university of groningen phenylketonuria mazzola, priscila · diaz jacques, melaine terra, giovana...
TRANSCRIPT

University of Groningen
PhenylketonuriaMazzola, Priscila
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2016
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Mazzola, P. (2016). Phenylketonuria: From body to brain [Groningen]: Rijksuniversiteit Groningen
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 01-12-2018

111
Chapter 6 Regular exercise prevents catecholamine and behavioral deficits in hyperphenylalaninemic rats
Priscila Nicolao Mazzola, Marcelo Xavier Cortes, Carlos Eduardo Diaz Jacques, Melaine Terra, Giovana Reche Dalazen, Emerson Andre Casali, Carlos Severo Dutra-Filho
Department of Biochemistry, Institute of Basic Health Sciences, Federal University of Rio Grande do Sul, Porto Alegre, Brazil
To be submitted

Chapter 6
112
Abstract Background: Phenylketonuria is characterized by accumulation of phenylalanine. Hyperphenylalaninemia (HPA) unbalances the availability of the other large neutral amino acids in tissues, thus impairing neurotransmitter and hormone synthesis. Impaired catecholaminergic system has been associated with the poor cognitive outcome found in patients and animal models of phenylketonuria. Regular exercise improves both central and peripheral synthesis of catecholamines. Therefore, this study evaluated the effects of regular exercise on behavior and catecholamine content in adrenals in rats with HPA. Methods: Fifteen-day-old wild-type Wistar rats were assigned to the groups saline (SAL) or HPA, and then to sedentary (Sed) or exercise (Exe) (n=14–20/group). HPA was chemically induced by subcutaneous injections of alpha-methyl phenylalanine and phenylalanine for 17 days, while SAL groups received saline. Exe groups ran on a treadmill for 20 min/day during 14 days. Animals performed the open field task twice, with a 24-h interval between the trials. Open field performance was analyzed to assess locomotor activity, indicative for anxiety-like behavior. After the last day of tests, the rats were sacrificed and adrenals collected for quantification of catecholamine content. Results: All groups showed similar exploratory activity during the first exposure to the open field. In the second open field exposure, HPA-induced rats did not show the expected decrease in locomotor activity, thus showing behavioral deficits possibly related to anxiety-like behavior. Moreover, HPA led to lower adrenal content of catecholamines. Regular exercise in HPA-induced rats improved rearings in the second open field exposure, and normalized the catecholamine content in the adrenals. Conclusions: Chemically induced HPA led to a deficit in regulating anxiety-like behavior and decreased catecholamine levels in adrenals of rats. Moreover, regular exercise was able to improve the regulation of anxiety-like behavior and restore catecholamine levels in adrenals.

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
113
6
Introduction Phenylketonuria (PKU, 261600) is an inborn error of amino acid
metabolism characterized by low to null activity of phenylalanine hydroxylase, which converts phenylalanine into tyrosine. With the lack of the enzymatic activity, phenylalanine accumulates to toxic levels while endogenous synthesis of tyrosine is compromised. Highly available in the blood, phenylalanine reaches the brain in elevated concentrations, being extremely neurotoxic. In order to prevent from severe and irreversible brain damage, PKU patients have to follow a phenylalanine-restricted diet (1). Despite effective in lowering phenylalanine levels, the dietary treatment is hard to adhere so that early-diagnosed patients still show hyperphenylalaninemia (HPA) (2). Thus, new treatment strategies are still needed in PKU.
HPA impairs the uptake of the other large neutral amino acids (LNAA) such as tyrosine by the tissues. Tyrosine is the precursor of the catecholamines synthetized in the brain and in the adrenals, and which central and peripheral levels are associated with brain functioning (3-5). Consequently, the HPA-related amino acid imbalance has shown to reduce the availability of tyrosine in the brain of PKU patients (6) and PKU mice (7). In addition, high phenylalanine levels may compromise tyrosine uptake by the adrenals since PKU patients have shown low levels of catecholamines in the blood (8, 9). In this way, low availability of tyrosine among other amino acids, followed by disturbed neurotransmitter synthesis have been proposed to underlie the cognitive dysfunction in PKU (10, 11).

