desencadeando a FHD, quando há uma
infecção secundária com sorotipo de vírus diferente. As manifestações clínicas
iniciais da FHD são as mesmas descritas para a DC, até que ocorra o
surgimento de manifestações hemorrágicas espontâneas ou provocadas por
algum processo de injúria (WHITEHORN; SIMMONS, 2011).

56
Figura 1.8: Vírus da Dengue. (a) Partícula viral imatura. Círculo
correspondendo a proteínas que sobressaem da superfície do capsídeo; (b)
Proteínas do envelope (E) de partícula viral madura. Cores azul, amarelo e
vermelho indicam os diferentes domínios das proteínas E. Verde indica as
junções dos domínios.
FONTE: Mukhopadhyay; Kuhn; Rossmann, 2005

57
1.2.5. Infecções Virais Hemorrágicas
As infecções virais geralmente são acompanhadas por quadros
hemorrágicos, sejam locais ou sistêmicos. Os adenovírus, por exemplo, podem
desencadear cistites, enterites, encefalites e conjuntivites hemorrágicas
(JOHANSSON et al., 2007; LENAERTS; NAESENS, 2006; STOCK et al.,
2006). Esses vírus também estão relacionados com a degeneração das
adenoides, de infecção respiratória aguda e febre faringo-conjuntiva.
Casos clínicos de encefalites hemorrágicas causadas por HSV-1 e HSV-
2 também não são raros na literatura. Geralmente, esses vírus causam
inflamação aguda no cérebro e, consequentemente, edema e hemorragia nos
lobos temporais (MUENCH et al., 2001). A evolução do processo hemorrágico
é acompanhada pela análise do fluido cerebroespinhal que, geralmente,
apresenta aumento nos teores de proteína e no número de linfócitos e
eritrócitos. As lesões hemorrágicas agudas são reveladas por sinais de alta
intensidade nas imagens de ressonância magnética (SALIH et al., 2009).
Em relação às doenças causadas por vírus envelopados de RNA que
causam hemorragias, a dengue pode ser destacada como uma das mais
importantes (FIGUEIREDO, 2006). Os DENVs infectam células da linhagem
dos monócitos/macrófagos, assim como as células dendríticas (GUGLANI;
KABRA, 2005). Na dengue clássica, várias células T são ativadas e esta
resposta é mediada por moléculas chamadas de citocinas, como interleucina-2
(IL-2), interleucina-8 (IL-8), interferon-γ (INFγ), fator de necrose tumoral α
(TNFα) etc. Há então, uma ativação tanto do sistema imune humoral quanto do
mediado por células T e, além disso, há produção de anticorpos para aquele
sorotipo de vírus específico (ROTHMAN; ENNIS, 1999). No entanto, durante
infecção secundária com diferente sorotipo do vírus, anticorpos provenientes
da primeira infecção se ligam ao segundo sorotipo, sendo que esta interação
não é suficiente para inativá-lo. Dessa forma, os vírus marcados com
anticorpos são internalizados com maior facilidade pelos macrófagos graças a
receptores Fc que reconhecem os anticorpos (GUGLANI; KABRA, 2005; LEE;
LIAO; LIN, 2005). Em seguida, estes macrófagos entram em processo de

58
ativação. Entretanto, até que se tornem maduros, há uma liberação exagerada
de TNFα o que leva à morte celular programada (apoptose) não só de células
infectadas, mas, também, de células endoteliais sadias causando, assim,
trombocitopenia, hemorragia e, em casos mais avançados, choque (LEE et al.,
2005). Estes eventos levam ao quadro clínico de FHD/SCD.
Em pneumonias virais podem ser observadas diferenças histológicas
significativas. Infecções brônquicas e alveolares podem estar presentes em
pacientes acometidos por vírus sincicial respiratório. Infecções com rinovírus
podem causar hiperplasia e descamação das células alveolares. Em adição,
análises patológicas de casos fatais de pneumonia causada por HMPV indicam
broncopneumonia hemorrágica bilateral (DONOSO et al., 2008). Essas lesões
anatomopatológicas, acompanhadas de quadros hemorrágicos, podem
também, ser observadas em frangos de corte infectados por metapneumovírus
aviário (AMPV). Essa infecção causa destruição do epitélio ciliar dos seios
nasais e da traqueia resultando em congestão, hemorragias petequiais, lesões
necróticas da mucosa da cavidade nasal, traqueal e fenda palatina, com
posterior surgimento de traqueíte e rinite com secreção purulenta (SALES,
2010).
1.2.6. Hemostasia
Hemostasia é definida como o controle da perda sanguínea a partir de
um vaso lesado. Esse processo é iniciado com as plaquetas, que aderem às
macromoléculas presentes nas regiões subendoteliais da área danificada e
sofrem agregação até a formação de tampão hemostático. Depois da adesão,
as plaquetas se tornam ativadas, expõem seus fosfolipídeos e alteram suas
glicoproteínas da superfície da membrana permitindo, assim, a agregação e
secreção do conteúdo dos grânulos. Concomitantemente, é realizada
estimulação da ativação local dos fatores plasmáticos de coagulação, gerando
coágulo de fibrina. Esse coágulo tem a função de reforçar o agregado

59
plaquetário, sendo degradado à medida que ocorre a cicatrização (MANN,
2009).
A coagulação ocorre por meio de ativação proteolítica sequencial de pró-
enzimas por proteases do plasma, resultando na formação de trombina que,
então, quebra a molécula de fibrinogênio em monômeros de fibrina (Figura
1.9). Por muito tempo foi utilizado um modelo que divide a coagulação em duas
vias disposta em “cascata”, via extrínseca e intrínseca (MACFARLANE, 1964;
DAVIE; RATNOFF; 1964). No entanto, como essas vias são bastante
intrincadas, muitos autores utilizam outro modelo que é dividido em iniciação,
amplificação e propagação (Figura 1.9). Esse modelo é indicado como mais
representativo para o entendimento da hemostasia in vivo (FERREIRA et al.,
2010).
A linha entre hemorragia e trombose é muito tênue, o que demonstra
que a hemostasia deve ser perfeitamente equilibrada. Quando a camada
subendotelial é exposta, devido a alguma lesão, uma glicoproteína chamada de
fator tissular (TF) entra em contato com o sangue e se liga ao fator VIIa (TF-
VIIa), formando um complexo denominado “tenase extrínseco”, que ativa o
fator X (KALAFATIS et al., 1994). O TF não é expresso constitutivamente nas
células endoteliais, mas está presente nas membranas das células ao redor do
músculo liso e nos fibroblastos. Assim, o fator Xa (fator X ativado) desencadeia
a formação de trombina a partir de protrombina que, por sua vez, cliva o
fibrinogênio em fibrina. Nesse ínterim, pequenas quantidades de trombina são
capazes de amplificar a ativação do fator X e produção de mais trombina pela
ativação dos fatores V, VII, IX e XI (FERREIRA et al., 2010).
A inibição das reações da coagulação é regulada pela inibição
enzimática ou pela modulação da atividade dos cofatores. Dentre outros, pode
ser destacado o inibidor da via do fator tecidual (TFPI), principal inibidor
fisiológico do complexo TF-VIIa e, portanto, da fase de iniciação da coagulação
(Figura 1.9).

60
Figura 1.9: Representação do modelo da coagulação baseado em superfícies
celulares, compreendendo as fases de iniciação, amplificação e propagação.FONTE: Vine, 2009.

61
Dentre os testes utilizados para avaliar os níveis de procoagulantes
envolvidos na fase de iniciação da coagulação, é destacado o teste do tempo
da trombina (TP). Além disso, o TTPa (Teste de tromboplastina parcial ativada)
é utilizado para avaliar os níveis de procoagulantes envolvidos na produção
amplificada de trombina na superfície das plaquetas ativadas (FERREIRA et
al., 2010). A análise do tempo de sangramento em ratos (FONSECA, 2009) e
humanos (LIND; KURKJIAN, 2007) tem sido bastante útil para analisar a
dinâmica in vivo da hemostasia. A partir dos resultados obtidos nesses
experimentos, é possível relacioná-los com o número e ativação das plaquetas.
Muitos autores relatam polissacarídeos com propriedade antiviral e
anticoagulante (CHATTOPADHYAY et al., 2007; DUARTE et al., 2001;
KOLENDER et al., 1997; LEE et al., 2007). Devido ao fato das doenças
cardiovasculares serem intimamente relacionadas à trombose, atualmente é
observado grande busca comercial para se obter novos anticoagulantes. O
mais utilizado é a heparina, descoberta em 1916 e, ainda hoje, utilizada
mundialmente como agente terapêutico (anticoagulante e antitrombótico) para
tratamento e prevenção da trombose venosa profunda pós-cirúrgica e pós-
parto (RODRIGUES, 2011). No entanto, as informações referentes aos quadros
hemorrágicos relacionados a infecções virais são sugestivas para a
investigação de polissacarídeos que apresentem não só, atividade antiviral,
como, também, atividade anti-hemorrágica.

62
1.3. OBJETIVOS
1.3.1. Gerais
Caracterizar físico-quimica e estruturalmente polissacarídeos obtidos de
folhas da planta Aloe barbadensis e avaliar suas atividades antiviral, citotóxica
e suas propriedades anti-hemorrágica, pró-coagulante e seus possíveis sinais
de toxicidade em camundongos.
1.3.2. ESPECÍFICOS
• Extrair os polissacarídeos totais (PT) de A. barbadensis e fracioná-los
através da cromatografia de troca iônica em colunas de DEAE-celulose;
• Avaliar o rendimento bruto dos PT e a composição química em ácidos
urônicos e grupos sulfato nos PT e nas frações PI e PII obtidas de DEAE-
celulose;
• Determinar a composição química dos monossacarídeos presentes nas
estruturas das frações PI e PII;
• Determinar a massa molecular e avaliar o grau de pureza, da fração
prioritária obtida de DEAE-celulose (PII), por procedimentos de
eletroforese em géis de poliacrilamida e agarose, respectivamente;
• Caracterizar estruturalmente por espectroscopia no infravermelho (IR) os
PT e as frações obtidas por DEAE-celulose (PI e PII), e por ressonância
magnética nuclear (RMN) a fração PII;

63
• Avaliar a citotoxicidade dos PT e das frações PI e PII em células de rim de
Cercopithecus aethiops (Vero), de larva de mosquito Aedes albopictus
(C6/36), de carcinoma cervical humano (HeLa), de carcinoma de laringe
humana (HEp-2) e de rim de macaco rhesus [Macaca mulata (LLC-MK2)];
• Avaliar a atividade antiviral dos PT e das frações PI e PII contra os vírus da
dengue (DENV-1), vírius herpes simples (HSV 1 e 2), metapneumovírus
(HMPV) e adenovírus (Ad-19 e Ad-41);
• Avaliar os efeitos anti-hemorrágico e pró-coagulante das frações PI e PII em
ratos;
• Avaliar a toxicidade dos PT por dose repetida (14 dias) em camundongos
através da análise de parâmetros bioquímicos hematológicos e
histopatológicos


65
2.1. MATERIAIS E MÉTODOS
2.1.1. Coleta e Identificação
Folhas verdes, túrgidas e não-injuriadas de A. barbadensis Miller
(Figura 2.1 A e B) (registrada sob n° 50925 no Herbário Prisco Bezerra,
Universidade Federal do Ceará, UFC, com a sinonímia Aloe vera L.
Burm.) foram coletadas no Horto de Plantas Medicinais da UFC,
município de Fortaleza, em abril de 2010 e 2011. Para isso, com auxílio
de estilete, as folhas foram removidas com corte realizado em suas bases
e submergidas em água, por 30 min, para eliminação do exudato
amarelado. Em seguida, foram lavadas em água corrente para eliminação
de eventuais sujidades e, por fim, com água destilada.
2.1.2. Obtenção dos Polissacarídeos Totais (PT) de A. barbadensis
Após o procedimento descrito anteriormente, a casca das folhas
frescas (7 Kg) foi removida, utilizando lâmina de aço inoxidável (Figura
2.1 C), e a polpa obtida (3,85 Kg) foi lavada com água destilada para
remoção do exudato amarelado (rico em compostos fenólicos). Em
seguida, foi adicionada água destilada na relação 1:1 (v/v), sendo a polpa
processada (processador Wallita) por 5 minutos e o homogenato
submetido à extração dos polissacarídeos totais segundo a metodologia
de Wu et al. (2006), com algumas modificações. Portanto, o homogenato
foi aquecido (70 °C, 2 h), seguido da filtração em tecido de nylon
acoplado à bomba de vácuo (Figura 2.1 D) e, ao filtrado obtido,
adicionado etanol comercial 95% (1:4 v/v). A mistura foi deixada em
repouso durante 12 h a 4 °C, centrifugada (12.000 x g, 30 min, 4 °C) e o
precipitado obtido lavado, sequencialmente, com etanol absoluto,
acetona e éter. Após lavagem, o precipitado foi redissolvido em água
destilada, dialisado contra o mesmo solvente, liofilizado e denominado
polissacarídeos totais (PT). A remoção de possível contaminantes
proteicos dos PT foi realizada de acordo com a metodologia descrita por
Yang et al. (1999). Para isto, os PT foram dissolvidos em água destilada

66
(1 mg/mL) e adicionado ácido tricloroacético a uma concentração final de
0,1 %, deixada em repouso por 2 h a temperatura ambiente. Em seguida,
as proteínas precipitadas foram removidas por centrifugação (12.000 x g,
30 min, 4 °C) e ao sobrenadante obtido, adicionado etanol comercial a
95 % (1:4 v/v) para precipitação dos PT livres de contaminação proteica.
Finalmente, os PT foram dialisados contra água destilada para remoção
do excesso de etanol e, em seguida, liofilizados (Figura 2.2).
Figura 2.1: (A) Fotografia do plantio de espécimes de Aloe barbadensis
em canteiro irrigado no Horto de Plantas Medicinais da UFC; (B) corte
transversal de uma folha, demonstrando a casca e a polpa interna; (C)
retirada da casca; (D) sistema de filtração a vácuo; (E) Classificação
taxonômica da espécie A. barbadensis.

67
Figura 2.2: Fluxograma de obtenção dos polissacarídeos totais (PT) de
Aloe barbadensis.

68
2.1.3. Determinação de Umidade da Polpa
Para determinação da umidade da polpa, pesa-filtros previamente
tarados, contendo amostras do mesófilo foliar (triplicata), foram colocados
em estufa a 110 °C por 24 horas. Os pesa-filtros, após serem removidos
da estufa, foram mantidos em dessecador até atingirem o equilíbrio com
a temperatura ambiente. Em seguida, foram pesados sucessivas vezes
para obtenção de uma leitura de massa constante. O teor de umidade foi
calculado pela diferença entre as massas inicial e final das amostras,
sendo esse valor expresso em percentagem.
2.1.4 Fracionamento dos PT por Cromatografia de Troca Iônica em
Coluna de DEAE-celulose
Os PT (200 mg/100 mL de tampão acetato de sódio 50 mM, pH
6,0) foram aplicados em coluna de DEAE-celulose (30 x 1,5 cm),
previamente equilibrada com o mesmo tampão e mantendo um fluxo
constante de 50 mL/h. A matriz foi inicialmente lavada com tampão de
equilíbrio seguida da eluição com o tampão de equilíbrio contendo
diferentes concentrações de NaCl (0,5; 1,0; 3,0 M). As frações obtidas
foram monitoradas em espectrofotômetro (modelo Amersham
Biosciences Ultrospec 1100 pro) a 490 nm utilizando o método do fenol-
ácido sulfúrico (DUBOIS, 1956). A fração majoritária foi dialisada
exaustivamente em água destilada, liofilizada e armazenada para uso
posterior. Depois de liofilizada, 3 mg das frações foram misturadas a 1
mL do azul de 1,9-dimetilmetileno (DMB) para analisar as propriedades
metacromáticas.

69
2.1.6. Caracterização Físico-química dos Polissacarídeos
2.1.6.1. Determinação de Carboidratos Totais (CT)
Os PT e as frações obtidas de DEAE-celulose (PI e PII) foram
submetidas à determinação de carboidratos totais pelo método do fenol-
ácido sulfúrico (DUBOIS et al., 1956). As leituras de absorbância foram
realizadas em espectrofotômetro a 490 nm, utilizando-se a D-galactose
como padrão.
2.1.6.2. Determinação de Ácidos Urônicos
Os PT e as frações PI e PII foram submetidas à determinação de
ácidos urônicos pelo método carbazol (DISCHE, 1974). As leituras de
absorbância foram realizadas em espectrofotômetro a 550 nm, utilizando
glucuronolactona como padrão.
2.1.6.3. Determinação do Teor de Sulfato
O teor de sulfato total foi determinado no hidrolisado ácido (2-4
mg/mL em HCl 1M, 5 horas, 105 °C) dos PT e das frações PI e PII
utilizando o método turbidimétrico da gelatina-bário (DODGSON, 1961).
As leituras de absorbância foram realizadas em espectrofotômetro a 360
nm, utilizando sulfato de sódio como padrão.

70
2.1.5.4. Determinação de Contaminantes Proteicos
A avaliação de contaminantes proteicos foi realizada segundo o método de
Bradford (1976), utilizando-se Albumina Sérica Bovina (BSA) como padrão.
2.1.5.5. Eletroforese em Gel de Agarose
Os PT e as frações PI e PII (20 µg) foram analisadas por eletroforese em
gel de agarose 0,5% em tampão 1,3-acetato diaminopropano 0,05 M (pH 9,0).
A corrida foi realizada em voltagem constante (100 V) durante 60 min. Após o
procedimento, as frações presentes no gel foram fixadas com solução de N-
cetil-N,N,N-brometo de trimetilamônio (cetavlon) 0,1% por 24 horas. Em
seguida, o gel foi corado com azul de toluidina 0,1% e descorado com uma
solução contendo etanol absoluto, água destilada e ácido acético concentrado
(4,95: 4,.5: 0,1; v/v/v) como descrito por Dietrich e Dietrich (1976). Em adição,
outra preparação eletroforética foi realizada com a finalidade de corar as
bandas dos polissacarídeos aniônicos metacromáticos (sulfatados) e não
metacromáticos (carboxilados). As frações presentes no gel, após fixadas com
uma solução de cetavlon 0,1%, por 24 horas, foram coradas com stainsall [1-
etil-2-(3-(1-etilnafto (1,2-d) tiazolina-2-ilideno)–2-metilpropenil) nafto (1,2-d)
tiazolium bromídeo] e o gel descorado com a mesma solução descrita
anteriormente. Os marcadores utilizados foram: condroitim 4-sulfato, dermatam
sulfato e heparam sulfato (Sigma-Aldrich, St. Louis, MO, USA).
2.1.5.6. Eletroforese em Gel de Poliacrilamida (PAGE)
Os PT e as frações PI e PII (20 µg) foram aplicados em gel de
poliacrilamida 6%, em tampão barbital sódico 0,02 M (pH 8,6). O gel foi
submetido à voltagem de 100 V por 40 min, em sistema de eletroforese
Mini – gel da Bio – Rad. Em seguida, o gel foi corado com azul de

71
toluidina 0,1% preparado em ácido acético 1%. Com a mesma finalidade,
conforme descrito anteriormente, outra corrida eletroforese foi realizada
também utilizando o mesmo corante stainsall. As massas moleculares
das frações polissacarídicas utilizadas foram estimadas por comparação
com a migração eletroforética de polissacarídeos marinhos tomados
como padrões (PAVÃO et al., 1998; PEREIRA; MULLOY; MOURÃO,
1999; SANTOS; MULLOY; MOURÃO, 1992), tais como: dextram sulfato
de alta massa molecular (~500 kDa), condroitim 6-sulfato de cartilagem
de tubarão (~60 kDa), condroitim 4-sulfato de cartilagem de baleia (~40
kDa) e dextram sulfato de baixa massa molecular (~8kDa), todos obtidos
da Sigma-Aldrich.
2.1.5.7. Cromatografia de Troca Iônica em Coluna de Mono-q
Acoplada a HPLC
A pureza da fração PII também foi monitorada através de
Cromatografia de troca-iônica Mono-q (Pharmacia) de acordo com os
padrões estabelecidos por Cinelli (2009). A coluna foi equilibrada em
tampão TRIS-HCl 20 mM, (pH 8,0), acoplada ao sistema de HPLC e
eluída na mesma solução de equilíbrio contendo concentrações
crescentes de NaCl ( 0,5 - 3 M). O volume das frações obtido foi de 0,5
mL, mantendo-se um fluxo constante de 30 mL/h.
2.1.5.8. Determinação da Composição Monossacarídica das Frações
Polissacarídicas
Inicialmente, as frações PI e PII (20 mg) foram hidrolisadas pela adição
de 0,5 mL de solução de ácido trifluoracético (TFA) 2 M, e mantidas a 100 °C,
por 3 h. Os hidrolisados obtidos foram submetidos a diálise exaustiva com
água destilada, para remoção do excesso de ácido. Em seguida, os
hidrolisados foram reduzidos pela adição de 0,5 mL de solução de hidróxido de

72
amônio (NH4OH) 1 M, contendo 1,0 mg de borohidreto de sódio (NaBH4),
mantidos a temperatura ambiente, por 1 h, seguido de acidificação por ácido
acético glacial 50%. Posteriormente, para remoção do excesso de ácido, foi
adicionado 1 mL de metanol, seguido de evaporação por fluxo de ar, repetindo
o processo por 5 vezes. Os produtos reduzidos obtidos (alditóis) foram
acetilados com solução de anidrido acético:piridina (10:1, v/v) e submetidos ao
aquecimento em banho-maria (100 °C,1 h) (KIRCHER, 1960). Finalmente foi
adicionado clorofórmio (CHCl3) (1:3 m/v) e os derivados alditol-acetato obtidos,
analisados por cromatografia gasosa acoplada a espectrômetro de massa (GC-
MS).
Para isso, foi utilizado o sistema Shimadzu GCMS-QP2010 Plus com
coluna Rtx-5MS (5 % fenil, 95 % dimetilpolissiloxano) da Restek® (30 m x 0.25
mm x 0.25 µm). O injetor foi mantido a 260 °C, com divisão de fluxo (split) na
razão de 1:1. A temperatura do forno da coluna foi elevada a 110-250 °C, com
taxa de aquecimento de 2 °C min-1 (ALVES et al., 1998). Hélio foi utilizado
como gás de arraste com velocidade linear de 37,4 cm s-1. Um volume de 1 µL
de amostra foi injetado no cromatógrafo. Para a detecção por espectrometria
de massa, foi utilizado detector contendo fonte de ionização por elétrons (EI-70
eV) e um analisador de massas quadrupolo. A identificação dos constituintes
da mistura foi feita por comparação dos seus espectros de massas com
espectros depositados na biblioteca NIST05, contida no computador do
espectrômetro de massa, assim como os seus tempos de retenção. A interface
foi mantida a 230 °C e a fonte de íons a 200 °C.
2.3.6. Caracterização Estrutural dos Polissacarídeos
2.3.6.1. Espectroscopia no Infravermelho Transformada de Fourier
(FTIR)
Os PT e as frações PI e PII foram avaliadas por espectroscopia no
infravermelho transformada por Fourier em Espectrômetro PERKIN ELMER
SPECTRUM 100, utilizando-se pastilhas de KBr, na região de 4000 a 400 cm-1.

73
2.1.6.2. Dessulfatação da Fração Polissacarídica Sulfatada (PII)
A dessulfatação da fração PII foi realizada como descrito por Vieira,
Mulloy e Mourão (1991). 20 mg da fração PII foram dissolvidos em 5 mL de
água destilada e adicionado 1 g de resina Dowex 50-W (H+, malha de 200-
400). Para liberação da fração dessulfatada da resina, a mesma foi
neutralizada pela adição de piridina, seguida da filtração a vácuo sendo o sal
piridina resultante, dissolvido em 2,5 mL de dimetil-sulfóxido:metanol (9:1, v/v),
seguido do aquecimento a 80 °C por 4 h e os produtos dessulfatados obtidos
dialisados exaustivamente em água destilada e liofilizados. A dessulfatação foi
mensurada pela razão molar de sulfato/açúcar total.
2.1.6.3 Espectroscopia de Ressonância Magnética Nuclear (RMN)
Os espectros de RMN da fração PII foram obtidos em
espectrômetro Bruker DRX 800 MHz utilizando uma sonda de tripla
ressonância de 5 mm. Para isso, 3 mg de Fração PII foram dissolvidos
em 0,5 ml de D2O 99,9%. Os experimentos foram realizados a 60 °C
com supressão de D2O por pré-saturação, sendo os unidimensionais,
obtidos de detecção direta de 1H, e os bidimensionais heteronucleares
por 1H/13C DEPT-HSQC.

