uva-dare (digital academic repository) bio-systematics … · famah sourassou, n...
TRANSCRIPT

UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl)
UvA-DARE (Digital Academic Repository)
Bio-systematics of predatory mites used for control of the coconut mite
Famah Sourassou, N.
Link to publication
Citation for published version (APA):Famah Sourassou, N. (2012). Bio-systematics of predatory mites used for control of the coconut mite
General rightsIt is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s),other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons).
Disclaimer/Complaints regulationsIf you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, statingyour reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Askthe Library: http://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam,The Netherlands. You will be contacted as soon as possible.
Download date: 28 Jul 2018

Bio-systematics of predatory mites used for control of the coconut mite
Nazer Fam
ah SourassouBio-system
atics of predatory mites used for control of the coconut m
ite
Nazer Famah Sourassou

Bio-systematics of predatory mites used for control of the coconut mite
Nazer-voorwerk.qxd 14-9-2012 10:34 Page 1

Famah Sourassou, N 2012. Bio-systematics of predatory mites used for control of the coconutmite
PhD thesis, University of Amsterdam, The Netherlands
The work presented in this thesis was funded by a grant from the Netherlands Organ-isation for Scientific Research (NWO; WOTRO project ‘Molecular and morphometriccharacterization of geographic populations of two species of predatory mites associ-ated with the coconut mite in Brazil, Benin and Sri Lanka’). Research was conductedpartly at the Institute for Biodiversity and Ecosystem Dynamics (IBED), University ofAmsterdam.
Publication of this thesis was financially supported by Koppert Biological Systems(www.koppert.nl)
ISBN: 978 94 91407 07 9
Cover design: Jan van Arkel (IBED)Lay-out: Jan Bruin (www.bred.nl)
Nazer-voorwerk.qxd 14-9-2012 10:34 Page 2

Bio-systematics of predatory mites used for control of the coconut mite
ACADEMISCH PROEFSCHRIFT
ter verkrijging van de graad van doctoraan de Universiteit van Amsterdamop gezag van de Rector Magnificus
prof. dr. D.C. van den Boomten overstaan van een door het college voor promoties
ingestelde commissie, in het openbaar te verdedigen in de Agnietenkapel
op vrijdag 26 oktober 2012, te 10:00 uur
door
Nazer Famah Sourassou
geboren te Paratao, Togo
Nazer-voorwerk.qxd 14-9-2012 10:34 Page 3

Promotiecommissie
Promotor: Prof. dr. M.W. Sabelis
Co-promotor:Dr. J.A.J. Breeuwer
Overige leden:Prof. dr. J.J.M. van Alphen Dr. R. HannaDr. A.RM. Janssen Prof. dr. S.B.J. MenkenDr. M. NavajasProf. dr. M.E. SchranzProf. dr. P.H. van Tienderen
Faculteit der Natuurwetenschappen, Wiskunde en Informatica
Nazer-voorwerk.qxd 14-9-2012 10:34 Page 4

Contents
6 Author addresses
7 1 – General introduction
15 2 – Morphological variation and reproductive incompatibility of three coconut-mite-associated populations of predatory mites identified as Neoseiuluspaspalivorus (Acari: Phytoseiidae)
N. Famah Sourassou, R. Hanna, I. Zannou, G.J. de Moraes, K. Negloh & M.W. Sabelis
31 3 – Endosymbiont Wolbachia and Cardinium bacteria in the predatory miteNeoseiulus paspalivorus
N. Famah Sourassou, R. Hanna, J.A.J. Breeuwer, K. Negloh, G.J. de Moreas & M.W. Sabelis
47 4 – Morphological, molecular and cross-breeding analysis of geographicpopulations of coconut-mite associated predatory mites identified as Neoseiulusbaraki: evidence for cryptic species?
N. Famah Sourassou, R. Hanna, I. Zannou, J.A.J. Breeuwer, G.J. de Moraes & M.W. Sabelis
67 5 – Females as intraguild predators of males in cross-pairing experiments withphytoseiid mites
N. Famah Sourassou, K. Negloh, R. Hanna, J.A.J. Breeuwer & M.W. Sabelis
77 6 – Predatory mites found under the bracts of coconuts and their potential forcontrolling Aceria guerreronis: review and hypotheses
N. Famah Sourassou, R. Hanna, I. Zannou, G.J. de Moraes, F.R. da Silva, K. Negloh, J.A.J. Breeuwer & M.W. Sabelis
93 7 – General discussion
97 Summary101 Samenvatting105 Curriculum vitae106 Acknowledgements
Nazer-voorwerk.qxd 14-9-2012 10:34 Page 5

Author addresses
Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, Science Park 904,1098 XH Amsterdam, The Netherlands
Johannes A.J. BreeuwerNazer Famah SourassouKoffi NeglohMaurice W. SabelisFernando R. da SilvaIgnace Zannou
International Institute of Tropical Agriculture (IITA), 08 BP 0932 Cotonou, Benin, West AfricaNazer Famah SourassouRachid HannaKoffi NeglohIgnace Zannou
Depto Entomologia e Acarologia, Escola Superior de Agricultura. 'Luiz de Queiroz', Universidadede Sao Paulo, Piracicaba-SP, 13418-900, Brazil
Gilberto J. de MoraesFernando R. da Silva
International Institute of Tropical Agriculture (IITA), BP 2008 (Messa), Yaoundé, CameroonRachid Hanna
6
Nazer-voorwerk.qxd 14-9-2012 10:34 Page 6

7
General introduction
Over the past 50 years there has been considerable progress in the use of phytosei-id predators for control of mite pests in agricultural crops (Helle and Sabelis 1985;
Lindquist et al. 1996; Gerson et al. 2003). Essential to the development of these biocon-trol methods is to provide a reliable description of the control agent (Moraes 1987;McMurtry 2010). A description can be obtained from taxonomic studies that are basedon morphological criteria for species identification. However, taxonomic identificationof species alone does not always suffice because populations of the same predatorspecies may largely differ in their capacity to control the pest. A recent example is thefinding that the predatory mite Phytoseiulus longipes from Brazil and Argentina can controlTetranychus evansi on tomato, but P. longipes from Chili and South-Africa cannot (Tixier etal. 2010). These between-population differences in control capacity appear to have agenetic basis. Another example, is the presence of five cryptic species from differenthosts and geographic localities (Euseius tularensis from citrus, E. hibisci from avocado, E.quetzali from deciduous fruits and nuts, E. obispensis from avocado in San Luis Obispocounty, E. stipulatus introduced from the Mediterranean region) in what was onceassumed to be a single species (E. stipulatus) (McMurtry and Badii 1985; McMurtry 2010).However, there may also be within-population genetic variation with respect to genetictraits relevant to biological control. For example, it is shown that at a scale of 0.25 m2
soil a population sample of the predatory mite Hypoaspis aculeifer exhibits a genetic poly-morphism in preference for two species of astigmatic mites, Tyrophagus putrescentiae andRhizoglyphus robini, that are pests in crops of lily bulbs (Lesna and Sabelis 1999, 2002).Thus, there is a need to identify and characterize the genetic entities (strain, population,biotype, ecotype, host race, geographic race, subspecies, cryptic species, species) that arerelevant for assessing traits relevant to biological control (McMurtry 2010).
Before deciding on relevant genetic entities it is useful to briefly introduce anddefine some terms because there is still room for confusion in the general literature.First and foremost, is the biological species concept (Mayer 1940; Mallet 1995), whichdefines a species as a group of organisms capable of interbreeding and producing fer-tile offspring of both sexes and separated from other such groups with which inter-breeding does not normally happen. With respect to the Phytoseiidae most species havebeen described solely based on morphological criteria (Chant and McMurtry 2007) andonly some species have been studied for their reproductive isolation. However, not allisolating mechanisms are immediately apparent. For example, the presence of endosym-bionts and their role in incompatibility between populations has long gone unnoticed,but now we know that this mechanism can isolate populations even when geographical-
1Nazer-chap1.qxd 14-9-2012 10:35 Page 7

ly very close to each other (see Hoy and Cave 1988 for an example in phytoseiid mites).Such isolating mechanisms cause reduced gene flow between populations and may giverise to genetic differences and incipient speciation (Telschow et al. 2005). This has ledto the notion of cryptic species (complex of species), which are defined by Stebbins(1950) as ‘population systems which were believed to belong to the same species untilgenetic evidence showed the existence of isolating mechanisms separating them’. Grant(1981) defined cryptic species as ‘good biological species that are virtually indistinguish-able morphologically. Cryptic species have been found in the Phytoseiidae based onmorphological studies combined with crossbreeding studies (Muma and Denmark 1976;Mahr and McMurtry 1979; McMurtry et al. 1985; Congdon and McMurtry 1985), withbehavioural studies (Beard 1999) and with molecular-genetic studies (Tixier et al. 2003,2004, 2006, 2008). Whether these cryptic species are also sibling species, requires indepth phylogenetic analysis using a variety of morphological, behavioural and genetictraits. Such an analysis is still lacking with respect to the Phytoseiidae. Instead, classifi-cations in the Phytoseiidae are purely based on morphological traits (Chant andMcMurtry 2005).
Geographically isolated populations may or may not be reproductively isolated.They are often referred to as geographic races, subspecies or semispecies, but their sta-tus as a species is unclear until it is known whether they are reproductively isolated whenthe races become sympatric (Diehl and Bush 1984). The same applies to terms like hostraces, ecotypes, biotypes and strains. These terms are used to indicate that populationsdiffer conspicuously in some genetically determined traits. Such differences are thoughtto result from reduced gene flow between populations and thus at least partial geneticisolation. However, the extent to which the partial isolation operates is not alwaysknown. Because gene flow is hard to assess and there is a practical need to distinguishpopulations with different traits (Clark and Walter 1995), the terms referred to abovewill persist, but it should be clear that the categories blend into one another (Diehl andBusch 1984).
For the purpose of biological control it is surely interesting to know whether pop-ulations differ in a genetic trait relevant to this objective. However, releasing such astrain, ecotype, biotype or geographic race in another area with a morphologically sim-ilar population is problematic if reproductive isolation is not ensured. This is why Ifocus on reproductive incompatibility between geographic populations and assess theirtraits as far as I think they are relevant to biological control.
Hybridization studies have been conducted between populations or strains of phy-toseiid species to test for reproductive compatibility. Such studies challenge taxonomiststo examine specimens more critically for morphological characters. This is illustrated bya case study on Typhlodromus exhilaratus (Ragusa), which appeared to harbour two crypticspecies, T. exhilaratus and T. phialatus (Athias-Henriot) (Tixier et al. 2006a). In other stud-
8
CHAPTER 1
Nazer-chap1.qxd 14-9-2012 10:35 Page 8

ies, synonymy was considered between phytoseiid mites previously identified as differentspecies but showing full reproductive compatibility, for instance synonymy betweenAmblyseius swirskii (Athias-Henriot) and A. rykei (Pritchard & Baker) (Zannou et al. 2011),or between Cydnodromus idaeus (Denmark & Muma) and C. picanus (Ragusa) (Tixier et al.2011).
Reproductive barriers may arise from prezygotic isolation mechanisms (e.g., matechoice) or or postzygotic mechanisms. Postzygotic incompatibility can result fromgenetic incompatibility (Navajas et al. 1999; Uesugi et al. 2003; Gotoh et al. 2005b; Prykeand Griffith 2008; Koevoets and Beukeboom 2009) or more specifically from endosym-bionts, such as Wolbachia and Cardinium (Laven 1967; O’Neill and Karr 1990; Perrot-Minnot et al. 1996; Breeuwer 1997; Gotoh et al. 2007) as well as their interactions withnuclear genes (Gotoh et al. 1995). Endosymbiont-mediated reproductive incompatibili-ty, so-called ‘cytoplasmic incompatibility’ is a sterility phenomenon that has been firstreported between strains of insects (Laven 1959, 1967), and is now known to occur ina wide range of taxa. Two basic forms of cytoplasmic incompatibility are known: uni-directional (nonreciprocal) cytoplasmic incompatibility and bidirectional cytoplasmicincompatibility. Unidirectional (nonreciprocal) cytoplasmic incompatibility occurs incrosses involving infected and uninfected populations and results in normal progeny inone direction but in few viable hybrids or no progeny in the other. Bidirectional cyto-plasmic incompatibility is observed in crosses between populations infected with differ-ent strains of endosymbionts (Perrot-Minnot et al. 1996). In the Phytoseiidae, unidirec-tional incompatibilities are the most common, and are associated with shrivelled eggs, alow number of eggs, low survival of immature stages and reduced fecundity in F1 indi-viduals (if present) (Johanowicz and Hoy 1998; Norhona and Moraes 2004). Althoughcompared to insects and isopods little is done to assess the cause of the reproductiveincompatibilities in the Phytoseiidae, there is one case study on Metaseiulus occidentalisNesbitt, demonstrating that Wolbachia mediates unidirectional incompatibility in phyto-seiid species (Hess and Hoy 1982; Hoy and Cave 1988; Johanowicz and Hoy 1998). Yet,in the Phytoseiidae, no study has reported Cardinium-mediated cytoplasmic incompati-bility, an endosymbiont well known from insects or other mite taxa (spider mites andfalse spider mites).
To discriminate between genetic entities a comprehensive biosystematicapproach is required (McMurtry et al. 1976; McMurtry 1980). Over the past decade,molecular methods are being emphasized in systematics as tools to discriminatebetween populations and/or species. Such techniques are complementary to morpho-logical analysis and crossbreeding studies, and vice versa. DNA-based tools are cur-rently applied to detect genetic variation at any taxonomic level and to measure relat-edness between taxa in mites. Mitochondrial DNA (mtDNA) provides useful mark-ers for the genetic characterization and studies of phylogenetic relationships of
9
General introduction
Nazer-chap1.qxd 14-9-2012 10:35 Page 9

organisms at different taxonomic levels (Hebert et al. 2003a,b; Tixier al. 2003, 2004,2006b, 2008a,b; Okassa et al. 2010). DNA barcoding is a method proposed to classi-fy animals to the species level using a fragment of mtDNA, usually the cytochromeoxidase subunit I (COI), well suited for population and species identification (Hebertet al. 2003a,b). The combination of morphological and molecular data has led to sep-aration of species that were hard to distinguish based on morphological criteria only(e.g., Tixier et al. 2006b)
In this thesis I will focus on geographic populations as genetic entities and willstudy whether they are reproductively isolated from other populations, whether they aremorphologically and/or genetically different and finally in which biocontrol-relevanttraits they differ. Hence, I will perform morphological and molecular analysis, cross-breeding experiments and – in case populations are not compatible – also experimentsto assess the causes of incompatibility (especially the role endosymbionts) (CHAPTERS 2-4). This approach has the practical advantage that genetic integrity of a promising pop-ulation of biocontrol agents is maintained even after it is introduced in the habitat ofanother conspecific population. Finally, I will determine differences between popula-tions and species in competitive abilities (CHAPTER 5) and in the traits thought to be rel-evant for biological control (CHAPTER 6).
Thesis outlineIn this thesis, I focus on mainly two species of predatory mites (Acari: Phytoseiidae) thatare found under the perianth of coconuts: Neoseiulus paspalivorus DeLeon and N. barakiAthias-Henriot. There, they feed on coconut mites (Aceria guerreronis Keifer) which inturn are plant parasites in that they consume the contents of meristematic cells in thetissue protected by the coconut perianth. Each of the two predator species has beenfound in the continents and countries explored for their fauna of predatory mites inhab-iting coconuts (Fernando et al. 2003; Lawson-Balagbo et al. 2008; Reis et al. 2008;Fernando and Aratchige 2009; Banerjee and Gupta 2011; Negloh et al. 2011; Zannou etal. unpublished; Fernando et al. unpublished). Thus, they occupy the same niche in thatthey share coconut mites as prey and they are likely to be each other’s competitor. Basedon morphological criteria these species belong to the same genus and they are closelyrelated systematically (Zannou et al. 2006; De Moraes et al. 2004). These two predatorspecies are considered to be candidates for the biological control of coconut mites. Toprovide a sound basis for identification and characterization of these natural enemiesfor utilization in biological control programs (Moraes et al. 1987; McMurtry 2010), Iinvestigated several geographic populations of each of the two species. I assessed thedegree of reproductive compatibility and examined the extent of morphological andgenetic variation between these populations of predatory mites. For each of these pop-ulations, I studied some traits thought to be relevant for biological control.
10
CHAPTER 1
Nazer-chap1.qxd 14-9-2012 10:35 Page 10

11
In CHAPTER 2 three populations of predatory mites identified as N. paspalivorus –from Benin, Ghana and Brazil – were subject to a test on reproductive compatibility andmultivariate analysis on 32 morphological characters. Multivariate morphometry is themost suitable approach for analyzing morphological variation on a wide set of morpho-metric and meristic traits. These methods are currently used to provide information oncharacter variability within and between phytoseiid mite populations and to contributeto the taxonomy knowledge of the studied group (i.e., Tixier al. 2003, 2004, 2006b,2008a,b; Okassa et al. 2010).
In CHAPTER 3 I investigate the genetic variation in the three populations identifiedas N. paspalivorus as well as the cause of their reproductive isolation. First, I used themitochondrial cytochrome oxidase subunit I (COI) primers to assess the genetic varia-tion between populations. Then, I used 16S rDNA primers to check for the presence ofendosymbiotic bacteria Wolbachia and Cardinium in those populations, and to analyze thephylogenetic relationships of each endosymbiont from each host population and fromother mite and insect populations of which the endosymbiont sequences are knownfrom GenBank. In addition, an antibiotic test was also conducted to determine whethereach symbiont can cause cytoplasmic incompatibility in its host.
In CHAPTER 4 I applied an integrative approach combining multivariate morphom-etry, crossbreeding and molecular analysis to four populations previously identified asN. baraki, one from Benin, two from Tanzania and one from Brazil.
In CHAPTER 5 I report on intraguild interactions observed in cross-pairing experi-ments conducted with the aim to assess reproductive barriers between N. baraki, N. pas-palivorus and N. neobaraki. These intraguild interactions may play a role in niche compe-tition between different species, but they are possibly also relevant in creating prezygot-ic barriers between populations of the same species.
In CHAPTER 6, I review the literature on predatory mites found under the perianthof coconuts and focus on the traits that are thought to be vital for their impact oncoconut mite control. First, I assess the features of numerical abundance of the preda-tor species. Then, I focus on interspecific and inter-population differences in size, lifehistory on a diet of coconut mites or alternative food (pollen), as well as predation.Finally, I present a phylogenetic analysis for all populations of the three predator speciesbased on COI sequences to test the hypothesis that they represent a complex species.
The general discussion (CHAPTER 7) of this Thesis is devoted to the question whichof the geographic populations may have the best combination of the traits for biologi-cal control of coconut mites. Moreover, I try to provide different alternative scenariosfor the worldwide invasion of the three species of predatory mites studied in thisThesis. Finally, I scrutinize the critical assumption underlying this Thesis that reproduc-tive isolation is a fixed trait, rather than a context-dependent plastic phenomenon.
General introduction
Nazer-chap1.qxd 14-9-2012 10:35 Page 11

AcknowledgementsI am very greatful to Maurice W. Sabelis for his valuable comments on earlier versions of thischapter.
ReferencesBanerjee D, Gupta SK (2011) Incidence and seasonal occurrence of mites (Acari) on coconut in
West Bengal, India. Zoosymposia 6:82–85.Breeuwer JAJ (1997) Wolbachia and cytoplasmic incompatibility in the spider mite Tetranychus urticae
and T. turkestani. Heredity 79:41–47.Chant DA, McMurtry JA (2007) Illustrated Keys and Diagnoses for the Genera and Subgenera of the
Phytoseiidae of the World (Acari: Mesostigmata). Indira Publishing House, West Bloomfield, USA.Clark AR, Walter GH (1995) Strains and the classical biological control of insect pests. Can J Zool
73:1777-1790Diehl SR, Bush GL (1984) An evolutionary and applied perspective of insect biotypes. Ann Rev
Entomol 29:471–504.Fernando LCP, Aratchige NS (2009) Status of coconut mite Aceria guerreronis and biological con-
trol research in Sri Lanka. In: M.W. Sabelis and J. Bruin (eds.) Trends in Acarology. Proceedings ofthe 12th International Congress. Springer, The Netherlands, pp. 419–423.
Fernando LCP, Aratchige NS, Peiris TSG (2003) Distribution pattern of coconut mite Aceria guer-reronis and its predator Neoseiulus aff. paspalivorus on coconut palms. Exp Appl Acarol31:71–78.
Gerson U, Smiley RL, Ochoa R (2003) Mites (Acari) for Pest Control. 539p., Blackwell Science, UK.Giordano R, O’Neill LS, Robertson HM (1995) Wolbachia infections and the expression of cyto-
plasmic incompatibility in Drosophila sechellia and D. mauritiana. Genetics 140:1307–1317.Gotoh T, Noda H, Ito S (2007) Cardinium symbionts cause cytoplasmic incompatibility in spider
mites. Heredity 98:13–20.Gotoh T, Noda H, Fujita T, Iwadate K, Higo Y, Saito S, Ohtsuka S (2005) Wolbachia and nuclear-
nuclear interactions contribute to reproductive incompatibility in the spider mite Panonychusmori (Acari: Tetranychidae). Heredity 94:237–246.
Gotoh T, Oku H, Moriya K, Odawara M (1995) Nucleus-cytoplasm interactions causing repro-ductive incompatibility between two populations of Tetranychus quercivorus Ehara et Gotoh(Acari: Tetranychidae). Heredity 74:105–114.
Grant V (1981) Plant Speciation. 2nd ed. New York: Columbia University Press.Hebert PND, Cywinska A, Ball SL, deWaard JR (2003a) Biological identifications through DNA
barcodes. Proc R Soc Lond B 270:313–321.Hebert PND, Ratnasingham S, deWaard JR (2003b) Barcoding animal life: cytochrome c oxidase
subunit 1 divergence among closely related species. Proc R Soc Lond B 270:S96–S99.Helle W, Sabelis MW (1985) Spider Mites: Their Biology, Natural Enemies and Control. World Crop
Pests Vol. 1B, Elsevier Amsterdam, 458 ppHess RT, Hoy MA (1982) Microorganisms associated with the spider mite predator Metaseiulus
(=Typhlodromus) occidentalis. Electron microscopic observations. J Invert Pathol 40:98–106.Hoy MA, Cave FE (1988) Premating and post-mating isolation among populations of Metaseiulus
occidentalis (Nesbitt) (Acarina: Phytoseiidae). Hilgardia 56:1-20.Johanowicz DL, Hoy MA (1999) Experimental induction and termination of non-reciprocal
reproductive incompatibility in a parahaploid mite. Entomol Exp Appl 87:51–58.Johanowicz DL, Hoy MA (1998) The manipulation of arthropod reproduction by Wolbachia
endosymbionts. Florida Entomol 83:310–317.Koevoets T, Niehuis O, Van de Zande L, Beukeboom LW (2011) Hybrid incompatibilities in the
parasitic wasp genus Nasonia. Heredity 108, 302–311.Laven H (1959) Speciation by cytoplasmic isolation in Culex strains. Evolution 5:370–375.
12
CHAPTER 1
Nazer-chap1.qxd 14-9-2012 10:35 Page 12

Laven H (1967) Evolution and speciation in Culex pipiens. In: Genetics of Insects vectors of disease. EdsJW Wright R Pal, Holland: Elsvier, pp 251-271.
Lawson-Balagbo ML, Gondim Jr MGC, Moraes GJ, Hanna R, Schausberger P (2008) Explorationof acarine fauna on coconut palm in Brazil with emphasis on Aceria guerreronis (Acari:Eriophyidae) and its natural enemies. Bull Entomol Res 98:83–96.
Lesna I, Sabelis MW (2002) Genetic polymorphism in prey preference at a small spatial scale: acase study of soil predatory mites (Hypoaspis aculeifer) and two species of astigmatic mites asprey. In: Acarid Phylogeny and Evolution: Adaptation in Mites and Ticks - Proceedings of the IVSymposium of the European Association of Acarologists (eds. F Bernini, R Nannelli, G Nuzzaci andE de Lillo), Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 49-64.
Lesna I, Sabelis MW (1999) Diet-dependent female choice for males with ‘good genes’ in a soilpredatory mite. Nature 401:581-584.
Lindquist EE, Sabelis MW, Bruin J (eds.) (1996) Eriophyoid Mites - Their Biology, Natural Enemies andControl. World Crop Pest Series Vol. 6, Elsevier Science Publishers, Amsterdam, TheNetherlands, 790 + xxxii pp.
Mallet J (1995) A species definition for the Modern Synthesis. TREE 10:294–299.Mayr E (1940) Speciation phenomena in birds. Am Nat 74:249–278.McMurtry JA (2010) Concepts of classification of the Phytoseiidae: Relevance to biological con-
trol of mites. In: MW Sabelis and J Bruin (Eds.) Trends in Acarology: Proceedings of the 12thInternational Congress. Springer, The Netherlands, pp. 393–407.
McMurtry JA, Badii MH, Congdon BD (1985) Studies on a Euseius species complex on avocadoin Mexico and Central America, with a description of a new species (Acari: Phytoseiidae). IntJ Acarol 26:107–116.
McMurtry JA (1980) Biosystematics of three taxa in the Amblyseius filandicus group from SouthAfrica with comparative life history (Acari: Phytoseiidae). Int J Acarol 6:147–156.
McMurtry JA, Mahr DL, Johson HG (1976) Geographic races in the predaceous mite, Amblyseiuluspotentillae (Acari: Phtyoseiidae). Int J Acarol 2:23–28.
Moraes GJ, Lopez PC, Fernando CP (2004). Phytoseiid mites (Acari: Phytoseiidae) of coconutgrowing areas in Sri Lanka, with description of three new species. J Acarol Soc Jpn 13:141-160.
Moraes GJ (1987) Importance of taxonomy in biological control. Insect Sci Appl 8:841–844.Navajas M, Gutierrez J, Lagnel L, Fauvel G, Gotoh T (1999) DNA sequences and cross-breeding
experiments in the hawthorn spider mite Amphitetranychus viennensis reveal high genetic differ-entiation between Japanese and French populations. Entomol Exp Appl 90:113–122.
Negloh K, Hanna R, Schausberger P (2011) The coconut mite, Aceria guerreronis, in Benin andTanzania: occurrence, damage and associated acarine fauna. Exp Appl Acarol 55:361-374.
Noronha ACS, Moraes GJ (2004) Reproductive compatibility between mite populations previous-ly identified as Euseius concordis (Acari: Phytoseiidae). Exp Appl Acarol 32:271–279.
Okassa M, Tixier M-S, Kreiter S (2010) Morphological and molecular diagnostics of Phytoseiuluspersimilis and Phytoseiulus macropilis (Acari: Phytoseiidae). Exp Appl Acarol 52:291–303.
O’Neill SL, Karr TL (1990) Bidirectional incompatibility between conspecific populations ofDrosophila simulans. Nature 348:178-180.
Perrot-Minnot M-J, Guo LR, Werren JH (1996) Single and double infection with Wolbachia in theparasitic wasp Nasonia vitripennis: Effects on compatibility. Genetics 143:961–972.
Pryke SR, Griffith SC (2008) Postzygotic genetic incompatibility between sympatric colormorphs. Evolution 63:793–798.
Reis AC, Gondim MGC, Moraes GJ, Hanna R, Schausberger P, Lawson-Balagbo LE, Barros R(2008) Population dynamics of Aceria guerreronis Keifer (Acari: Eriophyidae) and associatedpredators on coconut fruits in northeastern Brazil. Neotrop Entomol 37:457–462.
Telschow A, Hammerstein P, Werren JH (2005) The effect of Wolbachia versus genetic incompat-ibilities on reinforcement and speciation. Evolution 59:1607–1619.
13
General introduction
Nazer-chap1.qxd 14-9-2012 10:35 Page 13

Tixier M-S, Tsolakis H, Ragusa S, Poinso A, Ferrero M, Okassa M, Barbar Z, Ragusa S, Cheval B,Kreiter S (2011) Integrative taxonomy demonstrates the unexpected between two predatorymite species Cydnodromus idaeus and C. picanus (Acari: Phytoseiidae). Invert Syst 25:273–281.
Tixier M-S, Ferrero M, Okassa M, Guichou S, Kreiter S (2010) On the specific identity of speci-mens of Phytoseiulus longipes Evans (Mesostigmata: Phytoseiidae) showing different feedingbehaviours: morphological and molecular analysis. Bull Entomol Res 17:1–11.
Tixier M-S, Guichou S, Kreiter S (2008a) Morphological variation of the species Neoseiulus califor-nicus (McGregor) (Acari: Phytoseiidae): importance for diagnostic reliability and synonymies.Invert Syst 22:453–469.
Tixier M-S, Kreiter S, Croft BA, Cheval B (2008b) Kampimodromus aberrans (Acari: Phytoseiidae)from the USA: morphological and molecular assessment of its density. Bull Entomol Res98:125–134.
Tixier M-S, Kreiter S, Barbar Z, Ragusa S, Cheval B (2006a) The status of two cryptic species:Typhlodromus exhilaratus Ragusa and Typhlodromus phialatus Athias-Henriot (Acari:Phytoseiidae): consequences for taxonomy. Zoologica Scripta 35:115–122.
Tixier M-S, Kreiter S, Ferragut F, Cheval B (2006b) The suspected synonymy of Kampimodromushmiminai and Kampimodromus adrianae (Acari: Phytoseiidae): morphological and molecularinvestigations. Can J Zool 84:1216–1222.
Tixier M-S, Kreiter S, Croft BA, Cheval B (2004) Morphological and molecular differences in thegenus Kampimodromus Nesbitt. Implications for taxonomy. Phytophaga 14:361–375.
Tixier M-S, Kreiter S, Cheval B, Auger P (2003) Morphometric variation between populations ofKampimodromus aberrans Oudemans (Acari: Phytoseiidae). Implications for the taxonomy ofthe genus. Invert Syst 17:349–358.
Uesugi R, Goka K, Osakabe M (2003) Development of genetic differentiation and postzygoticisolation in experimental metapopulations of spider mites. Exp Appl Acarol 31:161–176.
Zannou ID, Hanna R (2011) Clarifying the identity of Amblyseius swirskii and Amblyseius rykei: arethey two distinct species or two populations of one species? Exp Appl Acarol 53:339–347.
Zannou ID, Moraes GJ, Ueckermann EA, Olivera AR, Yaninek JS, Hanna R (2006) Phytoseiidmites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from Sub-Saharan Africa. Int JAcarol 32:241–276.
14
CHAPTER 1
Nazer-chap1.qxd 14-9-2012 10:35 Page 14

15
Morphological variation and reproductiveincompatibility of three coconut-mite-associatedpopulations of predatory mites identified asNeoseiulus ppaspalivorus (Acari: Phytoseiidae)
ABSTRACT – Predatory mites identified as Neoseiulus paspalivorus DeLeon(Phytoseiidae) have been considered as agents for classical biological control ofthe coconut mite, Aceria guerreronis Keifer (Eriophyidae), in Africa and elsewhere.Preliminary identification of geographically distinct populations as belonging tothe same species (N. paspalivorus) was based on their morphological similarity.However, laboratory studies recently conducted have shown large differences infeeding behaviors and biological characteristics among individuals collected fromthree geographic origins: Brazil (South America), Benin and Ghana (West Africa).As morphologically similar specimens do not necessary belong to the samespecies, we evaluated under laboratory conditions, reproductive compatibilitybetween the specimens from three geographic locations to ascertain their con-specificity. Morphological measurements were also made to determine if there isa means of discriminating between them. Inter-population crosses showed com-plete reproductive isolation between the three geographic populations, but inter-population discontinuities in morphometric characters were absent. These resultsindicate that the tested specimens are distinct biological entities despite morpho-logical similarity. Further molecular genetic studies are therefore proposed,including screening for endosymbionts and assessment of genetic differentiation,to determine the cause of reproductive incompatibility and to clarify the taxo-nomic relationship between those populations.
Experimental and Applied Acarology (2011) 53: 323–238
Coconut, Cocos nucifera L., is one of the main crops in the Tropics. The coconut mite,Aceria guerreronis Keifer, has become one of the most important arthropod pests of
this crop around the world. It causes severe damage to the crop by attacking the meris-tematic tissues underneath the bracts. According to recent assessments the abundanceof the coconut mite and its damage to the crop may be lower, and the predator faunaassociated with coconut mite may be richer in certain areas of Brazil than in Africa(Benin, Ghana and Tanzania) and Sri Lanka (Fernando et al. 2003; Lawson-Balagbo etal. 2008; Reis et al. 2008; Negloh et al. 2010; Fernando et al. unpublished). Over the pasttwo decades, attempts were made to control the pest through the use of chemicals and
2Nazer-chap2.qxd 14-9-2012 10:35 Page 15

biopesticides. These control measures, however, have proven to be costly, often ineffec-tive and ecologically undesirable (Moore and Alexander 1987; Moore et al. 1989).Because of the exotic nature of the coconut mite in Africa (originated from SouthAmerica) (Navia et al. 2005), classical biological control, i.e., is the intentional importa-tion and release of natural enemies in the native range of the target pest, where the pestand its natural enemies co-evolved, for permanent establishment and self-sustained con-trol of the target pest, is thought to be a reasonable approach to combat the coconutmites.
Mites in the family Phytoseiidae are mostly predators that are well-known to play asignificant role in the biological control of arthropod pests such as mites and smallinsects (McMurtry et al. 1970; Helle and Sabelis 1985; Lindquist et al. 1996; Sabelis andVan Rijn 1997; Gerson et al. 2003). The predatory mite Neoseiulus paspalivorus (DeLeon)(Acari: Phytoseiidae) is the most common natural enemy associated with the coconutmite in Brazil (Lawson-Balagbo et al. 2008; Reis et al. 2008). In Benin and Ghana, WestAfrica, N. paspalivorus was also frequently reported on infested coconuts (Negloh et al.2010). However, recent laboratory experiments by Famah Sourassou et al. (in prep.)showed large differences in feeding behaviours and biological characteristics amongspecimens from the three geographic locations: Benin, Ghana (West Africa) and Brazil(Itamaraca, State of Pernambuco). All three geographic populations developed andreproduced on coconut mite prey, but their biological parameters differed. Furthermore,the specimens from Beninese and Brazilian populations were able to develop and repro-duce on Tetranychus urticae Koch (Acari: Tetranychidae), whereas the Ghanaian ones wereable to develop on this prey, yet unable to convert the food obtained from this prey intoeggs. Moreover, the specimens from Brazilian and Ghanaian populations were able todevelop on coconut pollen, whereas those from Beninese populations were not. Allthree geographic populations of N. paspalivorus did not produce any eggs on coconutpollen. Contrasting results with our data were previously reported by Lawson-Balagboet al. (2007), showing that N. paspalivorus specimens collected in Acarau (State Cesara,Brazil) could not feed and develop on T. urticae, whereas they were able to develop andreproduce on coconut pollen. Taken together, there are clear indications that the threeN. paspalivorus geographic populations are biologically different despite their morpho-logical similarity. Such biological differences between morphologically similar popula-tions may indicate cryptic species (Muma and Denmark 1969; Monetti and Croft 1997;Tixier et al. 2003, 2004, 2006a, 2008). To test whether the three allopatric populationsare ‘potentially able to interbreed and produce viable progeny’ (biological species sensuMayr 1940) is necessary to understand the evident biological differences between thethree geographic populations.
The study reported in this article aimed to evaluate reproductive compatibilityamong three geographic populations of predatory mites (Brazil, Benin, and Ghana) that
16
CHAPTER 2
Nazer-chap2.qxd 14-9-2012 10:35 Page 16