Chapter 6
114
Regular exercise can positively affect the synthesis of central and peripheral catecholamines. During exercise, the adrenals enhance the secretion of catecholamines, which act increasing blood supply and energetic sources to the active muscles (12) and the brain (13). Consequently, regular exercise can improve production and secretion of catecholamines by adrenals in healthy condition (14-17). Furthermore, physical training has shown to improve brain synthesis of dopamine in Parkinson’s disease (18, 19), which has possible relations with PKU. Therefore, regular exercise could represent a possible strategy to restore catecholamine levels in adrenals and possibly improve cognition in PKU. However, it is not yet know whether exercise can trigger these adaptations when phenylalanine is high as in PKU. In this way, this study aimed to verify the effects of regular exercise on behavioral performance and adrenal catecholamine content in rats subjected to a chronic model of HPA. Materials and Methods Reagents and equipments
All chemicals were purchased from Sigma/Aldrich (St. Louis, MO, USA). For centrifugation procedures, an Eppendorf 5417R (refrigerated version) and Eppendorf 5403 were used. A spectrophotometer Beckman DU™ 800 (Beckman Coulter, Inc., Fullerton, CA, USA), and a SpectraMax M5/M5 Microplate Reader (Molecular Devices, MDS Analytical Technologies, Sunnyvale, CA,

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
115
6
USA) were used for the measurements. A treadmill specifically for rodents was used (INBRAMED TK 01, Porto Alegre, Brazil). Animals
Fifteen day-old Wistar rats bred in the Department of Biochemistry, UFRGS, Brazil, were used according to sample size calculation with 80% of power and level of significance of 0.05 (MiniTab™). Rats stayed with theirs dams until weaning at 21 days of life, having free access to water and commercial chow (Germani, Porto Alegre, RS, Brazil). Temperature was maintained at 24 ± 1ºC, with a 12:12 h light/dark cycle. The study has been approved by the Local Ethics Committee, and the Principles of Laboratory Animal Care (NIH publication #85-23, revised 1985) were followed throughout the experiments.
The rats were randomly assigned into the groups saline (SAL) and HPA, and then into the subgroups sedentary (Sed) and exercise (Exe) (n=14–20/group). Chemically induced HPA
Rats from the HPA groups received twice-a-day subcutaneous injections of 1.6 µmol/g bw of alpha-methyl phenylalanine (a phenylalanine hydroxylase inhibitor) plus 2.1 µmol/g bw of phenylalanine between 0700-0900, followed by 2.1 µmol/g bw of phenylalanine alone between 1600-1800 (Hagen et al. 2002). The

Chapter 6
116
substances were dissolved in saline solution by heating. SAL groups received equivalent volumes of 0.9% saline solution.
In the first experiment, we measured phenylalanine levels in six 30-day-old sedentary rats subjected to the HPA model or saline solution after two days of the already mentioned injections. Animals were sacrificed one hour after the last injection and plasma and the total brain were collected for amino acid quantification. The HPA rats showed about 22 and four times higher phenylalanine concentrations in the plasma and in the brain, respectively if compared to control animals that received saline solution (Table 1). This data was comparable with findings from the genetic mouse model of phenylketonuria (PAHenu2), which have shown phenylalanine levels in the blood and brain between 19–25 and 7–17 times higher, respectively, than wild-type mice (7, 20, 21).
In the second experiment, we evaluated the effects of regular exercise on behavior and adrenal catecholamines in HPA-induced rats. For that, the HPA treatment lasted 17 days (from 15th to 31st day of life), and the rats were sacrificed 14 h after the last injection, thereby around 13 h after the last running session. Amino acid measurements
The concentration of free amino acid levels in plasma and brain deproteinized supernatants (first experiment) was determined according to Joseph and Marsden (22). The analysis was performed by reversed-phase high performance liquid chromatography system (HPLC) (Shimadzu, Japan) using a C18 column (25 cm × 4.6 mm x 5

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
117
6
µm, Restek - USA). The fluorescent detection was done using orthophtaldialdehyde and mercaptoethanol (fluorescence detector Shimadzu RF-535). The flow rate was 1.4 mL/min in a gradient of the mobile phase (methanol and phosphate buffer: buffer A, 20% methanol; buffer B, 80% methanol), and the duration of each analysis was 50 min. The LNAA: phenylalanine, tryptophan, tyrosine, leucine, isoleucine, histidine, methionine, and valine were quantitatively determined by calculating their chromatographic peak areas related to those obtained for a known standard mixture and to that of the internal standard homocysteic acid.