74
2.4. RESULTADOS E DISCUSSÃO
2.2.1. Extração e Rendimento dos Polissacarídeos Totais (PT) de A.
barbadensis
Antes do processo de extração a casca das plantas foi removida, a polpa
foi pesada (55% da massa foliar fresca) e a umidade da polpa foi estimada em
98%. Femênia et al. (1999 e 2003) descreveram valores de conteúdo de água
(umidade %) em plantas de A. barbadensis de 4 anos de idade, que variaram
de 98,5 a 99,55%. Apesar desses espécimes terem sido cultivados nas Ilhas
Baleares (Espanha), não diferenciaram muito das plantas descritas por
Campestrini (2007), crescidas no Estado do Paraná (Brasil), nem das do
presente trabalho, cultivadas no Horto de Plantas Medicinais (Fortaleza, Ceará,
Brasil). No entanto, as folhas das plantas do Horto apresentaram massa média
maior (ca. 350 g), quando comparadas com as cultivadas no estado do Paraná
(ca. 240,0 g). No trabalho realizado por Waller, Pelley e Strickland (2004),
também foi analisada a umidade da polpa de folhas de A. barbadensis obtidas
de três fazendas experimentais do Texas (EUA), cujos resultados obtidos
(99,89%) mostraram-se semelhantes aos obtidos nesse trabalho.
Os polissacarídeos totais (PT) de A. barbadensis foram extraídos por
extração aquosa a quente (70 °C) em pH neutro. Apesar da maioria dos
estudos citados na literatura terem referenciado a acemanana (manana
acetilada) como polissacarídeo majoritário, o presente trabalho objetivou obter
PT apresentando uma maior variedade estrutural de polissacarídeos. Muitos
polissacarídeos de reserva, pectinas e hemiceluloses, são obtidos por extração
aquosa a quente em pH neutro (FRY, 2011). Entretanto, hemiceluloses do tipo
xilanas e xiloglucanas requerem adição de álcalis durante a extração. Para este
trabalho, foi descartada a possibilidade de realizar a extração dessas
hemiceluloses, visto que o tratamento alcalino pode causar deacetilação dos
resíduos de manose e glucose, influenciando drasticamente na solubilidade
das frações obtidas (NI; YATES; TIZARD, 2004).

75
Os PT foram obtidos a partir de 3,85 Kg de polpa, provenientes de 7 Kg
de folhas frescas, seguido por precipitação com etanol comercial 95%. Os PT
obtidos da precipitação apresentaram rendimento de 5,5% em relação à massa
seca da polpa. Os PT, após tratados com ácido tricloroacético, apresentaram
uma redução de rendimento de 37,4%, chegando, assim, a um rendimento de
3,4%, em relação à massa seca da polpa. A primeira descrição de
polissacarídeos de folhas de A. barbadensis foi feita, provavelmente, por Rowe
e Parks (1941). Eles encontraram uma fração insolúvel em álcool (FIA) de
polissacarídeos obtidos a partir de extratos de folhas de A. barbadensis.
Femênia et al. (2003) ajustaram a quantidade de álcool para 6 volumes.
Gowda, Neelisidaiah e Anjaneyalu (1979) já haviam relacionado a obtenção da
FIA com as quantidades de manose e glucose. Mananos puros e
glucomananos com pouca glucose em sua estrutura tendem a ser precipitados
apenas com altas concentrações de etanol (proporção a partir de 3:1). A razão
para essa observação ainda não está muito clara, mas pode estar relacionada
ao grau de acetilação dos resíduos de manose e ao tamanho da molécula (NI;
YATES; TIZARD, 2004). A acetilação ocorre primariamente em resíduos de
manose, já que glucomananos ricos em unidades de glucose apresentam baixo
grau de acetilação. Os PT obtidos, no presente estudo, a partir da precipitação
com etanol, apresentaram rendimentos muito semelhantes como os descritos
por Campestrini (2007).
2.2.2. Cromatografia de Troca Iônica em Matriz de DEAE-celulose
O perfil cromatográfico dos PT (200 mg) aplicados na cromatografia de
troca iônica em coluna de DEAE-celulose (Figura 2.3), apresentou um pico
eluído com o tampão de equilíbrio, acetato de sódio 50 mM, pH 6,0 (PI) e, um
pico eluído com o mesmo tampão de equilíbrio contendo NaCl 0,5 M (PII). As
frações PI e PII apresentaram rendimentos de massa seca de 65,0 e 11,5%,
respectivamente, denotando, portanto, uma diferença em polissacarídeos de
53,5%. Esses resultados estão de acordo com alguns estudos descritos na
literatura, que relatam que os polissacarídeos neutros (pico não retido na

76
cromatografia em DEAE) constituem a fração majoritária dos carboidratos totais
de A. barbadensis (GRINDLAY; REYNOLDS, 1986; HAMMAN, 2008; LUTA;
DUNCAN; SINNOTT, 2009; REYNOLDS; DWECK, 1999; STRICKLAND;
PELLEY, 2004).
Chang, Feng e Wang (2011) e Chang, Chen e Feng (2011), realizaram
estudos de obtenção de polissacarídeos obtidos de diferentes partes da planta
(casca, polpa foliar e flores), para as espécies A. arborescens e A.
barbadensis, respectivamente, por procedimento de cromatografia, em coluna
de DEAE-Sephadex A-25. Nesse estudo, os autores obtiveram um rendimento
de uma fração polissacarídica obtida pelo procedimento de DEAE-sephadex A-
25 de 5% para os polissacarídeos negativamente carregados (PNC) presentes
na polpa. Esse resultado mostrou-se inferior ao obtido em nosso estudo
(11,5%).
Figura 2.3: Cromatografia de troca iônica em coluna de DEAE-celulose dos
PT de Aloe barbadensis. As frações PI e PII foram eluídas com tampão
acetato de sódio 50 mM, pH 6,0, sem NaCl e com o mesmo tampão contendo
0,5 M de NaCl, respectivamente. A cromatografia foi monitorada pelo método
fenol-ácido sulfúrico (DUBOIS et al., 1956). (-♦ -) Dosagem de carboidratos
totais; (-o-) concentração de NaCl (M).

77
Desde 1970, McAnalley et al. vêm realizando estudos com
polissacarídeos neutros de A. barbadensis. Em 1992, esse mesmo grupo isolou
a acemanana da mesma espécie de planta e utilizaram esse polissacarídeo
nos ensaios clínicos em pacientes portadores de AIDS. Esse estudo gerou uma
patente registrada sob numeração de 5.106.616 e, atualmente, este
polissacarídeo vem sendo comercializado como Acemannan® e Carrisyn®
(CARRINGTON LABORATORIES, 1992; STRICKLAND;PELLEY, 2004). Como
consequência, resultados obtidos, por outros autores, relacionados a outras
atividades biológicas obtidas para outros polissacarídeos neutros da mesma
espécie, foram atribuídos à acemanana (FEMENIA et al., 2003; LEE et al.,
2001; LEUNG et al., 2004; STRICKLAND; PELLEY, 2004). Leung et al. (2004),
isolaram 5 frações de diferentes massas moleculares (10, 20, 470, 1300 e
10000 kDa) por cromatografia de exclusão molecular em resina XAD-4,
acoplada a HPLC, sendo também nomeados de acemanana. Outros autores,
utilizando procedimentos cromatográficos de exclusão molecular em diferentes
matrizes cromatográficas, para A. barbadensis, encontraram somente um pico
que apresentou massa molecular que variou de acordo com a matriz
empregada: 45 kDa, resina Sephacryl S-400-HR (FEMENIA et al., 2003), 80
kDa, resina Beckman Spherogel TSK 2000 (STRICKLAND;PELLEY, 2004) e
>500 kDa, resina Biosep Sec H400 (LEE et al., 2001), onde os autores
atribuíram que essas moléculas, apesar de terem apresentado massas
moleculares diferentes são consideradas acemananas. Em paralelo, aos
estudos realizados por Lee et al. (2001), que, conforme descrito anteriormente,
isolaram uma acemanana de 500 kDa, Pugh et al. (2001), realizando estudos
de obtenção de polissacarídeos para a mesma espécie, também obtiveram um
polissacarídeo de massa semelhante (500 kDa), isolado a partir da resina
Shodex Ohpak KB-800, que foi nomeada de aloerídeo. Posteriormente,
Strickland e Pelley (2004), reuniram todos os relatos científicos obtidos da
literatura sobre acemanana e não concordaram com as afirmações de que
polissacarídeos de diferentes massas moleculares fossem considerados
acemanana.
Além disso, é citado na literatura que, além do procedimento de
exclusão molecular, para obtenção de polissacarídeos de A. barbadensis, estes

78
também foram obtidos pelo uso de cromatografia de troca iônica em matriz de
DEAE. Como por exemplo, t’Hart et al. (1989) foram os primeiros autores a
relatarem o isolamento de polissacarídeos de A. barbadensis, por DEAE-
celulose. Esses autores sugeriram que o PNC obtido era rico em resíduos de
manose e isento de ácidos urônicos. Outro grupo de pesquisadores utilizou o
procedimento de DEAE-celulose, somente com intuito de eliminar
contaminantes carregados negativamente dos PT de A. barbadensis (IM et al.,
2005; YAO et al., 2009).
No entanto, Young e Thompson (2005) isolaram de A. barbadensis uma
fração contendo PNC, através de cromatografia de troca iônica em matrizes de
DEAE-Sephadex G-25 e DEAE-celulose, realizadas separadamente. Os perfis
cromatográficos mostraram-se similares, sendo obtido somente uma única
fração de PNC, de massa molecular média de 50 kDa. Esses dados
corroboram com os de Chang, Feng e Wang (2011) e Chang, Chen e Feng
(2011).
Diferentemente dos resultados obtidos para obtenção de somente uma
única fração de PNC, tanto por DEAE-celulose como por DEAE-Sephadex G-
25, Chun-hui et al. (2007), ao submeterem a planta halófita A. barbadensis ao
processo de irrigação com água do mar, obtiveram duas frações de PNC, a
partir da aplicação de seus PT ao processo de cromatografia de troca iônica
em matriz de DEAE-52.
Adicionalmente, Aquino, Gravitol e Mourão (2011), realizaram estudos
com espécies de plantas halófitas, cultivadas na ausência e presença de
estresse salino, com a finalidade de avaliar a presença de polissacarídeos
sulfatados. Os resultados obtidos nesse estudo mostraram que os PT extraídos
da espécie Ruppia marítima, submetida ao estresse salino, ao serem
fracionados por cromatografia de exclusão molecular em coluna Superose 12,
o perfil cromatográfico obtido apresentou duas diferentes frações, sendo a de
massa molecular maior apresentando polissacarídeos sulfatados e a de menor
massa molecular contendo ácidos urônicos. Adicionalmente, os autores
sugerem que a presença de polissacarídeos sulfatados em plantas é uma
adaptação da planta a altas concentrações de sais no ambiente.

79
2.2.3. Caracterização Físico-química
2.2.3.1. Análises Químicas Colorimétricas
Os carboidratos totais (CT) presentes nos PT e nas frações PI e
PII foram quantificadas a partir do método descrito por Dubois et al.
(1956), sendo obtidas percentagens de 58,50; 65,65; e 27,74%,
respectivamente. O teor de proteínas solúveis contaminantes
apresentaram valores de 1,2; 0,5; e 0,8%, para as respectivas frações.
Para averiguar quais tipos de polissacarídeos estariam envolvidos na
interação da fração PII obtida na matriz de DEAE-celulose, foram
quantificados ácidos urônicos e sulfato livre para as diferentes frações.
Os ácidos urônicos dos PT e das frações PI e PII apresentaram
percentagens de 9,9; 10,0; e 8,0% (m/m), respectivamente. Em adição, o
teor de sulfato foi quantificado, correspondendo a 1,2; 0,0; e 12,0%
(m/m), referentes aos PT e às frações PI e PII, respectivamente.
Ácidos urônicos são muito comuns em polissacarídeos de plantas
(DUMVILLE; FRY, 2000; FANG; JIANG; WANG, 2006; GARRON;
CYGLER, 2010; MELTON; SMITH, 2001), inclusive em A. barbadensis
(NI; YATES; TIZARD, 2004; REYNOLDS; DWECK, 1999). Em contraste,
radicais sulfato são muito raros em plantas, sendo encontrados somente
em halófitas submetidas a estresse salino (AQUINO; GRATIVOL;
MOURÃO, 2011). Luta, Duncan e Sinnott (2009) encontraram sulfato no
extrato bruto total da folha (casca + polpa) de A. barbadensis, chegando
a concentrações que variaram de 1000 a 10000 ppm. No entanto, não
foram realizados estudos posteriores para elucidar se o sulfato detectado
fazia, ou não, parte da estrutura dos polissacarídeos. Na Tabela 2.1
podem ser observados os dados da composição química dos PT e das
frações PI e PII obtidas neste trabalho e, na Tabela 2.2, a relação dos
dados encontrados na literatura.

80
Tabela 2.1. Composição química de PT e das frações PI e
PII obtidos de folhas de Aloe barbadensis.
Frações Composição Química (% m/m)
PrTta CTsb AHUsc Sfd Sf/CTse
PT 1,20 58,50 9,90 1,20 0,02
PI 0,50 65,65 10,00 0,00 0,00
PII 0,80 27,74 8,00 12,00 0,32a: Proteínas Totais (PrTt);b: Carboidratos totais (CTs);c: Ácidos urônicos (AHUs);d: Sulfato (Sf);e: Razão em massa de sulfato/carboidratos totais (Sf/CTs)

81
Tabela 2.2. Composição química de frações de espécies do gênero Aloe.
Frações Espécies Composição Química (% m/m) ReferênciasPrTt CTsa AUsb Sfc Sf/CTsd
AIR Cascae A. barbadensis 7,56 60,34 14,05 n.d n.dAIR Gele 15,40 72,17 13,00 n.d n.d Femenia et al. (1999)
AcemananaSulfatada f A. barbadensis n.d. n.d. n.d. 30,00 n.d Lee et al. (2001)
AIR Gelf A. barbadensis 15,10 63,37 12,93 n.d n.d Femenia et al. (2003)PAC-Ig A. barbadensis n.d. 98,30 n.d. n.d. n.d.PAC-IIg (A. vera L. n.d. 98,07 n.d. n.d. n.d. Leug et al. (2004)PAC-IIIg var. chinensis) n.d. 95,63 n.d. n.d. n.d.BSWh A. barbadensis 1-2 90,00 n.d. n.d. n.d. Chow et al.(2005)APSi A. barbadensis n.d. 97,00 n.d. n.d. n.d. Yao et al. (2009)PT polpap <1 83,10 n.d. n.d. n.d.GN1j <1 n.d. - n.d. n.d.GN2j <1 n.d. - n.d. n.d.GA1k <1 n.d. 28,6 n.d. n.d.GA2k <1 n.d. n.d. n.d. n.d.PT cascap A. arborescens <1 1,10 n.d. n.d. n.d. Chang, Feng e WangSN1l <1 n.d. - n.d. n.d. (2011)SA1m <1 n.d. 31,20 n.d. n.d.SA2m <1 n.d. n.d. n.d. n.d.SA3m <1 n.d. n.d. n.d. n.d.PT floresp <1 28,70 n.d. n.d. n.d.FN1n <1 n.d. - n.d. n.d.FA1o <1 n.d. n.d. n.d. n.d.FA2o <1 n.d. 16,40 n.d. n.d.PT polpap <1 58,30 n.d. n.d. n.d.GN1j <1 n.d. - n.d. n.d.GN2j <1 n.d. - n.d. n.d.GA1k <1 n.d. 22,8 n.d. n.d.GA2k <1 n.d. 10,8 n.d. n.d.PT cascap A. barbadensis <1 37,30 n.d. n.d. n.d. Chang, Chen e FengSN1l <1 n.d. - n.d. n.d. (2011)SN2l <1 n.d. - n.d. n.d.SA1m <1 n.d. 21,40 n.d. n.d.SA2m <1 n.d. 18,60 n.d. n.d.PT floresp <1 23,40 n.d. n.d. n.d.FN1n <1 n.d. - n.d. n.d.FA1o <1 n.d. 20,80 n.d. n.d.
a: Carboidratos totais;b: Ácidos urônicos;c: Sulfato;d: Razão em massa de sulfato/carboidratos totais

82
(cont. Tabela 2.1)e: Fração de resíduos insolúveis em álcool (AIR);f: Acemanana sulfatada sinteticamente;g: Frações obtidas por cromatografia de filtração em gel acoplada a HPLC;h: Polissacarídeos solúveis em água (acemanana filtrada);i: Fração de resíduos insolúveis em álcool;j: Polissacarídeos neutros do gel;k: Polissacarídeos ácidos do gel (negativamente carregados);l: Polissacarídeos neutros da casca;m: Polissacarídeos ácidos da casca (negativamente carregados);n: Polissacarídeos neutros das flores;o: Polissacarídeos ácidos das flores (negativamente carregados);p: Polissacarídeos totais;n.d.: não determinado;– : ausente.
2.2.3.2. Eletroforese em Gel de Poliacrilamida (PAGE)
Os PT e as frações PI e PII foram submetidas a PAGE, para
determinação da massa molecular de polissacarídeos sulfatados. A
coloração das bandas obtidas foi realizada com azul de toluidina, corante
específico para compostos metacromáticos. As mobilidades
eletroforéticas das bandas obtidas foram comparadas com as de
diferentes marcadores moleculares. A fração PII mostrou migração
polidispersa e a massa molecular média aparente foi avaliada em 77 kDa
(Figura 2.4 A). A polidispersão observada pode ser atribuída à
heterogeneidade de massa molecular, característica comum de
polissacarídeos sulfatados (CINELLI, 2009; RODRIGUES, 2011).

83
Figura 2.4: Eletroforese em gel de poliacrilamida (PAGE) dos PT e das
frações PI e PII. (A) Coloração por azul de toluidina; (B) Coloração por
stainsall. M – Padrões dos GAGs dextram sulfato de baixa massa
molecular (~8 kDa) (M1), condroitim-4-sulfato (M2), condroitim-6-sulfato
(M3) e dextram sulfato de alta massa molecular (~500 kDa) (M4).
Como o resultado da PAGE sugere presença de sulfato na fração
PII, foi realizada nova eletroforese, sendo o gel corado com o reagente
stainsall (Figuras 2.4 B). De acordo com alguns autores, stainsall tem
sido utilizado tanto para corar polissacarídeos sulfatados quanto para
aqueles ricos em ácidos urônicos negativamente carregados (COPPA et
al., 2010; HIROKAWA; FUJIWARA; TSUZUKI, 2005; ORMISTON et al.,
2010; MARSH; CHANG; KING, 1992; VOLPI; MACCARI, 2007; VOLPI;
MACCARI; TITZE, 2005). Polissacarídeos sulfatados podem ser
detectados no gel pela coloração amarelada, amarronzada, avermelhada,
azul escuro ou, em geral, arroxeada (ORMISTON et al., 2010). Em
contraste, polissacarídeos poli-urônicos são apresentados com coloração
ciana ou lilás (ORMISTON et al., 2010; RIGOUIN et al., 2009).
A Figura 2.4 B mostra que PII continua apresentando uma
mobilidade polidispersa, o que também é observado para os PT. No
entanto, PII apresentou dois tipos de coloração, roxo e ciano. Esses

84
resultados sugerem a presença de polissacarídeos ricos em sulfato e em
ácidos urônicos, respectivamente (ORMISTON et al, 2010). Apesar de PT
e PI apresentarem percentagens de ácidos urônicos superiores a PII, não
foram observadas bandas eletroforéticas de cor ciana para essas frações.
De acordo com esses dados, pode-se sugerir que os ácidos urônicos
presentes nos PT e na fração PI estejam metil-esterificados, justificando,
assim, a não coloração com reagente stainsall e a não adsorção de PI na
matriz de DEAE.
2.2.3.3. Cromatografia de Troca Iônica em Coluna de Mono-q
Acoplada a HPLC
Com intuito de verificar a pureza da fração PII, foi realizada uma
cromatografia de troca iônica em coluna de Mono-q acoplada a um
sistema de HPLC. Um pico bem definido foi observado, com intensidade
de, aproximadamente, 400 mV, sendo obtido quando a amostra foi eluída
com um tampão em gradiente de concentração de 0,5 à 3 M de NaCl
(Figura 2.5). Além disso, dois picos pequenos de intensidade de 51 e 52
mV foram obtidos durante o processo de eluição por gradiente de
concentração. Apesar desses picos menores estarem presentes,
correspondem juntos apenas a 10,0% da área total dos picos, o que faz
com que as propriedades eletroforéticas em gel de poliacrilamida sejam
relacionadas ao pico majoritário. É sugerido que, apesar dos
contaminantes, não são dois grupos moleculares comigrando durante a
aplicação do campo elétrico, mas, sim, a um único grupo de moléculas
que apresentaram, simultaneamente, grupos sulfato e grupos
carboxilados.
Aquino, Gravitol e Mourão (2011), em estudo realizados com as
plantas halófitas Halodule wrightii e R. marítima cultivadas em ambientes
salinos de 38 e 15 ppt de sal, respectivamente, observaram que seus PT
quando aplicados em cromatografia de troca iônica em matriz de Mono-

85
Q, apresentaram um pico bem definido de polissacarídeos sulfatados,
eluído na concentração de NaCl 0,5M. Esses resultados corroboram com
os encontrados no presente estudo, visto que A. barbadensis também é
uma planta halófita,
Figura 2.5: Purificação dos polissacarídeos sulfatados de Aloe
barbadensis por cromatografia de troca iônica em matriz Mono-q,
acoplada a sistema de (HPLC), em gradiente de concentração de 0,5 – 3
M de NaCl (linha pontilhada).

86
2.2.3.4. Eletroforese em Gel de Agarose
Os PT e as frações PI e PII obtidos de DEAE-celulose ao serem
submetidos à eletroforese em gel de agarose, corado com azul de
toluidina, foi visualizada no início do procedimento de descoramento, para
PII, uma banda de coloração rósea que desapareceu completamente no
final do processo de descoramento (dado não mostrado). Este fato pode
ser justificado pela menor presença de grupos sulfatos na estrutura
química dos polissacarídeos na estrutura da molécula, conforme também
demonstrado por Pereira et al. (2005), para polissacarídeos sulfatados
de Gelidium crinale. Outra justificativa seria a interferência de grupos
carboxilados na mesma molécula, conforme demonstrado por Ormiston et
al. (2010), ao investigar o padrão de degradação de GAGs em ratos,
durante infecções pulmonares.
Com a finalidade de uma melhor visualização dos polissacarídeos
presentes em PII, foi realizado outro procedimento de eletroforese em gel
de agarose corado com stainsall (Figura 2.6 A e B). O resultado obtido
mostrou, no início do descoramento, duas bandas homogêneas de
colorações distintas, roxa e ciana, correspondentes a grupos sulfatos e
carboxilados, respectivamente (Figura 2.6 A), cujas bandas mostraram-se
com coloração roxa no final do descoramento (Figura 2.6 B).

87
Figura 2.6: Eletroforese em gel de agarose 0,5% da fração de polissacarídeoseluída com Tampão acetato de sódio 50 mM, pH 6,0, contendo 0,5M de NaCl(PII). (A) Coloração com stainsall (início do processo de descoramento). (B)Coloração com stainsall (final do processo de descoramento). M – Padrões dosglicosaminoglicanos (GAGs) heparam sulfato (HS), dermatam sulfato (DS) econdroitim sulfato (CS).