have been identified as N. paspalivorus based morphological similarity only. We also con-ducted morphological comparison to determine if there are morphological traits otherthan those used for their identification that may serve to distinguish these populations.
Material and methods
Source populations and rearing techniquesSpecimens of N. paspalivorus were collected from coconuts in October 2005 in Ouidah(06°21’N; 02°097’E), Southern Benin, in September 2006 in Itamaraca (07°46’S;34°52’W), State of Pernambuco, Northeastern Brazil and in December 2008 nearWinneba (05°22’907’’N, 00°38’685’’W), Southern Ghana. These three samples wereused for propagation in a rearing unit consisting of a black PVC tile (4 × 4 × 0.1 cm)placed on top of a foam pad (4 × 4 × 1 cm) resting in a Petri dish (14.5 cm in diame-ter and 1 cm in height). The edges of the tile were covered with a band of tissue paperthat also contacted the foam pad. To prevent mites from escaping, distilled water wassupplied to the Petri dish on a daily basis to keep wet the foam pad and the tissue paper.A tuft of hydrophobic cotton wool covered by a piece of transparent plastic was placedin the center of each rearing unit to serve as an oviposition site for the predators.Colonies of all populations were maintained in a climate-controlled room at 25-27 °C,70-90% relative humidity and L12:D12 photoperiod. The colonies were provided witha new supply of immature stages of T. urticae at three day intervals.
Crossing experimentsEach experiment started with a cohort of 1-day-old eggs obtained from gravid femalesof each of the three populations. To obtain them, one hundred females were confinedto rearing units similar to that described above and offered eggs of T. urticae as prey.After 24 h, each egg laid by the predator was transferred to a new unit, again similar tothat previously described above, except that the PVC was 2.5 cm diameter black PVCdisk and had a hole of 2 mm diameter in the center to serve as an oviposition site. Thenymphal stages of the three populations were reared on all stages of A. guerreronis sup-plied ad libitum. When reaching adulthood, predators were sexed and females and malesof each population were kept apart for the subsequent crosses.
Mating of all possible combinations of recently moulted females and males of thethree populations were considered to determine reproductive compatibilities. All cross-es were set up as a single pair mating between virgin females and males. Additionally, 10virgin females of each population were kept in isolation, to ascertain that mating is nec-essary for oviposition to take place. Each pair was observed daily and the number ofeggs laid was recorded for a period of 10 days. Eggs found in the arenas of a given male-female combination of geographic strains were pooled in a unit similar to that previous-
17
Morphological variation and reproductive incompatibility
Nazer-chap2.qxd 14-9-2012 10:35 Page 17

ly described (8.5 cm in diameter and 1.5 cm in height), and reared to adulthood to deter-mine egg viability, post-embryonic survivorship and sex-ratio. Offsprings of each crosswere backcrossed to assess hybrid fertility. All crossing experiments were carried outsimultaneously under the same environmental conditions as described previously (25-27°C, 70-90% relative humidity and L12:D12 photoperiod). At the end of the observationperiod, females and males of each population were preserved in 70% alcohol for mor-phological analysis.
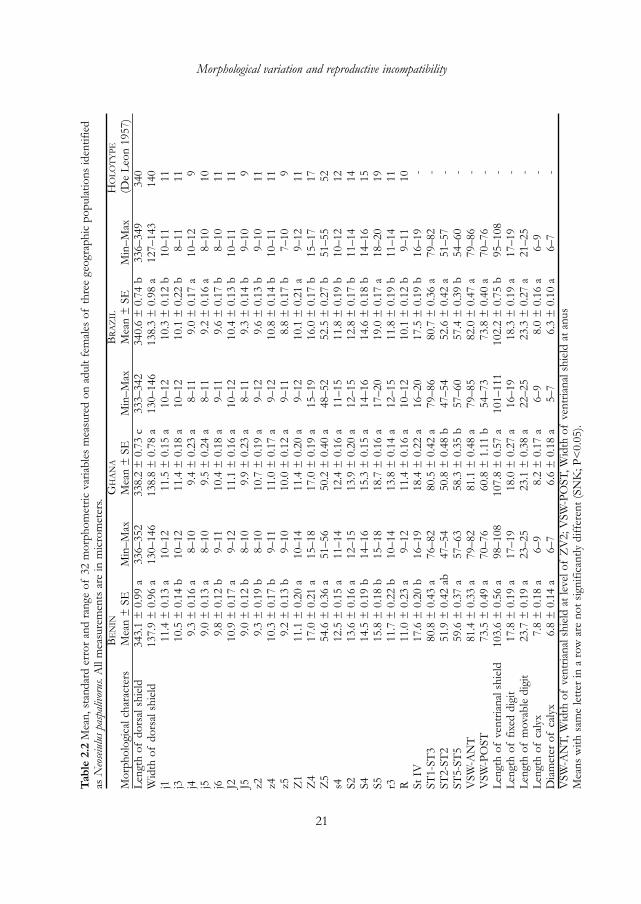
Morphological analysisTwenty adult females and ten adult males of each population were slide-mounted inHoyer’s medium for examination under a phase-contrast microscope. Measurementswere done with an ocular micrometer, at 400× magnification to measure body size andshield dimensions and at 1000× magnification to measure the length of setae, spermath-ecal calyx (spermatodactyl for male) and cheliceral digits. Females and males were char-acterized based on 32 morphological traits (Tables 2.2 and 2.3) that are used common-ly for the identification of phytoseiid mites (i.e., Chant and McMurtry 1994, 1995;Moraes et al. 2004; Zannou et al. 2007). Setal nomenclature follows that of Lindquistand Evans (1965), as applied to the phytoseiids by Rowell et al. (1978) and Chant andYoshida-Shaul (1991). For measurement of cheliceral digits and spermatodactyl, anoth-er set of at least 20 females and 20 males of each population were slide-mounted sepa-rately. The gnathosoma was cut from the rest of the body and a slight pressure wasapplied to the mount by touching the slide coverslip with the rear end of a small brushto help in distinguishing the two cheliceral digits. Holotype measurements, taken fromthe original description (De Leon 1957), were also included in the results for compari-son.
Data analysisCrossing dataA single-factor ANOVA was used to test the effects of crossing types on fecundity andduration of the pre-oviposition period. Data of the latter were log-transformed[ln(x+1)] for ANOVA. Means were compared using the Student-Newman-Keuls multi-ple range test (SNK) at P<0.05.
Morphological dataMorphological characters of females and males were analyzed separately with SAS soft-ware (SAS Institute 2003). For each population and each character examined, descrip-tive statistics (mean, standard error, maximum, minimum) were calculated using theMEANS procedure (Proc MEANS). Differences among geographic populations weretested by one-way analysis of variance (Proc ANOVA) followed by a Newman-Keuls
18
CHAPTER 2
Nazer-chap2.qxd 14-9-2012 10:35 Page 18

19
multiple comparison test at P<0.05. Next, a multifactorial analysis (PrincipalComponent Analysis) and a Canonical Discriminant Analysis were performed to assesspatterns of morphological variation and to identify morphological characters that con-tribute most to morphological differentiation among the three geographic populations.Prior to multifactorial analysis, a discriminant analysis was carried out to examine howmany specimens were correctly classified into their original populations.
Results
Cross-breeding experimentsAll mated females of intra-population crosses laid eggs, but the proportion of oviposit-ing females from inter-population crosses ranged between 80 and 100%. Unmatedfemales did not oviposit (Table 2.1). The pre-oviposition period of offspring from eachof the inter-population crosses (ranging between 2.9 and 5.4 days) was longer than thatof offspring from the intra-population crosses (<2.0 days) (P<0.001; Table 2.1).
Average fecundity was about two- to three-fold higher in intra-population than ininter-population crosses (P<0.001; Table 2.1). All eggs obtained from intra-populationcrosses were viable, with 71% producing females. All eggs produced by inter-populationcrosses were malformed and non-viable, similar to crosses between females from theBeninese population and males from the Brazilian population. In the crossings betweenfemales from Brazil and males from Benin, for which nearly 14% of the eggs produced(13 out of 92 eggs) had a normal shape, only 7.6% of the eggs (7 eggs) hatched givingrise to male progeny only. Eggs were not produced by females of any of the backcross-es involving hybrid males from Brazilian females and Beninese males or Beninese malesand Brazilian females (n = 3 for each combination), although mating was observed. Inthe control backcrosses (n = 3 for each combination), averages of 11.3 and 11.0 eggsper female were recorded for Brazilian and Beninese populations, respectively.
Morphological analysisMean, standard errors and range of the 32 characters are shown in the Tables 2.2 and2.3 for female and male populations, respectively. Significant differences were observedfor 22 and 15 out of the 32 characters measured, respectively among female and malepopulations. However, these differences were very small, except for the width of theventrianal shield at anus level (POST-WVS), which was smaller for the Ghanaianfemales and the length of seta S5, which was slightly shorter for Beninese females. Thestandard errors within populations were low and the differences between the minimaland the maximal values for each of the measured morphological variables (except forPOST-WVS of the Ghanaian females) were very small.
Morphological variation and reproductive incompatibility
Nazer-chap2.qxd 14-9-2012 10:35 Page 19

20
CHAPTER 2
Tabl
e 2.
1Out
com
e of
cros
ses (
mea
ns ±
SE
) inv
olvi
ng th
ree
geog
raph
ic po
pulat
ions
iden
tified
as N
eoseiu
lus p
aspa
livor
us.
CRO
SSn
%Pr
e-N
o.%
%
Im
mat
ure
Sex
Fem
ale ×
Male
ovip
ositi
ngov
ipos
ition
eggs
/de
form
edeg
gsu
rviv
orsh
ipra
tiofe
male
spe
riod
fem
aleeg
gs/f
emale
hatc
habi
lity
(% fe
male
s)I N
TRA
-PO
PULA
TIO
NCR
OSS
ES
Braz
il ×
Bra
zil
2010
01.
6 ±
0.1
9c12
.8 ±
0.6
4a0
100
99.0
± 0
.65
71.9
±3.
92Be
nin
× B
enin
2010
01.
8 ±
0.2
6c12
.2 ±
0.4
8a0
100
99.4
± 0
.51
71.8
±1.
11G
hana
× G
hana
1510
01.
9 ±
0.1
9c10
.1 ±
0.5
5b0
100
100
74.1
±4.
01I N
TER-
POPU
LATI
ON
CRO
SSE
SBr
azil
× B
enin
3083
.33.
8 ±
0.3
1b5.
2 ±
0.3
3c86
7.6
100
0.0*
Beni
n ×
Bra
zil
3093
.32.
9 ±
0.2
5b5.
7 ±
0.5
5c10
00.
0-
-Br
azil
× G
hana
1510
05.
4 ±
0.1
3a3.
8 ±
0.2
9c10
00.
0-
-G
hana
× B
razi
l15
93.3
5.2
± 0
.22a
4.2
± 0
.41c
100
0.0
--
Beni
n ×
Gha
na10
100
3.1
± 0
.10b
5.8
± 0
.66c
100
0.0
--
Gha
na ×
Ben
in10
805.
1 ±
0.5
4a3.
9 ±
0.8
4c10
00.
0-
-M
eans
follo
wed
by
the
sam
e let
ter i
n a
colu
mn
are
not s
igni
fican
tly d
iffer
ent (
SNK
;P>
0.05
)n:
num
ber o
fpa
ir cr
osse
s*:
all p
roge
ny a
re m
ales
Nazer-chap2.qxd 14-9-2012 10:35 Page 20
21
Morphological variation and reproductive incompatibilityTa
ble
2.2
Mea
n,st
anda
rd e
rror
and
rang
e of
32 m
orph
omet
ric v
ariab
les m
easu
red
on a
dult
fem
ales o
fth
ree
geog
raph
ic po
pulat
ions
iden
tified
as N
eoseiu
lus p
aspa
livor
us. A
ll m
easu
rem
ents
are
in m
icrom
eter
s.BE
NIN
GH
AN
ABR
AZ
ILH
OLO
TYPE
Mor
phol
ogica
l cha
ract
ers
Mea
n ±
SE
Min
–Max
Mea
n ±
SE
Min
–Max
Mea
n ±
SE
Min
–Max
(De
Leon
195
7)Le
ngth
of
dors
al sh
ield
343.
1 ±
0.9
9 a
336–
352
338.
2 ±
0.7
3 c
333–
342
340.
6 ±
0.7
4 b
336–
349
340
Wid
th o
fdo
rsal
shiel
d13
7.9
± 0
.96
a13
0–14
613
8.8
± 0
.78
a13
0–14
613
8.3
± 0
.98
a12
7–14
314
0j1
11.4
± 0
.13
a10
–12
11.5
± 0
.15
a10
–12
10.3
± 0
.12
b10
–11
11j3
10.5
± 0
.14
b10
–12
11.4
± 0
.18
a10
–12
10.1
± 0
.22
b8–
1111
j49.
3 ±
0.1
6 a
8–10
9.4
± 0
.23
a8–
119.
0 ±
0.1
7 a
10–1
29
j59.
0 ±
0.1
3 a
8–10
9.5
± 0
.24
a8–
119.
2 ±
0.1
6 a
8–10
10j6
9.8
± 0
.12
b9–
1110
.4 ±
0.1
8 a
9–11
9.6
± 0
.17
b8–
1011
J210
.9 ±
0.1
7 a
9–12
11.1
± 0
.16
a 10
–12
10.4
± 0
.13
b10
–11
11J5
9.0
± 0
.12
b8–
109.
9 ±
0.2
3 a
8–11
9.3
± 0
.14
b9–
109
z29.
3 ±
0.1
9 b
8–10
10.7
± 0
.19
a9–
129.
6 ±
0.1
3 b
9–10
11z4
10.3
± 0
.17
b9–
1111
.0 ±
0.1
7 a
9–12
10.8
± 0
.14
b10
–11
11z5
9.2
± 0
.13
b9–
1010
.0 ±
0.1
2 a
9–11
8.8
± 0
.17
b7–
109
Z1
11.1
± 0
.20
a10
–14
11.4
± 0
.20
a9–
1210
.1 ±
0.2
1 a
9–12
11Z
417
.0 ±
0.2
1 a
15–1
817
.0 ±
0.1
9 a
15–1
916
.0 ±
0.1
7 b
15–1
717
Z5
54.6
± 0
.36
a51
–56
50.2
± 0
.40
a48
–52
52.5
± 0
.27
b51
–55
52s4
12.5
± 0
.15
a11
–14
12.4
± 0
.16
a11
–15
11.8
± 0
.19
b10
–12
12S2
13.6
± 0
.16
a12
–15
13.9
± 0
.20
a12
–15
12.8
± 0
.17
b11
–14
14S4
14.5
± 0
.19
b14
–16
15.3
± 0
.15
a14
–16
14.6
± 0
.18
b14
–16
15S5
15.8
± 0
.18
b15
–18
18.7
± 0
.16
a17
–20
19.0
± 0
.17
a18
–20
19r3
11.7
± 0
.22
b10
–14
13.8
± 0
.14
a12
–15
11.8
± 0
.19
b11
–14
11R
11.0
± 0
.23
a9–
1211
.4 ±
0.1
6 a
10–1
210
.1 ±
0.1
2 b
9–11
10St
IV17
.6 ±
0.2
0 b
16–1
918
.4 ±
0.2
2 a
16–2
017
.5 ±
0.1
9 b
16–1
9-
ST1-
ST3
80.8
± 0
.43
a76
–82
80.5
± 0
.42
a79
–86
80.7
± 0
.36
a79
–82
-ST
2-ST
251
.9 ±
0.4
2 ab
47–5
450
.8 ±
0.4
8 b
47–5
452
.6 ±
0.4
2 a
51–5
7-
ST5-
ST5
59.6
± 0
.37
a57
–63
58.3
± 0
.35
b57
–60
57.4
± 0
.39
b54
–60
-V
SW-A
NT
81.4
± 0
.33
a79
–82
81.1
± 0
.48
a79
–85
82.0
± 0
.47
a79
–86
-V
SW-P
OST
73.5
± 0
.49
a70
–76
60.8
± 1
.11
b54
–73
73.8
± 0
.40
a70
–76
-Le
ngth
of
vent
riana
l shi
eld10
3.6
± 0
.56
a98
–108
107.
8 ±
0.5
7 a
101–
111
102.
2 ±
0.7
5 b
95–1
08-
Leng
th o
ffix
ed d
igit
17.8
± 0
.19
a17
–19
18.0
± 0
.27
a16
–19
18.3
± 0
.19
a17
–19
-Le
ngth
of
mov
able
digi
t23
.7 ±
0.1
9 a
23–2
523
.1 ±
0.3
8 a
22–2
523
.3 ±
0.2
7 a
21–2
5-
Leng
th o
fca
lyx7.
8 ±
0.1
8 a
6–9
8.2
± 0
.17
a6–
98.
0 ±
0.1
6 a
6–9
-D
iamet
er o
fca
lyx6.
8 ±
0.1
4 a
6–7
6.6
± 0
.18
a5–
76.
3 ±
0.1
0 a
6–7
-V
SW-A
NT,
Wid
th o
fve
ntria
nal s
hield
at l
evel
ofZ
V2;
VSW
-PO
ST,W
idth
of
vent
riana
l shi
eld a
t anu
sM
eans
with
sam
e let
ter i
n a
row
are
not
sign
ifica
ntly
diffe
rent
(SN
K;P
<0.
05).
Nazer-chap2.qxd 14-9-2012 10:35 Page 21

22
CHAPTER 2Ta
ble
2.3
Mea
n,st
anda
rd e
rror
and
rang
e of
32 m
orph
omet
ric v
ariab
les m
easu
red
on a
dult
male
s of
thre
e ge
ogra
phic
popu
latio
ns id
entif
ied a
sN
eoseiu
lus p
aspa
livor
us. A
ll m
easu
rem
ents
are
in m
icrom
eter
s.BE
NIN
GH
AN
ABR
AZ
ILH
OLO
TYPE
Mor
phol
ogica
l cha
ract
ers
Mea
n ±
SE
Min
–Max
Mea
n ±
SE
Min
–Max
Mea
n ±
SE
Min
–Max
(De
Leon
195
7)Le
ngth
of
dors
al sh
ield
257.
1 ±
1.7
3 a
250–
269
257.
7 ±
1.1
6 a
253–
263
256.
8 ±
2.1
1 a
241–
263
260
Wid
th o
fdo
rsal
shiel
d12
4.5
± 0
.67
a12
0–12
7 11
7.6
± 0
.87
b11
4 –1
2012
4.0
± 0
.56
a12
0–12
712
0j1
9.6
± 0
.19
a8–
109.
8 ±
0.2
5 a
9–11
10.0
± 0
.12
a9–
10-
j310
.0 ±
0.2
6 a
8–11
10.2
± 0
.12
a9–
1110
.3 ±
0.1
6 a
9–11
-j4
7.8
± 0
.25
b6–
97.
6 ±
0.2
2 b
6–9
8.5
± 0
.16
a7–
9-
j57.
8 ±
0.2
6 a
6–9
8.1
± 0
.21
a7–
98.
4 ±
0.1
9 a
7–9
-j6
8.6
± 0
.22
a7–
109.
4 ±
0.2
7 a
8- 1
19.
0 ±
0.3
6 a
7–10
-J2
9.2
± 0
.33
a8–
119.
6 ±
0.1
9 a
8–10
10.0
± 0
.26
a9–
11-
J57.
0 ±
0.2
0 b
6–8
6.6
± 0
.19
b6–
87.
6 ±
0.2
9 a
6–9
-z2
8.2
± 0
.27
b6–
99.
4 ±
0.2
0 a
8–10
8.6
± 0
.22
b7–
10-
z49.
1 ±
0.2
6 b
8–10
10.0
± 0
.18
a9–
119.
8 ±
0.2
2 a
9–11
-z5
7.3
± 0
.22
a6–
98.
0 ±
0.2
0 a
7–9
7.8
± 0
.19
a7–
9-
Z1
9.6
± 0
.19
a8–
109.
2 ±
0.4
2 a
7–11
9.5
± 0
.33
a9–
10-
Z4
14.4
± 0
.27
ab12
–15
14.0
± 0
.36
b13
–16
15.2
± 0
.31
a14
–16
-Z
540
.4 ±
0.6
4 a
36–4
441
.1 ±
0.6
5 a
38–4
440
.0 ±
0.6
9 a
37–4
5-
s410
.5 ±
0.2
7 a
9–11
11.2
± 0
.18
a10
–12
10.8
± 0
.19
a10
–11
-S2
11.2
± 0
.18
b10
–12
12.1
± 0
.19
a11
–13
11.4
± 0
.22
b10
–12
-S4
12.2
± 0
.31
b11
–14
13.1
± 0
.33
a11
–15
11.8
± 0
.20
a11
–13
-S5
12.7
± 0
.29
b11
–14
14.8
± 0
.34
a13
–16
15.0
± 0
.32
a13
–16
-r3
9.6
± 0
.19
b9–
1010
.2 ±
0.2
5 ab
9–11
10.8
± 0
.26
a10
–12
-R
9.1
± 0
.32
b8–
109.
5 ±
0.2
0 ab
8–10
10.2
± 0
.25
a9–
11-
St IV
15.0
± 0
.18
b14
–16
16.1
± 0
.22
a15
–17
15.3
± 0
.25
b14
–16
-ST
1-ST
367
.2 ±
0.6
3 a
63–7
066
.2 ±
0.5
6 a
63–7
065
.9 ±
0.4
2 a
63–6
6-
ST2-
ST2
40.2
± 0
.48
ab38
–41
40.0
± 0
.51
b38
–41
41.5
± 0
.31
a41
–44
-ST
5-ST
533
.6 ±
0.5
1 a
32–3
530
.7 ±
0.6
7 b
28–3
533
.0 ±
0.5
1 a
31–3
5-
VSW
-AN
T11
8.0
± 1
.62
a11
1–12
711
7.0
± 1
.31
a11
1–12
311
6.6
± 1
.31
a10
8–12
0-
VSW
-PO
ST76
.7 ±
1.5
5 a
70–8
275
.1 ±
1.0
6 a
70–7
978
.6 ±
0.9
2 a
73–8
2-
Leng
th o
fve
ntria
nal s
hield
92
.5 ±
0.9
2 a
89–9
894
.4 ±
0.6
3 a
92–9
892
.0 ±
1.0
5 a
85–9
5-
Leng
th o
ffix
ed d
igit
12.0
± 0
.20
b11
–12
12.8
± 0
.26
a11
–14
12.3
± 0
.25
ab11
–14
-Le
ngth
of
mov
able
digi
t17
.6 ±
0.3
0 a
16–1
917
.2 ±
0.2
5 a
16–1
917
.7 ±
0.2
2 a
16–1
9-
Leng
th o
fsp
erm
atod
.sha
ft11
.7 ±
0.2
0 a
11–1
212
.0 ±
0.2
0 a
11–1
211
.8 ±
0.2
1 a
11–1
2-
Leng
th o
fsp
erm
atod
.foo
t4.
6 ±
0.3
2 a
4–6
4.8
± 0
.25
a4–
64.
5 ±
0.2
7 a
4–6
-Sp
erm
atod
,Spe
rmat
odac
tyl
Mea
ns w
ith sa
me
lette
r in
a ro
w a
re n
ot si
gnifi
cant
ly di
ffere
nt (S
NK
;P<
0.05
).
Nazer-chap2.qxd 14-9-2012 10:35 Page 22

Predicted classification of specimens based on discriminant analysis is shown inTable 2.4. Overall, 86.7% of female specimens and 63.3% of male specimens were clas-sified in the population of origin. The majority of misclassifications for males con-cerned the Ghanaian and Brazilian specimens (7 of 11, or 63.6%), whereas for females50% of the misclassifications concerned the Beninese and Brazilian populations, andthe Ghanaian and the Brazilian populations. The results of discriminant analysis werecorroborated by the principal component analysis (Figure 2.1); substantial overlaps ofmeasurements of females occurred between the Brazilian and Beninese specimens, andbetween the Brazilian and the Ghanaian specimens. Overlaps were observed betweenthe Ghanaian and Beninese female specimens (Figure 2.1a). In relation to male speci-mens, considerable level of overlap was found between the measurements on theBrazilian and Beninese specimens (Figure 2.1b). The canonical discrimination analysisallowed a clear distinction between the females of the three populations (Figure 2.2a).The Brazilian and Beninese specimens are the most distant, while the Ghanaian speci-mens were intermediate between latter two. However, in males, only the Ghanaian spec-imens could be clearly distinguished from the two others (Figure 2.2b). The posteriorwidth of the ventrianal shield (VSW_POST) and the lengths of j1, Z5, S5 and r3 con-tributed the most to the morphological differentiation between females of the threegeographic populations, whereas the width of the dorsal and genital shields and lengthof seta S5 contributed the most to the morphological differentiation of males from thethree geographic populations.
Neoseiulus paspalivorus females from Ghana had a slightly narrower dorsal shield andrelatively longer ventrianal shield with a reduced width at the level of the anal shield(posterior width of ventrianal shield) and slightly shorter seta Z5, while males of thesame population had slightly narrower dorsal and genital shields. The two sexes ofBeninese specimens differed from those of Brazil and Ghana by having slightly short-er seta S5. Specimens from Brazil appeared to be intermediate, but much closer toBeninese specimens by having a similar size of ventrianal shield (for female) and geni-tal shield (for male) with the Beninese specimens and similar length of setae S5 with theGhanaian specimens.
23
Morphological variation and reproductive incompatibility
Table 2.4 Classification based on discriminant analysis of 32 morphological characters of threegeographic populations of Neoseiulus paspalivorus females and males.
FEMALE MALE% well- Benin Ghana Brazil % well- Benin Ghana Brazilclassified* classified*
Benin 90 18 0 2 60 6 2 2Ghana 85 0 17 3 80 0 8 2Brazil 85 2 1 17 50 0 5 5Total 86.7 20 18 22 63.33 6 15 9*Percentage of well-classified individuals in their original populations
Nazer-chap2.qxd 14-9-2012 10:35 Page 23

DiscussionThe results of univariate and multivariate analyses are consistent with the previous iden-tification of all three populations– from Benin, Ghana and Brazil– as belonging to thesame species. Overlapping morphometric values indicate great morphological similarityamong the geographic populations tested. The measurements of the specimens exam-ined were similar to the holotype measurements of N. paspalivorus (De Leon 1957) andmeasurements of females for the same species from Sri Lanka (Moraes et al. 2004).Differences in setal measurements were small, and the discrimination between the geo-
24
CHAPTER 2
Figure 2.1 Principal component analysis based on 32 morphometric characters measured onadult females (a) and (b) males from three geographic specimens identified as Neoseiulus pas-palivorus: Polygons formed based on the projection of the individuals of each population ontothe principal components 1and 2.
Nazer-chap2.qxd 14-9-2012 10:35 Page 24

graphic populations by multifactorial analyses was based on only some characters thathave low weight in species differentiation. McMurtry (1980) indicated that cautionshould be exercised in using relatively small differences in setal length to differentiatespecies. Tixier et al. (2006b) reported large differences in setal lengths betweenKampimodromus hmiminai (McMurtry and Bounfour) from France and Morocco and K.adrianae (Ferragut and Pena-Estévez) from the Canary Islands, but molecular analysisindicated that they are synonyms. However, the results of cross-breeding indicate repro-ductive isolation between the populations investigated, which, should therefore be con-sidered as separate species (Mayr 1940). The absence of oviposition in unmated N. pas-
25
Morphological variation and reproductive incompatibility
Figure 2.2 Canonical discrimination analysis of 32 morphometric characters measured on adultfemales (a) and (b) males from three geographic populations of Neoseiulus paspalivorus: Polygonsformed based on the projection of the individuals of each population onto the canonical vari-able 1 and 2.
Nazer-chap2.qxd 14-9-2012 10:35 Page 25

palivorus females observed in this study was in agreement with what is known to date forthe majority of phytoseiids (Croft 1970; McMurtry et al. 1976; McMurtry 1980; Moraesand McMurtry 1981; Noronha and Moraes 2002, 2004). Thus, the high proportion ofovipositing females observed in inter-population crosses imply that mating hadoccurred, therefore indicating the absence of pre-mating reproductive barriers betweenthe populations under test. The complete bidirectional post-mating reproductive incom-patibility observed between the geographic populations was expressed in the form ofreduced fecundity, zygotic mortality and a male-biased sex ratio among the few sterileoffspring.
Bidirectional incompatibility is a rare phenomenon, as unidirectional incompatibil-ities are the most common in phytoseiid and tetranychid mite populations (Hoy andCave 1988; Gotoh and Noguchi 1989; Gotoh et al. 1995; Breeuwer 1997; Johanowiczand Hoy 1998; Vala et al. 2000, 2002; Noronha and Moraes 2002, 2004). Our results ofbidirectional incompatibility among geographic populations of N. paspalivorus are there-fore quiet exceptional, but are similar to those reported by Monetti and Croft (1996)with respect to crosses between the morphologically similar phytoseiid species Neoseiuluscalifornicus (McGregor) and N. fallacis (Garman). Moreover, Klimov et al. (2004)observed postzygotic reproductive isolation between two cryptic species of Sancassaniamites, Sancassania salasi and S. ochoai (Klimov, Lekveishvili & OConnor), and molecularanalysis of these populations showed that they represent distinct species.
The causes of reproductive incompatibility are poorly studied in mites, particularlyin phytoseiids. Only in the phytoseiid Galendromus occidentalis (Nesbitt) and in some spi-der mites, the endosymbionts Wolbachia and more recently Cardinium have been demon-strated to mediate unidirectional reproductive incompatibility (Hess and Hoy 1982;Gotoh et al. 1995; Johanowicz and Hoy 1996; Breeuwer 1997; Vala et al. 2000, 2002,2003; Gotoh et al. 2006; Ros and Breeuwer 2009). Bidirectional incompatibility isassumed to be caused by either negative nuclear-nuclear genes interactions, as has beenreported in the spider mite Panonychus mori Yokoyama (Gotoh et al. 2005), or infectionby different strains of Wolbachia, as is well documented for insects (Laven 1959, 1967;Mercot et al. 1995; O’Neill and Karr 1990; Clancy and Hoffmann 1996). Althoughmales and females from different geographic origins were able to mate, we did notobtain F1 hybrids (except in the cross involving Beninese females and Brazilian maleswhere very few sterile males were produced). This apparent lack of gene exchangeamong the three geographically isolated populations revealed a pattern of reproductiveisolation among them. Moreover, the crossing data appeared to corroborate the previ-ously known biological differences observed among these three geographic populations(Famah Sourassou et al. in prep).
Taken together, we expect the reproductive incompatibility observed in this study tobe the result of genetic divergence due to allopatric differentiation among the popula-
CHAPTER 2
26
Nazer-chap2.qxd 14-9-2012 10:35 Page 26

tions investigated (Hurt and Hedrick 2003; Dettman et al. 2008) rather than to be causedby Wolbachia. Even if endosymbionts would be present, there is no guarantee that theywere the cause of reproductive isolation because some Wolbachia strains are incapable ofinducing reproductive incompatibility in their host (Giordano et al. 1995; Turrelli andHoffmann 1995; Gotoh et al. 2005). For example, Gotoh et al. (2005) observed bidirec-tional reproductive incompatibility between two Japanese populations of P. mori (fromHanayama and Toyama) although harbouring the same Wolbachia strain. In this case, theprocedure suggested by Breeuwer (1997), i.e., crossing experiments in combination withantibiotic treatment should be used to demonstrate whether they are involved in thereproductive incompatibility. Reproductive isolation is generally thought to develop bythe gradual accumulation of genetic differences between populations as a by-product ofother adaptive or neutral genetic changes that take place in allopatry (Mayr 1963;Charlesworth et al. 1987; Coyne 1992, 1993; Wu and Davis 1993; Drès and Mallet 2002;Gavrilets 2003; Coyne and Orr 2005; Dettman et al. 2008). Moreover, reproductive iso-lation has been shown to represent an initial step in the speciation process by preventingor greatly reducing gene flow between populations (Laven 1959, 1967; Conner and Saul1986; Thompson 1987; 1995; Dettman et al. 2008), and our data are therefore consistentwith incipient allopatric speciation (Drès and Mallet 2002; Gavrilets 2003; Coyne and Orr2005) among these three geographically isolated specimens identified as N. paspalivorus.
In conclusion, we found that three geographically isolated populations of predato-ry mites preliminarily identified as N. paspalivorus, show reproductive isolation indicatingthat they are distinct biological species despite morphological similarity. Based on thecrossing data reported in this study, we propose that there is every reason to screen forthe presence of endosymbionts and to perform molecular characterization in order totest the hypothesis on allopatric speciation. It would be also informative to use molec-ular methods for determining the origin and invasion route of these predatory mites incomparison with what is already known for A. guerreronis (Navia et al. 2005), i.e the preyof these predatory mites and the pest of coconut palms. By including more geograph-ic populations, our work may provide a basis for inferring the intercontinental andbetween-country invasion of these predatory mites and creates a solid basis for findingthe best strains for developing biological control of coconut mites.
AcknowledgementsWe thank R. Houndafoche, P. Sovimi, C. Kededji and B. Bovis for their assistance in maintainingthe predator colonies and conducting experiments. We also thank A. Onzo, D. Gnanvossou andM. Toko for their encouragements and advices. Many thanks go to Sam Korie for providing assis-tance on the statistical analysis of the data, and to Fabien Hountondji for his comments on themanuscript. The authors also thank M.G.C. Gondim Jr, for helping to locate and for supplyingthe specimens of N. paspalivorus from Brazil. This research was supported by the InternationalInstitute of Tropical Agriculture (IITA) and the University of Amsterdam, the Netherlands,
27
Morphological variation and reproductive incompatibility
Nazer-chap2.qxd 14-9-2012 10:35 Page 27

through a grant from the Board of the Netherlands Foundation for the advancement of TropicalResearch (WOTRO). This study is part of a PhD thesis of the senior author.
ReferencesBreeuwer JAJ (1997) Wolbachia and cytoplasmic incompatibility in the spider mites Tetranychus urticae
and T. turkestani. Heredity 79:41–47.Chant DA, McMurtry JA (1994) A review of the subfamilies Phytoseiinae and Typhlodrominae
(Acari: Phytoseiidae. Int J Acarol 20(4): 223–310.Chant DA, McMurtry JA (2005) Review of the subfamily Amblyseiinae Muma (Acari:
Phytoseiidae) Part IV. The tribe Euseiini N. tribe, subtribes Typhlodromalina, N. subtribe,Euseiina N. subtribe and Ricoseiina N. subtribe. Int J Acarol 31:187–222.
Chant DA, Yoshida-Shaul E (1991) Adult ventral setal patterns in the family Phytoseiidae (Acari:Gamasina). Int J Acarol 17(3):187–199.
Charlesworth B, Coyne JA, Barton NH (1987) The relative rates of evolution of sex chromosomesand autosomes. Am Nat 130:113–146.
Clancy DJ, Hoffmann AA (1996) Cytoplasmic incompatibility in Drosophila simulans: evolving com-plexity. Trends Ecol Evol 11:145–146.
Conner GW, Saul GB (1986) Acquisition of incompatibility by inbred wild-type stocks ofMormoniella. J Heredity 77:211–213.
Coyne JA (1992) Genetics and speciation. Nature 355: 511–515.Coyne JA (1993) The genetics of an isolating mechanism between two sibling species of Drosophila.
Evolution 47: 778–788.Coyne JA, Orr AH (2005) Speciation in the 21st century. Heredity 95:105–109.Croft BA (1970) Comparative study on four strains of Typhlodromus occidentalis (Acarina: Phytosei-
idae). I. Hybridization and reproductive isolation studies. Ann Entomol Soc Am 63:1559–1563.De Leon D (1957) Three new Typhlodromus from southern Florida (Acarina: Phtyoseiidae). Florida
Entomol 40:141–144.Dettman J, Aderson J, Kohn L (2008) Divergent adaptation promotes reproductive isolation
among experimental populations of the filamentous fungus Neurospora. BMC Evol Biol 1:35.Drès M, Mallet J (2002) Host races in plant-feeding insects and their importance in sympatric spe-
ciation. Phil Trans R Soc Lond B 357:471–492.Fernando LCP, Aratchige NS, Peiris TSG (2003) Distribution pattern of coconut mite Aceria guer-
reronis and its predator Neoseiulus aff. paspalivorus on coconut palms. Exp Appl Acarol 31:71–78.Gavrilets S (2003) Models of speciation. What have we learned in 40 years? Evolution 57:2197–2215.Giodarno R, O’Neill SO, Robertson HM (1995) Wolbachia infection and the expression of cytoplas-
mic incompatibility in Drosophila sechellia and Drosophila mauritiana. Genetics 140:1307–1317.Gerson U, Smile RL, Ochoa R (2003) Mites (Acari) for Pest Control. 539p., Blackwell Science, UKGotoh T, Noda H, Ito S (2006) Cardinium symbionts cause cytoplasmic incompatibility in spider
mites. Heredity 98:13-20.Gotoh T, H Noda, T Fujita, K Iwadate, Y Higo, S Saito, S Ohtsuka (2005) Wolbachia and nuclear-
nuclear interactions contribute to reproductive incompatibility in the spider mite Panonychusmori (Acari: Tetranychidae). Heredity 94:237–246.
Gotoh T, Nugochi O (1989) Developmental success and reproductive incompatibility among popula-tions of the European red mite Panonychus ulmi (Acari: Tetranychidae). Exp Appl Acarol 10:157–165.
Gotoh T, Oku H, Moriya K, Odawara M (1995) Nucleus-cytoplasm interactions causing repro-ductive incompatibility between two populations of Tetranychus quercivorus Ehara et Gotoh(Acari: Tetranychidae). Heredity 74:105–114.
Helle W, Sabelis MW (1985) Spider Mites: Their Biology, Natural Enemies and Control. World CropPests Vol. 1B, Elsevier Amsterdam, 458 pp.
28
CHAPTER 2
Nazer-chap2.qxd 14-9-2012 10:35 Page 28