Chapter 6
118
Table 1 Amino acid concentrations in the plasma and brain of young adult rats subjected to two days of the hyperphenylalaninemia (HPA) model by alpha-methylphenylalanine and phenylalanine versus controls. Results are expressed as mean ± SD (t test, n=3/group). LNAA, large neutral amino acids; NS, not significant. * other than phenylalanine, tryptophan, and tyrosine (i.e. summed levels of leucine, isoleucine, histidine, methionine, and valine). 2-day HPA Control p-value Plasma (µmol/L) Phenylalanine 1489 ± 225 69 ± 12 <0.001 Tryptophan 88 ± 18 180 ± 38 0.009 Tyrosine 491 ± 46 713 ± 156 0.012 Other LNAA* 983 ± 105 1065 ± 100 NS Brain (µmol/L) Phenylalanine 43 ± 10 12 ± 2 0.004 Tryptophan 3.7 ± 1.4 3.7 ± 2.0 NS Tyrosine 26 ± 6 259 ± 64 0.012 Other LNAA* 116 ± 16 167 ± 8 0.008 Exercise protocol
Animals of the Exe groups were habituated to the treadmill apparatus for three days, and then ran for 20 min at 12 m/min for 14 consecutive days (23, 24) between 1700 and 1900. Sed groups were kept in their home cages while the Exe groups were running.

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
119
6
Open field task In the morning of the last exercise training (1 h after the HPA
injection), rats were introduced to the open field task for assessment of anxiety-like behavior (25). Rats were placed at the center of a 120-cm diameter black cylinder and allowed to freely explore it for 3 min. About 24 h later, animals were re-exposed to the task at the same conditions. The cylinder was virtually divided into inner and outer zones using the software Any-Maze® version 4.63, keeping track of the time the animal spent in those zones, distance traveled, number of line crossing, and rearings. Catecholamine content
Total catecholamine content (adrenaline and noradrenaline) in the adrenals was carried out accordingly the trihydroxyindole fluorescence method (Kelner et al. 1985; Scomparin et al. 2009). Both adrenals were removed and homogenized in 350 µl acetic acid 10% and centrifuged at 10,000 Xg for 1 min. Fifty microliters of the supernatant fraction were mixed with 250 µl of 0.5 M phosphate buffer (pH 7.0) and 25 µl of potassium ferricyanate 0.5%, followed by a 20-min incubation. The reaction was stopped with 500 µl of ascorbic acid dissolved in NaOH. A standard curve of adrenaline was used to calculate catecholamine content of the samples in a spectrofluorometer (420 nm excitation and 510 nm emission). Data are expressed as µg of catecholamine per gland.

