vaccine discovery and development: lessons from covid-19
TRANSCRIPT

Vaccine Discovery and Development:
Lessons from COVID-19
Sponsored by:

Translate Insights Into Novel Strategies for Vaccine ResearchInnovative Solutions for Extraordinary TimesThe COVID-19 pandemic has scientists developing novel life-saving treatments at breakneck speed. Sartorius provides innovative high-throughput & automated solutions for faster vaccine discovery, characterization & development.
Our groundbreaking Octet®, iQue® and Incucyte® advanced cell & protein analysis platforms provide insights at unprecedented speed, depth & scale, streamlining workflows to accelerate results - when it counts.
www.sartorius.com/vaccine-discovery

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 1
Vaccine Discovery and Development Lessons from COVID-19This eBook was developed and produced by Current Protocols, a Wiley Brand, in partnership with Sartorius
eBook written by Katherine L. Molnar-Kimber, Ph.D.
Cover image © Sartorius
Copyright © 2021 Wiley Periodicals, Inc. All rights reserved. No part of thispublication may be reproduced, stored or transmitted in any form or by anymeans without the prior permission in writing from the copyright holder.

2 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
Table of ContentsTable of Contents
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3History of vaccine development for Emerging Infectious Diseases . . . . . . . . . . . . . 4
The COVID-19 Challenge . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6Introduction to COVID-19 disease and its causative virus, SARS-CoV-2 . . . . . . . . . . 6Vaccine Discovery & Development Workflow . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
Vaccine platforms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8COVID-19 Vaccines that Received Emergency Approval in 11 Months . . . . . . . . . 9
Target Discovery . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .10Comparison with similar viruses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .10
Viral protein(s) and cell receptor(s) essential for viral entry and infection . . . . .10Antibody responses from convalescent COVID-19 patients . . . . . . . . . . . . . . . . . . .11
Antigen/Immunogen discovery . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .13
Discovery Applications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .14Biology Discovery Processes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .14
Cell Viability /Proliferation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .15Infectivity assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .16Viral titers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .16Cellular-viral interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .16Pseudotyped viruses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .17Viral variants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .17
Assessment of immune responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .17Antibody responses and characterization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .17Epitope binning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .18Macrophages . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .18T cell responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .18Antiviral Therapeutics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .19
Summary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .21
For more information: . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .22
Selected References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .23
Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 3
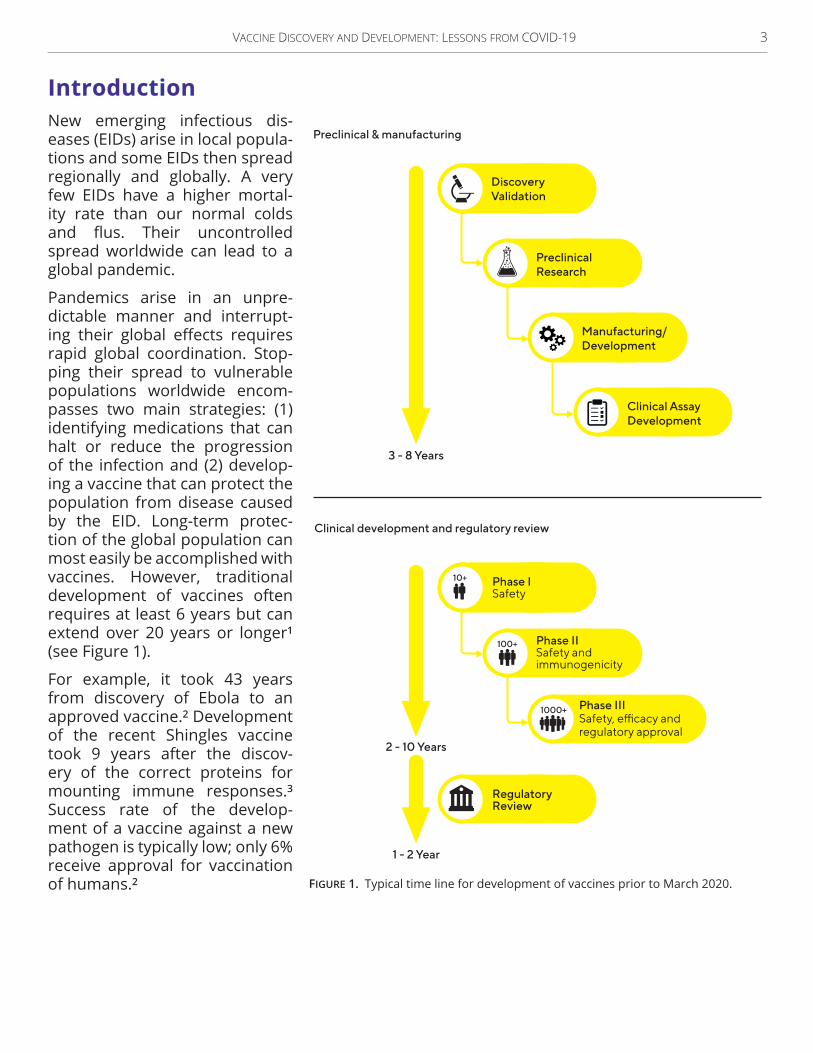
Introduction Introduction New emerging infectious dis-eases (EIDs) arise in local popula-tions and some EIDs then spread regionally and globally . A very few EIDs have a higher mortal-ity rate than our normal colds and flus. Their uncontrolled spread worldwide can lead to a global pandemic .
Pandemics arise in an unpre-dictable manner and interrupt-ing their global effects requires rapid global coordination . Stop-ping their spread to vulnerable populations worldwide encom-passes two main strategies: (1) identifying medications that can halt or reduce the progression of the infection and (2) develop-ing a vaccine that can protect the population from disease caused by the EID . Long-term protec-tion of the global population can most easily be accomplished with vaccines. However, traditional development of vaccines often requires at least 6 years but can extend over 20 years or longer1
(see Figure 1) .
For example, it took 43 years from discovery of Ebola to an approved vaccine .2 Development of the recent Shingles vaccine took 9 years after the discov-ery of the correct proteins for mounting immune responses .3 Success rate of the develop-ment of a vaccine against a new pathogen is typically low; only 6% receive approval for vaccination of humans .2
Preclinical & manufacturing
Clinical development and regulatory review
3 - 8 Years
2 - 10 Years
DiscoveryValidation
PreclinicalResearch
Clinical AssayDevelopment
Manufacturing/Development
1 - 2 Year
RegulatoryReview
Phase ISafety
Phase IISafety andimmunogenicity
Phase IIISafety, efficacy andregulatory approval
1000+
100+
10+
Figure 1. Typical time line for development of vaccines prior to March 2020 .

4 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
History of vaccine development for Emerging Infectious Diseases History of vaccine development for Emerging Infectious Diseases Vaccines have been used for centuries . Highlights of some discoveries and milestones for devel-opment of the different types of vaccines are shown in Figure 2.
16th centuryChinese description
inoculations (smallpox)
1853 UKMandatory smallpox
vaccinations
1892-1898Discovery First virus
1944First Flu vaccine
1950-1956 First combinationvaccine (DTP)
1960’sFirst vaccine grownon human cells inculture (Ad4, Ad7)
2019First RecombinantVSV based vaccine
(Ebola)
2020First Ad26-based
vaccine (Ebola)
2021Ad26-based vaccine
Emergency use(SARS-CoV-2)
1796 Jenner:1st scientificmethod vaccination
1885 Pasteur First live virusvaccine (rabies)
1920-1926TB, diphtheria, tetanus,Whooping cough vaccines
1951- 2000+Polio vaccines grown
in cultured Monkey cells,amended
1986 First genetically
engineered vaccine (HBV)
2019First NonReplicatingsmallpox vaccine
2020 First mRNA vaccines;Emergency use(SARS-CoV-2)
Figure 2. Major milestones in the history of vaccine development . Ad4, adenovirus serotype 4; Ad7, adenovirus type 7; Ad26, adenovirus serotype 26; DTP, diphtheria-tetanus-pertussis vaccine; HBV, Hepatitis B virus; TB, tuberculosis; UK, United Kingdom; VSV, recombinant vesicular stomatitis virus vector.