Hess RT, Hoy MA (1982) Microorganisms associated with the spider mite predator Metaseiulus(=Typhlodromus) occidentalis. Electron microscopic observations. J Invert Pathol 40:98–106.
Hoy MA, Cave FE (1988) Premating and post-mating isolation among populations of Metaseiulusoccidentalis (Nesbitt) (Acarina: Phytoseiidae). Hilgardia 56:1–20.
Hurt CR, Hedrick PW (2003) Initiation stages of reproductive isolation in two species of theendangered sonorant topminnow. Evolution 57(12): 2835-2841.
Johanowicz DL, Hoy MA (1998) Experimental induction and termination of non-reciprocalreproductive incompatibility in a parahaploid mite. Entomol Exp Appl 87(1):51–58.
Klimov PB, Lekveishvili M, Dowling APG, OConnor BM (2004) Multivariate analysis of mor-phological variation in two cryptic species of Sancassania (Acari: Acaridae) from Costa Rica.Ann Entomol Soc Am 97(2):322–345.
Laven H (1959) Speciation by cytoplasmic isolation in Culex strains. Evolution 5:370–375.Laven H (1967) Evolution and speciation in Culex pipiens. In Genetics of Insects Vectors of Disease.
Eds JW Wright, R Pal, pp. 251–271. Holland: Elsevier.Lawson-Balagbo ML, Gondim Jr MGC, Moraes GJ, Hanna R, Schausberger P (2007) Life histo-
ry of the predatory mites Neoseiulus paspalivorus and Proctolaelaps bickleyi, candidates for bio-logical control of Aceria guerreronis. Exp Appl Acarol 34:49–61.
Lawson-Balagbo ML, Gondim Jr MGC, Moraes GJ, Hanna R, Schausberger P (2008)Exploration of acarine fauna on coconut palm in Brazil with emphasis on Aceria guerreronis(Acari: Eriophyidae) and its natural enemies. Bull Entomol Res 98:83–96.
Lindquist EE, Evans GO (1965) Taxonomic concept in the Ascidae, with a modified setal nom-enclature for the idiosoma of the Gamasina (Acari: Mesostigmata). Mem Entomol Soc Can47:1–64.
Lindquist EE, Sabelis MW, Bruin J (eds.) (1996) Eriophyoid Mites - Their Biology, Natural Enemies andControl. World Crop Pest Series Vol. 6, Elsevier Science Publishers, Amsterdam, TheNetherlands, 790 + xxxii pp.
Mayr E (1940) Speciation phenomena in birds. Am Nat 74:249–278.Mayr E (1963) Animal Species and Evolution. Harvard University Press, Gambridge, MA.McMurtry JA (1980) Biosystematics of three taxa in the Amblyseius finlandicus group from South
Africa, with comparative life history studies (Acari: Phytoseiidae). Int J Acarol 6:147–156.McMurtry JA, Huffaker CB, Van de Vrie M (1970) Ecology of tetranychid mites and their natu-
ral enemies: A review: I. Tetranychidae enemies: their biological characters and the impactof spray practices. Hilgardia 40:331–390.
McMurtry JA, Mahr DL, Johnson HG (1976) Geographic races in the predaceous mite,Amblyseius potentillae (Acari: Phytoseiidae). Int J Acarol 2:23–28.
Mercot H, Llorente B, Jacques M, Atlas A, Montchamp-Moreau C (1995) Variability within theSeychelles cytoplasmic incompatibility system in Drosophila simulans. Genetics 141:1015–1023.
Monetti LN, Croft BA (1997) Mating cross-mating and related behaviors of Neoseiulus californicusand Neoseiulus fallacies (Acari: Phytoseiidae). Exp Appl Acarol 21: 67–74.
Moore D, Alexander L (1987) Stem injection of vamidothion for control of coconut mite,Eriophyes guerreronis Keifer in St Lucia. Crop Protection 6:329–333.
Moore D, Alexander L, Hall RA (1989) The coconut mite, Eriophyes guerreronis Keifer in St. Lucia:yield losses and attempts to control it with acaricide, polybutene and Hirsutella fungus. TropPest Manag 35:83–98.
Moraes GJ, Lopes PC, Fernando LCP (2004) Phytoseiid mites (Acari: Phytoseiidae) of coconutgrowing areas in Sri Lanka, with description of 3 new species. J Acarol Soc Jpn 13(2):141–160.
Moraes GJ, McMurtry JA (1981) Biology of Amblyseius citrifolius (Denmark and Muma) (Acarina –Phytoseiidae). Hilgardia 49:1–29.
Muma MH, Denmark HA (1969) Sibling species of Phytoseiidae (Acari: Mesostigmata). FlaEntomol 52:67–72.
29
Morphological variation and reproductive incompatibility
Nazer-chap2.qxd 14-9-2012 10:35 Page 29

Navia D, de Moraes GJ, Roderick G, Navajas M (2005) The invasive coconut mite Aceria guerrero-nis (Acari: Eriophyidae): origin and invasion sources inferred from mitochondrial (16S) andnuclear sequence. Bull Entomol Res 95:105–116.
Negloh K, Hanna R, Schausberger P (2010) Season- and fruit age-dependent population dynam-ics of Aceria guerreronis and its associated predatory mite Neoseiulus paspalivorus on coconut inBenin. Biol Contr 54: 349–358.
Noronha ACS, Moraes GJ (2002) Variaçãoes morfológicas intra e interpopulacionaes de Euseius citrifoliusDanmark & Muma e Euseius concordis Chant (Acari, Phytoseiidae). Revta Bras Zool 19(4):1111–1122.
Noronha ACS, Moraes, GJ (2004) Reproductive compatibility between mite populations previous-ly identified as Euseius concordis (Acari: Phytoseiidae). Exp Appl Acarol 32:271–279.
O’Neill SL, Karr TL (1990) Bidirectional incompatibility between conspecific populations ofDrosophila simulans. Nature 348:178–180.
Reis AC, Gondim MGC, Moraes GJ, Hanna R, Schausberger P, Lawson-Balago LE, Barros R(2008) Population dynamics of Aceria guerreronis Keifer (Acari: Eriophyidae) and associatedpredators on coconut fruits in Northeastern Brazil. Neotrop Entomol 37(4):457–462.
Ros VID, Breeuwer JAJ (2009) The effects of, and interactions between, Cardinium and Wolbachiain the doubly infected spider mite Bryobia sarothamni. Heredity 102:413–422.
Rowell HJ, Chant DA, Hansel RIC (1978) The discrimination of setal homologies and setal patternson the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can Entomol 110:859-876.
Sabelis MW, Van Rijn PCJ (1997) Predation by insects and mites. In: T. Lewis (Ed.) Thrips as CropPests. CAB-International, pp. 259–354.
SAS (2003) SAS Institute version 9.1 Inc., Cary, NC, USA.Thompson JN (1987) Symbiont-induced speciation. Biol J Linn Soc 32:385–393.Tixier M-S, Guichou S, Kreiter S (2008) Morphological variation of the species Neoseiulus califor-
nicus (McGregor) (Acari: Phytoseiidae): importance for diagnostic reliability and synonymies.Invert Syst 22:453–469.
Tixier M-S, Kreiter S, Barbar Z, Ragusa S, Cheval B (2006a) The status of two cryptic species:Typhlodromus exhilaratus Ragusa and Typhlodromus phialatus Athias-Henriot (Acari: Phytosei-idae): consequences for taxonomy. Zool Scripta 35:115–122.
Tixier M-S, Kreiter S, Ferragut F, Cheval B (2006b) The suspected synonymy Kampimodromus hmim-inai and Kampimodromus adrianae (Acari: Phytoseiidae): morphological and molecular investi-gations. Can J Zool 84:1216–1222.
Tixier M-S, Kreiter S, Cheval B, Auger P (2003) Morphometric variation between populations ofKampimodromus aberrans Oudemans (Acari: Phytoseiidae). Implications for the taxonomy ofthe genus. Invert Syst 17(2):349–358.
Tixier M-S, Kreiter S, Croft B A, Cheval B (2004) Morphological and molecular differences in thegenus Kampimodromus Nesbitt. Implications for taxonomy. Phytophaga 14:361–375.
Turelli M, HofFmann AA (1995) Cytoplasmic incompatibility in Drosophila simulans: dynamicsand parameters estimates from natural populations. Genetics 140:1319–1338.
Vala F, Breeuwer JAJ, Sabelis MW (2003) No variation for Wolbachia-induced hybrid breakdownin two populations of spider mite. Exp Appl Acarol 29:1–12.
Vala F, Weeks A, Claessen D, Breeuwer JAJ, Sabelis MW (2002) Within- and between-populationvariation in Wolbachia-induced reproductive incompatibility. Evolution 56:1331–1339.
Vala F, Breeuwer JAJ, Sabelis MW (2000) Wolbachia-induced ‘hybrid breakdown’ in the two-spot-ted spider mite Tetranychus urticae Koch. Proc R Soc Lond B 267:1931–1937.
Wu CI, Davis AW (1993) Evolution of postmating reproductive isolation: the composite natureof Haldane’s rule and its genetic bases. Am Nat 142:187–212.
Zannou ID, Moraes GJ, Ueckermann EA, Olivera AR, Yanineck JS, Hanna R (2006) Phytoseiidsmites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from Sub-Saharan Africa. Int JAcarol 32: 241–276.
CHAPTER 2
30
Nazer-chap2.qxd 14-9-2012 10:35 Page 30

31
Endosymbiont Wolbachia and Cardinium bacteria in the predatory mite Neoseiulus ppaspalivorus
ABSTRACT – Whereas endosymbiont-induced incompatibility is known to occur invarious arthropod taxa, such as spider mites, insects and isopods, it has been rarelyreported in plant-inhabiting predatory mites (Acari: Phytoseiidae). Recent cross-breeding studies with the phytoseiid mite Neoseiulus paspalivorus DeLeon revealed acomplete post-mating reproductive isolation between specimens collected fromthree geographic origins – Northeast Brazil (South America), Benin and Ghana(West Africa) – even though they are morphologically very similar. Here, we car-ried out a study to assess to what extent these populations exhibit genetic differ-ences and whether endosymbionts are involved in the incompatibility. First, weused the mitochondrial cytochrome oxidase I (COI) gene to assess genetic diver-sity among geographic populations. Second, we used a PCR-based method tocheck for the presence of Wolbachia and/or Cardinium in these mites, and we deter-mined their phylogenetic relationships using specific primers for Wolbachia andCardinium 16S rDNA genes. Third, we conducted a test using an antibiotic(Tetracycline) in an attempt to eliminate the symbionts and to evaluate their effectson the reproductive compatibility of their host. Based on the DNA sequences oftheir COI genes, specimens of the three populations appear to be genetically vir-tually identical. However, the 16S rDNA genes sequences of their associatedendosymbionts differed between the populations: the Benin and Brazil popula-tions harbour different strains of Wolbachia symbionts, while the Ghana populationharbours Cardinium symbionts. In response to antibiotic treatment females of eachof the three populations became incompatible with untreated males of their ownpopulation, similar to that observed in crossings between females from one geo-graphic population and males from another. We conclude that the three geograph-ic populations of N. paspalivorus used in our study represent a single species com-posed of genetically isolated populations, and that their associated bacterial sym-bionts are likely to be the cause of the post-mating reproductive isolation previ-ously observed among the three geographic populations. This insight is relevant tobiological control of coconut mites for which N. paspalivorus is an effective preda-tor, because introducing one geographic strain into the population of another (e.g.,in field releases or mass cultures) may cause population growth depression.
Unpublished manuscript
In the two past decades, our knowledge of the endosymbiotic bacteria – Wolbachia andCardinium – has considerably increased, as well as our understanding of reproductive
abnormalities they cause in their hosts, thus opening avenues for new research (Duron
3Nazer-chap3.qxd 14-9-2012 10:36 Page 31

et al. 2008). Yet, little attention has been paid to the occurrence of these endosymbioticbacteria in natural enemies widely used as biological control agents and the consequencethis may have on the outcome of classical biological control operations. For example,releasing an incompatible strain in the habitat of another may have consequences for theoutcome of the biological control (Stouthamer et al. 2000; Zindel et al. 2011).
Predatory mites (Acari: Phytoseiidae) are widely used for biological control ofplant-feeding mites. While there is a plethora of publications on the role of endosym-bionts in plant-feeding mites (i.e., Breeuwer 1997; Vala et al. 2000; Gotoh et al. 2003,2005a,b, 2007), very little is known about the presence and the role of endosymbiontsin plant-inhabiting phytoseiid mites that represent their most important natural enemies(Enigl and Schausberger 2007; Wu and Hoy 2012). In this article, we focus on the preda-tory mite Neoseiulus paspalivorus DeLeon, a candidate enemy for biocontrol of thecoconut mite, Aceria guerreronis Keifer, a key pest of coconut palms (Moraes et al. 2004;Lawson-Balagbo et al. 2008; Reis et al. 2008; Negloh et al. 2011). To better exploit theuse of this phytoseiid mite as biological control agent studies were conducted to deter-mine genetic diversity (cryptic species, biotypes and genotypes) among different popu-lations (McMurtry 2010). Crossing experiments between populations from three geo-graphic regions – Northeast Brazil (South America), Benin and Ghana (West Africa) –yielded no F1 offspring or of only a few sterile males and thus revealed a complete post-mating and bidirectional incompatibility (Famah Sourassou et al. 2011).
It is well known that post-zygotic incompatibility can result from genetic incompat-ibility (Navajas et al. 1999; Uesugi et al. 2003; Gotoh et al. 2005b; Pryke and Griffith2008; Koevoets and Beukeboom 2009), or from cytoplasmic incompatibilities due tointracellular bacteria such as Wolbachia and Cardinium (O’Neill and Karr 1990; Breeuwer1997; Gotoh et al. 2007). These two bacterial symbionts infect a wide range of arthro-pods and are known to cause reproductive abnormalities in their host, including cyto-plasmic incompatibility, feminization, parthenogenesis, hybrid breakdown and malekilling (Rigaud et al. 1997; Hoffmann and Turelli 1997; Hurst et al. 1999; Vala et al. 2000;Charlat et al. 2001). Cytoplasmic incompatibility, the most common effect of Wolbachiaand Cardinium infections, is a sterility phenomenon that has been described first ininsects and mites (Wade and Stevens 1985; Hoffmann et al. 1997; Hoffmann and Turelli1988; O’Neill 1989; Breeuwer and Werren 1990). Cytoplasmic incompatibility can beeither unidirectional or bidirectional. In the former, crosses between populations areincompatible in one direction, but in the reciprocal cross mating is fully compatible, asare matings between infected (by the same strain) individuals. In contrast, bidirectionalincompatibility usually occurs when the male and female carry different strains ofWolbachia (Laven 1959, 1967; Breeuwer and Werren 1990; Bordenstein et al. 2001, 2003;Telschow et al. 2005, 2007). Incompatibilities have been reported in crosses betweenpopulations of some phytoseiid species, but with unidirectional incompatibilities being
32
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 10:36 Page 32

most commonly observed (Hoy and Cave 1988; Johanowicz and Hoy 1998; Noronhaand Moraes 2002, 2004; Wu and Hoy 2012). However, the role of endosymbionts incausing these incompatibilities is still unclear, except in one case (Metaseiulus occidentalis;Hoy and Cave 1988; Wu and Hoy 2012).
In this article, our aim is to determine to what extent these populations exhibit genet-ic differences and whether endosymbionts are involved in the reproductive isolationbetween three geographic populations morphologically identified as N. paspalivorus (FamahSourassou et al. 2011). First, we assessed genetic diversity between the three populationsbased on the variability in the mitochondrial cytochrome oxidase I (COI) gene. Second,we used the 16S ribosomal DNA primers to detect the presence of the endosymbioticbacteria, Wolbachia and Cardinium, in the three populations and to analyse the phylogenet-ic relationships of these bacteria. Third, we performed an antibiotic test to determinewhether each of these bacteria is associated with cytoplasmic incompatibility in the host.
Material and methods
Populations studied and laboratory rearingThe three laboratory-reared populations – from Benin, Ghana and Brazil – used in thepresent study were the same as those used by Famah Sourassou et al. (2011). In addi-tion, specimens collected from weeds in Benin were included in genetic analyses forcomparative reasons. Details on the locality and date of collection, host plant and thenumber of colonizing females are listed in Table 3.1. The various populations weremaintained in rearing units consisting of a black PVC tile (4 × 4 × 0.1 cm) placed ontop of a foam pad (4 × 4 × 1 cm) resting in a Petri dish (14.5 cm diameter and 1 cmhigh). The edges of the tile (and part of the foam pad) were covered with a band of tis-sue paper. To prevent mites from escaping, distilled water was supplied to the Petri dishon a daily basis to keep the foam pad and the tissue paper wet. A small tuft ofhydrophobic cotton wool covered by a piece of transparent plastic was placed in thecenter of each rearing unit to serve as a resting place and as an oviposition substrate for
33
Endosymbiont bacteria in Neoseiulus paspalivorus
Table 3.1 Collection localities, host plant and date of collection of four geographic populationsof Neoseiulus paspalivorus.
No.Country Locality Host plant Latitude Longitude Collection date colonizing
femalesBenin Ouidah Cocos nucifera 06°21’66N 02°09’76E October 2005 52Benin Ouidah Cyperus esculentus 06°21’66N 02°09’76E October 2010 -Ghana Winneba Cocos nucifera 05°22’90N 00°38’68W December 2008 11Brazil Itamaraca Cocos nucifera 07°46’05N 34°52’08W September 2006 5
Nazer-chap3.qxd 14-9-2012 10:36 Page 33

the predators. All colonies were maintained in a room at 25-27 ºC, 70-90% relativehumidity and L12:D12 photoperiod. At three-day intervals the colonies were providedwith a fresh supply of prey consisting of eggs of Tetranychus urticae Koch and inevitablyalso the immatures that emerge from them.
Genetic analysis Two or three adult females of each colony were used for molecular analysis. DNA wasextracted using the Chelex method (Walsh et al. 1991). Mites were crushed using Precellys24 (Brevet Bertin Technology, France) in a sterile tube containing 100 µl of 5% chelexsolution and 4-5 sterile beads. Tubes were then vortexed, after which 5 µl of proteinaseK (20 mg/ml) was added. Tubes were subsequently incubated at 56 °C for 1 h, heated at95 °C for 8 min, and then briefly centrifuged and stored at -20 °C until later use.
A fragment of the mitochondrial cytochrome oxidase subunit one (COI) wasamplified and sequenced using primers specifically designed for tetranychid mites (5’-TGATTTTTTGGTCACCCAGAAG and 5’-TACAGCTCCTATAGATAAAAC;Navajas et al. 1996). The amplification reaction was carried out in a 25 µl reaction mix-ture containing 11.4 µl of sterile water, 2.5 µl 10X Super Taq Buffer (HTBioTechnology, Cambridge, UK), 5 µl dNTP mix (1 mM), 1.25 µl bovine serum albu-min (10 mg/ml), 1.25 µl MgCl2 (25 mM), 0.2 µl of each primer (10 µM), 0.2 µl of SuperTaq (5U/µL), and 3 µl of DNA template. The PCR amplification (PTC-200 thermalcycler, BioRad), was initiated with a 4-min incubation at 95 °C, followed by 35 cycles,each with 1 min at 94 °C, 1 min at 48 °C and 1 min at 72 °C, and then an extension for4 min at 72 °C. PCR products were purified using the Invitek DNA extraction kit(Invitek, UK) and the sequence was determined by the ABI Prism Big Dye TerminatorSequencing method (Applied Biosystems, The Netherlands). Quality check of sequencechromatograms and construction of contigs were performed with BioEdit version 5.0.6(Hall 1999). Sequences were aligned with ClustalW (as implemented in BioEdit), and aconstruction of the distance matrix and the Neighbour-Joining Tree (with 1000 boot-straps) using the Jukes-Cantor model (Jukes and Cantor 1969) was performed withMEGA® software version 5.
Screening for endosymbiontsDNA was extracted from a pool of 8-10 adult females of each population using thesame method as described above. A PCR-based approach was used to check for thepresence/absence of the two symbionts in each population. For detection of Wolbachiaand Cardinium, the 16S Wolbachia rDNA primers (76F–5’-TTGTAGCCTGCTATG-GTATAACT and 1012R– 5’-GAATAGGTATGATTTTCATGA; O’Neil et al. 1992),and the 16S Cardinium rDNA primers (CLO-f1–5’-GGAACCTTACCTGGGCTA-GAATGTATT and CLO-r–5’-GCCACTGTCTTCAAGCTCTACCAAC; Kurtti et al.
34
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 10:36 Page 34

35
1996) were used. Twenty-five µl PCR reactions were set up using 11.4 µl H2O, 1.25 µlMgCl2 (50 mM), 1.25 bovine serum albumin (10 mg/ml), 2.5 µl PCR buffer, 5 µl dNTPsmix (1 mM each), 0.2 µl each primer (20 µM each) (HT Bio Technology), 0.2 µl Taqpolymerase (5U/µL) and 3 µl of DNA extract. For Cardinium, neither MgCl2 nor bovineserum albumin was added. PCR amplification was carried out in a PTC-200 thermalcycler (BioRad). The thermal profile used for 16S Wolbachia was: 1 min at 94 °C, fol-lowed by 35 cycles each of 40 s at 94 °C, 30 s at 48 °C and 1 min at 72 °C, and 94 °C.For the Cardinium 16S rDNA (CLO-f/r), the thermal conditions were as follow: 2 minat 94 °C, followed by 34 cycles of 40 s at 94 °C, 30 s at 51 °C and 45 s at 72 °C, 5 minat 72 °C, and a final extension for 10 min at 10 °C. PCR products (2 µl) were visualizedon 1% agarose gel stained with ethidium bromide in 0.5X TBE buffer (45 mM Trisbase, 45 mM boric acid, and 1 mM EDTA pH 8.0). Eggs and immatures of T. urticaeused as prey for rearing these populations in our laboratory, as well as all stages of A.guerreronis, the natural prey of the predatory mite under study, were also tested for theoccurrence of endosymbionts. PCR products were purified using the Invitek DNAextraction kit (Invitek, UK) and sequenced along both strands by the ABI Prism BigDye Terminator Sequencing method (Applied Biosystems, The Netherlands). Sequencealignment and construction of contigs were performed using the software BioEdit 5.0.6with ClustalW.
Phylogenetic analysis of the 16S rDNA sequencesA NCBI BLAST search on January 15, 2012 reported our 16S Wolbachia rDNAsequences to be most similar with the existing 16S Wolbachia rDNA sequences in theNCBI database. A high similarity score (98-100%) was observed with the first fifty dis-played 16S Wolbachia rDNA sequences in the NCBI database. Representative Wolbachia16S rDNA sequences were retrieved from the NCBI database. These sequences, in addi-tion to the ones obtained in this study, were automatically aligned by the aid of ClustalW(Thompson et al. 1994), whereupon short entries were discarded resulting in a globalalignment of approximately 695 base-pairs, consisting of 31 sequences retrieved fromthe GenBank and of the three sequences obtained in this study. This final data set of 34sequences was used for phylogenetic analysis.
A BLAST search for the Cardinium 16S rDNA sequence in the NBCI also showeda similarity score of 95-97% with the existing 16S Cardinium rDNA sequences. For thephylogenetic analysis of the 16S Cardinium rDNA sequence obtained in this study, thesame procedure was used, and a global alignment of 395base-pairs was obtained, con-sisting of 27 sequences retrieved from the NBCI database and of the unique sequenceobtained in the present study. Phylogenetic trees based on Maximum Likelihood (ML)method were constructed in MEGA5 using the Juke-Cantor evolution model with 1000bootstrap replications.
Endosymbiont bacteria in Neoseiulus paspalivorus
Nazer-chap3.qxd 14-9-2012 10:36 Page 35

Antibiotics testPresence of Wolbachia or Cardinium is no guarantee that they are the cause of reproduc-tive isolation because some Wolbachia strains do not induce incompatibility in their host(Giordano et al. 1995; Navajas et al. 1999; Gotoh et al. 2005b). Our objective here wasto determine whether the symbionts found are associated with cytoplasmic incompati-bility of their host. Based on the common knowledge that incompatibility occursbetween uninfected females and infected males, we performed a test in which tetracy-cline-treated females from each population were mated with untreated males of thesame population. To obtain virgin females, gravid females were removed from thecolony and confined in a rearing unit as described above. Twenty-four hours later, eggslaid were transferred each to a separate experimental unit and reared to adulthood on adiet consisting of all stages A. guerreronis. To administer tetracycline to females, newexperimental units were placed on a cotton bed soaked with tetracycline solution (0.5%w/v in 10% glucose solution) in Petri dishes similar to the ones described above (10discs per Petri dish). In addition, 3-5 droplets of the tetracycline solution were deposit-ed on the PVC disk representing the experimental unit. The experimental units did notharbour any food source. In this way, each female was maintained on tetracycline for 24hours, thereafter mated with untreated males taken from the same colony as that of thefemale. Female F1 progeny from this cross was used directly in crossing experiments ofBenin and Ghana population. In case of the Brazilian population F1 females were treat-ed once more with antibiotics and mated with males from the infected base population.Progeny from this cross were then used in the crossing experiments. Tetracycline-treat-ed females were PCR-checked for the efficiency of the antibiotic treatment.
To test whether compatibility can be restored in inter-population crosses, bothmales and females of each population were tetracycline-treated for three successive gen-erations using the same procedure as described above. Individuals that were producedafter the last treatment were not treated and kept individually for one week before thecrossing experiments. Inter-population crosses were set up as a single pair matingbetween tetracycline-lines of the Benin and Brazil populations. Each pair was observeddaily and the number of eggs laid was recorded for a period of 10 days. Per cross alleggs produced from all mating pairs were pooled in a single arena.
Results
Genetic analysisWe amplified approximately 500 base pairs of the COI gene from individuals from eachof three N. paspalivorus populations. A 455-bp section was aligned and used for furtheranalyses. Similar nucleotide composition was observed for all populations studied
36
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 10:36 Page 36

(30.7% Adenine, 43.5% Thymine, 14.3% Guanine, and 11.4% Cytosine). A BLASTsearch in the GenBank database showed that the sequences aligned with others COIsequences of Phytoseiidae. The three geographic populations tested varied between 0and 0.2% nucleotide diversity.
Detection of endosymbionts The Benin and Brazil populations were PCR-positive for the 16S Wolbachia rDNA gene(Figure 3.1a), whereas the Ghana population was positive for the 16S Cardinium rDNAgene (Figure 3.1b). This suggests the presence of Wolbachia symbionts in both Benin andBrazil populations, and the presence of Cardinium symbionts in the Ghana population.However, the bands displayed in those populations were always weak compared withthose of positive controls, Bryobia sp. (for Wolbachia) and Brevipalpus sp. (for Cardinium),for each of which a single mite was used to extract DNA. This could be due to low bac-terial densities or inefficient PCR. Moreover, neither Wolbachia nor Cardinium was detect-ed in the prey used for rearing those populations in the laboratory (eggs and immaturesof T. urticae), as in their natural prey (all stages of A. guerreronis) collected from Ouidah,Benin. This rules out the possibility that positive PCRs in the predatory mites was duethe presence of bacteria in their digestive tract.
Phylogenetic relationships of the 16S rDNA genesAfter alignment and trimming of approximately 1000 base pairs of the 16S WolbachiarDNA and 500 base pairs of the Cardinium 16S rDNA gene, the final dataset contained880 bp of the former gene and 450 bp of the latter gene. The nucleotide sequences ofthe 16S Wolbachia rDNA gene amplified from the two Beninese populations – fromweeds and coconut fruits – were 100% identical. However, the nucleotide sequence ofthe 16S Wolbachia rDNA gene obtained from the Brazil population collected fromcoconut fruits differed by 4.5% from those nucleotides of the two Benin populations.
37
Endosymbiont bacteria in Neoseiulus paspalivorus
1 2 3 4 5 6 7 8 9 10
(a) Wolbachia 16S rDNA (b) Cardinium 16S rDNA
~1000
1 8 3 4 1 2 3 4 5 6 7 8 9 10
~1000
~500
(c)
Figure 3.1 PCR assay for Wolbachia and Cardinium infections based on the sequence of the 16SrDNA. Lane L, ladder (>1000 bp in Figure 3.1a, 1000 bp in Figure 3.1b,c); 1, Benin (fromcoconut); 2, Benin (from weeds); 3, Ghana; 4, Brazil; 5, T. urticae eggs; 6, T. urticae immaturestages; 7, A. guerreronis; 8, Bryobia sp. (positive control for Wolbachia); 9, Brevipalpus sp. (positivecontrol for Cardinium); 10, negative control (no DNA); (a) Wolbachia 16S rDNA; (b) Cardinium16S rDNA; (c) Tetracycline-treated individuals from 1, 3 and 4 (8, positive control)
Nazer-chap3.qxd 14-9-2012 10:36 Page 37

The phylogenetic tree based on the 16S Wolbachia sequences indicated that the sym-bionts identified in the Beninese and Brazil populations formed a monophyletic groupwith other 16S rDNA Wolbachia present in the GenBank. Moreover, there was a cleardifferentiation, with 100% bootstrap support, between the 16S Wolbachia rDNAsequences from the two Benin populations together, and that from the Brazil popula-tion (Figure 3.2).
38
CHAPTER 3
Figure 3.2 Phylogenetic tree of 16S rDNA sequences of Wolbachia based on MaximumLikelihood procedure in MEGA5. Numbers on the nods indicate bootstrap values (%, valuesabove 60 are displayed). Wolbachia strains are designated by their host names (in parentheses)and bear GenBank accession number. 1, Benin population collected from coconut; 2, Beninpopulation collected from weeds; 3, Brazil population collected from coconut.
Nazer-chap3.qxd 14-9-2012 10:56 Page 38

The phylogenetic analysis of the 16S Cardinium rDNA clearly indicated that thesymbiont detected in the Ghana population of N. paspalivorus was placed in the samegroup with other Cardinium symbionts present in the GenBank (Figure 3.3). TheCardinium in the Ghana population of N. paspalivorus seemed to be closely related to theCardinium symbionts from other phytoseiid mites.
39
Endosymbiont bacteria in Neoseiulus paspalivorus
Figure 3.3 Phylogenetic tree of 16S rDNA sequences of Cardinium based on MaximumLikelihood procedure in MEGA5. Numbers on the nods indicate bootstrap values (%, valuesabove 60 are displayed). Cardinium strains are designated by their host names (in parentheses)and bear GenBank accession number. Arrow indicates Cardinium sequence obtained in thisstudy.
Nazer-chap3.qxd 14-9-2012 10:36 Page 39