88
Esses resultados corroboram com os encontrados em PAGE,
corado com stainsall, onde a banda roxa é indicativa de grupos sulfato, e
a de coloração ciana, indicativa de grupos carboxilados, que comigraram
juntas no processo eletroforético, justificando a presença de uma única
molécula polissacarídica. Nesse procedimento, também foi observado,
que a banda eletroforética obtida migrou com a mesma intensidade do
padrão heparam sulfato. Isso pode ser justificado por o heparam sulfato
apresentar maior carga líquida negativa, em comparação aos outros
padrões utilizados, dermatam sulfato e condroitim sulfato. De acordo com
Dietrich e Dietrich (1976), no procedimento em eletroforese em gel de
agarose, a migração dos polissacarídeos sulfatados depende da sua
interação com grupos NH3+ presentes no tampão de corrida.
2.2.3.5. Análise da Composição Monossacarídica das Frações
PI e PII por GC-MS
Para melhor compreensão da composição monossacarídica das
frações PI e PII, estas foram submetidas à GC-MS. Os perfis
cromatográficos obtidos apresentaram 4 sinais com tempo de retenção
similares aos monossacarídeos padrões manose, glucose, galactose
correspondendo a 78,4; 7,3 e 2,1%, respectivamente (Figura 2.7 A) e
similar ao padrão fucose correspondendo a 2,8% (Figura 2.8 A). Os
espectros de fragmentação confirmaram a presença desses açúcares
(Figura 2.7 B,C,D; Figura 2.8 B). Em suma, para a fração PI, a proporção
de Man:AHU:Glu:Gal:Fuc foi de 39:5:4:1:1.
Em relação à determinação da composição monossacarídica da
fração PII, foram obtidos 5 sinais similares aos padrões de manose,
glucose e galactose, correspondendo a 39,2; 22,2; e 26,3%,
respectivamente (Figura 2.9 A), arabinose e xilose, correspondendo a
3,8% e 1,1%, respectivamente (Figura 2.10 A). Os espectros de
fragmentação confirmaram a presença desses açúcares (Figura 2.9;

89
Figura 2.10). Resumidamente, para a fração PII, a proporção de
Man:Gal:Glu:AHU:Ara:Xil foi de 20:13:11:4:2:1.
A avaliação das massas molares de ácidos urônicos presentes nas
frações PI e PII, foi obtida através da razão entre açúcares simples
neutros obtidos por GC-MS com os açúcares ácidos obtidos por análises
químicas (subitem 2.2.3.1). Na Tabela 2.3 pode ser observada a
comparação dos dados obtidos nesse trabalho com outros da literatura.
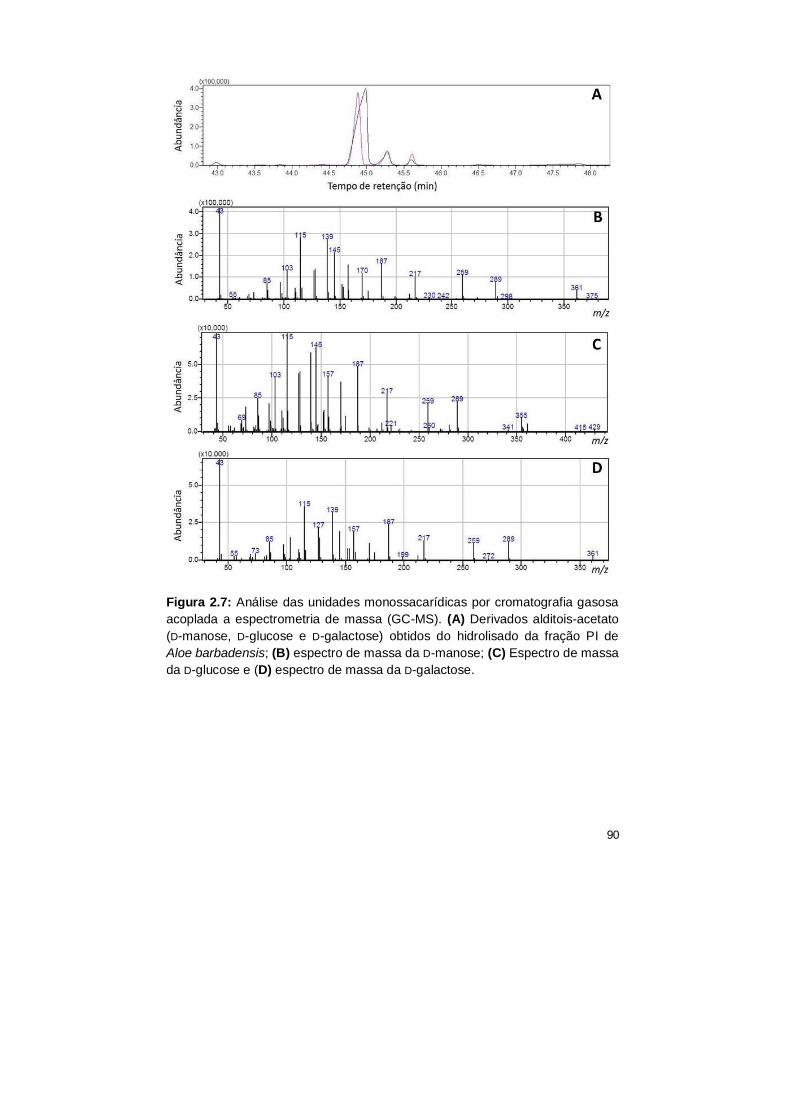
90
Figura 2.7: Análise das unidades monossacarídicas por cromatografia gasosaacoplada a espectrometria de massa (GC-MS). (A) Derivados alditois-acetato(D-manose, D-glucose e D-galactose) obtidos do hidrolisado da fração PI deAloe barbadensis; (B) espectro de massa da D-manose; (C) Espectro de massada D-glucose e (D) espectro de massa da D-galactose.

91
Figura 2.8: Análise das unidades monossacarídicas por cromatografia gasosaacoplada a espectrometria de massa (GC-MS). (A) Derivados alditois-acetatode L-fucose obtidos do hidrolisado da fração PI de Aloe barbadensis; (B)espectro de massa da L-fucose.

92
Figura 2.9: Análise das unidades monossacarídicas por cromatografia gasosaacoplada a espectrometria de massa (GC-MS). (A) Derivados alditois-acetato(D-manose, D-glucose e D-galactose) obtidos do hidrolisado da fração PII deAloe barbadensis; (B) espectro de massa da D-manose; (C) Espectro de massada D-glucose e (D) espectro de massa da D-galactose.

93
Figura 2.10: Análise das unidades monossacarídicas por cromatografia gasosaacoplada a espectrometria de massa (GC-MS). Derivados alditois-acetato de L-arabinose (A) e D-xilose (C) obtidos do hidrolisado da fração PII de Aloebarbadensis; (B) espectro de massa da L-arabinose; (D) Espectro de massa daD-xilose.

94
Tabela 2.3: Composição de monossacarídeos das frações PI e PII de Aloe barbadensis
comparadas com as obtidas para as espécies A. barbadensis e A. arborescens por outros
autores.
Frações Espécies Composição Monossacarídica (% mol) ReferênciasMana Gala Glua Araa Xila Fuca
PI A. barbadensis 78,4 2,1 7,3 - - 2,8 Presente TrabalhoPII 39,2 26,3 22,2 3,8 1,1 -AIR Casca A. barbadensis 30,1 8,4 2,9 5,9 11,7 2,5AIR Gel 52,8 3,5 5,3 1,2 1,4 0,6 Femenia et al. (1999)Aloerídeo A. barbadensis 19,5 23,9 37,2 10,3 - - Pugh et al. (2001)AIR Gel A. barbadensis 26,4 3,9 5,7 0,9 2,6 0,5 Femenia et al. (2003)BSW A. barbadensis 83,9 4,4 6,2 1,4 0,7 1,5 Chow et al.(2005)GAPS-1b 96,0 2,4 1,6 - - -SAPS-1b A. barbadensis 88,9 0,3 10,8 <0,1 1,0 - Chun-hui et al. (2007)AVG Pico 1c 99,0 - 1,0 - - -AVG Pico 2c 47,0 8,0 42,0 - 4,0 - Luta, Ducan e SinnottAVG Pico 3c A. barbadensis <1 - 75,0 - - - (2009)AVG pico 4c - - 100,0 - - -GN1 15,3 3,1 81,6 - - -GN2 13,6 5,4 81,0 - - -GA1 2,0 37,5 30,6 - 1,3 -GA2 n.d n.d. n.d. - n.d -SN1 6,9 63,7 29,4 - - - Chang, Feng e WangSA1 A. arborescens 21,8 11,0 27,3 - 8,7 - (2011)SA2 n.d. n.d. n.d. - n.d. -SA3 n.d. n.d. n.d. - n.d. -FN1 23,5 31,0 40,1 - 3,3 -FA1 n.d. n.d. n.d. - n.d. -FA2 14,6 29,8 20,1 - 7,9 -GN1 82,5 - 17,5 - - -GN2 82,9 - 17,1 - - -GA1 25,2 21,8 16,8 - 5,0 -GA2 26,2 33,7 23,6 - 3,4 - Chang, Chen e FengSN1 83,3 - 16,7 - - - (2011)SN2 A. barbadensis 83,1 - 16,9 - - -SA1 12,4 11,2 45,3 - 1,2 -SA2 14,0 15,3 42,5 - 2,0 -FN1 24,8 37,2 27,3 - 24,8 -FA1 12,8 27,9 25,7 - 12,8 -
a: manose (Man), glucose (Glu), galactose (Gal), arabinose (Ara), Fucose (Fuc) e Xilose (Xil).b: Polissacarídeos isolados do gel (GAPS-1) e casca (SAPS-1) através de DEAE-52.c: Frações obtidas de cromatografia de exclusão molecular acoplada a HPLC.n.d.: não determinado;– : ausente.

95
2.2.4. Análises Estruturais
2.2.4.1. Espectroscopia de Absorção na Região do
Infravermelho
Com o objetivo de analisar os grupamentos funcionais presentes
nas estruturas dos PT e das frações PI e PII, os mesmos foram
submetidos à espectroscopia de absorção na região do infravermelho
(IR), transformada por Fourier.
O perfil espectroscópico obtido, para os PT e para as frações PI e
PII, apresentou sinais em 3430,7; 2930,7 e 2888,8 onda.cm-1,
respectivamente, que representam estiramentos assimétricos para o
grupo hidroxila, a ligação C−H, e para os grupos −CH3 e −CH2−,
presentes em compostos alifáticos (Figura 2.11) (LAMBERT et al., 2007).
Os sinais característicos do estiramento de ligação de ácidos
urônicos foram detectados nos PT e na fração PI (Figura 2.12). Os PT e a
fração PI, mostraram um sinal ~1738 onda.cm-1 que corresponde ao
estiramento da ligação C−O de carboxilas metil-esterificadas (FANG;
JIANG; WANG, 2006). Os PT e a fração PI também apresentaram sinais
de ácidos urônicos carregados negativamente (1635,9 e 1426,9 onda.cm-
1). Apesar da fração PI ter apresentado estes sinais, esta não interagiu
com a matriz DEAE-celulose, durante o processo de cromatografia de
troca iônica, assim como, não apresentou uma banda quando submetida
aos procedimentos de PAGE e eletroforese em agarose, cujos géis foram
corados com o corante stainsall. Isso pode ser justificado pelo fato do
sinal obtido na região de ~1635 onda.cm-1, que conforme descrito acima,
corresponde ao estiramento de ácidos urônicos carregados
negativamente, este sinal também, pode estar relacionado ao grupo C=O
de aldeídos de cadeias alifáticas (CAMPESTRINI, 2007). Dessa forma,
podemos sugerir que os ácidos urônicos presentes na fração PI
encontram-se na forma metil-esterificados.
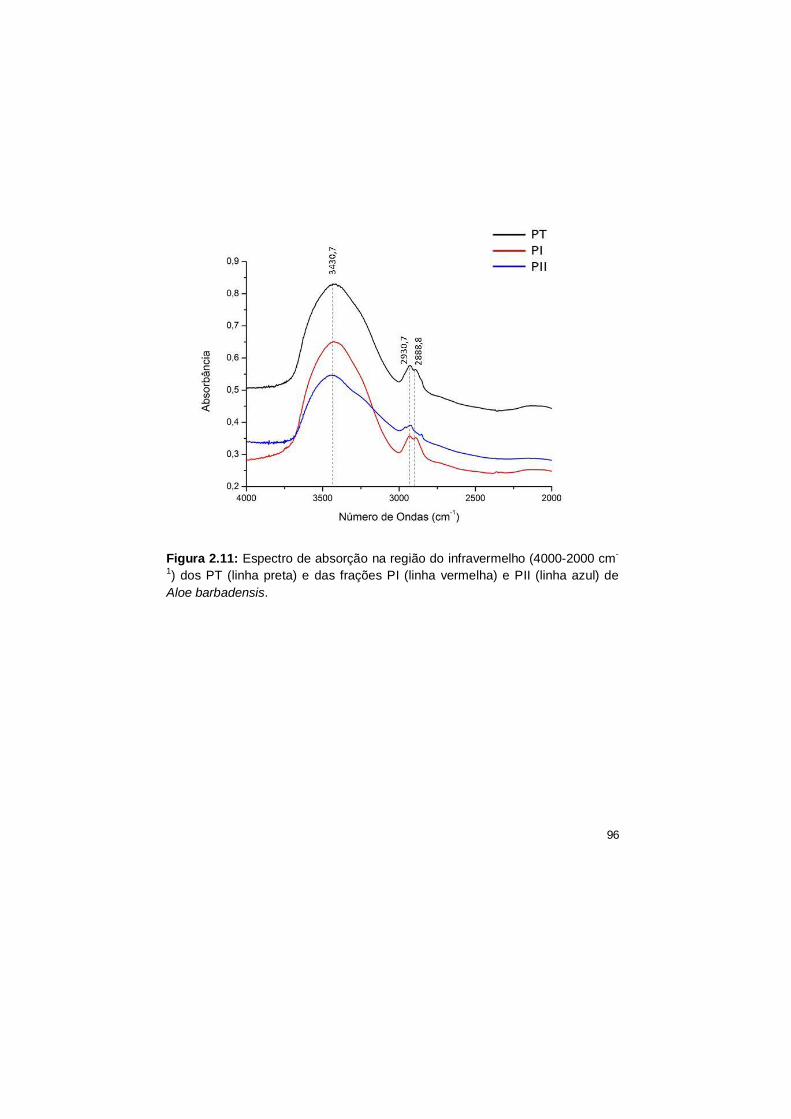
96
Figura 2.11: Espectro de absorção na região do infravermelho (4000-2000 cm-
1) dos PT (linha preta) e das frações PI (linha vermelha) e PII (linha azul) deAloe barbadensis.
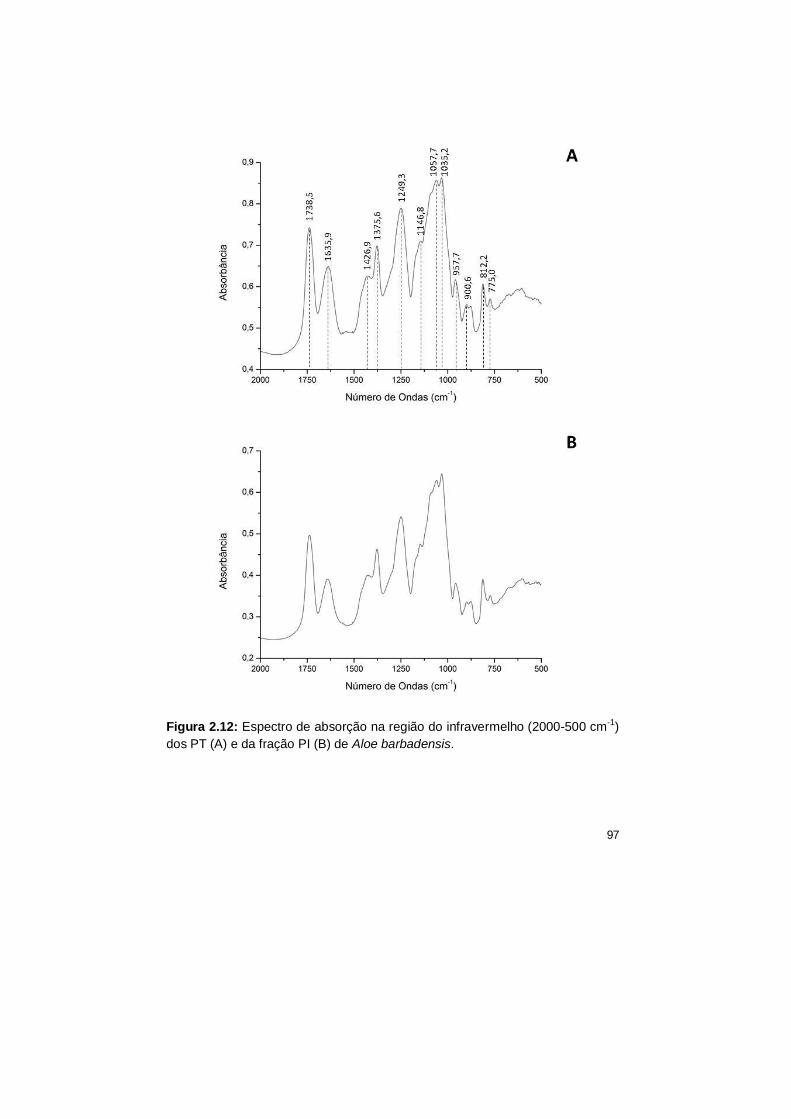
97
Figura 2.12: Espectro de absorção na região do infravermelho (2000-500 cm-1)dos PT (A) e da fração PI (B) de Aloe barbadensis.

98
Na avaliação dos dados espectrais da fração PII (Figura 2.13), os
resultados obtidos apresentaram sinais 1421,1-1741,6 onda.cm-1, onde o
sinal ~1635 onda.cm-1 apresentou uma intensidade maior do que o sinal
1741,6 onda.cm-1, indicando a presença de ácidos urônicos
negativamente carregados. Isso justifica sua interação com o trocador
aniônico da matriz de DEAE e sua coloração com stainsall, conforme
descrito anteriormente. Em adição, o espectro de IR da fração PII sugere,
ainda, a presença de grupamentos funcionais sulfatados. De acordo com
Prado-Fernandez et al. (2003), sinais de 1329,5 e 1260,9 onda.cm-1
podem estar relacionados a presença de grupos éster sulfato (O=S=O).
Além disso, nos PT e nas frações PI e PII foram detectados picos
entre ~1035-1047 onda.cm-1 (Figuras 2.12 e 2.13). De acordo com
Prashanth et al. (2006), esses sinais estão relacionados ao estiramento
da ligação C−O quando ligados a grupos C−O−C, indicando presença de
grupos acetilados. Ni, Yates e Tizard (2004) resumiram as evidências,
presentes na literatura, sobre a presença do grupo acetil em
polissacarídeos do gênero Aloe. Esse processo acontece primariamente
nos resíduos monossacarídicos de manose e, secundariamente, nos de
glucose. Nos PT e na fração PI, foram obtidos sinais espectrais de 812,2
e 1057,7 onda.cm-1, que são característicos de resíduos de manose
(Figura 2.12). Na fração PII, foi observado um sinal de 1014,7 onda.cm-1,
que também corresponde a resíduos de manose e um sinal 1078,2
onda.cm-1, correspondente a resíduos de galactose (Figura 2.13). Esses
dados corroboram com os relatados obtidos para a mesma espécie de
planta, por Chang, Chen e Feng (2011).
A intensidade de bandas por volta de 1375, 1249 e 1146 onda.cm-1
foram obtidas nos PT e nas frações PII e podem estar relacionadas aos
grupos −CH3, C−O−C e ligação glicosídica, respectivamente (CHUN-HUI
et al., 2007; GU et al., 2010; KAÈURÁKOVÁ et al., 2000). Em adição, o
sinal ~960 onda.cm-1 encontrado, pode corresponder a anéis
piranosídicos e o intervalo ~900-915 onda.cm-1 pode estar relacionado a
configuração â (CHUN-HUI et al., 2007). A Figura 2.14 apresenta um

99
comparativo dos espectros de IR, entre 2000 e 500 onda.cm-1, das três
frações estudadas.
Figura 2.13: Espectro de absorção na região do infravermelho (2000-500 cm-1)da fração PII de Aloe barbadensis.

100
Figura 2.14: Comparação dos espectros de absorção dos PT (linha preta) edas frações PI (linha vermelha) e PII (linha azul) de Aloe barbadensis na regiãodo infravermelho (2000-500 cm-1).

101
2.2.4.2. Ressonância Magnética Nuclear (RMN)
Com o intuito de determinar parcialmente a estrutura dos
polissacarídeos sulfatados da fração PII e constatar a presença de sulfato na
mesma, foram empregados experimentos de espectroscopia de RMN
unidimensional de 1H e bidimensional heteronuclear 1H/13C DEPT-HSQC.
Os espectros de 1H-RMN de PII nativa (sulfatada) (Figura 2.15 A)
mostram sinais similares aos encontrados por Leung et al. (2004), para espécie
A. vera L. var. chinensis (Haw.) Berg. De acordo com os mesmos autores, os
espectros entre äH 4,2 e 3,6 ppm representam os prótons dos resíduos de â-
manopiranose, o que corrobora com os dados encontrados nas análises
químicas e de IR, no presente estudo. Em adição, nesse espectro, foram
observados o sinal äH ~5,1 ppm, referentes à á-D-glucopiranose, e o sinal äH
~3,8 ppm, relacionado a ácidos urônicos negativamente carregados (COO-).
Além disso, foi demonstrado ausência do sinal äH 4,3 ppm que indica a
ausência de ácidos urônicos neutros metil-esterificados. Esses resultados
corroboram com os obtidos por McConaughy et al. (2008), para as pectinas de
A. barbadensis.
Na avaliação da região anomérica (äH 5,3-4,1 ppm) para fração PII
(Figura 2.16 A), esta região mostrou-se diferente quando comparada com o
espectro de 1H RMN obtido por Leung et al. (2004), para espécie A. vera L. var.
chinensis (Haw.) Berg. Nesta região foram observados os sinais äH ~5,2; 5,0;
4,9; 4,7; 4,45 e 4,4 ppm que se mostraram, portanto, ausentes no espectro
obtido da espécie A. vera L. var. chinensis (Haw.) Berg. Assim, os três
primeiros sinais podem estar relacionados a ácido á-glucurônico (KANG et al.,
2011), ácido á-galacturônico (TANAKA et al., 2010) e â-L-arabinose
(JAHANBIN et al., 2011), respectivamente, reforçando, portanto os resultados
positivos encontrados nas análises físico-químicas de determinação de ácidos
urônicos e determinação da composição monossacarídica, que mostrou a
presença de resíduos de arabinose. O pico mais intenso (äH 4,7 ppm) foi
atribuído aos prótons de H2O. Foram observados ainda sinais de prótons äH
4,45 e 4,4 ppm, que podem representar uma mistura de H-1 dos â-anômeros

102
de resíduos de galactopiranose e, sinais H-4 do anel de hexose (POMIN,
2008).
Figura 2.15: Espectros 1D de 1H-RMN em 800 MHz da fração PII nativa (A) e
dessulfatada (B) de Aloe barbadensis.

103
Figura 2.16: Espectros 1D de 1H-RMN em 800 MHz dos sinais
correspondentes aos prótons anoméricos de polissacarídeos da fração PII
nativa (A) e dessulfatada (B) de Aloe barbadensis.

104
Como pode ser observado nas Figuras 2.15 B e 2.16 B, os espectros da
região anomérica de PII dessulfatada diferem dos espectros da nativa. Na
fração PII dessulfatada foi observado um único sinal deslocado de äH 4,5 ppm,
diferindo, portanto, da nativa que apresentou dois sinais de intensidades äH
4,45 e 4,4 ppm. Além disso, na fração dessulfatada não foi observada a
presença do sinal äH 4,9 ppm, uma diminuição nas intensidades dos sinais
entre äH 4,2 e 5,0 ppm e o aparecimento de dois outros sinais no intervalo äH
5,4 e 5,6 (Figura 2.16 B). Todas essas evidências são indicativas da presença
de sulfato na estrutura dos polissacarídeos de PII nativa. Como os dados de1H-RMN não foram suficientes para confirmação da presença de sulfato na
estrutura dos polissacarídeos presentes na fração PII nativa, tornou-se
necessário a análise dessa fração por RMN bidimensional heteronuclear 1H/13C
DEPT-HSQC.
A análise comparativa dos espectros 2D heteronuclear 1H/13C DEPT-
HSQC das fração PII nativa (Figura 2.17) e dessulfatada (Figura 2.18) de A.
barbadensis, foi observado que alguns sinais presentes na fração nativa,
mostraram-se ausentes na fração dessulfatada.
Para análise mais detalhada da estrutura dos polissacarídeos presentes
na fração PII, torna-se necessário que estudos posteriores sejam realizados
utilizando-se as técnicas de RMN 2D homonucleares (COSY e TOCSY), com
intuito de assinalar os deslocamentos químicos.

105
Figura 2.17: Espectros 2D heteronuclear 1H/13C DEPT-HSQC da fração PII
nativa de Aloe barbadensis.

106
Figura 2.18: Espectros 2D heteronuclear 1H/13C DEPT-HSQC da fração PII
dessulfatada de Aloe barbadensis.

107
2.5. CONCLUSÕES
A extração aquosa (70 °C) de polissacarídeos totais (PT) de A.
barbadensis, seguido de precipitação dos PT por etanol e remoção dos
contaminantes proteicos com TCA apresentou um rendimento de 3,4%.
A cromatografia de troca iônica em DEAE-celulose foi eficiente no
fracionamento dos PT, onde foram obtidas as frações PI e PII.
A caracterização físico-química mostrou que a fração PI é composta do
resíduo monossacarídico majoritário manose (78,4%), dos resíduos glucose,
galactose, fucose e ácidos urônicos e isenta de grupos éster sulfato. Enquanto,
a fração PII é constituída por resíduos monossacarídicos de manose, glucose,
galactose, arabinose, xilose, ácidos urônicos e grupos éster sulfato.
Na revelação das bandas polissacarídicas da fração PII, obtidas por
PAGE e gel de agarose corados com azul de toluidina e stainsall foi constatada
a presença de duas bandas que apresentaram diferentes colorações, roxa e
ciana, correspondentes à presença de grupos sulfato e carboxilados,
respectivamente.
Na análise estrutural das frações PI e PII, por espectroscopia no IR, foi
demonstrado que a fração PI apresenta unidades monossacarídicas de â-
manose O-acetiladas e ácidos urônicos neutros em sua estrutura.
Diferentemente, a fração PII, mostrou-se ser constituída por unidades
monossacarídicas de manose, galactose, ácidos urônicos carregados
negativamente e éster sulfato.
Na avaliação estrutural da fração PII por RMN foi comprovada a
presença do grupo éster sulfato, onde análises estruturais posteriores deverão
ser realizadas para confirmação da posição desses grupos nas unidades
monossacarídicas.
A obtenção, caracterização físico-química e estrutural dos
polissacarídeos de A. barbadensis e, a detecção de grupos ésteres sulfato em
sua estrutura, até o presente não citada na literatura, é de grande valia para

108
estudos posteriores para detecção e investigações estruturais de outros
polissacarídeos sulfatados presentes em outras espécies do gênero Aloe,
assim como em outras espécies de plantas superiores.