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 5
Scientists continue to use recombinant DNA techniques to expand the information around pathogens and the host-pathogen interactions . Here is a brief summary of milestones that involved recombinant DNA techniques in vac-cine development:
• In 1986 for safety reasons, the originally approved Hepatitis B virus (HBV) vaccine which contained human Hepatitis B Sur-face antigen particles isolated from infected humans was replaced by Hepatitis B Surface antigen particles produced by recombinant DNA technology in yeast cells .
• In 2019, the FDA approved the first nonrep-licating vaccinia virus (cowpox) as a vaccine for smallpox .
• In 2019, the first recombinant attenuated replication competent Vesicular Stomatitis virus (VSV) vectored vaccine was approved by European Medicines Agency and the FDA .4 It expresses Ebola glycoprotein G and protects against Ebolavirus induced disease .
• In 2020, the first recombinant Adenovirus type 26 based vaccine was approved for marketing by the European Medicine Agency for Ebola. It is a two dose regimen with the first dose being Ad26 vector expressing Ebola glycoprotein G. The second dose was the nonreplicating vaccinia vector express-ing glycoproteins from 3 distinct strains of Ebola and the nucleoprotein from a fourth Ebola strain .
• In 2020, the first mRNA vaccines were approved for Emergency Use by the FDA to prevent against COVID-19 disease caused by SARS-CoV-2 .
• In 2021, two Adenovirus serotype (Ad26) based vaccines were approved for Emer-gency Use by the FDA or World Health Organ-ization (WHO) to prevent against COVID-19 disease caused by SARS-CoV-2 .

6 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
The COVID‑19 Challenge The COVID‑19 Challenge Here we present the story of COVID-19 which began disrupting global health in early 2020 . The WHO declared COVID-19 as a Pandemic on Mar 11, 2020 .5 International cooperation among researchers at government institutes, foundations, biotech companies, pharmaceu-tical firms, and publishers helped accomplish the development of several efficacious vac-cines that received European Medicine Agency and US FDA approval for emergency use within 15 months and subsequent approval from other countries (eg, Australia, Japan) . The coor-dinated development process greatly bene-fited from quick open access to all articles on COVID-19 . Its main preclinical steps included:
• Discovery and characterization of the causa-tive agent SARS-CoV-2,
• Characterization of the immune responses of people who recovered,
• Engineering and evalu-ation of various vaccine strategies, and
• Use of supportive research tools and equipment.
We further describe high-lights of the COVID-19 story to illustrate how researchers can streamline their devel-opmental cycles for vaccines and accomplish the develop-ment of efficacious vaccines, possibly within 1-2 years.
Introduction to Introduction to COVID‑19 disease and COVID‑19 disease and its causative virus, its causative virus, SARS‑CoV‑2SARS‑CoV‑2In late 2019, several cases of a new type of pneumonia were described in China .6‑8 The emerging infectious disease (EID) was identified as a new type of beta coronavirus on January 9, 2020. It was initially
called 2019-nCoV and eventually was formally named SARS-CoV-2 .8 Within a month, more than 2000 cases had been confirmed.9 The ini-tial sequences of the SARS-CoV-2 genome were released on Jan 10 by the Chinese Center for Disease Control and Prevention website and were globally disseminated on Jan 12th by the Global Initiative on Sharing all Influenza Data (GISAID) .10 The complete sequences were pub-lished online on Jan 29, 20209 and Feb 3, 2020 .11
This positive strand RNA genome of 29 .9kb is protected by its nucleoprotein (N) and a lipid bilayer envelope containing three proteins: the spike protein (S), membrane protein (M) and envelope protein (E), as shown in Figure 3.
Its sequence also encodes for at least 16 non-structural proteins that are involved in replica-tion of the genome, production of its structural
Envelope proteins
Spike protein
Nucleocapsid proteinscovering RNA genome
Lipid membrane
Membrane protein
Structure of SARS-CoV-2
Figure 3. Structure of SARS-CoV-2 virus .

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 7
proteins, and modulation of host functions such as splic-ing, translation, and protein trafficking.12 Similar to other viruses, concentration and purification of coronavirus and its protein components can be accomplished with ultrafiltra-tion for further analysis .13
To gauge the needed immune response for a vaccine, scien-tists measured the immune responses of patients who recovered from COVID-19 . Convalescent patients con-tained neutralizing antibod-ies to the spike protein and T cell responses .14 The titers of antibodies in the convalescent sera were compared to titers induced by vaccine-inoculated participants in the clinical trial . Thus, concrete goal posts for developing SARS-CoV-2 vac-cine were now known.
Vaccine Discovery & Vaccine Discovery & Development WorkflowDevelopment WorkflowBecause of the urgency for developing a vaccine to pro-tect against COVID-19 disease, many companies rushed to use their unique vaccine plat-form to develop a vaccine can-didate to prevent COVID-19 disease . Five of 7 strategies have been used to produce vaccines against other dis-eases. Two strategies (DNA vaccines and RNA vaccines) are being developed as vac-cine platforms against other diseases as well as COVID-19. Here we summarize the seven basic strategies being used to develop vaccines against COVID-19:
SARS-CoV-2
Immune responsemachinery
Immune response
Anti-coronavirusantibodies are
generated
T cell immunity
Infection
Weakenedvirus
Inactivated virus
Replicating virusvector (eg. VSV)coated with viral protein
Non replicating virusvector (eg. AAV) thatexpresses viral proteins
Spike (S)protein
DNA vaccine(with S gene)
mRNA vaccine(with S message)
Vaccine strategies
Strategies for COVID-19 Vaccine Development
Figure 4. Universal vaccine strategies directed against COVID-19.

8 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
Vaccine platforms Vaccine platforms Weakened or attenuated viruses have been used as vaccines against numerous diseases including smallpox and polio . In some cases, like smallpox, a related virus (cowpox, vaccinia) with less ability to cause severe disease is used as the inoculum . Inoculated people then had some protection against the more virulent smallpox . Traditionally attenuated viruses were devel-oped and selected by serial passages through appropriate hosts (specific cell lines or species). Several recent efforts have involved analysis of the sequenced genome, hypotheses on dis-ease-associated genes, synthesis of potentially attenuated viral genome, packaging in virions, and extensive testing . The testing includes char-acterization of its ability to (1) infect and cause preferably minimal disease (pathogenesis), (2) induce a protective immune response against the parent virus, and (3) exhibit an acceptable safety profile, stability during manufacturing, and high yield during manufacturing .
Inactivated virus vaccines have been used for decades, including the Salk polio vaccine which emphasized the importance of complete inac-tivation. To maintain efficacious inactivation, current practices include removing any clumps in the batch before the inactivation process . Subsequently, a sample of each manufactured lot undergoes thorough testing for safety and efficacy.
Replicating viral vector that expresses an immunogenic protein of the disease-causing virus such as Ebola often can induce neutral-izing antibodies and cellular immunity . Advan-tages of the replicating viral vector platforms include their ability to immunologically process the specific target protein from a pathogenic virus in infected cells without exposure to the whole pathogenic virus. A second advantage is that some vectors can accept and express 3 exogenous proteins, thereby protecting against 3 distinct viruses .15
Nonreplicating viral vector, such as a recom-binant Adenovirus 26 vector and /or a recom-binant, modified vaccinia vector, can express an immunogenic protein of the disease-caus-ing virus such as Ebola 16, 17 or SARS-CoV-2 .18‑21
Advantages of nonreplicating viral vector plat-forms are their abilities to induce neutralizing antibodies and cell mediated immunity and their lack of risk for overgrowth in immunosup-pressed patients .
Protein‑based vaccines, such as Hepatitis B virus (HBV) surface antigen particles, have been used for decades to elicit immunity to HBV . The immunogenic Spike protein of SARS-CoV-2 can be formulated in nanoparticles with an appropriate adjuvant,22 or 60 spike receptor-binding domains can self-assemble into a nan-oparticle .23 Different formulations of liposomes appear to increase antigen retention at the injection site, recruitment of immune cells, antigen uptake by phagocytosis, and trafficking to the draining lymph nodes .24 A major advan-tage of protein-based vaccines includes no risk of aberrant infection in immunocompromised individuals .
DNA‑based vaccines can be delivered by intradermal injection followed by electropora-tion and have been shown to induce humoral and cellular immune responses in animal mod-els . Advantages of DNA-based vaccines include that the manufacturing is scalable and the development of the candidate vaccine uses the DNA sequence of the target immunogen, such as the spike protein .25
RNA‑based vaccines contain modified mRNA encoding an immunogenic protein such as the S or spike protein of SARS-CoV-2 .26 They are often formulated in lipid nanoparticles .26 Advantages include that their engineering can begin as soon as the viral genome is known and manufacturing is easily scalable. Two mRNA vaccines targeting the S protein of SARS-CoV-2 were >94% efficacious in clinical trials10, 27 and were granted FDA approval on an emergency basis in 2020 .
Within 7 months, 338 million doses of a COVID-19 vaccine (eg, two RNA-based vaccines, one Ad26-based vaccine) have been adminis-tered in the US28 under emergency approval by the FDA . In Europe, 436 .5 million doses28 of four COVID-19 vaccines (two RNA-based vac-cines and two Ad26-based vaccines) have been administered under emergency approval by