Cytoplasmic incompatibilityBoth the Benin and Brazil populations showed the typical pattern of unidirectionalincompatibility: the cross between uninfected (treated) females and infected males pro-duced no or very few offspring and all other possible cross combinations showed sim-ilar egg production, survival and sex ratios (Table 3.2). However, the incompatibility wasexpressed in different ways. In the Brazil population, incompatibility resulted in very lowfecundity. Only one out of the 13 F2 females tested produced eggs; two eggs during the10 days of observation. In the Benin population, on the other hand, the incompatiblecross produced only slightly, but significant, fewer eggs, but none of these eggs hatched.
Antibiotic treatment of infected females from the Ghana base population yieldedvery few offspring. Possibly the Cardinium is essential for host fitness (Bandi et al. 2001;Vavre et al. 2002; Pannebakker et al. 2007). We therefore decided to use all female off-spring in the cross with infected males, which is the expected incompatible cross. Thiscross produced fewer eggs compared to the infected base population and all eggs shriv-elled and failed to hatch (Table 3.2). Although not all possible cross combination couldbe made, it suggests that the infection in the Ghana population can also cause unidirec-tional incompatibility.
In crossing experiments offspring were used from antibiotic treated mothers. It ispossible that antibiotics may still have unwanted side effects on the fitness of the off-spring. This was not the case since, crosses between uninfected females and males didnot differ in egg production and hatchability of the eggs compared to the (IxI) crossesbetween infected individuals from the base population. A PCR check for Wolbachia andCardinium confirmed that all uninfected females used in the crosses were symbiont-free.
In addition, we carried out crosses between uninfected Brazil and Benin popula-tions. Previous crossing experiments between mites from the Brazil and Benin popula-tions did not produce any offspring (Famah Sourassou et al. 2011). At that time thepresence of Wolbachia in these populations was not known. The crossing experimentsbetween uninfected individuals of the two populations revealed a different pattern(Table 3.3). Crosses between uninfected Brazil females and Benin males were fully com-patible producing on average 1.3 eggs per day with 96% egg hatching rate. The recipro-cal cross between Benin females and Brazil males, however, did not differ from earliercrosses between infected individuals (Famah Sourassou et al. 2011); females producedon average 0.2 eggs per day and all eggs failed to hatch.
DiscussionThe main question addressed in the present study was whether the three reproductive-ly isolated populations identified as N. paspalivorus – from Benin, Ghana and Brazil –represent a single species (Famah Sourassou et al. 2011). The key results of this studydemonstrate that all three geographic populations of N. paspalivorus studied represent a
40
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 10:36 Page 40

single species composed of at least three isolated populations, and that the post-matingreproductive isolation observed between these populations is most likely due to theirassociated endosymbiotic bacteria.
The three populations are morphologically identical (Famah Sourassou et al. 2011)and the genetic dissimilarity between the three populations of based on the mitochon-drial cytochrome oxidase I (COI) gene is low, less than 0.2%, and far less than intra-spe-cific genetic distances reported in other mite species such as Kampimodromus species(1.4%) (Tixier et al. 2008) and Euseius sp. (0-5%) (Okassa et al. 2009). This suggests that
41
Endosymbiont bacteria in Neoseiulus paspalivorus
Table 3.2 Cytoplasmic incompatibility patterns in intra-population crosses between uninfected(U) and infected (I) individuals of Neoseiulus paspalivorus. Note that all eggs per cross werepooled for egg hatch, survival and sex ratio measurements. Percentage survival is individual sur-vival from hatching to adulthood. Sex ratio is the percentage female offspring. Not all crosscombinations within the Ghana population were done because tetracycline-treated females pro-duced very few offspring. Means ± SE followed by different letters are significantly different(SNK test, P<0.05).CROSS n % gravid Fecundity % egg % Sex ratio female × male females (eggs/female/day) hatchability survival (% females)BENINI × I 10 100 1.4 ± 0.02 a 100 100 70U × I 10 100 1.1 ± 0.08 b 0 0 -U × U 10 100 1.2 ± 0.09 b 100 100 52.4I × U 8 100 0.9 ± 0.08 c 100 100 69.4BRAZILI × I 10 100 1.4 ± 0.09 a 100 100 70.8U × I 13 95.4 0.2 ± 0.23 b 100 100 54U × U 10 100 1.7 ± 0.31 a 100 100 58I × U 10 10 1.6 ± 0.06 a 100 100 60GHANAI × I 10 100 1.2 ± 0.07 a 100 100 69.1U × I 10 100 0.7 ± 0.08 b 0 100 -
Table 3.3 Compatibility patterns of crosses between infected (I) and uninfected (U) individualsof the Benin (Be) and Brazil (Br) populations of Neoseiulus paspalivorus. For comparison, crossesbetween infected individuals from the base populations are shown, but were done in a previousstudy (Famah Sourassou et al. 2011). Means ± SE followed by different letters within columnare significantly different (SNK test, P<0.05). Statistical testing only includes crosses from thisstudy.CROSS n % gravid Fecundity % egg % Sex ratio female × male females (eggs/female/day) hatchability survival (% females)UBr × UBe 15 100 1.3 ± 0.12 a 96 100 78.8UBe × UBr 16 100 0.2 ± 0.02 b 0 – –IBr × IBe 30 83 0.5 ± 0.03* 7.6 100 0IBe × IBr 30 93 0.6 ± 0.05* 0 – –*Famah Sourassou et al. (2011)[email protected]
Nazer-chap3.qxd 18-9-2012 2:48 Page 41

they belong to the same biological species N. paspalivorus. Nevertheless, crosses betweenthese populations produced no offspring (Table 3.3; Famah Sourassou et al. 2011).
Pre-zygotic isolation does not appear to explain the absence of offspring in cross-es between these three populations of N. paspalivorus. The proportions of gravidfemales in inter-population matings were as high as the proportions gravid females inthe intra-population matings (Table 3.3; Famah Sourassou et al. 2011). In pseudo-arrhenotokous mites, females only become gravid with eggs after copulation and suc-cessful sperm transfer (see Toyoshima et al. 2000). The presence of gravid females canthus be used as an indication that mating has taken place.
Moreover, the two Benin populations of N. paspalivorus from weeds and coconutshad identical Wolbachia 16S rDNA sequences, whereas the Brazil population had a differ-ent sequence, as supported by 100% bootstrap value in the phylogenetic analyses (Figure3.2). Thus the Benin and Brazil mite populations are infected with different Wolbachiastrains. Both Wolbachia and Cardinium symbionts can cause unidirectional incompatibilityin all three populations of N. paspalivorus (Table 3.2).
Interestingly, the timing of incompatibility differs between these populations. Inthe Wolbachia infected Benin and Cardinium infected Ghana populations, incompatibilitywas expressed early in offspring development resulting in shrivelling of the eggs, where-as in the Wolbachia infected Brazil population, incompatibility resulted in very low fecun-dity (Table 3.2). Host-symbiont interactions can vary as a result of genetic differencesof both host and symbiont (Bordenstein and Werren 2003). The Wolbachia infections inthe Brazil and Benin populations are, at least based on 16S gene, different, but also thetwo mite populations appear to be genetically different, as reciprocal crosses betweenuninfected individuals from both populations did not yield offspring (Table 3.3). Onepossible mechanistic explanation for the difference in timing of the visibility of incom-patibility is that Wolbachia concentrations in the Benin population are lower than in theBrazil population. As a result, incompatibility is less severe in incompatible crosses thatinvolve males with the lower bacterial density (Breeuwer and Werren 1993;Engelstaedter et al. 2007). Regardless of whether Wolbachia concentrations actually dif-fer, host-symbiont interactions can vary due to genetic differences between Wolbachiaand/or hosts (see Engelstaedter and Hurst 2009).
In addition, infected females tend to produce a more female biased sex ratio thanuninfected females in compatible crosses. This was most evident in Wolbachia infectedfemales. An increase in the proportion of male offspring was detected in crosses involv-ing treated females. Female bias sex ratio shift is one of the common effects of Wolbachiainfection (i.e., Breeuwer 1997; Rigaud et al. 1997; Kajeyama et al. 2002). From an evolu-tionary point of view, symbionts that are maternally transmitted are expected to bias sexratios toward the transmitting – female – sex.
42
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 11:00 Page 42

43
Bidirectional incompatibility was observed in reciprocal crosses between mitesfrom the Benin and Brazil populations that are both infected with Wolbachia. Bidi-rectional reproductive incompatibility often occurs in crosses between populations har-boring different Wolbachia strains. Our finding of different bacterial symbionts in thethree populations is consistent with this hypothesis. Compatibility was restored in cross-ings between tetracycline-treated females of the Benin population and tetracycline-treat-ed males of the Brazil population. However, incompatibility could not be restored in thereciprocal cross between tetracycline-treated males from Benin and tetracycline-treatedfemales from Brazil. We have only screened for two symbionts in each population andwe cannot rule out the possibility that another bacterium is involved (Duron et al. 2008;Enigl and Schausberger 2007). This needs further investigation. An alternative but notmutually exclusive explanation is that cyto-nuclear incompatibilities may exist betweenthe Benin nuclear genome and the Brazil cytoplasm. Such cyto-nuclear incompatibilitieshave been reported between closely related species of the parasitoid wasp Nasonia(Breeuwer and Werren 1995; Koevoets et al. 2011), but also between populations of thespider mite Tetranychus quercivorus (Gotoh et al. 2005).
We have only looked at three mite populations, and found that they are infectedwith two types of symbionts that have effects on sex ratio and incompatibility. It wouldbe interesting to determine the infection patterns throughout the distribution range ofN. paspalivorus and determine the nature of host-symbiont interactions.
Although morphologically and genetically (based on COI) similar, geographic pop-ulations of the predatory mite N. paspalivorus have developed reproductive isolation(Famah Sourassou et al. 2011), which is associated with (and perhaps also due to)endosymbiotic bacteria found in them. Evidently, this isolation has not (yet) led to mor-phological differences (as far as they are used for taxonomic classification) and to genet-ic differences at the level of the mitochondria. It is yet to be investigated whether thesepopulations differ in other traits that are relevant to biological control of coconut miteson palm trees. Another practical implication could be that introducing an incompatiblestrain from one geographic area into the habitat of another strain may initially create adepression in population growth due to failures in producing viable offspring. The samemay occur when different strains are mixed up to initiate a mass rearing of the predato-ry mites. These are all good reasons to check for endosymbionts in biocontrol agentsand carefully study their role in reproductive isolation between populations, as well asthe genetic differentiation that is expected to arise from this.
AcknowledgementsMany thanks go to M.G.C. Gondim Jr for supplying the specimens of N. paspalivorus from Brazil,and R. Houndafoche, P. Sovimi, C. Kededji and B. Bovis for their assistance in maintaining thepredator colonies and conducting crossing experiments. This research was supported by theInternational Institute of Tropical Agriculture (IITA) and the University of Amsterdam, The
Endosymbiont bacteria in Neoseiulus paspalivorus
Nazer-chap3.qxd 14-9-2012 11:00 Page 43

44
Netherlands, through a grant from the Board of the Netherlands Foundation for theAdvancement of Tropical Research (WOTRO).
ReferencesBandi C, Dunn AM, Hurst GD, Rigaud T (2001) Inherited microorganisms, sex-specific virulence
and reproductive parasitism. Trends in Parasitology 17:88–94.Bordenstein SR (2003) Symbiosis and the origin of the species. In: Bourtzis K, Miller T, eds. Insect
Symbiosis. CRC Press, Boca Raton, pp. 283–304.Bordenstein SR, O’Hara FP, Werren JH (2001) Wolbachia-induced bidirectional incompatibility
precedes other hybrid incompatibility in Nasonia. Nature 409:707–710.Breeuwer JAJ, Werren JH (1993) Cytoplasmic incompatibility and bacterial density in Nasonia vit-
ripennis. Genetics 135:565-574.Breeuwer JAJ, Werren JH (1995) Hybrid breakdown between two haplodiploid species: the role
of nuclear and cytoplasmic genes. Evolution 49:705–717.Breeuwer JAJ (1997) Wolbachia and cytoplasmic incompatibility in the spider mite Tetranychus urticae
and T. turkestani. Heredity 79:41–47.Breeuwer JAJ, Werren JH (1990) Microorganisms associated with chromosome destruction and
reproductive isolation between two insect species. Nature 346:558–560.Charlat S, Bourtzis K, Mercot H (2001) Wolbachia-Induced cytoplasmic incompatibility. In
Seckbach J (ed) Symbiosis: Mechanism and Model Systems. KAP, The Netherlands, pp. 621–644.Duron O, Bouchon D, Boutin S, Bellamy L, Zhou L, Engelstädter J, Hurst GD (2008) The diver-
sity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol 6: 27.Enigl M, Schausberger P (2007) Incidence of the endosymbionts, Wolbachia, Cardinium and
Spiroplasma in phytoseiid mites and associated preys. Exp Appl Acarol 42:75–85.Engelstaedter J, Hurst GDD (2009) The ecology and evolution of microbes that manipulate host
reproduction. Ann Rev Ecol Evol Syst 40:127-149.Engelstaedter J, Hammerstein P, Hurst GDD (2007) The evolution of endosymbiont density in
doubly infected host species. J Evol Biol 20:685-695.Famah Sourassou N, Hanna R, Zannou I, Moraes GJ, Negloh K, Sabelis MW (2011) Morphological
variation and reproductive incompatibility of three coconut-mite-associated populations of preda-tory mites identified as Neoseiulus paspalivorus (Acari: Phytoseiidae). Exp Appl Acarol 53:323–338.
Giordano R, O’Neill LS, Robertson HM (1995) Wolbachia infections and the expression of cyto-plasmic incompatibility in Drosophila sechellia and D. mauritiana. Genetics 140:1307–1317.
Giordano R, Jackson JJ, Robertson HM (1997) The role of Wolbachia bacteria in reproductive incom-patibilities and hybrid zones of Diabrotica beetles and Gryllus crickets. PNAS 94:11439–11444.
Gotoh T, Noda H, Ito S (2007) Cardinium symbionts cause cytoplasmic incompatibility in spidermites. Heredity 98:13–20.
Gotoh T, Noda H, Fujita T, Iwadate K, Higo Y, Saito S, Ohtsuka S (2005a) Wolbachia and nuclear-nuclear interactions contribute to reproductive incompatibility in the spider mite Panonychusmori (Acari: Tetranychidae). Heredity 94:237–246
Gotoh T, Oku H, Moriya K, Odawara M (2005b) Nucleus-cytoplasm interactions causing repro-ductive incompatibility between two populations of Tetranychus quercivorus Ehara & Gotoh(Acari: Tetranychidae). Heredity 74:105–114.
Gotoh T, Noda H, Hong XY (2003) Wolbachia distribution and cytoplasmic incompatibility basedon a survey of 42 spider mite species (Acari: Tetranychidae) in Japan. Heredity 91:208–216.
Hall TA (1999) BioEdit. A user-friendly biological sequence editor and analysis program forWindows 95/98/NT. Nucleic Acids Symposium Series 41:95-98.
Hoffmann AA, Turelli M (1997) Cytoplasmic incompatibility in insects. In: O’Neil SL, HoffmannAA, Werren JH (eds) Influential Passengers: Inherited Microorganisms and Arthropod Reproduction.Oxford University Press, Oxford, pp 42–80.
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 11:00 Page 44

45
Hoffmann AA, Turelli M (1988) Unidirectional incompatibility in Drosophila simulans: inheritance,geographic variation and fitness effects. Genetics 119:435–444.
Hoy MA, Cave FE (1988) Premating and post-mating isolation among populations of Metaseiulusoccidentalis (Nesbitt) (Acarina: Phytoseiidae). Hilgardia 56:1–20.
Hurst JDD, Jiggins FM, Schulenburg JH, Bertrand D, West SA, Goriceheva II, Zakharov IA,Werren JH, Stouthamer R, Majerus MEN (1999) Male killing Wolbachia in two species ofinsects. Proc R Soc Lond B 266:735–740.
Johanowicz DL, Hoy MA (1998) Experimental induction and termination of non-reciprocalreproductive incompatibility in a parahaploid mite. Entomol Exp Appl 87:51–58.
Jukes TH, Cantor CR (1969) Evolution of protein molecules. Pp. 21–132 In: Munro HN (Ed.)Mammalian Protein Metabolism. Academic Press, NY, USA.
Kajeyama D, Nishimura G, Hoshizaki S, Ishikawa Y (2002) Feminizing Wolbachia in an insect,Ostrinia furnacalis (Lepidoptera: Crambidae). Heredity 88:444–449.
Koevoets T, Niehuis O, van de Zande L, Beukeboom LW (2011) Hybrid incompatibilities in theparasitic wasp genus Nasonia. Heredity 108:302–311.
Koevoets T, Beukeboom LW (2009) Genetics of postzygotic isolation and Haldane’s rule in hap-lodiploid. Heredity 102:16–23.
Kurtti TJ, Munderloh UG, Andreadis TG, Magnarelli LA (1996) Tick cell culture isolation of anintracellular prokaryote from the tick Isodes scapularis. J Invert Path 6:7318–312.
Laven H (1959) Speciation by cytoplasmic isolation in Culex strains. Evolution 5:370–375Laven H (1967) Evolution and speciation in Culex pipientis. In Genetics of Insects Vectors of Disease.
Eds JW Wright R Pal. Holland: Elsvier, pp 251–271.Lawson-Balagbo ML, Gondim MGC, Moraes GJ, Hanna R, Schausberger, P. (2008) Exploration
of acarine fauna on coconut palm in Brazil with emphasis on Aceria guerreronis (Acari:Eriophyidae) and its natural enemies. Bull Entomol Res 98:83–96.
McMurtry JA (2010) Concepts of classification of the Phytoseiidae: Relevance to biological con-trol of mites. In: Trends in Acarology - Proceedings of the 12th International Congress, MW Sabelis,J Bruin (Eds), pp. 339–397. Springer, Dordrecht, The Netherlands.
Moraes GJ, Lopez PC, Fernando LCP (2004) Phytoseiid mites (Acari: Phytoseiidae) of coconutgrowing areas in Sri Lanka, with description of three new species. J Acar Soc Jpn 13:141–160.
Navajas M, Gutierrez J, Lagnel L, Fauvel G, Gotoh T (1999) DNA sequences and cross-breedingexperiments in the hawthorn spider mite Amphitetranychus viennensis reveal high genetic differ-entiation between Japanese and French populations. Entomol Exp Appl 90:113–122.
Navajas M, Gutierrez M, Langnel J (1996) Mitochondrial cytochrome oxidase I in tetranychidmites: a comparison between molecular phylogeny and change of morphological and life his-tory traits. Bull Entomol Res 86:407–417.
Negloh K, Hanna R, Schausberger P (2011) The coconut mite, Aceria guerreronis, in Benin andTanzania: occurrence, damage and associated acarine fauna. Exp Appl Acarol 55:361–374.
Noronha ACS, Moraes GJ (2004) Reproductive compatibility between mite populations previous-ly identified as Euseius concordis (Acari: Phytoseiidae). Exp Appl Acarol 32:271–279.
Noronha ACS, Moraes GJ (2002) Variaçãoes morfológicas intra e interpopulacionaes de Euseius citrifo-lius Denmark & Muma e Euseius concordis Chant (Acari, Phytoseiidae). Rev Bras Zool 19:1111–1122.
O’Neill SL (1989) Cytoplasmic incompatibility in Tribolium confusum. J Invert Pathol 53:132–134.O’Neill SL, Karr TL (1990) Bidirectional incompatibility between conspecific populations of
Drosophila simulans. Nature 348:178–180.Okassa M, Tixier M-S, Cheval B, Kreiter S (2009) Molecular and morphological evidence for new
species status within the genus Euseius (Acari: Phytoseiidae). Can J Zool 87:689–698.Pannebakker BA, Loppin B, Elemans CPH, Humblot L, Vavre F (2007) Parasitic inhibition of cell
death falicitates symbiosis. PNAS 104:213–215.Pryke SR, Griffith SC (2008) Postzygotic genetic incompatibility between sympatric color
morphs. Evolution 637:93–798.
Endosymbiont bacteria in Neoseiulus paspalivorus
Nazer-chap3.qxd 14-9-2012 11:00 Page 45

Reis AC, Gondim MGC, Moraes GJ, Hanna R, Schausberger P, Lawson-Balagbo ML, Barros R(2008) Population dynamics of Aceria guerreronis Keifer (Acari: Eriophyidae) and associatedpredators on coconut fruits in Northeastern Brazil. Neotrop Entomol 37:457–462.
Rigaud T, Juchault P, Mocquard JP (1997) The evolution of sex determination in isopod crus-taceans. BioEssays 19:409–416.
Stouthamer R, Jochemsen P, Platner GR, Pinto JD (2000) Crossing incompatibility betweenTrichogramma minutum and T. platneri (Hymenoptera: Trichogrammatidae): implications forapplications in biological control. Environ Entomol 29:832–837.
Stouthamer R, Breeuwer JAJ, Hurst GDD (1999) Wolbachia pipientis: microbial manipulator ofarthropod reproduction. Ann Rev Microbiol 53:71–102.
Telschow A, Hammerstein P, Werren JH (2005) The effects of Wolbachia versus genetic incompat-ibilities on reinforcement and speciation. Evolution 59:1607–1619.
Telschow A, Flor M, Kobayashi Y, Hammerstein P, Werren JH (2007) Wolbachia-induced unidirection-al cytoplasmic incompatibility and speciation: Mainland-Island Model. PLoS ONE 2(8):e701.
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTALW: improving the sensitivity of pro-gressive multiple sequence alignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Research 22:4673–4680.
Tixier M-S, Kreiter S, Croft BA, Cheval B (2008) Kampimodromus aberrans (Acari: Phytoseiidae) fromthe USA: morphological and molecular assessment of its density. Bull Entomol Res 98:125–134.
Toyoshima S, Nakamura M, Nagahama Y, Amano H (2000) Process of egg formation in thefemale body cavity and fertilization in male eggs of Phytoseiulus persimilis (Acari: Phytoseiidae).Exp Appl Acarol 24:441–451.
Uesugi R, Goka K, Osakabe M (2003) Development of genetic differentiation and postzygoticisolation in experimental metapopulations of spider mites. Exp Appl Acarol 31:161–176.
Vala F, Breeuwer JAJ, Sabelis MW (2000) Wolbachia-induced ‘hybrid breakdown’ in the two-spot-ted spider mite Tetranychus urticae Koch. Proc R Soc Lond B 267:1931–1937.
Vavre F, Fleury F, Varaldi J, Fouillet P, Boulétreau M (2002) Infection polymorphism and cytoplas-mic incompatibility in Hymenoptera-Wolbachia associations. Heredity 88:361–365.
Wade MJ, Steven L (1985) Microorganism-mediated reproductive incompatibility in four beetles(Genus Tribolium). Science 227:527–528.
Walsh PS, Metzger DA, Higuchi R (1991) Chelex 100 as medium for simple extraction of DNAfor PCR-based typing for forensic material. Biotechniques 10:506–13.
Werren JH (1998) Wolbachia and speciation. In: Howard D, Berlocher S, eds. Endless Forms: Speciesand Speciation. Oxford University Press, pp 245–260.
Wu K, Hoy, MA (2012) Cardinium is associated with reproductive incompatibility in the predato-ry mite Metaseiulus occidentalis (Acari: Phytoseiidae). J Invert Path 110:359–365.
Zindel R, Gottlieb Y, Aebi A (2011) Arthropod symbioses: a neglected parameter in pest- and dis-ease-control programmes. J Appl Ecol 48:864–872.
46
CHAPTER 3
Nazer-chap3.qxd 14-9-2012 11:00 Page 46

47
Morphological, molecular and cross-breedinganalysis of geographic populations of coconut-mite associated predatory mites identified asNeoseiulus bbaraki: evidence for cryptic species?
ABSTRACT – Surveys were conducted in Brazil, Benin and Tanzania to collectpredatory mites as candidates for control of the coconut mite Aceria guerreronisKeifer, a serious pest of coconut fruits. At all locations surveyed, one of the mostdominant predators on infested coconut fruits was identified as Neoseiulus barakiAthias-Henriot, based on morphological similarity with regard to taxonomicallyrelevant characters. However, scrutiny of our own and published descriptionssuggests that consistent morphological differences may exist between the Beninpopulation and those from the other geographic origins. In this study, we com-bined three methods to assess whether these populations belong to one speciesor a few distinct, yet closely related species. First, multivariate analysis of 32 mor-phological characters showed that the Benin population differed from the otherthree populations. Second, DNA sequence analysis based on the mitochondrialcytochrome oxidase subunit I (COI) showed the same difference between thesepopulations. Third, cross-breeding between populations was unsuccessful in allcombinations. These data provide evidence for the existence of cryptic species.Subsequent morphological research showed that the Benin population can be dis-tinguished from the others by a new character (not included in the multivariateanalysis), viz. the number of teeth on the fixed digit of the female chelicera.
Experimental and Applied Acarology (2012) 57: 15–26
Mites of the family Phytoseiidae are important predators of phytophagous mites inagricultural crops worldwide (McMurtry et al. 1970; Helle and Sabelis 1985;
Lindquist et al. 1996; Sabelis and Van Rijn 1997; McMurtry and Croft 1997; Gerson etal. 2003). Conventionally, morphological characters such as body size, length of dorsalsetae, leg chaetotaxy, shape and size of spermatheca, cheliceral length and dentition havebeen used to describe species in this family (see Chant and McMurtry 1994, 2006, 2007,2009; Tixier et al. 2008). However, since the set of morphological characters is limited,there may still be variation expressed at the genetic, physiological and behavioural levelthat may justify splitting into more species. Such cryptic species or species complexeshave been shown to exist in the Phytoseiidae (Muma and Denmark 1969; Rosen 1978;McMurtry et al. 1985; Monetti and Croft 1997; Tixier et al. 2003, 2004, 2006a, 2008a),
4Nazer-chap4.qxd 14-9-2012 10:36 Page 47

and also in other mite taxa (Krantz and Mellot 1972; Cicolani et al. 1981; Cicolani andDi Sabatino 1991; Bernini et al. 1991; Athias-Binche et al. 1993; Klimov et al. 2004).Hence, molecular and other tools may be helpful for taxonomic identification (Tixier etal. 2009). These techniques have allowed researchers to look beyond morphological sim-ilarities by searching for genetic and other differences to answer taxonomic questions(Navajas et al. 1996, 1998; Hinomoto et al. 2001, 2007; Jeyaprakash and Hoy 2002;Noronha et al. 2003; Ramadan et al. 2004; Klimov et al. 2004; Tixier et al. 2006a, 2010;Okassa et al. 2009, 2010, 2011; Kanouh et al. 2010; Ros and Breeuwer 2010). Here, weaddress the question whether predatory mites collected from different geographic local-ities and previously identified as Neoseiulus baraki Athias-Henriot represent more thanone closely related species. These predatory mites are considered to be potential candi-dates for biological control of the coconut mite Aceria guerreronis Keifer, a serious pestof the coconut palm (Cocos nucifera L.) in Africa, Americas and part of Asia (Moore2000). Whether differences between populations or cryptic species are relevant toachieve improvements in biological control remains to be seen, but to assess its rele-vance accurate identification is required (Moraes 1987). How to define and identifyspecies has been a long-standing debate. The most accepted definition, the biologicalconcept (Mayr 1940), defines a species as a group of organisms capable of interbreed-ing and producing fertile offspring of both sexes and separated from other such groupswith which interbreeding does not normally happen (Mayr 1940; see also Mallet 1995).
In surveys conducted in coconut production areas in Brazil, Benin, Tanzania, andSri Lanka, populations of predatory mites on Aceria-infested coconut fruits were sam-pled and most of the individuals were assigned to N. baraki based on their morpholog-ical similarity (Moraes et al. 2004; Lawson-Balagbo et al. 2008; Reis et al. 2008; Neglohet al. 2011). However, scrutiny of the morphological descriptions published by Moraeset al. (2004) for a Sri Lanka population on coconut and by Zannou et al. (2006) for pop-ulations sampled from weeds in Africa, and observed by one of us (NFS) for Benin andBrazil populations from infested coconuts, revealed morphological differences betweenthe Beninese specimens from coconuts or weeds and those from coconuts at other geo-graphic localities. For instance, the Sri Lankan (as well as the Brazilian; NFS pers obs)specimens had 10-11 teeth on their fixed cheliceral digits (Moraes et al. 2004), whereasBeninese specimens from weeds and coconuts had 7-8 teeth (Zannou et al. 2006; NFS,pers obs). Moreover, demography and diet breadth of the Benin and Brazil populationsdiffered (Negloh et al. 2008; Tixier et al. 2010; Zannou and Hanna 2011). Hence, we car-ried out a multidisciplinary approach to clarify the taxonomic status of these popula-tions by using three methods. First, we used a morphometric approach using multivari-ate analysis, a method applied successfully to solve taxonomic problems in mites (Boyceet al. 1990; Tixier al. 2003, 2004, 2006b, 2008a,b; Akimov et al. 2003; Klimov et al. 2004;Navia et al. 2009; Vidovic et al. 2010; Okassa et al. 2010). Second, we sequenced part of
48
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 48

the mitochondrial cytochrome oxidase subunit I (COI), as widely used for phylogenet-ic studies of Phytoseiidae and Tetranychidae (Navajas et al. 1996, 1998; Toda et al. 2000;Hinomoto et al. 2001, 2007; Navajas and Boursot 2003; Ramadan et al. 2004; Klimov etal. 2004; Tixier et al. 2006a,b; Ros and Breeuwer 2010; Yang et al. 2010). Third, we con-ducted cross-breeding experiments to assess the degree of reproductive isolationbetween the populations under study.
Material and methods
Populations under study and laboratory propagationThe origins of the specimens collected from infested coconut fruits in Brazil, Benin andTanzania are described in Table 4.1. They were propagated in the laboratory using themethod described in detail by Negloh et al. (2008; see also Famah Sourassou et al. 2011).The rearing unit consisted of a black PVC tile (4 × 4 × 0.1 cm) placed on top of a foampad (4 × 4 × 1 cm) resting in a Petri dish (14.5 cm in diameter and 1 cm in height). Theedges of the tile were covered with a band of tissue paper that also contacted the foampad. To prevent mites from escaping, distilled water was supplied to the Petri dish on adaily basis to keep the foam pad and the tissue paper wet. A tuft of hydrophobic cot-ton wool covered by a piece of transparent plastic was placed in the center of each rear-ing unit to serve as oviposition substrate for the predators. All colonies were maintainedin a room at 25-27 ºC, 70-90% relative humidity and a 12-12h light-dark cycle. At three-day intervals the colonies were provided with a fresh supply of prey consisting of imma-ture stages of T. urticae. The lab-reared populations were maintained at least one yearprior to the start of the experiments.
Morphological analysisA total of 30 adult females and 20 adult males of each colony were subject to morpho-metric analysis. Specimens were slide-mounted in Hoyer’s medium and examined withrespect to 32 morphometric characters (see list of characters in Tables 4.2 and 4.3). Inaddition, the number of teeth on the movable and fixed cheliceral digits was assessedfor all female specimens whose chelicerae were in an adequate position (lateral) to allow
49
Is Neoseiulus baraki a cryptic species?
Table 4.1 Collection localities of the specimens of predatory mites identified as Neoseiulus bara-ki studied.Country Locality District/Province/State Latitude LongitudeBenin (West Africa) Gbehoue Department of Atlantique 06°21’S 01°55’ETanzania (Eastern Africa) Nachenjele District of Mtwara 10°26’S 40°08’ETanzania (Eastern Africa) Singino District of Kilwa Masoko 08°78’S 39°41’EBrazil (South America) Itamaraca State of Pernambuco 07°46’S 34°52’W
Nazer-chap4.qxd 14-9-2012 10:36 Page 49

visualization of the teeth (i.e., at least 10 individuals). Observation was also made on theornamentation of the dorsal shield, and the shape of the spermatheca (if female) andthe spermatodactyl (if male). Measurements and counts were done with a phase-con-trast microscope at 40× or 100× magnification. Observations were made on the char-acters considered essential and sufficient for the description of phytoseiid species(Moraes et al. 2004; Zannou et al. 2006; Chant and McMurtry 2005, 2007; Tixier et al.2008a,b). We followed the setal nomenclature of Lindquist and Evans (1965), as adapt-ed by Rowell et al. (1978) and Chant and Yoshida-Shaul (1991) for the phytoseiids.Unfortunately, we did not have specimens from Sri Lanka at our disposal, and thereforewe relied on measurements given by Moraes et al. (2004). To enable comparison we alsorelied on measurements, given by Zannou et al. (2006) for the weed-associated speci-mens from Benin, and the measurements of the holotype (Athias-Henriot 1966). Allmeasurements are given in micrometers.
Morphological characters of males and females were analyzed separately. First, aone-way analysis of variance (Proc ANOVA) followed by Student-Newman-Keuls mul-tiple range comparison tests (α = 0.05) was performed to test differences between pop-ulations for each of the 32 metric characters measured. Second, a discriminant analysiswas conducted to evaluate how many individuals where correctly assigned into theiroriginal group. Third, a canonical discrimination analysis was performed to determinepatterns of morphological variation and to identify morphological characters that con-tribute most to the morphological differentiation between populations. All statisticalanalyses were performed with SAS version 9.2 (SAS Institute 2005).
Crossing experimentsOur preliminary investigations showed that the two Tanzania populations were repro-ductively compatible. Therefore, only one Tanzania population (Nachenjele) was includ-ed in the crossing experiments. For each of the three populations, experiments werestarted with a cohort of eggs, 1-day old since they were laid by 100 females on diet ofT. urticae eggs in rearing units as described above. To rear them to adulthood, these pred-ator eggs were transferred each separately to an experimental unit consisting of 2.5 cmdiameter black PVC disk and provided with all stages of A. guerreronis as prey. Whenadult, each predator was sexed and kept on their experimental unit until it was used forcrossing experiments.
Intra- and inter-population crosses were set up as single pair matings between virginfemales and males in experimental units similar to those described previously. All cross-es were conducted simultaneously and under the same laboratory conditions as where theparental colonies were maintained. Additionally, 10 virgin females of each populationwere kept in isolation, to ascertain that mating is necessary for oviposition (Nguyen andAmano 2010). Ten experimental units from the same cross experiment were put togeth-
50
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 50

51
er into one Petri dish (13.5 cm diameter and 1.7 cm high) filled with water-saturated cot-ton to prevent mites from escaping. Each pair was observed daily and the number ofeggs laid was recorded for a period of 10 days. Eggs laid in arenas with the same combi-nation of geographic origins for male and female were pooled in a rearing unit similar tothat previously described (8.5 cm in diameter and 1.5 cm in height), and reared to adult-hood to determine egg viability, post-embryonic survivorship and sex-ratio.
A single factor ANOVA and Student-Newman-Keuls multiple range test (SAS v. 9.2;SAS Institute 2005) were used to test the effect of crossing combination on fecundity.
Molecular analysisDNA was extracted using the Chelex method (Walsh et al. 1991). This method is sim-ple, fast, cost-effective and appropriate for reducing contamination risks because theentire extraction procedure is performed in a single tube. Per laboratory culture, 2-3individuals were used for molecular analysis. Single female mites were crushed (usingPrecellys 24, Brevet Bertin Technology, France) in a sterile tube containing 100µl of 5%chelex solution and 4-5 sterile beads. Tubes were then vortexed, after which 5µl of pro-teinase K (20 mg/ml) was supplied and centrifuged briefly. Tubes were subsequentlyincubated at 56 °C for 1 h, and then heated at 95 °C for 8 min. Tubes were briefly cen-trifuged and stored at -20 °C until later use.
A fragment of the mitochondrial cytochrome oxidase subunit one (COI) was PCR-amplified and sequenced. The COI primers used were those of Navajas et al. (1996),specifically designed for tetranychid mites (5’-TGATTTTTTGGTCACCCAGAAG and5’-TACAGCTCCTATAGATAAAAC). The amplification reaction was carried out in a25 µl reaction mixture containing 9 µl of sterile water, 2.5 µl 10X Super Taq Buffer (HTBioTechnology, Cambridge, UK), 5 µl dNTP mix (1 mM), 1.25 µl bovine serum albu-min (10mg/ml), 1.25 µl MgCl2 (25 mM), 0.4 µl of each primer (10 µM), 0.2 µl of SuperTaq (5U/µL), and 5 µl of DNA template. The PCR amplification was performed in aPTC-200 thermal cycler (BioRad) programmed as follow: 4 min at 95 °C, 35 cycles – 1min at 94 °C, 1 min at 48 °C, and 1 min at 72 °C –, and 4 min at 72 °C. PCR productswere purified using the Invitek DNA extraction kit (Invitek, UK). Both strands of thepurified DNA were sequenced with the same COI primers as used in the PCR amplifi-cation, using the ABI Prism Big Dye Terminator cycling Sequencing Kit (AppliedBiosystems, Nieuwerkerk a/d IJssel, The Netherlands). Sequence products wereresolved on an ABI 3700 DNA sequencer.
Quality checks of sequence chromatograms and construction of contigs were per-formed with BioEdit version 5.0.6 (Hall 1999). The forward and reverse sequences werepairwise aligned and checked manually to determine the consensus sequence. All fourconsensus sequences were aligned using ClustalW multiple alignment (Thompson et al.1994). Each of the four consensus sequences was checked for the stop codons using the
Is Neoseiulus baraki a cryptic species?
Nazer-chap4.qxd 14-9-2012 10:36 Page 51

invertebrate mitochondrial code (tansl_table=5; Elzanowski and Ostell 2010). Thealigned sequences were used to construct a distance matrix using the Jukes-Cantor model(Jukes and Cantor 1969) (Transition/Transversion rate ~ 1) in MEGA® software v. 5.
Results
Morphological analysisWith respect to females, significant differences were observed between the four popu-lations for most of the characters measured (P<0.05; Table 4.2), with the exception ofthe lengths of setae z2 and Z5 and the size (length, width) of the spermathecal calyx.With respect to males, also small differences were found between the four populations(P<0.05; Table 4.3), but not for the lengths of setae j4, j5, z5, Z1, R1 and the foot andshaft length of the spermatodactyl (Table 4.3). However, for both sexes, the differenceswere small, and the measurements of most characters analyzed did overlap.
According to discriminant analysis, 92.5% of the female specimens and 78.7% of themale specimens of the four populations together were assigned to the population of theirorigin (Table 4.4). The specimens from Benin were most well-defined because no individ-uals from either of the two sexes were misclassified, followed by those from Brazil withonly one misclassified female and no misclassified male. For Tanzania populations, therewere 13.3% misclassifications in females, and 40-45% misclassifications in males.However, these misclassified specimens from the Brazil and the two Tanzania populationswere never assigned to the Benin population, but to the Brazil or Tanzania population.
The results from the discriminant analysis with respect to data on females yieldeda complete separation of two groups (Figure 4.1a), one corresponding to the Beninpopulation, and the other group formed by the remaining populations. With respect tomale specimens, the polygons for the Benin population, the two Tanzania populationstogether and the Brazil population were all non-overlapping (Figure 4.1b). For bothsexes, the Benin population and the Brazil population were the most distant. The lengthof the dorsal shield, the lengths of setae j1, R1, j3, Z4, S4, S5, and the length of che-licerae contributed the most to the morphological differentiation between female spec-imens. In males, the most important characters in differentiating specimens were thelengths of setae j1, j3, Z4, S5, STIV, the length and width of the dorsal shield and theposterior width of the ventrianal shield.
All in all, we conclude that – besides having most of their setae relatively shorter –female specimens from the Benin population differed from those of the other threepopulations mainly by having 2-3 teeth less on their fixed digit. However, no morpho-logical discontinuity could be detected between populations from Tanzania and Brazil.
52
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 52