109
Capítulo 3Avaliação das Atividades
Antiviral, Prócoagulante e Anti-hemorrágica e Toxicidade

110
3.1. MATERIAIS E MÉTODOS
3.1.1. CITOTOXICIDADE
3.1.1.4. Culturas de Células
Células HEp-2 (linhagem de células provenientes de carcinoma de
laringe humana) e células HeLa (carcinoma cervical humano) foram mantidas
em meio mínimo essencial de Eagle (Sigma), modificado por Dulbecco (DMEM-
Eagle), acrescido de 10% de soro fetal bovino (SFB) e incubadas a 37 °C a 5%
de CO2.
Células Vero [linhagem de células de rim de macaco verde africano
(Cercopithecus aethiops)] foram mantidas em meio mínimo essencial de Eagle
(MEM - Eagle). Posteriormente, foram suplementados com 2 mM de L-
glutamina (Sigma), 50 µg/mL de garamicina (Shering-Plough), 2,5 µg/mL de
fungizona (Gibco), 10% de SFB e mantidas a 37°C, em atmosfera contendo 5%
de CO2.
Células LLC-MK2 [linhagem de células de rim de macaco rhesus
(Macaca mulata)] foram crescidas em DMEM-Eagle suplementado com 3 mM
L-glutamina, 50 mg/mL de garamicina, 2,5 mg/mL de fungizona, bicarbonato de
sódio a 0,25%, 10% de soro fetal bovino (SFB) e mantidas a 37 °C em
atmosfera contendo 5% de CO2.
Células C6/36 (linhagem de células de larvas de mosquito Aedes
albopictus) foram mantidas em meio L-15 (Leibovitz) + RPMI (1:1) e, em
seguida, suplementado com 0,02% de glutamina, 1% de MEM (solução de
aminoácidos não-essenciais) e 5% de SFB.
Todos os meios de cultura foram submetidos à filtração em membrana
Millipore (0,22 µm), transferidos para recipientes previamente estéreis e
mantidos a 4°C, para estocagem. Além disso, as células foram crescidas em
garrafas de vidro especializadas em cultura celular, contendo meio de cultura,
até que a confluência da monocamada preenchesse pelo menos 80% de uma
das faces da garrafa de cultivo celular. As condições de temperatura e
tratamento com CO2 seguem como descrito anteriormente. Em seguida, a

111
suspensão celular foi transferida para placas de 96 poços, mantidas até atingir
confluência 80% e tratadas com as frações de A. barbadensis.
3.1.1.5. Materiais Testados
Os PT e as frações PI e PII obtidas de A. barbadensis foram
solubilizadas em dimetil sulfóxido (DMSO) para uma concentração final de 1%
(m/v) e, em seguida, solubilizadas em água destilada, sendo obtidas soluções
de concentrações de 5000 µg/mL (PT e PI) e 2000 µg/mL (PII). Em seguida, as
soluções foram filtradas em membrana Millipore (0,22 µm), repassadas para
tubos rosqueados previamente esterilizados e conservadas a –20 oC.
3.1.1.6. Toxicidade das Frações para Cultura de Células
A citotoxicidade foi determinada de acordo com análises feitas das
alterações morfológicas (DE CLERCQ et al., 1980) e da viabilidade celular
(BOREFREUND; PUERNER, 1985).
3.1.1.6.1. Verificação da Alteração da Morfologia Celular
As células foram tratadas com diluições seriadas das frações, seguindo
concentrações de 2500 a 9,7 µg/mL, para os PT e PI, e de 1000 a 3,9 µg/mL,
para PII, usando meio de cultura sem SFB como diluente. Posteriormente, o
sistema foi incubado por 48 horas, a 27 °C em ambiente com 5% de CO2.
Decorrido esse tempo, as células foram observadas em microscopia de luz
invertido e comparadas com o controle (sem amostra, apenas meio de cultura).
A maior concentração de substância observada que não alterou a morfologia
celular foi denominada de Concentração Máxima Não Tóxica (CMNT) e, em
adição, concentrações iguais e menores que a CMNT foram utilizadas para os
experimentos posteriores.

112
3.1.1.6.2. Verificação da Viabilidade Celular
O efeito dos PT e das frações PI e PII na viabilidade celular foi
determinado através da técnica denominada dye-uptake (BOREFREUND;
PUERNER, 1985), com pequenas modificações. A técnica consiste na
incorporação do corante vermelho neutro pelas células vivas e sua posterior
quantificação por leitura em espectrofotômetro, em comprimento de onda de
492 nm. A percentagem de células viáveis foi obtida pela fórmula: [(DO do
extrato)-(DO do controle de células)]/[DO do Vermelho Neutro]-(DO do controle
de células)] x 100, sendo calculada a concentração que reduz a contagem de
células viáveis em 50% (CC50).
3.1.2. ATIVIDADE INIBITÓRIA SOBRE OS ADENOVÍRUS (Ad) E OS VÍRUS
HERPES SIMPLES (HSV)
3.1.2.4. Amostras Celulares e Virais
Para esse experimento foram utilizadas as células HEp-2 e Vero (5 x 105
células/mL) para adenovírus e HSV, respectivamente. As amostras de
adenovírus tipo 19 e 41 (Ad-19 e Ad-41) foram cedidas pelo Laboratório de
Viroses Respiratórias, Entéricas e Oculares e as de herpes 1 e 2 (HSV-1 e
HSV-2) pelo Laboratório Experimental de Drogas Antivirais e Citotóxicas
(LEDAC), ambos do Departamento de Virologia do Instituto de Microbiologia
Paulo de Góes (IMPPG) da Universidade Federal do Rio de Janeiro (UFRJ).
Todos os experimentos foram realizados nos referidos laboratórios.
3.1.2.5. Titulação dos Vírus
A titulação viral foi realizada com o objetivo de se estabelecer a dose
infectante para 50% dos cultivos celulares (TCID50) de acordo com o cálculo
estabelecido por REED e MUENCH (1938).

113
3.1.2.6. Avaliação da Atividade Antiviral por Redução do Título
Viral
As culturas de células foram inoculadas com 100 TCID50 da suspensão
viral na presença das frações, obedecendo a CMNT, e na ausência (controle
de vírus). Células não tratadas e não inoculadas serviram como controle
negativo. Dessa forma, as culturas de células foram incubadas a 37 °C por 48
horas e, após incubação, os sobrenadantes dos testes e do controle foram
titulados com base no método estatístico de Reed e Muench (1938).
O grau de atividade antiviral foi expresso em índice de inibição viral (IIV)
e percentagem de inibição (I%). O IIV foi obtido pela fórmula proposta por
Lagrota (1978):
IIV = B – A
Onde: “B” é o título do vírus na cultura de células sem amostra (controle).
“A” é o título do vírus na cultura de células com amostra.
I% foi calculada de acordo com a fórmula proposta por Nishimura, Toku e
Fukuyashu (1977):
I%=[1-(antilog T/antilog C)] X 100
Onde: “T” corresponde às unidades infecciosas na cultura de células tratadas
com amostra.
“C” corresponde às unidades infecciosas na cultura de células não
tratadas com amostra (controle).
A dose efetiva capaz de inibir 50% do efeito citopático viral (ED50) foi
calculada por regressão linear dose resposta. O índice de seletividade (IS) foi
determinado pela razão de CC50/EC50.

114
3.1.3. ATIVIDADE INIBITÓRIA SOBRE O METAPNEUMOVÍRUS (HMPV) E O
VÍRUS DENGUE TIPO 1 (DENV-1)
3.1.2.5. Amostras Celulares e Virais
Foram utilizadas células LLC-MK2 e C6/36, na concentração de 5 x 105
células/mL, para experimentos com HMPV e DENV-1, respectivamente. Uma
amostra de HMPV NL/1/00 foi gentilmente cedida por ViroNovative BV,
Erasmus University Rotterdam (The Netherlands) e de DENV-1 pelo LEDAC da
UFRJ. Pelo fato da replicação viral de HMPV ser dependente de tripsina, esta
enzima foi adicionada ao meio de cultura para concentração final de 1 µg/mL
em todos os experimentos. A atividade antiviral foi realizada pelo Laboratório
Experimental de Drogas Antivirais e Citotóxicas do Departamento de Virologia
do IMPPG da UFRJ.
3.1.2.6. Atividade antiviral
Células LLC-MK2 foram tratadas com os PT e com as frações PI e PII
em concentrações escolhidas de acordo com os resultados da CMNT. Em
seguida, 100 µL de uma suspensão de HMPV diluída a 10-1 foram adicionados
às células tratadas e não tratadas com as frações. Células não-tratadas e não-
inoculadas serviram como controle negativo. Todas as células foram incubadas
a 34 °C durante 7 dias em ambiente contendo 5% de CO2. Todos os
experimentos foram feitos em triplicata. Após incubação, os sobrenadantes das
células tratadas e não tratadas foram removidos e lisados com tampão
isotiocianato de guanidina. O RNA viral foi extraído e a reação de qRT-PCR em
tempo real realizada para a detecção do genoma do HMPV. A atividade
antiviral foi avaliada comparando o número de cópias do genoma viral, obtido
dos sobrenadantes das culturas de células na presença dos PT e das frações
PI e PII, com aquele do controle de vírus, não tratado com as frações. O

115
mesmo procedimento foi realizado para células C6/36 infectadas pelo DENV-1,
com exceção da temperatura de incubação que foi de 27 °C.
3.1.2.7. Extração do RNA viral
O RNA viral foi extraído dos sobrenadantes lisados, usando um KIT
comercial Totally RNA™ (Applied Biosystems/Ambion, USA), de acordo com as
instruções do fabricante.
3.1.2.8. qRT-PCR em tempo real
A transcrição reversa foi realizada em 10 µL de uma mistura de reação
contendo 5 µL do RNA extraído e 360 nM do primer antisense, usando uma
transcriptase reversa ImProm-II (Promega Madison, USA). A PCR em tempo
real foi realizada em 24 µL de uma mistura de reação contendo 5 µL de cDNA,
900 nM de primer sense, 300 nM de primer antisense e 12 µL de Power
SYBR-Green qPCR mix (Ludwig Biotec), através de um Step-One Real Time
PCR System (Applied Biosystems, USA). As condições da PCR
compreenderam uma ativação inicial a 94 °C por 5 minutos e 45 ciclos de 10 s
a 94 °C, 10 s a 60 °C e 10 s a 72 °C, seguido por uma curva Melting. A
quantificação do RNA do HMPV foi realizada usando uma curva padrão gerada
pelos valores Ct (Threshold cycle) obtidos de diluições logarítmicas dos
transcritos in vitro, contendo diluições variando de 100 a 107 do estoque original
de vírus. Cada diluição foi quantificada usando o Quant-IT™ RNA Assay Kit
(Invitrogen), que nos permitiu correlacionar à quantidade de RNA, isto é, o
número de cópias do genoma com o Ct de cada diluição. Os resultados foram
analisados em triplicata e o número médio de cópias do RNA viral foi calculado.

116
3.1.4. HEMOSTASIA
3.1.4.1. Ensaios de Coagulação in vitro
3.1.4.1. Tempo de tromboplastina parcial ativada (TTPa)
O teste do TTPa foi realizado segundo especificações do fabricante
(CLOT, Bios diagnóstica, Sorocaba, SP). Para isso 50 µL de plasma humano
foram incubados a 37°C por 3 minutos com 10 µL da solução das frações PI ou
PII (concentrações de 0,1; 0,25; 0,50; 0,75; 1 mg/mL) e 50 µL do reagente
TTPa. Após a incubação, foram adicionados ao sistema 50 µL de cloreto de
cálcio 25 mM para ativar a cascata de coagulação. Os ensaios foram
realizados em duplicata, sendo o tempo de coagulação registrado
automaticamente em coagulômetro (DRAKE, modelo QUICK-TIMER).
A atividade anticoagulante foi expressa em termos de unidades
internacionais (UI) por mg de polissacarídeo utilizando curva-padrão de
heparina não-fracionada (193 UI mg-1), oriunda do Instituto Nacional de Padrão
Biológico e Controle (Potters Bar, Herts, UK).
3.1.4.2. Tempo de Protrombina (TP)
O teste do TP foi realizado segundo especificações do fabricante (CLOT,
Bios disgnóstica, Sorocaba, SP). Para isso, 50 µL de plasma humano foram
misturados com 10 µL da solução das frações PI ou PII (concentrações de 0,1;
0,25; 0,50; 0,75; 1 mg/mL) e incubados a 37 °C durante 3 minutos. Em seguida,
50 µL do reagente TP foram adicionados à mistura para ativar a cascata de
coagulação. Os ensaios foram realizados em duplicata, sendo o tempo de

117
coagulação registrado, automaticamente, em um coagulômetro (DRAKE,
modelo QUICK-TIMER).
3.1.4.2. Ensaios de Coagulação ex vivo
3.1.4.2.1. Animais
Foram utilizados para os experimentos de hemostasia camundongos
Swiss macho e fêmea (20-25 g) e ratos machos Wistar (250-280 g), fornecidos
pelo Biotério Central da Universidade Federal do Ceará – Campus do Pici. Os
animais receberam água e alimentação ad libitum. Todos os esforços foram
feitos para minimizar o número e o sofrimento dos animais utilizados.
Os ensaios com os animais seguiram os padrões exigidos de ética e
biossegurança, acreditando na sua aprovação justa, devido a sua relevância e
respeito aos princípios dos 3 R’s da experimentação animal (“reduction,
replacemaent and refinemaent”), recebendo aprovação da comissão de ética
de pesquisa com animais de laboratório (CEPA n°33/09).
3.1.4.2.2. Avaliação do Efeito Hemorrágico
Ratos Wistar foram anestesiados com injeção intramuscular de cetamina
5% (König, Avellaneda, Argentina) na dose de 100 mg/kg e 16 mg/kg de
xilasina 2% (König, Avellaneda, Argentina). Uma cânula foi inserida na artéria
carótida direita para a administração intravascular das frações PI ou PII (doses
0,5; 1,0; 1,5 mg/Kg). Animais tratados com salina serviram como controle. Após
5 minutos de circulação da amostra, o sangue foi coletado em tubos citratados,
para análise de TTPa ex vivo, e um segmento de 3 mm da cauda do rato foi
cortado (Figura 3.1). Logo em seguida, a cauda foi inserida cuidadosamente

118
numa proveta, contendo 40 mL de água destilada. A perda sanguínea foi
determinada após 60 minutos, mensurando o nível de hemoglobina dissolvida
na água pelo método espectrofotométrico descrito por Herbet, Bernat e
Maffrand (1992). O volume de sangue perdido foi determinado através de uma
curva padrão baseada na A540nm.
Figura 3.1: Representação esquemática do modelo de análise do efeito
hemorrágico por tempo de sangramento. Nesse modelo, o volume de sangue
perdido é calculado a partir da concentração de hemoglobina dissolvida na
água destilada contida na proveta. FONTE: Fonseca, 2009.

119
3.1.4.2.3. Tempo de tromboplastina parcial ativada (TTPa) ex vivo
Sangue de rato citratado foi coletado 5 minutos após a administração
das frações PI e PII, nas doses descritas no item seguinte, através de uma
cânula posicionada dentro da artéria carótida. Posteriormente, o sangue foi
centrifugado, para obtenção do plasma, e submetido ao ensaio TTPa, como
descrito no item 3.1.4.1.
3.1.4.3. Avaliação Sistêmica dos PT de A. barbadensis na toxicidade
subcrônica em camundongos.
Para avaliação preliminar das propriedades tóxicas, os PT foram
administrados (10,0 mg kg-1; i.p.) em camundongos Swiss machos e fêmeas,
divididos em grupos de seis animais, durante quatorze dias consecutivos.
Durante a fase experimental, foram diariamente pesados, sendo mantidos no
biotério do Departamento de Bioquímica da UFC com livre acesso à água e
ração. Durante este período, foram observados sinais físicos como coçar
focinho, lamber as patas, rodopiar, ereção de pelos e, em adição, sinais
comportamentais como apatia ou agressividade.
No 15° dia, os animais foram anestesiados e, a partir do plexo orbital,
amostras de sangue foram coletadas em tubos citratados para, imediatamente,
realizar análise dos parâmetros hematológicos. Em adição, amostras de
sangue foram centrifugadas e o plasma obtido foi mantido a -20°C para
posteriores análises bioquímicas.
Em seguida, os camundongos foram submetidos à eutanásia por
deslocamento cervical seguido da retirada e pesagem dos órgãos fígado, rim,
coração, baço, timo e linfonodos. As massas dos órgãos foram correlacionadas
com as respectivas massas corpóreas. Todos os parâmetros toxicológicos
observados foram comparados ao grupo controle, animais normais tratados
com solução salina (NaCl 0,15 M), seguindo as mesmas condições.

120
3.1.4.4. Parâmetros Hematológicos e Bioquímicos
O objetivo desse estudo foi avaliar parâmetros hematológicos [número de
leucócitos (WBC), número de eritrócitos (RBC), concentração de hemoglobina
(HGB), hematócrito (HCT), volume corpuscular médio (MVC), hemoglobina
corpuscular média (MCH), concentração média de hemoglobina corpuscular
(MCHC), número de plaquetas (PLT), razão percentual de linfócitos (W-SCR),
razão percentual de monócitos (W-MCR), razão percentual de neutrófilos (W-
LCR), número de linfócitos (W-SCC), número de monócitos (W-MCC), número
de neutrófilos (W-LCC), amplitude da distribuição dos eritrócitos (RDW-SD),
coeficiente de variação ao redor da média do volume das células vermelhas
(RDW-CV), índice de amplitude de distribuição do tamanho das plaquetas
(PDW), volume plaquetário médio (MPV) e percentagem de plaquetas gigantes
(P-LCR)]. Esses parâmetros foram analisados com intuito de investigar se
houve alterações nos perfis hematológicos quando camundongos foram
administrados com polissacarídeos de A. barbadensis.
Além disso, foi realizada análise dos parâmetros bioquímicos a partir das
dosagens séricas de uréia, creatinina, fosfatase alcalina e das transaminases
glutâmico oxalacética e pirúvica (TGO e TGP). A metodologia foi seguida de
acordo com o fabricante (Labtest®).
3.1.4.5. Análises Histológicas
Os órgãos foram examinados macroscopicamente e, em seguida, foram
pesados e fixados em formol 10%. Após 24 horas, os órgãos foram submetidos
a sucessivas trocas com álcool etílico (70% até absoluto), colocados em
lâminas de vidro e corados com hematoxilina e eosina. As lâminas foram
examinadas por meio de microscopia de luz (aumento 400x).

121
3.1.4.6. Análises Estatísticas
Os dados obtidos foram expressos como média ± E.P.M. (erro padrão da
média). Para a verificação das diferenças estatísticas entre os grupos de
animais foi utilizado a Análise de Variância (ANOVA) e, quando observada
diferença entre as médias, foi aplicado o teste de Bonferroni. p<0,05 foi
considerado como significante.

122
3.2. RESULTADOS E DISCUSSÕES
3.2.1. Citotoxicidade e Viabilidade Celular
A citotoxicidade é definida como um conjunto de alterações da
homeostase celular, que provoca uma série de modificações e interferem na
capacidade adaptativa da célula, bem como na sua sobrevivência,
multiplicação e realização de suas funções metabólicas (NARDONE, 2006).
Essas modificações podem ser evidenciadas pela desorganização da
monocamada celular, acompanhada de aspecto granuloso e arredondado das
células (STREISSLE; SCHWOBEL; HEWLETT, 2006). Embora a maioria dos
polissacarídeos oriundos de planta apresente baixa toxicidade em células de
mamíferos (TALYSHINSKY; SOUPRUN; HULEIHEL, 2002), é indispensável à
avaliação prévia de sua possível citotoxicidade.
Células LLC-MK2, HEp-2, HeLa, Vero e C6/36 foram submetidas a
diferentes concentrações dos PT e das frações PI e PII de A. barbadensis
(Figura 3.2). Com exceção da linhagem LLC-MK2, todas as células
apresentaram viabilidade celular de 100% nas maiores concentrações
utilizadas (Tabela 3.1). Dessa forma, é sugerido que a concentração que reduz
a contagem de células viáveis em 50% (CC50) das amostras dos PT e PI seja
>2500 µg/mL [ppm (partes por milhão)]. Em adição, a CC50 de PII foi >1000
ppm, que foi a maior concentração obtida durante o processo de diluição, para
esse experimento. Além disso, a CMNT dessas amostras, para as células
citadas, também está acima das concentrações testadas, visto que não foi
observado nenhum efeito tóxico nessas concentrações. Em contraste, as
frações apresentaram citotoxicidade para as células LLC-MK2 nas
concentrações maiores. Com isso, foram obtidos valores da CC50 para os PT e
para as frações PI e PII, correspondendo a 203, 200 e 182 ppm,
respectivamente (Tabela 3.1).

123
Figura 3.2: Fotomicrografia das células de: A – rim de macaco rhesus [Macaca
Mulatta (LLC-MK2)]; B – carcinoma de laringe humana (HEP-2); C – carcinoma
cervical uterino HeLa; D – rim de macaco verde africano [Cercopithecus
aethiops (Vero)]; E – larva de mosquito Aedes albopictus (C6/36). (Aumento
100x)
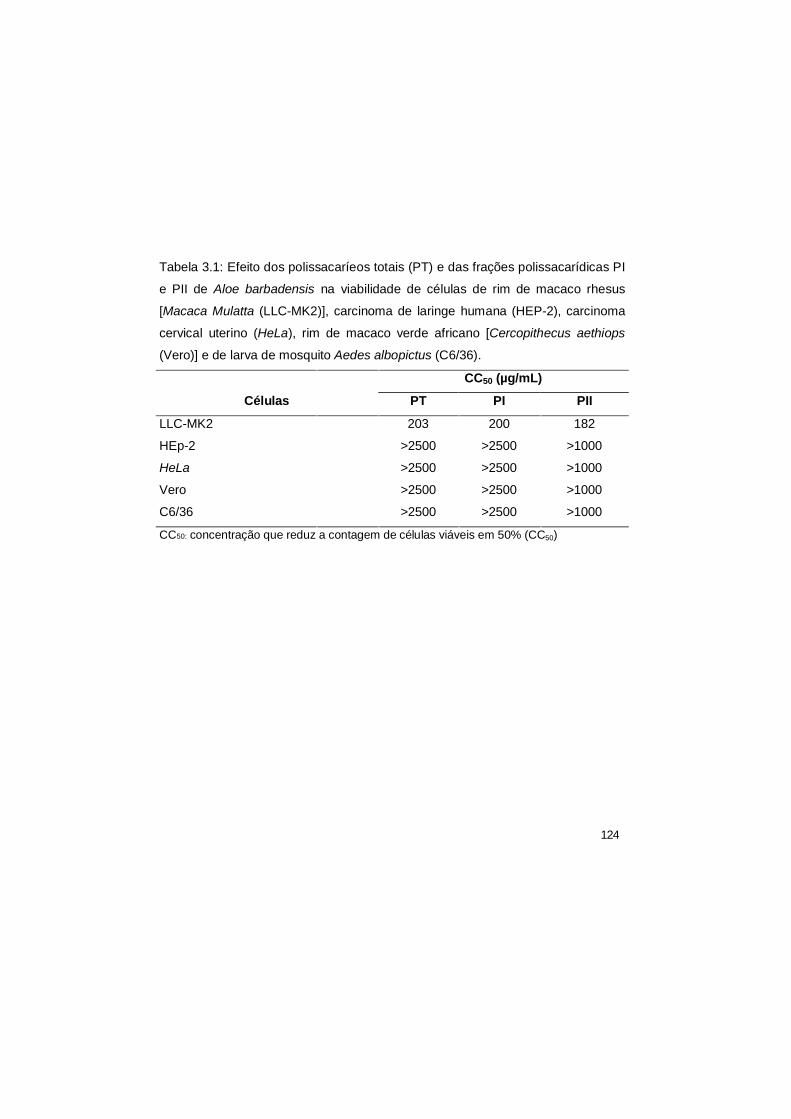
124
Tabela 3.1: Efeito dos polissacaríeos totais (PT) e das frações polissacarídicas PI
e PII de Aloe barbadensis na viabilidade de células de rim de macaco rhesus
[Macaca Mulatta (LLC-MK2)], carcinoma de laringe humana (HEP-2), carcinoma
cervical uterino (HeLa), rim de macaco verde africano [Cercopithecus aethiops
(Vero)] e de larva de mosquito Aedes albopictus (C6/36).
CC50 (µg/mL)
Células PT PI PII
LLC-MK2 203 200 182
HEp-2 >2500 >2500 >1000
HeLa >2500 >2500 >1000
Vero >2500 >2500 >1000
C6/36 >2500 >2500 >1000
CC50: concentração que reduz a contagem de células viáveis em 50% (CC50)

125
A análise de citotoxicidade em extratos foliares de espécies do gênero
Aloe tem sido, em sua maioria, relacionada ao extrato bruto da polpa ou dos
compostos fenólicos do exsudato amarelado da casca (IASC, 2012). Brasher et
al. (1969) misturaram ao gel de A. barbadensis a indometacina e predinisolona
e, em seguida, analisaram a citotoxicidade da mistura em células HeLa e
fibroblastos de rim de coelho. Foi verificado que, concentrações com diluições
a partir de 10-1 não foram tóxicas às células. Winters, Benavides e Clouse
(1981) concluíram que o gel de A. barbadensis foi tóxico em células normais e
tumorais. Testes posteriores mostraram que essa citotoxicidade se deve,
geralmente, aos compostos do exsudato da casca (DANOF; McANALLEY,
1983). É sabido, também, que Aloctin A, lectina mais estudada do gênero Aloe,
inibe o crescimento de fibrosarcoma in vitro e células do baço in vivo (LEE,
2006).
Adicionalmente, foi relatado que acemanana, polissacarídeo presente
nos extratos de A. barbadensis, apresentou citotoxicidade significativa em
macrófagos ativados de cultura de células da medula óssea de galinha
(KARACA; SHARMA; NORDGREN, 1995). Esses resultados entram em
contradição com os descritos por LEE (2006), no qual os polissacarídeos não
apresentaram citotoxicidade, apenas a fração proteica e fenólica. Esses
resultados corroboram com os encontrados por Brasher et al. (1969), onde o
Aloe Vera Gel (nome comercial dado ao extrato da polpa de A. barbadensis),
oriundo de diferentes empresas, apresentaram diferentes resultados
relacionados à citotoxicidade, sendo que alguns apresentaram toxicidade
extremamente baixa ou inexistente.
No presente estudo, a contaminação fenólica foi, em sua grande maioria,
excluída durante o processo de tratamento exaustivo com etanol, acetona e
éter (item 2.1.2). De acordo com Campestrini (2007), isso pode ser
acompanhado por uma diminuição drástica do sinal 775,0 ondas.cm-1 da
absorção do infravermelho, indicativo de compostos fenólicos (Figura 2.13). Em
adição, a insignificante contaminação por componentes não glicídicos
presentes nos PT e nas frações PI e PII sugerem que a citotoxicidade em
células LLC-MK2 esteja realmente relacionada aos seus polissacarídeos.