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 9
the European Medicines Agency .29 More than 3 .6 billion doses of one of the COVID-19 vac-cines (eg, inactivated vaccine,30 Ad26-based vaccines, two RNA-based vaccines) have been administered globally,28 under the emergency use listing by the World Health Organization or the local national health authorities . Higher vaccine coverage appeared associated with a reduction in incidence of COVID-19 cases between January and July 2021.28
COVID‑19 Vaccines that Received COVID‑19 Vaccines that Received Emergency Approval in 11 Months Emergency Approval in 11 Months Within 11 months of the sequence of SARS-CoV-2 being posted online, 2 pharma-ceutical teams had developed and tested an RNA vaccine against SARS-CoV-2 infection and progression to COVID-19 disease .26, 27 Two vac-cines that use a replication-incompetent Ad26 vector were developed within 14 months.18,
20, 21 The general steps to develop these vac-cines included:
• Target discovery (identified viral proteins and cellular receptors involved in entry in fine detail)
• Engineered constructs using vaccine platform • Generated formulation • Characterized in vitro and in vivo mRNA and protein products
• Established assays for measuring and char-acterizing neutralizing antibodies and T cell responses (from convalescent humans, in-vitro cell culture and animal models, humans in clinical trials)
• Determined immune responses from conva-lescent individuals and set as goals for vac-cine-induced immune responses
• Tested in rodent models and nonhuman pri-mate models for efficacy and safety
• Initiated & completed Phase 1/2 trials • Completed multicenter, randomized, strati-fied, placebo-controlled, observer-blinded Phase 3 trials in adults
To streamline the timeline, the execution of initial 6 steps in characterization of target and
immune responses of convalescent patients were overlapped as much as possible. Because of their high efficacy in preventing symptomatic COVID-19 illness, both RNA vaccines received FDA approval for emergency use in Dec, 2020 .
The mRNA‑1273 vaccine (Moderna) showed 94.1% efficacy in participants who received two 100 mcg dose of mRNA-1273 at a 28 day interval in a multicenter, randomized, strati-fied, placebo-controlled, observer-blinded Phase 3 trial . The phase 3 trial enrolled 30,420 adults with ages ranging from 18 to 95 years.27 Symptomatic COVID-19 infection occurred in 185 placebo-treated participants (56 .5 per 1000 participants) and 11 participants in the mRNA-1273 group (3 .3 per 1000 participants) .The safety profile included transient local and systemic reactions, mostly mild-to-moderate severity; frequency and severity increased after second dose .27
BNT162b2 mRNA vaccine (Pfizer/BioNTech) showed 95.0% efficacy for two 30 mcg doses of BNF162b2 administered 4 week apart in a ran-domized, observer-blind placebo-controlled trial of 45,548 adults older than 16 year .10 In subgroup analysis, efficacy in participants with hypertension was similar (94.6%). Symp-tomatic COVID-19 infection was significantly more frequent in placebo group (n=162) than in the BNF162b2 group who received 2 doses and had at least 7 days to mount an immune response after the second dose (n=8 partici-pants) .10 The BNT162b2 vaccinated participants mounted T cell responses and neutralizing antibody responses .10 Safety profile included transient local mild-to-moderate pain at injec-tion site, fatigue and headache, and BNT162b and placebo groups experienced similar rates of incidence of serious adverse events (0.6% and 0.5%, respectively).10
These results illustrate that vaccine develop-ment from viral discovery (Jan 10, 2020) to emergency approval of RNA-based vaccine can be accomplished in less than 1 year with suffi-cient resources . This report describes many of the processes that helped streamline the vac-cine development timeline by years .

10 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
Target DiscoveryTarget Discovery
Comparison with similar viruses Comparison with similar viruses With COVID-19 disease, SARS-CoV-2 is closely related to the SARS virus discovered in 2003 . Thus, the discovery process for revealing its replication cycle, the viral and host proteins involved in infection, and its proteins that trig-ger a protective immune response, required less time than a totally novel pathogen . Many aspects of the replication cycle of the SARS-CoV-2 are similar to the 2003 SARS virus, which is now called SARS-CoV or SARS-CoV-1. The 2003 SARS-CoV virus uses its spike protein to bind to the ACE2 on host cells and begin its infection .
Viral protein(s) and cell receptor(s) Viral protein(s) and cell receptor(s) essential for viral entry and infectionessential for viral entry and infectionTo test whether SARS-CoV-2 viruses also use the Spike protein to bind ACE2 receptors to enter cells, Alexandra Walls, a postdoctoral fel-low and colleagues in David Veesler’s labora-tory first engineered expression of the Spike protein from each SARS-CoV-1 and SARS-CoV-2 into a murine leukemia virus pseudotyping sys-
tem .31 Thus, they could investigate the proper-ties of the S protein without the risk of infection with the actual SARS-CoV-2.
The S protein bound to the ACE2 receptor on target cells . SARS-CoV-2 and SARS-CoV-1 showed comparable affinity for ACE2 with simi-lar kinetic rate constants and a slightly higher off-rate for the SARS-CoV-1 by Bio-Layer Inter-ferometry measured with an Octet®.31
Sequence comparison between the S genes from SARS-CoV-1 and 144 SARS-CoV-2 isolates indicated 50% identity of amino acids in the receptor-binding motif .31 Using cryo-electron microscopy, they provided detailed structural images of the SARS-CoV-2 S trimers in two con-figurations: about half had all three S proteins in the closed position and about half of the trimers had one S protein in the open position and the other two S proteins in the closed posi-tion .31 These fine structural details can help scientists design inhibitors of the S protein binding to the ACE2 receptor on cells .
Multiple laboratories have demonstrated that the SARS-CoV-2 Spike protein binds to ACE2 on susceptible cells and is essential for viral entry, the first step in viral infection. Elucidation of the
Figure 5. Bio-layer interferometry of S protein binding to ACE2 receptor. This figure was reprinted with permission from Cell, Vol . 183, Walls AC, Park YJ, Tortorici MA, et al . Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein, p . 283 . Copyright Elsevier (2020) .31

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 11
immunogenic surfaces on the SARS-CoV-2 spike protein and its receptor-binding domain (RBD) provides fundamental knowledge essential for identifying the immunogen for development of potent vaccines and antibody therapeutics .
As a tool, Hattori and colleagues develop a Spike receptor-binding domain (RBD) mutant that does not bind ACE2 (RBD-T) for character-izing anti-RBD antibodies from convalescent sera of COVID-19 patients .32 To maximize effi-ciency, they perform cell- and bead-based bind-ing assays with the iQue® system for advanced high-throughput flow cytometry. Most clones that bound the spike protein from a human antibody phage display library (89%) did not bind to the receptor-binding domain (eg, clone A4-2, Figure 6 left panel) . The biased screen for RBD binders identi-fied mostly antibodies that targeted the RBD and still bound the RBD-T mutant (eg, clone B3-4). Only 12% of antibodies that bound to spike protein and RBD recognize the ACE2 interacting site (eg, clone C3-3, Figure 6 right panel). The data showed that the surface of the RBD which inter-acts with the ACE2 is less immunogenic than other regions of the S protein and the RBD .32 Taken together, these data sug-gest that the immunogen for a component vaccine should not be the RBD fragment but instead the intact S protein trimer . Fur-
ther testing of the immunogenicity of open or closed positions of the S protein trimer may be beneficial.
Antibody responses from Antibody responses from convalescent COVID‑19 patientsconvalescent COVID‑19 patientsUnderstanding the quantity, quality, and dura-tion of antibody and cellular immune responses to SARS-CoV-2 in convalescent individuals set the goalposts for the development of vaccines . Multiple approaches revealed the antigenic-ity of the Spike glycoprotein of SARS-CoV-2 in humans .
Figure 6. Bead-based assay for neutralizing activity of spike specific antibodies shows three distinct results: A4-2 showed no binding of RBD, B3-4 bound RBD and mutant that does not bind ACE2 (RBD-T), and C3-3 bound region of RBD that binds ACE2 (but not mutant RBD-T), This figure was reprinted with permission from J Mol Biol, Vol. 433, 166748, Hattori T, Koide A, Noval MG, et al. The ACE2-binding Interface of SARS-CoV-2 Spike Inherently Deflects Immune Recognition, p.5. Copyright Elsevier (2021).32
Figure 7. Bead-based assay on iQue® correlate with results of ELISA, is more automated and has broader dynamic range . This figure was reprinted with permission from Commun Biol, Vol. 4, 129. Dogan M, Kozhaya L, Placek L, et al. SARS-CoV-2 specific antibody and neutralization assays reveal the wide range of the humoral immune response to virus, p . 3 . Copyright Springer Nature (2021) .33