Chapter 6
120
Statistical analysis Student’s t-test, paired t-test, and two-way ANOVA followed by
Tukey post-hoc test were performed. The Statistical Package for the Social Sciences (SPSS) software was employed. A value of p<0.05 was considered to be statistically significant. Results
In order to evaluate the effects of regular exercise on central and peripheral parameters that are affected in PKU, we conducted a study using forced exercise (treadmill running) in rats chemically subjected to HPA.
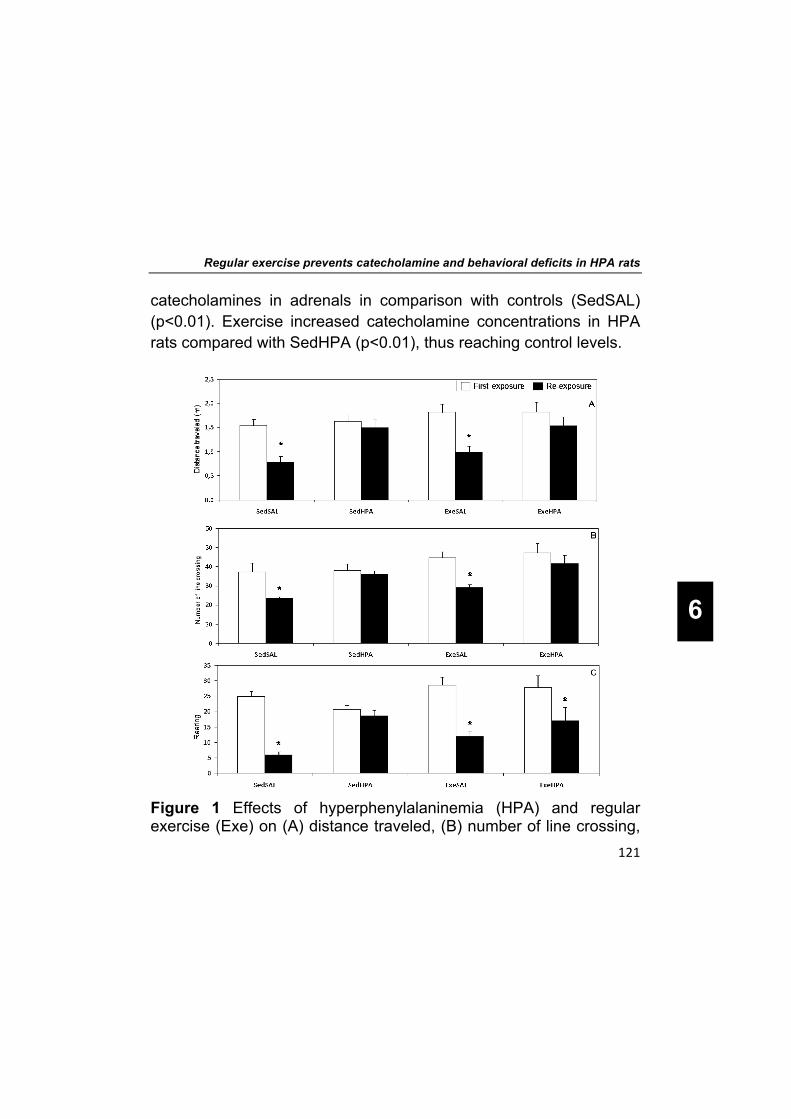
Regarding exploratory behavior in the first time in the open field task, all groups showed similar values for distance traveled, number of line crossing and number of rearings (Figure 1). Only the SAL groups showed lower exploratory behavior between first time and re-exposure to the same task. In SedSAL and ExeSAL the values for distance traveled, the number of line crossings, and the number of rearings between first time and re-exposure were decreased for SedSAL (both groups p<0.001, p<0.005, p<0.001, respectively). The SedHPA group did not show any difference in exploration between the trials. The ExeHPA group showed lower number of rearings in the re-exposure trial (p=0.018), while no differences were found in distance traveled and number of line crossing.
Peripherally, total catecholamine content of adrenals was affected by the factors physical activity and HPA treatment (both p<0.001). As shown in Figure 2, the HPA model led to lower levels of
Regular exercise prevents catecholamine and behavioral deficits in HPA rats
121
6
catecholamines in adrenals in comparison with controls (SedSAL) (p<0.01). Exercise increased catecholamine concentrations in HPA rats compared with SedHPA (p<0.01), thus reaching control levels.
Figure 1 Effects of hyperphenylalaninemia (HPA) and regular exercise (Exe) on (A) distance traveled, (B) number of line crossing,

Chapter 6
122
and (C) rearings in the first time and re-exposure to the open field task. Data are expressed as mean ± SEM (n=6–10/group). * indicates p<0.05 compared with the first time (paired t-test).
Figure 2 Effects of chronic hyperphenylalaninemia (HPA) and regular exercise (Exe) on adrenal catecholamine content in comparison to sedentary (Sed) controls (SAL). Data are expressed as mean ± SEM (n=7/group), * indicates p<0.01 compared with SedSAL; # indicates p<0.01 compared with SedHPA.

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
123
6
Discussion This study investigated the effects of regular exercise on central
and peripheral markers in rats subjected to HPA. Our findings showed that the forced exercise training was able to improve some behavioral and hormone parameters that were impaired by HPA. Moreover, HPA-induced rats were more responsive to exercise than the controls, since any parameter was affected by the same exercise protocol in control animals.
The rats subjected to HPA showed normal locomotor activity but disturbed exploratory behavior and low adrenal catecholamine content. Therefore, HPA possibly triggered anxiety-like behavior, which is compatible with findings in PKU mice (26). While the control rats reduced exploratory behavior in the re-exposure to the same apparatus, the HPA-induced rats showed similar exploratory behavior in the first- and re-exposure to the same open field arena. The similar activity between trials seen in the HPA rats may represent behavioral deficits possibly related to anxiety-like behavior (25). Anxiety is generally associated to aggressive behavior and may impair cognition (27). Anxiety-like behavior can be triggered by oxidative stress (28) and disturbances in the dopaminergic and serotonergic systems (29, 30). HPA is associated to oxidative stress (31) as well as disturbs amino acid concentrations in the brain (26). Furthermore, the low availability of tyrosine and consequently low brain catecholamine synthesis can importantly impair cognition (32). In addition, although in humans, peripheral catecholamine levels have shown to affect mood and cognition (4, 5). In this way, the