126
3.2.2. Atividade Inibitória Sobre os Vírus Adenovírus (Ad) e os Vírus
Herpes Simples (HSV)
As frações obtidas de A. barbadensis foram testadas nas CMNTs em
microplacas de 96 poços, com adição do polissacarídeo antes da adsorção dos
vírus Ad 19 e 41. Não houve diferença significativa entre efeito citopático do
controle positivo e células tratadas e inoculadas. Com base nos controles, foi
concluído que as frações estudadas não apresentam atividade antiviral direta
para os vírus não envelopados testados.
Alves et al. (2004) já haviam relatado que as antraquinonas de A.
barbadensis, aloe-emodina e aloe-barbaloína, não apresentam atividade contra
vírus não envelopados, como os adenovírus e rinovírus. No entanto, alguns
artigos relatam a participação dos polissacarídeos de A. barbadensis na
imunomodulação contra vírus não envelopados. Acemanana, por exemplo,
diminuiu a imunossupressão causada pelos poliomavírus e reovírus (RITCHIE,
et al., 1994; SHARMA; KARACA; PERTILE, 1994) e uma polimanose
aumentou as concentrações de anticorpos anti-coxsackivírus (GAUNTT et al.,
2000). Adicionalmente, em experimentos em cultura de células, foi observado
que extratos brutos de A. barbadensis apresentaram atividade contra vírus
envelopados como Newcastle (CHINNAN et al., 1992), citomegalovírus (SAOO
et al., 1996), herpes (ZANDI et al., 2007) e influenza A (SYDISKIS et al., 1991).
Com base nessas informações, foi realizada análise da atividade
antiviral dos PT e das frações PI e PII em 4 vírus envelopados (HSV-1, HSV-2,
DENV-1 e HMPV). Para os HSV-1 e HSV-2, vírus envelopados de DNA, foram
utilizadas concentrações de 250 ppm para as devidas frações, já que não
foram observadas alterações significativas quando comparadas com a CMNT
(2500 ppm). Após o período de incubação de 48 h, foi possível observar a
inibição do efeito citopático (ECP) (Figura 3.3). Esse efeito é caracterizado pelo
aparecimento de células com dimensões variáveis, arredondadas, bastante
brilhantes (Figura 3.3 A), mais refringentes, algumas vezes separadas e, mais
frequentemente, ligadas umas às outras por prolongamentos citoplasmáticos
(Figura 3.3 C). Regiões com essa característica recebe o nome de “foco da

127
infecção” (Figura 3.3 B), que se estendem rapidamente pela superfície do local
de infecção. As lesões citoplasmáticas aparecem como formações
policarióticas (sincícios e células gigantes) devido à fusão dos citoplasmas das
células adjacentes infectadas (Figura 3.3 D). O ECP observado foi semelhante
aos encontrados por MÜLLER, 2006.
As percentagens de inibição dos PT e das frações PI e PII podem ser
observadas na Tabela 3.2. Esses resultados mostram que os PT apresentaram
I% de 85,9%, para HSV-1, e 94,4%, para HSV-2. As maiores I% obtidas foram
dos PT, o que indica que PI e PII agem de maneira sinergística,
potencializando o efeito antiviral. Isso pode ser observado, principalmente, em
PII, que não apresentou efeito para HSV-1, mas potencializou a atividade dos
PT. Em contraste, PII inibiu a infecção causada por HSV-2 em 85,9%.
Alguns autores já haviam descrito a atuação de polissacarídeos
sulfatados no bloqueio da etapa de adsorção viral (GEMIN, 2008; MAZUNDER
et al., 2002). Isso é justificado pelo fato da infecção causada pelos HSV
requerer interação entre a glicoproteína viral gC e moléculas de heparan sulfato
celular. Dessa forma, os polissacarídeos sulfatados agem, competitivamente,
com sítio de ligação de gC, impedindo o processo de adsorção viral. Damonte
(2004) já havia relatado que a ação antiviral de polissacarídeos sulfatados não
depende só de sua carga líquida negativa, mas, também, da disposição dos
grupos sulfato na sua estrutura.

128
Figura 3.3: Efeito citopático em células de rim de macaco verde africano
Cercopithecus aethiops (Vero) infectadas com HSV-1. (a) Células
arredondadas desprendidas da monocamada (seta); (b) Foco de infecção em
monocamada de células (círculo); (c) Célula arredondada com tamanho
superior à células normais (seta). Pode ser observado prolongamentos
citoplasmáticos; (d) Células sinciciais gigantes (asterisco).

129
Tabela 3.2: Percentagem de inibição dos polissacarídeos totais (PT) e das
frações PI e PII de Aloe barbadensis contra os vírus HSV-1 e HSV-2 em células
de rim de macaco verde africano Cercopithecus aethiops (Vero).
Substância
CC50
(µg/mL)
Concentraçãoempregada
(µg/mL)
HSV-1
IIV I%
HSV-2
IIV I%
PT > 2500 250 0,9 85,9 1,3 94,4
PI > 2500 250 0,5 68,4 0,9 85,9
PII > 1000 250 0,0 0,0 0,9 85,9
CC50 = concentração citotóxica para 50% das células em cultura; HSV-1 =
Herpes simples vírus tipo 1; HSV-2 = Herpes simples vírus tipo 2; IIV = Índice
de Inibição Viral; I% = Percentagem de Inibição. Os resultados foram obtidos a
partir de experimentos desenvolvidos em triplicatas.

130
Embora seja relatado que muitos polissacarídeos sulfatados apresentem
atividade antiviral contra os HSV (BANDYOPADHYAY et al., 2011;
CASSOLATO et al., 2008; COSTA et al., 2010; GHOSH et al., 2009; HAYASHI
et al., 2008; MANDAL et al., 2008), isso não explica a ação exercida pelos
polissacarídeos neutros presentes em PI. Alguns polissacarídeos neutros,
porém sulfatados por modificações químicas, têm mostrado relevantes
atividades anti-herpéticas (LIU et al, 2011; MANDAL et al., 2010). Dessa forma,
outros mecanismos de interação/inibição devem estar envolvidos.
É sabido que a proteína gD, presentes no envelope viral dos HSV, é
importante no processo de adsorção e fusão viral. (BARBOSA, 2009). Essa
proteína pode interagir com quatro tipos distintos de receptores celulares: o
mediador de entrada do herpesvirus (HVEM – mediador de entrada de HSV),
um membro da superfamília dos receptores TNF/NGF (fator de necrose
tumoral/fator de crescimento neural); os receptores da família da Nectina 1 e 2,
membros da superfamília das imunoglobulinas; e moléculas de heparan sulfato
modificadas pela ação enzimática da 3-O-sulfotransferase (SPEAR et al., 2006;
TIWARI et al., 2005).
Já é descrito na literatura que polissacarídeos neutros de A. barbadensis
apresentam atividades imunomoduladoras. Em estudos com macrófagos, foi
observado que aloerídeo e acemanana induziram a expressão de mRNA que
codificam para IL(interleucina)-1, TNF-á e interferon- γ (IFN-γ), que são
potentes agentes na ativação de monócitos/macrófagos. Essa atividade está
associada aos receptores de manose presentes na membrana celular dos
macrófagos (LIU et al., 2006; TIZARD; RAMAMOORTHY, 2004) e, esses
receptores, são classificados como lectinas, proteínas ou glicoproteínas com
capacidade se ligar a carboidratos (VAN DAMME et al., 1998).
Em estudos realizados com células Vero, células comumente utilizadas
para estudo de infecções causadas pelo HSV, Rapoport et al. (2006)
observaram que as mesmas apresentam receptores lectínicos capazes de
reconhecer resíduos de galactose, manose, oligossacarídeos sulfatados e
ácido siálico. Segundo Šedý, Spear e Ware (2008), qualquer reconhecimento
celular mediado por um desses receptores podem alterar, significantemente, as

131
vias de sinalização da célula. Uma dessas alterações é a modificação do
padrão qualitativo e quantitativo de receptores de TNF/NGF e nectinas na
membrana plasmática celular (MANNI; ALOE, 2009; ŠEDÝ, SPEAR; WARE,
2008; SHUKLA; SINGH; SHUKLA, 2009).
Dessa forma, é sugerido que polissacarídeos neutros de PI estejam
interagindo com receptores manosil-ligantes da membrana plasmática de
células Vero, desencadeando mudança no perfil de receptores membranares.
Consequentemente, o processo de adesão e fusão viral, são importantes para
que a infecção pelos HSV, seja inibida.
Com base nos valores de CC50 e ED50 foi calculado o valor do índice de
seletividade (IS), que representa o grau de segurança para a utilização de um
composto testado. Esse parâmetro farmacológico foi calculado através da
razão entre CC50 e ED50 das frações. Ou seja, quanto maior o valor de IS, mais
promissora é a substância, indicando a viabilidade de realização de estudos
posteriores mais detalhados, tanto in vitro quanto in vivo (MÜLLER, 2006). Foi
observado que os maiores IS foram obtidos dos PT e da fração PI para o HSV-
1 e o HSV-2, respectivamente (Tabela 3.3). Esses valores foram equivalentes a
zalcitabina (IS=110), dideoxinucleotídeo (ddC) bastante utilizado como antiviral
comercial (BARBOSA, 2009).

132
Tabela 3.3: Atividade inibitória dos polissacarídeos totais (PT) e das frações PI
e PII de Aloe barbadensis sobre os vírus HSV-1 e HSV-2 em células de rim de
macaco verde africano Cercopithecus aethiops (Vero).
Substâncias CC50
(µg/mL)
HSV-1
ED50
(µg/mL)
HSV-2
ED50
(µg/mL)
IS
HSV-1
IS
HSV-2
PT > 2500 18,6 191,2 > 134,4 > 13,0
PI > 2500 58,0 25,1 > 43,1 > 99,6
PII > 1000 0,0 50,4 0,0 > 19,8
CC50 = concentração citotóxica para 50% das células em cultura; HSV-1 = vírus
Herpes simples tipo 1; HSV-2 = vírus Herpes simples tipo 2; ED50 = Dose
efetiva capaz de inibir 50% da propagação viral; IS = Índice de seletividade. Os
resultados foram obtidos a partir de experimentos desenvolvidos em triplicatas.

133
1.2.1. Atividade Inibitória Sobre o HMPV e o DENV-1
Como mostrado na Tabela 3.4, a fração contendo polissacarídeos
sulfatados (PII) apresentou atividade sobre o HMPV, próxima a 100%, a partir
da concentração de 3,9 µg/mL. Esses resultados podem ser acompanhados
pela amplificação quase inexistente do RNA viral, através de análises feitas por
qRT-PCR. Em adição, foram obtidos valores para CC50 de 182 µg/mL e ED50
0,48 µg/mL, o que implica em IS alto, chegando a valores de 379 (Tabela 3.5).
Embora nessas concentrações sejam observados efeitos citopáticos (Figura
3.4), o IS obtido é sugestivo para afirmarmos a segurança do uso de PII como
agente antiviral, na concentração de 3,9 µg/mL, para as células LLC-MK2.
Em adição, os PT apresentaram atividade contra HMPV, chegando a
inibir 85,77% da infecção viral (Tabela 3.4). Em relação a PI, foi observada
amplificação de RNA viral similar ao grupo controle, sugerindo, assim, que não
houve inibição do vírus. Dessa forma, os resultados sugerem que os grupos
carregados negativamente de PII estejam envolvidos na inibição da infecção
causada por HMPV.
A adesão e penetração são passos essenciais que os vírus utilizam para
obter sucesso no processo de infecção. Geralmente, a adesão é facilitada pela
interação eletrostática de moléculas negativamente carregadas da célula e
moléculas positivamente carregadas do envelope viral. Liu e Thorp (2002) já
haviam descrito o efeito antiviral que muitos polissacarídeos similares aos
glicosaminoglicanos possuem contra vírus envelopados (FEUILLET, 2012).
Wyde et al. (2004; 2005) relataram que NMSO3, lipídeo sialil sulfatado,
apresentou atividade contra HMPV e RSV, obtendo valores de ED50 de 6
µg/mL, para ambas amostras. Nesse mesmo trabalho, análises comparativas
com ribavirina, antiviral comercial muito utilizado para infecções causadas por
HMPV, mostraram IS de 84, bem menor do que PII, utilizada no presente
trabalho. Compostos sulfatados também são capazes de inibir outras espécies
de vírus da mesma família, Paramixoviridae, como RSV (LUNDIN et al., 2012)
e parainfluenza vírus (DE CLERCQ, 2004).

134
Tabela 3.4: Efeito inibitório dos polissacarídeos totais (PT) e das
frações PI e PII de Aloe barbadensis sobre o HMPV em células de rim
de macaco rhesus [Macaca Mulatta (LLC-MK2)].
Substância Concentração(µg/mL)
Inibição (%)
PT 7,8 85,77
3,9 79,51
1,9 66,52
0,9 17,83
PI 200,0 -
100,0 -
50,0 -
25,0 -
PII 3,9 96,60
1,9 87,64
0,9 65,33
0,4 44,91
(-) Não houve inibição

135
Tabela 3.5: Atividade inibitória dos polissacarídeos totais (PT) e
das frações PI e PII de Aloe barbadensis sobre o HMPV em
células de rim de macaco rhesus [Macaca Mulatta (LLC-MK2)].
Substância CC50
µg/mL
ED50
µg/mL
IS
PT 203 1,5 135
PI 200 - -
PII 182 0,48 379
CC50 = concentração citotóxica para 50% das células em
cultura; ED50 = Dose efetiva capaz de inibir 50% da propagação
viral; IS = Índice de seletividade; (-) Não houve inibição.

136
Figura 3.4: Efeito citopático de HMPV em células de rim de macaco rhesus
[Macaca Mulatta (LLC-MK2)]. Flechas indicam aglomeração celular.

137
Em suma, PII, por apresentar unidades monossacarídicas carregadas
negativamente, podem mimetizar os efeitos promovidos por GAGs, como
descrito na literatura. Os resíduos aniônicos, presentes na cadeia
polissacarídica de PII, podem estar se ligando a proteínas virais de adesão,
como a Proteína G, através de interações eletrostáticas múltiplas e,
consequentemente, inibindo a adesão do vírus na célula.
Embora essa mesma justificativa seja, também, utilizada para
polissacarídeos sulfatados que apresentam atividade antiviral contra sorotipos
do vírus da dengue (HIDARI et al., 2008; KATO et al., 2010; TALARICO et al.,
2005; TISCHER et al., 2006), as frações estudadas no presente trabalho não
foram capazes de inibir DENV-1. Alguns GAGs com alto grau de sulfatação tais
como heparina e condroitim sulfato, apresentam potente inibição da infecção
causada por flavivírus (SU et al., 2001). O grau de sulfatação tem sido
relacionado a esses efeitos inibitórios, já que podem interagir competitivamente
com a proteína viral E presente no envelope viral, capaz de reconhecer
resíduos de heparan sulfato presentes na membrana celular hospedeira
(DUARTE et al., 2004). No entanto, Kato et al. (2010) relataram que condroitim
sulfato E apresentou atividade inibitória contra os quatro sorotipos do vírus da
dengue, enquanto que condroitim sulfato D, com mesmo grau de sulfatação,
não inibiu a infecção. Esses resultados corroboram com os obtidos por
Damonte (2004), onde a disposição dos grupos sulfato na estrutura do
polissacarídeo é tão importante quanto sua carga líquida, para inibição viral.
Em adição, é sabido que a atividade dos polissacarídeos sulfatados contra o
vírus da dengue é dependente do sorotipo e da célula hospedeira (TALARICO
et al., 2005).
Dessa forma, outros estudos deverão ser desenvolvidos para elucidar
melhor a relação da atividade antiviral com a disposição estrutural dos grupos
sulfato e grau de sulfatação de polissacarídeos.

138
1.2.2. Hemostasia
3.2.4.1. Atividades Prócoagulante e Anti-hemorrágica das Frações
PI e PII
As atividades de coagulação TTPa e TP in vitro foram realizadas com
objetivo de averiguar se as frações PI e PII apresentam atividades anti- ou
prócoagulante. Foi observado que as frações PI e PII, nas concentrações
testadas, não foram capazes de alterar valores do TTPa. No entanto, a fração
PI, a partir de 0,5 mg/mL, diminui o TP de 40 para 22 segundos, permanecendo
constante nas concentrações 0,75 e 1 mg/mL. Nenhuma alteração nos valores
de TP foi observada quando aplicada a fração PII nas diferentes concentrações
testadas.
Para avaliar um possível efeito anti-hemorrágico das frações PI e PII, em
diferentes concentrações, foi utilizado um modelo experimental em ratos para
determinar o tempo de sangramento. A perda sanguínea é avaliada 60 minutos
após a aplicação das frações PI ou PII, mensurando, assim, o nível de
hemoglobina dissolvida na água, através da leitura da absorbância por
espectrofotometria. De acordo com os resultados obtidos (Figura 3.5 A), a
fração PI reduziu a perda sanguínea em 55,5%, quando comparado com os
animais que foram tratados apenas com salina. Em contraste, a aplicação
intravenosa da fração PII em ratos não alterou a mesma. Além disso, os
resultados obtidos de TTPa nos ensaios in vivo corroboram com os obtidos nos
ensaios in vitro, não havendo, assim, alteração significativa (Figura 3.5 B).
De acordo com os resultados obtidos, é sugerido que os polissacarídeos
presentes na fração PI estejam relacionados na ativação do processo inicial da
coagulação, visto que diminuiu TP. Em contraste, estes polissacarídeos não
estão envolvidos no processo de amplificação, já que não alteraram os valores
de TTPa, tanto in vitro quanto in vivo.

139
Figura 3.5: Avaliação do efeito anti-hemorrágico (A) e do tempo de
tromboplastina parcial ativada (TTPa) (B) em ensaios in vivo em ratos, após
tratamento com as frações PI e PII. * p<0,05 indica diferença significativa de PI
(1,5 mg/Kg) comparada ao grupo controle.

140
Liu et al. (2006) obtiveram resultados semelhantes aos do presente
trabalho, quando testaram alguns polissacarídeos sulfatados, de diferentes
fontes comerciais, em testes de coagulação e de perda sanguínea. Foi
observado que esses polissacarídeos não alteraram o TTPa, mas foram
potentes em diminuir TP e a perda sanguínea. Nesse trabalho, os testes de
coagulação foram realizados utilizando plasma sanguíneo proveniente de
humanos hemofílicos e normais, chegando à conclusão que os polissacarídeos
sulfatados testados poderiam estar inibindo o TFPI (inibidor da via do fator
tecidual), implicando, assim, na produção aumentada de fator tecidual (TF).
É sabido que TF é uma glicoproteína de membrana expressa,
constitutivamente, por fibroblastos subjacentes ao endotélio vascular. Células
endoteliais e monócitos, que, normalmente, não expressam o fator tecidual,
podem expressá-lo na vigência de lesão endotelial e na presença de TNF-á e
IL-1. Já foi descrito que polissacarídeos de A. barbadensis estão envolvidos na
ativação de monócitos/macrófagos e na liberação de TNF-á e IL-1 (LIU et al.,
2006; TIZARD; RAMAMOORTHY, 2004). Além disso, a ação que esses
polissacarídeos têm em ativar fibroblastos (CHUNG; CHOI, 2006), células que
contêm TP constitutivamente, pode estar envolvida no processo prócoagulante
e anti-hemorrágico.

141
3.2.4.2. Avaliação Sistêmica dos PT de A. barbadensis na toxicidade
subcrônica em camundongos.
Durante o período experimental das análises de toxicidade, não foi
verificada mortalidade ou qualquer alteração física ou comportamental dos
animais tratados com os PT. Contudo, houve diferença significativa (p<0,05) da
massa do baço (Tabela 3.6) dos camundongos e aumento do número de
plaquetas em, aproximadamente, 36 % para os machos e 50 % para as fêmeas
(Tabela 3.7). Além disso, não houve alterações significativas em relação aos
demais parâmetros bioquímicos e hematológicos, indicando que os PT não
foram tóxicos na concentração de 10 mg/Kg para os animais testados. Esses
dados corroboram, também, com os encontrados nas análises histopatológicas
realizadas (Figura 3.6), nas quais não apresentaram alterações irreversíveis.
Fogleman et at. (1992) já haviam relatado a baixa toxicidade de
acemanana quando administrada em camundongos, ratos e cachorros, na
concentração de 20 mg/Kg. Diferente do presente estudo, os autores relataram
que não houve nenhuma alteração nos órgãos, inclusive no baço. O aumento
do baço, no presente estudo, pode estar relacionado com o aumento da
hematopoese, implicando em uma maior retenção de eritrócitos na polpa
vermelha e de leucócitos na polpa branca do devido órgão (MURPHY;
TRAVERS; WALPORT, 2010). O aumento do número de linfócitos no baço é
indicativo de imunoestimulação (RODRIGUES, 2011). Outro relato de análises
de toxicidade de extratos de espécies do gênero Aloe, foi realizado por
Matsuda et al. (2007). Nesse trabalho ratos foram tratados com o gel extraído
de A. arborescens, durante um período de 1 ano, não aumentando o baço
aumentado, mas apresentando aumento no número de leucócitos (WBC).
Apesar de, no presente trabalho, ter sido observado aumento
plaquetário, esses valores estão distantes de indicar trombocitose. Schimitt et
al. (2001) compararam parâmetros plaquetários de seres humanos e
camundongos e mostraram que concentrações até 1500 x 10-3uL são
aceitáveis.

142
Tabela 3.6: Análise de toxicidade por dose repetida dos PT de Aloebarbadensis em camundongos machos e fêmeas Swiss.
Parâmetros Tratamento (10 mg/Kg; i.p.)Machos Fêmeas
Salina PT Salina PTMassa corporal (g)iniciala 25,39±0,89 25,38±0,41 24,55±0,69 26,17±0,85
Massa corporal (g)finala 29,09±1,51 31,34±0,73 29,70±0,71 30,57±0,75
Fígado (%)b
6,02±0,34 5,25±0,17 5,24±0,08 5,44±0,07
Rim (%)b
0,91±0,04 0,91±0,02 0,78±0,02 0,80±0,04
Coração (%)b0,60±0,02 0,58±0,01 0,66±0,03 0,55±0,03
Baço(%)b0,42±0,03 0,63±0,04* 0,48±0,03 0,67±0,02*
Timo (%)b0,30±0,02 0,37±0,01 0,41±0,02 0,46±0,02
Linfonodo (%)b
0,13±0,01 0,16±0,02 0,24±0,01 0,30±0,02
TGO (U/L)c
31,82±8,38 15,92±2,10 49,14±12,62 39,96±3,36
TGP (U/L)c7,16±0,71 8,95±1,15 5,80±1,10 7,13±0,97
Uréia(mg/dL)c36,31±1,00 36,25±1,55 29,37±2,78 27,22±1,40
Creatinina (mg/dL)c
0,26±0,04 0,20±0,05 2,65±0,83 2,41±0,35
Fosfatase alcalina(U/L)c 68,16±10,04 67,18±5,89 75,16±12,64 95,14±16,96
a- massa corporal
b- massa dos órgãos
c- dosagens bioquímicas
* Diferença significativa (P<0,05)

143
Tabela 3.7: Análises hematológicas de sangue de camundongos Swiss machos
e fêmeas tratados com os PT de Aloe barbadensis.
Parâmetros Tratamento (10 mg/Kg; i.p.)
Machos Fêmeas
Salina PT Salina PT
WBC (x103/uL) 3,81±0,54 3,10±0,51 2,08±0,36 1,41±0,17
RBC (x106/uL) 8,86±0,18 8,19±0,27 8,22±0,24 8,47±0,18
HGB (g/dL) 14,25±0,29 12,90±0,47 13,18±0,36 13,50±0,45
HCT (%) 45,78±1,27 41,62±1,57 40,78±1,37 42,93±1,45
MCV (fL) 51,68±1,09 50,80±0,70 49,60±0,42 50,62±0,71
MCH (pg) 16,10±0,20 15,75±0,31 16,05±0,18 15,93±0,20
MCHC (g/dL) 31,17±0,32 31,02±0,27 32,35±0,28 31,47±0,18
PLT (x103/uL) 944,00±51,19 1285,00±89,19* 823,50±70,72 1248,00±52,26*
W-SCR (%) 89,43±1,47 75,20±13,37 81,62±1,97 85,18±1,41
W-MCR (%) 0,00±0,00 0,00±0,00 0,00±0,00 0,00±0,00
W-LCR (%) 10,57±1,47 11,63±0,57 18,27±2,01 14,82±1,41
W-SCC (x103/uL) 3,36±0,44 2,75±0,46 1,73±0,31 1,21±0,16
W-MCC (x103/uL) 0,00±0,00 0,00±0,00 0,00±0,00 0,00±0,00
W-LCC (x103/uL) 0,45±0,10 0,30±0,02 0,35±0,06 0,20±0,02
RDW-SD (fL) 26,53±0,59 26,92±0,51 24,80±0,42 26,30±0,62
RDW-CV (%) 13,93±0,15 14,43±0,34 13,23±0,46 14,17±0,23
PDW (fL) 6,36±0,86 7,38±0,14 6,36±0,86 7,42±0,12
MPV (fL) 6,87±0,45 6,34±0,11 6,87±0,45 6,32±0,10
P-LCR (%) 6,80±0,55 6,50±0,62 6,80±0,55 4,60±0,25Número de leucócitos (WBC), número de eritrócitos (RBC), concentração de hemoglobina
(HGB), hematócrito (HCT), volume corpuscular médio (MVC), hemoglobina corpuscular média
(MCH), concentração média de hemoglobina corpuscular (MCHC), número de plaquetas (PLT),
razão percentual de linfócitos (W-SCR), razão percentual de monócitos (W-MCR), razão
percentual de neutrófilos (W-LCR), número de linfócitos (W-SCC), número de monócitos (W-
MCC), número de neutrófilos (W-LCC), amplitude da distribuição dos eritrócitos (RDW-SD),
coeficiente de variação ao redor da média do volume das células vermelhas (RDW-CV), índice
de amplitude de distribuição do tamanho das plaquetas (PDW), volume plaquetário médio
(MPV) e percentagem de plaquetas gigantes (P-LCR). * Diferença significativa (p<0,05).

144
Figura 3.6: Fotomicrografia de cortes histológicos, corados com hematoxilina eeosina, dos órgãos: coração (A e B), fígado (C e D), rim (E e F), baço (G e H),timo (I e J) e linfonodo (K e L) de camundongos machos Swiss tratados comsalina (figuras da esquerda) e 10 mg/kg dos PT de A.barbadensis (figuras dadireita). Os resultados referentes às fêmeas se mostraram idênticos aosmachos. Aumento 400x.

145
3.3. CONCLUSÃO
Os PT e as frações PI e PII não apresentaram citotoxicidade para às
células Vero, C6/36, HEp-2 e HeLa. No entanto, para células LLC-MK2, os PT
e as frações PI e PII apresentaram citotoxicidade para as concentrações
maiores testadas.
Os PT e as frações PI e PII não foram eficientes para inibir a infecção
causada pelos vírus não envelopados Ad-19 e Ad-41.
Os resultados com os HSV, vírus envelopados de DNA, mostraram que
os PT e a fração PI foram capazes de inibir os vírus tipo 1 e 2. Em contraste,
PII inibiu apenas HSV-2.
Em relação ao HMPV, vírus envelopados de RNA-negativo, a fração PII
apresentou inibição de, aproximadamente, 100%, enquanto PI, não apresentou
atividade antiviral. Essa inibição foi acompanhada pela apresentação de IS
4,5 vezes mais alto do que a ribavirina.
Os PT e as frações PI e PII não apresentaram atividade antiviral para
DENV-1, vírus envelopado de RNA-positivo.
A fração PI apresentou efeito anti-hemorrágico e prócoagulante
diminuindo o TP, agindo, provavelmente, na fase inicial da coagulação.
Os PT não apresentaram toxicidade em camundongos quando foram
analisados os parâmetros bioquímicos, hematológicos e histopatológicos.
Os PT aumentaram o baço e o número das plaquetas de camundongos.
Pode ser concluído que polissacarídeos de A. barabdensis além de
apresentar atividade inibitória contra os vírus HSV-1, HSV-2 e HMPV, podem
também apresentar efeito anti-hemorrágico e prócoagulante, propriedades
essas, importantes, visto que a complicação de muitas viroses leva a quadros
hemorrágicos. Além disso, os PT de A. barbadensis não apresentaram
toxicidade expressiva, podendo ser utilizada como agente terapêutico seguro e
eficaz.


147
ABED, Y.; BOIVIN, G. Treatment of respiratory virus infections. Antiviral
Research, v. 70, p. 1-16, 2006.
ACOSTA, E. G.; CASTILLA, V.; DAMONTE, E. B. Infectious dengue-1 virus
entry into mosquito C6/36 cells. Virus Research, v. 160, p. 173-179, 2011.
ALBERSHEIM, P.; DARVILL, A. G.; O’NEILL, M. A.; SCHOLS, H. A.;
VORAGEN, A. G. J. An hypothesis: the same six polysaccharides are
components of the primary cell walls of all higher plants. Progress in
biotechnology: Pectins and Pectinases, v. 14, p. 47-55, 1996.
ALVES, A. P.; MULLOY, B., MOY, G. W.; VACQUIER, V. D.; MOURÃO, P. A.
Females of the sea urchin Strongylocentrotus purpuratus differ in the strutures
of their egg jelly sulfated fucans. Glycobiology, v. 8(9), p. 939-946, 1998.
ALVES, D. S.; PÉRZ-FONS, L.; ESTEPA, A.; MICOL, V. Membrane-related
effects underlying activity of the anthraquinones emodin and barbaloin.
Biochemical Pharmacology, v. 68, p. 549-561, 2004.
AQUINO, R. S.; GRAVITOL, C.; MOURÃO, P. A. S. Rising from the sea:
correlations between sulfated polysaccherides and salinity in plants. Plos One,
v. 6 (4), p. 1-7, 2011.
AQUINO R. S.; LANDEIRA-FERNANDEZ, A. M.; VALENTE, A. P.; ANDRADE,
L. R.; MOURÃO, P. A. S. Ocurrence of sulfated galactans in marine
angiosperms: evolutionary implications. Glycobiology, v. 15(1), p. 11-20, 2005.

148
ARENA, A.; MAUGERI, T. L.; PAVONE, B.; IANNELO, D.; GUGLIANDOLO, C.;
BISIGNANO, G. Antiviral and immunoregulatory effect of a novel
exopolysaccharide from a marine thermotolerant Bacillus licheniformis.
International Immunopharmacology, v. 6, p. 8-13, 2006.
ASHLEY, R.; WALD, A. Genital herpes: review of the epidemic and potential
use of type-specific serology. Clin. Microbiol. Rev., v.12, p. 1-8, 1999.
AVIGAD, G.; DEY, P. M. Carbohydrate metabolismo: storage carbohydrate. In:
Dey, P. M. e Harborne, J. B. (Eds). Plant Biochemistry, Academic Press,
California, p. 143-204, 1997.
BANDYOPADHYAY, S. S.; NAVID, M. H.; GHOSH, T.; SCHNITZLER, P.; RAY,
B. Structural features and in vitro antiviral activities of sulfated polysaccharides
from Sphacelaria indica. Phytochemistry, v. 72, p. 276-283, 2011.
BARBOSA, J. E. F. Estudo da atividade citotóxica e antiviral (anti-HSV-1 e
anti-HIV-1) dos derivados N-amino-Triazóis. 131p. 2009. Dissertação
(Mestrado em Neuroimunologia), Universidade Federal Fluminense, Niterói,
2009.
BOREFREUND, E.; PUERNER, J. Toxicity determined in vitro by morphological
alterations and neutral red absorption. Toxicol. Lett., v. 24, p. 119–124, 1985.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding.
Analytical Biochemistry, v. 72, p. 248-254, 1976.

149
BRADY, R.; BERNSTEIN, D. Treatment of herpes simplex infections. Antiviral
Research, v. 61, p. 73-81, 2004.
BRASHER, W. J.; ZIMMERMANN, E. R.; COLLINGS, C. K. The effects of
prednisolone, indomethacin, and Aloe vera gel on tissue culture cells. Oral
Surgery, Oral Medicine and Oral Pathology, v. 27, p. 122-128, 1969.
BUCKERIDGE, M. S.; SANTOS, H. P.; TINÉ, M. A. S. Mobilisation of storage
cell wall polysaccharides in seeds. Plant Physiol. Biochem., v. 38, p. 141-156,
2000.
CAMPESTRINI, L. H. Aloe barbadensis Miller: análise do perfil metabólico
e estudos dos efeitos vasculogênicos e angiogênicos do extrato do
parênquima de reserva, da fração polissacarídica (FP) e da acemanana.
164f. 2007. Dissertação (Mestrado em Biotecnologia) – Universidade Federal
de Santa Catarina, Florianópoles, 2007.
CAMPO, V. L.; KAWANO, D. F.; SILVA JÚNIOR, D. B.; CARVALHO, I.
Carrageenans: Biological properties, chemical modifications ans structural
analysis – A review. Carbohydrate Polymers, v. 77, p. 167-180, 2009.
CARPITA, N. C.; GIBEAUT, D. M. Structural models of primary cell walls in
flowering plants: consistency of molecular structure with the physical properties
of the walls during growth. Plant J., v. 3, p. 1-30, 1993.

150
CARPITA, N. C.; Mc CANN, M. The cell wall. In: Buchanan, B. B.; Gruissem,
W.; Jones, R. L. (Eds.). Biochemistry and Molecular Biology of Plants.
American Society of Plant Physiologist, Maryland, pp. 81, 2000.
CARRINGTON LABORATORIES. Bill H. McAnalley. Robert H. Carpenter.
Harley R. McDaniel. Administration of acemannan. US 005106616 – A, 21 de
Abril de 1992.
CASSOLATO, J. E. F.; NOSEDA, M. D.; PUJOL, C. A.; PELLIZZARI, F. M.;
DAMONTE, E. B.; DUARTE, M. E. R. Chemical structure and antiviral activity of
the sulfated heterorhamnan isolated from the green seaweed Gayralia
oxysperma. Carbohydrate Research, p. 343, p. 3085-3095, 2008.
CHANG, X. L.; CHEN, B. Y.; FENG, Y. M. Water-soluble polysaccharides
isolated from skin juice, gel juice and flower of Aloe vera Miller. Journal of the
Taiwan Institute of Chemical Engineers, v. 42, p. 197-203, 2011.
CHANG, X. L.; FENG, Y. M.; WANG, W. H. Comparison of the polysaccharides
isolated from skin juice, gel juice and flower of Aloe arborescens tissues.
Journal of the Taiwan Institute of Chemical Engineers, v. 42, p. 13-19,
2011.
CHATTOPADHYAY, K.; MATEU, C. G.; MANDAL, P.; PUJOL, C. A.;
DAMONTE, E. B.; RAY, B. Galactan sulfate of Grateloupia indica: Isolation,
structural features and antiviral activity. Phytochemistry, v. 68, p. 1428-1435,
2007.

151
CHINNAH, A. D.; BAIG, M.; TIZARD, I. R.; KEMP, M. C. Antigen-dependent
adjuvant activity of a polydispersed b-(1,4)-linked acetylated mannan. Vaccine
v. 10, p. 551–558, 1992.
CHOW, J. T-N.; WILLIAMSON, D. A.; YATES, K. Y.; GOUX, W. J. Chemical
characterization of the immunomodulating polysaccharide of Aloe vera L..
Carbohydrate Research, v. 340, p. 1131-42, 2005.
CHUNG, M. H.; CHOI, S. W. The wound healing effect. In: Park, Y. I. e Lee, S.
K. eds, New Perspective on Aloe, Springer, Nova York, p. 63-82, 2006.
CHUN-HUI, L.; CHANG-HAI, W.; ZHI-LIANG, X., YI, W. Isolation, chemical
characterization and antioxidant activities of two polysaccharides from the gel
and the skin of Aloe barbadensis Miller irrigated with sea water. Process
Biochemistry, v. 42, p. 961-970, 2007.
CINELLI, L. P. Glicoconjugados sulfatados presentes na matriz gelatinosa
dos óvulos de ouriço-do-mar. 128f. 2009. Tese (Doutorado em Ciências) –
Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2009.
CIPRIANI, T. R.; MELLINGER, C. G.; GORIN, P. J.; IACOMINI, M. An
arabinogalactan isolated from the medicinal plant Maytenus ilicifolia. J. Nat.
Prod., v. 67, p. 703-706, 2004.
COPPA, G. V.; BUZZEGA, D.; ZAMPINI, L.; MACCARI, F.; GALEAZZI, T.;
PEDERZOLI, F.; GABRIELLI, O.; VOLPI, N. Effect of 6 years of enzyme
replacement therapy on plasma and urine glycosaminoglycans in attenuated
MPS I patients, Glycobiology, v. 20, n. 1259-1273, 2010.

152
COSGROVE, D. J. Growth of the plant cell wall. Nature Reviews – Molecular
Cell Biology, v. 6, p. 850-861, 2005
COSTA, L. S.; FIDELIS, G. P.; CORDEIRO, S. L.; OLIVEIRA, R. M.; SABRY, D.
A.; CÂMARA, R. B. G.; NOBRE, L. T. D. B.; COSTA, M. S. S. P.; ALMEIDA-
LIMA, J.; FARIAS, E. H. C.; LEITE, E. L.; ROCHA, H. A. O. Biological activities
of sulfated polysaccharides from tropical seaweeds. Biomedicine &
Pharmacotherapy, v. 64, p. 21-28, 2010.
CSEKE, G.; MAGINNIS, M. S.; COX, R. G.; TOLLEFSON, S. J.; PODSIAD, A.
B.; WRIGHT, D. W.; DERMODY, T. S.; WILLIAMS, J. V. Integrin alphavbeta1
promotes infection by human metapneumovirus. Proc. Natl. Acad. Sci. USA, v.
106, p. 1566-1571, 2009.
CUI, S. W.; WANG, Q. Cell wall polysaccharides in cereals: chemical structures
and functional properties, Structural Chemistry, v. 20, p. 291-297, 2009.
CUNNINGHAM, A.; GRIFFITHS, P.; LEONE, P.; MINDEL, A.; PATEL, R.;
STANBERRY, L.; WHITLEY, R. Current management and recommendations for
access to antiviral therapy of herpes labialis. Journal of Clinical Virology, v.
53, p. 6-11, 2012.
DAMONTE, E. B.; MATULEWICZ, M. C.; CEREZO, A. S.; COTO, C.E. Herpes
simplex virus-inhibitory sulfated xylogalactans from the red seaweed
Nothogenia fastigiata. In: DUARTE, M. E. R.; CAUDURO, J. P.; NOSEDA, D.
G.; NOSEDA, M. D.; GONÇALVES, A. G.; PUJOL, C. A.; DAMONTE, E. B.;
CEREZO, A. S. The structure of the agaran sulfate from Aconthophora spicifera

153
(Rhodomelaceae, Ceramiales) and its antiviral activity in agarans.
Carbohydrate Research, v. 339, p. 335-347, 2004.
DANOF, I. E.; McANALLEY, W. Stabilised Aloe vera: effect on human skin
cells. Drugs and Cosmetic Industry, v. 133 (52), p. 105-106, 1983.
DAVIE, E. W.; RATNOFF, O. D. Waterfall sequence for intrinsic blood clotting.
Science, v. 145, p. 1310-1312, 1964.
DE CLERCQ, E. D. Antivirals and antiviral strategies. Nature Reviews –
Microbiology, v. 2, p. 704-720, 2004.
DE CLERCQ, E. D.; DESCHAMPS, J.; VERHELST, G.; WALKER, R. T.;
JONES, A. S.; TORRENCE, P. F.; SHUGAR, D. Comparative efficacy of
antiherpes drugs against different strains of herpes simplex virus. J. Infect.
Dis., v.141, p.563-574, 1980.
DIETRICH, C. P.; DIETRICH, S. M. C. Electrophoretic behavior of acidic
mucopolysaccharides in diamine buffers. Analytical Biochemistry, v. 70, n.2,
p. 645-647, 1976.
DINAND, E.; VIGNON, M. R. Isolation and characterization of a (4-O-metil-D-
glucurono)-D-xylan from sugar beet pulp. Carbohydr. Res., v. 330, p. 285-288,
2001.
DISCHE, Z. A new specific color reaction of hexuronic acids. J. Biol. Chem., v.
167, p. 189-198, 1974.

154
DODGSON, K. S. Determination of inorganic sulphate in studies on the
enzymatic and non-enzymatic hydrolysis of carbohydrate and other sulphate
esters. Biochemistry Journal, v. 78, p. 312-319, 1961.
DONG, C-X.; HAYASHI, K.; MIZUKOSHI, Y.; LEE, J-B.; HAYASHI, T.
Structures and anti-HSV-2 activities of neutral polysaccharides from and edible
plant, Basella rubra L. International Journal of Biological Macromolecules,
v. 50, p. 245-249, 2012.
DONOSO A. F.; LEÓN, J. A.; CAMACHO, J. F.; CRUCES, P. I.; FERRÉS, M.
Fatal hemorrhagic pneumonia caused by human metapneumovirus in na
immunocompetent child. Pediatr. Intern., v. 50, p. 589-591, 2008.
DUARTE, M. E. R.; CAUDURO, J. P.; NOSEDA, D. G.; NOSEDA, M. D.;
GONÇALVES, A. G.; PUJOL, C. A.; DAMONTE, E. B.; CEREZO, A. S. The
structure of the agaran sulfate from Aconthophora spicifera (Rhodomelaceae,
Ceramiales) and its antiviral activity in agarans. Carbohydrate Research, v.
339, p. 335-347, 2004.
DUARTE, M. E. R.; NOSEDA, D. G.; NOSEDQA, M. D.; TULIO, S.; PUJOL, C.
A.; DAMONTE, E. B. Inhibitory effect of sulfated galactans from the marine alga
Botrychia montagnei on herpes simplex virus replication in vitro.
Phytomedicine, v. 8 (1), pp. 53-58, 2001.
DUBOIS, C. Colorimetric method for determination of sugars and related
substances. Analytical Chemistry, v. 28, p. 350– 356, 1956.

155
DUMVILLE, J. C.; FRY, S. C. Uronic acid-containing oligosaccharins: Their
biosynthesis, degradation and signaling roles in non-diseased plant tissues.
Plant Physiol. Biochem., v. 38 (1/2), p. 125-140, 2000.
EO, S-K., KIM, Y-S., LEE, C-K., HAN, S-S. Antiherpetic activities of various
protein bound polysaccharides isolated from Ganoderma lucidum. Journal of
Ethnopharmacology, v. 68, p. 175-181, 1999.
ESUA, M. F.; RAUWALD, J-W. Novel bioactive maloyl glucans from Aloe vera
gel: isolation, structure elucidation and in vitro bioassays. Carbohydrate
Research, v. 341, p. 355-354, 2006.
FANG, X.; JIANG, B.; WANG, X. Purification and partial characterization of an
acidic polysaccharide with complement fixing ability from the stems of Avicennia
Marina. Journal of Biochemistry and Molecular Biology, v. 39, p. 546-555,
2006.
FATAHZADEH, M.; SCHWART, R. A. Human herpes simplex virus infections:
Epidemiology, pathogenesis, symptomatology, diagnosis, and management. J.
Am. Acad. Dermatol., v.57 (5), p. 737-763, 2007.
FEMENIA, A.; SÁNCHEZ, E. S.; SIMAL, S.; ROSSELLÓ, C. Compositional
features of polysaccharides from Aloe vera (Aloe barbadensis Miller) plant
tissues. Carbohydrate Polymers, v. 39, p. 109-117, 1999.
FEMENIA, A.; GARCÍA-PASCUAL, P.; SIMAL, S.; ROSSELLÓ, C. Effects of
heat treatment and dehydratation on bioactive polysaccharide acemannan and

156
cell wall polymers from Aloe barbadensis Miller. Carbohydrate Polymers, v.
51, p. 397-405, 2003.
FERREIRA, C. N.; SOUSA, M. O.; DUSSE, L. M. S.; CARVALHO, M. G. O
novo modelo da cascata de coagulação baseado nas superfícies celulares e
suas implicações. Revista Brasileira de Hematologia e Hemoterapia, v. 32
(5), p. 416-421, 2010.
FEUILLET, F.; LINA, B.; ROSA-CALATRAVA, M.; BOIVIN, G. Ten years of
human metapneumovirus research. Journal of Clinical Virology, v. 53, p. 97-
105, 2012.
FIGUEIREDO, L. T. M. Febres hemorrágicas por vírus no Brasil. Revista da
Sociedade Brasileira de Medicina Tropical, v. 39 (2), p. 203-210, 2006.
FOGLEMAN, R. W.; CHAPDELAINE, J. M.; CARPENTER, R. H.; McANALLEY,
B. H. Toxicologic evaluation of injectable acemannan in the mouse, rat and dog.
Vet. Hum. Toxicol., v. 34(3), p. 201-205, 1992.
FONSECA, R. J. C. Desafios no estudo de novos polissacarídeos
antitrombóticos: o condroitim sulfato fucosilado. 194p. 2009. Tese
(Doutorado em Química Biológica), Universidade Federal do Rio de Janeiro,
Rio de Janeiro, 2009.
FRIEDMAN, H.; SPECTER, S.; BENDINELLI, M. In vitro models of HIV
disease and control. Springer US Publisher, p. 336-364, 2006.

157
FRY, S. C. Cell wall polysaccharide composition and covalent crosslinking. In:
Ulvskov, P. (Ed.). Annual Plant Reviews, Blackwell Publishing Ltd, First
edition, v. 41, p. 1-42, 2011.
FRY, S. C.; YORK, W. S.; ALBERSHEIM, P.; DARVILL, A.; HAYASHI, T.;
JOSELEAU, J. P.; KATO, Y.; LORENCES, E. P.; MacLACHLAN, G. A.;
McNEIL, M.; MORT. A.; REID. J. S. G.; SEITZ, H. U.; SELVENDRAN, R. R.;
VORAGEN, A. G. J.; WHITE, A. R. An unambiguous nomenclature for
xyloglucan-derived oligosaccharides. Physiol. Plant., Denmark, v. 89, p. 1-3,
1993.
GABRIELLI, I.; GATENHOL, P.; GLASSER, W. G.; JAIN, R. K.; KENNE, L.
Separation, characterization and hydrogel-formation of hemicellulose from
aspen wood. Carbohydrate Polymers, v. 43, p. 367-374, 2000.
GAINOTTI, R.; RICARTE, C.; EBEKIAN, B.; VIDELA, C.; CARBALLAL, G.;
DAMONTE, E. B.; ECHAVARRÍA, M. Real time PCR for rapid determination of
susceptibility of adenovírus to antiviral drugs. Journal of Virological Methods,
v. 164, p. 30-34, 2010.
GARRON, M-L.; CYGLER, M. Structural and mechanistic classification of uronic
acid-containing polysaccharide lyases. Glycobiology, v. 20, p. 1547-1573,
2010.
GAUNTT, C. J.; WOOD, H. J.; McDANIEL, H. R.; McANALLEY. Aloe
polymannose enhances anti-coxsackievirus antibody titres in mice.
Phytotherapy Research, p. 14 (4), p. 261-266, 2000.