12 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
Dogan and colleagues developed highly sensi-tive and specific assays for detecting SARS-CoV-2 antibodies to the spike and the nucleocapsid protein, their components, and their neutraliza-tion capabilities .33 Screening for antibody bind-ing to SARS-CoV-2 proteins was performed on an iQue® system for advanced high-through-put flow cytometry (Figure 7 left panel). Dogan et al. showed that bead-based assay correlated well with results from an ELISA-based antibody assay (r=0.86, Figure 7 left panel). Importantly, their bead-binding assay on the iQue® has a greater dynamic range than the ELISA-based assay (Figure 7 right panel) .33
The authors used the assays to probe the humoral immune responses across COVID-19 patients with different severity: outpatient, hos-pitalized, and hospitalized ICU. They detected IgG antibodies to different regions of S pro-tein or nucleocapsid (N) protein at dilutions up to 100,000-fold of plasma samples from SARS-CoV-2 patients (see Figure 8) . A robust neutralization assay using SARS-CoV-2 spike protein pseudovirus detected neutralization in both convalescent and COVID-19 plasma at up to 10,000-fold dilution . These assays also revealed that patients with severe disease had up to 3,000-fold higher antibody and neutrali-
zation titers compared to outpatient or conva-lescent plasma donors .33
Two common strategies help characterize the antibodies from COVID-19 patients and conva-lescent patients .
First, the rapid establishment of viral entry and neutralization assays help assess the protective antibody immune responses of SARS-CoV-2, preferably in a less restrictive containment laboratory . Many groups have engineered pseudotyped viruses expressing S protein for establishing neutralization assays: several viral backbones have been used in pseudotyped viruses .14, 31, 34, 35 The development of the pseu-dotyped virus based on VSV that expresses the VSV-dG –SARS-CoV-2 Spike protein in 1 week is described in great detail34 and is discussed in a subsequent section.
The second common strategy is to develop human monoclonal antibodies from B cells iso-lated from convalescent COVID-19 patients .35, 36 Isolation of the monoclonal antibodies allows production of sufficient pure stocks for charac-terization of the specificity, affinity, and avidity of multiple clones, with the advantage of iden-tifying clones with superior binding character-istics that may be useful as a therapeutic .35
Mor et al . analyzes the cellular, molecular, and functional characteristics of the antibody response in convalescent blood from severe versus mild SARS-CoV-2 donors .35 They used the Incucyte® S3 Live-Cell Analysis System to moni-tor cell death and cell-to-cell fusion induced by SARS-CoV-2 infection and the effect of isolated human anti-SARS-CoV-2 antibodies .35 The data show that severe COVID-19 donors exhibit high titers of anti-SARS-CoV-2 IgG against spike receptor-binding domain (RBD), and higher B-cell expansion than convalescent donors . Six out of 22 isolated monoclonal antibodies potently neutralize the live virus and prevent syncytia formation in live-cell assays . Figure 9 shows the effects of monoclonal antibody TAU-2212 and TAU-2230 in comparison to the vis-ible red fluorescent virally infected cells with the antibody isotype control, G053) .35
Figure 8. Titers of antibodies specific for S protein and nucleocapsid protein in SARS-CoV-2 patients. This figure was reprinted with permission from Commun Biol, Vol . 4, 129 . Dogan M, Kozhaya L, Placek L, et al. SARS-CoV-2 specific antibody and neutralization assays reveal the wide range of the humoral immune response to virus, p . 3 . Copyright Springer Nature (2021) .33

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 13
The virally induced cytotoxicity increased with length of incubation and several antibodies (eg, TAU-2212 and TAU-2230 and combination) reduced cell death, showing their neutralizing ability . Peptide libraries, computational meth-ods and mutagenesis analysis revealed the mechanism of antibody binding .
Antigen/Immunogen discovery Antigen/Immunogen discovery Multiple groups have used Bio-Layer Interfer-ometry by the Octet® to further elucidate how neutralizing antibodies can block the interac-tion between the S protein and the ACE2 cel-lular receptor .22, 36, 37 For example, Zost and colleagues analyzed 389 SARS-CoV-2 S pro-tein-specific human monoclonal antibodies obtained using B cells from 2 convalescent patients .36 Forty monoclonal antibodies bound to the RBD fragment and neutralized wildtype SARS-CoV-2 in a screening assay . The major antigenic sites in the RBD were determined by competitive binding analysis with Bio-Layer Interferometry using the Octet® HTX. Admin-istration of the two most potent neutralizing antibodies reduced infection in preclinical pro-phylactic and therapeutic mouse models .36
Library screening can help identify potential antibody or single-domain antibodies for their
potential feasibility as therapeutics .38 To probe the antibody epitopes in the receptor-binding domain, Wu et al .38 developed a phage display library of single-domain antibodies and tested their binding specificity and affinity to the spike protein and their ability to block its binding to ACE2 . Binding competition assays using the Octet® revealed that the single-domain anti-bodies with neutralizing activity could recog-nize 5 distinct epitopes on the spike protein, near the receptor-binding domain (RBD) . Bind-ing competition assays with Bio-Layer Inter-ferometry indicated that 2 epitopes located outside the RBD were required for binding to ACE2 but were not neutralized with the single-domain antibodies .38 One relatively immuno-genic epitope within the RBD (Group A) was not neutralizing .38
Taken together, these studies illustrate the versatility of Bio-Layer Interferometry and the Octet® in identifying the fine details of S protein – antibody and ACE2 protein interac-tions .36, 38 These interactions were visualized with cryo-electron microscopy.23 Additional examples include elucidation of a specific type of glycosylation critical for entry of a distinct coronavirus, OC43 that causes mild respiratory symptoms .39
Figure 9. Cocktails of isolated neutralizing antibodies prevented viral replication in cell culture. This figure was reprinted with permission from PLoS Pathology, Vol . 17, e1009165 . Mor M, Werbner M, Alter J, et al . Multi-clonal SARS-CoV-2 neutralization by antibodies isolated from severe COVID-19 convalescent donors, p . 3 . Copyright Creative Commons Attrition License (2021) .35

14 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
Discovery Discovery Applications Applications The continual possibility of an emerging infectious disease-causing severe dis-ease and mortality region-ally and globally creates an urgent need for strategies that can accelerate discov-ery of vaccines and thera-peutics. Here we discuss overlapping strategies to streamline the time line for vaccine development and identification of therapeutic candidates in culture .
Biology Discovery Biology Discovery Processes Processes The 3 main biology discov-ery processes are charac-terization of EID and its disease, sequencing and genomic analysis with com-parison to related patho-gens, and characterization of the immune responses in convalescent and severely diseased patients . Biotech firms often can employ platforms for develop-ing pseudotyped viruses for establishment of neu-tralization assays and their vaccine platforms. Use of high-throughput assays at the indicated steps can rapidly accumulate suffi-cient data to inform deci-sions on target proteins for in vivo testing of vaccine candidates and selection of neutralizing antibodies and cocktails of therapeu-tic monoclonal antibodies . The high-throughput assays are also used to character-ize the responses in the in vivo models .
Emerging infectious disease
Biology Discovery Processes
EID characterization Sequencing/genome analysis Convalescent adults
EID surface protein
Pseudovirusplatform
Vaccine platform
Human monoclonalantibodies selection*Neutralizing activity*
Convalescent seraNeutralizing titers*
T cellresponses*
Cell viability*Cytopathic effect*Target cells* Infectivity*
Neutralization assays*
EID target protein(s)* &Receptor binding domain*
EID target protein(s)*
Affinity* avidity* epitope mapping* Binning*
mAb discovery* fortherapeutics
2+ Vaccine constructs expressing 1 or more EID proteinsTest in rodent models, and both or best construct in NHP model
Therapeutic modelsin rodents & NHP
Prophylacticvaccine models
Neutralization assays*
Pseudoviruswith EIDsurface proteins
Figure 10. Pathways in the Biology Discovery Processes for Emerging Infectious Diseases. *Amenable to high-throughput assays. EID, emerging infectious disease; mAb, monoclonal antibody; NHP, nonhuman primate.