Chapter 6
124
reduced levels of catecholamines in the adrenals may have contributed to the disturbed behavior shown by the HPA-induced rats. Accordingly, PKU mice have shown poor cognitive function such as flawed latent- (33), iconic-, and short-term memory, and defective habit learning (34, 35). In this way, our findings corroborate previous studies where HPA led to impairments on behavior, also hypothesizing the contribution of peripheral catecholamines to the brain status.
Regular exercise was able to improve one behavioral parameter and to increase catecholamine content in the adrenals of HPA-induced rats, thus reaching control levels. Exercise is well known to improve cognition in several populations (36) as well as in animal models of neurodegenerative diseases (37, 38). Furthermore, exercise positively affects mood disorders including anxiety-like behavior (39, 40). Concerning mechanisms, exercise-related adaptations on brain functioning have been associated to enhanced catecholaminergic activity. Each exercise session markedly increased sympathetic activity (fight-or-flight reaction), which is mediated by catecholamines (41). Consequently, exercise training can strengthen the catecholaminergic system by increasing the activity and expression of tyrosine hydroxylase, which is the rate-limiting enzyme of catecholamine biosynthesis (16, 42, 43). Although we did not measure catecholamine levels in the blood or in the brain, the increased values in adrenal content points out to improvements in catecholamine synthesis. As the adrenals are the main source of circulating catecholamines, exercise-induced improvements in their

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
125
6
content may represent an interesting improvement for central and peripheral status in PKU.
Taken together, we demonstrated that regular exercise improved anxiety-like behavior besides preventing HPA-related disturbances in peripheral catecholamines. In this way, exercise-related adaptations in central and peripheral organs can be of interest in PKU. Acknowledgements
This work was supported by the research grants from “Conselho Nacional de Desenvolvimento Científico e Tecnológico” (CNPq), “Programa de Núcleos de Excelência” (PRONEX), “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior” (CAPES), “Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul” (FAPERGS), and “Rede Instituto Brasileiro de Neurociência” (FINEP) (IBN-Net #01.06.0842-00).

Chapter 6
126
References 1. Blau N, van Spronsen FJ, Levy HL (2010) Phenylketonuria. Lancet, 376(9750):
1417-27
2. Vilaseca MA, Lambruschini N, Gomez-Lopez L, Gutierrez A, Fuste E, Gassio R, Artuch R, Campistol J (2010) Quality of dietary control in phenylketonuric patients and its relationship with general intelligence. Nutr Hosp, 25(1): 60-6
3. Fernstrom JD, Fernstrom MH (2007) Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J Nutr, 137(6 Suppl 1): 1539S-1547S; discussion 1548S
4. Rief W, Mills PJ, Ancoli-Israel S, Ziegler MG, Pung MA, Dimsdale JE (2010) Overnight changes of immune parameters and catecholamines are associated with mood and stress. Psychosom Med, 72(8): 755-62
5. Brummett BH, Boyle SH, Kuhn CM, Siegler IC, Williams RB (2009) Positive affect is associated with cardiovascular reactivity, norepinephrine level, and morning rise in salivary cortisol. Psychophysiology, 46(4): 862-9
6. de Groot MJ, Hoeksma M, Reijngoud DJ, de Valk HW, Paans AM, Sauer PJ, van Spronsen FJ (2013) Phenylketonuria: reduced tyrosine brain influx relates to reduced cerebral protein synthesis. Orphanet J Rare Dis, 8: 133
7. Pascucci T, Giacovazzo G, Andolina D, Conversi D, Cruciani F, Cabib S, Puglisi-Allegra S (2012) In vivo catecholaminergic metabolism in the medial prefrontal cortex of ENU2 mice: an investigation of the cortical dopamine deficit in phenylketonuria. J Inherit Metab Dis, 35(6): 1001-9
8. Schulpis KH, Papassotiriou I, Tsakiris S, Vounatsou M, Chrousos GP (2005) Increased plasma adiponectin concentrations in poorly controlled patients with phenylketonuria normalize with a strict diet: evidence for catecholamine-mediated adiponectin regulation and a complex effect of phenylketonuria diet on atherogenesis risk factors. Metabolism, 54(10): 1350-5
9. Schulpis KH, Papassotiriou I, Vounatsou M, Karikas GA, Tsakiris S, Chrousos GP (2004) Morning preprandial plasma ghrelin and catecholamine concentrations in patients with phenylketonuria and normal controls: evidence