158
GEMIN, E. Determinação da atividade antiviral de galactomananas
sulfatadas contra o vírus herpes simplex tipo 1 (HSV-1), 111p. 2008.
Dissertação (Mestrado em Ciências Biológicas) – Universidade Federal do
Paraná, Curitiba, 2008.
GHOSH, T.; CHATTOPADHYAY, K.; MARSCHALL, M.; KARMAKAR, P.;
MANDAL, P.; RAY, B. Focus on antivirally active sulfated polysaccharides:
From structure-activity analysis to clinical evaluation. Glycobiology, v. 19, p. 2-
15, 2009.
GIBEAUT, D. M.; PAULY, M.; BACIC, A.; FINCHER, G. B. Changes in cell wall
polysaccharides in developing barley (Hordeum vulgare) coleoptiles. Planta,
v. 221, p. 729 – 738, 2005
GOWDA, D. C.; NEELISIDAIAH, B.; ANJANEYALU, Y.V. Structural studies of
polysaccharides from A. vera. Carbohydrate research, v. 72, p. 201-205,
1979.
GRINDLAY, D.; REYNOLDS, T. The Aloe vera phenomenon: a review of the
properties and modern uses of the leaf parenchyma gel. Journal of
Ethnopharmacology, v. 16, p. 117-151, 1986.
GU, W.; SONG, H.; WEN, X.; WANG, Y.; XIA, W.; FANG, Y. Binding interaction
between aloe polysaccharide and alizarin red by spectrophotometry and its
analytical application. Carbohydrate Polymers, v. 80, p. 115-122, 2010.
GUGLANI, L.; KABRA, S. K. T cell immunopathogenesis of dengue virus
infection. Dengue Bulletin, v. 29, p. 58-69, 2005.

159
HABEEB, F.; SHAKIR, E.; BRADBURY, F.; CAMERON, P.; TARAVATI, M. R.;
DRUMMOND, A. J.; ALEXANDER, I. G.; FERRO, V. A. Screening methods
used to determine the anti-microbial properties of Aloe vera inner gel. Methods,
v. 42, p. 315-20, 2007.
HAMMAN, J. h. Composition and applications of Aloe vera leaf gel. Molecules,
v. 13, p. 1599-1616, 2008.
HARRIS, P. J; SMITH, B. G. Plant cell walls and cell-wall polysaccharides:
structures, properties and uses in food products. International Journal of
Food Science and Technology, v.41, p. 129-143, 2006.
HAYASHI, K.; NAKANO, T.; HASHIMOTO, M.; KANEKIYO, K.; HAYASHI, T.
Defensive effects of a fucoidan from brown alga Undaria pinnatifida against
herpes simplex virus infection. International Immunopharmacology, v. 8, p.
109-116, 2008.
HERBET, J. M.; BERNAT, A.; MAFFRAND, J. P. Importance of platelets in
experimental venous thrombosis in rats. Blood, v. 80, p. 2281-2286, 1992.
HERMOS, C. R.; VARGAS, S. O.; McADAM, A. J. Human Metapneumovirus.
Clin. Lab. Med., v. 30, p. 131-148, 2010.
HIDARI, K. I. P. J.; TAKAHASHI, N.; ARIHARA, M.; NAGAOKA, M.; MORITA,
K.; SUZUKI, T. Structure and anti-dengue vírus activity of sulfated

160
polysaccharide from a marine alga. Biochemical and Biophysical Research
Communications, v. 376, p. 91-95, 2008.
HIROKAWA, Y.; FUJIWARA, S.; TSUZUKI, M. Three types of acidic
polysaccharides associated with coccolith Pleurochrysis haptonemofera:
comparison with Pleurochrysis carterae and analysis using fluorescein-
isothiocyanate-labeled lectins, Marine Biotechnology, v. 7, p. 634-644, 2005.
HOFFMAN, M.; JIA, Z.; PEÑA, M. J.; CASH, M.; HARPER, A.;
BLACKBURN II, A. R.; DARVILL, A.; YORK, W. S. Structural analysis of
xyloglucans in the primary cell wall of plants in the subclass Asteridae.
Carbohydr. Res., v. 340, p. 1826-1840, 2005.
HUCK, B.; EGGER, M.; BERTZ, H.; PEYERL-HOFFMAN, G.; KERN, W. V.;
NEUMANN-HAEFELIN, D.; FALCONE, V. Human metapneumovirus infection in
a hematopoietic stem cell transplant recipient with relapse multiple myeloma
and rapidly progressing lung cancer. J. Clin. Microbiol., v. 44, p. 2300-2303,
2006.
IASC – THE INTERNATIONAL ALOE SCIENCE COUNCIL. Certification
program, IASC, Disponível em: www.iasc.org. Acessado em 07/01/2012.
IM, S.; OH, S-T.; SONG, S.; KIM, M-R.; KIM, D-S.; WOO, S-S.; JO, T. H.;
PARK, Y. I.; LEE, C-K. Identification of optimal molecular size of modified Aloe
polysaccharides with maximum immunomodulatory activity. International
Immunopharmacology, v. 5, p. 271-279, 2005.

161
ISHII, T. O-acetylated oligosaccharides from pectins of potato tuber cell walls.
Plant Physiology, v. 113, p. 1265-1272, 1997.
JAHANBIN, K.; GOHARI, A. R.; MOINI, S.; EMAM-DJOMEH, Z.; MASI, P.
Isolation, structural characterization and antioxidant activity of a new water-
soluble polysaccharide from Acanthophyllum bracteatum roots. International
Journal of Biological Macromolecules, v. 49, p. 567-572, 2011.
JANI, G. K.; SHAN, D. P.; PRAJAPATI, V. D.; JAIN, V. C. Gums and mucilages:
versatile excipients for pharmaceutical formulations. Asian Journal of
Pharmaceutical Sciences, v. 4(5), p. 309-323, 2009.
JARVIS, M. C. Plant cell walls: supramolecular assembly, signalling and stress.
Structural Chemistry, v. 20, p. 245-253, 2009.
JIAO, G.; YU, G.; ZHANG, J.; EWART, H. S. Chemical structures and
bioactivities of sulfated polysaccharides from marine algae. Marine Drugs, v. 9,
p. 196-223, 2011.
JOHANSSON, S. M. C.; NILSSON, E. C.; ELOFSSON, M.; AHLSKOG, N.;
KIHLBERG, J.; ARNBERG, N. Multivalent sialic acid conjugates inhibit
adenovirus type, 37 from binding to and infecting human corneal epithelial cells.
Antiviral Research, v. 73, p. 92-100, 2007.
JONES, M. S.; HARRACH, B.; GANAC, R. D.; GOZUM, M. M.; DE LA CRUZ,
W. P.; RIEDEL, B.; PAN, C.; DELWART, E.; SCHNURR, D. New adenovirus
species found in a patient presenting with gastroenteritis. J. Virol., v. 81, p.
5978–5984, 2007.

162
JÚNIOR, A. V. Alfa e beta-amilase no metabolismo do amido durante o
amadurecimento da banana: clonagem, expressão e caracterização
molecular. 91f. 2006. Tese (Doutorado em Ciências Farmacêuticas) –
Universidade de São Paulo, São Paulo, 2006.
KABAKUS, N.; GURGOZE, M. K.; YILDIRIM, H.; GODEKMERDAN, A.; AYDIN,
M. Acute hemorrhagic leukoencephalitis manifesting as intracerebral
hemorrhage associated with herpes simplex virus type I. Journal of Tropical
Pediatrics, v. 51 (4), p. 245-249, 2005.
KAÈURÁKOVÁ, M.; CAPEK, P.; SASINKOVÁ, V.; WELLNER, N.;
EBRINGEROVÁ, A. FT-IR study of plant cell wall model compounds: pectin
polysaccharides and hemicelluloses. Carbohydrate Polymers, v. 43, p. 195-
203, 2000.
KALAFATIS, M.; SWORDS, N. A.; RAND, M. D,; MANN, K. G. Membrane-
dependent reactions in blood coagulation: role of the vitamin K-dependent
enzyme complexes. Biochim. Biophis. Acta, v. 1227, p. 113-129, 1994.
KANG, J.; CUI, S. W.; PHILLIPS, G. O.; CHEN, J.; GUO, Q., WANG, Q. New
studies on gum ghatti (Anogeissus latifolia) Part III: Structure characterization of
a globular polysaccharide fraction by 1D, 2D NMR spectroscopy and
methylation analysis. Food Hydrocolloids, v.25, p. 1999-2007, 2011.
KARACA, K.; SHARMA, J. M.; NORDGREN, R. Nitric oxide production by
chicken macrophages activated by acemannan, a complex carbohydrate

163
extracted from Aloe vera. International Journal of Immunopharmacology, v.
17, p. 183-188, 1995.
KATO, D.; ERA, S.; WATANABE, I.; ARIHARA, M.; SUGIURA, N.; KIMATA, K.;
SUZUKI, Y.; MORITA, K.; HIDARI, K. I. P. J.; SUZUKI, T. Antiviral activity of
chondroitin sulphate E targeting dengue vírus envelope protein. Antiviral
Research, v. 88, p. 236-243, 2010.
KHAN, M. T. H.; ATHER, A.; THOMPSON, K. D.; GAMBARI, R. Extracts and
molecules from medicinal plants against herpes simplex viruses. Antiviral
Research, v. 67, p. 107-119, 2005.
KIRCHER, H. W. Gas-liquid chromatography of methylated sugar. Anal.
Chem., v. 32, p. 1103-1106, 1960.
KIM, Y-S., EO, S-K., OH, K-W., LEE, C-K., HAN, S-S. Antiherpetic activities of
acidic protein bound polysaccharide isolated from Ganoderma lucidum alone
and in combinations with interferons. Journal of Ethnopharmacology, v. 725,
p. 451-458, 2000.
KLEYMANN, G. Novel agents and strategies to treat herpes simplex virus
infections. Expert Opinion, v. 12, p. 165-183, 2003.
KNIPE; D. M.; HOWLEY, P. M. Fields Virology. In: GEMIN, E. Determinação
da atividade antiviral de galactomananas sulfatadas contra o vírus herpes
simplex tipo 1 (HSV-1), 111p. 2008. Dissertação (Mestrado em Ciências
Biológicas) – Universidade Federal do Paraná, Curitiba, 2008.

164
KITAYAMA, T.; MORIMOTO, M.; TAKATANI, M.; TAMURA, Y.; TAKUMI, M.;
OKAMOTO, T. Isolation of hemicellulose from a sorghum, Andropogon
sorghum brot, Kumadake no. 263, and determination of its constituent sugars.
Carbohydrate Res., v. 325, p. 230-232, 2000.
KOLENDER, A. A.; PUJOL, C. A.; DAMONTE, E. B.; MATULEWICZ, M. C.;
CEREZO, A. S. The system of sulfated á-(1à 3)-linked D-mannans from the red
seaweed Nothogenia fastigiata: structures, antiherpetic and anticoagulant
properties. Carbohydrate Research, v. 304, p. 53-60, 1997.
KURT-JONES, E. A.; CHAN, M.; ZHOU, S.; WANG, J.; REED, G.; BRONSON,
R.; ARNOLD, M. M.; KNIPE, D. M.; FINBERG, R. W. Herpes simplex virus 1
interaction with Toll-like receptor 2 contributes to lethal encephalitis.
Proceedings of the National Academy of Sciences, v. 101 (5), p.1315-1320,
2004.
LAGROTA, M. H. C. Atividade antiviral in vitro de substâncias sintéticas e
naturais. 1978. Dissertação (Mestrado em Microbiologia) – Universidade
Federal do Rio de Janeiro (UFRJ), Rio de Janeiro, 1978.
LAMBERT, J. B.; SHURVELL, H. F.; LIGHTNER, D. A.; COOKS, R. G. Organic
structural spectroscopy. In: CAMPESTRINI, L. H. Aloe barbadensis Miller:
análise do perfil metabólico e estudos dos efeitos vasculogênicos e
angiogênicos do extrato do parênquima de reserva, da fração
polissacarídica (FP) e da acemanana. 164f. 2007. Dissertação (Mestrado em
Biotecnologia) – Universidade Federal de Santa Catarina, Florianópoles, 2007.

165
LEE, C-J.; LIAO, C-L.; LIN, Y-L. Flavivirus activates phosphatidylinositol 3-
kinase signaling to block caspase-dependent apoptotic cell death at the early
stage of virus infection. Journal of Virology, v. 79 (13), p. 8388-8399, 2005.
LEE, C. K. Immunomodulatory activity. In: Park, Y. I. e Lee, S. K. eds, New
Perspective on Aloe, Springer, Nova York, p. 155-167, 2006.
LEE, J-B.; HOU, X.; HAYASHI, K.; HAYASHI, T. Effect of partial desulfation and
oversulfation of sodium spirulan on the potency of anti-herpetic activities.
Carbohydrate Polymers, v. 69, p. 651-658, 2007.
LEE, J. K., LEE, M. K., YUN, Y-P., KIM, Y., KIM, J. S., KIM, Y. S., KIM, K.,
HAN, S. S., LEE, C-K. Acemannan purified from Aloe vera induces phenotypic
and functional maturation of immature dendritic cells. International
Immunopharmacology, v. 1, p. 1275-84, 2001.
LEE, S. K. Overview of Aloe study. In: Park, Y. I. e Lee, S. K. eds, New
Perspective on Aloe, Springer, Nova York, p. 1-5, 2006.
LEFLORE, S.; ANDERSON, P. L.; FLETCHER, C. V. A risk-benefit evaluation
of acyclovir for the treatment and prophylaxis of herpes simplex virus infections.
Drug. Saf., v. 23, p. 131-142,, 2000.
LENAERTS, L.; NAESENS, L. Antiviral therapy for adenovirus infections.
Antiviral Research, v. 71, p. 172-180, 2006.

166
LEUNG, M. Y. K.; LIU, C.; ZHU, L. F.; HUI, Y. Z.; YU, B.; FUNG, K. P. Chemical
and biological characterization of a polysaccharide biological response modifier
from Aloe vera L. var. chinensis (Haw.) Berg. Glycobiology, v. 14 (6), p. 501-
510, 2004.
LEUNG, T. K. H.; BROWN, M. Block in entry of enteric adenovirus type 41 in
HEK293 cells. Virus Research, v. 156, v. 54-63, 2011.
LIMA, D. U. Polissacarídeos de reserva de parede cellular em sementes.
Estrutura, metabolismo funções e aspectos ecológicos. Revista Brasileira de
Fisiologia Vegetal, v. 12, p. 137-162, 2000.
LIND, S. E.; KURKJIAN, C. D. The bleeding time. In: Michelson, A. D. (Ed)
Platelets, Elsevier, pp. 485-493, 2007.
LING, Z.; TRAN, K. C.; TENG, M. N. The human respiratory syncytial virus
nonstructural NS2 protein antagonizes the activation of interferon-{beta}
transcription by interacting with RIG-I. J. Virol., v. 83 (8), p. 3734-3742, 2009.
LIU, J. J., HUANG, T. S., HSU, M. L., CHEN, C. C., LIN, W. S., LU, F. J.
Antitumor effects of the partially purified polysaccharides from Antrodia
camphorata and the mechanism of its action. Toxicol. Appl. Pharmacol., v.
201, p. 186–93, 2004.
LIU, J.; THORP, S. C. Cell surface heparan sulfate and its roles in assisting
viral infections. Med. Res. Rev., v. 22, p. 1-25, 2002.

167
LIU, T.; SCALLAN, C. D.; BROZE Jr., G.; PATARROYO-WHITE, S.; PIERCE,
G., F.; JOHNSON, K. W. Improved coagulation in bleeding disorders by Non-
anticoagulant Sulfated polysaccharides (NASP). Blood Coag., Fibrin. And
Cellular Haem., v. 95, p. 68-76, 2006.
LIU, X-X.; WN, Z-J.; SHI, L.; LU, X-X. Preparation and antiherpetic activities of
chemically modified polysaccharides from Polygonatum cyrtonema Hua.
Carbohydrate Polymers, v. 83, p. 737-741, 2011.
LUNDIN, A.; BERGSTRÖM, T.; ANDRIGHETTI-FRÖHNER, C. R.;
BENDRIOUA, L.; FERRO, V.; TRYBALA, E. Potent anti-respiratory syncytial
virus activity of a cholestanol-sulfated tetrasaccharide conjugate. Antiviral
Research, v. 93, p. 101-109, 2012.
LUTA, G.; DUNCAN, C.; SINNOTT, R. Chemical characterization of
polysaccharide-rich ingredientes from Aloe vera, Larix occidentalis, and Undaria
pinnatifida. In: 6th Annual Natural Supplements Conference, 2009, San Diego,
California. Disponível em: http://mannatechscience.org/files/Luta%20Scripps
%202009%20formposter.pdf. Acesso em: 12/12/2011.
MACFARLANE, R. G. An enzyme cascade in the blood clotting mechanism,
and its function as a biological amplifier. Nature, v. 202, p. 498-499, 1964.
MANDAL, P.; PUJOL, C. A.; CARLUCCI, M. J.; CHATTOPADHYAY, K.;
DAMONTE, E. B.; RAY, B. Anti-herpetic activity of a sulfated xylomannan from
Scinaia hatei. Phytochemistry, v. 69, p. 2193-2199, 2008.

168
MANDAL, P.; PUJOL, C. A.; DAMONTE, E. B.; GHOSH, T.; RAY, R. Xylans
from Scinaia hatei: Structural features, sulfation and anti-HSV activity.
International Journal of Biological Macromolecules, v. 46, p. 173-178, 2010.
MANN, K. G. Thrombin formation. In: FONSECA, R. J. C. Desafios no estudo
de novos polissacarídeos antitrombóticos: o condroitim sulfato
fucosilado. 194p. 2009. Tese (Doutorado em Química Biológica), Universidade
Federal do Rio de Janeiro, Rio de Janeiro, 2009.
MANNI, L.; ALOE, L. Role of IL-1â and TNF-á in the regulation of NGF in
experimentally induced arthritis in mice. Rheumatol. Int., v. 18, p. 97-102,
1998.
MARSH, M. E.; CHANG, D. K.; KING, G. C. Isolation and characterization of a
novel acidic polysaccharide containing tartrate and glyoxylate residues from the
mineralized scales of a unicellular coccolithophorid alga Pleurochrysis carterae.
The Journal of Biological Chemistry, v. 267, p. 20507-20512, 1992.
MARINHEIRO, J. C. Variabilidade genética da adenovírus humano da
espécie B, associados a casos de infecção respiratória aguda, em São
Paulo, de 1995 a 2006. 69p. 2009. Tese (Doutorado em Ciências –
Microbiologia), Universidade de São Paulo, São Paulo. 2009.
MATHUR, V.; MATHUR, N. K. Fenugreek and lesser known legume
galactomannan-polysaccharides: Scope for developments. Journal of
Scientific & Industrial Research, v. 64, p. 475-481, 2005.

169
MATSUDA, Y.; YOKOHIRA, M.; SUZUKI, S.; HOSOKAWA, K.; YAMAKAWA,
K.; ZENG, Y.; NINOMIYA, F.; SAOO, K.; KUNO, T.; IMAIDA, K. One-year
chronic toxicity of Aloe arborescens Miller var. natalensis Berger in Wistar
Hannover rats. A pilot study. Food and Chemical Toxicology, v. 46(2), p. 733-
739, 2007.
MAY, C. D. Pectins. In: Phillips, G. O.; Williams, P. A. (Eds.). Handbook of
Hydrocolloids. Cambridge: Wood-head Publishig, p. 169-188, 2000.
MAZUNDER, S.; GHOSAL, P. K.; PUJOL, C. A.; CARLUCCI, M. J.; DAMONTE,
E. B.; RAY, B. Isolation, chemical investigation and antiviral activity of
polysaccharides from Gracilaria corticata (Gracilariaceae, Rhodophyta). Int. J.
of Biol. Macromol., v. 31, p. 87–95, 2002.
McCONAUGHY, S. D.; STROUD, P. A.; BOUDREAUX, B.; HESTER, R. D.;
McCORMICK, C. L. Structural characterization and solution properties of a
galacturonate polysaccharide derived from Aloe vera capable of situ gelation.
Biomacromolecules, p. 472-480, 2008.
MEDICALTRUE. Herpes Simplex Virus (HSV). Disponível em:
http://medicaltrue.com/herpes-infection-caused-by-herpes-simplex-virus-hsv.
Acessado em 07/02/2012.
MEHNERT, D. U.; CANDEIAS, J. A. N. Herpesvírus. In: Trabulsi, L. R. and
Alterthum, F. (Eds), Microbiologia, Editora Atheneu, 4a. edição, p. 599-606,
2005.

170
MELLINGER, C. G. Caracterização estrutural e atividade biológica de
carboidratos de Phyllanthus niruri (Quebra-pedra). 139f. 2006. Tese
(Doutorado em Ciências Biológicas) – Universidade Federal do Paraná,
Curitiba, 2006.
MELTON, L. D.; SMITH, B. G. Determination of the uronic acid content of plant
cell walls using a colorimetric assay. Current Protocols in Food Analytical
Chemistry, p. E3.3.1-3.3.4, 2001.
MENDES, G. S.; SOARES, A. R.; MARTINS, F. O.; ALBUQUERQUE, M. C. M.;
COSTA, S. S.; YONESHINGUE-VALENTIN, Y.; GESTINARI, L. M. S.;
SANTOS, N.; ROMANOS, M. T. V. Antiviral activity of the green marine alga
Ulva fasciata on the replication of human metapneumovirus. Rev. Inst. Med.
Trop. S. Paulo, v. 52 (1), p. 3-10, 2010.
MIAO, B., GENG, W., LI, J., LI, F., CHEN, H., GUAN, H., DING, J. Sulfated
polymannuroguluronate, a novel anti-acquired immune deficiency syndrome
(AIDS) drug candidate, targeting CD4 in lymphocytes. Biochemical
Pharmacology, v. 98, p. 641-649, 2004.
MILLER, C. S.; DANAHER, R. J.; JACOB, R. J. Molecular aspects of herpes
simplex virus latency, reactivation and recurrence. Crit. Rev. Oral Biol. Med.,
v. 9, p. 541-562, 1998.
MOLLER, I.; SORENSEN, I.; BERNAL, A. J.; BLAUKOPF, C.; LEE, K.; OBRO,
J., PETTOLINO, F.; ROBERTS, A.; MIKKELSEN, J. D.; KNOX, J. P.; BACIC,
A.; WILLATS, W. G. High-throughput mapping of cell-wall polymers within and
between plants using novel microarrays. Plant Journal, v. 50, p. 1118-1128,
2007.

171
MONTANER, J. K.; GILL, J.; SINGER, J.; RABOUD, J.; ARSENEAU, R.;
MCLEAN, B. D.; SCHECHTER, M. T.; RUEDY, J. Doubleblind placebo-
controlled pilot trial of acemannan in advanced human immunodeficiency virus
disease. Journal of Aquired Immune Deficiency Syndromes and Human
Retrovirology, v. 12, p. 153–157, 1996.
MUENCH, J.; VERDIECK, A.; LOPEZ-VASQUEZ, A.; NEWELL, M. crossing
diagnostic borders: Herpes encephalitis complicated by cultural and language
barriers. JABFB, v. 14(1), p. 46-50, 2001.
MUKHOPADHYAY, S.; KUHN, R. J.; ROSSMANN, M. G. A structural
perspective of the flavivirus life cycle. Nature Reviews – Microbiology, v. 3, p.
13-22, 2005.
MÜLLER, V. D. M. Triagem antiviral de extratos vegetais: Fracionamento
biomonitorado de Ilex paraguariensis A. St. Hil. Aquifoliaceae (erva-mate).
116p., 2006. Dissertação (Mestrado em Farmácia), Universidade Federal de
Santa Catarina, Florianópolis, 2006.
MURPHY, K.; TRAVERS, P.; WALPORT, M. Imunubiologia de Janeway.
Artmed, 908p., 7a. edição, 2010.
MURRELL, S.; WU, S-C.; BUTLER, M. Review of dengue virus and the
development of a vaccine. Biotechnology Advances, v. 29, p. 239-247, 2011.

172
NARDONE, R. M. Toxicity testing in vitro. In: MÜLLER, V. D. M. Triagem
antiviral de extratos vegetais: Funcionamento biomonitorado de Ilex
paraguariensis A. St. Hil. Aquifoliaceae (erva-mate). 116p. 2006.
Dissertação (Mestre em Farmácia), Universidade Federal de Santa Catarina,
Florianópolis, 2006.
NELSON, D.; COX, M. M. Lehninger – Principles of Biochemistry. 5. Ed.
New York: W. H. Freeman and Company, pp. 1100, 2008.
NEWTON, L. E. Aloe in habitat. In: Renolds, T. (Ed.). Aloes – The genus Aloe,
CRC Press LLC, New York, p. 1-13, 2004.
NI, Y.; TURNER, D.; YATES, K. M.; TIZARD, I. Isolation and characterization of
structural components of Aloe vera L. leaf pulp. International
Immunopharmacology, v. 4, p. 1745-1755, 2004.
NI, Y.; YATES, K. M.; TIZARD, I. R. Aloe polysaccharides. In: Renolds, T. (Ed.).
Aloes – The genus Aloe, CRC Press LLC, New York, p. 73-85, 2004.
NISHINARI, K.; TAKEMASA, M.; ZHANG, H.; TAKAHASHI, R. Storage plant
polysaccharides: xyloglucans, galactomannans, glucomannans. In: Kamerling,
J. P. (Ed.). Comprehensive Glycoscience – From Chemistry to Systems
Biology, Elsevier, Utrecht University, v. 4, p. 614-646, 2007.
NISHIMURA, T.; TOKU, K.; FUKUYASU, H. Antiviral compounds. XII. Antiviral
activity of aminohydrazones of alkoxyphenyl substituted carbonyl compounds
against influenza virus in eggs and mice. Kitasato Arch. Exp. Med., v. 50, p.
39-46, 1977.