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 15
Cell Viability /ProliferationCell Viability /ProliferationReal-time monitoring of cell growth and marker expression by the Incucyte® can inform on optimal conditions for set up of a cell viability assay: optimal seeding densities coupled with incubation times, morphology changes during activation or stem cell differentiation, and mon-oclonality after seeding with limiting dilution. In Figure 11, the three time points (0 hrs, 24 hrs, 48 hrs) illustrate the growth of the human umbili-cal vein endothelial cells (HUVECs) by using phase contrast microscopy of the Incucyte®.
Furthermore, subsequent assays for trans-fection, transduction, infection, or sensitiv-ity to potential therapeutic compounds are dependent on a consistent percentage of cell
confluency or cell growth in 3D cultures (eg, spheroids) for reproducibility . The use of cell viability assays for screening and characteriza-tion of therapeutic compounds is discussed in that respective section .
The Incucyte® system provides multiple analy-sis programs for different types of cells. Since mycelial growth and branching of fungi can resemble neuronal growth patterns, Wurster and colleagues repurposed the NeuroTrack processing module of the Incucyte®. They used the time-lapse feature to measure myce-lial growth branching of six different patho-genic fungi including Candida albicans which is shown in Figure 12.40
Figure 11. Incucyte® monitoring of growth. This figure was reprinted with permission from Sartorius.
Figure 12. Growth of the pathogenic fungi, Candida albicans.40 This figure was reprinted with permission from mBio, Vol. 10, e00673-19. Wurster S, Kumaresan PR, Albert ND, et al. Live Monitoring and Analysis of Fungal Growth, Viability, and Mycelial Morphology Using the Incucyte NeuroTrack Processing Module, p. 5. Copyright American Society for Microbiology and American Academy of Microbiology (2019) .40

16 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
Infectivity assaysInfectivity assaysMonitoring the progressing cytopathic effects on infected cells can inform on the type of EID . Cytopathic effects can include changes in size, shape, and attachment status of the infected cells . They are performed in the appropriate Biosafety Laboratory (BSL) level, often BSL3 or BSL4, and traditionally are monitored daily . As a real-time, live-cell monitoring equipment, the Incucyte® can be set to automatically image the 96- or 384 well tissue culture plates throughout the course of an experiment, allowing more frequent data collection, maintain culture con-ditions which supports reproducibility, and use no manpower for capturing the images.
To maximize safety, efficiency, and data acqui-sition (number of monitored time points), groups are using the Incucyte® (Figure 13.) to acquire data on 3 or 4 hr intervals and ana-lyze the data off-site due to the pandemic. As Touscane Fouriè and Dr . Franck Touret from Unite des Virus Emergents (Emerging Virus Unit), Marseille explained, after an assay is set up in the BSL3 laboratory in the Incucyte®, the acquisition of data at specified time intervals over a five day period is automated. Because they do not have to spend time to visit the BSL3 lab to image the plates, they are able to set up additional plates of positive samples, process-ing approx . 30,000 to 36,000 positive samples
per week. Data analysis is downloaded and analyzed at a convenient location outside of the BSL3 laboratory .
Viral titers Viral titers This high-throughput assay can measure viral titers per mL to reduce the labor-intensive manual counting of viral foci .41 The Incucyte® generated titer provided a reproducible bench-mark for screening antiviral therapeutics .41 This method may be used to screen EID variants for production of higher numbers of infectious viruses per infected cell .
As an extension, the expression of 3 infectious disease immunogens from live turkey herpes-virus vectored vaccine from various passages were compared at multiple time points in the Incucyte® and shown to be stable.15
Cellular-viral interactions Cellular-viral interactions A viral infection assay using the Incucyte® live-cell analysis system helps characterize cellular responses to Maraba oncolytic virus infection, a recent candidate for oncolytic treatment of cancer .42 Secondly, infectivity assays of Human Papilloma virus infection revealed that the neu-tralizing antibody-targeted L1 viral capsid pro-tein trafficks with the genome to the nucleus by monitoring at multiple time points with the Incucyte® system.43
To identify how viral proteins of SARS-CoV-2 infection affected lung cells, Stukalov et al. used state-of-the-art proteomics and detected interactions between 1146 host proteins and 24 SRS-CoV-2 proteins .44 Viral infection had altered multiple mediators involved in gene expression (transcriptome), protein abundance (proteome), protein ubiquitination, intracellu-lar trafficking, and degradation (ubiquitinome), and signaling pathways (phosphoproteome).
Similar techniques that elucidate host-viral interactions are applicable to other viruses including influenza A virus,45 Bursal Disease virus,46 and herpes simplex virus .47
Figure 13. Incucyte® provides programmable real-time, live-cell monitoring. This figure was reprinted with permission from Sartorius .

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 17
Pseudotyped virusesPseudotyped virusesInfectious EIDs surrogates allow studies to be performed in more convenient laboratory containment settings . Pseudotyped viruses express one or more EID surface proteins and can help probe viral entry, characterize effective immune responses of convalescent humans, and screen for inhibitors of the EID infection . For their binding studies, Walls et al . engineered a pseudotyped murine leukemia virus to express S protein from SARS-CoV-2 .48
Many pseudotyped viruses also express the green fluorescent protein for ease of live-cell monitoring, and cytotoxicity monitor-ing. Pseudotyped fluorescent VSV expressing the S protein of SARS-CoV-2 can be gener-ated and infection assays developed with the use of the Incucyte® in one week with the recently published protocol by Shehadeh’s group .34 If appropriate cell lines that express the viral cell receptor are not available, Con-dor Capcha et al . (2020)34 also describes how to generate susceptible cell lines in basically 2 steps: (1) construct lentiviruses to express essential proteins for the infectivity of the EID (SARS-CoV-2), and (2) transduce and isolate cell lines that express those essential proteins (eg, ACE2 receptor, Furin or TMPSS2 enzyme) for the infection assays . Pseudotyped viral assays performed with the susceptible cells are quan-titative and readily scalable for measurement of neutralizing antibodies, and monitoring of immune responses of vaccine candidates . Cou-pled with real-time, live-cell analysis platforms (Incucyte®), pseudotyped viral systems with a fluorescent reporter gene can rapidly advance screening for promising drugs against the EID disease .34
For example, a single cycle pseudotyped influ-enza virus with the enhanced green fluorescent protein was coated with Ebola glycoprotein in a stable producer line .49 Its advantages include no genetic material from Ebola virus, similar viral entry process to wildtype Ebola, infection restricted to single cycle in cells, handling in BSL1/2, and sufficient production yields. Sci-entists used analogous pseudotyped systems coupled with iQue® advanced flow cytometry or the Incucyte®49, 50 live-cell analysis platform
to screen more than 1200 pharmacologically active compounds against viral entry of Ebola .49 Pseudotyped viruses expressing the S protein are used for many coronaviruses, including a coronavirus that causes the mild respiratory infection, OC4339 and 17 variants of SARS-CoV2 detected globally .14
Viral variantsViral variantsEIDs can evolve as they spread globally . Vari-ants of SARS-CoV-2 are emerging in the United Kingdom, Brazil, South Africa, and elsewhere.51 The SARS-CoV-2 variants have one or more sub-stitutions in the Spike protein . High-through-put neutralization assays using pseudotyped viruses with the S gene from each of the vari-ants can be rapidly developed .34 Neutralizing ability of some convalescent sera were about 2 to 9 fold lower against variants.51 However, the sera from 5 BNT162b1-vaccinated people neutralized each of 17 variants expressed in pseudoviruses .14
Assessment of immune responsesAssessment of immune responsesDevelopment of a vaccine strategy against a new EID begins with identifying the target cells (which cells become infected with the patho-gen of EID), the essential components of the pathogen EID that attach to the target cell and promote entry, the cellular receptor (how the pathogen binds and enters the target cell), and the immune response of convalescent adults .
Antibody responses and characterizationAntibody responses and characterizationMeasuring the neutralization titers of poly-clonal sera from the convalescent adults pro-vides a target for neutralizing titers induced by vaccine candidates .33 Piccoli et al . charac-terized the neutralizing antibodies from more than 600 convalescent patients by multiple assays, including ELISAs, pseudovirus neutrali-zation assays, competition binding assays with Bio-Layer Interferometry with the Octet®, and cryo-electron microscopy .52 About 90% of the neutralizing activity in these sera recognized the RBD of Spike protein, and hospitalized patients developed higher neutralizing antibody titers . They identified 2 distinct antigenic sites in the RBD of the spike protein with cryo-electron