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
127
6
for catecholamine-mediated ghrelin regulation. J Clin Endocrinol Metab, 89(8): 3983-7
10. Surtees R, Blau N (2000) The neurochemistry of phenylketonuria. Eur J Pediatr, 159 Suppl 2: S109-13
11. van Spronsen FJ, Hoeksma M, Reijngoud DJ (2009) Brain dysfunction in phenylketonuria: is phenylalanine toxicity the only possible cause? J Inherit Metab Dis, 32(1): 46-51
12. Smilios I, Pilianidis T, Karamouzis M, Tokmakidis SP (2003) Hormonal responses after various resistance exercise protocols. Med Sci Sports Exerc, 35(4): 644-54
13. Kitaoka R, Fujikawa T, Miyaki T, Matsumura S, Fushiki T, Inoue K (2010) Increased noradrenergic activity in the ventromedial hypothalamus during treadmill running in rats. J Nutr Sci Vitaminol (Tokyo), 56(3): 185-90
14. Kjaer M, Galbo H (1988) Effect of physical training on the capacity to secrete epinephrine. J Appl Physiol, 64(1): 11-6
15. Levenson CW, Moore JB (1998) Response of rat adrenal neuropeptide Y and tyrosine hydroxylase mRNA to acute stress is enhanced by long-term voluntary exercise. Neurosci Lett, 242(3): 177-9
16. Erdem SR, Demirel HA, Broxson CS, Nankova BB, Sabban EL, Tumer N (2002) Effect of exercise on mRNA expression of select adrenal medullary catecholamine biosynthetic enzymes. J Appl Physiol, 93(2): 463-8
17. Desgorces FD, Chennaoui M, Gomez-Merino D, Drogou C, Bonneau D, Guezennec CY (2004) Leptin, catecholamines and free fatty acids related to reduced recovery delays after training. Eur J Appl Physiol, 93(1-2): 153-8
18. Sasco AJ, Paffenbarger RS, Jr., Gendre I, Wing AL (1992) The role of physical exercise in the occurrence of Parkinson's disease. Arch Neurol, 49(4): 360-5
19. Dibble LE, Addison O, Papa E (2009) The effects of exercise on balance in persons with Parkinson's disease: a systematic review across the disability spectrum. J Neurol Phys Ther, 33(1): 14-26

Chapter 6
128
20. Smith CB, Kang J (2000) Cerebral protein synthesis in a genetic mouse model of phenylketonuria. Proc Natl Acad Sci U S A, 97(20): 11014-9
21. Mazzola PN, Bruinenberg V, Anjema K, van Vliet D, Dutra-Filho CS, van Spronsen FJ, van der Zee EA (2015) Voluntary Exercise Prevents Oxidative Stress in the Brain of Phenylketonuria Mice. JIMD Rep,
22. Joseph MH, Marsden CA, Amino acids and small peptides., in HPLC of small peptides., C.K. LIM, Editor. 1986, Oxford. p. 13-27.
23. Scopel D, Fochesatto C, Cimarosti H, Rabbo M, Bello-Klein A, Salbego C, Netto CA, Siqueira IR (2006) Exercise intensity influences cell injury in rat hippocampal slices exposed to oxygen and glucose deprivation. Brain Res Bull, 71(1-3): 155-9
24. Mazzola PN, Terra M, Rosa AP, Mescka CP, Moraes TB, Piccoli B, Jacques CE, Dalazen G, Cortes MX, Coelho J, Dutra-Filho CS (2011) Regular exercise prevents oxidative stress in the brain of hyperphenylalaninemic rats. Metab Brain Dis, 26(4): 291-7
25. Prut L, Belzung C (2003) The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur J Pharmacol, 463(1-3): 3-33
26. Martynyuk AE, van Spronsen FJ, Van der Zee EA (2010) Animal models of brain dysfunction in phenylketonuria. Mol Genet Metab, 99 Suppl 1: S100-5
27. Vytal K, Cornwell B, Arkin N, Grillon C (2012) Describing the interplay between anxiety and cognition: from impaired performance under low cognitive load to reduced anxiety under high load. Psychophysiology, 49(6): 842-52
28. Bouayed J, Rammal H, Soulimani R (2009) Oxidative stress and anxiety: relationship and cellular pathways. Oxid Med Cell Longev, 2(2): 63-7
29. Rylands AJ, Hinz R, Jones M, Holmes SE, Feldmann M, Brown G, McMahon AW, Talbot PS (2012) Pre- and postsynaptic serotonergic differences in males with extreme levels of impulsive aggression without callous unemotional traits: a positron emission tomography study using (11)C-DASB and (11)C-MDL100907. Biol Psychiatry, 72(12): 1004-11

Regular exercise prevents catecholamine and behavioral deficits in HPA rats
129
6
30. Patki G, Atrooz F, Alkadhi I, Solanki N, Salim S (2015) High aggression in rats is associated with elevated stress, anxiety-like behavior, and altered catecholamine content in the brain. Neurosci Lett, 584: 308-13
31. Hagen MEK, Pederzolli CD, Sgaravatti AM, Bridi R, Wajner M, Wannmacher CM, Wyse AT, Dutra-Filho CS (2002) Experimental hyperphenylalaninemia provokes oxidative stress in rat brain. Biochim Biophys Acta, 1586(3): 344-52
32. Landvogt C, Mengel E, Bartenstein P, Buchholz HG, Schreckenberger M, Siessmeier T, Scheurich A, Feldmann R, Weglage J, Cumming P, Zepp F, Ullrich K (2008) Reduced cerebral fluoro-L-dopamine uptake in adult patients suffering from phenylketonuria. J Cereb Blood Flow Metab, 28(4): 824-31
33. Tang YP, Shimizu E, Dube GR, Rampon C, Kerchner GA, Zhuo M, Liu G, Tsien JZ (1999) Genetic enhancement of learning and memory in mice. Nature, 401(6748): 63-9
34. Zagreda L, Goodman J, Druin DP, McDonald D, Diamond A (1999) Cognitive deficits in a genetic mouse model of the most common biochemical cause of human mental retardation. J Neurosci, 19(14): 6175-82
35. Sarkissian CN, Boulais DM, McDonald JD, Scriver CR (2000) A heteroallelic mutant mouse model: A new orthologue for human hyperphenylalaninemia. Mol Genet Metab, 69(3): 188-94
36. Szuhany KL, Bugatti M, Otto MW (2015) A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J Psychiatr Res, 60: 56-64
37. Harrison DJ, Busse M, Openshaw R, Rosser AE, Dunnett SB, Brooks SP (2013) Exercise attenuates neuropathology and has greater benefit on cognitive than motor deficits in the R6/1 Huntington's disease mouse model. Exp Neurol, 248: 457-69
38. Souza LC, Filho CB, Goes AT, Fabbro LD, de Gomes MG, Savegnago L, Oliveira MS, Jesse CR (2013) Neuroprotective effect of physical exercise in a mouse model of Alzheimer's disease induced by beta-amyloid(1)(-)(4)(0) peptide. Neurotox Res, 24(2): 148-63

Chapter 6
130
39. Salim S, Sarraj N, Taneja M, Saha K, Tejada-Simon MV, Chugh G (2010) Moderate treadmill exercise prevents oxidative stress-induced anxiety-like behavior in rats. Behav Brain Res, 208(2): 545-52
40. DeBoer LB, Powers MB, Utschig AC, Otto MW, Smits JA (2012) Exploring exercise as an avenue for the treatment of anxiety disorders. Expert Rev Neurother, 12(8): 1011-22
41. de Diego AM, Gandia L, Garcia AG (2008) A physiological view of the central and peripheral mechanisms that regulate the release of catecholamines at the adrenal medulla. Acta Physiol (Oxf), 192(2): 287-301
42. Tumer N, Demirel HA, Serova L, Sabban EL, Broxson CS, Powers SK (2001) Gene expression of catecholamine biosynthetic enzymes following exercise: modulation by age. Neuroscience, 103(3): 703-11
43. Kjaer M (1998) Adrenal medulla and exercise training. Eur J Appl Physiol Occup Physiol, 77(3): 195-9