173
O’LOONEY, N.; FRY, S. C. Oxaziclomefone, a new herbicide, inhibits wall
expansion in maize cell-cultures without affecting polysaccharide biosynthesis,
xyloglucan transglycosylation, peroxidase action or apoplastic ascorbate
oxidation. Annals of Botany, v.96, 1097-1107.
O’NEILL, M. A.; YORK, W. S. The composition and structure of plant primary
cell walls. In: Rose, J. K. C. (Eds.). The plant cell wall. Annual Plant review,
CRC Press, v.8, p. 1-54, 2003.
ORMISTON, M. L.; SLAUGHTER, G. R. D.; DENG, Y., STEWART, D. J.;
COURTMAN, D. W. The enzymatic degradation of hyaluronan is associated
with disease progression in experimental pulmonary hypertension. Am J
Physiol Lung Cell Mol Physiol, v. 298, p. L148-L157, 2010.
PAVÃO, M. S. G.; AIELLO, K. R. M.; WERNECK, C. .; SILVA, L. C. F.;
VALENTE, A. P.; MULLOY, B.; COLWELL, N. S.; TOLLEFSEN, D. M.;
MOURÃO, P. A. S. Highly sulfated dermatan sulfate from Ascidians: structure
versus anticoagulant activity of these glycosaminoglycans. J. Biol. Chem., v.
273, p. 27848-27857, 1998.
PEREIRA, M. G.; BENEVIDES, N. M. B., MELO, M. R. S.; VALENTE, A. P.;
MELO, F. R.; MOURÃO, P. A. S. Structure and anticoagulant activity of a
sulfated galactan from the red alga, Gelidium crinale. Is there a specific
structural requirement for the anticoagulant action? Carbohydrate Research,
Oxford, v. 340, n.12, p. 2015-2023, 2005.
PEREIRA, M. S.; MULLOY, B.; MOURÃO, P. A. S. Structure and
anticoagulante activity of sulfated fucans. Comparision between the regular,
repetitive and linear fucans from echinoderms with the more heterogeneous and

174
branched polymers from brow algae. J. Biol. Chem., v. 274, p. 7656-7667,
1999.
PEUSER, M. Os capilares determinam o nosso destino: Aloe, imperatriz
das plantas medicinais, fonte de vitalidade e saúde, Diadema, SP, Hubertus
Produtos Naturais, 333p., 2003.
POMIN, V. H. Fucanas e galactanas sulfatadas: estrutura, biologia e
filogenia. 246p. 2008. Tese (Doutorado em Química Bilógica) – Universidade
Federal do Rio de janeiro, Rio de Janeiro, 2008.
POPPER, Z. A. Evolution and diversity of green plant cell walls. Current
Opinion in Plant Biology, v. 11, p. 197-199, 2008.
PRADO-FERNANDES, J.; RODRIGUEZ-VÁZQUEZ, J. A.; TOJO, E.;
ANDRADE, J. M. Quantitation of κ, ι and λ- carrageenans by mid-infrared
spectroscopy and PLS regression. Analytica Chimica Acta, v. 480, p. 23-37,
2003.
PRASHANTH, M. R. S.; PARVATHY, K. S.; SUSHEELAMMA, N. S.;
PRASHANTHY, K. V. H.; THARANATHAN, R. N.; CHA, A.; ANILKUMAR, G.
Galactomannan esters – A simple, cost-effective method of preparation and
characterization. Food Hydrocolloids, v. 20, p. 1198-1205, 2006.
PUGH, N.; ROSS, S.A.; ELSOHLY, M. A.; PASCO, D. S. Characterization of
aloeride, a new high-molecular-weight polysaccharide from Aloe vera with
potent immunostimulatory. J. Agric. Food Chem., v. 49, p. 1030-1034, 2001.

175
PULSE, T. L.; UHLIG, E. A significant improvement in a clinical pilot study
utilizing nutritional supplements, essential fatty acids and stabilized Aloe vera
juice in 29 HIV seropositive, ARC and AIDS patients. Journal of Advancement
in Medicine, v. 3, 209–230, 1990.
RÁCZ, M. L. Adenovírus. In: Trabulsi, L. R. and Alterthum, F. (Eds),
Microbiologia, Editora Atheneu, 4a. edição, p. 587-590, 2005.
RÁCZ, M. L.; MENCK, C. F. M. Propriedades Gerais dos Vírus. In: Trabulsi, L.
R. and Alterthum, F. (Eds), Microbiologia, Editora Atheneu, 4a. edição, p. 509-
514, 2005.
RAPOPORT, E. M.; MOCHALOVA, L. V.; GABIUS, H-J.; ROMANOVA, J.;
BOVIN, N. V. Search for additional influenza vírus to cell interactions.
Glycoconj., v. 23, p. 115-125, 2006.
REED, L. J.; MUENCH, H. A simple method of estimating fifty percents
endpoints. Am. J. Hyg., v. 27, p. 493-497, 1938.
REYNOLDS, T.; DWECK, A. C. Aloe vera leaf gel: a review update. Journal of
Ethnopharmacology, v. 68, p. 3-37, 1999.
RICHMOND, T. A.; SOMERVILLE, C. R. The cellulose synthase superfamily.
Plant Physiology, v.124, p. 495-498, 2000.

176
RIGOUIN, C.; LADRAT, C. D.; SINQUIN, C.; COLLIEC-JOUAULT, S.; DION, M.
Assessment of biochemical methods to detect enzymatic depolymerization of
polymerization of polysaccharides. Carbohydrate Polymers, v.76, p. 279-284,
2009.
RITCHIE, B. W.; NIAGRO, F. D.; LATIMER, K. S.; PRITCHARD, N.;
GREENLACRE, C.; CAMPAGNOLI, R. P.; LUKERT, P. D. Antibody response
and local reactions to adjuvant avian polyomavirus vaccines in psittacine birds.
Journal of the Association of Avian Veterinarians, v. 8, p. 21-26, 1994.
ROBINSON, C. M.; SHARIATI, F.; ZAITSHIK, J.; GILLASPY, A. F.; DYER, D.
W.; CHODOSH, J. Human adenovirus type 19: Genomic and bioinformatics
analysis of a keratoconjunctivitis isolate. Virus Research, v. 139, p. 122-126,
2009.
ROBYT, J. F. Polysaccharides: Energy storage. Encyclopedia of Life
Sciences, p. 1-8, 2001.
RODRIGUES, J. A. G. Atividades biológicas de uma fração polissacarídica
sulfatadas da alga marinha verde Caulerpa cupressoides var. lycopodium
(Vahl) C. Agardh. 159f. 2011. Tese (Doutorado em Biotecnologia) –
Universidade Federal do Ceará, Fortaleza, 2011.
ROTHMAN, A. L.; ENNIS, F. A. Immunopathogenesis of Dengue Hemorrhagic
Fever. Virology, v. 257, p. 1-6, 1999.
ROWE, T. D.; PARKS, L. M. Phytochemical study of Aloe vera leaf. Journal of
the American Pharmaceutical Association, v. 30, p. 262-266, 1941.

177
RUOHOLA, A.; WARIS, M.; ALLANDER, T.; ZIEGLER, T.; HEIKKINEN, T.;
RUUSKANEN, O. Viral etiology of common cold in children. Finland. Emerge.
Infect. Dis., v. 15, p. 344-346, 2009.
RUUSKANEN, O.; LAHTI, E.; JENNINGS, L. C.; MURDOCH, D. R. Viral
pneumonia. Lancet, v. 377, p. 1264-1275, 2011.
SALIH, M. A. M.; EL KHASHAB, H. Y.; HASSAN, H. H.; KENTAB, A. Y.; AL
SUBAEI, S. S.; ZEIDAN, R. M.; AL-NASSER, M. N.; OTHMAN, S. A. A study on
herpes simplex encephalitis in 18 children, including 3 relapses. The open
Pediatric Medicine Journal, v. 3, p. 48-57, 2009.
SANTOS, J. A.; MULLOY, B.; MOURÃO, P. A. S. Structural diversity among
sulfated alpha-L-galactans from ascidians (tunicates). Studies on the species
Ciona intestinalis and Herdmania monus. Eur. J. Biochem., v. 204, p. 669-677,
1992.
SALES, T. C. Metapneumovirus aviário: levantamento soroepidemiológico
e caracterização molecular em criações industriais e de galinhas de
quintal no pólo avícola da Bahia. 97p. 2010. Dissertação (Mestrado em
Saúde Animal). Universidade Federal da Bahia, Salvador, Bahia, 2010.
SAOO, K., MIKI, H., OHMORI, M., WINTERS, W. D. Antiviral activity of Aloe
extracts against cytomegalovirus. Phytotherapy Research, v. 10, p. 348–358,
1996.

178
SAVALIA, C. V.; SINDHI, S. H. Threat and strategic controlo f dengue infection
in man. Veterinary World, v. 1(5), p. 155-156, 2008.
SCHEIBLE, W. R.; PAULY, M. Glycosyltransferases and cell wall biosynthesis:
novel players and insights. Curr. Opin. Plant Biology, v. 7, p. 285-295, 2004.
SCHMITT, A.; GUICHARD, J.; MASSÉ, J-M.; DEBILI, N.; CRAMER, E. M. Of
mice and men: comparison of the ultrastructure of megakaryocytes and
platelets. Experimental Hematology, v. 29, p. 1295-1302, 2001.
ŠEDÝ, J. R.; SPEAR, P. G.; WARE, C. F. Cross-regulation between
herpesviruses and the TNF superfamily members. Nature Reviews –
Immunology, v.8, p. 861-873, 2008.
SERRANO, M.; VALVERDE, J. M.; GUILLÉN, F.; CASTILLO, S.; MARTÍNEZ-
ROMERO, D.; VALERO, D. Use of Aloe vera gel coating presenves the
funcional properties of table grapes. J. Agric. Food Chem., v. 54, p. 3881-86,
2006.
SENNI, K.; PEREIRA, J.; GUENICHE, F.; DELBARRE-LADRAT, C.; SINQUIN,
C.; RATISKOL, J.; GODEAU, G.; FISCHER, A-M.; HELLEY, D.; COLLIEC-
JOUAULT, S. Marine polysaccharides: a source of bioactive molecules for cell
therapy and tissue engineering. Marine Drugs, v. 9, p. 1664-1681, 2011.
SHARMA, J. M.; KARACA, K.; PERTILE, T. Virus-induced immunosuppression
in chickens. Poultry Science, v. 73, p. 1082–1086, 1994.

179
SHUKLA, S. Y.; SING, Y. K.; SHUKLA, D. Role of nectin-1, HVEM, and PILR-á
in HSV-2 entry into human retinal pigment epithelial cells. Invest. Ophyt. &
Visual Sciec., v. 50 (6), p. 2878-2887, 2009.
SPEAR, P. G.; MANOJ, S.; YOON, M.; JOGGER, C. R.; ZAGO, A.;
MYSCOFSKI, D. Different receptors binding to distinct interfaces on herpes
simplex virus gD can trigger events leading to cell fusion and viral entry.
Virology, v. 344, p. 17-24, 2006.
SILVA, M. A. C. R. Isolamento e caracterização dos polissacarídeos das
paredes celulares da azeitona. 1993. Tese (Doutorado em Química – Área de
Concentração em Bioquímica) – Universidade de Aveiro, 1993.
SINGH, R. B.; SHELLEY. Polysaccharide structure of degraded glucomannan
from Abrus precatorius Linn. seeds. Journal of Environmental Biology, v.
38(2), p. 461-464, 2007.
STOCK, R.; HARSTE, G.; MADISCH, I.; HEIM, A. A rapid quantitative PCR-
based assay human adenoviruses demonstrates type specific differences in
ribavirin activity. Antiviral Research, v. 72, p. 34-41, 2006.
STREISSLE, G.; SCHWOBEL, W.; HEWLETT, G. Evaluation of antiviral
compounds in cell cultures with acute or persistent virus infection. In: Triagem
antiviral de extratos vegetais: Funcionamento biomonitorado de Ilex
paraguariensis A. St. Hil. Aquifoliaceae (erva-mate). 116p. 2006.
Dissertação (Mestre em Farmácia), Universidade Federal de Santa Catarina,
Florianópolis, 2006.

180
STRICKLAND, F. M.; PELLEY, R. P. Plant saccharides and the prevention of
sun-induced skin cancer. In: Renolds, T. (Ed.). Aloes – The genus Aloe, CRC
Press LLC, New York, 46p., 2004.
SU, C. M.; LIAO, C. L.; LEE, Y. L.; LIN, Y. L. Highly sulfated forms of heparina
sulfate are involved in japanese encephalitis vírus infection. Virology, v. 286, p.
206-215, 2001.
SUNG, C. K. The history of Aloe. In: Park, Y. I. e Lee, S. K. eds, New
Perspective on Aloe, Springer, Nova York, pp. 7-17, 2006.
SYED, T., AFZAL, M., ASHFAX, A., HOLT, A., ALI, A., AHMAD, S.
Management of genital herpes in men with 0.5% Aloe vera extract in a
hydrophilic cream: a placebo-controlled double blind study. Journal of
Dermatological Treatment, v. 8, p. 99-102, 1997.
SYDISKIS, R. J.; OWEN, D. G.; LOHR, J. L.; ROSLER, K. H.; BLOMSTER, R.
N. Inactivation of enveloped viruses by anthraquinones extracted from plants.
Antimicrobial Agents and Chemotherapy, v. 35, p. 2463-2466, 1991.
TALARICO, L. B. e DAMONTE, E. B. Interference in dengue virus adsorption
and uncoating by carrageenans. Virology, v. 363, p. 473-85, 2007.
TALARICO, L. B.; PUJOL, C. A.; ZIBETTI, R. G. M.; FARÍA, P. C. S.; NOSEDA,
M. D.; DUARTE, M. E. R.; DAMONTE, E. B. The antiviral activity of sulfated
polysaccharides against dengue vírus is dependente on vírus serotype and host
cell. Antiviral Research, v. 66, p. 103-110, 2005.

181
TALYSHINSKY, M. M.; SOUPRUN, Y. Y.; HULEINEL, M. M. Anti-viral activity of
red microalgal polysaccharides against retroviruses. Cancer CellInternational,
v. 2, p. 2-8, 2002.
TANAKA, L. Y. A.; OLIVEIRA, A. J. B.; GONÇALVES, J. E.; CIPRIANI, T. R.;
SOUZA, L. M.; MARQUES, M. C. A.; WERNER, M. F. P.; BAGGIO, C. H.;
GORIN, P. A. J.; SASSAKI, G. L.; IACOMINI, M. An arabinogalactan with anti-
ulcer protective effects isolated from Cereus peruvianus. Carbohydrate
Polymers, v. 82, p. 714-721, 2010.
T’HART, L.A.; VAN DEN BERG, A. J. J.; KUIS, L., VAN DIJK, H.; LABADIE, R.
P. An anticomplementary polysaccharide with immunological adjuvant activity
from the leaf parenchyma gel of Aloe vera. Planta Medica, v. 55, p. 509-512,
1989.
THAMMAWAT, S.; SADLON, T. A.; HALLSWORTH, P. G.; GORDON, D. L.
Role of cellular glycosaminoglycans and charged regions of viral G protein in
human metapneumovirus infection. Journal of Virology, v. 82 (23), p. 11767-
11774, 2008.
TISCHER, P. C. S. F.; TALARICO, L. B.; NOSEDA, M. D.; GUIMARÃES, S. M.
P. B.; DAMONTE, E. B.; DUARTE, M. E. R. Chemical structure and antiviral
activity of carragenans from Meristiella gelidium against herpes simplex and
dengue virus. Carbohydrate Polymers, v. 63, p. 459-465, 2006.

182
TIWARI, V,; O DONELL, C. D.; OH, M. J.; NAGY, T. V.; SHUKLA, D. A role for
3-O-Sulfotransferase isoform-4 in assisting HSV-1 entry spread. Biochem.
Bioph. Res. Co., v. 338, p. 930-937, 2005.
TIZARD, I. R.; RAMAMOORTHY. Aloes and the immune system. In: Renolds,
T. (Ed.). Aloes – The genus Aloe, CRC Press LLC, New York, 22 pp., 2004.
TOLLEFSON, S. J.; COX, R. G.; WILLIAMS, J. V. Studies of culture conditions
and environmental stability of human metapneumovirus. Virus Research, v.
151, p. 54-59, 2010.
TONOMURA, Y.; KATAOKA, H.; YATA, N.; KAWAHARA, M.; OKUCHI, K.;
UENO, S. A successfully treated case of herpes simplex encephalitis
complicated by subarachnoid bleeding: a case report. Journal of Medical Case
Reports, v. 310, p.1-4, 2010.
UCKUN, F. M.; PENDERGRASS, S.; QAZI, S.; SAMUEL, P.
VENKATACHALAM, T. K. Phenyl phosphoramide derivatives of stavudine as
anti-HIV agents with potent and selective in-vitro antiviral activity against
Adenovirus. European Journal of Medicinal Chemistry, v. 39, p. 225-234,
2004.
VAN DAMME, E. J. M.; PEUMANS, W. J.; PUSXTAI, A.; BARDOCZ, S.
Handbook of Plant lectin: Properties and Biochemical applications. John
Willey and Son, Chichester, UK, 452 pp., 1998.
VAN DE VELDE, F.; KNUTSEN, S. H.; USOV, A. I.; ROLLEMA, H. S.;
CEREZO, A. S. 1H and 13C high resolution NMR spectroscopy of carragenans:

183
applications in research and industry. Trends in Food Science & Technology,
v. 13, n. 3, p. 73-92, 2002.
VAN DEN HOOGEN, B. G.; DE JONG, J. C.; GROEN, J.; KUIKEN, T.; DE
GROOT, R.; FOUCHIER, R. A.; OSTERHAUS, A. D. A newly discovered
human pneumovirus isolated from young children with respiratory tract disease.
Nat. Med., v.7, p. 719-724, 2001.
VAN DEN HOOGEN, B. G.; VAN DOORNUM, G. J.; FOCKENS, J. C.;
CORNELISSEN, J. J.; BEYER, W. E.; GROOT, R. R.; OSTERHAUS, A. D.;
FOUCHIER, R. A. Prevalence and clinical symptoms of human
metapneumovirus infection in hospitalized patients. J. Infect. Dis., v. 188, p.
1571-1572, 2003.
VIEIRA, R. P.; MULLOY, B.; MOURÃO, P. A. Structure of a fucose-branched
chondroitin sulfate from sea cucumber. Evidence for the presence of 3-O-sulfo-
â-D-glucuronosyl residues. J. Biol. Chem., v. 266, p. 13530-13536, 1991.
VILLARREAL, E. Current and potential therapies for the treatment of herpes
simplex infections. Prog Drug Res, v. 60, p. 263-307, 2003.
VILELA-SILVA, A. C. E. S.; CASTRO, M. O.; VALENTE, A. P.; BIERMANN, C.
H.; MOURÃO, P. A. S. Sulfated fucans from the egg jellies of the closely related
sea urchins Strongylocentrotus droebachiensis and Strongylocentrotus pallidus
ensure species-specific fertilization. J. Biol. Chem., v. 277, p. 379-387, 2002.
VINE, A. K. Recent advances in haemostasis and thrombosis. Retina, v. 29 (1),
p. 1-7, 2009.

184
VOLPI, N.; MACCARI, F. Characterization of a low-sulfated chondroitin sulfate
isolated from the hemolymph of the freshwater snail Planorbarius corneus. Mar
Biol, v. 152, p.1003-1007, 2007.
VOLPI, N.; MACCARI, F., TITZE, J. Simultaneous detection of submicrogram
quantities of hyaluronic acid and dermatan sulfate on agarose-gel by sequential
staining with toluidine blue and Stains-All. Journal of Chromatography B, v.
820, p. 131-135, 2005.
WALLER, T. A.; PELLEY, R. P.; STRICKLAND, F. M. Industrial processing and
quality control of Aloe barbadensis (Aloe vera) gel. In: Renolds, T. (Ed.). Aloes
– The genus Aloe, CRC Press LLC, New York, 2004.
WHITE, D. O.; FENNER, F. J. Medical Virology. Academic Press, New York,
603 p., 1994.
WHITEHORN, J.; SIMMONS, C. P. The pathogenesis of dengue. Vaccine, v.
29, p. 7221-7228, 2011.
WILLATS, W. G. T.; KNOX, P.; MIKKELSEN, J. D. Pectin: new insights into an
old polymer are starting to gel. Trends in Food Science & Tecnology, v. 17, p.
97-104, 2006.
WINTERS, D.; BENAVIDES, R.; CLOUSE, W. J. Effects of Aloe extracts on
human normal and tumor cells in vitro. Eco. Bot., v. 35, p. 89-95, 1981.

185
WU, J. H.; XU, C.; SHAN, C. Y.; TAN, R. X. Antioxidant properties and PC12
cell protective effects of APS-1 a polysaccharide from Aloe vera var. chinensis.
Life Sciences, v. 78, p. 622-630, 2006.
WYDE, P. R.; CHETTY, S. N.; JEWELL, A. M.; BOIVIN, G.; PIEDRA, P. A.
Comparison of the inhibition of human metapneumovirus and respiratory
syncytial virus by ribavirin and immune serum globulin in vitro. Antiviral
Research, v. 60, p. 51-59, 2003.
WYDE, P. R.; CHETTY, S. N.; JEWELL, A. M.; SCHOONOVER, S. L.; PIEDRA,
P. A. Development of a cotton rat-human metapneumovirus (hMPV) model for
identifying and evaluating potential hMPV antivirals and vaccines. Antiviral
Research, v. 66, p. 57-66, 2005.
WYDE, P. R.; MOYLETT, E. H.; CHETTY, S. N.; JEWELL, A.; BOWLIN, T. L.;
PIEDRA, P. A. Comparison of the inhibition of human metapneumovirus and
respiratory syncytial virus by NMSO3 in tissue culture assays. Antiviral
Research, v. 63, p. 51-59, 2004.
XEPAPADAKI, P.; PSARRAS, S.; BOSSIOS, A.; TSOLIA, M.; GOURGIOTIS,
D.; LIAPI-ADAMIDOU, G.; CONSTANTOPOULOS, A. G.; KAFETZIS, D.;
PAPADOPOULOS, N. G. Human metapneumovirus as a causative agent of
acute bronchiolitis in infants. Journal of Clinical Virology, v. 30, p. 267-270,
2004.
YANG, Z.; HUTTUNEN, E.; STAAF, M.; WIDMALM, G.; TENHU, H. Separation,
purification and characterisation of extracellular polysaccharides produced by
slime-forming Lactococcus lactis ssp. cremoris strains. International Dairy
Journal, v.9, p. 631–638, 1999.

186
YAO, H.; CHEN, Y.; LI, S.; HUANG, L.; CHEN, W.; LIN, X. Promotion
proliferation effect of a polysaccharide from Aloe barbadensis Miller on human
fibroblasts in vitro. International Journal of Biological Macromolecules, v.
45, p.152-156, 2009.
YOUNG & THOMPSON. Willem Van Dijk. Annelize F. Goedbloed. Floris J. R.
Koumans. Negatively charged polysaccharide derivable from Aloe vera. US
2005/0019433 – A1, 27 de Janeiro de 2005.
YU, C-M.; LI, R.-P.; CHEN, X.; LIU, P.; ZHAO, X-D. Replication and
pathogenicity of attenuated human metapneumovirus F mutants in severe
combined immunodeficiency mice. Vaccine, v. 30, p. 231-236, 2012.
ZANDI, K.; ZADEH, M. A.; SARTAVI, K.; RASTIAN, Z. Antiviral activity of Aloe
vera against herpes simplex virus type 2: An in vitro study. African Journal of
Biotechnology, v. 6 (15), p. 1770-1773, 2007.
ZIBETTI, R. G. M. Determinação da estrutura química de polissacarídeos
isolados de algas pertencentes à ordem Halymeniales (Rhodophyta). 199f.
2005. Tese (Doutorado em Ciências) – Universidade Federal do Paraná,
Curitiba, 2005.
ZHU, W., CHIU, L. C. M., OOI, V. E. C., CHAN, P. K. S., ANG Jr., P. O. Antiviral
property and mechanisms of a sulphated polysaccharide from the brown alga
Sargassum patens against Herpes simplex virus type 1. Phytomedicine, v. 13,
p. 695-701, 2006.