18 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
microscopy and confirmed with Bio-Layer Inter-ferometry. Several antibodies with neutralizing activity bind sites outside the RBD .52
To characterize individual human antibodies, monoclonal antibodies are generated from B cells isolated from convalescent adults . The monoclonal antibodies with neutralizing activ-ity are characterized for binding specificity, affinity, avidity, and competitive binding.35, 52 The binding specificity of the monoclonal anti-bodies is mapped to specific amino acids in the target protein which comprise the epitope. Most neutralizing antibodies will block the interaction between the receptor-binding domain of the viral S protein and its cellular receptor (ACE2 for SARS-CoV-2) . Many groups use a Bio-Layer Interferometry assay on the Octet® to determine the affinity and avidity of the antibodies and perform competitive bind-ing assays .22, 36‑38, 52, 53 The binding locations of single monoclonal antibodies and promising cocktails are often visualized with cryo-electron microscopy .37, 52
Surprisingly, two neutralizing human mAb from natural infection bound to the N-terminal domain of the S protein as determined with a library of S protein mutants in which each amino acid was individually substituted with alanine .54 The binding assay with the iQue® high-throughput flow cytometry mapped the epitope to 5 specific residues for COV2-2489 and a single residue for COV2-2676 . These mono-clonal antibodies required Fc- effector function for neutralizing activity, and showed protection in a hACE2 transgenic mouse model .54
Epitope binning Epitope binning Epitope binning groups monoclonal antibodies with overlapping epitopes by competitive bind-ing assays and is often performed using Bio-Layer Interferometry on an Octet®. Binding competition assays using the Octet® revealed that the single-domain antibodies with neutral-izing activity bound to one of 5 epitopes near the receptor-binding domain . The therapeutic antibodies in a mixture usually bind to distinct epitopes for a specific target in a noncom-petitive manner . Epitope binning also can be
applied to characterization of antibodies tar-geting a protein important in other therapeutic areas, such as anti-cancer activity .55
Macrophages Macrophages Macrophage phagocytosis can be readily quantified by using a real-time imaging plat-form set for 10 minute intervals (Incucyte®).56 Phagocytosis helps induce and amplify T cell and antibody immune responses . As a brief review, native antibodies contain both the anti-gen-binding region (Variable region) and the Fc region, which determines the isotype and innate functions including phagocytosis .57
T cell responses T cell responses Adaptive antiviral immune responses usu-ally involve both antibody responses and T cell responses . Yu, Seshadri and colleagues thoroughly profiled the functions of T cells and antibodies of convalescent, hospitalized, or not hospitalized patients with multiparam-eter flow cytometry and systems serology.58 They investigated the responses to the Spike, envelope, and the nucleocapsid proteins of SARS-CoV-2 by using the iQue® advanced flow cytometry platform to characterize the antibody-dependent cellular phagocytosis, antibody-dependent neutrophil phagocytosis, and antibody-dependent complement fixa-tion. Key findings showed that hospitalized individuals with comorbid illnesses displayed consistently higher magnitude and breadth of CD4 T cell responses and antibody responses to SARS-CoV-2 .58 After accounting for potential confounding factors, non-hospitalized patients exhibited a more coordinated and function-ally diverse response between antibodies and T cells targeting SARS-CoV-2 .
The protective role of T cells against SARS-CoV-2 was confirmed in macaques by McMahan, Baarouch and colleagues at Harvard Medi-cal School .59 They observed that CD8+ T cells augmented the protective effect of subopti-mal neutralizing antibody levels against SARS-CoV2, supporting the role of cellular immunity in abrogating the progression of SARS-CoV-2 infection .59

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 19
Sample quantity often limits the number of variables for testing from patient samples or samples from animal studies. Using kits and the iQue® high-throughput flow cytom-eter, scientists can efficiently characterize the stages of T cell development (activation, kill-ing activity, and exhaustion) and the memory compartment (naïve, stem cell memory, cen-tral memory, effector memory, and terminal memory T cells) in proportion of T cells spe-cific to S protein and RBD region. Comparison of unstimulated and stimulated T cells moni-tored at multiple time points by the iQue® are shown in Figure 14 above:
Antiviral TherapeuticsAntiviral TherapeuticsThe possible emergence of novel viral patho-gens causing severe disease creates an urgent need for strategies to accelerate discovery antiviral therapeutics .60 Gilchuk et al . pre-sented a workflow with integrated technology to obtain ZIKA specific monoclonal antibody therapeutics rapidly . The steps include gen-erate human B-cell derived monoclonal anti-bodies from convalescent ZIKA virus patients and select neutralizing clones, characterize the antibodies, epitope map, and assess in animal models in 90 days . First, they screened con-
valescent ZIKA virus patients for the highest responders of neutralizing Ab using an iQue® flow cytometer. After rapid generation of mon-oclonal antibodies, microscale production and purification of Mab, the lead candidates were screened for neutralizing activity and binding specificity with use of the iQue® flow cytom-eter, antibody-mediated phagocytosis by neutrophil and monocytes, complement dep-osition, and competitive binding analysis and epitope mapping . Preclinical studies involved challenge studies in mice and nonhuman pri-mates and assessment of the responses . The workflow was accomplished in 78 days.60
Dr . Pavlo Gilchuk probes the mechanisms of antibodies and the synergy of antibody cock-tails for blocking viral infections and spread . Gilchuk and colleagues at Vanderbilt use high-throughput data collection, analysis, and visu-alization of trends to elucidate the additive and synergistic interactions for characterizations of potential therapeutic mixtures . Adapting the binding assays for the iQue®, Dr. Gilchuk, senior staff scientist at Vanderbilt explains that it “definitely provides a technical benefit for antibody analysis . …We can consider it high-throughput two-dimensional analysis…and it was essential for our work.”
Figure 14. Monitoring of PBMC activation using phase contrast microscopy and live-cell immunocytochemistry with anti-CD71 FabFluor-488. PBMCs (30,000/well) were treated with vehicle control (nonactivated) or CD3 and IL-2. A) Phase contrast imaging captures morphological differences and increased expression of CD71 in activated cells. Quantitative analysis shows time-dependent increase in cell size (B), cell eccentricity (C), and expression of CD71 (D). This figure was reprinted with permission from Sartorius .

20 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
This methodology is used to analyze antibod-ies against SARS-CoV-2,36, 61 Ebolavirus,62 and ZIKA.60 When human neutralizing monoclonal antibodies are used as therapeutic agents, the Fc effector functions of neutralizing antibodies to SARS-CoV-2 are essential for optimal protec-tion .57 The identification of therapeutic anti-body cocktails may appear to be more complex in some diseases because of antigenic drift in HIV, filoviruses, and influenza.
Stukalov et al. showed that SARS-CoV-2 infec-tion had altered expression of numerous medi-ators involved in regulation of transcription, DNA damage response, and stress pathways.44 They used the Incucyte® Live-Cell Analysis system to monitor the effects of 48 currently marketed medications every 6 hrs on viral rep-lication and cell fusion . While several medica-tions increased viral replication, several other medications suppressed viral replication .44