53
Is Neoseiulus baraki a cryptic species?Ta
ble
4.2
Mea
n,st
anda
rd e
rror
and
rang
e of
32 m
orph
omet
ric c
hara
cter
s mea
sure
d an
d th
e nu
mbe
r of
teet
h on
the
cheli
cera
l dig
its re
cord
edon
fem
ales s
pecim
ens i
dent
ified
as N
eoseiu
lus b
arak
ifro
m fo
ur g
eogr
aphi
c or
igin
s.A
ll m
easu
rem
ents
are
giv
en in
micr
omet
ers.
Gbe
houe
(Ben
in)
Nac
henj
ele (T
anza
nia)
Sing
ino
(Tan
zani
a)Ita
mar
aca
(Bra
zil)
PM
ean
SEM
in–M
axM
ean
SEM
in–M
axM
ean
SEM
in–M
axM
ean
SEM
in–M
ax(α
= 0
.05)
Dor
sal s
hield
leng
th36
1.3
b0.
9335
2–37
036
0.1
b0.
7835
1–36
736
1.8
b0.
8734
8–36
837
3.9
a0.
5136
7–37
70.
0001
Dor
sal s
hield
wid
th15
9.1
c0.
3815
5–16
716
1.8
b0.
5015
8–16
516
1.4
b0.
5215
8–16
816
7.3
a0.
4616
2–17
10.
0001
j110
.0 c
0.21
9–12
12.7
b0.
329–
1513
.1 b
0.20
12–1
513
.9 a
0.28
12–1
60.
0001
j312
.8 c
0.26
9–15
15.9
b0.
3913
–19
16.5
ab0
.29
13–1
917
.2 a
0.29
15–1
90.
0001
j49.
2 b
0.17
7–10
9.3
b0.
146–
99.
6 a
0.10
9–12
10.0
a0.
219–
130.
0025
j59.
8 ab
0.18
9–12
9.2
b0.
176–
109.
4 c
0.10
6–10
10.2
a0.
249–
130.
0004
j611
.3 b
0.29
9–12
11.9
ab0
.24
10–1
312
.3 a
0.23
9–15
12.7
a0.
3010
–15
0.00
34J2
10.7
b0.
289–
1211
.1 b
0.29
10–1
312
.1 a
0.30
9–15
11.4
ab0
.28
9–13
0.00
96J5
9.8
b0.
179–
1210
.3 b
0.26
10–1
310
.2 b
0.24
9–12
12.1
a0.
219–
130.
0001
z29.
6 a
0.18
6–12
10.0
a0.
219–
1210
.1 a
0.23
9–12
10.3
a0.
269–
130.
14z4
11.7
b0.
279–
1213
.5 a
0.26
13–1
613
.4 a
0.36
10–1
613
.4 a
0.32
10–1
50.
0001
z58.
0 b
0.29
6–9
8.3
b0.
286–
99.
6 a
0.10
9–12
10.1
a0.
239–
130.
0001
Z1
12.1
a0.
269–
1511
.3 a
0.32
9–15
11.3
a0.
3610
–15
12.2
a0.
2510
–15
0.03
8Z
418
.7 c
0.17
15–1
922
.3 b
0.28
19–2
523
.2 a
0.38
19–2
822
.7 a
b0.2
122
–25
0.00
01Z
575
.2 a
0.45
69–7
9 75
.7 a
0.43
70–7
975
.8 a
0.47
73–7
976
.1 a
0.31
73–7
90.
57s4
14.2
c0.
2912
–15
15.4
b0.
2013
–16
15.9
b0.
2813
–19
17.2
a0.
3213
–19
0.00
01S2
15.0
c0.
3012
–19
16.4
b0.
3013
–19
15.9
b0.
3513
–19
17.5
a0.
2915
–19
0.00
01S4
20.4
c0.
2919
–22
26.8
b0.
2925
–28
25.8
a0.
3022
–28
27.1
a0.
2925
–29
0.00
01S5
19.4
b0.
2019
–22
25.5
a0.
3022
–28
25.7
a0.
3322
–28
25.6
a0.
3022
–28
0.00
01r3
13.1
c0.
2012
–15
15.7
b0.
3212
–19
15.9
b0.
2313
–19
17.1
a0.
3213
–19
0.00
01R1
12.8
b0.
1412
–15
16.4
a0.
2613
–19
16.3
a0.
3013
–19
16.4
a0.
2613
–19
0.00
01St
IV32
.7 d
0.48
28–3
838
.5 c
0.37
35–4
136
.6 b
0.54
32–4
140
.1 a
0.27
38–4
20.
0001
ST1-
ST3
83.4
b0.
3879
–89
84.2
b0.
4279
–89
84.4
b0.
3882
–88
87.1
a0.
2985
–89
0.00
01ST
2-ST
253
.8 b
0.23
51–5
7 53
.2 b
0.35
51–5
753
.3 b
0.34
51–5
755
.8 a
0.32
54–6
00.
0001
ST5-
ST5
59.1
c0.
4454
–63
61.5
b0.
3957
–66
60.8
b0.
3857
–66
63.7
a 0
.35
60–6
60.
0001
VSW
-AN
T96
.7 c
0.36
92–1
0110
1.0
b0.
3998
–105
100.
7 b
0.60
95–1
0410
3.9
a0.
3610
1–10
80.
0001
VSW
-PO
ST81
.2 c
0.83
63–8
985
.9 a
0.46
82–9
284
.2 b
0.47
79–8
987
.2 a
0.49
82–9
20.
0001
Vent
.Shi
eld le
ngth
116.
5 b
0.47
111–
120
117.
2 b
0.41
114–
120
115.
1 b
0.37
111–
120
122.
0 a
0.54
117–
127
0.00
01Fi
xed
digi
t len
gth
21.9
c0.
1419
–22
23.5
b0.
3622
–28
25.8
a0.
3622
–28
25.1
a0.
3022
–28
0.00
01M
ovab
le di
git l
engt
h27
.5 c
0.26
25–2
830
.1 b
0.29
28–3
230
.6 b
0.27
28–3
232
.1 c
0.29
28–3
50.
0001
Leng
th o
fca
lyx9.
8 a
0.17
9–12
9.9
a0.
209–
1210
.0 a
0.21
9–12
10.0
a0.
219–
120.
87D
iamet
er o
fca
lyx3.
8 a
0.23
3–6
3.7
a0.
203–
63.
5 a
0.17
3–6
3.7
a0.
213–
60.
76N
o.te
eth
on fi
xed
digi
t7-
810
-11
10-1
110
-11
No.
teet
h on
mov
.dig
it2
22
2V
SW-A
NT,
wid
th o
fth
e ve
ntria
nal s
hield
at t
he le
vel o
fZ
V2;
VSW
-PO
ST,w
idth
of
the
vent
riana
l shi
eld a
t the
leve
l of
anus
;mea
ns w
ithin
aro
w w
ith d
iffer
ent l
ette
rs a
re si
gnifi
cant
ly di
ffere
nt (S
NK
,P<
0.05
)
Nazer-chap4.qxd 14-9-2012 10:36 Page 53

54
CHAPTER 4
Tabl
e 4.
2Co
ntin
ued.
Sri L
anka
*Be
nin-
Buru
ndi-K
enya
**H
olot
ype
Mea
nM
in–M
axM
ean
Min
–Max
(Ath
ias-H
enrio
t 196
6)D
orsa
l shi
eld le
ngth
350
330–
375
353
336–
365
337
Dor
sal s
hield
wid
th16
014
6–17
515
714
2–16
815
0j1
1413
–16
1513
–19
13j3
1713
–19
1513
–18
15j4
119–
129
8–10
11j5
1110
–18
108–
1111
j613
11–1
511
8–14
13J2
1210
–13
1311
–14
13J5
1110
–13
108–
1111
z211
10–1
311
6–16
10z4
1410
–15
1311
–16
13z5
109–
159
8–10
10Z
112
11–1
413
11–1
613
Z4
2119
–25
1811
–24
20Z
569
60–7
361
53–6
777
s416
14–1
714
13–1
815
S215
14–1
616
11–2
415
S425
23–2
720
16–2
920
S525
21–2
825
19–3
020
r315
13–1
615
11–1
816
R113
12–1
512
11–1
410
St IV
3831
–42
3429
–40
35ST
1-ST
383
78–8
780
75–8
580
ST2-
ST2
5149
– 53
54
51–5
952
ST5-
ST5
5855
–60
5956
–62
56V
SW-A
NT
8771
–118
8782
–93
92V
SW-P
OST
8373
–95
8677
–99
87V
SL11
288
–120
117
107–
125
112
Fixe
d di
git l
engt
h31
30–3
325
25–3
6–
Mov
able
digi
t len
gth
2927
–31
2121
–22
–Le
ngth
of
calyx
109–
1112
11–1
210
Diam
eter
of
calyx
22–
33
3–3
2N
o.te
eth
on fi
xed
digi
t10
-11
7–
No.
teet
h on
mov
.dig
it2
1–
*Spe
cimen
s fro
m S
ri La
nka
on c
ocon
ut,m
easu
rem
ents
take
n fr
om M
orae
s et a
l.(2
004)
;**S
pecim
ens f
rom
Ben
in o
n w
eeds
,mea
sure
men
tsta
ken
from
Zan
nou
et a
l.(2
006)
V
SW-A
NT,
wid
th o
fth
e ve
ntria
nal s
hield
at t
he le
vel o
fZ
V2;
VSW
-PO
ST,w
idth
of
the
vent
riana
l shi
eld a
t the
leve
l of
anus
;mea
ns w
ithin
aro
w w
ith d
iffer
ent l
ette
rs a
re si
gnifi
cant
ly di
ffere
nt (S
NK
,P<
0.05
)
Nazer-chap4.qxd 14-9-2012 10:36 Page 54

55
Is Neoseiulus baraki a cryptic species?Ta
ble
4.3
Mea
n,st
anda
rd e
rror
and
rang
e of
32 m
orph
omet
ric c
hara
cter
s mea
sure
d on
male
spec
imen
s ide
ntifi
ed a
s Neos
eiulus
bar
akif
rom
four
geog
raph
ic or
igin
s.A
ll m
easu
rem
ents
are
giv
en in
micr
omet
ers.
Gbe
houe
(Ben
in)
Nac
henj
ele (T
anza
nia)
Sing
ino
(Tan
zani
a)Ita
mar
aca
(Bra
zil)
PM
ean
SEM
in–M
axM
ean
SEM
in–M
axM
ean
SEM
in–M
axM
ean
SEM
in–M
ax(α
= 0
.05)
Dor
sal s
hield
leng
th27
5.9
c0.
8426
8–28
227
6.2
c0.
8326
9–28
227
9.1
b1.
0127
3–28
528
8.1
a0.
9128
2–29
50.
0001
Dor
sal s
hield
wid
th14
1.5
c0.
4113
6–14
314
1.5
c0.
6613
3–14
514
5.2
b0.
9613
9–15
514
8.5
a0.
5214
6–15
20.
0001
j19.
3 c
0.15
6–10
10.6
b0.
419–
1511
.1 b
0.36
9–13
12.5
a0.
279–
160.
0001
j311
.2 c
0.36
9–13
13.4
b0.
5510
–15
13.8
b0.
4710
–16
16.2
a0.
3913
–19
0.00
01j4
8.4
a0.
346–
99.
3 a
0.15
6–10
8.4
a0.
346–
98.
5 a
0.33
6–9
0.33
j58.
5 a
0.33
6–9
9.0
a0.
256–
109.
2 a
0.22
6–10
9.0
a0.
256–
100.
64j6
9.2
b0.
216–
1010
.4 a
b0.3
310
–13
10.3
a0.
319–
129.
9 a
0.25
9–12
0.01
2J2
9.0
b0.
256–
99.
9 a
0.25
10–1
310
.4 a
0.33
10–1
39.
6 ab
0.15
9–12
0.01
9J5
9.0
a0.
256–
108.
4 ab
0.34
6–9
7.6
b0.
356–
99.
3 a
0.15
6–9
0.00
03z2
8.5
b0.
336–
98.
7 b
0.31
6–9
9.5
a0.
226–
129.
8 a
0.21
9–12
0.00
40z4
10.1
c0.
299–
1211
.4 a
b0.3
59–
1312
.3 a
0.39
10–1
510
.6 b
c0.3
49–
130.
0001
z56.
8 a
0.25
6–9
7.1
a0.
316–
97.
4 a
0.34
6–9
7.7
a0.
366–
90.
20Z
110
.3 a
0.38
7–13
10.6
a0.
349–
1310
.7 a
0.35
9–12
9.9
a0.
349–
120.
42Z
415
.5 c
0.50
10–1
919
.5 a
0.25
19–2
519
.3 a
0.39
16–2
217
.9 b
0.52
13–2
20.
0001
Z5
56.2
b0.
6051
–63
58.3
a0.
6254
–63
59.7
a0.
4757
–63
59.6
a0.
4957
–63
0.00
01s4
12.2
b0.
2510
–13
15.4
b0.
2013
–16
14.2
b0.
3613
–16
13.6
a0.
3313
–16
0.00
02S2
13.1
b0.
4110
–16
14.2
a0.
3613
–16
14.6
a0.
3513
–16
15.2
a0.
3713
–19
0.00
25S4
16.4
b0.
2915
–19
20.3
a0.
2919
–22
20.6
a0.
3619
–22
19.9
a0.
4616
–22
0.00
01S5
16.0
b0.
2715
–19
19.6
a0.
2919
–22
19.9
a0.
3319
–22
19.3
a0.
2119
–22
0.00
01r3
11.4
b0.
359–
1312
.5 a
0.27
9–15
12.5
a0.
279–
1312
.9 a
0.31
9–15
0.00
45R1
11.8
a0.
319–
1312
.6 a
0.32
9–15
12.3
a0.
319–
1312
.8 a
0.16
12–1
50.
10St
IV26
.4 d
0.34
25–2
831
.7 b
0.45
28–3
529
.8 c
0.62
25–3
534
.5 a
0.45
32–3
80.
0001
ST1-
ST3
69.1
b0.
3766
–72
69.8
b0.
3666
–72
69.7
b0.
3266
–73
72.1
a0.
3170
–73
0.00
01ST
2-ST
240
.7 b
0.25
38–
41
41.0
b0.
3638
–44
41.5
b0.
3138
–44
43.2
a0.
3441
–44
0.00
01ST
5-ST
532
.9 b
0.35
32–3
534
.7 a
0.53
32–3
833
.1 b
0.36
32–3
535
.8 a
0.3
334
–38
0.00
01V
SW-A
NT
122.
8 b
0.91
114–
130
121.
1 b
1.06
111–
133
123.
9 b
0.85
117–
130
126.
6 a
0.96
120–
133
0.00
11V
SW-P
OST
79.2
c0.
8273
–85
79.5
c0.
8267
–85
85.1
b0.
9879
–95
88.7
a0.
5185
–91
0.00
01V
SL10
4.9
b0.
8598
–111
103.
3 b
0.62
98–1
0810
4.1
b0.
4710
1–10
810
9.8
a0.
5310
4–11
40.
0001
Fixe
d di
git l
engt
h13
.1 c
0.25
12–1
614
.7 b
0.41
12–1
916
.1 a
0.39
13–1
915
.7 a
0.15
13–1
60.
0001
Mov
able
digi
t len
gth
18.2
b0.
3116
–19
18.7
ab0
.31
16–2
219
.8 a
0.38
16–2
219
.8 a
0.31
19–2
20.
0014
Sper
mat
odac
tyl_
FL8.
8 a
0.17
6–12
9.2
a0.
216–
108.
8 a
0.29
6–9
9.6
a0.
159–
120.
14Sp
erm
atod
acty
l_SL
3.3
a0.
153–
63.
8 a
0.29
3–6
3.4
a0.
213–
63.
7 a
0.21
3–6
0.43
FL;f
oot l
engt
h;SL
;sha
ft len
gth;
VSL
,ven
trian
al sh
ield
lengt
h;m
eans
with
in a
row
with
diff
eren
t let
ters
are
sign
ifica
ntly
diffe
rent
(SN
K,
P<0.
05)
Nazer-chap4.qxd 14-9-2012 10:36 Page 55

Cross-breeding analysisMating is a prerequisite for oviposition, because none of the virgin females (n = 10)from each of the three populations produced eggs. All single pair matings within pop-ulations produced offspring (Table 4.5). In contrast, in inter-population single pair mat-ings, not all females produced eggs: the percentage of ovipositing females rangedbetween 6 and 100%. Significant differences were observed in fecundity between differ-ent combinations of females and males (F8,109 = 56.7; P<0.0001). The average totalnumber of eggs produced per female in inter-population crosses ranged between 0.06and 4.4 eggs per female over the 10-day observation period, compared with the rangeof 9.4 to 11.8 eggs per female recorded in the intra-population crosses. All eggs pro-duced in the intra-population crosses were viable and gave rise to 59-74% female prog-eny. In contrast, eggs produced in inter-population crosses were not viable: all of theseeggs shriveled and did not hatch, except in crosses between females of the Brazil pop-ulation and males from the Benin population, where the four normal eggs laid gave riseonly to males of which three died just after reaching the adult stage and only one sur-vived much longer (Table 4.5). Back-crosses could not be performed because F1 prog-eny was not produced.
Molecular comparisonsThe length of the amplified COI gene was approximately 480 bp. No differences werefound between forward and reverse sequences of the same amplicon. A final sequencelength of 444 bp was used for further analysis (Figure 4.2). Neither stop codons norindels were found in any of the sequences examined, indicating the absence of nuclear
56
CHAPTER 4
Table 4.4 Classification based on discriminant analysis of 32 morphometric characters evaluat-ed on female and male specimens identified as Neoseiulus baraki from four geographic origins.
% well- Gbehoue Nachenjele Singino Itamaracaclassified*FEMALES
Gbehoue (Benin) 100 30 0 0 0Nachenjele (Tanzania) 86.67 0 26 4 0Singino (Tanzania) 86.67 0 2 26 2Itamaraca (Brazil) 96.67 0 0 1 29Total 92.50 30 28 27 31
MALESGbehoue (Benin) 100 20 1 0 0Nachenjele (Tanzania) 55 1 11 8 0Singino (Tanzania) 60 0 7 12 1Itamaraca (Brazil) 100 0 0 0 20Total 78.75 21 18 20 21*Percentage of individuals correctly classified in their original populations
Nazer-chap4.qxd 14-9-2012 10:36 Page 56

copies of mtDNA (Numts) in our sequences. Sequences were deposited in GenBankunder the following accession numbers: N. baraki from Brazil: JQ609092, N. baraki fromTanzania: JQ609093, N. baraki from Benin: JQ609094. The aligned sequences were sim-ilar in nucleotide composition and A-T rich (26.8% Adenine, 43.9% Thymine). TheBLAST search in Genbank and EMBL data bases showed that all COI sequencesmatched COI sequences of Phytoseiidae, with identity score ranging between 83-86%.The best score was obtained with Phytoseiulus persimilis Athias-Henriot (AccessionNumber GQ222414; Dermauw et al. 2010), followed by N. californicus (AccessionNumber AB500128; Hinomoto et al. 2010), and Euseius sp. (Accession numbersFJ404589; FJ404590; Okassa et al. 2009), and by Kampimodromus sp. (Accession numbersEF372604, EF372605; Tixier et al. 2008b). A genetic dissimilarity of 12.7-13% wasfound between the Benin population and those of Brazil and Tanzania, whereas thegenetic dissimilarity between the populations of Brazil and Tanzania was 0-0.2% (Table4.6).
57
Is Neoseiulus baraki a cryptic species?
Table 4.5 Oviposition and offspring viability of crossings involving males and females of three geo-graphic populations identified as Neoseiulus barakai.CROSS n % No. % % Immature %Female × Male ovipositing eggs/ shriveled egg survivorship female
females female eggs hatchability offspringINTRA-POPULATION CROSSESBrazil × Brazil 15 100 11.8 ± 0.84 a 0.0 100 100 69.0Benin × Benin 15 100 11.7 ± 0.75 a 0.0 100 100 59.3Tanzania × Tanzania 11 100 9.4 ± 1.41 b 0.0 100 100 74.4INTER-POPULATION CROSSESBrazil × Benin 15 53.3 1.0 ± 0.30 d 73.3 25 100 0.0*Benin × Brazil 15 6.0 0.06 ± 0.06 d 100 0.0 – –Brazil × Tanzania 10 100 3.7 ± 0.39 c 100 0.0 – –Tanzania × Brazil 10 90.0 1.7 ± 0.36 d 100 0.0 – –Benin × Tanzania 10 70.0 4.4 ± 0.52 c 100 0.0 – –Tanzania × Benin 10 10.0 0.1 ± 0.11 d 100 0.0 – –*Few male progeny only; Means within a column with different letters are significantly different (SNK,P<0.05)
Table 4.6 Pairwise nucleotide distance among specimens of predatory mites identified asNeoseiulus baraki from four geographic localities based upon 444 bp fragment of the COI.
Gbehoue Nachenjele Singino ItamaracaGbehoue (Benin) -Nachenjele (Tanzania) 0.1266 -Singino (Tanzania) 0.1297 0.0023 -Itamaraca (Brazil) 0.1266 0 0.0023 -
Nazer-chap4.qxd 14-9-2012 10:36 Page 57

DiscussionIn this article on different geographic populations – from Brazil, Benin and Tanzania –of predatory mites previously identified as N. baraki, we present three lines of evidencethat the Benin population should have a taxonomic status different from the other pop-ulations under study. Below we will discuss each of the three lines and we will argue thatthe Benin population is more close to the holotype and should be designated as N. bara-ki. If so, the other populations formerly designated as N. baraki should be re-assignedto a new species.
Multivariate analyses of the 32 metric characters showed a clear separation betweenthe Benin population on coconut and the other populations on coconut in Tanzania (this
58
CHAPTER 4
High value of DSL
High values of j1, STIV, j3, FCD, R1, Z4, S5, S4
(a)
(b)Low values of S4, S5, Z4
High values of j3, j1, DSL, STIV
Figure 4.1 Canonical discrimination analysis of 32 morphometric characters measured onfemale (a) and male (b) populations of predatory mites identified as Neoseiulus baraki from fourgeographic origins: Polygons formed based on the projection of the individuals onto the canon-ical variables 1 and 2. The amount of variation explained by each of the two canonical variablesis shown in parentheses. DSL, dorsal shield length; FCD, fixed cheliceral digit (length).
Nazer-chap4.qxd 14-9-2012 10:36 Page 58

article), Brazil (this article) and Sri Lanka (Moraes et al. 2004). This separation was con-sistent with the number of teeth on the fixed cheliceral digit, which was similar for theBrazilian, Tanzanian and Sri Lankan (Moraes et al. 2004) females, but different for theBeninese females from coconut and weeds (Zannou et al. 2006). These differences can-not be a simple consequence of allometry. The three African populations (one Benineseand two Tanzanian) have similar lengths and widths of the dorsal shield which is a goodmeasure of their size, but the Brazil population stands out as being somewhat larger(Table 4.2). Thus, allometry would predict similar length of setae and smaller number ofteeth for all the African populations. Clearly, this was not the case: only the Benin pop-ulation had relatively shorter setae and fewer teeth on their fixed cheliceral digit.
The problem that now arises is which of these populations (the Beninese or theothers) is closer to the holotype. The species N. baraki was first described by Athias-Henriot (1966) from specimens collected on Phalaris plants in Algeria, and subsequent-ly recorded in China, Puerto Rico, Taiwan, Thailand, Africa (Moraes et al. 2004; Zannouet al. 2006) and more recently in Africa, Brazil and Sri Lanka from coconut palms.Athias-Henriot’s (1966) original description of N. baraki was not sufficiently detailed,based on an insufficient number of specimens (one female) and contained no informa-tion on the cheliceral dentition. Also in the re-description provided by Ehara andBandhoufalck (1977), the number of teeth on the fixed digit was not provided, but theauthors did record 2 teeth on the movable digit, which corresponds to the 2 teethrecorded in the present and previous studies (Moraes et al. 2004). Based on multivariateanalyses of the 32 characters studied, the Benin populations (either from coconut orweeds) are closer to the holotype than are the populations from Brazil, Tanzania and SriLanka (see Figure 4.1). The single tooth on the movable digit recorded from specimens
59
Is Neoseiulus baraki a cryptic species?
Figure 4.2 COI sequences of specimens identified as Neoseiulus baraki from four geographiclocalities. Dots indicate identity with the first sequence.
Nazer-chap4.qxd 14-9-2012 10:36 Page 59

collected from weeds by Zannou et al. (2006) was probably a mistake (see Table 4.2).What is still needed is scrutiny of the number of teeth on the movable cheliceral digitfor the specimens from weeds in Benin, and the number of teeth on the fixed chelicer-al digit for the specimens from the same geographic origin as the holotype.
The molecular analysis of specimens from the four populations studied alsoshowed that the Benin population differed from all others, in line with the morpholog-ical differences. The genetic divergence of the Benin population had a magnitude simi-lar to that observed for phytoseiid species. The genetic distance found in the presentstudy is within the range (11.9-18.4%) reported for different species of the genusKampimodromus Nesbitt (Acari: Phytoseiidae) (Tixier al. 2008b) on the same DNA frag-ment, low compared to those reported for different Euseius species (17%-25%) (Okassaet al. 2009), and similar to that between Tetranychus and Bryobia species (Acari:Tetranychidae) (Navajas et al. 1998; Ros et al. 2008, 2010). However, the genetic distancefound between the Tanzanian and Brazilian populations is far less than the intra-specif-ic genetic distance observed for Kampimodromus species (Tixier et al. 2008b) and withinthe range observed for Euseius sp. (0-5%) (Okassa et al. 2009).
In addition to genetic differences observed, the cross-breeding experimentsshowed that populations studied represent distinct biological entities (biological species,sensu Mayr 1940), as has been observed for several phytoseiid and phytophagous mitespecies (McMurtry et al. 1976; Mahr and McMurtry 1979; Monetti and Croft 1997;Klimov et al. 2004; Noronha and Moraes 2004; Famah Sourassou et al. 2011). There areseveral explanations for the absence of offspring in crosses between populations. Theycan roughly be divided in pre- and post-zygotic isolation mechanisms. Pre-zygotic iso-lation would result in a lack of egg production, since virgin females do not produceeggs. We cannot rule out the possibility of non-mating (Hoy and Cave 1988) because wedid not make these behavioral observations. Nevertheless, the net result would be thatthe populations represent different ‘species’ as a result of pre-zygotic isolation. In caseof post-zygotic isolation, reduced numbers of offspring can be the result of geneticincompatibilities or cytoplasmic incompatibilities due to intracellular bacteria such asWolbachia or Cardinium (O’Neill and Karr 1990; Perrot-Minnot et al. 1996; Breeuwer1997; Gotoh et al. 2007). The incompatibility between the Benin population and thosefrom Brazil and Tanzania could be the result of their genetic divergence, or from mutu-al interactions with reproductive parasites. Whether due to genetic divergence orendosymbionts or a combination of both, reproductive isolation is evident between theBenin population and the other two. Crossing experiments in combination with antibi-otic treatment may help to clarify whether bacteria are the cause of the observed incom-patibilities (Breeuwer 1997).
Based on COI sequences, specimens from Brazil and Tanzania (and possibly thosefrom Sri Lanka because of the close morphological similarity) may belong to the same
60
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 60

species, whereas specimens from Benin may represent a distinct species, the females ofwhich can be separated from those of Brazil and Tanzania based mainly on the numberof teeth on the fixed cheliceral digit. Three cases have been reported earlier where mor-phological analysis and molecular methods, cross-breeding tests, or behaviour studiesled to separation in cryptic species in the Phytoseiidae. First, the original morphologicaldescriptions of Typhlodromus exhilaratus and T. phialatus Athias-Henriot were virtually thesame, but molecular analysis and cross-breeding experiments showed differences thatwere later confirmed by the discovery of a new taxonomic character to separate the twospecies: the shape of the spermatheca (Tixier et al. 2006a). Second, using a combinationof molecular markers and morphological characters, Tixier et al. (2008) showed that thespecies of Kampimodromus they considered in their study could be separated by chelicer-al dentition. Third, the original morphological descriptions of N. cucumeris Oudemansand N. bellinus (Womersley) were similar, but behavioural observations revealed differ-ences, that were later confirmed by the discovery of new taxonomic characters to sep-arate the two species: the number of teeth on the fixed cheliceral digit and the setal andcheliceral lengths (Beard 1999). In the study by Lofego et al. (2009) considerable varia-tion in the number of cheliceral teeth between specimens of N. benjamini Schicha froma same plant and locality (or even between right and left chelicerae of the same individ-ual) and has been reported, but no further studies have been carried out to elucidate thecause of this variation. Other cases where an integrative approach was also used to iden-tify cryptic species (or complex of species) have been reported in other mite taxa. Forexample, Heethoff et al. (2011) used integrative taxonomy, combining morphological,molecular and chemical methods to delineate species in the parthenogeneticTrhypochthonius tectorum complex. Klimov et al. (2004) employed the same methods, asused in the present study (morphological, cross-breeding and molecular) to identify twocryptic species of Sancassania mites, Sancassania salasi Klimov, Lekveishvili & OConnorand S. ochoai Klimov, Lekveishvili & OConnor.
Summarizing the three approaches we applied, the four geographic populations ofN. baraki are similar in most of morphological characters examined, in that differencesobserved are small and the measurements overlap for most of the characters. The mainmorphological difference between the Benin population and the other populations is thenumber of teeth on the fixed cheliceral digit of female specimens. In the morphometricanalysis, the Benin population is distinct from the other three, therefore indicating theexistence of two different morphological groups, which is consistent with the differencesin the number of teeth on the fixed cheliceral digit. This close relationship is also sup-ported by the molecular analysis showing a species-level genetic difference between theBenin population and the three others. Moreover, cross-breeding tests indicate that thepopulations studied represent different biological entities. The species morphologicallymost close to N. baraki (as far as reported in the literature) are N. paspalivorus DeLeon
61
Is Neoseiulus baraki a cryptic species?
Nazer-chap4.qxd 14-9-2012 10:36 Page 61

and N. mumai Denmark (Moraes et al. 2004), which, however, differed from the formerspecies by a number of morphological characters. We conclude that the population fromBenin and those from the other geographic origins represent cryptic species. The deep-er lesson of all these cases of cryptic (or complexes of) species is that an integrativeapproach combining morphological, molecular, behavioural or compatibility level isrequired to distinguish between these taxonomic entities and that – only if these distinc-tions are made – we can begin to assess whether they are relevant to biological control.
AcknowledgementsThe present study which is a part of PhD Thesis of the senior author was supported by theInternational Institute of Tropical Agriculture (IITA) and the University of Amsterdam, TheNetherlands, through a grant from the Board of the Netherlands Foundation for the advance-ment of Tropical Research (WOTRO). We are grateful to Betsie Voetdijk, Peter Kuperus, and LinDong for their availability and assistance during the molecular work. We thank Sam Korie for thestatistical analysis of the data, and R. Houndafoche, P. Sovimi, C. Kededji and B. Bovis for theirassistance in maintaining the predator colonies and conducting experiments. We also thank K.Negloh, A. Onzo and A. Bokonon-Ganta for their useful advices.
ReferencesAkimov IA, Benedyk SV, Zaloznaya LM (2003) Complex analysis of morphological characters of
gamasid mite Varroa destructor (Parasitiformes, Varroidea). Vestnik Zoologii 38(5):57-66.Athias-Binche F, Schwerz HH, Meierhofer I (1993) Phoretic association of Neoseius novus Ouds
(Acari: Uropodina) with Necrphorus spp. (Celeoptera: Silphidae): a case of sympatric specia-tion? Int J Acarol 19:75-86.
Athias-Henriot C (1966) Contribution a l’étude des Amblyseius palearctiques (Acariens anactinotri-ches, Phytoseiidae). Bulletin Scientifique de Bourgogne 24:181-230.
Beard JJ (1999) Taxonomy and biological control: Neoseiulus cucumeris (Acari: Phytoseiidae), a casestudy. Aust J Entomol 38:51–59.
Bernini F, Avanzati AM, Barati M (1991) Biochemical and morphological systematics in the genusAmerus (Acarida: Oribatida). In: Dusbabek F and Bukva V (Eds) Modern Acarology, volume 2.Prague: Academia and The Hague: SPB Academic Publishing, pp. 213-217.
Boyce W, Elliot L, Clark R, Jessup D (1990) Morphometric analysis of the Psoroptes spp. mitesfrom bighorn sheep, mule deer, cattle, and rabbits. J Parasitol 76(6):823–828.
Breeuwer JAJ (1997) Wolbachia and cytoplasmic incompatibility in the spider mite Tetranychus urticaeand T. turkestani. Heredity 79:41–47.
Chant DA, McMurtry JA (1994) A review of the subfamilies Phytoseiinae and Typholodrominae(Acari: Phytoseiidae). In J Acarol 20(4):223–310.
Chant DA, McMurtry JA (2005) Review of the subfamily Amblyseiinae Muma (Acari:Phytoseiidae) Part IV. The tribe Euseiini N. tribe, subtribes Typhlodromalina, N. subtribe,Euseiina N. subtribe and Ricoseiina N. subtribe. Int J Acarol 31:187–222.
Chant DA, McMurtry JA (2006) A review of the subfamily Amblyseiinae Muma (Acari:Phytoseiidae) Part IX. An overview. Int J Acarol 32:125–152.
Chant DA, McMurtry JA (2007) Illustrated Keys and Diagnoses for the Genera and Subgenera of the Phytoseiidaeof the World (Acari: Mesostigmata). Indira Publishing House, West Bloomfield, MI, USA.
Chant DA, Yoshida-Shaul E (1991) Adult ventral setal patterns in the family Phytoseiidae (Acari:Gamasina). Int J Acarol 17(3):187–199.
62
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 62

Cicolani B, Di Sabatino A (1991) Genetic differentiation between Macrocheles penciliger andMacrocheles minervae (Acari: Mesostigmata). In: Dusbabek F and Bukva V (Eds) Modern Acarologyvol. 2. Prague: Academia and The Hague: SPB Academic Publishing, pp. 437-432.
Cicolani B, Bullini APB, Bullini L (1981) Morphological and genetic differentiation betweenMacrocheles glaber and Macrocheles perglaber. Int J Acarol 7:221–224.
Dermauw W, Vanholme B, Tirry L, Van Leeuwen T (2010) Mitochondrial genome analysis of thepredatory mite Phytoseiulus persimilis and a revisit of the Metaseiulus occidentalis mitochondrialgenome. Genome 53(4):285-301.
Ehara S, Bandhoufalck A (1977) Phytoseiid mites of Thailand (Acarina: Mesostigmata). J Fac Educ,Tottori Univ, Nat Sc 27:43–82.
Elzanowski A, Ostell J (2010) The Genetic Codes. Natl Center for Biotechnology Information(NCBI), Bethesda, MD, USA. http://www.ncbi.nlm.nih.gov/Taxonomy/Utils/wprintgc.cgi.
Famah Sourassou N, Hanna R, Zannou I, Moraes GJ, Negloh K, Sabelis MW (2011)Morphological variation and reproductive incompatibility of three coconut-mite-associatedpopulations of predatory mites identified as Neoseiulus paspalivorus (Acari: Phytoseiidae). ExpAppl Acarol 53:323–338.
Gerson U, Smiley RL, Ochoa, R (2003) Mites (Acari) for Pest Control. Blackwell Science, UK, p 539.Gotoh T, Noda H, Ito S (2007) Cardinium symbionts cause cytoplasmic incompatibility in spider
mites. Heredity 98:13–20.Hall TA (1999) BioEdit. A user-friendly biological sequence editor and analysis program for
Windows 95/98/NT. Nucl Acids Symp Ser 41:95-98.Heethoff M, Laumann M, Weigmann G, Raspotnig G (2011) Integrative taxonomy: combining
morphological, molecular and chemical data for species delineation in the parthenogeneticTrhypochthonius tectorum complex (Acari, Oribatida, Trhypochtoniidae). Front Zool 8(1):2.
Helle W, Sabelis MW (1985) Spider Mites: Their Biology, Natural Enemies and Control. Vol. 1B. Elsvier,Amsterdam, The Netherlands.
Hinomoto N, Osakabe M, Gotoh, T, Takafuji A (2001) Phylogenetic analysis of the green and redforms of the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae), inJapan, based on the mitochondrial cytochrome oxidase subunit I sequences. Appl EntomolZool 36(4): 459-564.
Hinomoto N, Tran PD, Pham AT, Le, TBN, Tajima R, Ohashi K, Osakabe M, Takafuji A (2007)Identification of spider mite (Acari: Tetranychidae) by DNA sequence: A case study in NorthVietnam. Int J Acarol 33:53-60.
Hinomoto N, Shintaku T, Amano H (2010) Comparison of genetic diversity among three phyto-seiid mite species in Japan by Mitochondrial DNA analysis. J Acarol Soc Jpn 19(1):1–14.
Hoy MA, Cave FE (1988) Premating and post-mating isolation among populations of Metaseiulusoccidentalis (Nesbitt) (Acarina: Phytoseiidae). Hilgardia 56: 1-20.
Jeyaprakash A and Hoy MA (2002) Mitochondrial 12S rRNA sequences used to design molecularladder assay to identify six commercially available phytoseiids (Acari: Phytoseiidae). BiolControl 25:136–142.
Jukes TH and Cantor CR (1969) Evolution of protein molecules. Pp. 21-132 In: Munro HN (Ed.)Mammalian Protein Metabolism. Academic Press, New York, USA.
Kanouh M, Tixier M-S, Okassa M, Kreiter S (2010) Phylogenetic and biogeographic analysis ofthe genus Phytoseiulus (Acari: Phytoseiidae) Zool Scripta 39(5):450–461.
Klimov PB, Lekveishvili M, Dowling APG, OConnor BM (2004) Multivariate analysis of mor-phological variation in two cryptic species of Sancassania (Acari: Acaridae) from Costa Rica.Ann Entomol Soc Am 97(2):322–345.
Krantz GW, Mellot JL (1972) Study on phytoseiid specificity in Macrocheles mycotrupetes and M.peliotrupetes Krantz and Mellot (Acari: Macrochelidae) associates of geotrupine Scarabicidae.Acarologia 14:317–344.
63
Is Neoseiulus baraki a cryptic species?
Nazer-chap4.qxd 14-9-2012 10:36 Page 63