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 21
SummarySummaryThe rapid development of vaccines for COVID-19 demonstrated that various steps could be overlapped to shorten the overall timeline in the case of a pandemic . Furthermore, assays performed in high-throughput equipment can efficiently monitor numerous variables in small sample volumes at multiple time points which increases reproducibility, provides additional insights, and free-ups the valuable time of sci-entists. In addition, the downloaded analysis can be performed in a less restrictive biosafety setting, thereby providing safety for colleagues during a pandemic . Scientists often use high-
throughput assays based on advanced flow cytometry analysis, real-time, live-cell analy-sis, and/or label-free binding analysis to effi-ciently determine host-pathogen interactions and identify target antigens for vaccine devel-opment . After constructing vaccine candidate, they use these high-throughput assays to measure immune responses in animal models and clinical trials. Using these innovative high-throughput and automated Incucyte®, iQue®, and Octet® cell & protein analysis platforms can accelerate your research and the develop-ment of your novel, efficacious therapeutics and vaccines (Table 1) .
Incucyte® iQue® Octet®Determining Pathogen-Host Interactions
Live‑Cell Analysis Assays Advanced Flow Cytometry Assay
Label‑Free Binding Analysis
Host cells Cell Health & Viability Apoptosis, Cell cycle, Morphology, Proliferation, Cell monitoring & workflows: Cell culture QC, Cell-by-cell analysis, Immunocytochemistry, Reporter genes, Organoid culture QCCytotoxicity, Phagocytosis
Cell-based kits Cell Proliferation, Cell Encoding, Cell Viability, Cell Membrane Integrity, Cell Cycle
Receptor-ligand interactionsBiomolecular binding kinetics assays Biomolecular interactions
Infection, replica‑tion, and spread
Live –cell Assays Kinetics of Cytopathic effects, Necrosis, apoptosis, antibody internalization, Immune cell clustering & proliferation, Immune cell killing, NETosis
Antibody Titer, iso-type, kinetics Cytokine profilingNK KillingT cell activationT cell exhaustion
Host-Pathogen interactionsViral-Host receptor recognition analysisInhibitor binding analysisAntibody titer
Innate immunity Chemotaxis, cell migrationCell invasionEfferocytosisImmune cell killing NETosisPhagocytosis
Immunophenotyping and Function Antibody-dependent cellular phagocytosisAntibody-dependent complement fixationCytokine profilingNK killing
Binding of Fc region of therapeutic antibodies to complement
Antibody & T cell immunity
Live-cell immunochemistry IgG titer, isotype, and protein specificity T cell activation analysis
Antibody titerAntibody screeningCompetitive bindingEpitope binning
Post‑Infection Immune cell proliferation and killing
Antibody titerT cell memoryT cell exhaustion
Antibody titer kinetics
Protective immunity
ProliferationViability
Neutralizing antibodies titerAntibodies titerT cell memory
Neutralizing antibody analysis and titersEpitope binning
NK, Natural Killer cells; QC, quality control;
Table 1. Incucyte®, iQue® and Octet® advanced cell & protein analysis platforms provide insights at unprecedented speed, depth & scale, streamlining workflows to accelerate results

22 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
For more information:For more information:Incucyte®, iQue®, Octet® Evaluating Viral Infections Across the Entire Host-Pathogen Life Cycle: Find out how you can maximize your in vitro virology studies in minimal time . www.sartorius.com/en/pr/COVID-19-solutions/COVID-19-research-products/virology
Find out how you can enhance your antibody interaction analyses with advanced flow cytom-etry and live-cell analysis . Antibody Internaliza-tion: Advanced Flow Cytometry and Live-Cell Analysis Give Rich Insights During Antibody Pro-filing Lab Supplies for Coronavirus Testing, Air Monitoring, and Vaccine Research | Sartorius
Learn more about how Octet® systems are used in COVID-19 research at Octet COVID-19 Research (sartorius .com)
Explore our solutions for your downstream vaccine research, including harvesting, concen-tration and purification of viruses and proteins by using ultrafiltration.
Viral Vaccines & Vectors (sartorius .com)

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 23
Selected ReferencesSelected References1 . Heaton PM . The COVID-19 vaccine-development
multiverse . N Engl J Med. 2020;383(20):1986-1988.
2 . Mullard A . COVID-19 vaccine development pipe-line gears up . Lancet. 2020;395(10239):1751-1752.
3 . Ho RJY . Warp-speed COVID-19 vaccine devel-opment: beneficiaries of maturation in biop-harmaceutical technologies and public-private partnerships . J Pharm Sci. 2021;110(2):615-618.
4 . Santoro F, Donato A, Lucchesi S, et al . Human transcriptomic response to the VSV-vectored Ebola vaccine . Vaccines (Basel). 2021;9(2).
5 . Rawat K, Kumari P, Saha L. COVID-19 vaccine: A recent update in pipeline vaccines, their design and development strategies . Eur J Pharmacol. 2021;892:173751.
6 . Chen N, Zhou M, Dong X, et al. Epidemio-logical and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study . Lancet. 2020;395(10223):507-513.
7 . Zhou P, Yang XL, Wang XG, et al. A pneumonia out-break associated with a new coronavirus of prob-able bat origin . Nature. 2020;579(7798):270-273.
8 . Hu B, Guo H, Zhou P, et al . Characteristics of SARS-CoV-2 and COVID-19 . Nat Rev Microbiol. 2021;19(3):141-154.
9 . Lu R, Zhao X, Li J, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding . Lancet. 2020;395(10224):565-574.
10 . Polack FP, Thomas SJ, Kitchin N, et al. safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N Engl J Med. 2020;383(27):2603-2615.
11 . Wu F, Zhao S, Yu B, et al. A new coronavirus asso-ciated with human respiratory disease in China. Nature. 2020;579(7798):265-269.
12 . Wang MY, Zhao R, Gao LJ, et al . SARS-CoV-2: structure, biology, and structure-based thera-peutics development . Front Cell Infect Microbiol. 2020;10:587269.
13 . Landmann H, Menzel K, Green A, et al. Concentra-tion and Purification of Viruses by using Ultrafiltra-tion, Incl. Coronavirus – a Short Review. Goettingen: Sartorius Lab Instruments GmbH & Co;2021.
14 . Sahin U, Muik A, Derhovanessian E, et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses . Nature. 2020;586(7830):594-599.
15 . Tang N, Zhang Y, Sadigh Y, et al . Generation of a triple insert live avian herpesvirus vectored vac-cine using CRISPR/Cas9-based gene editing . Vac-cines (Basel). 2020;8(1):97.
16 . Pollard AJ, Launay O, Lelievre JD, et al . Safety and immunogenicity of a two-dose heterolo-gous Ad26 .ZEBOV and MVA-BN-Filo Ebola vac-cine regimen in adults in Europe (EBOVAC2): a randomised, observer-blind, participant-blind, placebo-controlled, phase 2 trial . Lancet Infect Dis. 2021;21(4):493-506.
17 . Mutua G, Anzala O, Luhn K, et al. Safety and Immunogenicity of a 2-dose heterologous vaccine regimen with Ad26.ZEBOV and MVA-BN-Filo Ebola vaccines: 12-month data from a phase 1 rand-omized clinical trial in Nairobi, Kenya. J Infect Dis. 2019;220(1):57-67.
18 . Bos R, Rutten L, van der Lubbe JEM, et al . Ad26 vector-based COVID-19 vaccine encoding a pre-fusion-stabilized SARS-CoV-2 Spike immunogen induces potent humoral and cellular immune responses . NPJ Vaccines. 2020;5:91.
19 . Sadoff J, Le Gars M, Shukarev G, et al. interim results of a phase 1-2a trial of Ad26 .COV2 .S COVID-19 vaccine . N Engl J Med. 2021:384(19): 1824-1835 .
20 . Ramasamy MN, Minassian AM, Ewer KJ, et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): a single-blind, randomised, controlled, phase 2/3 trial . Lancet. 2021;396(10267):1979-1993.
21 . Sadoff J, Gray G, Vandebosch A, et al. Safety and efficacy of single-dose Ad26.COV2.S vaccine against COVID-19 . N Engl J Med. 2021;384(23):2187-2201.
22 . Tian X, Li C, Huang A, et al. Potent binding of 2019 novel coronavirus spike protein by a SARS corona-virus-specific human monoclonal antibody. Emerg Microbes Infect. 2020;9(1):382-385.
23 . Walls AC, Fiala B, Schafer A, et al . Elicitation of potent neutralizing antibody responses by designed protein nanoparticle vaccines for SARS-CoV-2 . Cell. 2020;183(5):1367-1382 e1317.
24 . Agbayani G, Jia Y, Akache B, et al . Mechanistic insight into the induction of cellular immune responses by encapsulated and admixed archae-osome-based vaccine formulations . Hum Vaccin Immunother. 2020;16(9):2183-2195.