Lawson-Balagbo ML, Gondim MGC, Moraes GJ, Hanna R, Schausberger P (2008) Explorationof acarine fauna on coconut palm in Brazil with emphasis on Aceria guerreronis (Acari:Eriophyidae) and its natural enemies. Bull Entomol Res 98:83–96.
Lindquist EE, Sabelis MW, Bruin J (eds.) (1996) Eriophyoid Mites - Their Biology, Natural Enemies andControl. WCP Vol. 6, Elsevier Science, Amsterdam, The Netherlands, 790 + xxxii pp.
Lindquist EE, Evans GO (1965) Taxonomic concept in the Ascidae, with a modified setal nomen-clature for the idiosoma of the Gamasina (Acari: Mesostigmata). Mem Entomol Soc Can 47:1–64
Lofego AC, Demite PR, Kishimoto RG, Moraes GJ (2009) Phytoseiid mites on grasses in Brazil(Acarai: Phytoseiidae) Zootaxa 2240:41–59.
Mahr DL, McMurtry JA (1979) Cross-breeding studies involving populations of Typhlodromus citriGarman and McGregor, T. arboreus Chant, and a sibling species of each (Mesostigmata:Phytoseiidae). Int J Acarol 5:155–161.
Mallet J (1995) A species definition for the Modern Synthesis. TREE 10(7):294–299.Mayr E (1940) Speciation phenomena in birds. Am Nat 74:249–278.McMurtry JA, Badii MH, Congdon B D (1985) Studies on a Euseius species complex on avocado
in Mexico and Central America, with a description of a new species (Acari: Phytoseiidae). IntJ Acarol 26:107–116.
McMurtry JA, Mah DL, Johnson HG (1976) Geographic races in the predaceous mite Amblyseiuspotentillae (Acari: Phytoseiidae). Int J Acarol 2:23–28.
McMurtry JA, Croft BA (1997) Life-styles of phytoseiid mites and their roles in biological con-trol. Annu Rev Entomol 42:291–321.
McMurtry JA, Huffaker CB, Van de Vrie M (1970) Ecology of tetranychid mites and their natu-ral enemies: a review. I. Tetranychid enemies: their biological characters and the impact ofspray practices. Hilgardia 40(11):331–390.
Monetti LN, Croft BA (1997) Mating, cross-mating and related behaviours in of Neoseiulus califor-nicus and Neoseiulus fallacis (Acari: Phytoseiidae) Exp Appl Acarol 21:67–74.
Moore D (2000) Non-chemical control of Aceria guerreronis on coconuts. Biocontrol News andInformation 21 (3):83-88.
Moraes GJ (1987) Importance of taxonomy in biological control. Insect Sci Appl 8:841–844.Moraes GJ, Lopez PC, Fernando LCP (2004) Phytoseiid mites (Acari: Phytoseiidae) of coconut grow-
ing areas in Sri Lanka, with description of three new species. J Acarol Soc Jpn 13(2): 141–160.Muma MH, Denmark HA (1969) Sibling species of Phytoseiidae (Acari: Mesostigmata). Fla
Entomol 52(2):67–72.Navajas M, Boursot P (2003) Nuclear ribosomal DNA monophyly versus mitochondrial DNA
polyphyly in two closely related mite species: Influence of life history and molecular drive.Proc R Soc B 270:124–127
Navajas M, Gutierrez M, Langnel J (1996) Mitochondrial cytochrome oxidase I in tetranychidmites: a comparison between molecular phylogeny and change of morphological and life his-tory traits. Bull Entomol Res 86:407–417.
Navajas M, Langnel J, Gutierrez M, Boursot P (1998) Species wide homogeneity of nuclear ribo-somal ITS2 sequences in the spider-mite Tetranychus urticae contrasts with extensive mitochon-drial COI polymorphism. Heredity 87:742–752
Navia D, Moares GJ, Querino RB (2009) Geographic pattern of morphological variation of thecoconut mite, Aceria guerreronis Keifer (Acari: Eriophyidae), using multivariate morphometric.Braz J Biol 69(3): 773–783.
Negloh K, Hanna R, Schausberger P (2011) The coconut mite, Aceria guerreronis, in Benin andTanzania: occurrence, damage and associated acarine fauna. Exp Appl Acarol 55:361-374.
Negloh K, Hanna R, Schausberger P (2008) Comparative demography and diet breadth ofBrazilian and African populations of the predatory mite Neoseiulus baraki, a candidate for bio-logical control of coconut mite. Biol Cont 46:523–531.
64
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 64

Nguyen TTP, Amano H (2010) Sperm reception and egg production of mating-interrupted, sin-gle-mated and multi-mated females of Neoseiulus californicus (McGregor) (Acari: Phytoseiidae)at different temperatures. J Acarol Soc Jpn 19(2):97–106.
Noronha ACS, Moraes GJ (2004) Reproductive incompatibility between mite populations previ-ously identified as Euseius concordis (Acari: Phytoseiidae). Exp Appl Acarol 32:271–279.
Noronha ACS, Mota A, Moraes GJ, Coutinho LL (2003) Caracterização Molecular de Populaçõesde Euseius citrifolis Denmark and Muma e Euseius concordis Chant (Acari: Phytoseiidea) utilizan-do o seqüciamento das Regiões ITS1 e ITS2. Neotrop Entomol 32(4): 591–596.
O’Neil SL, Karr TL (1990) Bidirectional incompatibility between conspecific populations ofDrosophila simulans. Nature 348:178-180.
Okassa M, Tixier M-S, Cheval B, Kreiter S (2009) Molecular and morphological evidence for newspecies status within the genus Euseius (Acari: Phytoseiidae). Can J Zool 87:689–698.
Okassa M, Tixier M-S, Kreiter S (2010) Morphological and molecular diagnostics of Phytoseiuluspersimilis and Phytoseiulus macropilis (Acari: Phytoseiidae). Exp Appl Acarol 52:291–303.
Perrot-Minnot M-J, Guo LR, Werren JH (1996) Single and double infection with Wolbachia in theparasitic wasp Nasonia vitripennis: Effects on compatibility. Genetics 143:961–972.
Ramadan HAI, El-Banhawy EM, Hassan AA, Afia SI (2004) Genetic variation in the predaciousphytoseiid mite, Amblysieus swirskii (Acari: Phytoseiidae): Analysis of specific mitochondrialand nuclear sequences. Arab J Biotech 7(2):189–196.
Reis AC, Gondim MGC, Moraes GJ, Hanna R, Schausberger P, Lawson-Balagbo ML, Barros R(2008) Population dynamics of Aceria guerreronis Keifer (Acari: Eriophyidae) and associatedpredators on coconut fruits in Northeastern Brazil. Neotrop Entomol 37(4):457–462.
Ros VID, Breeuwer JAJ (2010) Spider mite (Acari: Tetranychidae) mitonchondrial COI phyloge-ny reviewed: host plant relationships, phylogeography, reproductive parasites and barcoding.Exp Appl Acarol 49:239–262.
Ros VID, Breeuwer JAJ, Menken SBJ (2008) Origins of asexuality in Bryobia mites (Acari:Tetranychidae). BMC Evol Biol 8:153.
Rosen D (1978) The importance of cryptic species and specific identifications as related to bio-logical control. In: Biosystematics in Agriculture, Vol. II (ed. JA Romberger), pp. 23–25. Wiley,New York, USA.
Rowell HJ, Chant DA, Hansel RIC (1978) The discrimination of setal homologies and setal pat-terns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can Entomol110:859-876.
Sabelis MW, Van Rijn PCJ (1997) Predation by insects and mites. In: T Lewis (ed.) Thrips as CropPests. CAB-International, UK, pp. 259–354.
SAS (2005) SAS Institute, version 9.2 Inc., Cary.Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTALW: improving the sensitivity of pro-
gressive multiple sequence alignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucl Acids Res 22:4673–4680.
Tixier M-S, Ferrero M, Okassa M, Guichou S, Kreiter S (2010) On the specific identity of speci-mens of Phytoseiulus longipes Evans (Mesostigmata: Phytoseiidae) showing different feedingbehaviours: morphological and molecular analyses. Bull Entomol Res 17:1–11.
Tixier M-S, Okassa M, Kreiter S (2009) On the way to the molecular diagnostic of species ofPhytoseiidae. Euraac Newsletter 1+2.
Tixier M-S, Guichou S, Kreiter S (2008a) Morphological variation of the species Neoseiulus califor-nicus (McGregor) (Acari: Phytoseiidae): importance for diagnostic reliability and synonymies.Invert Syst 22:453–469.
Tixier M-S, Kreiter S, Croft BA, Cheval B (2008b) Kampimodromus aberrans (Acari: Phytoseiidae)from the USA: morphological and molecular assessment of its density. Bull Entomol Res98:125–134.
65
Is Neoseiulus baraki a cryptic species?
Nazer-chap4.qxd 14-9-2012 10:36 Page 65

Tixier M-S, Kreiter S, Barbar Z, Ragusa S, Cheval B (2006a) The status of two cryptic species:Typhlodromus exhilaratus Ragusa and Typhlodromus phialatus Athias-Henriot (Acari:Phytoseiidae): consequences for taxonomy. Zool Scripta 35:115–122.
Tixier M-S, Kreiter S, Ferragut F, Cheval B (2006b) The suspected synonymy of Kampimodromushmiminai and Kampimodromus adrianae (Acari: Phytoseiidae): morphological and molecularinvestigations. Can J Zool 84:1216–1222.
Tixier M-S, Kreiter S, Croft BA, Cheval B (2004) Morphological and molecular differences in thegenus Kampimodromus Nesbitt. Implications for taxonomy. Phytophaga 14:361–375.
Tixier M-S, Kreiter S, Cheval B, Auger P (2003) Morphometric variation between populations ofKampimodromus aberrans Oudemans (Acari: Phytoseiidae). Implications for the taxonomy ofthe genus. Invert Syst 17(2):349–358.
Toda S, Osakabe M, Kumazaki S (2000) Interspecific diversity of mitochondrial COI sequence inJapanese Panonychus species (Acari: Tetranychidae). Exp Appl Acarol 24:621–629.
Vidovic B, Stanisavljevic L, Petanovic R (2010) Phenotypic variability in five Aceria spp. (Acari:Prostigmata: Eriophyiddae) inhabiting Cirsium species (Asteracea) in Serbia. Exp Appl Acarol52:169–181.
Walsh PS, Metzger DA, Higuchi R (1991) Chelex 100 as medium for simple extraction of DNAfor PCR-based typing for forensic material. Biotechniques 10(4):506-13.
Yang B, Cai J, Cheng X (2010) Identification of astigmatid mites using ITS2 and COI regions.Parasitol Res 108:497–503.
Zannou ID, Hanna R (2011) Clarifying the identity of Amblyseius swirskii and Amblyseius rykei: arethey two distinct species or two populations of one species? Exp Appl Acarol 53(4):339–347.
Zannou ID, Moraes GJ, Ueckermann EA, Olivera AR, Yaninek JS, Hanna R (2006) Phytoseiidmites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from Sub-Saharan Africa. Int JAcarol 32:241–276.
66
CHAPTER 4
Nazer-chap4.qxd 14-9-2012 10:36 Page 66

67
Females as intraguild predators of males incross-pairing experiments with phytoseiid mites
ABSTRACT – Studies on intraguild interactions between phytoseiid species haveshown that intraguild predation occurs and is most commonly manifested asadult females of one species feeding on juveniles of another. Whether suchintraguild interactions can also occur between adult females of one species andadult males of another, is not known. Herein, we report on intraguild interactionsbetween adults of the two sexes in cross-breeding experiments involving threerelated phytoseiid species (Neoseiulus paspalivorus, N. baraki and N. neobaraki) thatare potential candidates for controlling the coconut mite Aceria guerreronis, a seri-ous pest of coconut palms in tropical countries. For comparative reasons, theexperiments were repeated with larvae instead of males, and with only males oronly females of two different species together. In the presence of an ample sup-ply of prey, females of N. neobaraki never fed on individuals of their own species,yet appeared to be very aggressive against males, as well as larvae of the other twophytoseiid species. They also fed on females of N. paspalivorus, but rarely onfemales of N. baraki. Males of N. neobaraki did not suffer mortality when togeth-er with females of either of the two other phytoseiid species. Males of N. barakidid not suffer predation from females of N. paspalivorus, but males of N. pas-palivorus suffered some mortality (15%) from N. baraki females. Larvae of each ofthe three species were vulnerable to intraguild predation by heterospecificfemales, except for N. neobaraki larvae when together with N. baraki females. Theabsence or presence of intraguild predation is largely explained by the size ratiosof the individuals that were put together: large individuals feed on smaller ones,but never the reverse. For each stage and sex, size declines in the following order:N. neobaraki > N. baraki, > N. paspalivorus. Moreover, for each species, females arelarger than males and males are larger than larvae. Strikingly, however, females didnot kill males and larvae of their own species. We propose that niche competitionbetween related phytoseiid species is not only determined by intraguild predation,but also by the limited availability of males from the intraguild prey because phy-toseiid females being pseudo-arrhenotokous require insemination to produce off-spring of both sexes.
Submitted manuscript
There is a growing awareness that biological control agents can be involved inintraguild interactions. (Rosenheim et al. 1995; Zannou et al. 2005; Negloh et al.
2012). Intraguild predation may interfere with the goals of biological control, especial-ly when a less effective predator feeds on another, resulting in an overall decrease in pre-
5Nazer-chap5.qxd 14-9-2012 10:37 Page 67

dation pressure on the target pest (Rosenheim et al. 1995; Schausberger and Croft 2000;Zannou et al. 2005; Negloh et al. 2012).
Such interactions may be relevant especially when different predator species areused to control a single pest. This is the case with respect to the coconut mite Aceria guer-reronis Keifer, a serious pest of coconut palms in South and Central America, theCaribbean and Africa, because there are three candidate predators of the target prey:Neoseiulus paspalivorus DeLeon, N. baraki Athias-Henriot and N. neobaraki Zannou,Moraes and Oliveira. The first two species were reported from surveys conducted inAfrica, South America and Asia, while the latter was found in Africa only (Moraes et al.2004; Lawson-Balagbo et al. 2008; Negloh et al. 2011; Zannou et al. unpubl. data). Thethree species, particularly N. baraki and N. neobaraki, are morphologically very similar,and can be separated only on the basis of a few morphological characters (Moraes et al.2004; Zannou et al. 2006; Famah Sourassou et al. 2012).
As part of a series of studies on their taxonomic relationships (Moraes et al. 2004;Zannou et al. 2006), we carried out cross-pairing experiments with the aim to assessreproductive barriers between N. baraki and N. neobaraki. Instead of interspecific mat-ing and hybridization, however, we observed that females of one species exhibited hightendency to kill males of the other in all pairings (Famah Sourassou, personal observa-tions). Field surveys conducted in Benin and Tanzania showed that the three species arerarely observed together under the bracts of Aceria-infested coconut fruits, although itis possible to find them sharing the same palm (Negloh et al. 2011; I. Zannou, pers.comm.). This behaviour, together with the spatially segregated distribution of the threephytoseiids species on coconut palms, underscores the need to investigate intraguildinteractions between females and males of the three phytoseiid species. In this article,we report on intraguild predation between females and males in cross-pairing experi-ments with N. neobaraki, N. baraki and N. paspalivorus.
Material and methodsThe predators used originate from colonies of the three species N. neobaraki, N. barakiand N. paspalivorus (see Table 5.1 for information about collection), maintained at 25-27ºC, 70-90% r.h. and L12:D12 h, at the IITA-Benin Acarology Laboratory. Their rear-ing units consisted of a black PVC tile (4 × 4 × 0.1 cm) on top of a foam pad (4 × 4 ×1 cm) in a Petri dish (14.5 cm diameter, 1 cm height). The edges of the tile (and part ofthe foam pad) were covered with a band of tissue paper. To prevent mites from escap-ing, distilled water was supplied to the Petri dish to create a barrier and this was main-tained on a daily basis to keep the foam pad and the tissue paper wet. A small tuft ofhydrophobic cotton wool covered by a piece of transparent plastic was placed in thecenter of each rearing unit to serve as an oviposition site for the predators. All colonieswere provided with a fresh supply of prey consisting of eggs of Tetranychus urticae Koch.
68
CHAPTER 5
Nazer-chap5.qxd 14-9-2012 10:37 Page 68

All possible combinations involving females and males of the three species werecreated in cross-pairing experiments in the presence of A. guerreronis as prey. Followingthe findings from the first set of experiments on the high aggressiveness of N. neobara-ki females, we performed the following treatments (see Table 5.6): a) N. neobarakifemales were paired with N. baraki male and the mortality was recorded for 48 h; b) alldead males were removed and replaced by N. neobaraki males; c) After 48 h, survivingN. neobaraki males were removed and replaced by N. baraki males; d) N. neobarakifemales were paired with two different males, a conspecific and a N. baraki male. Forcomparative reasons, we also examined pairings between males (‘male × male’) andbetween females (‘female × female’) of the three species under study, as well as pairingsbetween females and heterospecific larvae (‘female × larva’). The experiments were con-ducted under laboratory conditions (25-27ºC, 70-90% r.h. and L12:D12 h), in arenasconsisting of 2.5 cm diameter black PVC tiles placed on moist cotton wool in a 14.5 cmdiameter Petri dish. The experiments were replicated with new pairs of individuals for20-40 times. Occurrence of intraguild predation was assessed after 48 h. The number ofheterospecifics (males, female or larvae) killed after 48 h was used as a proxy forintraguild predation.
Occurrence of intraguild predation for each cross-pairing experiment is expressedas a percentage calculated from the ratio of the scored number of pairings yielding indi-viduals (males, or females, or larvae) killed (i.e., the scored number of pairings with deadindividuals) to the total number (N) of pairings for that cross-pairing experiment. Wearranged data in 2 × 2 contingency tables and performed a Chi-square test of independ-ence (using SAS version 9.2, SAS Institute 2005) to test whether the occurrence ofintraguild predation and cross-pairing type (interspecific pairings vs. intraspecific pair-ings) are independent.
69
Females as intraguild predators of males
Table 5.1 Collection localities, host plant and date of collection of predatory mites used in thestudy.Species Country Locality Host plant Latitude Longitude Collection No.
date colonizingfemales
Neoseiulus paspalivorusBenin Ouidah Cocos nucifera 06°21’N 02°09’E Oct 2005 52Ghana Winneba Cocos nucifera 05°22’N 00°38’W Dec 2008 11Brazil Itamaraca Cocos nucifera 07°46’N 34°52’W Sept 2006 5
Neoseiulus barakiBenin Gbehoue Cocos nucifera 06°21’S 01°55’E Feb 2009 4Tanzania Nachenjele Cocos nucifera 08°78’S 39°41’E June 2010 5Brazil Itamaraca Cocos nucifera 07°46’S 34°52’W July 2006 8
Neoseiulus neobarakiBenin Gbehoue Cocos nucifera 06°21’S 01°55’E Nov 2005 11Tanzania Kiparanganda Cocos nucifera 07°07’S 39°12’E Jun 2010 Unknown
Nazer-chap5.qxd 14-9-2012 10:37 Page 69

ResultsIn pairings with heterospecific males of N. baraki and N. paspalivorus, N. neobarakifemales killed all males (100% male mortality; Table 5.2). Moreover, when paired withfemales of the two other species, N. neobaraki females killed all N. paspalivorus females,but only 10% of the N. baraki females (Table 5.3). In pairings with N. paspalivorus males,N. baraki females killed 15% of the males. However, N. paspalivorus females did not killany males of N. baraki. When pairing either only females or only males of the latter twospecies, no mortality was recorded (Tables 5.3 and 5.4). Male mortalities recorded ininterspecific pairings were significantly different from those in intraspecific pairings(control) (Table 5.2), as were female mortalities in interspecific pairings when testedagainst intraspecific pairings (Table 5.3).
70
CHAPTER 5
Table 5.2 Percent mortality in interspecific and intraspecific pairings between females and malesof three phytoseiid species. The 2 × 2 contingency table and Chi-square test are given below.CROSS-PAIRINGS
Female × male n Males killed Females killedINTER-SPECIES PAIRINGS
N. neobaraki × N. paspalivorus 40 100 0N. neobaraki × N. baraki 40 100 0N. baraki × N. paspalivorus 40 15 0N. baraki × N. neobaraki 40 0 0N. paspalivorus × N. baraki 40 0 0N. paspalivorus × N. neobaraki 40 0 0
INTRA-SPECIES PAIRINGSN. paspalivorus × N. paspalivorus 20 0 0N. baraki × N. baraki 20 0 0N. neobaraki × N. neobaraki 20 0 0
CHI-SQUARE TESTCross Dead Alive Total X2 df PIntra-specific pairings 86 154 240 30.1 1 <<0.0001Inter-specific pairings 0 240 240Total 86 394 480
Table 5.3 Percent mortality in intra- and interspecific pairings between females of three phyto-seiid species. The 2 × 2 contingency table and Chi-square test are given below.
n N. paspalivorus N. baraki N. neobarakiN. paspalivorus 20 0N. baraki 20 0 0N. neobaraki 20 100 10 0CHI-SQUARE TEST
Cross Dead Alive Total X2 df PIntra-specific pairings 22 18 40 42.3 1 <<0.0001Intra-specific pairings 0 60 60Total 22 78
Nazer-chap5.qxd 14-9-2012 10:37 Page 70

71
To assess whether the conditions under which we observed intraguild predationamong adults, also allowed for predation by females of one species on larvae of theother, we paired N. neobaraki females with larvae of N. baraki or larvae of N. paspalivorusand found 100% larval mortality within 24 h, in all cases. Also, N. baraki females killedall N. paspalivorus larvae, but they killed none of the N. neobaraki larvae. Finally N. pas-palivorus females killed only few N. neobaraki larvae (15%) and not many N. baraki larvae(35%) (Table 5.5). Larval mortalities recorded in interspecific pairings were significant-ly different from those recorded in control pairings (Table 5.5).
To assess the individual ability of N. neobaraki females to discriminate interspecifi-cally, we carried an additional series of experiments in which we confronted them witheither heterospecific or conspecific males or both in a sequence of 4 experimentalrounds. First, N. neobaraki females paired with N. baraki males killed all heterospecificmales within 48 h (Table 5.6a). Second, after removal of the dead heterospecific males
Females as intraguild predators of males
Table 5.4 Mortality (%) in intra- and interspecific pairings between males of three phytoseiidspecies.
n N. paspalivorus N. baraki N. neobarakiN. paspalivorus 20 0N. baraki 20 0 0N. neobaraki 20 0 0 0
Table 5.5 Mortality (%) in inter- and intraspecific pairings between females and larvae of threephytoseiid species. The 2 × 2 contingency table and Chi-square test are given below.CROSS-PAIRINGS
Female × male n Mortality (%)INTERSPECIFIC PAIRINGS
N. neobaraki × N. paspalivorus 40 100N. neobaraki × N. baraki 40 100N. baraki × N. paspalivorus 40 100N. baraki × N. neobaraki 40 0N. paspalivorus × N. baraki 40 35N. paspalivorus × N. neobaraki 40 15
INTRASPECIFIC PAIRINGSN. neobaraki × N. neobaraki 40 2.5N. baraki × N. baraki 40 0
Table 5.6 Male mortality (%) in a series of experiments involving Neoseiulus neobaraki females (n= 20) on the one hand and conspecific and/or heterospecific (N. baraki) males on the other.
(a) (b) (c) (d)1 male 1 male 1 male 1 male + 1 maleN. baraki N. neobaraki N. baraki N. baraki N. neobaraki
Female N. neobaraki 100 0 100 100 + 0
Nazer-chap5.qxd 14-9-2012 10:37 Page 71

and replacement by conspecific males, none of the latter males was killed (Table 5.6b).Third, after removal of the conspecific males and replacement by heterospecific malesagain, all of the latter males were killed (Table 5.6c). Fourth, when given a simultaneouschoice between a conspecific male and an heterospecific male, all N. baraki males werekilled, whereas all conspecific males survived (Table 5.6d). We conclude that females ofN. neobaraki can discriminate between conspecific and heterospecific males, because theykill only the latter.
DiscussionTo the best of our knowledge (see Croft 1970; Menotti and Croft 1997; McMurtry et al.1976; Hoying and Croft 1977; Mahr and McMurtry 1979; Congdon and McMurtry 1986)our results are the first to show that phytoseiid females can act as intraguild predators ofheterospecific males, whereas they do not kill conspecific males (Table 5.2). No malekilling (or predation) was observed in intraspecific crosses between geographic popula-tions of each of the two species (Famah Sourassou et al. 2011, 2012). In terms of aggres-siveness between species, N. neobaraki was superior over N. baraki and, N. baraki was supe-rior over N. paspalivorus. When paired with a conspecific male and a heterospecific maletogether, each N. neobaraki female discriminated and killed only the heterospecific male(Table 5.6). This implicates species recognition prior to the decision to attack an intraguildprey. Strikingly, we never observed an attempt of males to mount a heterospecific female,let alone to mate with her. Thus, discrimination probably occurs prior to courtship.
We have no reason to suppose that phytoseiid females specifically aim to attack het-erospecific males. This is because they also attack heterospecific females and heterospe-cific larvae (Tables 5.3 and 5.5). Moreover, size differences matter to who is theintraguild predator and who is the intraguild prey (Lucas et al. 1998; Hindayana et al.2001; Sato et al. 2003; Janssen et al. 2007): large individuals feed on smaller ones, butnever the reverse. For each stage and sex, size declines in the following species order:N. neobaraki > N. baraki, > N. paspalivorus (Figure 5.1). Moreover, for each species,females are larger than males and males are larger than larvae. We conclude that the datain Tables 5.2-5.5 largely confirm that the intraguild predator is larger in size than theintraguild prey and that predation of females on heterospecific males is no exception tothis rule. However, we cannot explain why N. baraki females did not prey on N. neobara-ki larvae. This requires scrutiny.
When sizes differed only little, as in pairings involving females of N. neobaraki andfemales of N. baraki, mortality was very low. While this again confirms the rule that dif-ferential size matters, behavioural observations on these females also showed that thereis an alternative to being killed: N. neobaraki females stayed in the centre of the discswhere the prey was placed, but N. baraki females moved along the edges of the discs.This may be interpreted as escape behaviour, a response commonly observed in
72
CHAPTER 5
Nazer-chap5.qxd 14-9-2012 10:37 Page 72

intraguild prey (Hindayana et al. 2001; Yasuda et al. 2001; Michaud and Grant 2003;Janssen et al. 2007). In our experiments individuals cannot escape, but, under field con-ditions, individuals may move away (e.g., out of the area under the perianth) uponencountering an intraguild predator. Thus, caution should be exercised when extrapolat-ing lab observations to the field.
Usually, intraguild predation decreases as the density of the shared, extraguild preyincreases (Schausberger 2003; Zannou et al. 2005). Indeed, females of N. paspalivorusbecome less aggressive towards larvae of N. neobaraki when the density of coconutmites, their prey, increases (Negloh et al. 2012). However, in our study where we offeredcoconut mites as prey to mimick conditions where they feed and compete for prey, N.neobaraki females exhibited intraguild predation on heterospecific larvae, females andmales despite the presence of prey. This implies that niche competition between phyto-seiid species is not only determined by food competition and intraguild predation onsmaller species, but also by imposing greater mortality on males than on females of theother species. Since phytoseiid females reproduce by pseudo-arrhenotoky and thusrequire insemination to produce daughters and sons (Sabelis et al. 2002), smaller phyto-seiid species may go extinct when males become too rare due to intraguild predation,unless there is a refuge where they can escape from this. These mechanisms may explainwhy the three phytoseiid species under study were not, or rarely, found co-occurringunder the same perianth of an Aceria-infested coconut fruit. In Tanzania where N. neo-baraki is prevalent, only one out of the 18 locations surveyed, had N. neobaraki and N.paspalivorus occurring together on two nuts from two different palms (Negloh et al.2011), but unfortunately no information was provided on the sex of the co-occurringindividuals of the two species.
If it is true that intraguild predation limits co-occurrence of phytoseiid speciesunder the perianth of the same coconut, then there would be little room for synergistic
73
Females as intraguild predators of males
Figure 5.1 Ratio body length (a) and width (b) of female/male between Neoseiulus paspalivorus,N. baraki and N. neobaraki. Measurements from 20 specimens of each species.
Nazer-chap5.qxd 14-9-2012 10:37 Page 73

effects between the three phytoseiid species and therefore also no reason for strategiesto combat coconut mites with all three species together. However, if there are refugesfrom intraguild predation under the perianth (e.g., N. paspalivorus occupying below-peri-anth space too narrow for the other two species), then coexistence may be possible, andpossibly also synergistic effects of two or more phytoseiid species on suppression ofcoconut mites. We consider this to be a key issue for future research on multiple pred-ator species that are used to control coconut mites.
AcknowledgementsWe thank R. Houndafoche and P. Sovimi for their assistance in maintaining the predator coloniesand conducting experiments. This research was supported by the International Institute ofTropical Agriculture (IITA) and the University of Amsterdam, the Netherlands, through a grantfrom the Board of the Netherlands Foundation for the advancement of Tropical Research(WOTRO). This study is part of the PhD Thesis of the senior author.
ReferencesCongdon BD, McMurtry JA (986) The distribution and taxonomic relationships of Euseius quetza-
li McMurtry in California (Acari: Phytoseiidae). Int J Acarol 12: 7–11.Croft BA (1970) Comparative studies on four strains of Typhlodromus occidentalis Nesbitt (Acarina:
Phytoseiidae). I. Hybridization and reproductive isolation studies. Ann Entomol Soc Am63:1528–1531.
Famah Sourassou N, Hanna R, Zannou I, Breeuwer JAJ, Moraes G, Sabelis MW (2012)Morphological, molecular and cross-breeding analysis of geographic populations of coconutmite-associated predatory mites identified as Neoseiulus baraki: evidence for cryptic species?Exp Appl Acarol 57:15–36.
Hindayana D, Meyhöfer R, Scholz D, Poehling H-M (2001) Intraguild predation among the hov-erfly Episyrphus balteatus de Geer (Diptera: Syrphidae) and other aphidophagous predators.Biol Control 20:236–246.
Hoying SA, Croft BA (1977) Comparisons between populations of Typhlodromus longipilus Nesbittand T. occidentalis Nesbitt: taxonomy, distribution and hybridization. Ann Entomol Soc Am 70:150–159.
Janssen A, Sabelis MW, Magalhães S, Montserrat M, Van der Hammen T (2007) Habitat structureaffects intraguild predation. Ecology 88:2713–2719.
Lawson-Balagbo ML, Gondim MGC, Moraes GJ, Hanna R, Schausberger P (2008). Explorationof acarine fauna on coconut palm in Brazil with emphasis on Aceria guerreronis (Acari:Eriophyidae) and its natural enemies. Bull Entomol Res 98:83–96.
Lucas É, Coderre D, Brodeur J (1998) Intraguild predation among aphid predators: characteriza-tion and influence of the extraguild prey density. Ecology 79:1084–1092.
Mahr DL, McMurtry JA (1979) Crossbreeding studies involving populations of Typhlodromus citriGarman and T. arboreus Chant and a sibling species of each. Int J Acarol 5:155–161.
McMurtry JA Mahr DL, Johnson HG (1976) Geographic races in the predaceous mite, Amblyseiuspotentillae (Acari: Phytoseiidae). Ann Entomol Soc Am 57:649–655.
Michaud JP, Grant AK (2003) Intraguild predation among ladybeetles and a green lacewing: dothe larval spines of Curinus coeruleus (Coleoptera: Coccinellidae) serve as a defensive function?Bull Entomol Res 93:499–505.
Monetti LN, Croft BA (1997) Mating, cross-mating and related behaviours in of Neoseiulus califor-nicus and Neoseiulus fallacis (Acari: Phytoseiidae). Exp Appl Acarol 21:67–74.
74
CHAPTER 5
Nazer-chap5.qxd 14-9-2012 10:37 Page 74

Moraes GJ, Lopez PC, Fernando LCP (2004) Phytoseiid mites (Acari: Phytoseiidae) of coconutgrowing areas in Sri Lanka, with description of three new species. J Acarol Soc Jpn13:141–160.
Negloh K, Hanna R, Schausberger P (2012) Intraguild predation and cannibalism between thepredatory mites Neoseiulus neobaraki and N. paspalivorus, natural enemies of the coconut miteAceria guerreronis. Exp Appl Acarol 58(3) DOI 10.1007/s10493-012-9581-6.
Negloh K, Hanna R, Schausberger P (2011) The coconut mite, Aceria guerreronis, in Benin andTanzania: occurrence, damage and associated acarine fauna. Exp Appl Acarol 55:361-374.
Rosenheim JA, Kaya HK, Ehler LE, Marois JJ, Jaffee BA (1995) Intraguild predation among bio-logical control agents; theory and evidence. Biol Control 5:303–335.
Sabelis MW, Nagelkerke CJ, Breeuwer JAJ (2002) Sex ratio control in arrhenotokous and pseudo-arrhenotokous mites. In: Sex Ratios, Concepts and Research Methods (ICW Hardy, ed.), Cam-bridge University Press, Cambridge, UK, pp. 235-253.
SAS (2005) SAS Institute, version 9.2 Inc., Cary.Sato S, Dixon AFG, Yasuda H (2003) Effects of emigration on cannibalism and intraguild preda-
tion in aphidophagous ladybirds. Ecol Entomol 28:628–633.Schausberger P (2003) Cannibalism among phytoseiid mites: a review. Exp Appl Acarol
29:173–191Schausberger P, Croft B (2000) Cannibalism and intraguild predation among phytoseiid mites: Are
aggressiveness and prey preference related to diet specialization? Exp Appl Acarol 24:709–725.Yasuda H, Kikuchi T, Kindlmann P, Sato S (2001) Relationships between attack and escape rates,
cannibalism, and predation in larvae of two predatory ladybirds. J Ins Behav 14:373–384.Zannou ID, Moraes GJ, Ueckermann EA, Olivera AR, Yaninek JS, Hanna R (2006) Phytoseiid
mites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from Sub-Saharan Africa. Int JAcarol 32:241–276.
Zannou I, Hanna R, Moraes JG, Kreiter S (2005) Cannibalism and interspecific predation in aphytoseiid predator guild from cassava field in Africa: Evidence from the laboratory. ExpAppl Acarol 37:27–42.
75
Females as intraguild predators of males
Nazer-chap5.qxd 14-9-2012 10:37 Page 75

Nazer-chap5.qxd 14-9-2012 10:37 Page 76

77
Predatory mites found under the bracts ofcoconuts and their potential for controlling Aceria gguerreronis: review and hypotheses
ABSTRACT – The coconut mite Aceria guerreronis feeds on the meristematic tissueunder the bracts of coconuts and is a serious pest of coconut palms. Predatorymites found on Aceria-infested coconuts are reviewed based on extensive surveysin Benin, Tanzania, Brazil, Colombia, Mexico, Venezuela, Sri Lanka, Oman andIndia, carried out to explore possibilities for biological control of coconut mites.Three closely related species, Neoseiulus paspalivorus, N. baraki and N. neobaraki,stand out as being most abundant and closely associated with coconut mitesunder the bracts of coconuts. Unlike N. neobaraki found to occur in Africa only,the other two predator species were present in all continents explored, but geo-graphic populations of these two species were reproductively isolated due toendosymbiont infections and/or genetic factors. A COI-based phylogeneticanalysis as well as morphological analysis of geographic populations of each ofthe three species led to the hypothesis that the predatory mite N. paspalivorus hasfollowed the same route of continental invasions as coconut mites, their prey,whereas N. baraki (especially from Benin) and N. neobaraki belong to a complexspecies that had much of their early evolutionary history on weeds and moved tococonut palms more recently. Based on their reproductive incompatibilities weused geographic populations as the basic genetic entities for evaluating the bio-logical characteristics of these two predator species for their use in biologicalcontrol. We found differences in life history traits and ability to utilize pollen suchthat there was a tendency for relatively high population growth capacities to beassociated with lower abilities to utilize pollen. Although this trait association ispremature and needs statistical validation, it poses a testable hypothesis on atrade-off between population growth and alternative food utilization and thishypothesis is crucial for determining which geographic race is the best candidatefor coconut mite control.
Unpublished manuscript
Based on socio-economic impact the coconut palm, Cocos nucifera L., has been classi-fied among the most important crops in the world and especially in the Tropics
(Vietmeyer 1986). However, its yield is significantly reduced by several arthropods thatfeed on this plant (Lever 1969; Krantz et al. 1978). The coconut mite, Aceria guerreronisKeifer (Acari: Eriophyidae), is one of the most important arthropod pests of this cropworldwide. It was originally described from specimens collected on coconut fruits in
6Nazer-chap6.qxd 14-9-2012 10:37 Page 77

Guerrero, Mexico (Keifer 1965). It is a specialist herbivore of young coconuts, attack-ing mainly the meristematic zone covered by the perianth (Moore et al. 1989). Damageto coconut production ranges between 67-85% in Benin (West Africa) and 43-81% inTanzania (East Africa) (Negloh et al. 2011). Increasing public concern over the use ofinsecticides, together with the fact that several insecticides have become ineffective ortheir application became impracticable (Moore and Alexander 1987; Moore et al. 1989)has provided the impetus to develop more sustainable and environmentally safe strate-gies for controlling the coconut mite, such as biological control. Predatory mites arewidely distributed in most terrestrial ecosystems, and effective biological control agentsof a wide range of harmful mites and insects have been reported (Helle and Sabelis1985; Lindquist et al. 1996; McMurtry and Croft 1997).
This paper provides a review of the predatory mites reported from surveys con-ducted in several African, Asian, South-, Central- and North-American countries. Wefocus on the three most abundant and apparently most important mite predators of A.guerreronis. Based on the distribution of these predator species over the continents, theirgenetic and morphological variability in each of the continents and an earlier assessmentof the invasion history of coconut mites (Navia et al. 2005) we develop a hypothesis onthe invasion routes (or the absence thereof) for each of the three predator species. Next,we consider genetic variation among reproductively isolated, geographic populations ofthese predators with particular emphasis for traits relevant to biological control ofcoconut mites.
MethodologyA total of 9 countries from 3 continents (Africa, Asia, and America) were surveyed forthe occurrence of natural enemies of A. guerreronis on coconuts while still attached tothe palm (excluding fallen coconuts, leaves, branches and inflorescences) (Table 6.1).Data from surveys conducted in Brazil (Lawson-Balagbo et al. 2008a; Reis et al. 2008),Colombia, Mexico and Venezuela (Silva et al. unpublished), Benin and Tanzania (Neglohet al. 2011; Zannou et al. unpublished), India (Banerjee and Gupta 2011), Oman(Hountondji et al. 2010), Sri Lanka (Moraes et al. 2004; Fernando and Aratchige 2009)were put together not only to provide a species list but also to assess the relative abun-dance of each species in each country.
Predatory mites associated with Aceria gguerreronisAltogether, about 60 predatory mite species belonging to 6 families have been reportedfrom surveys conducted in Africa, the Americas and Asia (Table 6.2). The most fre-quently-found species belong to the Phytoseiidae and Ascidae sensu lato (Lindquist et al.2009). Other less abundant predatory mites belong to Bdellidae, Cheyletidae, Cunaxidaeand Tydeidae (Table 6.2, Figure 6.1).
78
CHAPTER 6
Nazer-chap6.qxd 14-9-2012 10:37 Page 78

PhytoseiidaeOverall, 32 phytoseiid species belonging to eleven genera have been collected on infest-ed coconut fruits (Table 6.2), representing 53% of all predatory mite species (Figure6.1). In each of the locations surveyed three closely related phytoseiid species, namelyNeoseiulus paspalivorus De Leon, N. baraki Athias-Henriot and N. neobaraki Zannou,Moraes & Oliveira are by far the most abundant and the most frequently-found in closeassociation with A. guerreronis under the bracts of coconuts (Table 6.3). Evidently, theirsize permits them to move into the space between the bracts and the fruit surface.Because of their their exclusive occurrence on coconut in association with A. guerreronis
79
Predators under coconut bracts: review and hypotheses
Table 6.1 Continents and countries surveyed for natural enemies of Aceria guerreronis.Continent Country ReferencesAfrica Benin (1) Negloh et al. 2011; (2) Zannou et al. unpublished data
Tanzania (1) Negloh et al. 2011; (2) Zannou et al. unpublished dataAmericas Brazil (3) Lawson-Balagbo et al. 2008; (4) Reis et al. 2008; Oliveira et al. 2012
Colombia (5) Silva et al. unpublished dataMexico (5) Silva et al. unpublished dataVenezuela (5) Silva et al. unpublished data
Asia India (6) Banerjee and Gupta 2011Oman (7) Hountondji et al. 2010Sri Lanka (8) Moraes et al. 2004; (9) Fernando and Aratchige 2011
Numbers in parentheses refer to the superscripts in Table 6.2.
Figure 6.1 Overall percent relative abundance of species in the most important families ofpredatory mites reported from survey conducted in Africa, South America and Asia. Percentswere calculated based on data provided by Lawson-Balagbo et al. (2008), Reis et al. (2008),Negloh et al. (2011), Banerjee and Gupta (2011), Fernando and Aratchige (2011), Silva et al.(unpublished), and Zannou et al. (unpublished). The percent relative abundance of mite speciesin each family was calculated by dividing the number of mite species recorded for that family inall these locations surveyed by the total of all mite species recorded.
Nazer-chap6.qxd 14-9-2012 10:37 Page 79

80
CHAPTER 6Ta
ble
6.2
Ext
ant o
fpr
edat
ory
mite
spec
ies re
porte
d in
ass
ociat
ion
with
A. g
uerr
eroni
sKeif
er a
t diff
eren
t loc
atio
ns su
rvey
ed.
Fam
ilyA
frica
Am
erica
s A
siaG
ener
a an
d Sp
ecies
Beni
n1,2
Tanz
ania1
,2Br
azil3
,4Co
lom
bia5
Mex
ico5
Vene
zuela
5In
dia6
Om
an7
Sri L
anka
8,9
Phyt
oseii
dae
Amb
lyseiu
s adh
atod
ae M
uma
+-
--
--
--
-A
mblys
eius a
eriali
sMum
a-
-+
--
--
--
Amb
lyseiu
sherb
icolus
Chan
t+
--
--
--
--
Amb
lyseiu
s lar
goen
sisM
uma
++
++
++
++
+A
mblys
eius t
amat
aven
sisBl
omm
ers
--
+-
--
--
-A
mblys
eiuss
p.-
--
+-
--
--
Cydn
oseiu
s nege
viSw
irski
& A
mita
i-
--
--
--
+-
Eus
eius a
latus
DeL
eon
--
+-
--
--
-E
useiu
s ben
inen
sis M
orae
s & M
cMur
try
+-
--
--
--
-E
useiu
s doss
ei Pr
itcha
rd &
Bak
er+
--
--
--
--
Gale
ndro
muss
p +
-+
--
--
--
Iphis
eius d
egene
rans
Berle
se+
--
++
--
--
Meye
rius s
p.-
--
--
--
+-
Neos
eiulus
bar
akiA
thias
-Hen
riot
++
++
++
--
+N
eoseiu
lus n
eobar
akiZ
anno
u,M
orae
s & O
liver
a+
+-
--
--
--
Neos
eiulus
pas
paliv
orus
DeL
eon
++
++
++
++
+N
eoseiu
lus re
cifen
sisG
ondi
m Jr
.& M
orae
s-
- +
--
--
--
Neos
eiulus
mum
aiD
enm
ark
--
-+
--
--
-N
eoseiu
lussp
.-
--
+-
+-
--
Phyto
scutu
s wies
ei U
ecke
rman
n &
Loo
ts+
--
--
--
--
Prop
riosei
opsis
cann
aens
isM
uma
--
+-
--
--
-Pr
oprio
seiop
sis m
exica
nusG
arm
an-
--
++
+-
--
Prop
riosei
opsis
sp.
--
-+
++
--
-Ty
phlod
romi
na su
btro
pica
Mum
a &
Den
mar
k-
-+
++
+-
--
Typh
lodro
mips
cana
neien
sis(G
ondi
m Jr
.& M
orae
s-
-+
--
--
--
Typh
lodro
mus (
A.)
aspe
roset
osus Z
anno
u et
al.
+-
--
--
--
-Ty
phlod
romu
s aff.
vulga
risE
hara
--
+-
--
--
-Ty
phlod
romu
s dalf
ardic
usD
anes
hvar
--
--
--
-+
-Ty
phlod
romu
s orn
atus
Den
mar
k &
Mum
a-
-+
--
--
--
Ueck
erman
nseiu
s eas
tafri
caeM
orae
s et a
l.+
--
--
--
--
Ueck
erman
nseiu
s qui
licii
Uec
kerm
ann
& K
reite
r-
+-
--
--
--
Ueck
erman
nseiu
s neoh
avu
Mor
aes e
t al.
+-
--
--
--
-+
Foun
d;-N
ot fo
und
Supe
rscr
ipts
beh
ind
coun
tries
:see
refe
renc
es in
Tab
le 6.
1
Nazer-chap6.qxd 14-9-2012 10:37 Page 80

81
Predators under coconut bracts: review and hypothesesTa
ble
6.2
Cont
inue
d.Fa
mily
Afr
icaA
mer
icas
Asia
Gen
era
and
Spec
iesBe
nin1
,2Ta
nzan
ia1,2
Braz
il3,4
Colo
mbi
a5M
exico
5Ve
nezu
ela5
Indi
a6O
man
7Sr
i Lan
ka8,
9
Asc
idae
s.lat
o1A
sca fo
xiD
eLeo
n-
-+
--
--
--
Asca
sp+
--
-+
+-
-Bl
attis
ocius
keeg
aniF
ox-
-+
--
--
--
Gam
asell
odes
sp+
--
--
--
--
Hop
loseiu
ssp.
--
--
--
--
-La
siosei
us su
bterr
aneu
sCha
nt+
-+
--
--
--
Lasio
seius
sp.B
erles
e+
+-
-+
++
--
Meli
char
es aff
. agil
isH
erin
g-
-+
--
--
--
Proct
olaela
ps b
ickley
iBra
m+
-+
++
+-
--
Proct
olaela
ps co
ffeae
Kar
g &
Rod
rigue
z-
-+
--
--
--
Proct
olaela
ps in
terme
diusA
thias
-Hen
riot
--
+-
--
--
-Pr
octola
elaps
bulbo
sus M
orae
s et a
l.-
-+
--
--
--
Proct
olaela
ps sp
.-
-+
++
++
+-
Chey
letid
aeCh
eletom
imus
sp.
--
+-
--
--
-Ch
eletom
imus
gracil
is Fa
in,B
ochk
.& C
orp.
-Rar
os-
--
--
--
+-
Cheyl
etus m
alacce
nsis
Oud
eman
s-
--
--
-+
--
Hem
icheyl
etia
sp-
-+
--
--
--
Mex
echele
ssp.
--
--
--
--
-Pa
rach
elytia
sp.
--
-+
++
--
-Bd
ellid
aeBd
ella
distin
ctaBa
ker a
nd B
alogh
++
+-
--
--
-Bd
ella
sp.
--
-+
--
--
-Sp
inibd
ella
sp.
-+
+-
--
--
-Cy
tasp
.-
-+
--
--
--
Cuna
xida
eA
rmas
cirus
sp.
--
+-
--
--
-Ty
deid
aeA
frotyd
eus s
p.+
--
--
--
--
Meta
pron
emat
us sp
.-
-+
--
--
--
Pron
emat
us sp
.-
+-
--
--
--
1 Thi
s gro
up w
as re
cent
ly sp
lit in
to th
ree
fam
ilies
(Lin
dqui
st e
t al.
2009
);ac
cord
ing
to th
is sy
stem
,Asca
and
Gam
asell
odes
are
plac
ed in
Asc
idae
,Bl
attis
ocius
,Hop
loseiu
sand
Las
ioseiu
sare
plac
ed in
Blat
tisoc
iidae
,whe
reas
Meli
char
es an
d Pr
octola
elaps
are
plac
ed in
Meli
char
idae
Nazer-chap6.qxd 14-9-2012 10:37 Page 81

and their demonstrated ability to feed and reproduce on a diet of coconut mite alone(Lawson-Balagbo et al. 2007; Negloh et al. 2008), these species are generally consideredthe most important predators of coconut mites.
The Amblyseius species are the second most abundant group of predators collectedfrom Aceria-infested coconuts. Amblyseius largoensis (Muma) was reported from all loca-tions surveyed, but in very low numbers per coconut, and occasionally found under thebracts of Aceria-infested fruits. This species is larger than the three above-mentionedNeoseiulus species found under the bracts, what suggests that its size limits access to thearea under the bracts. It is found much more abundantly elsewhere on the coconutpalm, especially on coconut leaves and fallen coconuts (Fernando and Aratchige 2009;Negloh et al. 2011; Lawson-Balagbo et al. 2008a; Silva et al. unpublished).
Specimens identified as belonging to Proprioseiopsis, Typhlodromina and Typhlodromipswere reported from South America, but not from Africa or Asia. Species of Typhlodromuswere recorded only in South America and Africa. According to I. Zannou (personalcommunication) a specimen from Benin mistakenly identified as a Galendromus species(Negloh et al. (2011), was upon examination of additional specimens observed to cor-respond to a Typhlodromus species (Anthoseius). Other genera rarely represented wereIphiseius from Benin, Colombia and Mexico and Meyerius and Cydnoseius from Oman.
Ascidae sensu latoThis group was recently split into three families (Lindquist et al. 2009); according to thisnew system, Asca and Gamasellodes were kept in the Ascidae, Blattisocius, Hoploseius andLasioseius were placed in Blattisociidae, whereas Melichares and Proctolaelaps were placed inMelicharidae. Given the considerable morphological and behavioural similaritiesbetween them, we retain in this paper the old classification system (Lindquist and Evans1965), referring to them as Ascidae sensu lato. This group accounts for 22% of the preda-tory mite species found on coconuts attached to the palm (Figure 6.1). Among them,Proctolaelaps species dominate, but they have not been found in Tanzania, Oman and SriLanka. These species are much less abundant on the coconuts attached to the palm treethan they are on fallen coconuts on which they can reach large numbers. Bracts on fall-en coconuts stand more distantly from the fruit surface, thereby providing Proctolaelapspredators with access to the space under the bracts despite being larger than the three
82
CHAPTER 6
Table 6.3 Percent relative abundance of Neoseiulus paspalivorus, N. baraki and N. neobaraki at dif-ferent locations surveyed.
Benin Tanzania Brazil Colombia Mexico VenezuelaN. paspalivorus 84.5 15.3 61 77 21.3 71.5N. baraki 12.5 22.7 26 5.8 11.1 4.2N. neobaraki 1.7 59 0 0 0 0Others 1.3 3 13 7.2 66.7 4.3
Nazer-chap6.qxd 14-9-2012 10:37 Page 82

Neoseiulus species. The larger body size is probably a major limitation for Proctolaelapsbickleyi (Bram) and P. bulbosus (Moraes, Reis and Gondim Jr.) to become effective pred-ators of A. guerreronis on fruits attached to palms despite their relatively voracity andpopulation growth (Lawson-Balagbo et al. 2007; Galvão et al. 2011b; Lima et al. 2012).The second most abundant ascid group belongs to Lasioseius. It is reported fromcoconuts in South America and Africa, but not from Oman and Sri Lanka. No detailedinvestigation about the potential of species from this group has been carried out.
Bdellidae, Chelytidae, Tydeidae and CunaxidaeBdellidae account for 10% of all predatory mite species collected from coconut. Bdellaspp. have been reported in Brazil, Benin, Tanzania and Colombia, whereas Spinibdellaspp. have been reported from Tanzania and Brazil and Cyta spp. from Brazil. Cheyletidaerepresent 7% of all predatory mite species collected from coconut, and were reportedfrom South America and Asia. Cunaxidae represent 7% of all species and were repre-sented by Armascirus species in Brazil. Tydeidae were rare and represented by Afrotydeussp. in Benin, Pronematus sp. in Tanzania, and Metapronematus sp. in Brazil. All these non-phytoseiid species were collected mainly from the exposed surface of coconut fruits.
Candidate biocontrol agents of coconut mitesThree phytoseiid species, Neoseiulus paspalivorus, N. baraki and N. neobaraki, stand out ascandidate biocontrol agents of coconut mites, because they are the most abundantspecies associated with coconut mites. The first two are common on all continentsincluded in the surveys, but the third species is only found in Africa and is most preva-lent in Tanzania. Depending on location surveyed, the three species comprise up to 80-98% of the phytoseiids associated with A. guerreronis under the bracts. The relativelysmall, flat and elongated body shape of these mites facilitates their access under thebracts of coconuts, being the microhabitat inhabited by the target prey (Moraes et al.2004).
Neoseiulus paspalivorus was first described by De Leon (1957) from specimens col-lected from a Paspalum grass species in association with Steneotarsonemus spp., on which itcould be feeding (De Leon 1957). This predator has been reported from all countrieswhere surveys were made of the predators under the bracts of Aceria-infested coconuts,in some occasions also in association with Steneotarsonemus spp. (Moraes et al. 2004;Lawson-Balagbo et al. 2008a; Reis et al. 2008; Negloh et al. 2011; Fernando andAratchige 2009; Hountondji et al. 2010; Banerjee and Gupta 2011, Silva et al. unpub-lished). It is by far the most dominant predator in all countries surveyed, except inTanzania (Africa) and Mexico (North America) (Lawson-Balagbo et al. 2007; Negloh etal. 2011; Silva et al. unpublished data) (Table 6.3). Compared to other Neoseiulus speciesfound on coconut, N. paspalivorus has the smallest size and the shortest legs (Figure 6.2).
83
Predators under coconut bracts: review and hypotheses
Nazer-chap6.qxd 14-9-2012 10:37 Page 83

Neoseiulus baraki was first described by Athias-Henriot (1966) from specimens col-lected from weeds (Phalaris sp.) in Algeria (Africa). Further morphological informationabout this species was provided by Ehara and Bandhufalck (1977), Schicha (1981),Moraes et al. (2004) and Famah Sourassou et al. (2012). This species is slightly largerthan N. paspalivorus and has slightly longer legs (Figure 6.2). It has been the second mostfrequently reported phytoseiid species in all countries surveyed, except in India andOman where this species was not found (Hountondji et al. 2010; Banerjee and Gupta2011). Neoseiulus baraki is the most dominant phytoseiid in Sri Lanka, almost as domi-nant as N. neobaraki in Tanzania (Zannou et al. unpublished data), but the second mostprevalent phytoseiid species in Brazil and Benin (Negloh et al. 2011; Lawson-Balagbo etal. 2008; Zannou et al. unpublished data).
Neoseiulus neobaraki is so far found in Benin and Tanzania only, being the mostprevalent species in the latter country. It is the largest of the three Neoseiulus species(Figure 6.2). It was described by Zannou et al. (2006) from specimens collected from theweed Andropogon gayanus and from coconut, and is morphologically close to N. baraki.
Climatic requirementsSurveys conducted in Brazil (Lawson-Balagbo et al. 2008; Reis et al. 2008) and Benin(Negloh et al. 2011; Zannou et al. unpublished data) suggest that N. paspalivorus thrivesin dry areas, but N. baraki and N. neobaraki in humid areas. However, a study conduct-ed in Sri Lanka reported N. paspalivorus prevailing at relatively humid areas and N. bara-ki in relatively dry areas, both species co-occurring in areas with intermediate climaticconditions (Fernando and Aratchige 2009). To understand these contrasting observa-tions there is a need to quantify the extent to which an area is relatively dry/humid, but
84
CHAPTER 6
Figure 6.2 Length and width of the dorsal shield of Neoseiulus paspalivorus, N. baraki and N. neo-baraki. Proctolaelaps bickleyi was included for comparison.
Nazer-chap6.qxd 14-9-2012 10:37 Page 84

also the extent to which populations in different geographic areas are genetically similaror dissimilar. In addition, it should be noted that these species live exclusively under thebracts where the microclimate differs dramatically from the ambient climate.
Biology and behaviourBased on data availability and the supposed relevance to biological control, we focus ontwo traits of the Neoseiulus predators: (1) the population growth under conditions ofample supply of coconut mites as prey, and (2) the ability to utilize coconut pollen asfood source for survival, development and reproduction. We assessed these data for N.paspalivorus populations from Benin, Ghana and Brazil. All assessments were done at25±1 °C, 70-90% RH and 12:12h light-dark cycle. All N. paspalivorus populations wereobserved to feed, develop and reproduce successfully on A. guerreronis (Table 6.4), butthey differed in population growth rates and in ability to utilize coconut pollen for sur-vival, and development. In particular, none of the populations could reproduce oncoconut pollen. Moreover, the population from Benin was not able to develop on a dietof coconut pollen, whereas the other populations studied did so successfully. Theseresults suggest other possible differences between these reproductively isolated popula-tions. The data also indicated a negative relation between the ability to utilize pollen fordevelopment and adult survival and the population growth on a diet of coconut mites,as shown in Figure 6.4 where the intrinsic rate of population increase (rm) is plottedagainst the developmental rate (inverse of developmental time) and longevity on a dietof coconut pollen. We will further refer to this trend as the negative trade-off hypoth-esis. A special note should be made with respect to data on a population of N. pas-palivorus from Brazil (Acaraú) (Lawson-Balagbo et al. 2007). The results from that studydiffer strikingly from the results from the present study in that the predators were ableto reproduce on pollen. However, we emphasize that these data cannot be comparedwith ours because reproduction on a diet of coconut pollen was measured using femalesthat completed their development on a diet of coconut mites. In our study (where wefound no such egg production) we used females that completed their development oncoconut pollen. Complementary studies are needed to resolve this issue.
For populations previously identified as N. baraki, the population growth rates on adiet of coconut mites and the ability to utilize pollen were published by Negloh et al.(2008). That study showed that population growth on a diet of coconut mites was higherin the Brazil population, but the ability to utilize pollen was higher for the Benin popula-tion whose females could reproduce (after reaching adulthood and mating on coconutpollen). This is again in agreement with the hypothesis of a negative trade-off betweenpopulation growth rate on a diet of coconut mites and the ability to utilize coconut pollenas food. Unfortunately, nothing is known about the life history of N. neobaraki on differ-ent diets because these data could also serve to test the trade-off hypothesis.
85
Predators under coconut bracts: review and hypotheses
Nazer-chap6.qxd 14-9-2012 10:37 Page 85

86
CHAPTER 6
Tabl
e 6.
4Su
mm
ary
ofth
e lif
e hi
stor
y pa
ram
eter
s of
thre
e N
eoseiu
lus p
aspa
livor
usge
ogra
phic
popu
latio
ns o
n th
ree
diffe
rent
food
s typ
es.
Egg
-Adu
lt1Su
rviv
al2%
fem
ales
Long
evity
3Pr
e-O
vipo
sitio
n3Po
st-
Fecu
ndity
4Se
x ra
tio5
r mgr
avid
ovip
ositi
on3
ovip
ositi
on3
A. g
uerr
eroni
sBe
nin
5.6
± 0
.05
b10
0 (5
0)10
0 (3
4)37
.3 ±
2.5
b1.
8 ±
0.1
b23
.1 ±
1.1
a12
.2 ±
2.1
b1.
4 ±
0.0
4 b
69.8
0.19
1G
hana
5.6
± 0
.05
b98
(50)
100
(31)
35.9
± 3
.2 b
2.6
± 0
.3 a
26.1
± 1
.9 a
7.4
± 1
.8 c
1.1
± 0
.05
c69
.10.
149
Itam
arac
a6.
2 ±
0.0
8 a
100
(50)
100
(30)
44.3
± 1
.8 a
2.1
± 0
.1 a
b18
.4 ±
0.8
b23
.1 ±
1.9
a1.
8 ±
0.0
6 a
69.5
0.20
7Co
conu
t pol
lenBe
nin
13.6
± 1
.11
a7.
1 (7
0)0.
0 (3
)9.
6 ±
2.5
b–
––
––
Gha
na9.
9 ±
0.1
0 b
91.4
(70)
0.0
(34)
26.7
± 2
.1 a
––
––
–Ita
mar
aca
11.1
± 0
.22
b70
(70)
0.0
(24)
13.1
± 1
.3 b
––
––
–D
iffer
ent l
ette
rs in
dica
te si
gnifi
cant
diff
eren
ces p
er fo
od ty
pe a
mon
g pr
edat
or p
opul
atio
ns (p
airw
ise S
tude
nt’s
t-tes
t;P<
0.05
);1 T
otal
dura
tion
ofde
velo
pmen
t (in
day
s);2
Perc
enta
ge re
achi
ng a
dulth
ood;
3 Mea
n in
day
s;4 E
ggs p
er fe
male
per
day
;5Pe
rcen
tage
fem
ale o
ffspr
ing;
r m,i
ntrin
sic ra
teof
popu
latio
n in
crea
se
Tabl
e 6.
5Pa
irwise
dist
ance
mat
rix w
ithin
- and
bet
wee
n th
ree
coco
nut m
ite-a
ssoc
iated
Neos
eiulus
Hug
hes.
Spec
iesPo
pulat
ions
12
34
56
78
910
N. p
aspa
livor
us1.
Braz
il2.
Gha
na0.
003
3.Be
nin
(1)
0.00
00.
003
4.Be
nin
(2)
0.00
00.
003
0.00
0N
. bar
aki
5.Br
azil
0.36
30.
363
0.36
30.
363
6.Ta
nzan
ia0.
363
0.36
30.
363
0.36
30.
000
7.Ta
nzan
ia0.
363
0.36
30.
363
0.36
30.
000
0.00
08.
Beni
n0.
350
0.35
40.
350
0.35
00.
127
0.12
70.
127
N. n
eobar
aki
9.Be
nin
0.36
30.
363
0.36
30.
363
0.13
30.
133
0.13
30.
118
10.T
anza
nia
0.33
80.
342
0.33
80.
338
0.14
20.
142
0.14
20.
152
0.08
9Pr
octola
elaps
bick
leyi(
outg
roup
)11
.Ben
in0.
322
0.32
60.
322
0.32
20.
363
0.36
.30.
363
0.34
20.
346
0.34
2Th
e di
stan
ce m
atrix
was
con
stru
cted
usin
g th
e Ju
kes-
Cant
or m
odel
(Juke
s and
Can
tor 1
969)
in M
EG
A®
softw
are
vers
ion
5.
Nazer-chap6.qxd 14-9-2012 10:37 Page 86

Phylogenetic relationshipsUsing published data on cytochrome oxidase subunit I (COI) sequences for N. baraki(Famah Sourassou et al. 2012; GenBank accession numbers JQ609092/93/94), Neoseiuluscalifornicus McGregor (GenBank AB500127/29/30/31), Neoseiulus womersleyi (Schicha)(GenBank AB500132/33/34/35/36) and Iphiseius degenerans (Berlese) (GenBankFM210120) and data on N. paspalivorus (to be submitted to GenBank) and N. neobaraki(to be submitted to GenBank) we carried out a phylogenetic analysis, the results of which
87
Predators under coconut bracts: review and hypotheses
Figure 6.3 Phylogenetic relationships of predatory mites identified as Neoseiulus paspalivorus, N.baraki and N. neobaraki. 1Singino; 2Nachenjele; 3Specimens from coconut; 4Specimens fromweeds; A search was performed in the GenBank in an attempt to retrieve COI sequences of allNeoseiulus species available. Our search conducted on Mars 13, 2012 resulted in only 9 COIsequences from 2 Neoseiulus species. The 9 COI sequences from the GenBank, in addition tothe 10 COI sequences from this study, were automatically aligned by the aid of ClustalW(Thompson et al. 1994). COI sequences of Iphiseius degenerans from the GenBank was added inthe phylogenetic analysis as out-group. A global alignment of approximately 361 base pairs wasreached with the 20 COI sequences, which was then used for phylogenetic analysis. Tree basedon Neighbor-Joining method was constructed in MEGA5 using the Juke-Cantor model with1000 bootstrap replications.
Nazer-chap6.qxd 14-9-2012 10:37 Page 87

are presented in Figure 6.3 and Table 6.5. Neoseiulus paspalivorus appeared genetically themost distant from N. baraki and N. neobaraki as indicated by the 34-36% genetic dissim-ilarity. However, the 12-15% genetic dissimilarity between specimens identified as N.baraki and those identified as N. neobaraki is comparable to that between specimens fromBenin and those from Tanzania and Brazil previously identified as ‘N. baraki’, but nowconsidered to be a cryptic species. A genetic dissimilarity of 9% was found between spec-imens from Benin and those Tanzania identified as N. neobaraki. In the phylogenetic tree,N. paspalivorus formed a completely separate monophyletic group, whereas specimensidentified as N. baraki from Benin, from Tanzania and Brazil, and N. neobaraki fromBenin and Tanzania formed together another monophyletic group (Figure 6.3).
Regarding the taxonomic status of the three phytoseiid species so far examined, thephylogeny based on COI sequences suggests that a complex of species may occur in Africa,comprising specimens identified as N. baraki from Benin, specimens identified as N. barakifrom Tanzania, and specimens identified as N. neobaraki from Benin and Tanzania. Only onespecies from this complex in Africa is also found in Brazil and in Sri Lanka.
Invasion routeA study by Navia et al. (2005) has shown that South America is likely to be the area oforigin of the pest A. guerreronis, from where it invaded first Africa, and then Sri Lankaand India. This prompts the question: what has been the invasion route for the preda-tory mites associated with A. guerreronis? Did the predators follow the same route as theirprey or are they endemic on other host plants and moved to coconut palms later on?Summarizing the patterns of geographic distribution, together with morphological andmolecular data presented above, we propose that only N. paspalivorus followed the inva-sion route of its prey, whereas N. baraki is a complex of species that had a prior evolu-tionary history on an endemic host plant before moving onto coconut palms. This con-clusion is reached based on: (a) N. paspalivorus is the only species found in all geograph-
88
CHAPTER 6
Figure 6.4 Trend between the intrinsic rate of population increase (rm) on measured diet ofAceria guerreronis and and the developmental rate (inverse of developmental time) and longevityon a diet of coconut pollen. Different letters indicate significant differences between popula-tions in rm and longevity on diets of A. guerreronis and coconut pollen (pairwise Student’s t-test;P<0.05).
Nazer-chap6.qxd 14-9-2012 10:37 Page 88

ic locations/continents surveyed; (b) specimens of N. paspalivorus from the differentgeographic localities surveyed are morphologically and genetically similar, but clearlymore samples from a wide range of geographic localities are needed to critically test ourhypothesis;(c) specimens identified as N. baraki and N. neobaraki from Africa are aspecies-complex with only one species (from Tanzania) being morphologically andgenetically similar to the one population studied from Brazil and another from SriLanka. Yet again, a more detailed study involving samples from a wider geographicrange is still needed to fully determine the invasion route of the species in this complex.
DiscussionTo date, exploration for predatory mites on coconuts infested by A. guerreronis has yield-ed about 60 species of predatory mites, with Phytoseiidae being represented by 32species and being the most abundant group in all locations surveyed. The phytoseiidspecies N. paspalivorus, N. baraki and N. neobaraki stand out because they are (1) closelyassociated with coconut mites under the bracts of the coconut (in contrast to manyother species recorded, such as Amblyseius spp. and species of Ascidae sensu lato,Bdellidae, Cheyletidae, Cunaxidae and Tydeidae) and (2) by far the most dominant. Inaddition, except for N. neobaraki being found in Africa only, N. paspalivorus and N. bara-ki are found in samples of all continents. Predatory mites identified as N. paspalivoruswere found in all locations surveyed in Africa, Asia and South America, either alone orwith predatory mites identified as N. baraki and/or N. neobaraki. However, not includ-ed in this review, is a recently published survey conducted in Southeastern Brazil (SãoPaulo), which yielded neither N. paspalivorus nor N. baraki (Oliveira et al. 2012). This isprobably due to the fact that (1) coconut cultivation was only relatively recently intro-duced into that area, (2) A. guerreronis was probably only recently introduced to that area,and (3) the climatic conditions in Sao Paulo are very different from areas where this mitehas been found in other parts of the world.
Based on phylogenetic and morphological analyses of the species identified as N. pas-palivorus, N. baraki or N. neobaraki we propose that N. paspalivorus has followed the sameinvasion route as the coconut mites (from South-America to Africa and then to Asia)because it is found on all locations and all continents explored and because their geo-graphic populations on different continents are morphologically and genetically similar.We also propose that species previously identified as N. baraki moved from weeds tococonut palms after their introduction into the area and from Africa to other continentsbecause they are part of a highly differentiated species complex occurring in Africa andbecause specimens identified as N. baraki have been found on weeds in Africa (FamahSourassou et al. 2012; this study; Zannou, pers. comm.). These two proposals are no morethan hypotheses that need to be tested much more extensively by sampling from a widerrange of geographic localities in all three continents. Such extended surveys and analyses
89
Predators under coconut bracts: review and hypotheses
Nazer-chap6.qxd 14-9-2012 10:37 Page 89

could help to test whether N. paspalivorus has its greatest genetic diversity in SouthAmerica (as expected from our hypothesis) and whether Asian and South American pop-ulations identified as N. baraki are less genetically diversified than those in Africa.
These hypotheses on invasion routes are helpful in making decisions on releases ofpopulations from one continent into another. If the area of origin of the predatorymites is indicated by higher genetic diversity, then it follows that exploration of thisgenetic diversity for traits relevant to biological control may yield candidate strains forimproving coconut mite control in areas elsewhere. We hypothesize that the followingpredator traits are determinants of their impact on coconut mite populations: (1) size-dependent access to the space under the bracts of coconuts (Aratchige et al. 2007; Limaet al. 2012), (2) the functional and numerical response to the density of coconut mites(Lima et al. 2012) and (3) the ability to survive on alternative food, such as coconutpollen (Lawson-Balagbo et al. 2007; Negloh et al. 2008). Our data suggest that thecapacity for population increase on a diet of coconut mites is negatively related to theability to utilize coconut pollen (Figure 6.4). While this is no more than a trend basedon small sample, this hypotehesis is worthy testing by more in-depth research, especial-ly because the phytoseiids at large also show such a negative relation (McMurtry andCroft 1997). If this negative trade-off holds, then we need to determine which combi-nation of the two traits involved in the trade-off is best for biological control. This isan important issue for future research. It is also not yet clear how important climaticadaptations are because the mites live under the bracts of coconuts and thus in a pro-tected environment. However, in Sri Lanka N. paspalivorus inhabits relatively humid areasand N. baraki lives in dry areas whereas the two species co-occur in areas with an inter-mediate climate (Fernando and Aratchige 2009). This suggests that climatic adaptationsdo play a role, but it is not a proof and more research is needed to establish this.
We consider size-dependent access to the space under the coconut bracts as one ofthe most critical features of the predator-prey system (Aratchige et al. 2007; Lima et al.2012). The smaller size of A. guerreronis in relation to the three Neoseiulus species mostcommonly found associated with it gives it a head start in moving under the tightlyappressed bracts of young coconuts. Thus predatory mites are only expected to be ableto exert their impact on coconut mites by the time the bracts stand sufficiently distantfrom the fruit. As this process of morphological change of the bracts takes time(Aratchige et al. 2007), it comes as no surprise that in field experiments conducted inBenin and Brazil it was found that the coconut mites infest the fruits when they are only1-month old, whereas the predatory mites, N. paspalivorus and N. baraki, invade infestednuts two-three months later (Negloh et al. 2011; Galvão et al. 2011a). This delay in pred-ator impact poses a serious limitation on biological control of coconut mites. We sug-gest that this delay may be shortened by selecting coconut varieties with bracts thatexhibit a faster morphological response to coconut mite infestation.
90
CHAPTER 6
Nazer-chap6.qxd 14-9-2012 10:37 Page 90

AcknowledgementsThis research was supported by the International Institute of Tropical Agriculture (IITA)and the University of Amsterdam, the Netherlands, through a grant from the Board ofthe Netherlands Foundation for the advancement of Tropical Research (WOTRO). This studyis part of the PhD Thesis of the senior author.
ReferencesAratchige NS, Sabelis MW, Lesna I (2007) Plant structural changes due to herbivory: Do changes
in Aceria-infested coconut fruits allow predatory mites to move under perianth? Exp ApplAcarol 43: 97-107.
Athias-Henriot C (1966) Contribution à l’étude des Amblyseius paléarctiques (Acariens anactinotri-ches, Phytoseiidae). Bull Scientifique de Bourgogne 24:181-230.
Banerjee D, Gupta SK (2011) Incidence and seasonal occurrence of mites (Acari) on coconut inWest Bengal, India. Zoosymposia 6:82–85.
De Leon D (1957) Three new Typhlodromus from southern Florida (Acarina: Phytoseiidae). FlaEntomol 40:141–144.
Ehara S, Bandhoufalck A (1977) Phytoseiid mites of Thailand (Acarina: Mesostigmata). J Fac Educ,Tott Univ, Nat Sci 27:43–82.
Famah Sourassou N, Hanna R, Zannou I, Breeuwer JAJ, Moraes GJ, Sabelis MW (2012)Morphological, molecular and cross-breeding analysis of geographic populations ofcoconut-mite associated predatory mites identified as Neoseiulus baraki: evidence for crypticspecies? Exp Appl Acarol 57:15–36.
Fernando LCP, Aratchige NS (2009) Status of coconut mite Aceria guerreronis and biologicalcontrol research in Sri Lanka. Trends in Acarology - Proceedings of the 12th International Congress.In: MW Sabelis and J Bruin (eds.), Springer, Dordrecht, The Netherlands, pp. 419–423.
Galvão AS, Gondim MGC, Moraes GJ, Melo JWS (2011a) Distribution of Aceria guerreronis and Neo-seiulus baraki among and within coconut bunches in northeast Brazil. Exp Appl Acarol 54: 373–384.
Galvão AS, Gondim Jr. MGC, Moraes GJ (2011b) Life history of Proctolaelaps bulbosus feeding onthe coconut mite Aceria guerreronis and other possible food types occurring on coconut fruits.Exp Appl Acarol 53:245–252.
Helle W, Sabelis MW (1985) Spider Mites: Their Biology, Natural Enemies and Control. Vol. 1B. Elsvier,Amsterdam, The Netherlands.
Hountondji FCC, Moraes GJ, Al-Zawamri H (2010) Mites (Acari) on coconut, date palm andassociated plants in Oman. Syst Appl Acarol 15:228–234.
Jukes TH, Cantor CR (1969) Evolution of protein molecules. Mammalian Protein Metabolism. InMunro HN (ed.), Academic Press, New York, USA, pp. 21-132.
Keifer HH (1965) Eriophyid Studies B-14. Bureau of Entomology, California Department ofAgriculture, Sacramento, CA, USA, 20 p.
Krantz J, Schmutterer K, Koch W (1978) Disease, Pests and Weeds in Tropical Crops. Wiley, Chichester, 666p.Lawson-Balagbo LM, Gondim Jr, MGC, De Moraes GJ, Hanna R, Schausberger P (2008b)
Compatibility of Neoseiulus paspalivorus and Proctolaelaps bickleyi, candidate biocontrol agents ofthe coconut mite Aceria guerreronis: Exp Appl Acarol 45:1–13.
Lawson-Balagbo ML, Gondim Jr MGC, Moraes GJ, Hanna R, Schausberger P (2007) Life histo-ry of the predatory mites Neoseiulus paspalivorus and Proctolaelaps bickleyi, candidates for biolog-ical control of Aceria guerreronis. Exp Appl Acarol 34:49–61.
Lawson-Balagbo ML, Gondim MGC, Moraes GJ, Hanna R, Schausberger P (2008a) Explorationof acarine fauna on coconut palm in Brazil with emphasis on Aceria guerreronis (Acari:Eriophyidae) and its natural enemies. Bull Entomol Res 98:83–96.
91
Predators under coconut bracts: review and hypotheses
Nazer-chap6.qxd 14-9-2012 10:37 Page 91

Lever RJAW (1969) Pests of the Coconut Palm. FAO agricultural studies. No 77, Rome, 190p.Lima DB, da Silva Melo JW, Gondim MGC, Moraes GJ (2012) Limitations of Neoseiulus baraki and
Proctolaelaps bickleyi as control agents of Aceria guerreronis. Exp Appl Acarol 56:233–245.Lindquist EE, Evans GO (1965) Taxonomic concepts in the Ascidae, with a modified setal
nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem Entomol SocCan 47:1–64.
Lindquist EE, Sabelis MW, Bruin J (eds.) (1996) Eriophyoid Mites - Their Biology, Natural Enemies andControl. WCP Vol. 6, Elsevier Science, Amsterdam, The Netherlands, 790 + xxxii pp.
Lindquist EE, Krantz GW, Walter DE (2009) Order Mesostigmata. In: Krantz GW and Walter DE(Eds.), A Manual of Acarology. 3 ed., Texas Tech University Press, Lubbock, pp.124-232.
McMurtry JA, Croft BA (1997) Life-styles of phytoseiid mites and their roles in biological con-trol. Annu Rev Entomol 42:291–321.
Moore D, Alexander L (1987) Stem injection of vamidothion for control of coconut mite,Eriophyes guerreronis Keifer in St Lucia. Crop Protection 6:329–333.
Moore D, Alexander L, Hall RA (1989) The coconut mite, Eriophyes guerreronis Keifer in St. Lucia:yield losses and attempts to control it with acaricide, polybutene and Hirsutella fungus. TropPest Manag 35:83–98.
Moraes GJ, Lopez PC, Fernando CP (2004) Phytoseiid mites (Acari: Phytoseiidae) of coconutgrowing areas in Sri Lanka, with description of 3 new species. J Acarol Soc Jpn 13:141–160.
Navia D, Moraes GJ, Roderick G, Navajas M (2005) The invasive coconut mite Aceria guerreronis(Acari: Eriophyidae): origin and invasion sources inferred from mitochondrial (16S) andnuclear sequences. Bull Entomol Res 95:105–116.
Negloh K, Hanna R, Schausberger P (2012) Intraguild predation and cannibalism between thepredatory mites Neoseiulus neobaraki and N. paspalivorus, natural enemies of the coconut miteAceria guerreronis. Exp Appl Acarol 58(3) DOI 10.1007/s10493-012-9581-6.
Negloh K, Hanna R, Schausberger P (2011) The coconut mite, Aceria guerreronis, in Benin andTanzania: occurrence, damage and associated acarine fauna. Exp Appl Acarol 55:361-374.
Negloh K, Hanna R, Schausberger P (2010) Season- and fruit age-dependent population dynam-ics of Aceria guerreronis and its associated predatory mite Neoseiulus paspalivorus on coconut inBenin. Biol Control 54:349–358.
Negloh K, Hanna R, Schausberger P (2008) Comparative demography and diet breadth ofBrazilian and African populations of the predatory mite Neoseiulus baraki, a candidate for bio-logical control of coconut mite. Biol Control 46:523–531.
Oliveira DC, Moraes GJ de, Dias CTS (2012) Status of Aceria guerreronis Keifer (Acari: Eriophyidae)as a pest of coconut in the state of São Paulo, southeastern Brazil. Neotrop Entomol 41:315–323.
Reis AC, Gondim MGC, Moraes GJ, Hanna R, Schausberger P, Lawson-Balagbo LE, Barros R(2008) Population dynamics of Aceria guerreronis Keifer (Acari: Eriophyidae) and associatedpredators on coconut fruits in northeastern Brazil. Neotrop Entomol 37:457–462.
Sabelis MW (1985) Reproduction and sex allocation. In: W Helle and MW Sabelis (eds.) Spidermites - Their Biology, Natural Enemies and Control. Elsevier, Amsterdam, pp. 73-94.
Schicha E (1981) A new species of Amblyseius (Acari: Phytoseiidae) from Australia compared toten closely related species from Asia, America and Africa. Int J Acarol 7:203–2016.
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTALW: improving the sensitivity of pro-gressive multiple sequence alignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res 22:4673–4680.
Vietmeyer ND (1986) Lesser-known plants of potential use in agriculture and forestry. Science232:1379–1384.
Zannou ID, Moraes GJ, Ueckermann EA, Oliveira AR, Yaninek JS, Hanna R (2006) Phytoseiidmites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from Sub-Saharan Africa. Int JAcarol 32:241–276.
92
CHAPTER 6
Nazer-chap6.qxd 14-9-2012 10:37 Page 92

93
General discussion
In biological pest control it is not only important to identify and characterize the pestspecies, but also the natural enemies that are found in association with the target pest
species (Moraes et al. 1987; McMurtry 2010). However, the traits of the natural enemiesmay vary with the genetic entities considered. Such entities may be represented by geno-types, bio- or eco-type, geographic populations, cryptic species or species. In this thesis,I focused on geographically isolated populations and studied the extent to which thesepopulations are reproductively incompatible, because this has consequences for massrearing and for introducing a strain from one geographic area into the area of anotherstrain. For one thing mixing different geographic strains in mass culture may have neg-ative effects on population growth rates if the strains are reproductively incompatible(Stouthamer et al. 2000; Mochiah et al. 2002). For another, reproductive incompatibilityensures reproductive isolation, thereby possibly maintaining the genetic integrity of thegeographic strain even after it has been released in the geographic area of another strain.Having established the reproductive isolation between geographic populations of thenatural enemies (Chapters 2, 3 and 4), I reviewed and assembled data to demonstrate theextent to which these geographic populations exhibit genetic variation in traits relevantto biological pest control. Moreover, by reconstructing the invasion routes of the natu-ral enemies I tried to determine their geographic area of origin, which most likely is thearea where they exhibit the greatest genetic diversity. In this way it is possible to planreleases of natural enemies in other areas where their genetic variation is limited.
So, what can be learnt from the results presented in this thesis with respect to geo-graphic populations of mainly two species of predatory mites (Acari: Phytoseiidae) thatare found in close association with coconut mites under the bracts of coconuts? I havepresented evidence for reproductive isolation between geographic populations of thepredatory mite Neoseiulus paspalivorus and N. baraki and showed that this was due to thepresence of endosymbionts in one of the two predator species (N. paspalivorus)(Chapters 2 and 3). Moreover, I showed that the other predator species, N. baraki, isactually a species complex with two cryptic species, one from Benin and the otheroccurring in Tanzania, Brazil and Sri Lanka (Chapter 4). I also argued that a third pred-ator species, N. neobaraki, also belongs to this complex (Chapter 6). Phylogenetic analy-sis of all Neoseiulus species together (plus an outgroup) suggests that the centre of genet-ic diversity of Neoseiulus species is in Africa, but there were no differences in geneticdiversity among populations of N. paspalivorus from different countries (Chapter 6). Thisled to the hypothesis that N. baraki moved from weeds to coconut palms in Africa andthat one cryptic species from Africa invaded the other continents, whereas N. pas-
7Nazer-chap7.qxd 14-9-2012 10:37 Page 93

palivorus from South-America followed the same invasion route as proposed for its prey,the coconut mites (Navia et al. 2005). If this hypothesis holds, then it makes sense tointroduce geographic strains of N. baraki from Africa into other continents, but thereis as yet no reason to do that with strains of N. paspalivorus. Now the question ariseswhich traits are relevant to biological control and whether geographic populations dif-fer in these traits. While this still requires an in-depth analysis, it is good to identify somegeneral trends that may serve as a hypothesis. First, I assumed that two traits are of par-ticular importance: the population growth rate on a diet of coconut mites and the abil-ity to utilize pollen for survival. Then, I observed a trend in that high values of the firsttrait are associated with lower abilities concerning the second trait (and vice versa).Thus, it seems that there is no master of all trades (good in population growth and inpollen utilization) and that there is a trade-off between two traits that I assumed to becrucial for biological control. Unfortunately, there is no hard theory stating which com-bination of the two traits is best to achieve (short- or long-term) biological control ofcoconut mites. In absence of such a theory there is a need to perform experimental testswith strains exhibiting contrasting combinations of the two traits.
Apart from testing which combination of the two traits is best for biological con-trol there are several other issues that need scrutiny before accepting the hypothesesproposed in this Thesis. First, I would recommend doing research that establishes themetabolic and/or genetic reasons for a trade-off between population growth on a dietof coconut mites and the ability to utilize coconut pollen for survival and development.Since the presence of endosymbionts may come at a cost to reproduction (Stouthameret al. 1999; Bordenstein and Werren 2000), one may wonder how an individual predato-ry mite can profit from an infection by endosymbionts, unless the endosymbiont con-fers an advantage, e.g. in digestion of proteins from coconut pollen. Second, I wouldfavour carrying out a more extensive sampling program to test our hypotheses on thecentres of genetic diversity (and consequently the invasion routes). Third, I haveassumed that the reproductive isolation I have assessed indeed leads to reproductive iso-lation in the field. If a geographic strain from one area would be released in the area ofanother, can I assume that reproductive isolation is retained? Clearly, this is a questionthat needs further research because there is evidence that mating preferences depend onthe environment (e.g. diet) (e.g. Kokko et al. 2002) and it is still a major question whetherthe impact of endosymbionts is under control of the endosymbionts, their host or both.
ReferencesBordenstein SR, Werren JH (2000) Do Wolbachia influence fecundity in Nasonia vitripennis? Heredity,
84:54–62.Stouthamer R, Jochemsen P, Platner GR, Pinto JD (2000) Crossing incompatibility between
Trichogramma minutum and T. platneri (Hymenoptera: Trichogrammatidae): implications forapplications in biological control. Environ Entomol 29:832–837.
94
CHAPTER 7
Nazer-chap7.qxd 18-9-2012 2:51 Page 94

Moraes GJ (1987) Importance of taxonomy in biological control. Insect Sci Appl 8:841–844.Muma MH, Denmark HA (1969) Sibling species of Phytoseiidae (Acari: Mesostigmata). Fla
Entomol 52(2):67–72.McMurtry JA, Badii MH, Congdon BD (1985) Studies on a Euseius species complex on avocado
in Mexico and Central America, with a description of a new species (Acari: Phytoseiidae). IntJ Acarol 26:107–116.
Navia D, Moraes GJ, Roderick G, Navajas M (2005) The invasive coconut mite Aceria guerreronis(Acari: Eriophyidae): origin and invasion sources inferred from mitochondrial (16S) andnuclear sequences. Bull Entomol Res 95:105–116.
Mochiach MB, Ngi-Song AJ, Overholt WA, Stouthamer R (2002) Wolbachia infection in Cotesiasesamiae (Hymenoptera: Braconidae) causes cytoplasmic incompatibility: implications for bio-logical control. Biol Control 25:74–80.
Kokko H, Brooks R, McNamara JM, Houston AI (2002) The sexual selection continuum. Proc RSoc Lond B 269:1331–1340.
95
General discussion
Nazer-chap7.qxd 14-9-2012 10:37 Page 95

Nazer-chap7.qxd 14-9-2012 10:37 Page 96

97
Summary
Ever since its discovery as a pest of coconut palms in Mexico in 1965, the coconutmite Aceria guerreronis Keifer has been reported from other countries, first in South-
America, then in Africa and finally also in Asia. With the aim to establish biological con-trol of this pest, a search for candidate natural enemies associated with this pest hasbeen conducted in several countries in each of these geographic areas. Prior to the workdescribed in this Thesis, two species of predatory mites (Acari: Phytoseiidae) werefound to co-occur in Brazil, Tanzania, Benin and Sri Lanka and, based solely on mor-phological criteria, they have been identified as Neoseiulus paspalivorus DeLeon and N.baraki Athias-Henriot. Because these species inhabit widely different geographic areas,there may be genetic variation between geographic populations of each of these twospecies that is worth to be explored in order to improve the effectiveness of biologicalcontrol. The aim of the work presented in this Thesis is to assess the genetic entities(biotypes, cryptic species, subspecies and species) that should be considered for theirimpact on biological control. Therefore, this Thesis is largely a biosystematic study thatserves as a basis for future studies on biological control. This is done by a combinationof morphological measurements, crossbreeding experiments and molecular analysis.
It was found that geographic populations of Neoseiulus paspalivorus were morpho-logically similar, yet reproductively incompatible (CHAPTER 2). Females of each popula-tion were able to mate with males from the other populations, but they laid fewer eggsand most of these eggs did not hatch because they shrivelled. Moreover (CHAPTER 3),the populations were genetically identical, based on the mitochondrial cytochrome oxi-dase subunit I (CO1-sequences). Then, using the 16S rDNA primers, the three popula-tions were checked for the presence of two endosymbiotic bacteria, Wolbachia andCardinium known to cause reproductive abnormalities in their hosts, and their phyloge-netic relationships were analysed (CHAPTER 3). Different Wolbachia strains were detect-ed in the populations from Benin and Brazil whereas a population from Ghana har-boured only Cardinium symbionts. An antibiotic test showed that removing the sym-bionts from females of each population made them incompatible with males of thesame population. Moreover, these tests restored compatibility between the populationsfrom Benin and Brazil in one direction, but not in the reciprocal cross, suggestingincompatibility arising from interactions between nuclear and cytoplasmic genes.Antibiotic-treated females of the population from Ghana produced hardly any progeny,so that we were not able to establish a Cardinium-free line and hence no crossings withother populations. Given the symbiont-associated reproductive isolation, genetic differ-entiation between the three populations remains a possibility worth to be explored for
Nazer-summary.qxd 14-9-2012 10:38 Page 97

traits relevant to biological control. However, we conclude that the three populationsbelong to a single species.
Following the same approach we found that geographic populations of predatorymites, previously identified as N. baraki, are reproductively incompatible, differ genetical-ly with respect to mitochondrial CO1 sequences and some morphological traits (i.e.,number of teeth on the fixed digit of female chelicera) and therefore do not belong toone species. In particular, the population from Benin was distinct from the other popu-lations studied (Brazil, Tanzania and possibly also Sri Lanka) and may represent a crypticspecies (CHAPTER 4). Because scrutiny of the holotype was not possible, it cannot beinferred which of the two cryptic species is the real N. baraki. Nevertheless, these genet-ic entities seem worth to be explored for genetic traits relevant to biological control.
Whereas in each of the geographic areas studied N. paspalivorus co-occurs with oneor the other cryptic species closely related to N. baraki and in Africa also with anotherpredator species, i.e., N. neobaraki, they may also co-occur in the same palm tree.However, they almost never co-occur within the same bunch of coconuts and on thesame coconut. This lack of coexistence at a local scale, yet apparent coexistence at alarger scale, may arise from priority effects emerging in Lotka-Volterra competitionmodels for two species competing for the same resource, and in models of reciprocalintraguild predation. However, the cross-pairing studies (CHAPTER 5) described in thisThesis also revealed another mechanism preventing local coexistence. Cross-pairingsbetween species revealed intraguild predation by females of one predator species on themales and larvae of the other. Since size determines who is the intraguild predator andwho is the intraguild prey, the largest of the three predator species (N. neobaraki) wasmore aggressive than the smallest (N. paspalivorus). Such intraguild interactions maydrive the intraguild prey extinct not only because of the mortality inflicted on theintraguild prey, but also because it reduces the chances for females of the intraguild preyto find a male.
In the final part of the Thesis (CHAPTER 6) an up-to-date review is provided of allspecies of predatory mites found under the bracts of coconuts. Out of a total of 60species, 32 species belong to the Phytoseiidae which in terms of their abundance repre-sent 53% of all specimens found. Among the Phytoseiidae three species stand out asbeing predominant: N. paspalivorus, N. baraki and N. neobaraki. In most geographic areasstudied the most dominant of these three is N. paspalivorus, possibly because it may havebetter access to the area under the bracts of the coconut due to its smaller size.However, in Tanzania, N. neobaraki, the largest of the three predator species, dominates,but the reasons for this dominance are still unclear. Virtually always the predators withintermediate abundance are the two cryptic species closely related to N. baraki, being thepredator of intermediate size. For two of the abundant predator species, the biologicalcharacteristics are summarised, as far as available in the literature, and supplied, where
98
Summary
Nazer-summary.qxd 18-9-2012 2:53 Page 98

no data are available. This shows that N. baraki and N. paspalivorus from Brazil tend tohave higher capacity for population increase on a diet of coconut mites than the sametwo species from Africa. However, the cryptic species closely related to N. baraki fromBenin is better able to survive and reproduce on coconut pollen than that from Brazil.Compared to species closely related to N. baraki, populations of N. paspalivorus haveequal or higher capacities for population increase on a diet of coconut mites and theyalso vary in the ability to utilize coconut pollen. The populations from Ghana and Brazil(Acarau) are relatively better in pollen utilization whereas that from Benin is relativelyworse. Overall, there is a trend for populations to have lower capacities for populationincrease (on a diet of coconut mites) when they are better in utilizing pollen. Which ofthese combinations is best for biological control of coconut mites is still to be deter-mined.
99
Summary
Nazer-summary.qxd 14-9-2012 10:38 Page 99

Nazer-summary.qxd 14-9-2012 10:38 Page 100

101
Samenvatting
Sinds zijn ontdekking in 1965 als plaag van kokospalm in Mexico is de kokosnootmijtAceria guerreronis Keifer gemeld van andere landen, eerst in Zuid-Amerika, toen in
Afrika en ten slotte ook in Azië. Met als doel de biologische bestrijding van deze plaag-mijt is in verscheidene landen op elk van de drie continenten gezocht naar natuurlijkevijanden die samen met A. guerreronis voorkomen. Voorafgaand aan het werk datbeschreven staat in dit proefschrift waren twee roofmijtsoorten (Acari: Phytoseiidae)gevonden in Brazilië, Tanzania, Benin en Sri Lanka – op basis van alleen morfologischekenmerken zijn de roofmijten geïdentificeerd als Neoseiulus paspalivorus DeLeon and N.baraki Athias-Henriot. Omdat deze soorten voorkomen in zeer verschillende geografi-sche gebieden zou er wel eens genetische variatie kunnen bestaan tussen geografischepopulaties, die van belang kan zijn voor verbetering van de effectiviteit van biologischebestrijding. Doel van het werk beschreven in dit proefschrift is het bepalen van degenetische eenheden (o.a. biotypes, cryptische soorten, ondersoorten, ‘echte’ soorten)die relevant zijn binnen de context van biologische bestrijding. Daarom beslaat ditproefschrift in de eerste plaats een biosystematische studie – bestaande uit een combi-natie van morfologische metingen, kruisingsproeven en moleculaire analyse – die alsbasis kan dienen voor toekomstig werk aan biologische bestrijding.
Geografische populaties van N. paspalivorus bleken weliswaar morfologisch eender,maar reproductief incompatibel: vrouwtjes van elke populatie waren in staat om teparen met mannetjes van de andere populaties, maar ze legden daarna minder eieren, diein meerderheid verschrompelden (HOOFDSTUK 2). Genetisch – op basis van mitochon-driale CO1 sequenties – bleken de populaties identiek (HOOFDSTUK 3). Vervolgens zijnde drie populaties met behulp van 16S rDNA primers onderzocht op het voorkomenvan twee endosymbiontische bacteriën, te weten Wolbachia en Cardinium, waarvan be-kend is dat ze reproductieve abnormaliteiten bij hun gastheren kunnen veroorzaken.Ook zijn de fylogenetische relaties geanalyseerd (HOOFDSTUK 3). VerscheideneWolbachia-stammen werden gevonden in de populaties uit Benin en Brazilië, terwijl eenpopulatie uit Ghana alleen Cardinium-symbionten bleek te bevatten. Uitschakeling van desymbionten door middel van antibiotica maakte vrouwtjes van elke populatie incompa-tibel met mannetjes van de eigen populatie (dat wil zeggen: paring van vrouwtjes enmannetjes leidde niet tot nakomelingen). Bovendien bleek dat met deze proeven decompatibiliteit tussen populaties uit Benin en Brazilië hersteld werd in één richting, maarniet in de reciproke richting (dus wel succesvolle paring tussen vrouwtjes van de ene enmannetjes van de andere populatie, maar niet tussen mannetjes van de ene en vrouwtjesvan de andere). Een dergelijke uitkomst wijst erop dat de incompatibiliteit het gevolg is
Nazer-samenvatting.qxd 14-9-2012 10:38 Page 101

van de wisselwerking tussen genen uit de kern en genen uit het cytoplasma. Met anti-biotica behandelde vrouwtjes van de Ghanese populatie produceerden nauwelijksnakomelingen, zodat we geen Cardinium-vrije lijn van mijten konden kruisen met anderepopulaties. De met de symbionten samenhangende reproductieve isolatie geeft aan datgenetische differentiatie tussen de drie populaties mogelijk is en dat het de moeite waardblijft om verschillen in eigenschappen te onderzoeken die van belang zijn voor de bio-logische bestrijding. Wij concluderen echter dat de drie populaties behoren tot eenenkele soort.
Op vergelijkbare wijze hebben we gevonden dat geografische populaties van deroofmijt N. baraki reproductief incompatibel zijn en genetisch verschillen op basis vanmt-CO1 sequenties en enkele morfologische eigenschappen (zoals het aantal tanden opde onbeweeglijke helft [fixed digit] van de vrouwelijke monddelen [chelicera]) – dezepopulaties behoren daarom niet tot een enkele soort. Vooral de Beninese populatie weekaf van de andere populaties (uit Brazilië, Tanzania en mogelijk ook Sri Lanka) en er zouwel eens sprake kunnen zijn van twee cryptische soorten (HOOFDSTUK 4). Omdatbestudering van het holotype niet mogelijk was, kunnen we niet beslissen welke van detwee cryptische soorten de echte N. baraki is. Hoe dan ook, deze twee genetische een-heden lijken relevant voor vervolgonderzoek naar verschillen in eigenschappen die vanbelang zijn voor de biologische bestrijding.
In elk van de geografische gebieden komt N. paspalivorus voor samen met (de eneof de andere cryptische soort die nauw verwant is met) N. baraki en in Afrika ook meteen tweede roofmijt, namelijk N. neobaraki. Deze soorten kunnen samen voorkomen opeen enkele palmboom, ze worden echter vrijwel nooit samen gevonden op een enkelekokosnoot of een enkele tros kokosnoten. Coexistentie op een regionale schaal, maarniet op lokale schaal, kan tot stand komen door voorrangseffecten (priority effects) bin-nen Lotka-Volterramodellen voor competitie tussen twee soorten om een enkele‘resource’, en in modellen voor wederkerige predatie binnen voedingsgilden (reciprocalintraguild predation). De paringsproeven beschreven in HOOFDSTUK 5 van dit proef-schrift brachten echter nog een ander mechanisme voor het tegengaan van plaatselijkecoexistentie aan het licht. Paringen tussen soorten toonden het bestaan aan van(intraguild) predatie door vrouwtjes van de ene roofmijtsoort op mannetjes en larvenvan de andere. Aangezien grootte bepalend is voor de rolverdeling tussen (intraguild)predator en prooi, was de grootste van de drie roofmijtsoorten (N. neobaraki) agressie-ver dan de kleinste soort (N. paspalivorus). Dergelijke ‘intraguild’ interacties kunnen lei-den tot het verdwijnen van de (intraguild) prooi, niet alleen door sterfte als gevolg vanvraat maar ook door het verkleinen van de kans voor vrouwtjes van de (intraguild) prooiom een mannetje te vinden om mee te paren.
In het laatste deel van het proefschrift (HOOFDSTUK 6) wordt een actueel overzichtgegeven van alle soorten roofmijten die gevonden worden in de nauwe ruimte onder de
102
Samenvatting
Nazer-samenvatting.qxd 14-9-2012 10:38 Page 102

huid van de kokosnoot. Van de in totaal 60 soorten behoren er 32 tot de familiePhytoseiidae (of te wel 53% van alle individuele roofmijten). Binnen de Phytoseiidaedomineren drie soorten: N. paspalivorus, N. baraki en N. neobaraki. In de meeste gebiedenoverheerst van deze drie soorten N. paspalivorus, mogelijk omdat mijten van deze soorthet kleinst zijn en dus het makkelijkst toegang hebben tot de nauwe ruimte onder dehuid van de kokosnoten. In Tanzania overheerst echter N. neobaraki, de grootste van dedrie roofmijtsoorten – de verklaring daarvoor is nog onduidelijk. Vrijwel in alle gevallenzijn de mijten van (de twee cryptische soorten nauw verwant aan) N. baraki zowel inter-mediair in grootte als in aantal. De biologische eigenschappen van de twee meestvoorkomende roofmijtsoorten worden opgesomd, voor zover bekend uit de literatuurof uit eigen onderzoek. Dit overzicht laat zien dat N. baraki en N. paspalivorus uit Braziliëeen grotere populatiegroeipotentie hebben op een dieet van kokosnootmijten dandezelfde twee soorten uit Afrika. Neoseiulus baraki (de twee cryptische soorten) uit Beninis echter beter in staat te overleven en zich voort te planten op een dieet van kokospalm-stuifmeel dan die uit Brazilië. Vergeleken met (de twee soorten nauw verwant aan) N.baraki hebben populaties van N. paspalivorus een gelijke of grotere populatiegroeipoten-tie op een dieet van kokosnootmijten en ze variëren in de mate waarin ze zich kunnenvoeden met kokospalmstuifmeel. De populaties uit Ghana en Brazilië (Acarau) kunnenverhoudingsgewijs goed overweg met stuifmeel, terwijl de populatie uit Benin er naarverhouding slecht in is. Al met al lijkt het erop dat populaties een kleinere groeipotentiehebben op een dieet van kokosnootmijten naarmate ze beter overweg kunnen met stuif-meel. Welke combinatie van eigenschappen het beste is voor de biologische bestrijdingvan de kokosnootmijt valt nog te bezien.
103
Samenvatting
Nazer-samenvatting.qxd 14-9-2012 10:38 Page 103

Nazer-samenvatting.qxd 14-9-2012 10:38 Page 104

105
Curriculum vitae
Nazer Famah Sourassou was born on December 31st, 1973 in Paratao (Togo). He start-ed his school education in 1979, and obtained his primary school certificate ‘C.E.P.D.’in 1985, and his secondary school certificate (B.E.P.C) in 1989. In the same year, hestarted his high school education at ‘Lyceé d’Enseignement General de Sokodé’ wherehe graduated with high school diploma ‘Baccalauréat D’. He started his university studiesat the University of Lomé (Lomé, Togo) in 1994, stayed one year at the Faculty ofMedical Science, before moving to the Faculty of Agriculture ‘Ecole Superieured’Agronomie’. In November 2001, he graduated with ‘Diplôme d’Ingénieur Agronome’. Heworked as research consultant at IITA-Benin till September 2005. In the same year hewas granted a scholarship by the Vlaamse Interuniversitaire Raad (VLIR) to undertakea 2-year master programme in molecular biology in Belgium conjointly organized by theUniversity of Brussels and the Catholic University of Leuven. He started in October2005 and graduated in September 2007 with a diploma ‘Master of Science in molecular biol-ogy’. In 2008, he was granted a fellowship by the Netherlands Foundation for theAdvancement of Tropical Research (WOTRO) and the International Institute ofTropical Agriculture (IITA). He started his PhD-project May 1st, 2008 at the Instituteof Biodiversity and Ecosystem Dynamics (IBED), University of Amsterdam. In 2011,he was awarded a Donald E. Johnston fellowship to participate in the InternationalAcarology Course held at Ohio State University, Columbus, USA.
Nazer-CV+Acknowl.qxd 14-9-2012 10:38 Page 105

Acknowledgements
Iwould like to express my thanks to a number of people who in one way or anothercontributed to the preparation and completion of this Thesis. It is to them that I owe
my deepest gratitude.First and foremost, I would like to express my deep and sincere gratitude to my
Promotor, Professor Maurice W. Sabelis, who despite his many other academic and pro-fessional commitments was available every time I need him. His detailed and construc-tive comments have provided a good basis for the present thesis. His personal guidanceand important support throughout this work have been of great value for me. I appre-ciated his great effort to explain things simply and clearly. Special thanks for giving theopportunity to attend the Acarology Summer Course in Columbus, Ohio, USA, and the7th European Association of Acarology (EURAAC) Congress in Vienna, Austria.
I am deeply grateful to my supervisor Dr. Rachid Hanna for giving me the oppor-tunity to do this PhD study. The good advice, support and guidance throughout thisstudy have been invaluable on both academic and personal level, for which I amextremely grateful. Thank you for your friendship, assistance over the past years.
I also owe much gratitude to Dr. Hans Breeuwer, for allowing conducting mymolecular work in his laboratory. His extensive and constructive discussions around mywork have been very helpful.
In the laboratories, I have been aided by many some people which I would like toexpress my gratitude: Richard Houndafoche, Pierre Sovimi, Bovis Bonaventure at thelaboratory of IITA-Benin for their assistance in maintaining predatory mites and con-ducting experiences; Betsie Voetdijk, Peter Kuperus and Lin Dong for their assistanceduring molecular work at the University of Amsterdam. I also deserve my sincerestthanks to my colleagues from the International institute of Tropical Agriculture (IITA-Benin), and from the Institute of Biodiversity of Ecosystem Dynamics (IBED),University of Amsterdam. Their friendship and assistance has meant more to me thanI could ever express.
The financial support of the Board of the Netherlands Foundation for theadvancement of Tropical Research (WOTRO) is gratefully acknowledged.
Finally, I would like to express my greatest thanks to my family members, especial-ly my wife Rahamatou Issaka and my sons Abdul-Raouf and Abdul-Madjid. Their infi-nite patience, encouragements and enthusiasm were important for the completion ofthis Thesis. A vous tous, je vous dis infinément merci.
Nazer-CV+Acknowl.qxd 14-9-2012 10:38 Page 106

Bio-systematics of predatory mites used for control of the coconut mite
Nazer Fam
ah SourassouBio-system
atics of predatory mites used for control of the coconut m
ite
Nazer Famah Sourassou