24 Vaccine DiscoVery anD DeVelopment: lessons from coViD-19
25 . Tebas P, Yang S, Boyer JD, et al . Safety and immu-nogenicity of INO-4800 DNA vaccine against SARS-CoV-2: A preliminary report of an open-label, Phase 1 clinical trial . EClinicalMedicine. 2021;31:100689.
26 . Vogel AB, Kanevsky I, Che Y, et al. BNT162b vac-cines protect rhesus macaques from SARS-CoV-2. Nature. 2021:592(7853): 283-289 .
27 . Baden LR, El Sahly HM, Essink B, et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine . N Engl J Med. 2021;384(5):403-416.
28 . Randall T, Sam C, Tartar A, et al . More than 3 .67 billion shots given: COVID-19 tracker . Bloomb-erg . July 20, 2021 . https://www.bloomberg.com/graphics/covid-vaccine-tracker-global-distribution/
29 . European Medicines Agency . COVID-19 vaccines: authorized . European Medicines Agency . https://www.ema.europa.eu/en/human-regulatory/overview/public-health-threats/coronavirus-disease-COVID-19/treatments-vaccines/vac-cines-COVID-19/COVID-19-vaccines-authorised . Published 2021 . Accessed July 21, 2021 .
30 . Jara A, Undurraga EA, Gonzalez C, et al. Effective-ness of an inactivated SARS-CoV-2 vaccine in Chile . N Engl J Med. 2021 .
31 . Walls AC, Park YJ, Tortorici MA, et al . Structure, function, and antigenicity of the SARS-CoV-2 Spike glycoprotein . Cell. 2020;181(2):281-292 e286.
32 . Hattori T, Koide A, Noval MG, et al. The ACE2-binding interface of SARS-CoV-2 Spike inher-ently deflects immune recognition. J Mol Biol. 2021;433(3):166748.
33 . Dogan M, Kozhaya L, Placek L, et al. SARS-CoV-2 specific antibody and neutralization assays reveal the wide range of the humoral immune response to virus . Commun Biol. 2021;4(1):129.
34 . Condor Capcha JM, Lambert G, Dykxhoorn DM, et al . Generation of SARS-CoV-2 Spike pseudo-typed virus for viral entry and neutralization assays: a 1-week protocol. Front Cardiovasc Med. 2020;7:618651.
35 . Mor M, Werbner M, Alter J, et al . Multi-clonal SARS-CoV-2 neutralization by antibodies isolated from severe COVID-19 convalescent donors . PLoS Pathog. 2021;17(2):e1009165.
36 . Zost SJ, Gilchuk P, Case JB, et al . Potently neutral-izing and protective human antibodies against SARS-CoV-2 . Nature. 2020;584(7821):443-449.
37 . Pinto D, Park YJ, Beltramello M, et al . Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody . Nature. 2020;583(7815):290-295.
38 . Wu Y, Li C, Xia S, et al. Identification of human single-domain antibodies against SARS-CoV-2 . Cell Host Microbe. 2020;27(6):891-898 e895.
39 . Tortorici MA, Walls AC, Lang Y, et al . Struc-tural basis for human coronavirus attachment to sialic acid receptors . Nat Struct Mol Biol. 2019;26(6):481-489.
40 . Wurster S, Kumaresan PR, Albert ND, et al. Live monitoring and analysis of fungal growth, viability, and mycelial morphology using the incucyte neu-rotrack processing module . mBio. 2019;10(3).
41 . Stewart H, Bartlett C, Ross-Thriepland D, et al. A novel method for the measurement of hepatitis C virus infectious titres using the Incucyte ZOOM and its application to antiviral screening . J Virol Methods. 2015;218:59-65.
42 . Hassanzadeh G, Naing T, Graber T, et al . Charac-terizing cellular responses during oncolytic Mar-aba virus infection . Int J Mol Sci. 2019;20(3).
43 . DiGiuseppe S, Bienkowska-Haba M, Guion LGM, et al . Human Papillomavirus major capsid pro-tein l1 remains associated with the incoming viral genome throughout the entry process . J Virol. 2017:91(16):e00537-17 .
44 . Stukalov A, Girault V, Grass V, et al . Multilevel pro-teomics reveals host perturbations by SARS-CoV-2 and SARS-CoV . Nature. 2021:594(7892):246-252 .
45 . Lim K, Kim M, Lee MK, et al. Biophysical char-acterization of sites of host adaptive mutation in the influenza A virus RNA polymerase PB2 RNA-binding domain . Int J Biochem Cell Biol. 2014;53:237-245.
46 . Cubas-Gaona LL, Diaz-Beneitez E, Ciscar M, et al. Exacerbated apoptosis of cells infected with Infectious Bursal Disease Virus upon exposure to interferon alpha . J Virol. 2018;92(11).
47 . Guo H, Omoto S, Harris PA, et al . Herpes simplex virus suppresses necroptosis in human cells . Cell Host Microbe. 2015;17(2):243-251.
48 . Walls AC, Park YJ, Tortorici MA, et al . Structure, function, and antigenicity of the SARS-CoV-2 Spike glycoprotein . Cell. 2020;183(6):1735.
49 . Xiao JH, Rijal P, Schimanski L, et al. Characteriza-tion of Influenza virus pseudotyped with Ebolavi-rus glycoprotein . J Virol. 2018;92(4).

Vaccine DiscoVery anD DeVelopment: lessons from coViD-19 25
50 . Wurster S, Sass G, Albert ND, et al . Live imaging and quantitative analysis of Aspergillus fumigatus growth and morphology during inter-microbial interaction with Pseudomonas aeruginosa. Viru-lence. 2020;11(1):1329-1336.
51 . Chen RE, Zhang X, Case JB, et al. Resistance of SARS-CoV-2 variants to neutralization by monoclo-nal and serum-derived polyclonal antibodies . Nat Med. 2021;27(4):717-726.
52 . Piccoli L, Park YJ, Tortorici MA, et al . Mapping neutralizing and immunodominant sites on the SARS-CoV-2 Spike Receptor-Binding Domain by structure-guided high-resolution serology . Cell. 2020;183(4):1024-1042 e1021.
53 . Wang C, Li W, Drabek D, et al . A human monoclo-nal antibody blocking SARS-CoV-2 infection . Nat Commun. 2020;11(1):2251.
54 . Suryadevara N, Shrihari S, Gilchuk P, et al . Neutralizing and protective human mono-clonal antibodies recognizing the N-terminal domain of the SARS-CoV-2 spike protein . Cell. 2021:184(9):2316-2331 .
55 . Koh XY, Koh XH, Hwang LA, et al. Therapeutic anti-cancer activity of antibodies targeting sulf-hydryl bond constrained epitopes on unglyco-sylated RON receptor tyrosine kinase . Oncogene. 2019;38(48):7342-7356.
56 . Kapellos TS, Taylor L, Lee H, et al. A novel real time imaging platform to quantify macrophage phago-cytosis . Biochem Pharmacol. 2016;116:107-119.
57 . Winkler ES, Gilchuk P, Yu J, et al . Human neutral-izing antibodies against SARS-CoV-2 require intact Fc effector functions for optimal therapeutic pro-tection . Cell. 2021 .
58 . Yu KK, Fischinger S, Smith MT, et al. Comorbid illnesses are associated with altered adaptive immune responses to SARS-CoV-2 . JCI Insight. 2021;6(6).
59 . McMahan K, Yu J, Mercado NB, et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature. 2021;590(7847):630-634.
60 . Gilchuk P, Bombardi RG, Erasmus JH, et al . Inte-grated pipeline for the accelerated discovery of antiviral antibody therapeutics . Nat Biomed Eng. 2020;4(11):1030-1043.
61 . Zost SJ, Gilchuk P, Chen RE, et al . Rapid isolation and profiling of a diverse panel of human mono-clonal antibodies targeting the SARS-CoV-2 spike protein . Nat Med. 2020;26(9):1422-1427.
62 . Gilchuk P, Murin CD, Milligan JC, et al . Analysis of a therapeutic antibody cocktail reveals determi-nants for cooperative and broad Ebolavirus neu-tralization . Immunity. 2020;52(2):388-403 e312.

Translate Insights Into Novel Strategies for Vaccine ResearchInnovative Solutions for Extraordinary TimesThe COVID-19 pandemic has scientists developing novel life-saving treatments at breakneck speed. Sartorius provides innovative high-throughput & automated solutions for faster vaccine discovery, characterization & development.
Our groundbreaking Octet®, iQue® and Incucyte® advanced cell & protein analysis platforms provide insights at unprecedented speed, depth & scale, streamlining workflows to accelerate results - when it counts.
www.sartorius.com/vaccine-discovery
Get to Your Destination Faster with the New iQue®3 When time is of the essence, the iQue®3 advanced flow cytometry platform offers a complete platform solution of instrument, software, and reagents. This user friendly walk-away system provides the fastest plate sampling combined with built in data analysis and novel data reduction tools that take you from samples to actionable answers in minutes. www.sartorius.com/covid-19-solutions
© 2021 Essen BioScience. All rights reserved. Intellicyt and iQue are registered trademarks and the property of Essen BioScience. Essen BioScience is a Sartorius Company.

Photo Credit © Sartorius
Sponsored by: