vlvri surwhlq surwhlqlqwhudfwlrqv … · wrsx. uli\wkhuhfrpelqdqw3,1. v...
TRANSCRIPT

Optimisation of expression of recombinant
puroindolines and analysis of
protein-protein interactions in planta
Azadeh Niknejad
This thesis is presented for the degree of
Doctor of Philosophy
June 2014
Department of Chemistry and Biotechnology
Faculty of Science, Engineering and Technology
Swinburne University of Technology
Melbourne, Australia

i
Abstract
The two puroindoline proteins of wheat (Triticum aestivum L.), puroindoline-a (PINA)
and puroindoline-b (PINB) are largely responsible for grain texture, a property
important in food technology and the wheat trade. Furthermore, it has been suggested
that PINs may have an in vivo role in seed pathogen defence and antimicrobial
properties, which make them very attractive as antimicrobial agents for novel medical,
pharmaceutical and food-industry applications. Soft grain texture in wheat grains
depends on the presence of both PIN proteins in their wild type and functional form.
Variations in either or both genes, or lack of expression of either gene, result in a hard
grain texture. Currently, the biochemical basis of the role of PIN proteins in endosperm
texture as well as in antimicrobial functions remains unclear. Plant expression systems
are being developed to produce recombinant PINA and PINB proteins, with the aim to
assess their structure and function. For therapeutic use, production and purification of
high quality PIN proteins is important. One aspect of this study focussed on the rapid
transient expression of PINs for better yield using the deconstructed tobacco mosaic
virus-based ‘magnICON®’ plant expression system. This system was used to test the
subcellular localisation of PINs in different compartments of Nicotiana benthamiana
cells (cytosol, chloroplast, apoplast and endoplasmic reticulum). The results indicated a
profound impact of the cellular compartment on the yield and stability of the PIN
proteins. The presence of recombinant PINA and PINB was confirmed using western
blot and ELISA analyses. Maximum yields of His-tagged recombinant PINA and PINB
occurred when proteins were targeted to the chloroplast. Each PIN protein was shown
to occur in both monomeric and oligomeric forms, which addresses some of the
observations in the literature related to their functionality in determining wheat grain
texture. Affinity purification systems using His-tag and hydrophobic interactions were
used to purify the recombinant PINs. The purified proteins exhibited antibacterial and
antifungal activities, suggesting they were correctly folded. There is evidence that PINs
may interact co-operatively or interdependently to confer the soft grain texture and
influence in pathogen protection. Therefore, as a second major focus of this work,
interactions between PINs were investigated using the Bimolecular Fluorescence
Complementation (BiFC) system in a plant system. Results based on the fluorescence
intensity obtained and confirmed in vivo interactions between PINA and PINB and also
between PINB and PINB in planta. In order to investigate the regions involved in the

ii
protein-protein interactions of PINs, clones constructed previously containing deletion
of the tryptophan-rich domain (TRD) in PINA and the hydrophobic domain (HD) in
both PINs were used for cloning into BiFC vectors. The result indicated that no
interaction occurred between the mutant PIN proteins. The finding shows that both the
TRD and HD regions may have roles in the interactions of PIN proteins, but may
contribute in different ways to their folding and stability.
The work has led to successful expression, purification and characterisation
methodology for obtaining PINs of appropriate quality essential for potential infection
control in diverse areas. Additionally, the work also confirmed their in planta
interactions. To our knowledge, this is the first report of functional PINs produced in N.
benthamiana based on transient expression, and it provides a starting point for
investigating a potential role for PINs on grain texture and antimicrobial abilities.

iii
Acknowledgments
All praises to Almighty God for the strength and His blessing in completing this work.
I would like to thank my principal supervisor, Professor Mrinal Bhave for giving me the
great opportunity of performing this research and for her support, guidance and patience
throughout this project. Thanks particularly for all the time that she has put on the
preparation of this thesis. I would also like to express my deepest appreciation and
gratitude to Dr Diane Webster for acting as a supervisor and being helpful and
supportive from the first day I stepped into the Plant Biotechnology laboratory in
Monash University for three years. Thanks for encouragement and generosity of time in
all meetings during this study.
I am also grateful to Swinburne University of Technology for awarding me the SUPRA
scholarship.
Thanks to Professor Michael Gilding and Dr Elena Verezub for advice and help and Ms
Angela McKellar in Swinburne University. I would also like to thank Professor David
Smyth and Mr Tezz Quon in Monash University for providing plasmid pNBV and
pMLBART. Thanks to Associate Professor Sureshkumar Balasubramanian and his
group for always welcoming me in SKB lab in Monash University.
Thank you to all my lab mates and friends in Monash University; Rasika, Kartika,
Claire, Robert, Indramohan, David, Vincent and Victor.
Thank you to past and present PhD students in Swinburne University; Dr Peter Gollan
and Dr Rebecca Alfred for always sharing their knowledge. Many thanks to Dr
Abirami Ramalingam, Dr Runyararo Hove, Dr Shanthi Joseph and Dr Kaylass Poorun
for all their help, encouragement and friendship. Thank you to my other friends; Bita,
Narges, Farnaz, Saifone, Atul, Yen, Guri, Nadin, Rasika, Jafar, June, Elisa, Dhivya,
Snehal, Rashida and Chris.

iv
Dedicated to
The memory of my dear father; Rasoul
and
My beloved mother; Zahra
For her loving support and always believing in me

v
Declaration
I, Azadeh Niknejad, declare that the PhD thesis entitled ‘Optimisation of expression of
recombinant puroindolines and analysis of protein-protein interactions in planta’ is no
more than 100,000 words in length, exclusive of tables, figures, appendices, references
and footnotes. This thesis contains no material that has been submitted previously, in
whole or in part, for the award of any other academic degree or diploma, and has not
been previously published by another person. Except where otherwise indicated, this
thesis is my own work.
Azadeh Niknejad
June 2014

vi
Abbreviations
BDT Big Dye Terminator
BiFC Bimolecular Fluorescence Complementation
bp Base pair
BSA Bovine Serum Albumin
C-terminal Carboxyl terminal (of a protein)
Cys (or C) Cysteine
DNA Deoxyribonucleic acid
dNTP Deoxyribonucleotide (A, T, G or C)
dpi Days post infiltration
EDTA Ethylenediaminetetraacetic acid
ELISA Enzyme linked immunosorbent assay
ER Endoplasmic Reticulum
g Centrifuge force
gDNA Genomic DNA
GFP Green Fluorescent Protein
GSP-1 Grain Softness Protein-1 (a component of friabilin)
His (or H) Histidine
IPTG Isopropyl β-D-1-thiogalactopyranoside
Kb Kilobase pairs; 1kb= 1000 bp
kDa KiloDalton
LB Luria Bertani broth
MALDI-TOF Matrix-assisted laser desorption ionisation-time of flight
mg Milligram
mL Millilitre
MW Molecular Weight
N-terminal Amino terminal (of a protein)
OD Optical Density
ORF Open Reading Frame
PCR Polymerase Chain Reaction
Pin Puroindoline genes
Pina Puroindoline-a gene
Pinb Puroindoline-b gene

vii
PIN(s) Puroindoline protein(s)
PINA Puroindoline-a protein (wild type)
PINB Puroindoline-b protein (wild type)
rPINA Recombinant PINA
rPINB Recombinant PINB
PPI Protein-Protein Interaction
RBCs Red Blood Cells
rpm Revolutions per minute
SDM Site-directed mutagenesis
SDS-PAGE Sodium dodecyl sulphate-poly acrylamide gel electrophoresis
TAE Tris acetate ethlenediaminetetraacetic acid buffer
TBSV Tomato Bushy Stunt Virus
Trp (or W) Tryptophan
TRD Tryptophan-rich domain
TSP Total Soluble Protein
UV Ultra Violet
X-gal 5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside
YFP Yellow Fluorescent Protein
YC C-terminal fragment of YFP
YN N-terminal fragment of YFP
°C Degree Celsius
List of databases/softwares
ApE A plasmid Editor
BioEdit Biological sequence alignment editor
BLAST Basic Local Alignment Search Tool
COTH Threading-recombination approach
ExPASY Expert Protein Analysis System
I-TASSER Iterative Threading Assembly Refinement
NCBI National Center for Biotechnology Information
PyMOL Python-enhanced molecular graphics tool
WoLF PSORT PSORT II program for protein subcellular location prediction

viii
Table of contents
Chapter 1 General introduction and literature review 1
1 General introduction 2
1.1 Introduction to wheat 2
1.1.1 Evolution of wheat species 3
1.1.2 Structure of the wheat grain 5
1.2 Grain texture in wheat 6
1.2.1 Friabilin 7
1.3 Puroindoline genes and grain texture 8
1.3.1 Mutations, duplications and deletions in Pin genes 8
1.3.2 Point mutations, duplications and deletions in Pina-D1 gene 9
1.3.3 Point mutations, duplications and deletions in Pinb-D1 gene 9
1.3.4 Pinb-2 genes 10
1.3.5 Pin-like genes in related species 12
1.4 Biochemical properties and structure of PIN proteins 13
1.4.1 Heterologous expression of recombinant PIN proteins 18
1.4.2 Evidence of transgenic Pin genes altering grain texture 20
1.5 Evidence of in vitro antimicrobial activity of PINs 22
1.5.1 Evidence of in vivo antimicrobial activity of PINs 23
1.5.2 Evidence of antimicrobial activity of PIN-based peptides 25
1.5.3 Possible mode of action for PINs as antimicrobial proteins and
peptides
25
1.5.4 Possible interactions between PINs 29
1.6 Plant expression system 30
1.6.1 Host plants 31
1.6.1.1 Nicotiana tabacum (Tobacco) 31
1.6.1.2 Nicotiana benthamiana 32
1.6.2 Transformation method and expression systems 32
1.6.2.1 Transient expression using viral vectors 33
1.6.3 Subcellular location in plant system for protein accumulation 34
1.6.3.1 Cytosol 35
1.6.3.2 Endoplasmic Reticulum (ER) 35

ix
1.6.3.3 Apoplast 35
1.6.3.4 Vacuole 35
1.6.3.5 Chloroplast 36
1.6.4 Plant systems for heterologous production of antimicrobial
peptide
36
1.7 Summary of the literature and aims of the research 39
1.7.1 Summary of the above literature 39
1.7.2 The aims of the project 40
Chapter 2 Materials and methods 41
Equipment and materials
2.1 Equipment 42
2.2 Commercial kits 43
2.3 Primary and secondary antibodies 44
2.4 Commonly used buffers, other solutions and media 44
2.4.1 Protein purification buffers 46
2.5 Microbial strains 46
2.5.1 Strains used as hosts for gene cloning 46
2.5.2 Strains used for testing of antimicrobial activity of
recombinant PINs
47
2.6 Plant material 47
2.6.1 Propagation of wheat seedlings 47
2.6.2 Propagation of Nicotiana benthamiana seedlings 47
2.7 General molecular methods 48
2.7.1 Genomic DNA extraction 48
2.7.2 Plasmid DNA purification 49
2.7.3 Spectrophotometric quantification of DNA 49
2.7.4 Agarose gel electrophoresis 49
2.8 The polymerase chain reactions (PCR) for gene amplifications 50
2.8.1 Design and synthesis of primers 50
2.8.2 Design of primers for amplification of full-length Pin genes 50
2.8.3 Design of primers for directional cloning into magnICON®
and BiFC vectors
51
2.8.4 Typical PCR conditions 51

x
2.8.5 PCR for directional cloning of Pin genes into plant expression
vectors
51
2.8.6 Purification of PCR products 52
2.9 Cloning and DNA sequencing 52
2.9.1 Restriction enzyme digestions of PCR products and vectors 52
2.9.2 Ligation reactions 53
2.9.3 Preparation of chemically competent cells of E. coli Mach1 53
2.9.4 Transformation of E. coli Mach1 competent cells by heat
shock
54
2.9.5 Screening of E. coli transformants containing recombinant
plasmids by colony PCR
54
2.9.6 Preparation of electro-competent A. tumefaciens cells 54
2.9.7 Electroporation of electro-competent A. tumefaciens cells 55
2.9.8 Screening of A. tumefaciens transformants containing
recombinant plasmids by colony PCR
55
2.9.9 DNA sequencing 56
2.10 Bioinformatics methods 56
2.10.1 Sequence alignments 56
2.10.2 Plasmid vector map 56
2.10.3 Predictions of the isoelectric point (pI) and molecular weight
(Mw) of PIN proteins
57
2.10.4 Prediction of protein subcellular localisation 57
2.10.5 Modeling and structure prediction of proteins 57
2.10.6 Protein-Protein complexes structure prediction 58
2.11 Biochemical methods for protein analysis 58
2.11.1 Protein concentration measurements 58
2.11.2 SDS-PAGE 59
2.11.3 Western blotting analysis for PIN proteins detection 59
2.11.4 Enzyme-linked immunosorbent assay (ELISA) for
approximate quantitation of recombinant PIN proteins
60
2.12 Testing of antimicrobial properties of the in planta expressed
PINs and a PIN-based synthetic peptide
61
2.12.1 Design of synthetic peptide based on PINA 61

xi
2.12.2 Antibacterial activity assay 61
2.12.3 Antifungal activity assay 62
2.12.4 Stability testing of recombinant PIN proteins 63
2.12.5 Haemolytic activity assay 63
Methods specific to Chapter 3
2.13 Expression of PIN proteins in N. benthamiana using
magnICON® viral vector and characterisation of their
biochemical and antimicrobial properties
64
2.13.1 Principles of the magnICON® viral vector systems for in
planta expression
64
2.13.2 Design of primers for directional cloning into the 3’ module
pICH11599
67
2.13.3 Amplifications of Pin genes, restriction enzyme digestion,
cloning into the 3’ module pICH11599 and sequencing of
clones
68
2.13.4 Electroporation of recombinant pICH11599 and other
modules into A. tumefaciens
70
2.13.5 Agroinfiltration of N. benthamiana using the magnICON®
viral vectors
71
2.13.6 GFP expression 72
Protein methods
2.13.7 Extraction of total soluble protein (TSP) from N. benthamiana
leaf
72
2.13.8 Protein concentration measurements, SDS-PAGE, western
blotting for detection and ELISA for approximate quantitation
of PINs
73
Methods for further PIN protein purification
2.13.9 His-tag purification under native conditions 73
2.13.10 His-tag purification under denaturing conditions 74
2.13.11 Hydrophobic interaction purification 74
2.13.12 Dialysis 75
2.13.13 Concentrate of dilute protein samples 75
2.13.14 Sodium phytate precipitation 75

xii
2.13.15 Testing of antimicrobial and haemolytic properties of the
expressed PINs
76
Methods specific to Chapter 4:
2.14 Protein-protein interactions of PIN proteins using BiFC
system
76
2.14.1 Principle of the Bimolecular Fluorescence Complementation
(BiFC) system
76
2.14.2 New BiFC Vector (pNBV) 77
2.14.3 Design of primers for directional cloning into BiFC vectors
(pNBVs)
78
2.14.4 Restriction enzyme digestion of PCR products 79
2.14.5 Cloning of PIN-YFP fragments into pMLBART vector 80
2.14.6 Electroporation into A. tumefaciens GV3101 81
2.14.7 Use of TBSV-P19 vector 81
2.14.8 Agroinfiltration of N. benthamiana using BiFC vectors 82
2.14.9 Fluorescence microscope and imaging 82
2.14.10 Protein methods for detection of BiFC 82
2.14.11 Native gel electrophoresis and detection of YFP fluorescence 83
2.15 Identification of any protein-protein interaction regions of
PINs using BiFC
85
2.15.1 Gifts of gene construct containing deletions of the tryptophan-
rich domain (TRD) of PINA and the hydrophobic domain
(HD) of PINA and PINB
85
2.15.2 Cloning of PINA∆TRD and PINA∆HD and PINB∆HD in
pNBV vectors
86
2.15.3 Cloning of PINA∆TRD and PINA∆HD and PINB∆HD into
the 3’ module pICH11599
87
Chapter 3 Results 88
3.1 Introduction 89
3.2 Expression of PINs using magnICON® viral vectors 91
3.2.1 Cloning PINA and PINB into magnICON® viral vector
system
91
3.2.2 Transient expression of PIN proteins in the viral vector system 94

xiii
3.2.3 Expression of Green Fluorescent Protein (GFP) by the viral
vector system
95
3.2.4 Expression of the recombinant PINA protein by the viral
vector system
97
3.2.5 Expression of the recombinant PINB protein by the viral
vector system
100
3.2.6 Optimisations of expression of His-tagged recombinant PINA
and PINB
102
3.3 Results of purification of PINA and PINB 105
3.3.1 His-tag purification under native conditions 105
3.3.2 His-tag purification under denaturing conditions 107
3.3.3 Hydrophobic interaction purification 108
3.3.4 Western blot analysis using anti His-tag for PIN proteins 111
3.4 Results of antimicrobial activity tests 112
3.4.1 Antibacterial activity 112
3.4.2 Antifungal activity 114
3.4.3 Protein stability at different temperature 116
3.4.4 Haemolytic activity 117
3.5 Discussion 118
3.5.1 Expression 118
3.5.2 Purification 120
3.5.3 Characterisation of bioactivity 122
Chapter 4 Results 125
4.1 Introduction 126
4.2A Interaction studies for wild type PINA and PINB 127
4.2.1 Cloning of wild type PINA and PINB into pNBV vectors of
the BiFC system
127
4.2.2 BiFC system for transient expression in N. bethamiana leaves 132
4.2.3 Post-transcriptional gene silencing (PTGS) in transient system 132
4.2.4 Fluorescence detection for wild type PIN protein-protein
interactions
133
4.2.5 Western blot analysis of wild type PIN protein-protein
interactions
135

xiv
4.2.6 Gel detection of YFP fluorescence for wild type PIN protein-
protein interactions
136
4.2.B Interaction studies of mutagenesed PINA and PINB 137
4.2.7 Cloning of the previously made constructs PINA∆HD and
PINB∆HD into pNBV vectors
137
4.2.8 Detection of mutant PINA∆HD and PINB∆HD protein-
protein interactions by fluorescence and western blot
141
4.2.9 Cloning of PINA∆TRD into pNBV vectors 142
4.2.10 Fluorescence detection for mutant PINA∆TRD protein-protein
interactions
144
4.2.11 Expression of mutant PIN proteins in magnICON® system 145
4.2.12 Bioinformatics analysis of PIN sequences for potential
interaction domain(s)
149
4.3 Discussion 155
Chapter 5 General discussion and future directions 159
5.1 General discussion 160
5.2 Future directions 163
References 165
Appendices 200

xv
List of Figures
1.1 Schematic representation of the evolutionary history of wheat
species (Triticum and Aegilops)
4
1.2 Composition of the wheat kernels: the endosperm, germ and bran 5
1.3 Alignment of amino acid sequences of GSP-1 from common wheat 8
1.4 Alignment of amino acid sequences of PINB-2v1 from common
wheat
10
1.5 Alignment of amino acid sequences of SINA and SINB in triticale 12
1.6 Alignment of the primary structure of PINA and PINB 14
1.7 Cysteine backbone and α-helix positioning in PINs and nsLTP 15
1.8 Structure prediction of PINA using I-TASSER 16
1.9 NMR solution structure of PuroA 17
1.10 Structure of PuroA in the presence of SDS (sodium dodecyl sulfate)
micelles
17
1.11 The predicted tertiary structures of PINA and mutants of PINA 18
1.12 Commonly cited models for antimicrobial peptide activity 26
2.1 Wheat and N. benthamiana seeds and plantlets 48
2.2 Diagram of indirect ELISA assay 61
2.3 Schematic showing the assembly of the three viral vector modules
and production of PIN proteins
64
2.4 The 3’module pICH11599 65
2.5 The other magnICON® viral vector modules 66
2.6 Generation of the Pins construct for magnICON® viral vectors 70
2.7 Agroinfiltration of N. benthamiana leaf 72
2.8 Principle of the BiFC assay 77
2.9 The pNBV (New BiFC Vector) modules 78
2.10 Plasmid map of pMLBART 80
2.11 Plasmid map of the P19 vector 81
2.12 Strategy used to construction of BiFC vectors for investigating
potential interactions of PIN proteins
84
2.13 Alignment of putative PINA and PINB 85

xvi
2.14 Generation of the mutant PINs construct for magnICON® viral
vectors
87
3.1 Second-round PCR of gene sections encoding mature PINs 91
3.2 Colony PCR for preliminary selection of clones 92
3.3 Example of double digests of clones in pICH11599 92
3.4 The 3’ module constructs encoding mature PIN with His-tag and
His/SEKDEL tags
93
3.5 N. benthamiana leaves 10 days post-infiltration 94
3.6 GFP production using two different targeting modules, in
N. benthamiana leaves (10 dpi)
95
3.7 Standard curve of BSA concentration for Bradford assay 96
3.8 Western blot analysis to determine GFP expression using anti-GFP
antibody
96
3.9 Coomassie Blue stained gel to analyse TSP from infiltrated leaves 97
3.10 Determination of PINA expression by western blotting of TSP
extracts from infiltrated leaves
98
3.11 Standard curve of 3% non-durum wheat dilutions for ELISA assay 99
3.12 Quantification of PINA proteins targeted to chloroplast, cytosol and
ER by ELISA
99
3.13 Coomassie Blue stained gel to analysis TSP from infiltrated leaves 100
3.14 Determination of PINB expressions by western blotting of TSP
extracts from infiltrated leaves
101
3.15 Quantification of PINB proteins targeted to chloroplast, cytosol and
ER by ELISA
102
3.16 Detection of PIN proteins harvested at different times post-
infiltration, by ELISA
102
3.17 Heat sensitivity of plant-expressed PINA and PINB detected by
western blot
103
3.18 Determination of PINA and PINB protein expression in infiltrated
leaves with viral vector module by western blot of TSP in expected
size
104
3.19 SDS-PAGE of TSP and wash steps of His-tag purification under
native conditions
105

xvii
3.20 SDS-PAGE and western blot analysis with Durotest antibody for
eluted fraction on Ni-NTA resin columns based on His-tag
purification under native conditions
106
3.21 SDS-PAGE and western blot analysis with Durotest antibody for
eluted fraction on columns with 6M urea in elution buffer
107
3.22 Fractions in the course of extraction procedure 108
3.23 SDS-PAGE for fractions eluted from the Butyl-Toyopearl column
based on hydrophobic interaction system
109
3.24 SDS-PAGE of eluted fraction on Butyl-Toyopearl column based on
hydrophobic interaction system and sodium phytate precipitation
109
3.25 Western blot of eluted fractions on Butyl-Toyopearl column based
on hydrophobic interaction system
110
3.26 Western blot analysis of PIN proteins purified with two different
systems
111
3.27 Example of Minimum Inhibitory Concentration (MIC) assay for
PuroA peptide and plant-made recombinant PIN proteins against
E. coli
113
3.28 Example of microdilution plate Minimum Inhibitory Concentration
(MIC) assay for filamentous fungi against R. solani
114
3.29 Haemolytic activity assays against sheep red blood cells (RBCs) 117
4.1 Second-round PCR of gene sections encoding mature PINs and
DNA plasmids of empty pNBV vectors
127
4.2 Colony PCR for preliminary selection of clones 128
4.3 The pNBV module constructs encoding mature PINA and PINB
proteins
129
4.4 Alignments sequence of PINA in pNBV construct 130
4.5 Alignments sequence of PINB in pNBV construct 131
4.6 BiFC visualisation in N. benthamiana leaves 133
4.7 BiFC visualisation of PINA-PINB interactions in N. benthamiana
leaves
134
4.8 BiFC visualisation of PINA-PINA and PINB-PINB interactions in
N. benthamiana leaves
135
4.9 Determination of interacting form of PINA+PINB and PINB+PINB 136

xviii
by western blot analysis from TSP extract
4.10 Native gel electrophoresis and UV imaging of YFP fluorescence of
PIN protein complexes
137
4.11 Amplification of inserts from PINA∆HD and PINB∆HD constructs 138
4.12 The pNBV module constructs generated encoding mutant PIN
proteins
139
4.13 Sequences of PINA∆HD in pNBV construct 139
4.14 Sequences of PINB∆HD in pNBV construct 140
4.15 Infiltrated mixtures of constructs into N. benthamiana leaves side by
side for detect and compare the interaction of wild type and mutant
form of PIN proteins
141
4.16 Western blot analysis to determine the interacting forms of wild
type and mutant PIN proteins
142
4.17 The pNBV module construct generated encoding PINA∆TRD
protein
143
4.18 Sequences of PINA∆TRD in pNBV construct 143
4.19 The 3’ module constructs encoding mutant PIN proteins with His-
tag
145
4.20 Sequences of PINA∆HD-His and PINA∆TRD-His in 3’ module
(pICH11599)
146
4.21 Sequences of PINB∆HD-His in 3’ module (pICH11599) 147
4.22 Determination of wild type and mutant PIN proteins expressions in
infiltrated leaves with chloroplast viral vector module by western
blotting of TSP extract
148
4.23 Structure prediction using PyMOL and prediction secondary
structure for PINA
150
4.24 Structure prediction using PyMOL and prediction secondary
structure for PINB
151
4.25 Modeling result of COTH for PINA and PINB Complex in
horizontal and vertical direction
153
4.26 Modeling result of COTH for PINA∆HD and PINB∆HD Complex 154

xix
List of Tables 1.1 The mutations identified in Pina-D1a and Pinb-D1a alleles in
common wheat (T. aestivum)
11
1.2 Reported recombinant PIN proteins 20
1.3 Reported antibacterial and antifungal activities of PIN proteins 23
1.4 Reported in vivo antimicrobial activity of PINs 24
1.5 Reported antibacterial and antifungal activities of PIN-based
peptides
25
1.6 Summary of factors affecting gene transcription, mRNA translation
and protein accumulation
31
1.7 AMPs heterologous produced in Nicotiana tabacum 38
2.1 Instruments and apparatus used 42
2.2 Commercial kits 43
2.3 Antibodies for western blot analysis 44
2.4 Composition of general buffers and solutions 44
2.5 Composition of protein purification buffers 46
2.6 Primer used for the wild type full length Pina-D1 and Pinb-D1
amplifications
51
2.7 Primers used in directional cloning into 3’ viral module, pICH11599 68
2.8 Genes cloned with His-tag and SEKDEL tag into pICH11599 68
2.9 Tri-partatite infiltration of magnICON® constructs into
N. benthamiana
71
2.10 Protein extraction buffers used in this study 73
2.11 Primers used in directional cloning into pNBV vectors 79
3.1 Quantification of TSP for targeted expression of GFP using
Bradford assay
96
3.2 ELISA data analysis for quantification of recombinant PINA
(rPINA) protein
99
3.3 ELISA data analysis for quantification of recombinant PINB
(rPINB) protein
101
3.4 Antibacterial activity of plant made recombinant PINA and PINB 113
3.5 Antifungal activity of plant made recombinant PINA and PINB 115

xx
3.6 Stability of recombinant PINs at different incubation temperature 116
3.7 Haemolytic activity of plant-made recombinant PINs 117
4.1 BiFC constructs for PIN proteins (wild type and mutant) to detect
fluorescence signal
144

CHAPTER 1
General introduction and literature review

Chapter 1 Introduction and literature review
- 2 -
1. General introduction
The information in this chapter provides an extensive review of the literature on
puroindoline (Pin) genes and puroindoline (PIN) proteins, their specific function in
controlling grain texture, as well as their possible biological roles as antimicrobial
proteins in wheat seeds.
Following the general overview, the introduction also highlights the importance of:
Understanding the roles of PIN proteins in lipid binding and impact on grain hardness
Identifying further PINs or PIN-based peptide structures
Plants as an alternative system for the expression of proteins in order to study the
structure and function of antimicrobial proteins and peptides
1.1 Introduction to wheat
Wheat (Triticum aestivum L.) is one of the most widely consumed grain crops in the
world. It is no doubt one of the most important cereal crops for humankind, with unique
end-use qualities that allow it to be processed into a range of flour-based foods such as
bread, biscuit and pastry products; additionally, it also has a role in the production of
starch and gluten (Li et al., 2012; Shewry, 2009). Wheat in the form of bread provides
essential nutrients like fibre, carbohydrates, protein, B vitamins, iron, calcium,
phosphorus, zinc, potassium and magnesium (Painter et al., 2002; Pena et al., 2002;
Wrigley, 2009). Wheat ranks third after maize (corn) and rice in terms of worldwide
production (Shewry, 2013).
Originating in the Middle East region about 10,000 years ago, wheat is the largest grain
crop based on area of cultivation (Dubcovsky and Dvorak, 2007; Matsuoka 2011;
Wrigley, 2009). Among agricultural crops, wheat is well adapted to a wide range of
soils, climates, and environmental conditions, unlike rice and maize that prefer tropical
environments (Gill et al., 2004). Durum wheat (Triticum turgidum L. var. durum) is
more adapted to the dry Mediterranean climate than bread wheat (Triticum aestivum L.)
(Shewry, 2009). Wheat varieties are categorised into six major classes based on
planting and harvesting dates, as well as hardness, shape and the colour of the kernels:
(1) hard red spring; (2) hard red winter; (3) soft red winter; (4) hard white wheat; (5)
soft white wheat; (6) durum (Rochelle, 2001).

Chapter 1 Introduction and literature review
- 3 -
1.1.1 Evolution of wheat species
A detailed understanding for the origin of cultivated wheat has extended our knowledge
for improving it genetically. Wheat (genus Triticum) belongs to the Poaceae (or
Gramineae) family. The genus has three subclasses of ploidy: diploid (2n=2x=14),
tetraploid (2n=4x=28) and hexaploid (2n=6x=42) (Eckardt, 2001; Feldman, 1995;
Feldman, 2001). Three genomes, designated A, B and D, are involved in the formation
of the polyploidy series with seven pairs of chromosomes (Feldmann, 2001). Bread
wheat (Triticum aestivum), the most widely cultivated wheat today, is hexaploid
AABBDD and its origin has most likely occurred through one or more rare
hybridisation processes. Wild einkorn T. urartu (AA) crossed through natural
hybridisation with Aegilops speltoides (BB) to produce wild emmer T. turgidum L. ssp.
durum (AABB) (Chantret et al., 2005; Matsuoka, 2011). T. urartu is the diploid
progenitor of the A genome in tetraploid and hexaploid wheats (Haider, 2013). Another
hybridization between the tetraploid T. turgidum L. ssp. dicoccum (AABB) and the
diploid donor of the D genome, Ae. tauschii (2n =14, DD) restored the Hardness (Ha)
locus in T. aestivum (Chantret et al., 2005; Wrigley, 2009). The D genome is present in
T. aestivum but not in T. turgidum L. ssp. durum. Probably as a result of transposable
element insertions and two large deletions caused by illegitimate recombination, the Ha
locus in the D genome of hexaploid wheat (T. aestivum) is 29 kb smaller than in the D
genome of diploid Ae. tauschii (Chantret et al., 2005; Haider, 2013). Numerous studies
have attempted and published reports about the origin of the B genome in cultivated
wheat; however, the B genome donor remains uncertain. It is believed that the B
genome is closely related to the S genome from the Sitopsis group of the genus Ae.
speltoides (Feldman, 2001; Haider, 2013). The formation of tetraploid and hexaploid
wheat is summarised in Figure 1.1.

Figure 1.1 Schematic representation of the evolutionary history of wheat species (Triticum and Aegilops). Source: modified from Chantret et al. (2005) and Dubcovsky and Dvorak, (2007).

Chapter 1 Introduction and literature review
- 5 -
1.1.2 Structure of the wheat grain
Wheat kernels consist of three distinct layers: the endosperm, germ and bran (Shewry,
2013) (Figure 1.2). The major part of wheat grain is endosperm and approximately
three-quarters of the endosperm includes starch, which has an important role in
determining the end-use of wheat grain. Differences in the endosperm texture of wheat
are an extremely important characteristic that have an effect on flour quality and yield
for human consumption. White flour contains the starchy endosperm, which contains a
high proportion of starch and gluten (Delcour and Hoseney, 2010; Turnbull and
Rahman, 2002).
Figure 1.2 Composition of the wheat kernels: the endosperm, germ and bran.
The germ is the embryo of the kernel and comprises only about 2.5% of the weight.
Wheat germ contains protein (~25%) and lipids (~8%), and is also an important source
of vitamins B and E (Cornell, 2003; Slavin, 2003). The bran layers surrounding the
endosperm are rich in vitamins and minerals and contains high-quality protein (19%), as
well as large amounts of insoluble dietary fibre (Wrigley et al., 2004). The bran layer
accounts for approximately 14.5% of the weight of the kernel. The aleurone layer is the
outermost layer of the endosperm and is located between the bran layer and endosperm.
The bran and aleurone layers play a role in protecting the embryo and nutrient-rich
endosperm against stress and pathogens (Gillies et al., 2012; Jerkovic et al., 2010).
Endosperm
Germ
Bran

Chapter 1 Introduction and literature review
- 6 -
1.2 Grain texture in wheat
Wheat hardness is a major quality trait and is defined as the force needed to crush the
kernels, which is required for the marketing and technological utilisation of wheat
(Morris, 2002), and also relates to the degree of hardness or softness of the grain. Grain
hardness is an essential factor, as it impacts milling and baking in the flour industry
(Bettge et al., 1995). Overall flour particle size, shape and flour density, starch damage,
water adsorption and milling yield are physical properties affected by hardness (Martin
et al., 2001). Grain hardness defines the quantitative variations within and across three
qualitative classes: ‘soft’ hexaploid wheats, ‘hard’ hexaploid wheats and the ‘very hard’
durum wheats. Moreover based on intermediate hardness, wheat has been classified as
‘medium hard’ and ‘medium soft’ (Morris, 2002). Bread wheat endosperm texture
varies from extremely soft to hard, while durum wheat has the hardest kernels of all
wheat species. In comparison with hard wheat, soft wheat generally yields flour with a
smaller average particle size and lower levels of damaged starch. Differences in
endosperm texture between hard and soft wheats are the most important factors for
functionality and marketing of wheat (Mikulikova, 2007). Generally, different types of
wheat are required by breeders, millers and bakers. Soft wheat flour with low protein
content is used to make cakes and pastries while hard wheat flour is suited for bread in
addition to other yeast raised products. Durum wheat with unique properties like high
protein content and gluten strength is used for pasta products and semolina (Douliez et
al., 2000; Morris, 2002).
Wheat endosperm hardness is controlled by genetic factors and biochemical factors
such as seed moisture, lipid and pentosan content. However, environmental conditions
also affect endosperm hardness (Konopka et al., 2005; Turnbull and Rahman et al.,
2002). Grain hardness has been linked to one major and several minor loci (Mattern et
al., 1973; Symes, 1965, 1969). The major locus was named Hardness (Ha) and shown
to be located at the distal end of the short arm of chromosome 5D (Doekes and
Belderok, 1976; Law et al., 1978; Mattern et al., 1973). Hard wheat has shown stronger
adhesion between starch granules and storage protein compared to soft wheat
(Simmonds et al., 1973). Studies conducted using scanning electron microscopy have
also revealed that more gluten adheres to the surface of starch granules isolated from

Chapter 1 Introduction and literature review
- 7 -
hard wheat than to the surface isolated from soft wheat (Barlow et al., 1973; Hoseney
and Seib, 1973). It has been suggested that the degree of adhesion between starch and
gluten proteins is one of the essential factors for differences in endosperm hardness
between hard and soft wheat (Barlow et al., 1973; Hoseney and Seib, 1973; Simmonds
et al., 1973; Pauly et al., 2013).
1.2.1 Friabilin
Endosperm hardness is related to the occurrence of a 15 kDa protein complex called
‘friabilin’ on the surface of water-washed starch granules (Greenwell and Schofield,
1986). The protein was named ‘friabilin’ as it was observed that soft wheat was more
friable than hard wheat, while durum wheat lacked this protein (Morrison et al., 1992;
Morris, 2002). The amounts of starch granule associated protein from water washed
starch differ in soft and hard wheats (Jolly, 1993; Jolly et al., 1996; Rahman et al.,
1994). The occurrence of friabilin on the surface of water-washed starch granules
appears to be related to the level of bound polar lipids (glycolipids and phospholipids).
Polar lipids are present in high levels on the starch granule surface in soft wheat, but in
low levels in hard wheat (Capparelli et al., 2003; Greenblatt et al., 1995). Furthermore,
Greenblatt et al. (1995) showed that friabilin can be extracted with a propan-2-ol/water
(90:10) mixture, which is effective for removing starch-bound polar lipids. However,
with the same buffer, when no lipids are removed earlier, only a very small amount of
friabilin can be extracted. Friabilin components associated with starch granules through
polar lipids were shown to be involved in ionic and hydrophobic interactions
(Greenblatt et al., 1995). Friabilin protein analysis revealed that it is likely a mixture of
several polypeptides (Blochet et al., 1993; Morris et al., 1994; Oda and Schofield,
1997). The two major friabilin components are puroindoline-a (PINA) and
puroindoline-b (PINB) (Gautier et al., 1994). Grain Softness Protein-1 (GSP-1) and α-
amylase inhibitors (Rahman et al., 1994) are the minor components (Blochet et al.,
1993; Morris et al 1994). The GSP-1 protein (Figure 1.3) with approximately 164
amino acids long exhibits 40% identity and 60% similarity to the PINs with 10 cysteine
backbone and TRD with two tryptophan residues (Bhave and Morris, 2008a; Rahman et
al., 1994). Elmorjani et al. (2013) have shown that the GSP-1 protein may undergo
different proteolytic cleavages in the N and C-terminal regions of the pre-pro-protein.

Chapter 1 Introduction and literature review
- 8 -
Figure 1.3 GSP-1 putative protein sequence from common wheat. (T. aestivum) GenBank accession number: S48186
1.3 Puroindoline genes and grain texture
The Puroindoline (Pin) genes Pina-D1 and Pinb-D1, responsible for encoding the wild
type puroindoline proteins, are part of the Ha locus on the short arm of chromosome 5D
in common wheat (Gautier et al., 1994; Ragupathy and Cloutier, 2008). However,
during evolution and after polyploidisation of cultivated polyploid durum wheat, Pin
genes were deleted from chromosomes 5A and 5B. The Pin (Pina-D1, Pinb-D1) and
Gsp-1 genes are located in a 60 kbp DNA fragment of the Ha locus of Ae. tauschii,
which is the D-genome donor of T. aestivum (Pauly et al., 2013). Pin genes are located
only on the D-genome, while GSP-1 is present on all three wheat genomes (A, B and D)
in diploid, tetraploid and hexaploid wheat (Chantret et al., 2004, Chantret et al., 2005;
Morris 2002; Rahman et al., 1994). Without the D genome and thus both Pin genes and
also PIN proteins, resulting in the very hard durum wheats (Chantret et al., 2005).
1.3.1 Mutations, duplications and deletions in Pin genes
The Puroindoline genes (Pina-D1, Pinb-D1) contain 447 base pairs (bp) without introns
and both genes are 70.2% identical in the coding regions, but only 53% identical in the
3’-untranslated region (Gautier et al., 1994). The presence of both Pina-D1 and Pinb-
D1 genes in functional forms results in soft endosperm texture; however, mutations in,
or absence of either genes result in low levels of PIN protein(s) and hard endosperm
(Giroux and Morris 1998, Wall et al, 2010). The most common form of mutations are
single nucleotide polymorphisms (SNPs), which may result in an amino acid
substitution, or replacement with a stop codon, or a frame shift caused by a single base

Chapter 1 Introduction and literature review
- 9 -
insertion or deletion (INDELs). The first report to show mutations in both Pin genes in
common wheats was by Chang et al. (2006). The mutations identified to date in Pina-
D1a and Pinb-D1a genes are listed in Table 1.1 (reviewed by Bhave and Morris, 2008a;
Nadolska-Orczyk et al., 2009).
1.3.2 Point mutations, duplications and deletions in Pina-D1 gene
The most common hardness-associated mutation to be reported in Pina-D1 was a null
mutation (Pina-D1b) and has a harder texture than other prevalent hardness alleles
(Giroux and Morris 1998; Morris, 2002. This results in the absence of Pina transcripts
and a lack of the PINA proteins in the kernel (Giroux and Morris 1998; Pauly et al.,
2013). Point mutations or SNPs have been reported in Pina-D1 genes (Chen et al.,
2006; Gazza et al., 2005; Massa et al., 2004). Chen et al. (2006) described different
SNPs in the mature protein sequence, which are: a) premature stop codon replacing
tryptophan at position 43 (Pina-D1n), b) proline to serine at position 35 (Pro35Ser)
(Pina-D1m). Mutations of serine in Pina-D1m may affect the lipid-binding ability of
the TRD due to the location of Ser in a loop between the first helix and the TRD (Bhave
and Morris, 2008a). Moreover, in Asian wheat cultivars, two types of deletion mutation
in Pina-D1 which results in PINA-null products have been reported. The 4.4kb deletion
mutant was designated Pina-D1r and the 10.5kb deletion was designated Pina-D1s
(Ikeda et al., 2010).
1.3.3 Point mutations, duplications and deletions in Pinb-D1 gene
Giroux and Morris (1997) were the first to report a single nucleotide change in Pinb-D1,
leading to the glycine-to-serine change at position 46 in PINB (G46S; Pinb-D1b), which
was associated with the hard texture in common wheat. This mutation has been
discovered widely in wheats around the world (Chen et al., 2006; Lillemo et al., 2006;
Pickering and Bhave, 2007; Tanaka et al. 2007; Xia et al., 2005). Wheats possessing
Pinb-D1b alleles were slightly softer than wheats with the Pina-D1b/Pinb-D1a (Morris,
2002). It has been suggested that this amino acid change affects lipid binding and starch
granule interaction, in addition to altering the tertiary structure of PINB at the TRD
(Bhave and Morris, 2008a). The point mutation of Pinb-D1c that causes a leucine-to-
proline change at position 60 (Leu60Pro) was first reported by Lillemo and Morris,

Chapter 1 Introduction and literature review
- 10 -
(2000). Among the point mutations of Pinb-D1, mutations frequently occur at position
44 of the mature protein sequence, e.g., a tryptophan to arginine change at position 44
(Trp44Arg; Pinb-D1d) (Lillemo and Morris, 2000), tryptophan to early stop codon at
position 44 (Trp44stop codon; Pinb-D1f) (Morris et al., 2001), as well as a Pinb-D1q
(Trp44Leu) (Chen et al., 2005).
1.3.4 Pinb-2 genes
The six alleles of Pinb-2 gene have been reported and designated Pinb-2v1 to Pinb-2v6
(Chen et al., 2010a; Chen et al., 2010c; Ramalingam et al., 2012; Wilkinson et al.,
2008), which provide a potential new resource for minor variation in grain texture.
Mapping of the Pinb-2 genes in wheat reported Pinb-2v1 to be located on chromosome
7D in bread wheat and absent in durum wheat, Pinb-2v2 and Pinb-2v3 are located on
chromosome 7B and Pinb-2v4 is located on chromosome 7A (Chen et al., 2010a; Chen
et al., 2010c). The reported transcript levels of Pinb-2 variants suggest they are only
expressed at 10% the levels of the Pinb-D1 genes in common wheat (Wilkinson et al.,
2008). Additionally, RNA sequencing has shown that relative to Pinb, Pinb-2v1
expression was at 1%, and Pinb-2v2 and Pinb-2v3 was at ~8% of the expression levels
detected, while Pinb-2v4 was undetectable (Giroux et al., 2013). Pinb-2 genes appear
to have a greater impact on the grain texture of soft wheat than hard wheat and are
associated with increased grain texture in soft wheats (Chen et al., 2010b). Moreover,
the putative TRD of the PINB-2v1 proteins with two tryptophan residues (Figure 1.4)
has shown antimicrobial activity in vitro against a number of bacteria and fungi
(Ramalingam et al., 2012).
Figure 1.4 PINB-2v1 putative protein sequence. (T. aestivum) GenBank accession number: ADA7764

*SNP: Single nucleotide polymorphism *NS: Non-synonymous mutation, a change in the nucleotide sequence that does not lead to a change in the amino acid sequence
PIN allele Phenotype PIN protein Change to protein sequence* Reference Pina-D1a Soft Wild-type - Gautier et al. (1994) Pina-D1b Hard PINA null Gene deletion Giroux and Morris (1998) Pina-D1f Hard PINA Three SNPs. Arg58Gln and NS Ala19+Leu52 Massa et al. (2004) Pina-D1k Hard PINA null Gene deletion (Associated with Pinb deletion) Tranquilli et al. (2002)
Ikeda et al. (2005) Pina-D1l Hard PINA truncated Single base deletion, frame shift Gln61Lys, then a premature stop codon at pos120 Gazza et al. (2005) Pina-D1m Hard PINA Single SNP. Pro35Ser Chen et al. (2006) Pina-D1n Hard PINA Single SNP. Trp43stop, premature stop codon Chen et al. (2006) Pina-D1p Hard PINA truncated Single base deletion, frame shift Cys110Ala, then premature stop codon Chang et al. unpublished Pina-D1q Hard PINA Two SNP. Asn111Lys+lle112Leu Chang et al. (2006) Pina-D1r Hard PINA null Gene deletion Ikeda et al. (2010) Pina-D1s Hard PINA null Gene deletion Ikeda et al. (2010) Pina-D1t Hard PIN A truncated Single SNP. Trp41stop, premature stop codon Ramalingam et al. (2012) Pinb-D1a Soft Wild-type - Gautier et al. (1994) Pinb-D1b Hard PINB Single SNP. Gly46Ser Giroux and Morris (1997) Pinb-D1c Hard PINB Single SNP. Leu60Pro Lillemo and Morris (2000) Pinb-D1d Hard PINB Single SNP. Trp44Arg Lillemo and Morris (2000) Pinb-D1e Hard PINB truncated Single SNP.Trp39Stop,premature stop codon Morris et al. (2001) Pinb-D1f Hard PINB truncated Single SNP.Trp44stop,premature stop codon Morris et al. (2001) Pinb-D1g Hard PINB truncated Single SNP.Cys56stop,premature stop codon Morris et al. (2001) Pinb-D1l Hard PINB Single SNP.Lys45Glu Pan et al. (2004) Pinb-D1p Hard PINB truncated Single base deletion, frame shift Lys42Asn, then a premature stop at pos60 Chang et al. (2006) Pinb-D1q Hard PINB truncated Single SNP.Trp44Leu Ikeda et al. (2005) Xia et al.
(2005) Chen et al. (2005) Pinb-D1r Hard PINB truncated Single base deletion, frame shift Glu14Gly, then a premature stop at pos48 Ram et al. (2005) Pinb-D1s Hard PINB truncated Single base deletion and SNP, frame shift Glu14Gly, then a premature stop at pos48 Ram et al. (2005) Pinb-D1t Hard PINB Single SNP. Gly47Arg Chen et al. (2006) Pinb-D1u Hard PINB truncated Single base deletion, frame shift Glu14Ser, then a premature stop at pos18 Chen et al. (2007) Pinb-D1v Hard PINB Two SNPs. Ala8Thr+Leu9lle Chang et al. (2006) Pinb-D1w Hard PINB Single SNP.Ser115lle Chang et al. (2006) Pinb-D1aa Hard PINB truncated Single SNP. Single based deletion, frame shift Lys42Asn, a premature stop codon Li et al. (2008a) Pinb-D1ab Hard PINB truncated Single SNP. Gln99stop, premature stop codon Tanaka et al. (2008)
Table 1.1 The mutations identified in Pina-D1a and Pinb-D1a alleles in common wheat (T. aestivum)

Chapter 1 Introduction and literature review
- 12 -
1.3.5 Pin-like genes in related species
Genes encoding puroindoline-like proteins do not occur in rice, maize or sorghum, but
are present in cereals related to wheat such as rye, barley and oat (Gautier et al., 2000;
Pauly et al., 2013). The PIN homologues in rye (Secale cereal L.) and triticale cultivar
are named secaloindoline a (SINA) and secaloindoline b (SINB) (Li et al., 2006;
Simeone and Lafiandra, 2005) (Figure 1.5). Until now, no relationship between
secaloindoline alleles and grain hardness has been reported in rye since rye cultivars
have generally soft endosperm texture with very low variation in kernel hardness
(Simeone and Lafiandra, 2005). Recently, Gasparis et al. (2013) reported RNAi-based
silencing of Sin genes resulted in a significant decrease in the level of transcripts and the
yield of both secaloindoline proteins but did not affect grain hardness in triticale cultivar
Wanad. Furthermore, barley (Hordeum vulgare L.) contains the PIN homologues
hordoindoline a (HINA) and hordoindoline b (HINB) and have been mapped on the
short arm of chromosome 5H (Beecher et al., 2002b). The Ha locus in barley has been
shown to be associated with variations in endosperm texture (Beecher et al., 2002b). In
addition, PIN homologues have been identified in oat (Avena sativa L.), designated
avenoindoline a and avenoindoline b (Gautier et al., 2000). Immunolocalisation studies
using Durotest antifriabilin antibody have detected tryptophanins (TRPs) proteins in oat
seed associated with the surface of starch granules (Mohammadi et al., 2007).
Figure 1.5 Alignment of amino acid sequences of SINA and SINB in triticale. GenBank accession numbers: AGO65289.1 and AGO65290.1 for SINA and SINB respectively

Chapter 1 Introduction and literature review
- 13 -
1.4 Biochemical properties and structure of PIN proteins
N-terminal sequencing identified the basic cysteine-rich proteins Puroindoline-a (PINA)
and Puroindoline-b (PINB) as the major components of friabilin (Blochet et al., 1993).
The name ‘puroindoline’ is derived from the Greek word ‘puros’ for wheat, and
‘indoline’, referring to the indoline ring of the tryptophan-rich domain (Gautier et al.
1994). The PINs and other proteins (for instance storage proteins and the enzyme
granule-bound starch synthase I) with affinity for defatted starch granules were also
isolated from wheat endosperm using the nonionic detergent Triton X-114 (Blochet et
al., 1993; Bako et al., 2007).
PINA and PINB contain tryptophan residues in the unique tryptophan-rich domain
(TRD) and a highly conserved cysteine-rich backbone of 10 cysteine residues (Blochet
et al., 1993). The TRD of PINA consists of five tryptophan residues (WRWWKWWK;
position without the putative signal paptide: 38 to 45), while in PINB, it consists of
three (WPTKWWK; position without the putative signal paptide: 39 to 45) (Blochet et
al., 1993; Gautier et al., 1994). The full length PIN proteins consist of 148 amino acids
with a similar molecular mass of 13 kDa (Gautier et al., 1994). PIN proteins are highly
basic, with a calculated isoelectric point (pI) of 10.5 for PINA and 10.7 for PINB and
show 55% similarity at the amino acid level (Gautier et al., 1994). Both PINs are
synthesized as pre-pro-proteins containing an N-terminal signal peptide, which
comprises the first 28 amino acid residues for PINA and the first 29 amino acid
residues
for PINB (Gautier et al., 1994) (Figure 1.6). These N-terminal signal peptides may have
a role in intracellular targeting (Bhave and Morris, 2008a; Gautier et al., 1994). C-
terminal processing has also been suggested for both PINs and can result from slightly
different post-translation processing pathways (Day et al., 2006). Raman spectroscopy
studies have shown that PINA and PINB have a similar secondary structure at pH 7.
Both PINs consist of approximately 30% α-helices, 30% ß-sheets and 40% unordered
structures (Le Bihan et al., 1996). Far-UV circular dichroism (CD) measurements have
confirmed similarity between PINs in secondary structure (Kooijman et al., 1997).

Chapter 1 Introduction and literature review
- 14 -
The 10 cysteine residues form five intramolecular disulphide bonds. Disulphide bonds
are essential for stabilizing the α-helical structure and to maintain the native structure
and solubility of PINA and PINB (Le Bihan et al., 1996). Eight of the cysteines form a
particular pattern known as the “eight cysteine motif” (8CM), which appears widely
among plant proteins and has a similar tertiary structure, including four α-helical
structures (Figure 1.6) and variable loops (Pauly et al., 2013). Proteins containing this
motif show a wide range of functions in storage, enzyme inhibition, lipid transfer, plant
defence and cell wall structure (José-Estanyol et al., 2004). The cysteine residues may
have an important role in forming a network of disulphide bridges, which are important
for the maintenance of the three-dimensional (3D) structure of the molecule, together
with the central helical core (José-Estanyol et al., 2004). The TRD is located between
two additional cysteine residues. Kooijman et al. (1997) suggested that the TRD of
PINs has a role in the stability of the structure. The TRD confers hydrophobicity for
strong affinity in PIN interactions with lipids (Douliez et al., 2000; Kooijman et al.,
1997).
Figure 1.6 Alignment of the primary structure of PINA and PINB. Signal peptide in both PINs is highlighted before potential N-terminal cleavage sites (after the 28th
residue for PINA and after the 29th
residue for PINB). The 4-alpha helices are as
determined by Le Bihan et al. (1996) with the following locations, respectively for PINA and PINB: residues 17-28 and 18-29 for H1, 49-62 and 50-63 for H2, 69-76 and 70-77 for H3, and 87-100 and 89-101 for H4.

Chapter 1 Introduction and literature review
- 15 -
The high resolution structure of PINs has some limitations due to the strong aggregative
properties at high protein concentration and difficulties in obtaining a stable non-
aggregated solution required for crystallisation and NMR characterisation (Clifton et al.,
2011a; Marion et al., 2007). The 3D structure of PINs has not yet been determined.
However, a considerable amount of literature has been published on predicted models.
The first of these was based on the folding pattern of the eight-cysteine motif in
nonspecific lipid transfer proteins (ns-LTP). The eight-cysteine motif, including the
Cys-Cys and Cys-arginine-Cys characteristic of both PINs, is also found in nsLTPs.
The similarities of the disulphide bonds support the suggestion that nsLTPs and
puroindolines are related in their primary and secondary structure, and display similar
folding properties (Douliez et al., 2000; Marion et al., 2007) (Figure 1.7). PINs have
five disulphide bonds while ns-LTPs have four. The TRD of PINs is located in a loop
between the first and second α-helix, outside the protein based on the 3D model for ns-
LTP (Doulies et al., 2000; Kooijman et al., 1997; Marion et al., 1994). In addition, the
variable loops connected to the α-helices are the functional regions of the eight-cysteine
motif proteins (José-Estanyol et al., 2004), which seems appropriate for PINs. There is
also a unique tyrosine residue (positions without the putative signal paptide: 23 and 24
for PINA and PINB respectively) in the first α-helix, which may be functionally
important. Tyrosine is not bound to a negatively charged carboxylate of the protein, but
is hydrogen-bonded to water molecules (Le Bihan et al., 1996).
Figure 1.7 Cysteine backbone and α-helix positioning in PINs and nsLTP. Source: Douliez et al. (2000)

Chapter 1 Introduction and literature review
- 16 -
Recently, a different 3D structure model of PINA using iterative threading assembly
refinement (I-TASSER) has been revealed for structure prediction by Lesage et al.
(2011) (Figure 1.8). The prediction result for PINA displayed the five disulphide
bonds: Cys20/Cys55 and Cys56/Cys104 are connected and readily form disulphide
bonds; Cys11/Cys66 and Cys68/Cys110 are positioned at close proximity (Lesage et al.
2011) and the TRD is an extension stabilised by a Cys28/Cys48 disulphide bridge in
PINA and a Cys29/Cys48 disulphide bridge in PINB (Le Bihan et al., 1996). PINs have
several features of membrane proteins, such as high levels of α-helices. In addition,
both PIN proteins can be extracted with non-ionic detergent like TX-114; however,
none of them contains a typical trans-membrane region. Moreover, after extraction in
the absence of detergents, puroindolines are fairly water-soluble, and act differently
from membrane proteins (Lesage et al., 2011).
Figure 1.8 Structure prediction of PINA using I-TASSER. Source: Lesage et al. (2011)
PINA forms aggregate under acidic and high ionic strength conditions and at low
temperatures, where the TRD plays an important role in the aggregate formation (Le
Bihan et al., 1996). Clifton et al. (2011a) confirmed this observation and has shown that
PINA forms monodisperse prolate ellipsoidal micelles in a solution consisting of 38
PINA molecules and is stable in a wide range of pH and temperatures. Self-assembly of
PINA is suggested to be driven by intermolecular hydrophobic forces between the
tryptophan at the TRD. Although PINB shows more than 50% amino acid similarity
with PINA, no self-assembly into micelles has been reported. This may be due to the
fewer tryptophan residues in the PINB TRD, resulting in weaker intermolecular
hydrophobic interactions (Clifton et al., 2011a). The structure of a PINA section, i.e.,

Chapter 1 Introduction and literature review
- 17 -
the peptide PuroA (FPVTWRWWKWWKG) was determined by NMR (Jing et al.,
2003) (Figure 1.9) and showed that all positively charged residues are located close to
the tryptophan indole rings (Jing et al., 2003). Furthermore, the 3D structure of micelle-
bound PuroA shows that all positively charged residues are placed close to the
tryptophan indole rings, which have a role in cation-interactions and allow PuroA to
penetrate and disrupt bacterial membrane (Chan et al., 2006) (Figure 1.10).
Figure 1.9 NMR solution structure of PuroA. Source: Jing et al. (2003). 13 amino acids corresponding to the tryptophan-rich domain (TRD) of puroindoline a. tryptophan (W), arginine (R) and lysine (K) residues are numbered based on their position in the PuroA peptide.
Figure 1.10 Structure of PuroA in the presence of SDS (sodium dodecyl sulfate) micelles. Source: Chan et al. (2006). The cation–π arrangement between tryptophan and arginine residues on the left side of the peptide are shown; tryptophan residues are highlighted in green, phenylalanine is show red, and lysine and arginine residues are presented in blue colour. The disulphide bridges are shown in yellow stick representation.

Chapter 1 Introduction and literature review
- 18 -
A prediction of the tertiary structure of wild type PINA (represented as ABC) and
mutants of PINA (contain two (ABBC) or three (ABBBC) copies of TRD) proteins was
obtained using the program Protein Homology/analogue Y Recognition Engine v2.0
(Phyre2) (Miao et al., 2012) (Figure 1.11). Furthermore, the secondary structures of the
purified recombinant proteins were estimated by Circular dichroism (CD) spectroscopy
(Miao et al, 2012).
Figure 1.11 The predicted tertiary structures of PINA and mutants of PINA. Source: Miao et al. (2012). a and d: Wild type PINA (ABC); b and e: mutant of PINA (ABBC); c and f: mutant of PINA (ABBBC). TRD(s) are shown in boxes; arrows point to the hydrophobic tryptophan residues (in red).
1.4.1 Heterologous expression of recombinant PIN proteins
Heterologous expression of PINs is an important strategy for scaling up their production
and functional analysis, and has been described by several authors (Capparelli et al.,
2006; Capparelli et al., 2007; Issaly et al., 2001; Miao et al., 2012; Sorrentino et al.,
2009). The yeast Pichia pastoris has been successfully used to express recombinant
PINA protein (Issaly et al., 2001). During fermentation the addition of Triton X-114
increased the production yield of the recombinant PINA by 10-fold to 14 mg/L (of
which 80% was soluble), compared to concentrations of recombinant PINA previously
detected without Triton X-114 (Issaly et al., 2001).
The cysteine-rich PIN proteins fused with an N-terminal His-tag (6 histidine amino
acids) were successfully expressed in Escherichia coli strains BL21 and BL21pLysS as

Chapter 1 Introduction and literature review
- 19 -
18 kDa proteins (Capparelli et al., 2006). Both types of cells expressed His-PINA and
His-PINB in the insoluble fraction, while His-PINs did not refold properly. His-protein
purification using Ni-NTA spin column under denaturing conditions allowed the
proteins to be obtained in purified form. Additionally, both PINs were expressed in E.
coli strain DH5α with a GST-tag (Glutathione-S-Transferase, known to increase the
solubility of the target protein) as 38 kDa proteins and correctly folded. Further, only
recombinant GST-PINs could be efficiently purified, refolded and cleaved from their
tag (Capparelli et al., 2006).
In summary, 1.5 mg recombinant His and GST tagged PINs were obtained from a 1 L
culture (Capparelli et al., 2006). In addition, Capparelli et al. (2007) described the
expression of PINA and PINB in the Origami strain of E. coli, with yields of the
purified proteins at 1.8 and 0.65 mg per litre of culture, respectively, the yield being 2.6
mg per litre of culture for each protein before cleaving the tag. Wild type PINA (ABC)
and mutants of PINA (with two (ABBC) or three (ABBBC) copies of TRD) were
expressed in E. coli strain Rosetta-gami (DE3), with a NusA fusion tag at the N-
terminal to promote the solubility. The strain Rosetta-gami (DE3) has trxB and gor
mutations, which enhance disulphide bond formation in the E. coli cytoplasm and
provides rare codon tRNAs, hence it can be assumed that the recombinant PINs
expressed in it were folded correctly. A low temperature pre-culture and induction
strategy was also used to substantially increase the yield of soluble, functional PINs
(Miao et al., 2012). Alongside the use of classical organism systems such as E. coli and
yeast to express recombinant PIN proteins, molecular farming approaches with plant
system have also been used. The mature sequence of PINs (lacking the signal peptide)
was fused with an N-terminal 3×FLAG tag (DYKDDDDK) and sub-cloned into a plant
expression vector to target both PINs in the apoplast and the PINB in the chloroplast of
Nicotiana tabacum cv. Bright Yellow 2 (BY-2) cells. PINB targeted to the chloroplast
showed a 20 kDa band based on immunoblot analysis using the MαFLAG-M2
monoclonal antibody and yields of 0.35% of the total soluble proteins. Nevertheless, in
stable suspension-cell cultures, no positive signals were observed for PINA and PINB
targeted to the apoplast in either the cell protein extract or the culture medium
(Sorrentino et al., 2009). The reported recombinant PIN proteins are listed in Table 1.2.

Chapter 1 Introduction and literature review
- 20 -
Table 1.2 Reported recombinant PIN proteins Protein Host Result References
PINA and PINB
Escherichia coli (BL21, BL21pLysS and DH5α)
Obtained His tagged and GST tagged PINs, 1.5 mg recombinant protein from 1 l culture
Capparelli et al. (2006)
Escherichia coli (Origami)
Obtained 1.8 mg l−1 PINA and 0.65 mg l−1
PINB yields of purified protein Capparelli et al. (2007)
PINA
Yeast (Pichia pastoris) Obtained 80% of soluble recombinant PINA Issaly et al.
(2001)
Escherichia coli (Rosetta-gami, DE3)
Expressed and purified PINA and two artificial mutants of PINA
Miao et al. (2012)
PINB Tobacco (Nicotiana tabacum) Expressed recombinant PINB; 0.35% of TSP Sorrentino et al.
(2009)
1.4.2 Evidence of transgenic Pin genes altering grain texture
PINs may be synthesised in the aleurone and transported to the endosperm (Gautier et
al., 1994). PIN localisation in wheat indicated that both proteins were co-localised in
the aleurone layer and the starchy endosperm of rehydrated mature seeds, however,
neither PIN could be found in root, leaf, shoot or hypocotyls of seedlings (Capparelli et
al., 2005; Digeon et al., 1999; Dubreil et al., 1998). Additionally, immunofluorescent
localisation study confirmed the presence of both PINs at the starch granule surface
(Feiz et al., 2009c). In transgenic wheat, both Pin genes were expressed in the starch
endosperm cells (Wiley et al., 2007). However, PINs have been found in protein bodies
during endosperm development by Lesage et al. (2011). It is currently hypothesised
that PINs determine wheat hardness by stabilising the amyloplast membrane on the
surface of starch granules during grain desiccation, thereby preventing total breakdown
of the lipids when the wheat kernel ripens resulting in soft genotypes containing more
phospholipids and glycolipids than hard textured genotypes (Feiz et al., 2009b; Kim et
al., 2012b).
Expression of both wild type Pin alleles (Pina-D1a and Pinb-D1a) in rice, under the
control of the maize ubiquitin promoter, resulted in reduced hardness of the transgenic
rice seeds texture and decreased levels of damaged starch granules, as well as the
average particle size during milling (Krishnamurthy and Giroux, 2001). A softer texture
in transgenic Japonica rice was also observed following Pinb-D1a gene expression
(Wada et al., 2010). Two different transgenic rice lines were also used for
histochemical analysis of the endosperm cell and immunodetection of wheat PINA and

Chapter 1 Introduction and literature review
- 21 -
PINB using the Durotest anti-friabilin antibody and the results suggest that they were
localised between starch granules in the rice endosperm cell (Fujiwara et al., 2014). In
transgenic corn, the expression of Pin genes (Pina-D1a and Pinb-D1a), resulted in an
increase in germ size, germ yield and seed oil content (Zhang et al., 2010a).
The expression of wild type Pinb-D1a in the hard wheat cv. Hi-Line resulted in soft
phenotype, increased friabilin yield and decreased damaged starch (Beecher et al.,
2002a). Further, in a similar study by Hogg et al. (2004), the transgenic hard wheat
cultivar ‘Hi-Line’ with wild type Pina-D1a, Pinb-D1a or both showed reduced softness;
however, expression of PINB resulted in softer kernel. Swan et al. (2006a) reported that
the crossing of the transgenic ‘Hi-Line’ with a soft wheat resulted in progenies
expressing different levels of PINs; those expressing PINB were softer and had more
starch-bound PINs compared to those expressing PINA which only had an increase of
starch-bound PINA. This work supports the need for both PINs in their wild form for
the soft endosperm texture and may indicate that PINB has a role in assisting PINA
binding to starch.
The over-expression of foreign Pina has been associated with co-suppression of the
endogenous Pina (silencing of Pina), resulting in significantly harder kernels (Xia et al.,
2008). The silencing of either Pin was found to decrease the expression of the other Pin
genes and increased grain hardness, based on RNA interference (RNAi) studies in
transgenic wheat (Gasparis et al., 2011). The introduction of Pina-D1a and Pinb-D1a
(Gazza et al., 2008) or Pina-D1a (Li et al., 2014) into durum wheat significantly
reduced its hardness value. Pin genes from soft wheat cv. Chinese Spring were
transferred through ph1b-mediated homoeologous recombination into durum wheat, and
these durum lines were stable and could be used in crossing the soft endosperm texture
to other durum wheat cultivars (Morris et al., 2011).

Chapter 1 Introduction and literature review
- 22 -
1.5 Evidence of in vitro antimicrobial activity of PINs
PIN proteins are located in the starchy endosperm and the aleurone cells of wheat seed,
and have a possible biological role as antimicrobial proteins. Therefore, a considerable
number of studies have been published on the in vitro antibacterial and antifungal
activities of PIN proteins (Capparelli et al., 2005; Capparelli et al., 2006; Capparelli et
al., 2007; Dubreil et al., 1998; Jing et al., 2003). However, the biological function of
PINs is not yet clear. PINA and PINB purified from wheat display in vitro antifungal
activity against Alternaria brassicola, Ascochyta pisi, Botrytis cineria, Fusarium
culmorum and Verticillium dahlia, with the antifungal activity of PINB higher than that
of PINA (Dubreil et al., 1998) (Table 1.3). Both wheat PINs show similar antibacterial
activity and it appears that synergy between the two proteins may enhance their
antimicrobial activity (Capparelli et al., 2005). Further, recombinant PINs expressed
and purified from bacterial expression systems (i.e., E. coli) folded similarly to the
native proteins purified from wheat and revealed the same extent of in vitro antibacterial
activity (Capparelli et al., 2006) (Table 1.3). Antimicrobial activity against E. coli (91%
growth inhibition) has been demonstrated for chloroplast targeted PINB in Nicotiana
tabacum cells (Sorrentino et al., 2009). Mutant forms of recombinant PINA that contain
an extra copy of the TRD have shown higher antibacterial activities (70 µg/mL against
E. coli) than the wild type PINA (90 µg/mL against E. coli) (Miao et al., 2012),
providing evidence that the TRD has a role in their antimicrobial activity. Other studies
have also proposed the TRD of PINs to have a role for their antibacterial and antifungal
activities (Evrard et al., 2008; Jing et al., 2003; Phillips et al., 2011). The PIN proteins
also seem to be a promising tool in topical antibiotics for bacterial infections.
Recombinant PINs have been shown to be effective in ectopic treatments for fungal skin
infections caused by Staphylococcus epidermidis without exhibiting any haemolytic
activity or toxicity to mouse macrophage cells in vitro, and ability to kill intracellular S.
epidermidis (Capparelli et al., 2007). Furthermore, Palumbo et al. (2010) showed
potential antibacterial activity against Listeria monocytogenes in a mouse model with
intravenously injected PIN proteins.

Chapter 1 Introduction and literature review
- 23 -
Table 1.3 Reported antibacterial and antifungal activities of PIN proteins
Puroindoline proteins purified from wheat
Protein Organism Antimicrobial activity (µg/mL) Reference
PINA Alternaria brassicicola Ascochyta pisi Fusarium culmorum Verticillium dahliae
100 200 100 100
Dubreil et al. (1998)
Escherichia coli Staphylococcus aureus Agrobacterium tumefaciens Pseudomonas syringae Erwinia carotovora Clavibacter michiganensis
30 30 35 50 50 50
Capparelli et al. (2005)
PINB Alternaria brassicicola Ascochyta pisi Fusarium culmorum Verticillium dahliae
20 70 40 30
Dubreil et al. (1998)
Escherichia coli Staphylococcus aureus Agrobacterium tumefaciens Pseudomonas syringae Erwinia carotovora Clavibacter michiganensis
30 30 35 50 50 50
Capparelli et al. (2005)
Puroindoline proteins expressed in bacterial cells
Protein Organism Antimicrobial activity (µg/mL) Reference
PINA Escherichia coli Staphylococcus aureus
30 30
Capparelli et al. (2006)
Staphylococcus epidermidis 30 Capparelli et al. (2007) Escherichia coli Staphylococcus aureus
90 150
Miao et al. (2012)
PINB Escherichia coli Staphylococcus aureus
30 30
Capparelli et al. (2006)
Staphylococcus epidermidis 30 Capparelli et al. (2007) Puroindoline protein expressed in Nicotiana tabacum Protein Organism Percentage growth inhibition Reference PINB Escherichia coli 91% Sorrentino et al.(2009)
1.5.1 Evidence of in vivo antimicrobial activity of PINs
PINs have been shown to induce a significant increase in plant resistance towards
microbial pathogens by over-expression and can therefore be applied in the preservation
of food products. However, no in vivo work had been reported until 2001, when the
earliest evidence of in vivo antimicrobial activity for PIN expression was reported.
Transgenic rice expressing PINA and/or PINB showed in vivo antimicrobial activity and
higher resistance against two major fungal pathogens in rice, Magnaportha grisea and
Rhizoctonia solani, which are causal agents of rice blast and sheath blight, respectively
(Krishnamurthy et al., 2001).

Chapter 1 Introduction and literature review
- 24 -
The durum wheat varieties ‘Luna’ and ‘Venusia’ which naturally lack Pina and Pinb
genes were transformed with the Pina-D1a gene, under the control of the maize
ubiquitin promoter, and both varieties showed significantly increased resistance to
Puccinia triticina (leaf rust fungus) and an increase in harvest yield (Luo et al., 2008).
Kim et al. (2012a) compared seed fungal resistance by overexpression of PINA, PINB
or both PINs in wheat to near-isogenic lines (NILs) with mutations in PINA or PINB
and found that lines expressing both PINs had reduced Penicillium sp. fungal infection
in seeds and increased germination. Expressing both Pin genes into corn was also
effective in reducing the symptoms of Cochliobolus heterostrophus, the corn southern
leaf blight pathogen, by 42% (Zhang et al., 2011).
PINs can also be expressed successfully in dicots. For example, two apple genotypes
transformed via Agrobacterium tumefaciens with Pinb-D1a under the control of the
cauliflower mosaic virus 35S promoter (CaMV35S) led to PINB yield to 0.24% of the
total soluble proteins. In addition, the expression of Pinb alone or in association with a
natural resistance gene Vf, induced partial resistance to scab, a fungal disease caused by
Venturia inaqualis (Faize et al., 2004). The in vitro antifungal activity of PINB against
V. inaqualis had previously been shown by Chevreau et al. (2001). The reported in vivo
antimicrobial activity of PINs is listed in Table 1.4.
Table 1.4 Reported in vivo antimicrobial activity of PINs
Gene Host Resistance against Disease Reference
Pina and/or Pinb
Transgenic rice Magnaportha grisea
and Rhizoctonia solani
rice blast and
sheath blight
Krishnamurthy et al. (2001)
Transgenic wheat lines Penicillium sp. fungal infection
in seeds Kim et al. (2012a)
Pina and Pinb
Transgenic corn Cochliobolus heterostrophus
southern corn leaf blight Zhang et al. (2011)
Pina Transgenic tetraploid wheat Puccinia triticina leaf rust Luo et al. (2008)
Pinb Transgenic apple lines Venturia inaqualis apple scab Faize et al. (2004)

Chapter 1 Introduction and literature review
- 25 -
1.5.2 Evidence of antimicrobial activity of PIN-based peptides
Several studies have utilised chemically synthesised peptides, which have been
designated to identify the bioactive domain of the PIN proteins. The PuroA
(FPVTWRWWKWWKG-NH2) and PuroB (FPVTWPTKWWKG-NH2) peptides were
designed based on the TRD of PINA and PINB, respectively, to identify the
antimicrobial activity. PuroA showed a bactericidal effect on both Gram positive and
Gram negative bacteria; in contrast, PuroB showed no activity even at the highest
concentration tested (Jing et al., 2003). Wild type and variant forms of synthetic
peptide were tested against bacteria, as well as phyto-pathogenic fungi (Table 1.5).
PuroB showed specific activity against Rhizoctonia sp., with minimum inhibitory
concentrations (MIC) 64 µg/mL, however, PuroA had a much higher antibacterial
activity than PuroB. The number of tryptophan and positively charged residues can be a
possible explanation for these different functions (Jing et al., 2003; Phillips et al., 2011).
Evrard et al. (2008) showed that two tryptophan residues (W41 and W44) in the TRD
were essential for PINA to interact with the membrane of Saccharomyces cerevisiae,
while the lysine residues (K42 and K45) in the TRD of PINB seem to be more
important for yeast membrane interaction.
Table 1.5 Reported antibacterial and antifungal activities of PIN-based peptides
Name Sequence Organism Activity (µg/mL) References
PuroA
FPVTWRWWKWWKG-NH2
Escherichia coli Staphylococcus aureus
14 33
Jing et al. (2003)
Escherichia coli Staphylococcus aureus Collectotrichum graminicola Drechslera brizae Rhizoctonia cerealis Rhizoctonia solani
16 16 250 250 64 32
Phillips et al. (2011)
PuroB FPVTWPTKWWKG-NH2 Rhizoctonia cerealis Rhizoctonia solani
64 64
Phillips et al. (2011)
1.5.3 Possible mode of action for PINs as antimicrobial proteins and peptides
Anti-microbial peptides or proteins (AMPs) identified from prokaryotes and eukaryotes
are usually less than 100 amino acid residues long and often have a high frequency of
positively charged amino acids (Silva et al., 2011). They are strongly cationic (pI: 8.9-
10.7) having a net charge of 2+ to 7+, mainly due to the presence of arginine, lysine and

Chapter 1 Introduction and literature review
- 26 -
histidine residues, heat-stable (100°C, 15 minutes), and additionally, they have may
have 50% hydrophobic amino acids and do not have any lysing/killing effects on
eukaryotic cells (Hancock and Chapple 1999; Li et al., 2012b).
AMPs have been shown to be an important and ancient mechanism of natural resistance,
a rapid and metabolically inexpensive first line of defence against pathogens (Brokaert
et al., 1997; Egorov et al., 2005). The antimicrobial activity of such peptides was
observed to involve ionic interactions between their basic residues and the negatively
charged bacterial membrane constituents, such as phospholipid head groups (Jenssen et
al. 2006; Melo et al., 2009; Shai, 2002). On the other hand, the outermost layer of the
eukaryotic membranes is composed of lipids with no net charge and zwitterionic
headgroups positioned within the inner layers; membrane-binding is controlled by
hydrophobic rather than ionic interactions (Lee et al., 2011a). The simplest models of
microbe permeation by antimicrobial peptides involve the formation of transbilayer
pores or channels, as shown by the models in Figure 1.12 (Wimley, 2010).
Figure 1.12 Commonly cited models for antimicrobial peptide activity. Source: Wimley (2010).

Chapter 1 Introduction and literature review
- 27 -
During the past 15 years, a wide range of AMPs have been shown to play essential roles
in plant defence systems. Their sizes range from 2 to 9 kDa and many of them have a
cysteine-rich structure with 6 or 8 cysteine residues involved in disulphide bridges; e.g.,
thionins, defensins, non-specific lipid transfer proteins (nsLTPs), hevein- and knottin-
like peptides, MBP1, IbAMP and snakins (Broekaert et al., 1997; Benko-Iseppon et al.,
2010; Egorov et al., 2005; García-Olmedo et al., 1998; Nawrot et al., 2014; Odintsova et
al., 2009). Some of the cysteine-rich AMPs, such as defensins, thionins and LTPs, are
observed in wheat plants (Egorov et al., 2005). The hevein-like peptide with a 10
cysteine motif isolated from Triticum kiharae has been shown to have antifungal and
antibacterial activity (Dubovskii et al., 2011; Odintsova et al., 2009).
The wheat puroindoline proteins share properties with other AMPs, e.g., the low
molecular weight and the disulphide bond structure (Chan et al., 2006; Schibli et al.,
2002). Dubreil et al. (1997) showed that PINA is able to interact tightly with both
wheat phospholipids and glycolipids while PINB interacts only with negatively charged
phospholipids. However, the integrity of the highly hydrophobic TRD is required for
interaction with neutral polar lipids. Using fluorescence emission and circular
dichroism (CD) spectroscopy the involvement of the TRD region in lipid binding
confers strong affinity for polar lipids to PINs (Kooijman et al., 1997). The TRD in
PINA is involved in insertion into the lipid biofilm and interacts with the lipid head
group region of anionic phospholipids, mostly due to external hydrophobic residues and
cationic side chains at the TRD (Kooijman et al., 1998). The involvement of tryptophan
and basic residues in the TRD of PINA has been proposed as important for lipid binding
and its insertion into membranes (Kooijman et al., 1997; Le Guerneve et al., 1998).
Tryptophan-rich antimicrobial peptides have shown unique amphipathic properties from
the side chain of tryptophan (Schiffer et al., 1992).
The in vitro antimicrobial properties of cationic peptides based on the TRD of the PINA
and/or PINB proteins (Capparelli et al., 2005; Charnet et al., 2003; Jing et al., 2003;
Phillips et al., 2011; Ramalingam et al., 2012) have been summarized above. PuroA
binds strongly with and penetrates deeper into negatively charged phospholipid head
groups of microbial membranes and disrupts the membrane bilayer structure by forming

Chapter 1 Introduction and literature review
- 28 -
cation interactions between arginine and lysine and the indole side chains of tryptophan
(Jing et al., 2003). Tryptophan and arginine residues play a key role in the antimicrobial
action of peptides such as tritrpticin and indolicidin (Chan et al., 2006). However, it has
been suggested that the position rather than the number of tryptophans is an important
factor (Bi et al., 2013). Membrane interaction studies in yeast have shown that
tryptophan-41 and tryptophan-44 at the TRD of PINA may be essential for interaction
with fungal plasma membranes. However, for PINB, none of the tryptophan residues
were required, whereas the positively charged lysine-42 and lysine-45 were essential for
yeast membrane interaction of PINB (Evrard et al., 2008). In addition, single
substitution at the TRD of PINB, of a glycine to a more polar serine residue (Gly46Ser)
and a tryptophan to a basic arginine residue (Trp44Arg) decreased the selectivity
towards anionic phospholipids (Clifton et al., 2007a; Clifton et al., 2007b), which
supports the hypothesis that tryptophan may influence membrane selectivity. Further,
Phillips et al. (2011) showed the antimicrobial properties of cationic peptides based on
the unique TRD of the PIN proteins. They have shown reduced activity of PINA
peptide upon substitution of tryptophan residues with phenylalanine within the TRD.
For antifungal activity of the PINB-TRD, lysine-45 and tryptophan-39 have indicated
important roles, while tryptophan-44 does not appear to be essential and substitution of
lysin-45 to glutamic acid in Pinb-L abolished any observable antifungal activity
(Phillips et al., 2011). Microbial membrane interaction via the polar lipids and
membranes may be required as a mode of action of the PIN proteins (Blochet et al.
1993). Using electrophysiological experiments on Xenopus oocytes and artificial planar
bilayers, channel formation mechanisms were shown to be important for puroindolines
antimicrobial activity (Charnet et al., 2003). Moreover, the PIN proteins appear to act
synergistically in combating pathogens not only with α-purothionins but also with one
another (Capparelli et al., 2005; Chan et al., 2006). The pore formations by PIN-based
peptides appear by a carpet-like model, following the intracellular mechanisms of
activity (Alfred et al., 2013b). Recently it has also been suggested that PIN-based
peptides may inhibit DNA synthesis (Haney et al., 2013). It is thus clear that further
work is required to determine the mechanism(s) of activity of the PIN proteins in
membrane interactions, relevant to both their effects on texture and any in vivo defence
roles

Chapter 1 Introduction and literature review
- 29 -
1.5.4 Possible interactions between PINs
A considerable number of studies on comparative genetic and protein expression have
suggested some co-operation or interdependence between the two PIN proteins. The
presence of both PINs in their functional form at the surface of starch is necessary for
the soft phenotype (Hogg et al., 2004) and with a single protein present; the result is an
intermediate grain texture (Feiz et al., 2009c; Wanjugi et al., 2007). It is likely that
PINA has a primary role in binding to the starch granule surface and also has a co-
operative role in stabilising the binding of PINB, as well as mediating its binding to
starch granules (Amoroso et al., 2004; Capparelli et al., 2003; Gazza et al., 2005).
Furthermore there is a possible minor role for PINB in assisting the binding of PINA to
the starch granule surface (Bhave and Morris, 2008b). Supportive roles for PINA and
PINB in binding to the starch granules have been hypothesised (Amoroso et al., 2004;
Swan et al., 2006b). Ziemann et al. (2008), using the yeast two-hybrid system, provided
evidence of physical interactions between PINA and PINB, suggesting that both
functional PINA and PINB are essential for soft texture and that interaction between
PINA and PINB may occur in the wheat kernel.
The requirement of both PIN proteins in polar lipid binding was shown in transgenic
wheat (Feiz et al., 2009c). PIN proteins were independent of each other in localising to
the surface of starch granules, but the greater binding to polar lipids was observed only
when both PINs were present (Feiz et al., 2009c). The occurrence of both wild type
PINs may explain the increase in the equilibrium surface pressure at air/water interphase
and the degree of penetration in lipid monolayers (Clifton et al., 2007a). Overall, the
different lipid binding behaviours of PIN proteins provide further insight into the impact
of hydrophobic and cationic amino acid residues on the functional properties of their
antimicrobial activity (Sanders et al., 2013). Based on these reports, the synergism
between wild type PINA and PINB seems important for grain hardness and possibly in
their antimicrobial activity.

Chapter 1 Introduction and literature review
- 30 -
1.6 Plant expression system
Among various expression systems such as bacteria, yeast, mammalian cells and
insects, plants are becoming an attractive system for the production of recombinant
proteins for pharmaceuticals, industrial proteins, monoclonal antibodies and vaccine
antigens due to many advantages including safe usage, rapid scale-up, long-term storage
and lower production cost (Daniell et al., 2001, 2009b; Fischer and Emans, 2000;
Fischer et al., 2003, 2004, 2013; Horn et al., 2004; Howard and Hood, 2005; Klimyuk et
al., 2008; Ma et al., 2003, 2005; Rybicki, 2009; Schmidt, 2004 Thomas et al., 2011). In
addition, plant cells have the ability to perform post-translational modifications (PTM)
of proteins which are typical of eukaryotic organisms (Gomord and Faye, 2004; Mett et
al., 2008; Vitale and Pedrazzini 2005). PTM affects the proteins activities and functions
such as folding, stability, and solubility, and dynamic interactions with other molecules,
the major type of PTM for plant-made recombinant proteins being glycosylation
(Marino, 1991; Stulemeijer and Joosten, 2008; Webster and Thomas, 2012).
A wide variety of promoters have been used for protein expression in plants, including
constitutive, inducible and tissue-specific, native and synthetic promoters (reviewed in
Egelkrout et al., 2012). The most widely used promoter is the cauliflower mosaic virus
35S promoter (CaMV35S), which promotes high-level of gene expression through the
transcriptional terminator of the Agrobacterium nopaline synthase gene (nos)
(Barampuram and Zhang, 2011; Lee et al., 2008). Several molecular events such as
gene transcription, mRNA translation and protein accumulation affect overall success of
protein expression (Table 1.6).

Chapter 1 Introduction and literature review
- 31 -
Table 1.6 Summary of factors affecting gene transcription, mRNA translation and protein accumulation Source: modified from Egelkrout et al. (2012) Factors affecting transcription rate Factors affecting translation rate Promoter selection Post-transcriptional gene silencing Transcription factor binding (TFB) sites Codon usage Upstream regulatory sequences (URS) mRNA stability
Intron-mediated sequences Recruitment and retention of translation-ralated protein
Transcriptional gene silencing (TGS) Factors affecting protein accumulation Pre-mRNA stability Protein stabilisation Transgene trait stacking Protein localisation The impact of the site of integration Protein modification The number of gene copies Cellular milieu
Successful recombinant protein accumulation in plant systems depends on several key
factors including suitable host tissue, sub-cellular location, the nature of the foreign
proteins, their possible effect on the host plant, as well as the post-translational
modifications and the degree of protein purification required, and based on these
factors, there are several options for protein expression in plant systems (Egelkrout et
al., 2012; Schiermeyer et al., 2004). These are discussed below.
1.6.1 Host plants
Protein expression platforms have been developed for a variety of host systems
including seed crops (corn, canola, soybeans, and rice) and leafy crops (tobacco, alfalfa,
lettuce), as well as plant cell cultures (tobacco, carrot, rice), hairy root cultures and
aquatic plants (Lemna minor), in addition to moss and green algae (Wilken and
Nikolov, 2012). One of the major plant systems that is used for the production of
recombinant proteins is tobacco, the advantage being high leaf biomass yields as it can
be cropped several times a year (Basaran and Rodriguez-Cerezo, 2008; Twyman et al.,
2003; Twyman, 2004; Wilken and Nikolov, 2012).
1.6.1.1 Nicotiana tabacum (Tobacco)
The genus Nicotiana belongs to the family Solanaceae, which includes 76 species
originating in North America, South America, Australia and Africa (Chase et al., 2003;
Knapp et al., 2004). Nicotiana tabacum is an amphidiploid/allotetraploid species with
48 chromosomes, generated from hybridisation between N. sylvestris and either N.
tomentosiformis Goodspeed or N. otophpra Grisebach, all of which originate from

Chapter 1 Introduction and literature review
- 32 -
South America (Kitamura et al., 2001; Lewis, 2011). Nicotiana tabacum as a model
plant in research provides many practical advantages for large-scale production of
recombinant proteins, including high biomass yield, low maintenance and high seed
production (Twyman, 2004). It is a non-food, non-feed crop, hence carries a reduced
risk of transgenic material contaminating food chains (Stoger et al., 2005; Twyman et
al., 2003; Twyman, 2004). Tobacco cultivars contain a high amount of nicotine and
other alkaloids; hence, it is necessary to remove these toxic components (Twyman,
2004). Ma et al. (1995, 1998) reported the first successful expression of the secretory
monoclonal antibody in transgenic N. tabacum. The two tobacco cell lines, BY-2
(bright yellow 2) and NT-1 (N. tabacum 1), are readily available for Agrobacterium-
mediated genetic transformation and have been optimised for rapid growth (Doran,
2013).
1.6.1.2 Nicotiana benthamiana
The non-cultivated tobacco species, Nicotiana benthamiana, is a unique Australian
native plant that may have resulted from hybridisation between N. suaveolens and N.
debneyi (Goodspeed, 1954; Goodin et al., 2008). It is an amphidiploid with 38
chromosomes (Goodspeed, 1954) and has been used as a host in the production of
monoclonal antibodies and vaccines. It is a suitable host species for producing
recombinant proteins using viral vectors such as the tobacco mosaic virus (TMV)
(Twyman, 2004; Yang et al., 2004). It can be transformed with high efficiency and
maintained easily, due to its short stature, short regeneration time and high seed
production, and is also useful for transient expression of genes (Goodin et al., 2008).
1.6.2 Transformation method and expression systems
Genetic transformation can be achieved by physical or biological methods. Physical
methods are usually used to transfer non-viral vectors by using a gene gun (biolistics)
and chemical polyethylene glycol (PEG) transformation or electroporation of
protoplasts (Barampuram and Zhang, 2011; Rao et al., 2009). The biological method of
genetic transformation of plant cells by Agrobacterium involves the transfer and
integration of a large tumor-inducing (Ti) or rhizogenic (Ri) plasmid resident in
Agrobacterium into the plant nuclear genome. Although several species of

Chapter 1 Introduction and literature review
- 33 -
Agrobacterium are known, two species are predominantly used for transformation;
Agrobacterium tumefaciens (T-DNA transfer for generation of transgenic plants) and
Agrobacterium rhizogenes (Ri-plasmid transfers necessary DNA to generate transgenic
hairy roots) (Gelvin, 2003; Gleba et al., 2014; Rao et al., 2009).
The production of recombinant proteins in plants, also known as molecular farming, is
classified into two expression systems, i.e., stable or transient. Stable expression
involves integration of foreign DNA into the plant genome and transfer to the next
generations (Sharma et al., 2005). The storage organs such as seeds have been used for
the stable expression of proteins of interest (Scheller and Conrad, 2004). The
chloroplast stable transformation requires more than two months and nuclear stable
transformation that requires several months (Patiño-Rodríguez et al., 2013).
Transient expression systems include Agrobacterium-mediated and viral-based
transformation, wherein the foreign gene is not integrated in the genome is expressed
only for a few days after being introduced into the plant cell/tissue. The strategy relies
on A. tumefaciens, which contains transgenes; the bacteria are infiltrated directly into
the organs of interest by injection or by using a vacuum chamber (agroinfiltration)
(Klimyuk et al., 2014; Leckie and Stewart, 2011; Leuzinger et al., 2013;
Vaghchhipawala et al., 2011). Transient expression is generally used to verify the
activity of the expression constructs and generate small amounts of recombinant protein
for functional analyses (Twyman et al., 2003). Many results have been obtained so far
with viral backbones such as those of the RNA viruses, Tobacco mosaic virus (TMV),
Potato virus X (PVX) and Cowpea mosaic virus (CPMV) (Gleba et al., 2014;
Marillonnet et al., 2005). Transient expression using viral vectors is one of the best and
rapid methods for obtaining a high yield of protein expression (Egelkrout et al., 2012).
1.6.2.1 Transient expression using viral vectors
Transient, viral-based systems are amongst the best available systems and have the
advantage of being a rapid method for large-scale production of proteins such as
antibodies and vaccine antigens (Gleba et al., 2007; Pogue et al., 2002). The viral
vector system is based on two strategies: ‘full virus’ vectors used to express long

Chapter 1 Introduction and literature review
- 34 -
polypeptides (at least 140 amino acids) and ‘deconstructed’ virus vectors which rely on
A. tumefaciens to deliver DNA copies of one or more viral RNA replicons to plant cells
(Gleba et al., 2007; Gleba and Giritch, 2012). One main system of deconstructed virus
vectors is the ‘magnICON®’ from ICON genetics, based on the tobacco mosaic virus
(TMV), but instead of supplying whole RNA or linear DNA, it is divided in different
modules captured between the left and right border of T-plasmid of A. tumefaciens
(Gleba et al., 2004; Gleba et al., 2014; Marillonnet et al., 2004). Three A. tumefaciens
cultures deliver different vector modules containing different viral components,
targeting and gene sequences and these pro vector 5’ and 3’ modules assemble within
the plant cell in the form of functional viral vector (Marillonnet et al., 2004). After
delivery by agroinfiltration, they assembled inside the cell with the help of a site-
specific recombinase to create an RNA replicon to produce the desired protein. The
advantage of this system is that the desired protein can be produced at high yield,
because of viral amplification (Marillonnet et al., 2004). Targeting the proteins into
different subcellular compartments can alter protein stability (Gils et al., 2005) e.g.,
chloroplasts can improve stability (Doran, 2006). The green florescence protein (GFP)
has been expressed in different sub cellular compartments of N. benthamiana using this
system, to produce recombinant protein at up to 80% total soluble protein (Marillonnet
et al., 2004; 2005). A considerable amount of research has been published on the use of
the magnICON® system (Gleba et al., 2004, 2005, 2007; Gils et al., 2005; Giritch et al.,
2006; Huang et al., 2006, 2010; Marillonnet et al., 2004, 2005; Santi et al., 2006, 2008;
Webster et al., 2009; Werner et al., 2011).
1.6.3 Subcellular location in a plant system for protein accumulation
The targeting of foreign proteins to the most appropriate subcellular compartment is
important for protein accumulation, stability, folding and posttranslational modification
(Twyman, 2004). The choice depends on the structural characteristics of the protein
and the use of appropriate signals (Benchabane et al., 2008; Faye et al., 2005). Several
subcellular compartments have been considered including the cytosol, endoplasmic
reticulum (ER), apoplast, vacuole and chloroplast (Benchabane et al., 2008; Ma et al.,
2003).

Chapter 1 Introduction and literature review
- 35 -
1.6.3.1 Cytosol
Recombinant proteins are retained in the cytosol when no targeting signals are used.
Several studies have revealed recombinant proteins that remain stable within the cytosol
(De Jaeger et al., 1999; Michaud et al., 1998; Marusic et al., 2007). However, the
cytosol may not be the best location for several reasons: high levels of protease and
ubiquitin activity, the negative redox potential that may affect the correct folding of
proteins with disulphide bonds, and a lack of important co- and post-translational
modifications, such as glycosylation, which may affect folding, assembly and the
tertiary structural stability (Benchabane et al., 2008; Faye et al., 2005).
1.6.3.2 Endoplasmic Reticulum (ER)
Targeting proteins to the ER can lead to high yields of recombinant proteins in planta
(Conrad and Fiedler, 1998; Wandelt et al., 1992). Recombinant proteins entering the
endoplasmic reticulum (ER) can be retained here by adding (SEK/H/K)DEL retention
sequences to the C-terminal (Benchabane et al., 2008; Michaud et al., 1998; Mainieri et
al., 2004), in most cases a (K/H)DEL sequence (Benchabane et al., 2008; Ma et al.,
2003). The presence of molecular chaperones in the ER can have a role in disulphide
bond formation (Benchabane et al., 2008; Faye et al., 2005; Nuttall et al., 2002).
1.6.3.3 Apoplast
Proteins can be targeted into the apoplast via the secretory pathway (Benchabane et al.,
2008). The plant apoplast includes the cell wall matrix and intercellular spaces and has
a wide range of physiological functions (Witzel et al., 2011). The lack of KDEL in
endosperm-expressed protein resulted in targeting to the apoplast, where mostly the
recombinant protein accumulates in the space under the cell wall (Christou et al., 2004).
1.6.3.4 Vacuole
The vacuole is one of the alternative destinations for recombinant proteins produced in
plants (Benchabane et al., 2008). Additionally, it has several key roles in planta e.g.,
control of cell turgidity, levels of macromolecules, accumulation of toxic secondary
metabolites and storage of high-energy compounds in vegetative tissues (Benchabane et
al., 2008; Twyman, 2004).

Chapter 1 Introduction and literature review
- 36 -
1.6.3.5 Chloroplast
The chloroplast is one of the cellular destinations in a plant expression system, through
the addition of an appropriate targeting signal peptide (Daniell and Dhingra, 2002;
Hyunjong et al., 2006). The recombinant proteins may be moved to the chloroplast.
Chloroplast transformation offers uniform expression rates, a high copy number of the
transformed genes (due to hundreds of plastids in a cell), co-expression of multiple
genes from the same construct, minimal transgene escape in the environment due to the
maternal inheritance of chloroplast DNA and minimal gene silencing (Daniell et al.,
2004). Post-translational modifications such as multimerisation and disulphide bridge
formation can occur in the stroma (Daniell, 2006; Parachin et al., 2012). The
chloroplast can thus be a suitable compartment without needing the cell secretory
pathway or modifications such as glycosylation; however, some endogenous proteases
can affect the stability and accumulation of recombinant proteins (Adam and Clarke,
2002). However, this may not be relevant issue for proteins expressed at very high
yield (Daniell, 2006).
In conclusion, plants can act as ‘biofactories’ for the production of recombinant
proteins, for reducing the production costs due to the low maintenance cost and high
quality of the therapeutic protein (Davies, 2010; Gomord and Faye, 2004; Gleba, 2007;
Ling et al., 2010; Mett et al., 2008; Melnik and Stoger, 2013). The use of plant systems
for production of AMPs is summarised as follows.
1.6.4 Plant systems for heterologous production of antimicrobial peptide
Antimicrobial peptides/proteins (AMPs) are unique biologically active molecules that
comprise defence systems against numerous pathogens such as bacteria, fungi, parasites
and/or viruses. Heterologous expression systems, including bacteria and fungi as host
cells, have been used for production of AMPs with different sizes, folds and
complexities (reviewed in Parachin et al., 2012). Plant systems have also been used as a
suitable source for the production of AMPs and can be directly used for crop
improvement without purifying the peptide or proteins (Desai et al., 2010; Giddings et
al., 2000). The heterologous production of radish Rs-AFP2 defensin in tobacco
protected the transgenic tobacco from Alternaria longipes (Terras et al., 1995), whereas

Chapter 1 Introduction and literature review
- 37 -
the alfalfa anti-fungal peptide (alfAPF) defensin reduced the infection in transgenic
potato plants against the fungal pathogen Verticillium dahliae (Gao et al., 2000).
Several AMPs cannot be produced in microbial systems due to their antimicrobial
activity and complex structures. For example, retrocyclin-101 (RC101) and Protegrin-1
(PG1) can be used as therapeutic agents against bacterial and/or viral infections; both
have not yet been produced in microbial systems, but have been expressed in the
tobacco chloroplast with a yield of around 20% of total soluble protein, and retention of
antiviral activity (Lee et al., 2011b). The large scale production of AMPs can be
applied in numerous industries such as biotechnology, pharmaceuticals, cosmetics and
food. However, the limiting factor is the high production costs of some AMPs due to
their molecular mass and protein folding characteristics. The AMP can aggregate or be
degraded by endopeptidases, which also limits their bioactivity. Choosing the right
system is thus essential for large scale production and functional recombinant AMPs.
Plant-based expression systems are a promising platform for further engineering, and
production of AMPs (Parachin et al., 2012). All AMPs heterologously produced in N.
tabacum are listed in Table 1.7.

Chapter 1 Introduction and literature review
- 38 -
Table 1.7 AMPs heterologous produced in Nicotiana tabacum AMP transformed Source Objective Resistance against References
protegrin-1 (PG-1)
Synthesized peptide
Production peptide and antimicrobial activity
Klebsiella pneumoniae, Staphylococcus aureus, Escherichia coli, Mycobacterium bovis, Candida albicans
Patiño-Rodríguez et al. (2013)
Dermaseptin Lysozyme C4V3
Phyllomedusa sauvagii, Gallus gallus, Synthetic
Production and isolation of the peptide and vaccine
Anti-HIV activity, induction of mammalian immune response
Rubio-Infante et al. (2012)
EtMIC2 Eimeria tenella Peptide production
Eimeria tenella (chicken coccidiosis)
Sathish et al. (2011)
Retrocyclin-101 and Protegrin-1
Artificial AMP based on rhesus monkey (Macaca mulatta) circular minidefensins
Chloroplast genome transformation aiming peptide production
Erwinia carotovora, tobacco mosaic virus
Lee et al. (2011b)
NmDEF02 Nicotiana megalosiphon
Proof of concept for crop improvement
Phytophthora parasitica var. nicotianae, Peronospora hyoscyami f.sp. tabacina, Alternaria solani, Phytophthora infestans
Portieles et al. (2010)
Trichokonins Trichoderma pseudokoningii
Proof of concept for crop improvement
Tobacco mosaic virus Luo et al. (2010)
Pal and Cpl-1 Phages infecting S.pneumoniae
Chloroplast genome transformation aiming peptide production
Streptococcus pneumoniae Oey et al. (2009)
MsrA2 (N-methionine-dermaseptin B1)
Phyllomedusa sauvagei and Phyllomedusa bicolor
Gene stacking for crop improvement
Fusarium solani, Fusarium oxysporum, Alternaria alternata, Botrytis cinerea, Sclerotinia sclerotiorum, Pythium aphanidermatum, Pectobacterium carotovorum
Yevtushenko and Misra, (2007)

Chapter 1 Introduction and literature review
- 39 -
1.7 Summary of the literature and aims of the research
1.7.1 Summary of the above literature
This chapter provided background information that addressed the basics of puroindoline
proteins (PINs) in wheat, one of the most widely used crops worldwide. The
relationship between Pin genes and grain texture was reviewed. The review also
addressed the antimicrobial activity and lipid binding properties of PIN proteins and
PIN-based peptides. However, the in vivo roles for both PIN proteins in wheat seed and
the effect of these hardness mutations in puroindolines on the grain hardness as well as
their antimicrobial activity remain unclear. Several studies have reported the co-
operative or interdependent interaction binding to the surface of starch granules to
confer the soft phenotype, the basis for which is also unclear.
PIN proteins aggregate in the presence of membrane lipids and in the absence of lipids,
these proteins can still aggregate, but no precise information is available on the
relationship between aggregation and interaction with lipids in the absence of three-
dimensional structure of the proteins. The high resolution structure of PINs are yet to
be solved, due to problems with their crystallisation and difficulties in obtaining a stable
non-aggregated solution required for NMR structural characterisation. It is important to
investigate the functionality of the PIN proteins in order to better understanding the
biological functions of this family of proteins for their use in biotechnology and
therapeutic applications. Plant molecular farming approach can be used for expressing
large quantities of proteins or peptide. The characterisation of the two PIN proteins in
expressed in such systems will be invaluable for addressing many of the above stated
issues.

Chapter 1 Introduction and literature review
- 40 -
1.7.2 The aims of the project
The specific aims of this project are as follows:
1. To clone the Pin genes encoding the putative mature PINA and PINB in to the
magnICON® viral vector system and utilise A. tumefaciens infiltrations, to express and
optimize their transient expression for high yield in the leaves of Nicotiana
benthamiana.
2. To test the antibacterial and antifungal activities of recombinant PIN proteins purified
with affinity purification using His-tag and hydrophobic interactions systems.
3. To analyse the protein-protein interactions among the PIN proteins in planta using
the BiFC system based on pNBV vectors and co-expression of the silencing suppressor
protein p19.
4. To test the significance of the tryptophan-rich domain (TRD) of PINA and the
hydrophobic domain (HD) of both PINs in any interactions between the PINs using the
BiFC system.

CHAPTER 2
Materials and methods

Chapter 2 Materials and methods
- 42 -
Equipment and materials
2.1 Equipment
The instruments and major apparatus used in this work are listed in Table 2.1.
Table 2.1 Instruments and apparatus used
Equipment Purpose Manufacturer
MyCyclerTM Conducting the Polymerase Chain Reactions (PCR)
Bio-Rad, California, USA
PowerPacTM HC power supply Nucleic acid and protein gel electrophoresis
Mini-protean® Tetra cell Polyacrylamide gel electrophoresis (PAGE or SDS-PAGE)
Westen blotting Mini-Protein II Trans blot module
Transfer of proteins from gels to membrane
SmartSpec 3000 Spectrophotometer
Determining bacterial concentration (Optical Density or OD)
Gene pluser II Electroporation system for transforming various cell types
NanoDropTM 1000 Spectrophotometer
Quantification of nucleic acid concentration
Thermo Fisher, USA
Finnpipette (0.5-10,5-50,20-200 and 100-1000µL) Dispensing liquid
Microplate reader Absorbance detection for ELISA assays and microbial growth turbidity
Microscope (Axioskop 2 MOT Plus)
Visualizing the bimolecular fluorescence complementation (BiFC) assay results Carl Zeiss,
Germany Microscope camera (AxioCam HR) Imaging the BiFC assay results
SNAP i.d Blocking, washing and antibody incubations for western blot Merck Millipore,
Australia Amicon® ultra 0.5 centrifugal filter (3K device - 3,000 NMWL)
Concentrating dilute protein samples
Spectroline Ultra Violet (UV) transilluminators
Visualisation of green or yellow fluorescent protein (GFP or YFP) expression
Spectroline, USA
Chemidoc system Visualisation of gel images UVP,California, USA
End-over-end mixer Mixing of samples for extractions and purification Ratek, Australia
Plant growth cabinet Growth of wheat plants under controlled conditions
Thermoline, Victoria, Australia
Film processor CP1000 Developer for illumination of antibody-HRP labeled western blot Agfa, Australia
Kodak, BioMax light film For western blotting analysis Sigma, USA

Chapter 2 Materials and methods
- 43 -
2.2 Commercial kits
The commercial kits and reagents used in study are listed in Table 2.2.
Table 2.2 Commercial kits
Kit/material Purpose Manufacturer Biomix™ Red (2×) PCR
Bioline, Australia HyperladderTMI (10000bp-200bp)
Molecular weight standard for agarose gel electrophoresis of DNA samples
Precision Plus Protein Dual Xtra Standards (250 kDa-2 kDa)
Molecular weight sizing on SDS-PAGE gels
Bio-Rad, California, USA
Butyl-Toyopearl Hydrophobic interaction protein purification
Tosoh Corporation, Japan
HiYield™ Gel/PCR Fragments Extraction Kit
Recover or concentrate DNA fragments from agarose gels, PCR or other enzymatic reactions
Real Biotech Corporation (RBC), Taiwan HiYield™ Plasmid Mini Kit Plasmid DNA isolation
HisPur™ Ni-NTA Resin Affinity protein purification Thermo Fisher, USA KOD Hot Start DNA polymerase PCR Novagen, Germany
PlatinumR Pfx DNA polymerase PCR Life Technologies,
Invitrogen, USA SYBR® Safe DNA gel stain Staining of DNA in agarose gels
RNase A RNA digestion during genomic DNA extraction Sigma, USA
Restriction endonucleases DNA digestion NEB, (Genesearch), Australia
Sheep blood Haemolysis assay Amyl Media, Australia
Calf intestinal alkaline phosphatase (CIAP) Dephosphorylation of DNA ends
Promega, Australia Wizard® Genomic DNA Purification Kit Genomic DNA extraction

Chapter 2 Materials and methods
- 44 -
2.3 Primary and secondary antibodies
Table 2.3 Antibodies for western blot analysis
Antibody name Purpose Specific Manufacturer and
Catalog number
Durotest® S* To detect friabilin proteins
Monoclonal antibody Host: mouse
R-Biopharm Rhone Ltd., Glasgow, UK; P10
Anti-His To detect histidine tag containing proteins
Monoclonal antibody Host: mouse
Life Technologies, Invitrogen, USA; 37-2900
Anti-GFP To detect GFP protein
Monoclonal antibody Host: rabbit
Life Technologies, Invitrogen, USA; G10362
Anti-mouse Secondary antibody Polyclonal antibody Host: goat Sigma, USA; A0168
Anti-Rabbit Secondary antibody Polyclonal antibody Host: goat Sigma, USA; A0545
*The Durotest® S has a monoclonal antibody raised to detect ‘friabilin’ protein mixture from bread wheat. The kit is detailed later under western blotting section.
2.4 Commonly used buffers, other solutions and media
The common buffers, solutions and media were prepared according to the instructions
in Sambrook and Russell (2001). These are listed in Table 2.4. All aqueous buffers and
solutions were prepared with MilliQ water (Millipore, Australia) that also autoclaved.
Solutions were sterilised by autoclaving (121°C for 20 minutes), or filter sterilizing
through a 0.22 µm syringe filter. All glassware and appropriate disposable plastic ware
(such as PCR tubes, centrifuge tubes, tips) were also autoclaved before use.
Table 2.4 Composition of general buffers and solutions
Buffer and Solution Composition
Coomassie Blue staining solution 40% methanol, 10% acetic acid, 0.05% Coomassie Blue R-250
Coating buffer 15 mM Na2CO3, 35 mM NaHCO3, (pH 9.6) De-stain solution 50% methanol, 10% acetic acid Infiltration buffer for magnICON® system
10 mM 2-(N-Morpholino)ethanesulfonic acid (MES), 10 mM MgSO4 (pH 5.5)
Infiltration buffer for BiFC system 10 mM MgCl2 and acetosyringone to a final concentration of 200 µM
Isopropyl-β-D-thiogalactopyranoside (IPTG) 0.1 M in sterile MilliQ water
Phosphate-buffered saline (PBS) 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4 (pH 7.4)
PBS containing Tween-20 (PBST) PBS with 0.1% (v/v) Tween 20 Milk-PBS-Tween 20 (MPBST) 0.5% milk powder, PBS, 0.05% Tween 20

Chapter 2 Materials and methods
- 45 -
Table 2.4 Composition of general buffers and solutions (continued)
Buffer and Solution Composition
Native-PAGE loading dye, 2x 125 mM Tris (pH 6.8), 50% glycerol, 0.02% bromophenol blue
SDS-PAGE loading dye, 5x 50 mM Tris-HCl (pH 8.8), 10% sodium dodecyl sulfate (SDS), 30% glycerol, 500 mM ß-mercaptoethanol, 0.02% bromophenol blue
TB buffer 10 mM Hepes, 15 mM CaCl2, 250 mM KCl, (pH 6.7) then add MnCl2 to a final concentration of 55 mM
TE buffer 10 mM Tris, 1 mM EDTA, (pH 8) Tris acetate EDTA (TAE) buffer, 50x 2 M Tris base, 6.5 M EDTA disodium salt (pH 8)
Tris glycine buffer, 10x 25 mM Tris, 192 mM glycine buffer (pH 8.8)
Transfer buffer, 1x 25 mM Tris, 192 mM glycine buffer, 20% (v/v) methanol
Running buffer, 1x 25 mM Tris, 192 mM glycine buffer, 0.1% SDS 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal) 5% (w/v) in dimethylformamide (DMF)
Composition of buffers and solutions for sequencing BDT reaction buffer, 5x 400 mM Tris (pH 9), 10 mM MgCl
2
Sequencing reactions clean-up solution 0.2 mM MgSO4 in 70% ethanol
Media Name Composition Luria Bertani (LB) broth 10 g/L tryptone, 5 g/L yeast extract, 5 g NaCl Luria Bertani (LB) agar LB with 15 g/L agar added (for plates) Luria Bertani (LB) glycerol LB with 30% glycerol Mueller-Hinton Broth (MHB) 21 g/L Potato Dextrose Agar (PDA) 39 g/L
Super Optimal broth (SOB) 0.5% yeast extract, 2% tryptone, 10 mM NaCl, 2.5 mM KCl, 10 mM MgCl2, 10 mM MgSO4
Super Optimal broth with Catabolite repression (SOC) SOB with 20 mM glucose (filter sterilized)
Antibiotic stock Carbenicillin 200 mg/mL in sterile MilliQ water Ampicillin
50 mg/mL in sterile MilliQ water Kanamycin Rifampicin Spectinomycin

Chapter 2 Materials and methods
- 46 -
2.4.1 Protein purification buffers
Various buffers were needed to be prepared as per the protein purification protocol and
systems applied. These are listed in Table 2.5.
Table 2.5 Composition of protein purification buffers
His-tag purification buffer (Native condition) [Thermo Fisher, USA] Equilibration buffer 20 mM sodium phosphate, 300 mM sodium chloride with 10 mM imidazole, pH 7.4 Wash buffer 20 mM sodium phosphate, 300 mM sodium chloride with 25 mM imidazole, pH 7.4 Elution buffer 20 mM sodium phosphate, 300 mM sodium chloride with 250 mM imidazole, pH 7.4 His-tag purification buffer (denaturing conditions) [PropurTM, Denmark] Equilibration buffer 20 mM sodium phosphate, 300 mM sodium chloride , 6 M urea with 10 mM imidazole, pH 7.4 Wash buffer 20 mM sodium phosphate, 300 mM sodium chloride, 6 M urea with 25 mM imidazole, pH 7.4 Elution buffer 20 mM sodium phosphate, 300 mM sodium chloride, 6 M urea with 250 mM imidazole, pH 7.4 Hydrophobic Interaction buffer [Lee et al, 2011b] Extraction buffer Saturated ammonium sulphate, pH 7.8; a final saturation of 60%, 70%, 80% Equilibration buffer 20% ammonium sulphate in 10 mM Tris–HCl, pH 7.8 Elution buffer 10 mM Tris–HCl, pH 7.8
2.5 Microbial strains
2.5.1 Strains used as hosts for gene cloning
Escherichia coli Mach1 strain was obtained from stock in Monash University and used
for general molecular cloning procedures. It was typically used for gene cloning.
Agrobacterium tumefaciens strain GV3101 was also used in transformation of plant
expression system vectors which is resistant to rifampicin due to a chromosomal
mutation (Lee and Gelvin, 2008).
The gift glycerol stock vectors which are detailed below were cloned in E. coli XL1-
Blue and A. tumefaciens strain LBA4404.

Chapter 2 Materials and methods
- 47 -
2.5.2 Strains used for testing of antimicrobial activity of recombinant PINs
The bacterial species Escherichia coli (ATCC 25922) and Staphylococcus aureus
(ATCC 25923) and the fungal species, Collectotrichum graminicola, Drechslera brizae,
Fusarium oxysporum, Rhizoctonia cerealis and Rhizoctonia solani were obtained from
the culture collection at Swinburne University of Technology. The bacterial cultures
were streaked on fresh LB plates and incubated overnight at 37°C and a single colony
was used to inoculate overnight cultures. The fungal cultures were maintained on PDA
plates (Table 2.4).
2.6 Plant material
2.6.1 Propagation of wheat seedlings
The soft common wheat (Triticum aestivum L cv. Rosella) was used for isolation of the
wild type alleles of Puroindoline a (Pina-D1a) and Puroindoline b (Pinb-D1a)
henceforth designated as Pina and Pinb. The seeds, kindly provided by the Australian
Winter Cereals Collection (AWCC, Tamworth, Australia) were soaked in sterile water
overnight then sown on a sterilised mixture of potting soil and vermiculite (2:1). The
seedlings were grown in a plant growth cabinet (Thermoline, Australia) with 16 hours
of light (25ºC) and 70% humidity (Figure 2.1). For DNA extractions, leaf tissue was
harvested from 4 week old plants for immediate use or snap-frozen in liquid nitrogen
and stored at -80ºC.
2.6.2 Propagation of Nicotiana benthamiana seedlings
Nicotiana benthamiana was used for plant transformations. The seeds for the wild type
N. benthamiana in Australia were kindly provided by Dr Diane Webster, Monash
University, Melbourne. The seeds were germinated in jiffy peat pellets. The plantlets
(Figure 2.1) were transferred to sterilised mixture of potting soil and vermiculite (2:1)
and grown in a greenhouse maintained at 25°C with a 16h light/8h dark photoperiod.
Young, fully expanded leaves of 8 week old seedlings were used for infiltration.

Chapter 2 Materials and methods
- 48 -
A B
C D Figure 2.1 Wheat and N. benthamiana seeds and plantlets. A: Wheat seeds (T. aestivum L cv. Rosella); B: Wheat plantlet (3 weeks); C: N. benthamiana seeds; D: N. benthamiana plantlet (3 weeks). 2.7 General molecular methods
2.7.1 Genomic DNA extraction
Genomic DNA (gDNA) was extracted from the leaves of the wheat (T. aestivum L cv.
Rosella) seedlings (grown as above) using the Wizard® Genomic DNA Purification Kit
(Promega, Australia), according to the protocol supplied. Approximately 100 mg of the
leaf tissue was snap-frozen in liquid nitrogen, ground to a powder using a mortar and
pestle, mixed with 600 µL Nuclei Lysis solution and incubated at 65°C for 15 minutes.
Then 3 μL of RNase A (4 mg/mL) was added and the solution incubated at 37°C for 10
minutes. 200 μL Protein Precipitation solution was added and the mixture centrifuged
at 10000×g for 5 minutes. The supernatant was carefully mixed with 600 µL
isopropanol and centrifuged as above for 5 minutes. The DNA pellet was washed in
400 μL of 70% ethanol and re-spun as above. The DNA pellet was air-dried, dissolved
in 70 μL TE buffer and stored at -20°C. Then 5 μL of the DNA solution was run on a
1% agarose gel in TAE buffer to determine quality, any degradation and approximate
size.

Chapter 2 Materials and methods
- 49 -
2.7.2 Plasmid DNA purification
The HiYield™ Plasmid Mini Kit (RBC, Taiwan) and the related instructions were used
for all plasmid preparations. A single colony of a bacterial culture was inoculated into 3
mL LB with relevant antibiotics (carbenicillin) and grown overnight at 37°C
temperature and 220 rpm. The cultures were pelleted for one minute at 200×g. The cell
pellets were resuspended in 200 µL PD1 buffer (Resuspension solution), mixed with
200 µL PD2 buffer (Cell Lysis solution) and incubated at room temperature (RT) for
two minutes. Subsequently, 300 µL PD3 buffer (neutralisation) was added to the lysate
and the solution centrifuged for three minutes at 200×g. The supernatant was
transferred to a spin-column assembly (provided with the kit) and centrifuged at 200×g
for one minute. The flow-through was discarded and the column washed twice with
diluted W1 and Wash buffers. The plasmid DNA was eluted by addition of 30 µL
Elution buffer to the column and centrifugation at 200×g for two minutes. DNA (5 µL)
was electrophoresed to check it quality and the DNA concentrations determined using a
Spectrophotometer. The methodology detailed above was applied to all non-
recombinant or recombinant plasmid preparations from transformants in E. coli Mach1
or E. coli XL-1 Blue.
2.7.3 Spectrophotometric quantification of DNA
Purified genomic DNAs were diluted 1:50 with sterile MilliQ water to determine the
DNA concentrations based on absorbance recorded at 260 nm and 280 nm using the
SmartSpec 3000 Spectrophotometer (Bio-Rad, USA), which also reported the
concentration based on absorbance of 1 at A260=50 μg/mL of double stranded DNA.
The A260/A280 absorbance ratio of 1.8 to 2 was used as an indication of high purity
(Sambrook and Russell, 2001). Purified plasmid DNA concentrations were determined
with 1 µL of sample using NanoDropTM 1000 Spectrophotometer.
2.7.4 Agarose gel electrophoresis
Agarose gel electrophoresis was used for multiple purposes such as qualitative
examination of the isolated gDNA, its approximate quantification, determining the sizes
of PCR products or restriction fragments, and purifying selected bands. Agarose gels
were typically prepared as 1% (w/v) gels in 1×TAE buffer with SYBR® Safe DNA gel

Chapter 2 Materials and methods
- 50 -
stain (0.5 μg/mL; Invitrogen) added to the cooled gel solution. Electrophoresis was
typically performed at 100V for 30-90 minutes. The DNA samples were mixed with a
5×loading dye and loaded onto gels. A molecular weight marker such as the
HyperladderTM
І (10,000-200bp; Bioline) was used for fragment size estimation and for
approximate estimation of DNA concentrations by comparison of the intensity of the
test DNA bands with that of the marker fragments. The gels were viewed on a UV
transilluminator and photographed using a Chemidoc system (UVP, USA).
2.8 The polymerase chain reaction (PCR) for gene amplifications
2.8.1 Design and synthesis of primers
Primers for gene amplifications by the polymerase chain reaction (PCR) were designed
using Netprimer (http://www.premierbiosoft.com/netprimer; last accessed October
2013) with the following criteria: length 18-25 bp (without tag), a 3’ G/C clamp,
percentage of GC content kept to approximately 50%, minimisation of secondary
structures such as dimers, cross-dimers and hairpins and the difference between
annealing temperatures of the forward and reverse primers limited to ±5°C. All primers
were synthesised commercially by Invitrogen (Australia) and supplied as desalted
pellets. The pellets were re-suspended in sterile MilliQ water to a stock concentration
of 2 μg/µL and stored at -20°C. The working concentration in standard PCR was 0.1
μg/µL (approximately 10 µM depending on primer length and sequence). The specific
primers designed for amplifying the Pina and Pinb genes and various protein expression
strategies are detailed in separate sections below.
2.8.2 Design of primers for amplification of full-length Pin genes
Primers Pina-D1-Forward and Pina-D1-Reverse were designed to amplify the entire
coding sequence (start to stop codon) of the wild type (WT) allele Pina-D1a of the Pina
gene (Gautier et al., 1994) (Genbank accession number DQ363911) (Table 2.6).
Likewise, Pinb-D1-Forward and Pinb-D1-Reverse were designed to amplify the entire
coding sequence of the WT allele Pinb-D1a (Gautier et al., 1994) (Genbank accession
number DQ363913).

Chapter 2 Materials and methods
- 51 -
Table 2.6 Primer used for the wild type full length Pina-D1 and Pinb-D1 amplifications Amplicons Primers sequence* Expected product size
Pina-D1 Pina-D1-Forward
5’-ATGAAGGCCCTCTTCCTCA-3’ 447bp (Genbank: DQ363911) Pina-D1-Reverse
5’-TCACCAGTAATAGCCAATAGTG-3’
Pinb-D1 Pinb-D1-Forward
5’-ATGAAGACCTTATTCCTCCTA-3’ 447bp (Genbank: DQ363913) Pinb-D1-Reverse
5’-TCACCAGTAATAGCCACTAGGGAA-3’ *Start and stop codons are shown in bold
2.8.3 Design of primers for directional cloning into magnICON® and BiFC vectors
These are described in chapter-specific methods sections below.
2.8.4 Typical PCR conditions
Routine amplifications such as those of Pina and Pinb genes from gDNA or inserts
from recombinant plasmids, were generally carried out in 25 µL volumes, consisting
12.5 μL of 2×BioMixTM Red (Bioline, Australia), 5-10 ng plasmid DNA or 100 ng of
first-round PCR products as templates, and 0.1 µg of each forward and reverse primer.
Negative control reactions were identical except that the template DNA was omitted.
All amplifications involved an initial denaturation at 94°C for 5 minutes. This was
followed by 35 cycles of denaturation at 94°C (30 seconds), annealing at primer-
specific temperature (30 seconds) generally 5°C below the theoretical annealing
temperatures for both primers, extension at 72°C (30 seconds) and then a final extension
at 72°C for 5 minutes. The extension time was estimated roughly using 2 kb/min as the
rate of Taq polymerase activity at 72°C. The BioMixTM Red contains an inert red dye
that permits easy visualisation and direct loading onto a gel. Five μL aliquots of the
PCR products were examined by gel electrophoresis.
2.8.5 PCR for directional cloning of Pin genes into plant expression vectors
The PCRs for amplifying the mature protein-encoding sections of Pin genes (i.e.,
without the putative signal peptides; Gautier et al., 1994) for directional cloning into
various expression vector systems (described later) were conducted using KOD Hot
Start DNA Polymerase (Novagen, Germany) for increased specificity. For each
reaction, the following was assembled in a 0.5 mL PCR tube: 5 µL 10×PCR buffer for
KOD Hot Start DNA polymerase, 0.2 mM dNTPs and 1 mM MgSO4, 200 ng gDNA or

Chapter 2 Materials and methods
- 52 -
5-10 ng plasmid DNA, 0.5 U/μL of KOD Hot Start DNA Polymerase, each primer to a
final concentration of 0.3 μM and sterile MilliQ water to a final volume 50 µL.
Negative controls (no template) were also included. The polymerase was activated by
heating for 2 minutes at 95°C followed by 35 cycles of denaturation at 95°C (20
seconds), annealing at 60°C (10 seconds) and extension at 72°C (15 seconds). The
results were assessed by agarose gel electrophoresis using 5 µL of each PCR products.
2.8.6 Purification of PCR products
The PCR products (when of a single size) or DNA bands of interest were purified using
HiYield™ gel/PCR fragments extraction kit (RBC, Taiwan). For purification from gels,
the desired band was excised and weighed (maximum 300 mg in one tube). The gel
slice was mixed with 500 µL of DF buffer and incubated at 55°C for 10 minutes to melt
the gel. The mixture was transferred to a spin column assembly (provided with the kit)
and centrifuged at 6,000×g for 30 seconds. The column was then washed with 500 µL
wash buffer. The assembly was centrifuged for additional 30 seconds at 10,000×g to
ensure wash buffer was removed completely. The elution buffer (20 µL) was then added
to the centre of the spin column, incubated for 2 minutes at RT and centrifuged at
10,000×g for 2 minutes. The eluted purified DNA was stored at -20°C. The kit was
also used for direct clean-up of PCR products, prior to restriction digestion and/or
cloning. The reaction (up to 100 µL) was transferred to a microcentrifuge tube. Five
volumes of DF buffer were added and mixed, followed by the steps as above. The
eluted DNAs were quantified by gel electrophoresis and/or NanoDropTM 1000
spectrophotometer.
2.9 Cloning and DNA sequencing
2.9.1 Restriction enzyme digestions of PCR products and vectors
A restriction digest typically contained 100-200 ng DNA substrate, 1 μL of a restriction
enzyme (typically 20 U/μL; New England Biolabs), 10 μL of the appropriate NEB
buffer, 1 μL BSA (10 mg/mL; New England Biolabs) and sterile MilliQ water. For in
planta expression of putative mature PIN proteins, the PCR products of full-length Pina
and Pinb genes were re-amplified with primers containing added restriction sites (and
additional tags) for directional cloning into various expression vectors (detailed in

Chapter 2 Materials and methods
- 53 -
sections 2.13.3 and 2.14.4). These second-round PCR products and the corresponding
vectors were double-digested with two appropriate enzymes together to enable their
ligations. All reactions were incubated at 37°C for 2 hours. An aliquot was tested on an
agarose gel to check the success of digestions.
2.9.2 Ligation reactions
Ligations were typically performed in 10 µL volumes containing 1 µL ligase buffer
(10x), 1 µL rapid T4-ligase (NEB), 50 ng digested plasmid DNA vector and 200 ng
insert (e.g., the purified or restriction-digested PCR products). For an efficient ligation,
the amount of plasmid vector should be one third of the insert, in terms of the number of
molecules. The amount of insert was calculated as per Sambrook and Russell (2001):
Insert (ng) = Vector (ng)×Insert (Kb)Vector (Kb)
×molar ratio InsertVector
.
Ligation reactions without insert were used as negative controls. The reactions were
held at 4°C overnight and then transformed into chemically competent E. coli Mach1
cells.
2.9.3 Preparation of chemically competent cells of E. coli Mach1
Chemically competent E. coli Mach1 cells were prepared according to Inoue et al.
(1990) with minor modifications. A single colony from a LB agar plate was inoculated
into 5 mL LB medium and incubated overnight at 37°C on a shaker (200 rpm). This
culture was used to inoculate 250 mL of SOB medium and incubated at 37°C with
shaking until the cell density reached OD600
= 0.5. The culture was cooled on ice for 10
minutes then 40 mL of it was transferred into 50 mL tubes, followed by centrifugation
at 2,500×g for 10 minutes at 4°C. The pellets were resuspended in 12 mL ice-cold TB
buffer and incubated on ice for 10 minutes. The washing step was repeated and the
cells were resuspended in 3 mL of TB buffer. This was followed by the addition of
DMSO to a final concentration of 7% (v/v) with gentle mixing and incubation on ice for
10 minutes. The cell suspension was dispensed into 0.1 mL aliquots into prechilled
tubes, which were then snap frozen in liquid nitrogen and stored at -80°C.

Chapter 2 Materials and methods
- 54 -
2.9.4 Transformation of E. coli Mach1 competent cells by heat shock
A tube of the above cells was thawed on ice for 5 minutes and 10 µL of a ligation
reaction was added to it. The suspension was held on ice for 20 minutes, followed by
heat-shock at 42°C for 50 seconds, then holding on ice for 2 minutes. The cells were
transferred into 0.5 mL SOC medium and incubated at 37°C for 1 hour with shaking
(200 rpm) then spread onto LB agar containing 100 µg/mL of carbenicillin and
incubated for 16 hours at 37°C.
2.9.5 Screening of E. coli transformants containing recombinant plasmids by
colony PCR
The recombinant colonies containing inserts in pICH-11599 or pNBV vectors cannot be
distinguished from non-recombinant ones with blue/white selection, as the pICH vector
does not carry the lacZ gene for the α-peptide of ß-galactosidase and the lacZ gene is
also not active in pNBV. Hence colony PCR was used, wherein half each of many
individual (all white) colonies from E. coli transformants were picked from the LB
plates and transferred into 25 µL 2x Red PCR mix and the DNA amplified using gene
specific primers (Table 2.7 and 2.11). The results were assessed using gel
electrophoresis. For the colonies which were found positive for inserts, the remaining
half colony was inoculated into 5 mL LB broth with the appropriate antibiotic and
grown overnight at 37°C with shaking (200 rpm) and used for plasmid purification
(Section 2.7.2).
2.9.6 Preparation of electro-competent A. tumefaciens cells
Agrobacterium strain GV3101 cells were prepared as per Mattanovich et al. (1989) with
minor modifications. The stock culture was streaked onto a non-selective LB agar agar
plate and grown for 1.5 days at 28°C with shaking (200 rpm) in 10 mL LB media
containing 50 µg/mL rifampicin. One mL of this culture was used to inoculate 100 mL
LB medium containing 50 µg/mL rifampicin and incubated at 28°C with shaking until
the cell density reached OD600=0.6. The culture was chilled before centrifugation of 25
mL of it at 2000×g for 20 minutes at 4°C. The supernatant was discarded and the cell
pellet resuspended in 25 mL of ice cold 1 mM HEPES buffer (pH 7) and centrifuged.
The cells were resuspended gently into 2 mL ice-cold glycerol (10%) and then a further

Chapter 2 Materials and methods
- 55 -
10 mL glycerol (10%). The cells were pelleted again and resuspended in 300 µL of
glycerol (10%) with gentle mixing then aliquoted into 45 µL batches in pre-chilled
tubes, which were stored at -80°C.
2.9.7 Electroporation of electro-competent A. tumefaciens cells
Electroporation was performed in 2 mm electroporation cuvettes using 1 µL of (non-
recombinant or recombinant) plasmid DNA and 45 µL electro-competent
Agrobacterium cells prepared as above. After mixing above, the cuvettes were
incubated on ice for 5 minutes, then put into the electro holder of Gene Pulser (Bio-Rad,
USA) and pulsed at 2.5kv (kilovolts), 200Ω (ohm), 25μF (Microfarad), 5.7msec
(millisecond). One mL LB was immediately added into the cuvette. Cells were
transferred into a culture tube and shaken at 28°C for 60 minutes. All non-recombinant
magnICON® modules (extracted earlier from their E. coli XL1 Blue hosts) and all
recombinant pICH11599 vectors were electroporated individually into A. tumefaciens,
plated on plates with 100 µg/mL carbenicillin plus 50 µg/mL rifampicin selections and
grown at 28°C for two days. Recombinant pMLBART vectors were also electroporated
and grown on 50 µg/mL spectinomycin plus 50 µg/mL rifampicin selections at 28°C for
two days.
2.9.8 Screening of A. tumefaciens transformants containing recombinant plasmids
by colony PCR
Colony PCR was used to identify colonies of A. tumefaciens containing inserts. Half
each of many individual colonies from A. tumefaciens transformants were picked from
the LB plates and transferred into 25 µL 2x Red PCR mix volume and the DNA
amplified using gene specific primers as above. The results were assessed using gel
electrophoresis. The remaining half-colonies of interest were inoculated into 5 mL LB
broth with the appropriate antibiotics and grown overnight at 28°C with shaking (200
rpm) for preparing the glycerol stock (750 µL of bacterial culture plus 750 µL LB
glycerol) and processing for infiltration to the plant.

Chapter 2 Materials and methods
- 56 -
2.9.9 DNA sequencing
All plasmids from E. coli Mach1 were prepared using HiYield™ plasmid mini kit
(RBC, Taiwan), as mentioned earlier. These were sequenced using the BigDye
Terminator (BDT) version 3.1 (Applied Biosystems, USA) according to the instructions
of the Australian Genome Research Facility (AGRF, Melbourne, Australia). The
reactions contained 1 μL BDT reagent, 2 μL 5×BDT buffer, 200-500 ng plasmids DNA
template and 3.2 pmol sequencing primer (Pin gene based primer), made up to 10 µL
with sterile MilliQ water. The cycling conditions were as follows: initial denaturation at
96°C for 2 minutes then 35 cycles of denaturation at 96°C for 10 seconds, primer
annealing at 50°C for 5 seconds and extension at 60°C for 4 minutes. Following this,
the reactions were cleaned up by the addition of 75 μL of 0.2 mM MgSO4
ethanol
solution, incubated at RT for 15 minutes and centrifuged at 200×g for 20 minutes. The
pelleted DNA was washed with 100 μL 70% ethanol followed by centrifugation using
the same conditions. The pellets were air-dried and resuspended in 20 μL sterile MilliQ
water. The solution was submitted to the AGRF (Melbourne) for capillary separation
using an AB3730x/DNA Analyser. The resulting chromatograms were inspected
visually using BioEdit (http://www.mbio.ncsu.edu/bioedit/bioedit.html) for quality
checking. Generally, sequencing was done in both forward and reverse directions.
2.10 Bioinformatics methods
2.10.1 Sequence alignments
The putative amino acid sequences were translated from the nucleotide sequence data
and aligned using Bioedit v 7.1.11 sequence alignment editor program. Bioedit also
was used for analysis of gene sequences of Pina-D1, Pinb-D1. The alignments of DNA
sequences were performed with homologous sequences found in Genbank
(http://www.ncbi.nlm.nih.gov/genbank).
2.10.2 Plasmid vector maps
Vector maps of pNBV and pMLBART plasmids were created using A plasmid Editor
(ApE) software v 8.5.2.0 (http://biologylabs.utah.edu/jorgensen/wayned/ape; last
accessed March 2014) for sequence annotation, virtual digests and circular schematic
sequence representation.

Chapter 2 Materials and methods
- 57 -
2.10.3 Predictions of the isoelectric point (pI) and molecular weight (Mw) of PIN
proteins
The pI and Mw of the putative mature PINA and PINB sequences were determined
using the ‘compute pI/Mw tool’ at the Expert Protein Analysis System (ExPAsy)
(http://web.expasy.org/compute_pi/).
2.10.4 Prediction of protein subcellular localisation
The subcellular locations of mature PINA and PINB were predicted for plant systems
using WoLF PSORT (http://www.genscript.com/psort/wolf_psort.html), which is an
extension of the PSORT II program for converting protein amino acid sequences into
numerical localisation features; based on sorting signals, amino acid composition and
functional motifs (Horton et al., 2007).
2.10.5 Modeling and structure prediction of proteins
The putative amino acid sequences of wild type PIN proteins were submitted to the
Iterative Threading Assembly Refinement (I-TASSER). Using this method, three
dimensional (3D) atomic models were generated from multiple threading alignments
and iterative structural assembly simulations (Roy et al., 2010; Zhang, 2008). The
structures were visualized using molecular visualisation system and the final structural
models with tryptophan-rich domain (TRD) and hydrophobic domain (HD) produced
using PyMOL (http://www.pymol.org/; last accessed January 2014) (Delano, 2002).
The output of the I-TASSER server included the following: five full length atomic
models (ranked based on cluster density), the images of the predicted models and their
estimated accuracy (including confidence score of all models and prediction of TM-
score and RMSD for the first model), the predicted secondary structures, predicted
solvent accessibility and 10 proteins in PDB (Protein Data Bank) which are structurally
closest to the putative models (Roy et al., 2010). The ‘C-score’ is a confidence score
for estimating the quality of the predicted models by I-TASSER and is calculated based
on the significance of the threading template alignments and the convergence
parameters of the structure assembly simulations and is highly correlated with the TM
(template modelling)-score and RMSD (root-mean-square-deviation). The latter two
are used as standards for measuring similarity between two structures, and are useful in

Chapter 2 Materials and methods
- 58 -
determining the accuracy of structure modelling when the native structure is unknown.
A TM-score > 0.5 indicates a model of correct topology and TM-scores < 0.17 indicates
random similarity (Roy et al., 2010).
2.10.6 Protein-Protein complexes structure prediction
Threading-recombination approach, (http://zhanglab.ccmb.med.umich.edu/COTH/)
(CO-Threader: COTH) (Mukherjee and Zhang, 2011) is based on to fold-recognition
and structural recombination of protein-protein complexes prediction by combining
tertiary structure templates with complex alignments. The sequences are first aligned to
complex templates using a modified dynamic programming algorithm. The monomer
templates are then structurally superposed to the dimer template to generate the final
models. The final protein-protein complex predictions for PINA+PINB and
PINA∆HD+PINB∆HD were produced using PyMOL.
2.11 Biochemical methods for protein analysis
2.11.1 Protein concentration measurements
Protein concentrations were measured according to Bradford (1976) using the Bradford
reagent (Bio-Rad, USA) for microtitre plate applications as described by the supplier.
The Bio-Rad dye reagent concentrate was diluted 1:5 with sterile MilliQ water and
filtered with Whatman filter paper 1. Bovine serum albumin (BSA) was used to
generate the standard curve using the range of 0.1, 0.2, 0.3, 0.4 and 0.5 mg/mL. Protein
samples (total soluble proteins or purified recombinant PIN proteins expressed in N.
benthamiana; described later) were diluted 1:10 and/or 1:50 in PBS. To each well in a
microtiter plate, 10 µL of each of the samples and standards (duplicates of each
dilution) was added, followed by 200 µL of diluted dye regent, and incubated for 10
minutes. The absorbance was measured using a Microplate reader (Thermo Fisher,
USA) at 595 nm wave length. The protein concentration of each unknown sample was
calculated from the standard curve.

Chapter 2 Materials and methods
- 59 -
2.11.2 SDS-PAGE
Sodium dodecyl sulphate (SDS)-polyacrylamide gel electrophoresis (PAGE) was used
to assess protein expression profiles, sizes of the (denatured) expressed PIN proteins
and purity at various stages of purification. Samples were mixed with 5 µL of 5x
protein loading dye and boiled for two minutes. The proteins of interest were separated
using a 15% ReadyGel® Tris-HCl gel (Bio-Rad, USA) in running buffer at 20 mA for 1
to 2 hours. The gel was immersed in Coomassie Blue staining solution with gentle
rocking for one hour then immersed overnight in de-staining solution. A molecular
weight marker (250 kDa to 2 kDa) was used as size standards (Table 2.2).
2.11.3 Western blotting analysis for PIN proteins detection
Western blot analyses were carried out using the Durotest®S kit (R-Biopharm, Rhone
Ltd., http://www.r-biopharm.com/wp-content/uploads/4173/DUROTEST-S-Follow-On-
P10.pdf; last accessed February 2014). The kit has a monoclonal antibody raised
against friabilin and it is highly specific for the PIN proteins, which are the two major
components of friabilin (Greenwell et al., 1992; Rakszegi et al., 2009; Wiley et al.,
2007). The kit is commonly used in the pasta industry for rapid detection of
adulteration of durum wheat (semolina, which lacks PINs) with common wheat flour,
which contains the friabilin (mostly PINs). The kit contains two standards for the test:
(i) 100% durum flour, and (ii) durum flour containing 3% non-durum flour (i.e.,
common wheat flour). The epitope recognised by the Durotest antibody has not been
mapped (Greenwell et al. 1992; Wiley et al., 2007).
The total soluble proteins or purified recombinant PINs expressed in N. benthamiana
(described later) were identified through western blotting as described by Rakszegi et al.
(2009) and Wiley et al. (2007) with modification. Proteins was fractionated by SDS-
PAGE then transferred to polyvinylidene difluoride (PVDF) transfer membrane
(Millipore, Australia) in transfer buffer. The membrane was blocked in 0.5% MPBST
for one hour at 4°C on shaker. The Durotest antibody was used at a dilution of 1 in
1000 in 3 mL 0.3% MPBST and incubated for ten minutes at RT by using SNAP i.d.®
protein detection system (Millipore, Australia). The membrane was washed three times
with PBST, then incubated with the secondary antibody (the anti-mouse horseradish

Chapter 2 Materials and methods
- 60 -
peroxidise (HRP) conjugate produced in goat) at a dilution of 1 in 1000 in 3 mL 0.3%
MPBST for ten minutes at RT by using SNAP id.® system. Following three washes
with PBST, the membrane was incubated in chemiluminescent detection reagents (ECL)
(Millipore, Australia) for five minutes and exposed to a Kodak, BioMax light film
(Sigma, USA) for 30 seconds or 1 minute and developed by AGFA CP1000 processor.
2.11.4 Enzyme-linked immunosorbent assay (ELISA) for approximate quantitation
of recombinant PIN proteins
Indirect ELISA involves two binding processes: binding of a primary antibody to the
antigen of interest coated to the microtitre plate surface, and binding of a conjugated
secondary antibody to the primary antibody, followed by assay of activity of the thus-
activated enzyme linked to the secondary antibody (Figure 2.2). The ELISA protocol
for PIN proteins was based on Zhang et al. (2010a) with modifications. The
concentration of PIN proteins expressed in the total soluble protein (TSP) extracts from
infiltrated N. benthamiana leaves (described later) were quantified approximately using
extract of 3% non-durum wheat (Durotest kit; R-Biopharm Rhone Ltd) with different
dilutions for developing a standard curve. The TSP samples were diluted 1/50, 1/100
and 1/200 in coating buffer. Each well (of a 96-well plate) was coated with 100 μL of
the diluted protein samples and left overnight at 4°C. The plate was washed three times
at RT with a phosphate buffered saline solution containing Tween-20 (PBST), then
blocked with 300 µL PBS containing 1% (w/v) bovine serum albumin (PBS-BSA) and
incubated for two hours at RT. Following three washes with PBST, 100 μL of the
Durotest antibody (diluted 1:1000 in PBS-BSA) was added and the plate incubated
overnight at 4°C. The plate was washed three times at RT with PBST. Then 100 μL
solution containing goat anti-mouse horseradish peroxidase (HRP) conjugated antibody
(Sigma, USA) was added (dilution 1:1000 in PBS-BSA) to the plate. The plate was
incubated at RT for one hour and then wells washed five times with PBST. One
hundred μL TMB (3, 3′, 5, 5′-Tetramethylbenzidine) (Sigma, USA) was added and
plates incubated for 15 minutes at RT. The reactions were stopped by addition of 100
μL H2SO4 (0.18 M) and absorbance was measured at 450 nm using a microplate reader
(Thermo Fisher, USA). The coating buffer and infiltrated leaf without Pin construct
were used as negative controls.

Chapter 2 Materials and methods
- 61 -
Figure 2.2 Diagram of indirect ELISA assay. The antigen (Ag) of interest is coated directly to the assay plate and detection of the antigen performed using enzyme-conjugated (E) to the unlabeled primary and conjugated secondary antibodies.
2.12 Testing of antimicrobial properties of the in planta expressed PINs and a PIN-
based synthetic peptide
2.12.1 Design of synthetic peptide based on PINA
The peptide PuroA (FPVTWRWWKWWKG-NH2) was modeled on the TRD of PINA
and used as control for antimicrobial activity, as it had been tested previously (Jing et
al., 2003; Phillips et al., 2011). The peptide was synthesised at >95% purity by
Biomatik Corporation (Ontario, Canada) by solid-phase method using N-(9-flurenyl)
methoxycarbonyl (Fmoc) chemistry, with C-terminal amidation (NH2). Stock solution
was prepared at 1 mg/mL in 0.01% glacial acetic and stored at -20°C.
2.12.2 Antibacterial activity assay
Escherichia coli (ATCC 25922) and Staphylococcus aureus (ATCC 25923) were used
to test the antibacterial activity of the PIN proteins expressed in N. benthamiana. The
minimum inhibitory concentration (MIC) was determined by the microtitre broth
dilution method (Wiegand et al., 2008) with modifications. Bacterial suspension from
an overnight culture was adjusted in Mueller-Hinton Broth (MHB; Oxoid, USA) to
1×108 CFU/mL
using a spectrophotometer (OD600= 0.08-0.13). The suspension was
diluted in MHB to 5×105
CFU/mL. Recombinant purified PIN, from the elution steps of
the His-tag purification and hydrophobic interaction systems (see later) in
approximately 1 mg/mL stock concentrations (measured by Bradford assays) were
added (25 μL) to the wells of sterile flat-bottomed 96-well plates (Corning, Australia)
Substrate
Ag
E
Primary antibody
Secondary antibody conjugate

Chapter 2 Materials and methods
- 62 -
and a two-fold serial dilution was carried out across each row with a starting
concentration of 250 μg/mL and final concentration of 0.5 μg/mL. The wells were then
inoculated with 75 μL of the bacterial suspension and incubated overnight at 37°C. The
MIC values were determined by visual observation of bacterial growth as well as
measuring absorbance at 595 nm using a microplate reader (Thermo Fisher, USA). All
protein samples were tested in triplicate. The MIC was defined as ‘the lowest
concentration of antimicrobial agent that prevents visible growth of microorganism
under defined conditions’ (Wiegand et al., 2008).
2.12.3 Antifungal activity assay
The antifungal activity of PIN proteins was determined by microbroth assay (Broekaert
et al., 1990) with modifications. Filamentous fungal cultures of Colletotrichum
graminicola, Drechslera brizae, Fusarium oxysporum, Rhizoctonia cerealis and
Rhizoctonia solani were grown in Potato Dextrose Broth (PDB; Difco-BD, USA) for
seven days at 25°C with shaking (200 rpm). Mycelia were vortexed vigorously and the
resulting suspension adjusted with PDB to a concentration of 0.8-5×106 mycelia
fragments/mL (OD600 = 0.15-0.17). The suspension was diluted 1:50 with PDB to a
final concentration 0.4-5×104 mycelia fragments/mL (Espinel-Ingroff and Cantón,
2007). Sterile water (~200 μL) was added to the outermost wells of plates when testing
filamentous fungi, so as to reduce the effects of evaporation from the test sample wells
during the longer incubation times. Recombinant purified PINs, from the elution steps
of His-tag purification and hydrophobic interaction systems in approximately 1 mg/mL
stock concentrations (measured by Bradford assays) in 25 μL volumes were added to
the first empty well and a two-fold dilution was carried out across each row with a
starting protein concentration of 500 μg/mL and a final of 2 μg/mL. Test wells were
subsequently inoculated with 75 μL of the fungal mycelia and incubated at 25°C in the
dark for 5-7 days. Then, MIC values were measured which is defined as the lowest
value to completely inhibit fungal hyphal growth (Espinel-Ingroff and Cantón, 2007).
All protein samples were tested in triplicate.

Chapter 2 Materials and methods
- 63 -
2.12.4 Stability testing of recombinant PIN proteins
Aliquots of the protein solutions (the same as those tested above) were stored at 37°C,
RT (between 20°C-25°C), 4°C, -20°C and -80°C for one week. Activity against E. coli
was then determined as above, each sample being tested in triplicate.
2.12.5 Haemolytic activity assay
The haemolytic activity of recombinant PINs, from the elution step of the His-tag
purification system in approximately 1 mg/mL (measured by Bradford assays) stock
concentrations was tested as described by Dathe et al. (1996) and Phillips et al. (2011).
The sheep red blood cells (RBCs) were isolated from defibrinated blood (Amyl Media,
Australia) by centrifugation of the whole blood at 1200×g at 4°C for 10 minutes. The
pellets were washed three times with PBS (pH 7.4) and resuspended in 2 mL PBS.
Then, 25 μL aliquots of the RBC suspension were mixed with different concentrations
of the purified PIN proteins and or the PuroA peptide (500-16 μg/mL) then incubated
for 1 hour at 37°C with gentle shaking. The samples were centrifuged at 3,000×g for 5
minutes and the absorbance of the supernatant measured at 540 nm. PBS-only (no
haemolysis expected) and 1% Triton X-100 (Sigma, USA) in PBS (100% haemolysis
expected) were included as negative control and positive control respectively.
Haemolytic concentration was defined as the concentration of the protein/peptide
required to haemolyse 50% of the RBCs (HD50).

Chapter 2 Materials and methods
- 64 -
Methods specific to Chapter 3:
2.13 Expression of PIN proteins in N. benthamiana using magnICON® viral vector
and characterisation of their biochemical and antimicrobial properties
2.13.1 Principles of the magnICON® viral vector systems for in planta expression
The magnICON® viral vectors system is based on tobacco mosaic virus (TMV) and
designed for in planta assembly from separate pro-vector modules to produce the
protein. The gene of interest is cloned in a 3’ vector module. Various 5’ modules are
then used for targeting the expressed protein to different parts of the plant cell (cytosol,
chloroplast or apoplast). An integrase module is used for recombination of 3’ and 5’
modules in planta. These modules recombine within the plant cell and the resulting
DNA is transcribed and the recombination sites (integrase) are spliced out, to create an
RNA replicon to produce the desired protein (Marillonnet et al., 2004) (Figure 2.3).
The protein is thus expressed in the appropriate subcellular compartment in the plant
cell (depending on the 5’ module used).
Figure 2.3 Schematic showing the assembly of the three viral vector modules and production of PIN proteins. The 5’ and 3’ modules with integrase in the plant cell allow recombining via the AttP and AttB sites. The intron is then spliced out. Viral replication occurs and translation and PIN protein processing occurs. Source: modified from Marillonnet et al. (2004).
+
5’ module P
3’ module PIN T
Integrase
5’ module P 3’ module PIN T
Recombination transcription
Splicing, export
Nucleus
5’ module 3’ module
Cytoplasm
Plant cell
Viral replication
Translation and protein processing
PIN
AttP AttB

Chapter 2 Materials and methods
- 65 -
AttB
intron
Splicing acceptor
site
KpnI NcoI EcoRI SacI BamHI HindIII
Nsil RB LB
The vectors were obtained from ICON Genetics (Halle, Germany;
http://www.icongenetics.com/html/home.htm; last accessed March 2014). The modules
are ampicillin or carbenicillin resistance in E. coli and come as follows (Figure 2.4 and
2.5).
(i) Two 3’ modules:
- [pICH11599]: 3’ module for cloning the gene of interest at the multi-cloning site
- [pICH7410]: Green Fluorescent Protein (GFP)-expressing 3’ module
Figure 2.4 The 3’ module pICH11599, for cloning in gene of interest at the multi-cloning site.
(ii) Various 5’ modules for targeting the expressed protein to different compartments of
the plant cell, including:
- [pICH15879]: untargeted (cytosol) 5’ module
- [pICH8420]: apoplast-targeting 5’ module
- [pICH12190]: chloroplast-targeting 5’ module
(iii) [pICH14011]: the integrase module for recombination of the 3’ and 5’ modules
within the plant cell
Tnos 3’ntr(cr) Tnos NPT Pnos
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F

Chapter 2 Materials and methods
- 66 -
Figure 2.5 The other magnICON® viral vector modules. Detailed vector maps for A: pICH15879, untargeted (cytosol) 5’ module; B: pICH8420, apoplast-targeting 5’ module; C: pICH12190, chloroplast-targeting 5’ module; D: pICH14011, the integrase module; E: pICH7410, GFP expressing 3’ module.
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F
pICH74107173 bp
Ap R
Kan
GFP
Intron
Trfa region
LB
RB
Pnos NTR
AttB
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (959)
BsaI (3859)
EcoRI (6983)
HindIII (1247)
Kpn I (6999)
PstI (1239)
Pvu II (6432)
XhoI (7010)
Nhe I (6785) Swa I (823)
Sca I (4278)
Nde I (1715)
pICH115996446 bp
Intron
Ap R
Trfa region
Kan
3'NTR
LB
AttB recombination site
NOS promoter
pUC ORI
RLK2 OriV
Nos term
Nos term
BamHI (7)
EcoRI (6434)
PstI (29)
PvuII (5741)
SacI (1)
ApaI (275)
HindIII (31)
NcoI (6426)
NheI (6094)
SalI (19)
ScaI (3587)
pICH842012679 bp
RdRp
AP r
Kan
MP
Intron (5' part)
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
AttP
Calreticulin apoplast targeting presequence
AtAct2Prom
ORI
oriV
Nos term
Nos term
omega
BsaI (2907)
Kpn I (6047)
Nco I (11277)
Pvu II (5480)
XhoI (12482)
Not I (4352)
EcoRV (7528)
pICH1219012758 bp
RdRp
MP
Intron 5' part
Ap R
Trfa region
Kan
LB
Pnos
AttP
Artificial dicots chloroplast targeting presequence
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
BsaI (2894)
EcoRI (12273)
HindIII (9446)
Kpn I (6022)
PstI (10488)
Pvu II (5467)
Sac I (11242)
ApaI (1)
Sph I (7266)
Ava I (8443)
EcoRV (7503)
pICH140118816 bp
AP r
Kan
C31 integrase
Nuclear localisation signal
Hsp81.1 promoter
RK2 TrfA
pUC ori
TDNA-LB
TDNA-RB
ORI
oriV
Nos term
Nos 3'
BamHI (1863)
ClaI (1155)
Kpn I (7925)
Nco I (1096)
PstI (12)
XbaI (6787)
XhoI (1)
Not I (6237) ApaI (2177)
Sph I (2342)
Nhe I (7718)
Sca I (5211)
FseI (1887)
Nde I (2648)
pICH1587913371 bp
MP
RdRp (middle fragment)
RdRP 5'
RdRP 3'
Intron 5'part
intron atbg3
intron atipl3
intron atsug1
intron atbg11
intron atatp2
Intron pet psk7-6-2
Ap R
Trfa region
Kan
LB
AttP
T-DNA RB
AtAct2Prom
pUC ORI
RLK2 OriV
Nos term
Nos term
omega
RdRp exon 10
RdRp exon 11
RdRp exon6
RdRp exon 7
BsaI (4219)
EcoRI (1022)
HindIII (11181)
Kpn I (7347)
Nco I (1)
PstI (12223)
Pvu II (6792)
SacI (13362)
ApaI (1326)
Sph I (8591)
Ava I (9768)
EcoRV (8828)
A B C
ED F

Chapter 2 Materials and methods
- 67 -
The above vector systems were used for the expression of the wheat Pina and Pinb
genes in N. benthamiana to study the localisation of the PIN proteins and purify and
characterise them. For this purpose, all above plasmids were extracted from their
glycerol stock cultures in E. coli XL1 Blue (provided by Dr Diane Webster, Monash
University) using the HiYield™ plasmid mini kit (Section 2.7.2). Four different 3’
expression modules; pICH-PINA-His, pICH-PINB-His, pICH-PINA-His-SEKDEL and
pICH-PINB-His-SEKDEL were then constructed in pICH-11599 such that they
contained the gene sections encoding the putative mature PINA or PINB protein, with a
C-terminal histidine tag (His-tag) or SEKDEL tag. The primer design and cloning
processes for this purpose are detailed below.
2.13.2 Design of primers for directional cloning into the 3’ module pICH11599
Primers were designed based on the open reading frame (ORF) encoding the mature
proteins, i.e., without the 28 and 29 residue signal peptides of PINA and PINB,
respectively (Gautier et al., 1994). For cloning of Pina, the nucleotide sequence
encoding the restriction site for EcoRI was incorporated at the 5’ end of the forward
primer and the sequence for BamHI incorporated at the 5’ end of the reverse primer.
For cloning of Pinb, the restriction site for NcoI was incorporated in the forward
primers and for SacI in the reverse primers. The reverse primer for both genes also
contained a sequence that would encode a C-terminal six-histidine tag (His-tag) in the
sense strand, to allow further protein purification. Further, another reverse primer was
designed for each gene such that it also encoded a further C-terminal SEKDEL (Ser-
Glu-Lys-Asp-Glu-Leu), the highly conserved eukaryotic endoplasmic reticulum (ER)
retention signal (Munro and Pelham, 1987; Wang et al., 2008). A BamHI site was
designed at the 5’ end of both the pICH-PINA-His-SEKDEL and pICH-PINB-His-
SEKDEL primers. A few extra spacer nucleotides were added to the primers before and
after the restriction site to allow proper digestions. The primers are listed in Table 2.7.
The expected PCR product sizes for both Pin genes with the His-tag and His-SEKDEL
tag are shown in Table 2.8.

Chapter 2 Materials and methods
- 68 -
Table 2.7 Primers used in directional cloning into 3’ viral module, pICH11599
Primer name Sequence of primers used* Cloning
sites
Pina-His Forward: 5’-GGTAGAATTCGATGGATGTTGCTGGCGGGGGTG-3’ Reverse: 5’-CCGGGATCCTCAATGGTGATGGTGATGGTGCCAGTAATAGCCAATAGTGCC-3’
EcoRI
BamHI
Pinb-His Forward: 5’-GAACCATGGAAGTTGGCGGAGGAGGTGG-3’ Reverse: 5’-CCGGAGCTCTCAATGGTGATGGTGATGGTGCCAGTAATAGCCACTAGG-3’
NcoI
SacI
Pina-His-
SEKDEL
Forward: 5’-GGTAGAATTCGATGGATGTTGCTGGCGGGGGTG-3’ Reverse: 5’CCGGGATCCTCATAGCTCATCTTTCTCAGAATGGTGATGGTGATGGTGCCAGTA3’
EcoRI
BamHI
Pinb-His-
SEKDEL
Forward: 5’-GAAGAATTCGATGGAAGTTGGCGGAGGAGGTGG-3’ Reverse: 5’CCGGGATCCTCATAGCTCATCTTTCTCAGAATGGTGATGGTGATGGTGCCAGTA3’
EcoRI
BamHI
* Cloning sites to be incorporated for directional cloning are shown in italics and shaded. Nucleotides in bold and shade are sequences encoding the His-tag in the sense strand. Nucleotides in bold and underline are sequences encoding SEKDEL in the sense strand. Nucleotides in italics and wave underline encode start and stop codons. The first residue of mature PINs is boxed.
Table 2.8 Genes cloned with His-tag and His-SEKDEL tag into pICH11599
Clones mature protein section of gene that was cloned into pICH11599 Final product size
pICH-PINA-His PINA ORF from DVAGGG to stop (363 bp) +His-tag (18 bp) 381 bp
pICH- PINA-His-SEKDEL
PINA ORF from DVAGGG to stop (363 bp) +His-tag (18 bp)+SEKDEL(18 bp) 399 bp
pICH-PINB-His PINB ORF from EVGGGG to stop (360 bp) +His-tag (18 bp) 378 bp
pICH-PINB-His-SEKDEL
PINB ORF from EVGGGG to stop (360 bp) +His-tag (18 bp)+SEKDEL (18 bp) 396 bp
2.13.3 Amplifications of Pin genes, restriction enzyme digestions, cloning into the
3’ module pICH11599, and sequencing of clones
The 447 bp full-length genes Pina-D1 and Pinb-D1 were amplified from the gDNA
of T. aestivum cv. Rosella (with primers given in Table 2.6).
These PCR products were then used as templates in second-round PCRs with the
primer combinations given in Table 2.7, to amplify the gene sections encoding the
mature PINs, with the His (and SEKDEL) tag(s) and the appropriate restriction sites.
Both second-round PCR products were purified using HiYield™ gel/PCR fragments
extraction kit (Section 2.8.6).

Chapter 2 Materials and methods
- 69 -
The purified PINA PCR products (PINA-His and PINA-His-SEKDEL) were each
double-digested with EcoRI and BamHI, as detailed in section (2.9.1).
The empty vector pICH11599 was purified from the stock culture in E. coli XL-1
Blue using HiYield™ Plasmid Mini Kit (Section 2.7.2).
An aliquot of pICH11599 was double-digested with EcoRI and BamHI.
The two double-digests of PINA were ligated separately with the EcoRI-BamH1
digested pICH11599.
Likewise, the purified PINB PCR product (PINB-His) and another aliquot of
pICH11599 were each digested with NcoI and SacI and then ligated with each other.
The purified PINB PCR product (PINB-His-SEKDEL) was double-digested with
EcoRI and BamHI and then ligated with the double-digested pICH11599.
The ligations were transformed into chemically competent cells of E. coli Mach1 (as
detailed in Section 2.9.4), and plated on LB agar supplemented with carbenicillin.
Several half-colonies per transformation were subjected to colony-PCR (Section
2.9.5) to identify those containing recombinant plasmids.
The other half-colonies of interet were cultured and plasmids were prepared from
these using HiYield™ plasmid mini kit (Section 2.7.2) and sequenced (Section
2.9.9)
The steps are summarized in Figure 2.6.

Chapter 2 Materials and methods
- 70 -
Figure 2.6 Generation of the Pin constructs in magnICON® viral vectors. A: the open reading frame of Pina and Pinb, encoding the mature proteins, followed by the His-tag sequence, ligated into; B: the magnICON® viral vector 3’ module; C: the open reading frame of Pina and Pinb, encoding the mature proteins, followed by the His-tag and SEKDEL sequence, ligated into magnICON® viral vector 3’ module. 2.13.4 Electroporation of recombinant pICH11599 and other modules into
A. tumefaciens
Following confirmation of correct DNA sequences encoding the mature PIN ORFs with
the C-terminal His or His-SEKDEL tags, the recombinant pICH11599 vectors were
electroporated into A. tumefaciens GV3101 (as described in section 2.9.7) and the
bacteria grown on LB medium containing 100 µg/mL carbenicillin plus 50 µg/mL
rifampicin selection at 28°C for two days. The colonies were screened by colony-PCR
for recombinant plasmids using specific primers (Table 2.7). All non-recombinant
vector modules were also electroporated individually into A. tumefaciens.

Chapter 2 Materials and methods
- 71 -
2.13.5 Agroinfiltration of N. benthamiana using the magnICON® viral vectors
Single colonies of A. tumefaciens cultures containing the different vectors were grown
in LB medium supplemented with carbenicillin (100 µg/mL) and rifampicin (50 µg⁄mL)
overnight at 28°C. Equal volumes (2.5 mL each) of three different cultures (Table 2.9)
(OD600=0.7 each) were mixed together in various combinations of (i) a 5’-module; (ii) a
3’-module; (iii) the integrase module. The mixture was then centrifuged at 600×g for
ten minutes. The bacterial pellet was resuspended in 10 mL of infiltration buffer (Table
2.4) and infiltrated into leaves of 8 week old N. benthamiana plants using a 1 mL
syringe without a needle (Figure 2.7). The injected area was marked with permanent
marker pen for ease of identification. Plants were allowed to grow for a further 8 to 10
days in green house conditions before the leaves were harvested and frozen in liquid
nitrogen then stored at -80°C.
Table 2.9 Tri-partatite infiltration of magnICON® constructs into N. benthamiana
Columns show specific infiltrated constructs. Rows show the Agrobacterium cultures containing different viral vector modules. The colors show the mixture of Agrobacterium cultures for specific infiltrated constructs. GFPc: GFP cytosol targeting; GFPch: GFP chloroplast targeting.
Agrobacterium Culture Infiltrated construct
Integrase Cytosol Chloroplast Apoplast GFP PINA
Or PINB
PINA-SEKDEL
Or PINB-
SEKDEL
Cytosol Chloroplast
Apoplast ER
GFPc GFPch
Negative

Chapter 2 Materials and methods
- 72 -
Figure 2.7 Agroinfiltration of N. benthamiana leaf. Lower surface of leaf was infiltrated with Agrobacterium suspension in infiltration buffer. 2.13.6 GFP expression
The green fluorescent protein (GFP) is 238 amino acid long and has MW of 26.9 kDa.
Shimomura et al. (1962) isolated GFP from the jelly fish Aequorea victoria and
demonstrated that it could exhibit bright green fluorescence when exposed to light in the
UltraViolet (UV) range. The gene for GFP was cloned by Prascher et al. (1992) and the
protein was expressed in E. coli by Chalfie et al. (1994). A 3’ viral vector containing
the GFP gene (pICH7410) was infiltrated into N. benthamiana leaves side by side with
the 5’ module for cytosol or the 5’ module for chloroplast. At 10 days post infiltration
(dpi), GFP expression in leaves was checked under UV light (Spectroline UV
transilluminator).
Protein methods
2.13.7 Extraction of total soluble protein (TSP) from N. benthamiana leaf
Pieces from the infiltrated area of the leaf (100 mg) were collected in pre-weighed two
mL tubes and the tubes weighed again. Stainless steel beads of 5 mm diameter (Qiagen,
USA) and 500 µL of extraction buffer [100 mM Ascorbic Acid, 0.1% Triton X-100,
EDTA-free protease inhibitor cocktail tablets in PBS, pH 7.4] were added to each tube.
Centrifugation at 600×g for ten minutes
Resuspended in 10 mL of infiltration buffer
Infiltrated behind the leaf of N. benthamiana using a 1 mL syringe
3’ module, 5’ module and integrase culture were mixed together
5’ module P 3’ module T
Att P Att B
Integrase

Chapter 2 Materials and methods
- 73 -
Leaf tissues were ground at 28 strokes per second for 1 minute using TissueLyserII
(Qiagen, USA), then centrifuged at 4°C at 10,000×g for 10 minutes. The extract was
filtered through Miracloth (Calbiochem, Australia) and centrifuged at 100×g for 5
minutes at 4°C. The clear supernatant was collected into a clean tube. The samples
were either stored at -80ºC or used directly for western blot analysis. In further
optimisations, different extraction buffers (Table 2.10) were used for TSP extraction
before western blot analyses.
Table 2.10 Protein extraction buffers used in this study
No Extraction buffers References
1 PBS containing 100 mM Ascorbic Acid, 0.1% Triton X-100 or Triton X-114, protease inhibitor cocktail tablet
Webster et al. (unpublished)
2 PBS containing 100 mM Ascorbic Acid, 20 mM EDTA, 0.1% Triton X-100, protease inhibitor cocktail tablet
Webster et al. (2009)
3 50 mM Tris (pH 8.8), 50 mM NaCl, 10 mM EDTA, 1 µg/mL leupeptin, protease inhibitor cocktail tablet
Sorrentino et al. (2009)
4 PBS containing 50 mM Tris (pH 8.8), 50 mM NaCL,
10 mM EDTA, 1 µg/mL leupeptin,100 mM ascorbic acid, 0.1% TritonX-100, protease inhibitor cocktail tablet
Tremblay et al. (2011)
2.13.8 Protein concentration measurements, SDS-PAGE, western blotting for
detection and ELISA for approximate quantitation of PINs
Protein concentrations of the TSP were measured by the Bradford assay (see section
2.11.1). SDS-PAGE was used to assess protein expression, the protein sizes and extent
of purification (Section 2.11.2). The PIN components in the TSP, or the purified
recombinant PINs from various methods described below, were identified through
western blotting (Section 2.11.3). The PINs expressed in the TSPs were quantified
approximately by ELISA (Section 2.11.4).
Methods for further PIN protein purification
2.13.9 His-tag purification under native conditions
The His-tagged recombinant PIN proteins expressed by the magnICON® viral vectors
were purified from the TSP using the immobilized metal affinity chromatography
(IMAC) system (Thermo Fisher, USA) which uses imidazole-containing buffers,
according to the manufacturer’s protocol with some modification. The Ni-NTA resin
was equilibrated with equilibration buffer (Table 2.5). TSP samples (20 mL) were

Chapter 2 Materials and methods
- 74 -
mixed with an equal volume of equilibration buffer, applied to the centrifuged tube with
pre-equilibrated resin and mixed on an end-over-end rotator for 30 minutes. The tube
was centrifuged at 700×g for 5 minutes at 4°C. The resin with the assumed bound His-
tagged protein was washed 3 times using two resin-bed volumes of wash buffer (Table
2.5) and centrifuged at 700×g for 2 minutes at 4°C. The bound proteins were eluted in
one resin-bed volume of elution buffer (Table 2.5) by centrifuging at 700×g for 2
minutes at 4°C. The eluted products were removed carefully and stored at -80°C.
Aliquots (50 µL) were taken at each step of purification for Bradford analysis, SDS-
PAGE and western blot analysis (as above).
2.13.10 His-tag purification under denaturing conditions
Urea [CO(NH2)2] is a powerful protein denaturant, as it disrupts the noncovalent bonds
in the proteins. Urea (6 M) was used for purification of the His-tagged recombinant
PINs, to solubilise and make the proteins more accessible for interactions with the Ni-
NTA resin. The TSP was purified using IMAC system (Nunc, Roskilde, Denmark)
under denaturing conditions with urea. The binding, wash and elution buffers with 6 M
urea and different concentration of imidazole were made up in PBS (pH 7.4) (Table
2.5). The TSP samples (20 mL) were applied to the pre-equilibrated nickel IMAC
column and centrifuged at 150×g for 30 minutes at 4°C. The column was washed 3
times using wash buffer and centrifuged at 500×g for 3 minutes at 4°C. Elution buffer
was added to the column and centrifuged at 500×g for 3 minutes at 4°C. The imidazole
and urea were removed from the final eluted product by dialysis (Section 2.13.12) and
stored at -80°C.
2.13.11 Hydrophobic interaction purification
The hydrophobic interaction purification method was used as described by Skosyrev et
al. (2003) and Lee et al. (2011) with modifications. Saturated ammonium sulphate (pH
7.8) was added to a final concentration of 60%, 70% or 80% to the TSP extract. This
suspension was then mixed vigorously with one-fourth the volume of ethanol (70%) for
2 minutes. The ethanol phase was separated by centrifugation at 2500×g at RT for 5
minutes and collected carefully to avoid disturbance of the interphase. Approximately
500 µL of 5 M sodium chloride (NaCl) solution was added to the ethanol phase and

Chapter 2 Materials and methods
- 75 -
mixed for 2 minutes with one-fourth volume of n-butanol (Sigma, USA). The solution
was centrifuged at 2500×g for 2 minutes and the lower phase containing the fusion
protein was carefully collected. The lower phase solution was adjusted to 20%
ammonium sulphate and loaded directly on the pre-equilibrated column with Butyl-
Toyopearl (Tosoh Corporation; Japan). The Butyl-Toyopearl were equilibrated and
washed with equilibration buffer (Table 2.5). The protein was eluted with elution buffer
(Table 2.5) and stored at -80°C for SDS-PAGE and western blot analysis.
2.13.12 Dialysis
The eluted proteins under native and denaturing conditions were exchanged into PBS
(pH 7.5) to avoid precipitating the protein in imidazole and urea. The proteins purified
under denaturing conditions (in 6 M urea) were dialyzed against buffers containing
decreasing concentrations of the denaturing agent (urea) as follows: PBS, 6 M urea;
PBS, 3 M urea; and PBS, 1 M urea. Samples were dialyzed at 4°C for more than 8
hours in each dialysis buffer. Following dialysis, the Amicon ultra 0.5 centrifugal filter
device (3K-3,000 NMWL) were used to remove the rest of imidazole buffer by
centrifugation at 4,000×g for 15 minutes at 4°C for eluted proteins under denaturing
condition. The Amicon device was used for eluted proteins under native conditions to
remove imidazole buffer and change the buffer into PBS.
2.13.13 Concentrate of dilute protein samples
All diluted purified proteins were concentrated using Amicon ultra 0.5 centrifugal filter
device (3K-3,000 NMWL). Spin cycles were performed at 14,000×g for 500 μL of
sample. To recover the concentrated protein, the device was placed upside down in a
clean microcentrifuge tube and centrifuged at 1,000×g for 2 minutes at 4°C. The
samples were applied and the column centrifuged until sufficient concentration was
achieved.
2.13.14 Sodium phytate precipitation
Sodium phytate precipitation was performed as per Krishnan and Natarajan (2009), to
remove Rubisco from samples purified with the hydrophobic interaction system. PINs
purified with the hydrophobic interaction system were treated with 10 mM CaCl2 and

Chapter 2 Materials and methods
- 76 -
10 mM sodium phytate (phytic acid sodium salt hydrate) for 10 minutes at 37°C, and
then centrifuged at RT for 10 minutes at 10000×g. The supernatant was removed to a
fresh tube for further analysis.
2.13.15 Testing of antimicrobial and haemolytic properties of the expressed PINs
The antibacterial and antifungal properties of the recombinant PINs purified with His-
tag under native and denaturing conditions and the hydrophobic interaction purification
system were tested as detailed above (Section 2.12).
Methods specific to Chapter 4:
2.14 Protein-protein interactions of PIN proteins using BiFC system
2.14.1 Principle of the Bimolecular Fluorescence Complementation (BiFC) system
The BiFC system is used to identify interactions between two proteins. The technique
is based on the use of two candidate proteins that are translationally fused to the coding
region fragments of two non-fluorescent fragments of the yellow fluorescent protein
(YFP); YC (C-terminal half) and YN (N-terminal half). Therefore, in a BiFC assay,
interaction of the candidate proteins leads to functional YFP reconstitution and
fluorescence which can be detected by monitoring fluorescence via fluorescence
microscopy techniques (Hu and Kerppola, 2003; Kerppola, 2006) (Figure 2.8). Initially,
this technique was used to demonstrate protein-protein interactions in living mammalian
cells (Hu et al., 2002; Hu and Kerppola, 2003) and then adapted for the use in plant
systems (Bracha-Drori et al., 2004; Walter et al., 2004).

Chapter 2 Materials and methods
- 77 -
Figure 2.8 Principle of the BiFC assay. X and Y were proteins that fused to N-(YN) or C-(YC) terminal of the fluorescent protein (YFP) fragments. If X and Y are interacting proteins the fluorescent protein will be reconstructed and fluorescent signal detected. But if X and Y are non-interacting proteins, the N-(YN) and C-(YC) terminals will not be fused to reconstruct the fluorescent protein and no signal will be detected.
The BiFC test was applied here to test and visualize the occurrence of any interactions
between PINA and PINB proteins in living plant cells. The pNBV vectors used for this
purpose are described below.
2.14.2 New BiFC Vector (pNBV)
The pNBV (Figure 2.9) were kindly provided by Professor David Smyth and Mr Tezz
Quon (Monash University, Melbourne). The vectors (pNBV) contain the gene for
ampicillin or carbenicillin resistance, a terminator of the Nos gene (NosT) and 35S
promoter of the cauliflower mosaic virus. In each pNBV vector pair, the gene of
interest fused either to the N-terminal 155 amino acid fragment (YN) or to the C-
terminal 86 amino acid fragment (YC) of Yellow Fluorescent Protein (YFP) (Hu et al.,
2002).
YN
YC
Y
YN
YC
X X Y
YN
YC
X Y
YN
YC
Y X
Co-expression
Interaction
Co-expression
Interaction
Fluorescent signal
No signal

Chapter 2 Materials and methods
- 78 -
A B
C D
Figure 2.9 The pNBV (New BiFC Vector) modules. Detailed vector maps for the, A: pNBV gene-YN; B: pNBV gene-YC; C: pNBV YN-gene; D: pNBV YC-gene. YN is the non-fluorescent N-terminal half and YC is the non-fluorescent C-terminal half of YFP. These need to come together for fluorescence to occur (see earlier section). Gene-YN refers to gene fused upstream of YN; YN-gene refers to gene fused downstream of YN. Gene-YC refers to gene fused upstream of YC; YC-gene refers to gene fused downstream of YC.
2.14.3 Design of primers for directional cloning into BiFC vectors (pNBVs)
Primers were designed based on the ORFs encoding mature PINA and PINB proteins
(without the putative signal peptides) for directional cloning into pNBV vectors, in-
frame and upstream or downstream of the YN and YC fragments. For cloning as gene-
YN or gene-YC fusions, the restriction site for XbaI was incorporated at the 5’ end of
the forward primers and the site for XmaI at the 5’ end of reverse primers. For cloning
as YN-gene or YC-gene fusions, the site for EcoRI was incorporated into the forward
primers and for BamHI into the reverse primers. For all forward primers the start codon
was added to Pin sequence. The native stop codon was deleted in the reverse primers
for cloning into two pNBV vectors (gene-YN and gene-YC). The primers listed in
Table 2.11.

Chapter 2 Materials and methods
- 79 -
Table 2.11 Primers used in directional cloning into pNBV vectors
Primer name Sequence of primers used * Cloning sites Pina-YN
and Pina-YC
Forward: 5’- GGTTCTAGAATGGATGTTGCTGGCGGGGGTG -3’ Reverse: 5’- CCGCCCGGGCCAGTAATAGCCAATAGTGCC -3’
XbaI
XmaI
Pinb-YN and
Pinb-YC
Forward: 5’- GAATCTAGAATGGAAGTTGGCGGAGGAGGTGG -3’ Reverse: 5’- CCGCCCGGGCCAGTAATAGCCACTAGG -3’
XbaI
XmaI
YN-Pina and
YC-Pina
Forward: 5’- GGTGAATTCATGGATGTTGCTGGCGGGGGTG -3’ Reverse: 5’- CCGGGATCCTCACCAGTAATAGCCAATAGTGCC -3’
EcoRI
BamHI
YN-Pinb and
YC-Pinb
Forward: 5’- GAAGAATTCATGGAAGTTGGCGGAGGAGGTGG -3’ Reverse: 5’- CCGGGATCCTCACCAGTAATAGCCACTAGG -3’
EcoRI
BamHI
* Cloning sites to be incorporated for directional cloning are shown in italics and shaded. Nucleotides in italics and wave underline encode start and stop codons. The first residue of mature PINs is boxed. 2.14.4 Restriction enzyme digestion of PCR products
The gene sections encoding the putative mature PINA and PINB, cloned previously
into 3’ viral module vector (pICH11599), were amplified with gene-specific primers
with the appropriate restriction sites and start/stop codons (Table 2.11).
An aliquot of the above Pina and Pinb PCR products was subjected to double-
digestion with XbaI and XmaI.
The four vectors pNBV (gene-YC and gene-YN) and pNBV (YC-gene and YN-
gene) were prepared from their stocks in E. coli Mach1.
The pNBV (gene-YN and gene-YC) were double-digested with XbaI and XmaI.
Four separate ligations were set up, as (i) Pina with pNBV gene-YN vector; (ii)
Pina with pNBV gene-YC; (iii) Pinb with pNBV gene-YN; (iv) Pinb with pNBV
gene-YC.
Another aliquot of the Pina and Pinb PCR products was double-digested with
EcoRI and BamHI.
The pNBV (YN-gene and YC-gene) were double-digested with EcoRI and BamHI.
Four more ligations were set up, as (v) Pina with pNBV YN-gene; (vi) Pina with
pNBV YC-gene; (vii) Pinb with pNBV YN-gene; (viii) Pinb with pNBV YC-gene.
All ligations were transformed into chemically competent cells of E. coli Mach1,
and plated on LB agar supplemented with carbenicillin.

Chapter 2 Materials and methods
- 80 -
Several half-colonies per transformation were subjected to colony-PCR to identify
those containing recombinant plasmids.
The half-colonies of interest were cultured and plasmids were prepared from these
using HiYield™ plasmid mini kit and sequenced.
The eight BiFC constructs were sequenced and the correct ligations and ORFs
(pNBV-YN-PINA, pNBV-YC-PINA, pNBV-PINA-YN, pNBV-PINA-YC, pNBV-
YN-PINB, pNBV-YC-PINB, pNBV-PINB-YN and pNBV-PINB-YC) were
confirmed.
2.14.5 Cloning of PIN-YFP fragments into pMLBART vector
The pMLBART vector (Figure 2.10) was kindly provided by Professor David Smyth
and Mr Tezz Quon (Monash University, Melbourne). It is commonly used as a plant
transformation vector for study of protein interactions, by using the NotI restriction site
for cloning and spectinomycin resistance as the selection marker.
Figure 2.10 Plasmid map of pMLBART. Plant transformation vector; NotI site show in red.
All eight BiFC constructs made as above were digested at the NotI site, to release the
section containing the 35S promoter of CaMV, the non-fluorescent N-terminal half of
YFP (YN) or the non-fluorescent C-terminal half of YFP (YC), PINA or PINB,
terminator of the Nos gene (NosT) and LacZ (see Figure 2.9). The inserts were cloned
at the NotI site of pMLBART, the ligation mixtures transformed into E. coli Mach1 and
spread onto LB plates containing 50 μg/mL spectinomycin as well as 0.5 mM IPTG and
pMLBART estimate
9777bp

Chapter 2 Materials and methods
- 81 -
80 μg/mL X-gal for ‘Blue-white screening’, (based on the principle of α-
complementation of the β-galactosidase (LacZ) gene) which were incubated at 37°C for
16 hours. The white colonies (potentially containing recombinant clones) were tested
by colony PCR. The plasmids were prepared from cultures of insert using the
HiYield™ plasmid mini kit, the inserts confirmed by PCR again and glycerol stocks of
clones prepared and stored at -80°C. The pNBV vector without a Pin insert was also
digested at the NotI site, transformed into E. coli Mach1 cell and the plasmid isolated
(to be used as negative control).
2.14.6 Electroporation into A. tumefaciens GV3101
One µL each of the plasmid DNAs of all eight clones and the negative control in the
pMLBART were each electroporated into A. tumefaciens GV3101 (Section 2.9.7)
grown on LB plates supplemented with 50 μg/mL spectinomycin and 50 µg/mL
rifampicin for two days at 28°C. The colonies were screened by PCR for the presence
of the inserted gene.
2.14.7 Use of TBSV-P19 vector
TBSV-P19 is based on co-expression of a viral suppressor protein of the Tomato Bushy
Stunt virus (TBSV-P19) called P19, that is able to prevent post-transcriptional gene
silencing (PTGS); hence the vector allows high levels of transient gene expression
(Voinnet et al., 2003) (Figure 2.11). It encodes kanamycin resistance and was
transformed previously into Agrobacterium strain LB4404 and stored as a glycerol
stock at -80°C (culture donated by Dr Diane Webster, Monash University).
Figure 2.11 Plasmid map of the P19 vector.

Chapter 2 Materials and methods
- 82 -
2.14.8 Agroinfiltration of N. benthamiana using BiFC vectors
The cultures of the eight clones in pMLBART in A. tumefaciens were grown overnight
at 28°C in LB supplemented with 50 μg/mL spectinomycin and 50 µg/mL rifampicin.
Agrobacterium tumefaciens LB4404 containing the P19 vector (see above) was grown
in LB medium supplemented with 50 μg/mL kanamycin and 50 µg/mL rifampicin.
Agrobacterium tumefaciens cultures of various combinations of one pMLBART-YN,
with one pMLBART-YC, and the P19 (all at OD600 of 1) were mixed in a ratio of
1:1:0.5 then the mixture centrifuged at 600×g for 10 minutes at RT. The mixed
bacterial pellet was resuspended in 10 mL of infiltration buffer (Table 2.4) and allowed
to stand for one hour at RT. The suspensions were infiltrated into leaves of 8 week old
N. benthamiana plants using a 1 mL syringe (without a needle) until the entire leaf air
space was filled with the Agrobacterium suspension. Plants were allowed to grow for a
further 3 days under greenhouse conditions before the leaves were harvested and
analysed by fluorescence microscopy.
2.14.9 Fluorescence microscope and imaging
Small sections from the infiltrated area of N. benthamiana leaves were removed and
placed in distilled water on glass slides and covered with cover slips. The lower leaf
epidermis was prepared for higher resolution of the intracellular distribution of any
expressed fluorescence. The samples were examined using ZEISS fluorescence
microscope equipped with AxioCam ICc3 (ZEISS) using a 460-480 nm excitation filter
and a 495-540 nm barrier filter. Images were captured using the AxioVision LE
software (ZEISS) and processed using Adobe Photoshop CS4 software.
2.14.10 Protein methods for detection of BiFC
Total soluble proteins (TSP) were isolated as described earlier (Section 2.13.7) from the
Agrobacterium-infiltrated areas of N. benthamiana leaves potentially expressing the
respective fusion proteins. Protein concentrations were measured by the Bradford
assay. The TSP samples were ultra-filtered and concentrated using an Amicon ultra 0.5
centrifugal filter device (3K) (Millipore, Australia) at 13,000×g and used for western
blots.

Chapter 2 Materials and methods
- 83 -
2.14.11 Native gel electrophoresis and detection of YFP fluorescence
The protocol followed Morell et al. (2007) with minor modifications. The TSP
fractions were electrophoresed at 4°C using a 15% ReadyGel® Tris-HCl gel (Bio-Rad,
USA) without SDS in any buffer. The gel was exposed to UV light using Spectroline
UV transilluminator to detect YFP fluorescence.
The overall strategy for construction of BiFC vectors and infiltration into plant cells is
summarized in Figure 2.12.

Chapter 2 Materials and methods
- 84 -
Figure 2.12 Strategy used to construction of BiFC vectors for investigating potential interactions of PIN proteins.
Putative PINA cloned in BiFC vectors (pNBV)
Putative PINB cloned in BiFC vector (pNBV)
The BiFC cassettes in pNBV (contains: 35S; YN or YC; PINB; NosT; LacZ) were released from pNBV by NotI digestion to cloned into the plant expression vector, pMLBART
Colony PCR and sequencing
Transformation into A. tumefaciens GV3101
Mixed cultures with bacteria containing P19 were infiltrated into N. benthamiana
ICON vectors used as template DNA for PCR
Four pNBV constructs prepared: PINA-YN/PINA-YC/YN-PINA/YC-PINA
Four pNBV constructs prepared: PINB-YN/PINB-YC/YN-PINB/YC-PINB
The BiFC cassettes in pNBV (contains: 35S; YN or YC; PINA; NosT; LacZ) were released from pNBV by NotI digestion to cloned into the plant expression vector, pMLBART
ICON vector (pICH11599) with putative mature PINB
cloned into MCS
ICON vector (pICH11599) with putative mature PINA
cloned into MCS

Chapter 2 Materials and methods
- 85 -
2.15 Identification of protein-protein interaction regions of PINs using BiFC
2.15.1 Gifts of gene construct containing deletions of the tryptophan-rich domain
(TRD) of PINA and the hydrophobic domain (HD) of PINA and PINB
The clones designated PINA∆TRD (deletion of TRD at position 34-46 in the mature
PINA protein; Figure 2.13A), PINA∆HD (deletion of HD at position 75-85 in the
mature PINA protein; Figure 2.13B) and PINB∆HD (deletion of HD at position 75-83
in the mature PINB protein; Figure 2.13C) were constructed previously in the pGBK
yeast 2-hybrid system vectors by Dr Rebecca Alfred using the QuikChange II site-
directed mutagenesis kit (Stratagene, California, USA) to delete the above gene sections
(Alfred, 2013 and Alfred et al., 2014). The primers used for introducing the above
deletions are listed in Appendix I (Table I-1). These clones were kindly provided by Dr
Alfred for the present in planta work.
Figure 2.13 Alignment of putative PINA and PINB. A: The tryptophan-rich domain (TRD) highlighted in pink (residues position 34-46) for PINA; B: The hydrophobic domain (HD) highlighted in blue (residues position 75-85) for PINA; C: The hydrophobic domain (HD) highlighted in blue (residues position 75-83) for PINB.
A
B
C

Chapter 2 Materials and methods
- 86 -
2.15.2 Cloning of PINA∆TRD and PINA∆HD and PINB∆HD into pNBV vectors
The PINA∆HD, PINB∆HD and PINA∆TRD-encoding genes cloned into pGBK were
used as templates in a second-round PCR (with primers detailed in Table 2.11) to enable
cloning into pNBV for interaction studies by BiFC assay.
Second-round PCR products were sequenced and then purified using HiYield™
gel/PCR fragments extraction kit.
The purified PCR products were double-digested with XbaI and XmaI.
The vector pNBV (gene-YC) were double-digested with XbaI and XmaI.
Two ligations were set up, as (i) PINA-∆HD with pNBV gene-YC (ii) PINA-∆TRD
with pNBV gene-YC.
The purified PCR product for PINB∆HD and the vector pNBV (YN-gene and YC-
gene) were double-digested with EcoRI and BamHI.
Two more ligations were set up, as (iii) PINB-∆HD with pNBV YN-gene (iv)
PINB-∆HD with pNBV YC-gene.
The ligations were transformed into E. coli Mach1 and plated on LB agar
supplemented with carbenicillin.
Several colonies per transformation were subjected to colony-PCR to identify those
containing recombinant plasmids.
The plasmids were prepared from cultures of this using HiYield™ plasmid mini kit
and sequenced.
The four PIN mutant BiFC constructs made as above were digested at the NotI site
and cloned into the NotI site of pMLBART. The ligation mixtures were
transformed into E. coli Mach1 and spread onto LB plates containing 50 μg/mL
spectinomycin, 0.5 mM IPTG and 80 μg/mL X-gal for ‘Blue-white screening’.
The white colonies (containing recombinant clones) were tested by colony PCR.
The plasmids were prepared from cultures of this using HiYield™ plasmid mini kit
and the inserts were confirmed by PCR again and glycerol stock of clones were
prepared and stored at -80°C. All further steps were as per the sections 2.14.6 and
2.14.8

Chapter 2 Materials and methods
- 87 -
2.15.3 Cloning of PINA∆TRD and PINA∆HD and PINB∆HD into the 3’ module
pICH11599
The above-made clones in pNBV (encoding PINA∆TRD, PINA∆HD and PINB∆HD)
were used as template in a second-round PCR with gene specific primers with the
appropriate restriction enzyme sites for pICH11599 (Table 2.7). Second-round PCR
products were purified using HiYield™ gel/PCR fragments extraction kit, double-
digested with appropriated restriction sites and ligated with the digested pICH11599
(Figure 2.14). The ligations were transformed into E. coli Mach1 and plated on LB agar
supplemented with carbenicillin. Several colonies per transformation were subjected to
colony-PCR to identify those containing recombinant plasmids. The plasmids were
prepared from cultures of E. coli using HiYield™ plasmid mini kit and sequenced. All
further steps were as per sections 2.13.4 and 2.13.5.
Figure 2.14 Generation of the mutant PIN constructs for magnICON® viral vectors. A: the region of PINA∆HD and PINB∆HD, encoding the PIN proteins without HD, followed by the His-tag sequence, ligated into; B: the magnICON® viral vector 3’ module; C: the region of PINA∆TRD encoding the PIN proteins without TRD ligated into magnICON® viral vector 3’ module.

CHAPTER 3
Expression, purification and antimicrobial activity of PINA and PINB proteins in Nicotiana benthamiana

Chapter 3 Protein expression
- 89 -
3.1 Introduction
Protein expression and purification is required in a number of biochemical
investigations such as protein identifications, sequencing, testing of functionality
including enzymatic activities and three dimensional structure determinations.
Therefore, transgenic plant technology, followed by biochemical and techniques, were
applied to PIN proteins to understand the roles they play in determining grain texture,
protection against pathogens and the mechanisms of action in both aspects. For the past
two decades, industrially and therapeutically important proteins have been successfully
expressed in plant systems. However, low yield and lack of efficient purification
methods are two major challenges for economical production of plant-made
recombinant proteins (Conley et al., 2011). Further biochemical characterisation of the
PIN proteins such as three-dimensional structure by X-ray crystallography is essential to
better understand their properties and functions. PINs could be an alternative to
conventional antibiotics to control plant and human diseases due to their antimicrobial
properties. Despite several investigations, there are no reports of PIN proteins purified
from wheat in high yield. However, a small number of studies have identified the lipid-
binding nature of PIN extracted and purified directly from wheat flour (Clifton et al.,
2007b; Clifton et al., 2008; Clifton et al., 2011b). Recombinant PINs have also been
purified from E. coli based expression systems, with varying degrees of success
(Capparelli et al., 2006; 2007; Miao et al., 2012). In addition, Sorrentino et al. (2009)
showed that PINB can be produced in transgenic tobacco cells, targeted to chloroplast,
with the maximum yield of only 0.35% of the total soluble proteins (TSP). In all these
cases the purpose of producing the recombinant PINs was to assess their antimicrobial
activity (detailed in Chapter 1).
The present work aimed to clone and transiently over-express PINA and PINB in
planta, using expression systems based on plant viral vectors. The magnICON® system
was used to produce C-terminal His-tagged fusion proteins in the cytosol, chloroplast,
apoplast and ER. In addition, various strategies were tested for optimising the
production of recombinant PIN within the TSP fraction, including the time of harvest
and extraction buffers. It was necessary to subsequently purify the proteins to avoid
effects of plant-based alkaloids during antimicrobial assays. Two purification systems

Chapter 3 Protein expression
- 90 -
based on His-tag affinity and hydrophobic interactions, were used. To investigate the
functionality of the in planta expressed PINs, antibacterial and antifungal assays were
then performed.
The results in this chapter are divided into 3 sections: expression, purification and
characterisation of the antimicrobial activity of the PINA and PINB.

Chapter 3 Protein expression
- 91 -
600bp 400bp
400bp 200bp
200bp
3.2 Expression of PINs using magnICON® viral vectors
3.2.1 Cloning PINA and PINB into magnICON® viral vector system
The 447 bp genes encoding the full-length putative PINA and PINB proteins were first
amplified from the gDNA of the soft wheat line Triticum aestivum cv. Rosella using the
primers given in Table 2.6 (results not shown). For directional cloning into the 3’
module of the magnICON® system, i.e., pICH11599, these products were used as
templates in second-round PCR (using primers listed in Table 2.7) to amplify the gene
sections encoding the putative mature PINA and PINB with C-terminal His-tags, or His-
SEKDEL-tags, and appropriate restriction sites. PCR products of ~400 bp obtained for
PINA-His and PINB-His (Figure 3.1) were consistent with sizes of the gene sections
encoding start codon (3 bp), the putative mature PINA (363 bp) or PINB (360 bp), and
the 6×His tag (18 bp) (Table 2.8). PCR products of ~400 bp were also obtained for
mature PINA-His-SEKDEL and mature PINB-His-SEKDEL, consistent with the gene
sections encoding mature PINA (363 bp) or PINB (360 bp) and the added start codon (3
bp) and tags (6×His, 18 bp; and SEKDEL, 18 bp) (data not shown).
Figure 3.1 Second-round PCR of gene sections encoding mature PINs. Lane 1: Negative control (no template); lane 2: PINA-His (mature protein-encoding section of Pina-D1a with His-tag); lane 3: PINB-His (mature protein-encoding section of Pinb-D1a with His-tag); lane M: DNA molecular weight marker, Hyperladder l. As per the primer design (Section 2.13.2), the PINA-His and PINA-His-SEKDEL PCR
products and the vector pICH11599 were double-digested with EcoRI+BamH1, for
ligations. The PINB-His PCR products were double-digested with NcoI+SacI and
PINB-His-SEKDEL PCR products with EcoRI+BamH1, and ligated with similarly-
1 2 3 M

Chapter 3 Protein expression
- 92 -
digested pICH11599. The four ligations were transformed into E. coli Mach1 cells.
Due to lack of blue/white colour-selection (lacZ) on pICH11599, colony PCR was
carried out using PIN primers (Table 2.7) to test for inserts (approximately 400 bp), as
exemplified in Figure 3.2. Plasmids were isolated from the above clones and double-
digested with the appropriate enzyme combination, to also ensure the presence of
correct-sized inserts. The results are exemplified in Figure 3.3.
Figure 3.2 Colony PCR for preliminary selection of clones. Lanes M: marker, Hyperladder 1; lanes 1-4: pICH-PINA-His products; lane 5-8: pICH-PINB-His products; lane 9: negative control (no template); lanes 10-11: pICH-PINA-His-SEKDEL products; lanes 12-13: pICH-PINB-His-SEKDEL.
Figure 3.3 Example of double digests of clones in pICH11599. Lane M: marker, Hyperladder 1; lanes 1, 2: undigested and EcoRI+BamHI digested pICH-PINA-His clone respectively; lanes 3, 4: undigested and NcoI+SacI digested pICH-PINB-His clone respectively; lanes 5, 6: undigested and EcoRI+BamHI digested pICH-PINA-His-SEKDEL clone respectively; lanes 7, 8: undigested and EcoRI+BamHI digested pICH-PINB-His-SEKDEL clone.
M 1 2 3 4 5 6 7 8

Chapter 3 Protein expression
- 93 -
For each construct, four potential clones were sequenced using gene-specific primers
and alignments of the data with the published sequences, Pina-D1a (DQ363911) and
Pinb-D1a (DQ363913) showed high identity with the corresponding sections. The
sequences are provided in Appendix II (Figure II-1 to II-4). The constructs were
designated pICH-PINA-His; pICH-PINB-His; pICH-PINA-His-SEKDEL; pICH-PINB-
His-SEKDEL (Figure 3.4). Each construct was electroporated into A. tumefaciens
GV3101 (Section 2.13.4). Screenings of clones containing inserts were carried out
using colony-PCR as above (data not shown). The various 5’ modules, the 3’ GFP
module and the integrase module (Section 2.13.1) were also transformed individually
into A. tumefaciens.
Figure 3.4 The 3’ module constructs encoding mature PIN with His-tag and His/SEKDEL tags. A: pICH-PINA-His; B: pICH-PINB-His; C: pICH-PINA-His-SEKDEL; D: pICH-PINB-His-SEKDEL.
His Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB EcoRI BamHI
PINA
His PINB Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB NcoI SacI
PINA His
SEKDEL EcoRI
Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB EcoRI BamHI EcoRI
PINB His
SEKDEL EcoRI
Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB EcoRI BamHI
A
B
C
D

Chapter 3 Protein expression
- 94 -
3.2.2 Transient expression of PIN proteins in the viral vector system
The A. tumefaciens transformants were used in combinations of three cultures, one with
a 5’ module (for targeting the expressed protein to different cellular compartments), one
with a 3’ module (empty, or with PIN or GFP inserts), and one with the integrase
module to allow recombination of the former two modules and hence recombinant
protein expression. The cytosol and chloroplast targeting GFP served as positive
control, while cultures of the 5’ cytosol and the integrase modules (and no 3’ module)
served as negative control. The combinations are detailed in section 2.13.5 (Table 2.9).
The cultures were mixed, pelleted, resuspended and infiltrated into the underside of N.
benthamiana leaves for transiently expressing the encoded proteins. Leaves were
harvested from three independent plants for each agro-infiltration, before and after
necrosis developed in that area. The necrosis suggested protein production, as the
negative control (5’+integrase modules only) developed marginal necrosis (Figure 3.5).
A B Figure 3.5 N. benthamiana leaves 10 days post-infiltration (different leaves on one plant). A: agro-infiltration with the 5’ cytosol+integrase modules only (negative control); B: agro-infiltration with 3’ (pICH-PINB-His)+integrase+cytosol modules. Agro-infiltrated areas are shown by arrows and dotted lines.
The total soluble protein (TSP) was extracted (Section 2.13.7) from each infiltration
area, GFP expression tested, and western blot and ELISA applied for PINs. Four
different batches were tested and extracts with the highest amounts of PINA or PINB
were preferentially used for further purification and test activity. The results are
detailed below.

Chapter 3 Protein expression
- 95 -
Chloroplast Cytosol
3.2.3 Expression of Green Fluorescent Protein (GFP) by the viral vector system
GFP is widely used as a reporter for gene expression and localisation (Ward, 2006), as
an in situ tag for fusion proteins (Cremazy et al., 2005; Wang and Hazelrigg, 1994), a
biosensor (Crone et al., 2013; Lalonde et al., 2005) or a probe for protein-protein
interactions (Magliery and Regan, 2006; Ventura, 2011). It is 238 amino acids long
(26.9 kDa) and forms a ß-barrel around one α-helix (Ormö et al., 1996; Yang et al.,
1996). GFP expression (Section 2.13.6) was used as a positive control to ensure the
viral vector-based heterologous expression was working correctly. In conjunction with
the 3’ GFP module and integrase, two separate 5’ modules (for cytosol or chloroplast),
were agro-infiltrated and production of GFP was observed over time, between 10-14
days post infiltration (dpi). Leaves were examined using UV light and GFP protein
showed bright fluorescence (Figure 3.6).
A Chloroplast B Cytosol
Figure 3.6 GFP production using two different targeting modules, in N. benthamiana leaves (10 dpi). A: Agro-infiltrated leaves viewed under natural light; B: GFP production in chloroplast and cytosol viewed under UV. The TSP extracted from infiltrated leaves using 3’ GFP module and different 5’
modules (cytosol or chloroplast) were quantified using Bradford assay. The absorbance
of each sample was read at 595 nm and the TSP concentration determined from the
standard curve (Figure 3.7), constructed using bovine serum albumin (BSA) standards
ranging from 0.1 to 0.5 mg/mL (Section 2.11.1). Table 3.1 shows the concentration of
TSP from leaves infiltrated with chloroplast-targeted GFP, cytosol-targeted GFP and
also negative controls. Approximately 93 µg and 98 µg of the quantified TSP samples
were run on a gel to analyse the GFP expression by SDS-PAGE gel (data not shown)
and western blot. Western blot analysis using anti-GFP antibody (Section 2.3)
confirmed the presence of GFP at approximately 25 kDa, at the expected size for GFP
(Figure 3.8).

Chapter 3 Protein expression
- 96 -
Figure 3.7 Standard curve of BSA concentration for Bradford assay. The standard curve was prepared using BSA in PBS buffer, and absorbance measured at 595 nm. Table 3.1 Quantification of TSP for targeted expression of GFP using Bradford assay
Protein Samples Average of test OD595
Protein calculation
based on standard curve (mg/mL)
Dilution factor
Amount of TSP
(mg/mL)
TSP loaded on SDS (µL)
Protein on gel (µg)
GFP Cytosol 0.698 ± 0.005 0.264 10 2.64 37 97.68 GFP Chloroplast 0.685 ± 0.004 0.252 10 2.52 37 93.24 Negative Cytosol 0.594 ± 0.007 0.168 10 1.68 37 62.16 Negative Chloroplast 0.577 ± 0.005 0.152 10 1.52 37 56.24
Figure 3.8 Western blot analysis to determine GFP expression using anti-GFP antibody. Lane 1: TSP from leaf infiltrated with 5’ chloroplast+integrase modules only (negative control); lane 2: TSP from leaf infiltrated with 5’ chloroplast+integrase+3’ GFP modules; lane 3: void lane (no sample loaded); lane 4: TSP from leaf infiltrated with 5’ cytosol+integrase+3’ GFP modules; lane 5: TSP from leaf infiltrated with 5’ cytosol+integrase modules only (negative control).
y = 1.0865x + 0.4124 R² = 0.9843
00.20.40.60.8
11.2
0 0.1 0.2 0.3 0.4 0.5 0.6Abs
orba
nce
(595
nm
) BSA concentration (mg/mL)
1 2 3 4 5
25kDa
93µg 98µg
37kDa
75kDa
56µg 62µg
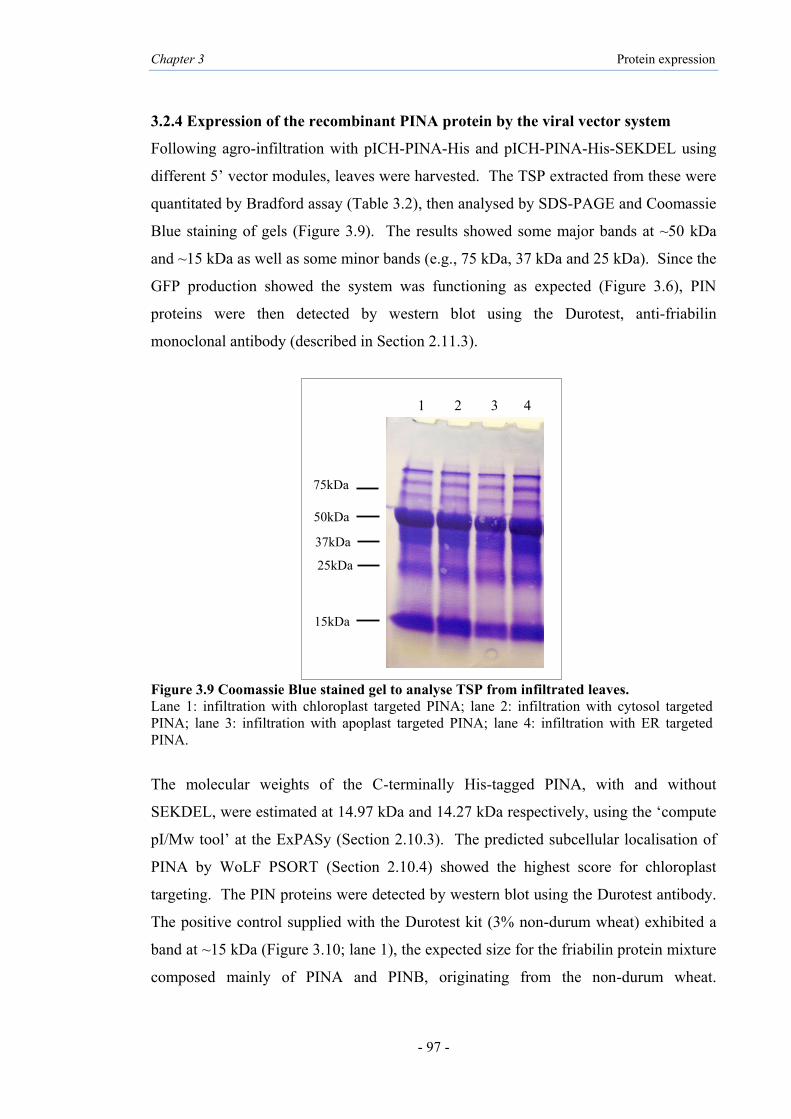
Chapter 3 Protein expression
- 97 -
3.2.4 Expression of the recombinant PINA protein by the viral vector system
Following agro-infiltration with pICH-PINA-His and pICH-PINA-His-SEKDEL using
different 5’ vector modules, leaves were harvested. The TSP extracted from these were
quantitated by Bradford assay (Table 3.2), then analysed by SDS-PAGE and Coomassie
Blue staining of gels (Figure 3.9). The results showed some major bands at ~50 kDa
and ~15 kDa as well as some minor bands (e.g., 75 kDa, 37 kDa and 25 kDa). Since the
GFP production showed the system was functioning as expected (Figure 3.6), PIN
proteins were then detected by western blot using the Durotest, anti-friabilin
monoclonal antibody (described in Section 2.11.3).
Figure 3.9 Coomassie Blue stained gel to analyse TSP from infiltrated leaves. Lane 1: infiltration with chloroplast targeted PINA; lane 2: infiltration with cytosol targeted PINA; lane 3: infiltration with apoplast targeted PINA; lane 4: infiltration with ER targeted PINA.
The molecular weights of the C-terminally His-tagged PINA, with and without
SEKDEL, were estimated at 14.97 kDa and 14.27 kDa respectively, using the ‘compute
pI/Mw tool’ at the ExPASy (Section 2.10.3). The predicted subcellular localisation of
PINA by WoLF PSORT (Section 2.10.4) showed the highest score for chloroplast
targeting. The PIN proteins were detected by western blot using the Durotest antibody.
The positive control supplied with the Durotest kit (3% non-durum wheat) exhibited a
band at ~15 kDa (Figure 3.10; lane 1), the expected size for the friabilin protein mixture
composed mainly of PINA and PINB, originating from the non-durum wheat.
1 2 3 4
75kDa
50kDa
37kDa
15kDa
25kDa

Chapter 3 Protein expression
- 98 -
Interestingly, the recombinant PINA was detected at the unexpected size of ~45 kDa
(between 50 kDa and 37 kDa) when targeted to different parts of the plant cell, i.e.,
chloroplast, cytosol and ER, but not detected in the apoplast, probably due to limited
protein delivery to this compartment. Further, the ~45 kDa bands were observed only
when the TSP samples were not heated prior to the western blot, and could not be
detected when the TSP samples were heated (as discussed later in section 3.2.6).
Figure 3.10 Determination of PINA expression by western blotting of TSP extracts from infiltrated leaves. Lane 1: 3% non-durum wheat as positive control for antibody; lane 2: negative control (5’+ integrase modules only); lane 3: cytosol targeted PINA; lane 4: apoplast targeted PINA; lane 5: chloroplast targeted PINA; lane 6: ER targeted PINA. The quantified TSP samples, as well as the negative controls, were used for enzyme-
linked immunosorbent assay (ELISA) analysis (Section 2.11.4) to measure the yield of
recombinant PINA in N. benthamiana leaves. High-binding ELISA plates were coated
with 0.3 µg TSP/well. The assay tested protein samples in three different dilutions
(1/50, 1/100 and 1/200). The absorbance of each sample was read at 450 nm, and the
PINA concentration determined from the 3% non-durum wheat (supplied in the
Durotest kit) standard curve, ranging from 0.0675-0.54 µg/mL (Figure 3.11).
1 2 3 4 5 6
50kDa
15kDa
75kDa
37kDa

Chapter 3 Protein expression
- 99 -
Figure 3.11 Standard curve of 3% non-durum wheat dilutions for ELISA assay.
Table 3.2 shows the concentration of the recombinant PINA (rPINA) in infiltrated
leaves for the 1/200 dilution test. The maximum yield obtained for PINA was ~0.836%
of TSP, or approximately ~51.7 µg/g of fresh weight using the 5’ module for
chloroplast (Figure 3.12). The PINA yields were ~0.642% TSP or ~40.5 µg/g of fresh
weight using the 5’ module for cytosol and ~0.63% TSP or ~39.7 µg/g of fresh weight
for ER.
Table 3.2 ELISA data analysis for quantification of recombinant PINA (rPINA) protein Sample name OD450
a
rPINA (µg/mL)
rPINA (mg/mL)b
TSP (mg/mL)
FWc (µg/g)
Yield of rPINA (%)
PINA-Chloroplast 0.663 ± 0.0126 5.173 0.00517 0.702 ± 0.059 51.7 0.836 ± 0.033
PINA-Cytosol 0.658 ± 0.010 4.052 0.00405 0.602 ± 0.029 40.5 0.642 ± 0.032 PINA-ER 0.667 ± 0.012 3.972 0.00397 0.612 ± 0.044 39.7 0.63 ± 0.033 PINA-Apoplastd 0.10877 ± 0.003 - - - - - Negatived 0.0547 ± 0.0015 - - - - - a average of at least 4 independent tests; b different dilutions were tested, data show 200-fold dilution. c FW: fresh weight; d data not applicable for apoplast and negative samples.
C h loro
p las t
C y tos o l
E R0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
P I N A
T a rg e tin g
Per
cen
tag
e
Figure 3.12 Quantification of PINA proteins targeted to chloroplast, cytosol and ER by ELISA.TSP extract from leaves harvested after infiltration was added to ELISA plate and PINA detected using the Durotest antibody. Bars show the mean percentage of protein extracts from four individual leaves and the error bars indicate the standard error of the mean (SEM).
y = 1.6016x-1.044 R² = 0.9927
0
0.2
0.4
0.6
0.8
1
1.2
0 0.1 0.2 0.3 0.4 0.5 0.6
Abs
orba
nce
(450
nm
)
3% non-durum wheat concentration (µg/mL)
0.836%
0.642% 0.63%

Chapter 3 Protein expression
- 100 -
3.2.5 Expression of the recombinant PINB protein by the viral vector system
The constructs pICH-PINB-His and pICH-PINB-His-SEKDEL made using different 5’
vector modules were infiltrated into N. benthamiana leaves. The TSPs were extracted
and analysed using SDS-PAGE (Figure 3.13) and quantitated by Bradford assay (Table
3.3).
Figure 3.13 Coomassie Blue stained gel to analysis TSP from infiltrated leaves. Lane 1: infiltration with chloroplast targeted PINB; lane 2: infiltration with cytosol targeted PINB; lane 3: infiltration with ER targeted PINB; lane 4: infiltration with apoplast targeted PINB.
The recombinant PINB (rPINB) was detected by western blot using the above antibody.
The molecular weights for C-terminally His-tagged PINB with or without the SEKDEL
were predicted, using the ‘compute pI/Mw tool’ at the ExPASy (Section 2.10.3) at
15.13 kDa and 14.43 kDa, respectively. The predicted subcellular localisation of PINB
by WoLF PSORT (Section 2.10.4) showed the highest score for chloroplast targeting.
The western blot showed the friabilin proteins at the expected size (~15 kDa), in the
positive control, 3% non-durum wheat. The recombinant PINB with or without the
SEKDEL was detected at the unexpected size of ~45 kDa bands (between 50 kDa and
37 kDa) (Figure 3.14), only when the TSP samples were not heated prior to western blot
(as discussed in section 3.2.6). The ~45 kDa PINB was noted when targeted to the
chloroplast, cytosol or ER, but not when targeted to apoplast.
1 2 3 4
75kDa
50kDa
37kDa
15kDa
25kDa

Chapter 3 Protein expression
- 101 -
Figure 3.14 Determination of PINB expressions by western blotting of TSP extracts from infiltrated leaves. Lane 1: 3% non-durum wheat as positive control for antibody; lane 2: negative control (5’+integrase modules only); lanes 3 and 9: ER targeted PINB; lanes 4 and 5: cytosol targeted PINB; lanes 6 and 7: chloroplast targeted PINB lane 8: apoplast targeted.
The TSP samples from freshly infiltrated leaves were quantified by Bradford assay and
analysed by ELISA (Section 2.11.4) with Durotest antibody to measure the yield of
recombinant PINB protein. ELISA plates were coated with 0.3 µg TSP/well. The assay
tested protein samples in three dilutions (1/50, 1/100 and 1/200). The absorbance was
read at 450 nm, and the PINB concentration determined from the 3% non-durum wheat
standard curve as above (Figure 3.11). Table 3.3 shows the concentration of PINB in
infiltrated leaves in 1/200 dilution test. As shown in Figure 3.15, the maximum yield
for PINB was ~0.894% of TSP, or approximately ~70 µg/g of fresh weight using the 5’
vector module for chloroplast. PINB yields of up to ~0.73% and ~0.68% TSP, or ~45
µg/g and ~40 µg/g of fresh weight were obtained for cytosol and ER targeting,
respectively.
Table 3.3 ELISA data analysis for quantification of recombinant PINB (rPINB) protein
Sample name OD450a
rPINB (µg/m
L)
rPINB (mg/mL)b
TSP (mg/mL)
FWc (µg/g)
Yield of rPINB (%)
PINB-Chloroplast 0.66 ± 0.011 6.294 0.00629 0.762 ± 0.045 62.9 0.894 ± 0.051 PINB-Cytosol 0.661 ± 0.012 4.533 0.00453 0.692 ± 0.060 45.3 0.734 ± 0.028 PINB-ER 0.66 ± 0.019 4.053 0.00405 0.615 ± 0.034 40.5 0.672 ± 0.023 PINB-Apoplastd 0.113 ± 0.0056 - - - - - Negatived 0.0547 ± 0.001 - - - - -
a average of at least 4 independent test; b different dilutions were tested, data show 200-fold dilution; c FW: fresh weight; d data not applicable for apoplast and negative samples.
1 2 3 4 5 6 7 8 9
50kDa
15kDa
75kDa
37kDa

Chapter 3 Protein expression
- 102 -
C h loro
p las t
C y tos o l
E R0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
P IN B
T a rg e tin g
Per
cent
age
Figure 3.15 Quantification of PINB proteins targeted to chloroplast, cytosol and ER by ELISA. TSP extract from leaves harvested after infiltration was added to ELISA plate and PINB detected using the Durotest antibody. Bars show the mean percentage of protein extracts from four individual leaves and the error bars indicate the standard error of the mean (SEM). 3.2.6 Optimisations of expression of His-tagged recombinant PINA and PINB
The optimal time of PIN harvest was investigated by infiltrating leaves with constructs
for targeting to the chloroplast and cytosol and harvesting at 8, 10 or 12 days post-
infiltration (dpi). One µg of TSP (as assayed by Bradford method) was used for ELISA
in two dilutions (1/100 and 1/200). Total soluble protein (TSP) from the infiltrated
leaves harvested at 10 dpi resulted in a slightly higher OD450 compared to 8 or 12 dpi
(Figure 3.16). Overall, the maximal yields of PINs were achieved at 10 dpi, using these
modules.
D a y 8
D a y 10
D a y 12
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
C y to s o l
H a rv e s t
OD
450n
m
P IN A
P IN B
N eg
D a y 8
D a y 10
D a y 12
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
C h lo r o p la s t
H a rv e s t
OD
450n
m
P IN A
P IN B
N eg
Figure 3.16 Detection of PIN proteins harvested at different times post-infiltration, by ELISA. Quantified TSP samples were added to ELISA plate and PINs detected using the Durotest antibody. The negative control was protein extracted from leaf material infiltrated with only the 5’+integrase modules. The bars indicate the mean OD450 of protein extracts from two individual leaves and the error bars indicate the standard error of the mean (SEM).
0.894%
0.73% 0.68%

Chapter 3 Protein expression
- 103 -
The quantified TSP samples (approximately 50 µg) were heated at 60°C, 65°C and
70°C for 15, 30 seconds and 1 minute to denature any recombinant PINs prior to
western blotting. No bands were observed at 70°C for the tested proteins at all three
time points (data not shown). The same result was observed for 60°C and 65°C as
shown in Figure 3.17. Heating at 65°C for 30 seconds led to faint bands with size shift
near 37 kDa compared to the unheated samples (Figure 3.17). No bands could be
detected when the samples were heated at 65°C for 1 minute (Figure 3.17). This
demonstrates that the plant-made recombinant PINs are heat-sensitive.
Figure 3.17 Heat sensitivity of plant-expressed PINA and PINB detected by western blot. Lane 1 and 2: chloroplast-targeted PINA and PINB, respectively, in TSP heated at 65°C for 30 seconds; lanes 3 and 4: these PINA and PINB in TSP heated at 65°C for one minute; lane 5 and 6: these PINA and PINB, in TSP not heated; lane 7: negative control (5’+integrase modules only).
1 2 3 4 5 6 7
50kDa
15kDa
75kDa
37kDa

Chapter 3 Protein expression
- 104 -
A number of different extraction buffers (Section 2.13.7; Table 2.10) were then tested
and the proteins detected by western blot. Extraction buffer 1 was used for all further
analyses, as other buffers did not improve the yield or extent of denaturation of PINs.
Further, in the optimisations, only one western blot confirmed the chloroplast-targeted
PINA and PINB at ~15 kDa (Figure 3.18), expected for His-tagged PINA and PINB
monomers, predicted to be 14.27 kDa and 14.43 kDa, respectively. These bands were
obtained using extraction buffer 1 which was without EDTA, and without heating the
TSP, and when approximately 90 µg of TSP samples were loaded on gels.
Figure 3.18 Determination of PINA and PINB protein expression in infiltrated leaves with viral vector module by western blot of TSP in expected size. Lane 1: chloroplast targeted PINA (~15 kDa); lane 2: chloroplast targeted PINB (~15 kDa); lane 3: negative control (5’+integrase modules only); lane 4: 3% non-durum wheat as positive control.
1 2 3 4
25kDa
15kDa
37kDa
50kDa

Chapter 3 Protein expression
- 105 -
3.3 Results for purification of PINA and PINB
3.3.1 His-tag purification under native conditions
The PINs were purified from TSP of leaf infiltrated with pICH-PINA-His or pICH-
PINB-His, in combination with the chloroplast 5’ module, due to the high yield of
recombinant PINs when targeted to the chloroplast. Purification was performed using
Ni-NTA resin column (Section 2.13.9). In order to increase the chances of C-terminal
His-tag being correctly folded and functional, the soluble PIN-His proteins were
purified under native conditions (without denaturing buffer) and at 4°C to minimize
protein degradation. Samples (approximately 50 µg) that had indicated chloroplast-
expressed PINs by western blots before purification, and the fractions passed over a Ni-
NTA resin column with wash buffer (Table 2.5), were run on 15% SDS-PAGE and
visualized by Coomassie Blue staining (Figure 3.19). The washed fractions with
imidazole (25 mM) in the wash buffer showed faint bands at 50 kDa. However, the 37
kDa and 15 kDa bands were absent on gel (Figure 3.19; lane 4, 5).
Figure 3.19 SDS-PAGE of TSP and wash steps of His-tag purification under native conditions. Lane 1: Dual Xtra standards (Bio-Rad); lane 2 and 3: TSP of chloroplast-expressed PINA and PINB respectively; lane 4 and 5: fractions from Ni-NTA column with imidazole (25 mM) in wash buffer for PINA and PINB purification respectively.
50
37
15
1 2 3 4 5
75
250
25
kDa

Chapter 3 Protein expression
- 106 -
His-tagged PINs were eluted with an elution buffer containing 250 mM imidazole
(Table 2.5) and then dialysed to remove imidazole, then concentrated (Sections 2.13.12
and 2.13.13) and quantitated using Bradford assays. This was followed by SDS-PAGE
and western blot analysis with ~48 µg and ~52 µg for PINA and PINB, respectively
(Figure 3.20). The fractions exhibited a few faint bands and strong bands at 37 kDa and
15 kDa for both PINs in the stained gel (lanes 2, 3) which may correspond to the
multimer and monomer sizes of His-tagged PINs, respectively. The results indicate that
the target proteins were retained on the resin and were only visible after elution with
high imidazole (250 mM) in the elution buffer. The western blot of the eluted fractions
(42 µg, 48 µg and 52 µg protein for negative control, PINA and PINB, respectively)
indicated a broad band at ~37 kDa, confirming the identity of 37 kDa bands as PIN
proteins (Figure 3.20; lanes 6, 7). However, the 15 kDa band not shown signal with
antibody which may not just detected with Durotest. The faint band above ~37 kDa
reacted with the Durotest antibody in PINA and PINB samples (lane 6 and 7) which was
probably observed as 45 kDa protein when TSP samples were analysed.
Figure 3.20 SDS-PAGE and western blot analysis with Durotest antibody for eluted fraction on Ni-NTA resin columns based on His-tag purification under native conditions. Lane 1 and 5: eluted fraction of negative control (5’+integrase modules only); lane 2 and 6: eluted fraction of PINA on SDS-PAGE and western blot respectively; lane 3 and 7: eluted fraction of PINB on SDS-PAGE and western blot respectively; lane 4: Dual Xtra standards (Bio-Rad).
75kDa
1 2 3 4 5 6 7
50kDa
37kDa
15kDa
42µg 48µg 52µg 42µg 48µg 52µg

Chapter 3 Protein expression
- 107 -
3.3.2 His-tag purification under denaturing conditions
His-tag purification was performed using the refolding method (Section 2.13.10) to
observe the effect of a protein denaturant such as urea, which disrupts the non-covalent
bonds and increases protein solubility. His-tag purification generally works well under
denaturing conditions (Waugh, 2005). Purification steps were typically performed at
4°C to minimize degradation. PINs were eluted from the Ni-NTA column with a buffer
containing 250 mM imidazole and 6 M urea (Table 2.5) then dialysed (Section 2.13.12)
to remove imidazole and urea, and protein concentrations measured by Bradford
assays. This was followed by SDS-PAGE and western blotting with ~32 µg and ~35
µg for eluted samples of PINA and PINB, respectively, and 30 µg for negative control.
A single band was observed for PINA and PINB at approximately 15 kDa on the gel
(Figure 3.21; lanes 2 and 3) which could be their putative monomer. The same
amounts of protein samples were analysed by western blot, however the denatured
monomeric forms of both PINs were not detected (lanes 6 and 7).
Figure 3.21 SDS-PAGE and western blot analysis with Durotest antibody for eluted fraction on columns with 6M urea in elution buffer. Lane 1: Dual Xtra standards (Bio-Rad); lane 2 and 6: eluted fraction of PINA on SDS-PAGE and western blot respectively; lane 3 and 7: eluted fraction of PINB on SDS-PAGE and western blot respectively; lane 4 and 8: eluted fraction of negative control (5’+integrase modules only) lane 5: 3% non-durum wheat as a positive control for antibody.
30 µg
1 2 3 4 5 6 7 8
37
25
20
15
kDa
32 µg 35 µg 30 µg

Chapter 3 Protein expression
- 108 -
3.3.3 Hydrophobic interaction purification
In a previous attempt to purify the PINs using His-tag, the proteins had not shown a
dominant band on SDS-PAGE (results in section 3.3.1). This may be due to the other
plant proteins binding to the Ni-NTA resin. Therefore, hydrophobic interaction
purification, a separation technique that uses the properties of hydrophobicity to
separate proteins from one another, was applied (Section 2.13.11). Proteins passing
through the column with hydrophobic amino acid side chains on their surfaces are able
to bind to the hydrophobic groups on the column. The interactions are too weak in
water (a polar solvent), however the additions of salts such as ammonium sulfate
[(NH4)2SO4] to the buffer result in stronger hydrophobic interactions. Ammonium
sulfate concentrations of 60%, 70% and 80% saturation were used for phase separation
by ethanol extraction (Figure 3.22).
Figure 3.22 Fractions in the course of extraction procedure. Protein was extracted into upper phase (ethanol) after addition of ammonium sulfate (60%, 70% and 80% saturation) and ethanol.
The proteins were eluted from Butyl-Toyopearl column, quantified by Bradford assay
and ~40 µg of each preparation was run on SDS–PAGE gel and visualized by
Coomassie Blue staining. 70% and 80% ammonium sulfate did not show clear bands of
PINs (data not shown). However, elution with reduced (60%) ammonium sulfate led to
pure proteins, seen as two bands at ~37 kDa and ~15 kDa, assumed to be a multimeric
and monomeric forms of PINs (Figure 3.23; lanes 2 and 3). It is notable that these
bands were only recovered by hydrophobic interaction purification. Further, strong
bands at about 50 kDa were observed on the gel for chloroplast PIN proteins and also
negative control (Figure 3.23; lanes 2, 3, 5). This band may represent the ribulose 1, 5-
60% 70% 80%

Chapter 3 Protein expression
- 109 -
bisphosphate carboxylase/oxygenase (RuBisCO) large subunit, which is the most
abundant protein in leaves (Bally et al., 2009).
Figure 3.23 SDS-PAGE for fractions eluted from the Butyl-Toyopearl column based on hydrophobic interaction system. Lane 1: Dual Xtra standards (Bio-Rad); lane 2 and 3: eluted fractions of PINA and PINB respectively; lane 4: no sample; lane 5: eluted fraction of negative control (5’+integrase modules only). Further purification was attempted in order to remove Rubisco. TSP fractions were
prepared as per previous purifications and passed over a Butyl-Toyopearl column.
After this, RuBisCO was removed using sodium phytate precipitation (Section 2.13.14).
The stained gel showed successful removal of Rubisco, but the bands of interest (37
kDa and 15 kDa) were also very faint (Figure 3.24; lanes 2 and 3).
Figure 3.24 SDS-PAGE of eluted fraction on Butyl-Toyopearl column based on hydrophobic interaction system and sodium phytate precipitation. Lane 1: Dual Xtra standards (Bio-Rad); lane 2 and 3: eluted fractions of PINA and PINB with sodium phytate precipitation respectively.
1 2 3 kDa
50
37
15
75
50
37
15
1 2 3 4 5
75
kDa

Chapter 3 Protein expression
- 110 -
Western blot analysis using the Durotest antibody resulted in a broad band at ~37 kDa,
confirming this as the multimer form of PINA and PINB (Figure 3.25; lanes 1 and 2).
The result confirmed that this antibody is specific for both PINs, but also suggest that it
is most likely not able to recognize the monomer forms of PINA or PINB, as seen
earlier (Figure 3.20).
Figure 3.25 Western blot of eluted fractions on Butyl-Toyopearl column based on hydrophobic interaction system. Lane 1 and 2: eluted fractions of PINA and PINB, respectively; lane 3: eluted fractions of negative control (5’+integrase modules only).
1 2 3
37kDa
25kDa
15kDa

Chapter 3 Protein expression
- 111 -
3.3.4 Western blot analysis using anti His-tag for PIN proteins
The TSP of infiltrated leaves using 3’ PINA and PINB modules and different 5’ vector
modules (cytosol or chloroplast) were subjected to western blot analysis with Durotest
antibody. The samples which showed a signal with this antibody were used for western
blot with anti-His antibody (Table 2.3). Different concentrations of proteins and anti-
His antibody were tested, but no signals detected. Further, after purification with two
different systems, i.e., His-tag purification under native conditions, and hydrophobic
interaction purification, purified PINs were used for western blots. Following His-tag
purification under native conditions, no bands for C-terminally His-tagged PINs were
observed (Figure 3.26; lanes 3 and 4), while the samples after hydrophobic purification
showed two bands, corresponding to the PIN multimer of ~37 kDa and monomer size of
~15 kDa (lanes 1 and 2).
Figure 3.26 Western blot analysis of PIN proteins purified with two different systems. Lane 1 and 2: eluted fractions of PINA and PINB from Butyl-Toyopearl columns respectively; lane 3 and 4: eluted fractions of PINA and PINB on Ni-NTA resin columns respectively; lane 5: eluted fraction of negative control (5’+integrase modules only). After purification the recombinant PIN proteins, to obtain an accurate mass PINs the
preliminary analysis has been done using Maldi-TOF MS (detailed in Appendix III).
However, the results were inconclusive as sequences of the peptide were determined to
be unknown.
1 2 3 4 5
37kDa
15kDa
25kDa

Chapter 3 Protein expression
- 112 -
3.4 Results of antimicrobial activity tests
3.4.1 Antibacterial activity
The chloroplast-expressed purified PINs with different purification systems were tested
for antibacterial activity (as detailed in Chapter 2), using the protein stock (~1 mg/mL).
The synthetic peptide PuroA (FPVTWRWWKWWKG-NH2) was used as positive
control (Section 2.12.1). The negative control used was the TSP of N. benthamiana
infiltrated by 5’ and integrase modules that was purified based on affinity and
hydrophobic purifications. The activities were tested against Escherichia coli (ATCC
25922; a Gram negative bacterium) and Staphylococcus aureus (ATCC 25923; a Gram
positive bacterium); the strains of both can cause a range of illnesses in humans. The
minimum inhibitory concentration (MIC) was defined ‘the lowest concentration of an
antimicrobial agent which will inhibit the growth of a microorganism after overnight
incubation’ (Wiegand et al., 2008). The MIC results were observed by eye and by
measuring the optical density at 595 nm (OD595) of each well after 18 hours incubation
of cells with the peptide/protein. Activity of PuroA was consistent with previous work
in our laboratory (Phillips et al., 2011) (Table 3.4). The results confirmed in vitro
antibacterial abilities of plant-made recombinant PINs purified by His-tag purification
under native conditions and also by hydrophobic interaction purification. The PINs
purified under denaturing conditions had no observable activity. Both PINA and PINB
His-tag purified under native conditions showed activity against E. coli (MIC 64
μg/mL) which was less than that of the PuroA peptide (MIC 16 µg/µL) (Table 3.4;
Figure 3.27). PINA was observed to be active against S. aureus (MIC 64 μg/mL),
however, PINB showed higher activity (MIC 125 µg/mL). With hydrophobic
purification, both PINA and PINB showed better activity against E. coli (32 μg/mL),
and PINA showed the same activity (MIC 32 μg/mL) against S. aureus while PINB was
less active against it (64 μg/mL). These observations confirm the in vitro antimicrobial
abilities of recombinant PINs. Further, the PINs purified based on hydrophobic
interaction had better activity compared to both the His-tag purification systems. The
antibacterial activity of mixed PINA and PINB (1:1) (~1 mg/mL stock) were also tested
against both bacteria. The result observed same activities for both purification systems
(32 μg/mL and 64 μg/mL) (Table 3.4).

Chapter 3 Protein expression
- 113 -
Figure 3.27 Example of Minimum Inhibitory Concentration (MIC) assay for PuroA peptide and plant-made recombinant PIN proteins against E. coli. Peptide/protein dilution series from column 1 to 8; A: negative control (extract of leaf infiltrated with 5’+integrase modules only); B: positive control (PuroA peptide); C: rPINA (His-tag purification under denaturing conditions); D: rPINA (His-tag purification under native conditions); E: rPINA (hydrophobic purification); F: rPINB (His-tag purification under denaturing conditions); G: rPINB (His-tag purification under native conditions); H: rPINB (hydrophobic purification). Wells were inoculated with E. coli (ATCC 25922) and MIC recorded as the lowest concentration to completely inhibit growth. GC column= growth control of E. coli.
Table 3.4 Antibacterial activity of plant made recombinant PINA and PINB
Protein/peptide name Purification method
MIC (µg/mL)a
E. coli S. aureus PuroA
Positive control Synthetic peptide
FPVTWRWWKWWKG-NH2 16 (±0) 16 (±0)
rPINA His-tag purification under denaturing conditions >250 >250
rPINA His-tag purification under native conditions 64 (±0) 64 (±0)
rPINA Hydrophobic interaction purification 32 (±0) 32 (±0)
rPINB His-tag purification under denaturing conditions >250 >250
rPINB His-tag purification under native conditions 64 (±0) 125 (±0)
rPINB Hydrophobic interaction purification 32 (±0) 64 (±0)
PINA+PINB His-tag purification under native conditions 32 (±0) 64 (±0)
PINA+PINB Hydrophobic interaction purification 32 (±0) 64 (±0)
Negative controlb His-tag purification under native conditions >250 >250
Negative controlb Hydrophobic interaction purification >250 >250 aDefined as the lowest concentration that completely inhibited bacterial growth/ the mean of triplicate assays. bTSP of N. benthamiana infiltrated by 5’ and integrase modules only.
µµg/mL250 125 64 32 16 8 4 2 GC A
B
C
E
G
D
F
H

Chapter 3 Protein expression
- 114 -
3.4.2 Antifungal activity
The purified recombinant PINs were also tested for activity against five common phyto-
pathogenic fungi (Section 2.12.3). Rhizoctonia cerealis and Rhizoctonia solani are
responsible for sharp eye spot in wheat and sheath blight in rice, respectively (Hamada
et al., 2011; Taheri and Tarighi, 2011). Colletotrichum graminicola is responsible for
anthracnose in many cereals including wheat and maize (Dean et al., 2012). Drechslera
brizae is a fungus that causes post-harvest rot in crops (Naureen et al., 2009). Negative
control wells contained purified TSP of N. benthamiana infiltrated by 5’ and integrase
modules, and positive control wells contained PuroA. The MIC was defined as the
lowest concentration of the protein/peptide to inhibit fungal growth (Espinel-Ingroff and
Cantón, 2007), which was observed visually only. The results are presented in Table
3.5. The PIN proteins purified under both the native condition of His-tag purification
and the hydrophobic interaction purification showed activity against R. solani (250
µg/mL and 125 µg/mL) but were less active than the PuroA peptide (32 µg/mL) (Table
3.5; Figure 3.28). Proteins purified under denaturing conditions had no observable
antifungal activity.
Figure 3.28 Example of microdilution plate Minimum Inhibitory Concentration (MIC) assay for filamentous fungi against R. solani. Peptide/protein dilution series from column 1 to 9; A: negative control (extract of leaf infiltrated with 5’+integrase modules only); B: positive control (PuroA peptide); C: rPINA (His-tag purification under denaturing conditions); D: rPINA (His-tag purification under native conditions); E: rPINA (hydrophobic interaction purification); F: rPINB (His-tag purification under denaturing conditions); G: rPINB (His-tag purification under native conditions); H: rPINB (hydrophobic interaction purification). GC column=growth control of R. solani.

Chapter 3 Protein expression
- 115 -
Table 3.5 Antifungal activity of plant made recombinant PINA and PINB
aDefined as the lowest concentration of the peptide or protein to inhibit fungal growth/ the mean of triplicate assays. bTSP of N. benthamiana infiltrated by 5’ and integrase modules only.
Protein/ peptide name Purification method
MIC (µg/mL)a C.Gramini
cola D.brizae F. oxy-sporum
R. solani
R. cerealis
PuroA Positive control
Synthetic peptides FPVTWRWWKWWKG-NH
2 250 (±0) 250 (±0)
>500
32 (±0)
125 (±0)
PINA His-tag purification under denaturing conditions
>500 >500 >500 >500 >500
PINA His-tag purification under native conditions 500(±0) 500(±0) >500 250(±0) 250(±0)
PINA Hydrophobic interaction purification 500(±0) 500(±0) >500 125(±0) 250(±0)
PINB His-tag purification under denaturing conditions
>500 >500 >500 >500 >500
PINB His-tag purification under native conditions 500(±0) 500(±0) >500 250(±0) 250(±0)
PINB Hydrophobic interaction purification 500(±0) 500(±0) >500 125(±0) 125(±0)
PINA+PINB His-tag purification under native conditions 500(±0) 500(±0) >500 125(±0) 125(±0)
PINA+PINB Hydrophobic interaction purification 500(±0) 500(±0) >500 125(±0) 125(±0)
Negative controlb
His-tag purification under native conditions
>500 >500 >500 >500 >500
Negative controlb
Hydrophobic interaction purification
>500 >500 >500 >500 >500

Chapter 3 Protein expression
- 116 -
3.4.3 Protein stability at different temperature
The effect of temperature on the activity and stability of PIN proteins purified by His-
tag purification under native conditions were tested against E. coli and determined by
MIC assay. PIN proteins were stored at 37°C, RT (between 20°C-25°C), 4°C, -20°C
and -80°C for one week, then tested against E. coli. The results showed stable activity
against E. coli when stored at -20°C and -80°C. PIN proteins showed better activity
against E. coli at day one (MIC 64 μg/mL) compared to when stored at 4°C (MIC 250
μg/mL). The PuroA peptides retained with their original activity after one week of
incubation at room temperature (between 20°C-25°C), 4°C, -20°C and -80°C for one
week (Table 3.6).
Table 3.6 Stability of recombinant PINs at different incubation temperature
Peptide/Protein name
Incubation Temperature
MIC (µg/mL)a against E. coli at day 1
MIC (µg/mL)* against E. coli at day 7
PuroA Allb 16 (±0) 16 (±0) rPINA/rPINB -80°C 64 (±0) 64 (±0) rPINA/rPINB -20°C 64 (±0) 64 (±0) rPINA/rPINB 4°C 64 (±0) 250(±5) rPINA/rPINB 20°C-25°C 64 (±0) >250 rPINA/rPINB 37°C 64 (±0) >250
a The mean of triplicate assays, b PuroA not tested at 37°C.

Chapter 3 Protein expression
- 117 -
3.4.4 Haemolytic activity
The haemolytic activity of the purified PINs against sheep RBCs was determined as a
measure of toxicity to eukaryotic cells (Section 2.12.5). Positive haemolytic activity
was defined as the recombinant protein/peptide dose required to lyse 50% of the RBCs
(HD50). PBS (pH 7.4) and 1% Triton X-100 were included as a negative (zero%
haemolysis) and positive (100% haemolysis) controls, respectively. The recombinant
PINs purified with His-purification system under native condition at 125 μg/mL, 64
μg/mL, 32 μg/mL and 16 μg/mL did not show any notable activity (Figure 3.29, Table
3.7), confirming their non-lysis of mammalian cells and specificity against microbial
membranes. However, the PINs at 500 μg/mL and 250 μg/mL appeared to show higher
haemolytic activity than the peptide PuroA.
Figure 3.29 Haemolytic activity assays against sheep red blood cells (RBCs). Dilution series from column 1 to 6: 500 µg/mL, 250 µg/mL, 125 µg/mL, 64 µg/mL, 32 µg/mL and 16 µg/mL. rPINA: recombinant plant made PINA; rPINB: recombinant plant made PINB; Pos: Positive control 100% RBC lysis (1% Triton X-100); Neg: Negative control 0% RBC lysis (PBS, pH 7.4). Table 3.7 Haemolytic activity of plant-made recombinant PINs
Protein/Peptide
Percent haemolysis of sheep RBCs by recombinant PINs Haemolysis (HD50)a 500
(µg/mL) 250
(µg/mL) 125
(µg/mL) 64
(µg/mL) 32
(µg/mL) 16
(µg/mL) PuroA 15 10 6 4 0 0 >500 rPINA 32 22 10 6 0 0 >125 rPINB 20 20 8 6 0 0 >125
a Results represent the average of three separate experiments conducted in duplicate.

Chapter 3 Protein expression
- 118 -
3.5 Discussion
3.5.1 Expression
A number of studies with transgenic Pin genes have reported, the purpose being to test
their causative roles of PIN proteins in altering grain texture (Beecher et al., 2002a;
Hogg et al. 2004; Krishnamurthy and Giroux, 2001; Wada et al., 2010; Xia et al., 2008)
or testing whether they can impart in vivo antimicrobial activity (Faize et al., 2004; Kim
et al., 2012a; Krishnamurthy et al., 2001; Luo et al., 2008; Zhang et al., 2011) as
detailed in Chapter 1 (Sections 1.4.2 and 1.5.1). Further, mature full-length PINA and
PINB have been reported to be over-expressed in bacterial expression systems
(Capparelli et al., 2006; Capparelli et al., 2007) and in BY-2 Nicotiana tabacum cell
suspension culture (Sorrentino et al., 2009). In the BY-2 culture, only the recombinant
PINB targeted to the chloroplast was observed with an apparent size of 20 kDa and
yield of 0.35% of TSP but no signal were observed using two constructs for apoplast
targeting for both PINs (Sorrentino et al., 2009). Further, peptides similar to the PINs in
terms of being cysteine-rich and antimicrobial, but smaller, expressed in transgenic
tobacco lines represented only 0.1% of the TSP, and their expression either intra- or
extra-cellularly did not result in increased fungal resistance (De Bolle et al., 1996).
Plants as an alternative production platform for active recombinant therapeutic proteins
offers several advantages including lower production costs and high biomass volume.
In this study we used a transient transformation technology, the magnICON® for
production of PINs using N. benthamiana. This system is based on the deconstructed
tobacco mosaic virus (TMV) and is distinguished by the use of independent modules
harboring binary vectors, encoding different virus components, and allowing for DNA
delivery into plant cells by A. tumefaciens. The in vivo recombination, mediated by the
integrase (pICH14011) is accompanied with assembly of various pro-vectors, followed
by intron splicing, rendering an active viral replicon capable of producing the protein of
interest, and yields of up to 40% of TSP (Gleba et al. 2007). The replicon can move
systemically through the plant (Gleba et al. 2005; Marillonnet et al. 2004). The
assembly of recombinant proteins in plant tissues is critical for viability the plant
systems as bioreactors for therapeutic proteins (Streatfield 2007). The magnICON®
system is considered to be fast, requiring approximately 10 days for transformation and

Chapter 3 Protein expression
- 119 -
less than a month for the recovery of the expressed protein. This is highly desirable
compared to other systems such as chloroplast transformation which requires more than
two months, and nuclear stable transformation that requires several months (Patiño-
Rodríguez et al., 2013).
The successful expression of GFP proteins was visualized by examination under UV
light and in western blot using the anti-GFP antibody. Different constructs were used to
produce the PIN proteins in leaves of N. benthamiana. Both PIN proteins produced the
highest amount 10 days post infiltration when targeted to the chloroplast and cytosol.
As a commercial standard is not available for PINs, the 3% non-durum wheat in
Durotest kit for standard in ELISA assay was used to quantify PIN proteins; hence the
quantify estimates for PINs are approximate. The results demonstrate PINA and PINB
were transiently expressed in N. benthamiana and accumulated to ~0.83% and ~0.89%
of the TSP, respectively, following targeting to chloroplast. Western blot analysis by
Durotest antibody confirmed the size at approximately 45 kDa when the chloroplast and
cytosol targeting 5’ modules were used, and also with C-terminal SEKDEL added for
retention of the expressed protein in the ER. This finding suggests the formation of
aggregates not only inside chloroplasts but also in the cytosol and ER compartments of
the plant cell.
PINA is known to form aggregates in solutions with high ionic strength and low pH (Le
Bihan et al., 1996), correct folding being dependent on pH and a contributor to low
protein solubility (Issaly et al., 2001). Further, an earlier study has noted the importance
of protein aggregates of recombinant soybean agglutinin (rSBA) in N. benthamiana
necessary for its biological activity (Tremblay et al., 2011). Similar folding pattern has
also been observed, when proteins containing multiple disulphide bonds were expressed
in chloroplasts, including CTB-proinsulin (Ruhlman et al., 2007) and interferon-a2b
(Arlen et al., 2007) RC101 and PG1 (Lee et al., 2011b).

Chapter 3 Protein expression
- 120 -
3.5.2 Purification
Both PINA and PINB are disulphide-bonded proteins with unique TRD. PINs contain
10-Cys backbone proposed to form five disulphide bonds that have an important role in
protein stability and activity (detailed in Chapter 1). PINA and PINB have been
expressed in bacterial systems using His-tag (Capparelli et al., 2006) and GST-tag
(Capparelli et al., 2006; Capparelli et al., 2007). Furthermore, 3×FLAG N-terminal tag
was added for western blot analysis using mouse anti-flag monoclonal antibody which
detected recombinant PINB at ~20 kDa (Sorrentino et al., 2009).
Affinity tags can have role for promoting the solubility of the expressed fusion proteins
(Hearn and Acosta, 2001; Terpe, 2003; Waugh, 2005). Ideally, the tags should not
affect the structure and stability of the proteins. On the other hand, they may affect
proteins advantageously by increasing expression, solubility and stability (Arnau et al.,
2006; Hearn and Acosta 2001; Smyth et al., 2003; Terpe, 2003) and promote proper
folding (Smyth et al., 2003; Waugh, 2005). The His-tag with metal IMAC systems is
currently the most commonly used affinity tag (Listwan et al., 2004; Waugh, 2005).
Generally His-tag can interact with a chromatographic matrix like Ni-NTA resin as it is
most attracted to immobilized metal ions, such as cobalt, nickel and copper (Terpe,
2003; Waugh, 2005) the selectivity being based on the fact that the density of His
residues in this tag is higher than that on the surface of many proteins (Gaberc-Porekar
and Menart 2001) and other potential electron donor side chains include tryptophan and
cysteine (Scopes, 1993). Since both PINs contain tryptophan and cysteine, they may be
capable of binding to metal-chelate columns, hence both were fused translationally to a
small His-tag.
After affinity purification using the His-tag for PINs expressed in the chloroplast,
western blot analysis confirmed two bands at 45 kDa and 37 kDa. However the ~45
kDa proteins were observed as faint bands (Figure 3.20; lanes 6 and 7). In addition, for
the PINs protein sample infiltrated using the chloroplast 5’-module, the western blot
detected the small and faint band at ~30 kDa before purification (Figure 3.10 and 3.17;
lane 5). The expected sizes of the PIN monomers with His-tag were ~15 kDa. Thus,
the bands may correspond to multimers. Further, the His-tag could have affected the

Chapter 3 Protein expression
- 121 -
folding of the recombinant PIN proteins or incomplete cleavage of the chloroplast
transit peptide (Bruce, 2000; Emanuelsson et al., 1999), resulting in the size
discrepancy. The incomplete cleavage of the chloroplast transit peptide has also been
reported for the PyMSP4/5 antigen expressed in plants using the magnICON® system
chloroplast 5’-module (Webster et al., 2009). The artificial dicot chloroplast targeting
presequence for pICH12190 chlorolast targeted 5’ module coding for
[massmlssaavvatrasaaqasmvapftglksaasfpvtrkqnnlditsiasnggrvqca] (Marillonnet et al.,
2004; Dr. S. Marillonnet, personal commincation, April 2014) with estimated size of 6
kDa using the ‘compute pI/Mw tool’ at the ExPASy.
The hydrophobic purification system was attempted for C-terminally His-tagged PINs,
as this is widely used for the isolation of hydrophobic membrane proteins and
antimicrobial cationic peptides (Skosyrev et al., 2003). Hydrophobic patches of
proteins interact with a hydrophobic stationary phase. Ammonium sulfate decreases
solubility of most proteins, due to the salting-out effect, hence ethanol was used, being
less polar. The organic solvents, i.e., ethanol or n-butanol, enable protein precipitation
by electrostatic aggregations. Protein solubility during extraction is affected by a
complex mechanism using organic solvents. The salt promotes solubility of proteins by
preventing their electrostatic aggregation in the organic solvent (Samarkina et al., 2009).
However, the ethanol phase is also saturated with ammonium sulfate. It is important to
add ethanol as soon as possible after ammonium sulfate so as to avoid nonspecific
precipitation of target protein with other proteins. The two phases were formed within
three to five minutes after addition of ethanol and vigorous shaking, and centrifugation
was started as soon as the phases began to form. The n-butanol, a less polar and more
hydrophobic organic solvent than water was added to the ethanol phase to recover the
PINs in the lower aqueous phase. The hydrophobic interaction purification thus
allowed fast processing and ability to concentrate the sample. Butyl-Toyopearl column
was used for removing any compounds that were not visible in Coomassie Blue stained
gels. Both PINs were detected at ~37 kDa with the Durotest antibody, and this was the
only technique that showed the PINs with anti-His antibody in both multimer (~37 kDa)
and monomer (~15 kDa) bands. This finding is most significant and suggests that
hydrophobic purification affected the folding of PINs. Further, the results confirmed

Chapter 3 Protein expression
- 122 -
that the Durotest antibody is likely not able to detect monomeric forms of PINs.
However this antibody detected the positive at ~15 kDa which is protein complex of
friabilin with PINs as major part (detailed in section 1.2.1). Prior studies (Greenwell,
1992; Wiley et al., 2007) noted that the Durotest antibody probably did not react with
PINs after reduction of disulphide bonds or denaturation during fixation for
immunocytochemical analyses. The protein extracts from grain of a bread wheat line
and two transgenic lines of durum wheat expressing PINA and PINB were tested by
western blot using this antibody and unreduced PIN detected at 15 kDa (Wiley et al.,
2007). Recently, this antibody was used for immunolocalisation of PIN proteins in the
endosperm cell of transgenic rice (Fujiwara et al., 2014).
3.5.3 Characterisation of bioactivity
PINA and PINB have potent activities against a broad spectrum of microorganisms
(detailed in section 1.5). The recombinant PINA and PINB have been expressed in E.
coli and affinity-purified using His and GST tags, and tested against E. coli (MIC 30
μg/mL) and S. aureus (MIC 30 μg/mL) (Capparelli et al., 2006). The activity of
recombinant PINA expressed in E. coli and purified by affinity purification using NusA
fusion tag was reported to be active against E. coli (MIC 90 μg/mL) and S. aureus (MIC
150 μg/mL) (Miao et al., 2012). Furthermore, chloroplast targeted PINB in N. tabacum
cells has activity against E. coli (91% growth inhibition) (Sorrentino et al., 2009).
Several studies have revealed the successful production of low-cost and functional
antimicrobial peptides and proteins in transgenic tobacco (Jacob and Zasloff, 1994; Lee
et al., 2011b; Oey et al., 2009). Previously the ability of the Potato Virus X (PVX)
vector to produce an antimicrobial protein in N. benthamiana leaves has been reported
(Saioth et al. 2001). The human lactoferrin (hLfN) expressed in N. benthamiana using a
PVX is also active against E. coli and S. aureus (Li et al., 2004). The magnICON®
technology was also used in N. tabacum for producing the antimicrobial peptide
protegrin-1 (PG-1), which inhibited the growth of several bacterial and fungal
pathogens of humans (Patiño-Rodríguez et al., 2013). Hence the goal of this study was
to produce and purify functional PINs in planta.

Chapter 3 Protein expression
- 123 -
In the current study, purified soluble protein extracts from transiently-transformed N.
benthamiana leaves with magnICON® system, have been used for biological activity
assays of PIN proteins and sharp decrease observed in the viability of different
pathogens. Purified extracts of leaves infiltrated with PIN transgenes were added to in
vitro bioassays and showed successful expression of bioactive proteins. The
antibacterial activity of the His-tag purified PINA under native conditions showed a
similar activity against S. aureus and E. coli (MIC 64 μg/mL), however the purified
PINA using hydrophobic interaction system was more active against both S. aureus and
E. coli (MIC 32 μg/mL). His-tag purified PINB under native conditions had similar
activity as PINA against E. coli (MIC 64 μg/mL) but not against S. aureus (MIC 125
μg/mL). The activity results indicate that the hydrophobic interaction purified proteins
displayed 2-fold more activity against E. coli for both recombinant PINs. No inhibition
of microbial growth was detected with extracts from infiltrated leaves without PINs,
purified with both systems. The peptide PuroA (based on the TRD of wild type PINA)
used as positive control in the assays showed strong activity towards E. coli and S.
aureus (MIC 16 μg/mL), supporting previous reports (Jing et al., 2003; Phillips at al.,
2011). It is particularly noteworthy that the growth inhibition of both Gram-positive
and Gram-negative bacteria by recombinant PINs was high for both affinity and
hydrophobic systems, suggesting correct formation of disulphide bonds in the plant-
made PINs. The PIN proteins lose antimicrobial activity in the presence of 6 M urea.
Many proteins require a particular three-dimensional structure and disulphide bonds to
function. If the disulphide bonds are not formed correctly, the protein can degrade or
accumulate as insoluble inclusion bodies (Lesk, 2010). PINs, with their 10-Cys
backbone that forms five disulphide bonds may require these bonds for protein stability
and activity.
The antifungal activity of PIN proteins has been shown in vitro (Dubreil et al., 1998)
and in planta when Pin genes have been introduced transgenically, e.g., in rice
(Krishnamurthy et al., 2001), apple (Faize et al., 2004), durum wheat (Luo et al., 2008)
and wheat seeds overexpressing Pins (Kim et al., 2012a). This work further examined
the activity of the recombinant PINs against the five common fungi. Recombinant
plant-made PINs purified under His-tag and hydrophobic purification showed activity

Chapter 3 Protein expression
- 124 -
against R. solani and R. cerealis. PINA and PINB, when purified by hydrophobic
interactions, showed better activity against most organisms tested, in comparison with
the His-tag purification under native conditions. A possible reason might be related to
the presence of more monomer forms of PINs in these preparations, or better protein
folding. Initially, the assay was carried out, which showed no activity against C.
graminicola, D. brizae and F. oxysporum at MIC 250 µg/mL. Hence, the assay was
repeated at a higher MIC (500 µg/mL) for both recombinant PINs. Activity was
observed only against C. graminicola and D. brizae (not for F. oxysporum) for PINA
and PINB purified with His-tag under native conditions and hydrophobic purification
systems. A possible explanation may be the stability of PINs and the time of incubation
(at 25°C in dark for 7 days). The results of stability testing indicate that the plant-made
PINs are not stable at 25°C or 37°C for 7 days. However, prior study in our laboratory
has noted the thermal stability of PuroA and PuroB peptides (Alfred et al., 2013a).
Previously in vitro studies indicated that the two proteins work synergistically with
enhanced activity against fungal pathogens (Capparelli et al., 2005). In this work when
PINA and PINB were mixed for the antimicrobial assay shown the inhibition against
bacteria and fungi and have shown synergy between the two proteins against fungi and
bacteria. The haemolytic activity assay displayed in PuroA at >500 µg/mL, however the
haemolytic activity of the recombinant PINs showed at >125 µg/mL. This variation
between PuroA and PIN proteins may be related to plant alkaloids in the preparations,
such as nicotine most abundant alkaloid in the genus Nicotiana and recently found to
exhibit haemolytic activity in vitro against human erythrocytes (Jasiewicz et al., 2014).
The findings in this chapter are highly promising in providing a methodology for
production of PINs as potential therapeutic agents against various mammalian and plant
pathogens as well as for use in hygiene and food industries. The two wild type PINs are
reported act in a co-operative manner when binding to the surface of starch granules.
The following chapter investigates PIN interaction in a plant system using the BiFC
system.

CHAPTER 4
Determination of the protein-protein interactions (PPI) of PIN proteins using Bimolecular Fluorescence
Complementation (BiFC) in planta

Chapter 4 Protein interaction
- 126 -
4.1 Introduction
The wild type PIN proteins (PINA and PINB) are causatively associated with soft
wheat, while mutations in one or both Puroindoline genes result in hard textures, as
summarised in Chapter 1 (Sections 1.3 and 1.4). Both PINA and PINB associate with
biological membranes through their lipid-binding properties, which are also likely
involved in grain hardness determinants (Gautier et al., 1994; reviewed in Bhave and
Morris, 2008b; Morris, 2002; Nadolska-Orczyk et al., 2009). Transgenic studies (Hogg
et al., 2004; Martin et al., 2006; Swan et al., 2006, Wanjugi et al., 2007) have shown
that the PINs undergo some interaction or act in an inter-dependent manner in their
effects on grain texture. The two wild type PINs were found to act co-operatively to
prevent break-down of lipids in soft grains, mutations in either gene leading to higher
polar lipid degradation (Kim et al., 2012a). Furthermore, the tryptophan-rich domain
(TRD) has been considered the most important domain for the membrane-active nature
of PIN proteins. Despite the above literature on PINs, the nature of the interdependence
of the two proteins, interactions of PINs and any role of the TRD in these are unclear.
Addressing these important questions could help to improve wheat quality. Previous
work in our laboratory using the yeast 2-hybrid system provided evidence of in vivo
protein-protein interactions (PPI) in PIN proteins expressed in yeast cells (Ziemann et
al., 2008). The bimolecular fluorescence complementation (BiFC) assay is based on
complementation approaches to detect protein associations occurring in-vivo, and allows
visualisation of PPI in living cells by fluorescent signal (Gell et al., 2012; Hu et al.,
2002). This chapter investigates (i) whether any physical interactions occur in PINA
and PINB in planta using BiFC system; (ii) the effect of deletions of the hydrophobic
domain (HD) domains from both PINs and the TRD of PINA on any interactions, and
expression based on magnICON® system. The aims have been addressed using the
methodologies explained in section 2.14.

Chapter 4 Protein interaction
- 127 -
4.2A Interaction studies for wild type PINA and PINB
4.2.1 Cloning of wild type PINA and PINB into pNBV vectors of the BiFC system
The ORF regions of the wild type alleles Pina-D1a (Genbank DQ363911), and Pinb-
D1a (Genbank DQ363913) encoding the mature PIN protein sections (lacking the N-
terminal signal peptide sequences) were amplified from the pICH-PINA-His and pICH-
PINB-His clones (detailed in Chapter 3). Gene-specific primers with the appropriate
restriction sites (Table 2.11) were used to introduce specific restriction sites at the 5’
and 3’ ends of the PCR fragments for cloning into the pNBV vectors (Section 2.14.2).
The start codon was added to all forward primers and the native stop codon was
removed for cloning into two pNBV vectors (gene-YN and gene-YC). The resulting
PCR fragments were 400 bp (Figure 4.1; lanes 1 and 2). These were double-digested
and cloned into four sets of double-digested pNBV vectors (gene-YN, gene-YC, YN-
gene and YC-gene), which allowed for fusions of each gene with the N-terminal 155
amino acids of YFP (the YN fragment) or the 84 amino acids of the C-terminal end of
YFP (the YC fragment). Empty pNBV vector DNAs were used to confirm the vector
size consistent with promoter, terminator and YFP fragments (~4 kb) (Figure 4.1; lanes
4 to 7). More details for pNBV vectors are shown in Figure 4.3.
Figure 4.1 Second-round PCR of gene sections encoding mature PINs and DNA plasmids of empty pNBV vectors. Lane M: DNA molecular weight marker, Hyperladder 1; lane 1: PINA (mature protein section of Pina-D1a with restriction sites); lane 2: PINB (mature protein section of Pinb-D1a with restriction sites); lane 3: negative control (no template); lane 4: DNA plasmid of pNBV vector (gene-YC); lane 5: DNA plasmid of pNBV vector (gene-YN); lane 6: DNA plasmid of pNBV vector (YN-gene); lane 7: DNA plasmid of pNBV (YC-gene).
M 1 2 3 4 5 6 7
4000bp
400bp
1000bp

Chapter 4 Protein interaction
- 128 -
The ligation mixtures of pNBV were transformed into E. coli Mach1 cells and grown on
LB plates with carbenicillin. Empty vectors were also transformed to be used as
controls. The presence of inserts was confirmed by colony-PCR (Figure 4.2) using
insert-specific primers (Table 2.11). Positive colonies (clones with inserts) were grown
in LB media containing carbenicillin. Plasmids were isolated from the positive clones
and used for PCR and double-digestion with restriction sites at the 5’ and 3’ ends of the
PINA and PINB fragments to confirm that the vectors contained the insert (data not
shown).
Figure 4.2 Colony PCR for preliminary selection of clones. Lane M: DNA molecular weight marker, Hyperladder 1; lane 1-3: pNBV-PINA product; lane 4 and 5: pNBV-PINB product; lane 6: negative control (no template). The above BiFC constructs were designated as pNBV-PINA-YN, pNBV-PINA-YC,
pNBV-YN-PINA and pNBV-YC-PINA for PINA and pNBV-PINB-YN, pNBV-PINB-
YC, pNBV-YN-PINB and pNBV-YC-PINB for PINB (Figure 4.3). They contain the
35S promoter of the cauliflower mosaic virus (862 bp), terminator of the Nos gene
(NosT: 264 bp), lacZ gene (215 bp), a linker between YN or YC and the gene (54 bp),
and yellow fluorescent protein (YFP) (YN=N-terminal fragment, amino acids 1 to 155
(465 bp), or YC=C-terminal fragment, amino acids 156 to 238 (249 bp)). The BiFC
cassettes in pNBV were released from pNBV by NotI digestion and inserted into the
plant expression vector (pMLBART; Figure 2.10). The full sequences of pNBV
cassettes without PINs are shown in Appendix IV.
M 1 2 3 4 5 6
600bp
400bp
1000bp

Chapter 4 Protein interaction
- 129 -
Figure 4.3 The pNBV module constructs encoding mature PINA and PINB proteins. 35S: promoter of the cauliflower mosaic virus; NosT: terminator of the Nos gene; YFP: yellow fluorescent protein (YN=N-terminal fragment 1 to 155 amino acid/YC=C-terminal fragment 156 to 238 amino acid). A1: pNBV-PINA-YN; A2: pNBV-PINA-YC; A3: pNBV-YN-PINA; A4: pNBV-YC-PINA; B1: pNBV-PINB-YN; B2: pNBV-PINB-YC; B3: pNBV-YN-PINB; B4: pNBV-YC-PINB.
LacZ PINA 35S NosT YFP-(YN)
NotI NotI XbaI XmaI
NosT 35S LacZ YFP-(YC)
NotI NotI XbaI XmaI
PINA
35S LacZ YFP-(YN)
NotI NotI
Linker
BamHI EcoRI
NosT PINA
35S LacZ YFP-(YC)
NotI NotI
Linker
BamHI EcoRI
NosT PINA
LacZ 35S NosT YFP-(YN)
NotI NotI XbaI XmaI
PINB
NosT 35S LacZ YFP-(YC)
NotI NotI XbaI XmaI
PINB
35S LacZ YFP-(YN)
NotI NotI
Linker
BamHI EcoRI
NosT PINB
35S LacZ YFP-(YC)
NotI NotI
Linker
BamHI EcoRI
NosT PINB
A1
A2
A3
A4
B2
B1
B4
B3

Chapter 4 Protein interaction
- 130 -
The constructs were transformed into E. coli strain cells and screened by colony PCR
with insert-specific primers (Table 2.11) followed by DNA sequencing. Alignment
with the reference sequences; Pina-D1a (DQ363911) (Figure 4.4) and Pinb-D1a
(DQ363913) (Figure 4.5) showed 100% identity.
Figure 4.4 Alignments sequence of PINA in pNBV construct. A: DNA sequence of mature putative Pina-D1a genes cloned in the pNBV (gene-YN) and pNBV (gene-YC) vectors. B: DNA sequence of mature putative Pina-D1a genes cloned in the pNBV (YN-gene) and pNBV (YC-gene) vectors. In both A and B, the line 1 is sequence of the reported WT Pina-D1a gene (DQ363911).
10 20 30 40 50 60. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ G A T G T T G C T G G C G G G G G T G G T G C T C A A C A A T G C C C T G T A G A G A C ApNBV-PINA-YN G G G G A C T C T A G A A T G G A T G T T G C T G G C G G G G G T G G T G C T C A A C A A T G C C C T G T A G A G A C ApNBV-PINA-YC ~ ~ ~ G A C T C T A G A A T G G A T G T T G C T G G C G G G G G T G G T G C T C A A C A A T G C C C T G T A G A G A C A
70 80 90 100 110 120. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature A A G C T A A A T T C A T G C A G G A A T T A C C T G C T A G A T C G A T G C T C A A C G A T G A A G G A T T T C C C GpNBV-PINA-YN A A G C T A A A T T C A T G C A G G A A T T A C C T G C T A G A T C G A T G C T C A A C G A T G A A G G A T T T C C C GpNBV-PINA-YC A A G C T A A A T T C A T G C A G G A A T T A C C T G C T A G A T C G A T G C T C A A C G A T G A A G G A T T T C C C G
130 140 150 160 170 180. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature G T C A C C T G G C G T T G G T G G A A A T G G T G G A A G G G A G G T T G T C A A G A G C T C C T T G G G G A G T G TpNBV-PINA-YN G T C A C C T G G C G T T G G T G G A A A T G G T G G A A G G G A G G T T G T C A A G A G C T C C T T G G G G A G T G TpNBV-PINA-YC G T C A C C T G G C G T T G G T G G A A A T G G T G G A A G G G A G G T T G T C A A G A G C T C C T T G G G G A G T G T
190 200 210 220 230 240. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature T G C A G T C G G C T C G G C C A A A T G C C A C C G C A A T G C C G C T G C A A C A T C A T C C A G G G G T C A A T CpNBV-PINA-YN T G C A G T C G G C T C G G C C A A A T G C C A C C G C A A T G C C G C T G C A A C A T C A T C C A G G G G T C A A T CpNBV-PINA-YC T G C A G T C G G C T C G G C C A A A T G C C A C C G C A A T G C C G C T G C A A C A T C A T C C A G G G G T C A A T C
250 260 270 280 290 300. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature C A A G G C G A T C T C G G T G G C A T C T T C G G A T T T C A G C G T G A T C G G G C A A G C A A A G T G A T A C A ApNBV-PINA-YN C A A G G C G A T C T C G G T G G C A T C T T C G G A T T T C A G C G T G A T C G G G C A A G C A A A G T G A T A C A ApNBV-PINA-YC C A A G G C G A T C T C G G T G G C A T C T T C G G A T T T C A G C G T G A T C G G G C A A G C A A A G T G A T A C A A
310 320 330 340 350 360. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature G A A G C C A A G A A C C T G C C G C C C A G G T G C A A C C A G G G C C C T C C C T G C A A C A T C C C C G G C A C TpNBV-PINA-YN G A A G C C A A G A A C C T G C C G C C C A G G T G C A A C C A G G G C C C T C C C T G C A A C A T C C C C G G C A C TpNBV-PINA-YC G A A G C C A A G A A C C T G C C G C C C A G G T G C A A C C A G G G C C C T C C C T G C A A C A T C C C C G G C A C T
370 380 390. . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature A T T G G C T A T T A C T G G T G A pNBV-PINA-YN A T T G G C T A T T A C T G G C C C G G G A T G G A G C A ApNBV-PINA-YC A T T G G C T A T T A C T G G C C C G G G A T G T A C
A
B

Chapter 4 Protein interaction
- 131 -
Figure 4.5 Alignments sequence of PINB in pNBV construct. A: DNA sequence of mature putative Pinb-D1a genes cloned in the pNBV (gene-YN) and pNBV (gene-YC) vectors. B: DNA sequence of mature putative Pinb-D1a genes cloned in the pNBV (YN-gene) and pNBV (YC-gene) vectors. In both A and B, the line 1 is sequence of the reported WT Pinb-D1a gene (DQ363913).
One each of the eight pMLBART constructs was individually transformed into the
Agrobacterium strain GV3101. Screening of the clones containing inserts was carried
out by colony-PCR using primer listed in Table 2.11 (data not shown).
10 20 30 40 50 60. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ G A A G T T G G C G G A G G A G G T G G T T C T C A A C A A T G T C C G C A G G A G C G GpNBV-PINB-YN ~ ~ ~ G A C T C T A G A A T G G A A G T T G G C G G A G G A G G T G G T T C T C A A C A A T G T C C G C A G G A G C G GpNBV-PINB-YC G G G G A C T C T A G A A T G G A A G T T G G C G G A G G A G G T G G T T C T C A A C A A T G T C C G C A G G A G C G G
70 80 90 100 110 120. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature C C G A A G C T A A G C T C T T G C A A G G A T T A C G T G A T G G A G C G A T G T T T C A C A A T G A A G G A T T T TpNBV-PINB-YN C C G A A G C T A A G C T C T T G C A A G G A T T A C G T G A T G G A G C G A T G T T T C A C A A T G A A G G A T T T TpNBV-PINB-YC C C G A A G C T A A G C T C T T G C A A G G A T T A C G T G A T G G A G C G A T G T T T C A C A A T G A A G G A T T T T
130 140 150 160 170 180. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature C C A G T C A C C T G G C C C A C A A A A T G G T G G A A G G G C G G C T G T G A G C A T G A G G T T C G G G A G A A GpNBV-PINB-YN C C A G T C A C C T G G C C C A C A A A A T G G T G G A A G G G C G G C T G T G A G C A T G A G G T T C G G G A G A A GpNBV-PINB-YC C C A G T C A C C T G G C C C A C A A A A T G G T G G A A G G G C G G C T G T G A G C A T G A G G T T C G G G A G A A G
190 200 210 220 230 240. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature T G C T G C A A G C A G C T G A G C C A G A T A G C A C C A C A A T G T C G C T G T G A T T C T A T C C G G C G A G T GpNBV-PINB-YN T G C T G C A A G C A G C T G A G C C A G A T A G C A C C A C A A T G T C G C T G T G A T T C T A T C C G G C G A G T GpNBV-PINB-YC T G C T G C A A G C A G C T G A G C C A G A T A G C A C C A C A A T G T C G C T G T G A T T C T A T C C G G C G A G T G
250 260 270 280 290 300. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature A T C C A A G G C A G G C T C G G T G G C T T C T T G G G C A T T T G G C G A G G T G A G G T A T T C A A A C A A C T TpNBV-PINB-YN A T C C A A G G C A G G C T C G G T G G C T T C T T G G G C A T T T G G C G A G G T G A G G T A T T C A A A C A A C T TpNBV-PINB-YC A T C C A A G G C A G G C T C G G T G G C T T C T T G G G C A T T T G G C G A G G T G A G G T A T T C A A A C A A C T T
310 320 330 340 350 360. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature C A G A G G G C C C A G A G C C T C C C C T C A A A G T G C A A C A T G G G C G C C G A C T G C A A G T T C C C T A G TpNBV-PINB-YN C A G A G G G C C C A G A G C C T C C C C T C A A A G T G C A A C A T G G G C G C C G A C T G C A A G T T C C C T A G TpNBV-PINB-YC C A G A G G G C C C A G A G C C T C C C C T C A A A G T G C A A C A T G G G C G C C G A C T G C A A G T T C C C T A G T
370 380. . . . | . . . . | . . . . | . . . . | . . . .
PINB-Mature G G C T A T T A C T G G T G A pNBV-PINB-YN G G C T A T T A C T G G C C C G G G A T G G A GpNBV-PINB-YC G G C T A T T A C T G G C C C G G G A T G
A
B

Chapter 4 Protein interaction
- 132 -
4.2.2 BiFC system for transient expression in N. bethamiana leaves
Agrobacterium harbouring BiFC constructs were infiltrated into eight week old N.
benthamiana leaves. The fluorescence started to appear 3-4 days post infiltration (dpi).
In general, the best observation time for BiFC fluorescence was 3 dpi, after which the
signal began to fade. The expression of the fusion proteins was observed using a highly
active promoter i.e., the CaMV 35S promoter. In order to ensure that the fluorescence
resulted from the specific PIN protein-protein interactions and not from non-specific
binding of the two YFP fragments, or from background or autofluorescence,
infiltrations with buffer only, or with just one PIN-YFP construct, or YN and YC
construct without PIN ligation, were used as negative controls. Any fluorescence
obtained with such constructs would reflect non-specific interactions of the expressed
proteins. In addition, non-treated leaves were used to detect background fluorescence.
4.2.3 Post-transcriptional gene silencing (PTGS) in transient system
N. benthamiana plants were transiently co-transformed with the desired construct and
p19. The p19 protein from tomato bushy stunt virus (TBSV) (Section 2.14.7), by co-
expression of a viral-encoded suppressor of gene silencing can prevent post-
transcriptional gene silencing (PTGS) in a transient assay system (Voinnet et al., 2003).
This strategy has been used in a number of transient assays to enhance transgene
expression (Grefen et al., 2008; Hellens et al., 2005; Wroblewski et al., 2005), including
BiFC analyses (Schütze et al., 2009; Walter et al., 2004; Waadt et al., 2008). To
determine if co-expression of BiFC constructs and the p19 could enhance PIN
expressions, an Agrobacterium LB4404 containing the p19 was co-infiltrated with BiFC
constructs into N. benthamiana leaves. This led to a strong increase in fluorescence
signal, as observed in individual cells. This improvement in signal and experimental
reliability was the reason for the p19 to be included in the subsequent transient BiFC
analyses.
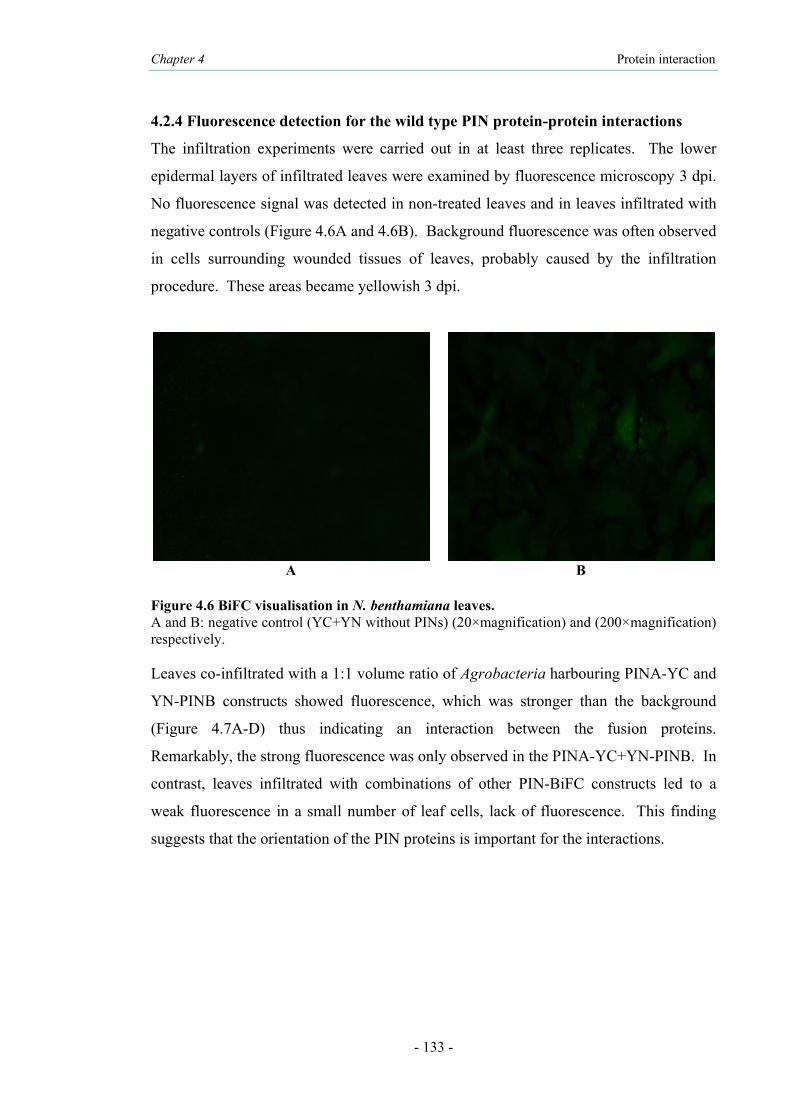
Chapter 4 Protein interaction
- 133 -
4.2.4 Fluorescence detection for the wild type PIN protein-protein interactions
The infiltration experiments were carried out in at least three replicates. The lower
epidermal layers of infiltrated leaves were examined by fluorescence microscopy 3 dpi.
No fluorescence signal was detected in non-treated leaves and in leaves infiltrated with
negative controls (Figure 4.6A and 4.6B). Background fluorescence was often observed
in cells surrounding wounded tissues of leaves, probably caused by the infiltration
procedure. These areas became yellowish 3 dpi.
A B
Figure 4.6 BiFC visualisation in N. benthamiana leaves. A and B: negative control (YC+YN without PINs) (20×magnification) and (200×magnification) respectively. Leaves co-infiltrated with a 1:1 volume ratio of Agrobacteria harbouring PINA-YC and
YN-PINB constructs showed fluorescence, which was stronger than the background
(Figure 4.7A-D) thus indicating an interaction between the fusion proteins.
Remarkably, the strong fluorescence was only observed in the PINA-YC+YN-PINB. In
contrast, leaves infiltrated with combinations of other PIN-BiFC constructs led to a
weak fluorescence in a small number of leaf cells, lack of fluorescence. This finding
suggests that the orientation of the PIN proteins is important for the interactions.

Chapter 4 Protein interaction
- 134 -
A B
C D
Figure 4.7 BiFC visualisation of PINA-PINB interactions in N. benthamiana leaves. A and B: interaction of PINA-PINB, in direction of PINA-YC+YN-PINB (20×magnification); C and D: interaction of PINA-PINB, in direction of PINA-YC+YN-PINB (200×magnification).
Since the structural basis of complex formations for PIN proteins is not clear, the two
interacting proteins were fused N- and C-terminally to the C- and N- terminal part of
YFP. Furthermore, potential homodimerisation was also investigated. Notably, the
fluorescence signals were observed in YN-PINB and YC-PINB constructs (Figure 4.8A
and 4.8B). However, no fluorescence was detected for homodimerisation in wild type
of PINA (Figure 4.8C and 4.8D) tested in different orientations (Table 4.1). From
images of fluorescence microscopy it seemed that PINA and PINB most likely
presented interactions in the cytoplasm. The small fluorescence dots illustrated likely
location in the chloroplast, and the small rings as Golgi compartments, for PINA-PINB
(Figure 4.7A-D) and especially for PINB-PINB (Figure 4.8A and B) interactions.

Chapter 4 Protein interaction
- 135 -
A B
C D
Figure 4.8 BiFC visualisation of PINA-PINA and PINB-PINB interactions in N. benthamiana leaves. A: interaction of PINB-PINB, in direction of YN-PINB+YC-PINB (20× magnification); B: interaction of PINB-PINB, in direction of YN-PINB+YC-PINB (200× magnification); C: interaction of PINA-PINA (20×magnification); D: interaction of PINA-PINA (200× magnification).
4.2.5 Western blot analysis of wild type PIN protein-protein interactions
Western blot analysis using the Durotest antibody was used to verify the co-expression
of the interacting forms of PINs. The leaves were harvested after 3 dpi and used for
microscopy. The total soluble protein (TSP) was extracted from leaves with the
fluorescence signal. The amounts of TSP were quantified using Bradford assay and
about 40 µg of TSP were loaded on 15% gel for further analysis by western blot. One
intense band at approximately 45 kDa was detected by western blot from the TSP of
infiltrated leaf with the PINA-YC+YN-PINB constructs (Figure 4.9; lanes 4 and 5).

Chapter 4 Protein interaction
- 136 -
The western blot also detected YN-PINB+YC-PINB interacting forms (Figure 4.9; lane
2); however the band size appeared to be slightly higher than PINA+PINB interactions.
Figure 4.9 Determination of interacting form of PINA+PINB and PINB+PINB by western blot analysis from TSP extract. Lane 1: YFP construct without PIN (negative control); lane 2: interacting form of recombinant PINB and PINB (YN-PINB+YC-PINB); lane 3: void lane (no sample loaded); lanes 4 and 5: interacting form of recombinant PINA and PINB (PINA-YC+YN-PINB); lane 6: 3% non-durum wheat as positive for antibody.
4.2.6 Gel detection of YFP fluorescence for wild type PIN protein-protein
interactions
In order to confirm the protein complexes, native electrophoresis was used on the same
samples as those used for western blot, for subsequent imaging of in-gel YFP
fluorescent under UV light. Fluorescent bands were detected for PINA-YC+YN-PINB
and for YN-PINB+YC-PINB interactions (Figure 4.10; lanes 1 and 2) which was also
confirming the above results of fluorescent microscopy and western blot. No bands
were detected for the negative controls (lanes 3 and 4).
1 2 3 4 5 6
50kDa
37kDa
75kDa
15kDa

Chapter 4 Protein interaction
- 137 -
Figure 4.10 Native gel electrophoresis and UV imaging of YFP fluorescence of PIN protein complexes. Lane 1: interacting form of recombinant PINA and PINB (PINA-YC+YN-PINB); lane 2: interacting form of recombinant PINB and PINB (YN-PINB+YC-PINB); lanes 3 and 4: YFP construct without PIN (negative).
4.2B Interaction studies of mutagenised PINA and PINB
4.2.7 Cloning of the previously made constructs PINA∆HD and PINB∆HD into
pNBV vectors
Site-Directed Mutagenesis (SDM) PCR had been earlier carried out by Dr R. Alfred
(Swinburne University of Technology) to delete the predicted hydrophobic domain
(HD) at positions 75-85 in the putative mature PINA (PINA∆HD) and at positions 75-
83 in the putative mature PINB (PINB∆HD) in yeast 2-hybrid system vectors pGBK
(details in section 2.15.1). These clones kindly provided by Dr R. Alfred, were used in
the present work as templates for PCR (Figure 4.11) to clone into pNBV vectors, so as
to see whether these deletions significantly affect the likely PPI of PINs. The primers
PIN-Forward and PIN-Reverse (Section 2.14.3) were used to introduce specific
restriction sites (XbaI and XmaI for PINA∆HD and EcoRI and BamHI for PINB∆HD)
at the 5’ and 3’ ends of the PCR fragments. PCR products of ~400 bp were obtained for
PINA∆HD and PINB∆HD (Figure 4.11), consistent with sizes of the gene sections
encoding putative mature PINA without HD (330 bp) or PINB without HD (333 bp) and
the added specific restriction sites (12 bp).
1 2 3 4

Chapter 4 Protein interaction
- 138 -
Figure 4.11 Amplification of inserts from PINA∆HD and PINB∆HD constructs. PCR products from clones in pGBK vectors using insert-specific primers. Lane 1: DNA molecular weight marker, Hyperladder l; lanes 2 and 3: PINA∆HD; lanes 4 and 5: PINB∆HD.
The PCR fragments were digested at restriction sites and cloned into cloning site of
pNBV vector to allow for translational fusions with the YN or YC fragments of YFP, as
above. The BiFC constructs were designated: pNBV-PINA∆HD-YC, pNBV-YN-
PINB∆HD and pNBV-YC-PINB∆HD (Figure 4.12). The cassettes in pNBV were
released from pNBV by NotI digestion, inserted into pMLBART transformed into E.
coli cells and screened by colony PCR with insert-specific primers (Table 2.11)
followed by DNA sequencing. Alignment with the reference sequences; Pina-D1a
(DQ363911) and Pinb-D1a (DQ363913) that showed 100% identity with the
corresponding sections (Figures 4.13 and 4.14).

Chapter 4 Protein interaction
- 139 -
Figure 4.12 The pNBV module constructs generated encoding mutant PIN proteins. 35S: promoter of the cauliflower mosaic virus; NosT: terminator of the Nos gene; YFP: yellow fluorescent protein (YN=N-terminal fragment 1 to 155 amino acid/YC=C-terminal fragment 156 to 238 amino acid). A: pNBV-PINA∆HD-YC; B: pNBV-YN-PINB∆HD; C: pNBV-YC-PINB∆HD.
Figure 4.13 Sequences of PINA∆HD in pNBV construct. DNA sequences of the mutated Pina-D1a genes cloned in pNBV vector. The top line (PINA) is the deduced protein sequence of the reported WT Pina-D1a (Genbank ABD72477, mature protein), line 2 is the DNA sequence of the reported Pina-D1a gene (DQ363911) and line 3 is the DNA sequence of PINA∆HD in pNBV (gene-YC) vector. The HD (IQGDLGGIFGF) is underlined.
NosT 35S LacZ YFP-(YC)
NotI NotI XbaI XmaI
PINA∆HD
35S LacZ YFP-(YN)
NotI NotI
Linker
BamHI EcoRI
NosT PINB∆HD
35S LacZ YFP-(YC)
NotI NotI
Linker
BamHI EcoRI
NosT PINB∆HD
A
B
C

Chapter 4 Protein interaction
- 140 -
Figure 4.14 Sequences of PINB∆HD in pNBV construct. DNA sequences of the mutated Pinb-D1a genes cloned in pNBV vector. The top line (PINB) is the deduced protein sequence of the reported WT Pinb-D1a (Genbank ABD72479, mature protein), line 2 is the DNA sequence of the reported Pinb-D1a gene (DQ363913), line 3 is the DNA sequence of PINB∆HD in pNBV-YN-gene vector, line 4 is the DNA sequence of PINB∆HD in pNBV-YC-gene. The HD (VIQGRLGGF) is underlined. Each pMLBART construct was transformed into the Agrobacterium strain GV3101.
Screening of the clones containing inserts was confirmed by antibiotic (spectinomycin
plus rifampicin) selection and colony PCR using primer listed in Table 2.11 (data not
shown).

Chapter 4 Protein interaction
- 141 -
4.2.8 Detection of mutant PINA∆HD and PINB∆HD protein-protein interactions
by fluorescence and western blot
The potential role of the hydrophobic domain (HD) in any PPI of PINA and PINB was
investigated through infiltration of the above constructs into N. benthamiana leaves side
by side on the same leaf (Figure 4.15), with the wild type PIN construct which had
shown the fluorescent signal for interactions (Figure 4.7). Since the PINs had shown
fluorescent signal in one orientation (PINA-YC+YN-PINB), the mutant PINs were
tested as same orientation, i.e., PINA∆HD-YC+YN-PINB∆HD. However the results
showed no fluorescence. The interaction between PINA∆HD and wild type PINB
(PINA∆HD-YC+YN-PINB) and also wild type PINA and PINB∆HD (PINA-YC+YN-
PINB∆HD) led to no fluorescence.
Figure 4.15 Infiltrated mixtures of constructs into N. benthamiana leaves side by side for detect and compare the interaction of wild type and mutant form of PIN proteins.
Further, TSP were extracted from infiltrated areas, quantified using Bradford assay and
about 40 µg of TSP electrophoresed. Western blot analysis resulted in very faint bands
at ~45 kDa (Figure 4.16; lanes 2, 3, 4) compared to the wild type controls (lanes 5, 7).
The results suggest that the HD deletion in the direction led to significant reduction in
interactions of PINA∆HD-YC with YN-PINB∆HD and also of the self-interacting
PINB (YN-PINB∆HD+YC-PINB∆HD) (Figure 4.16; lane 4).
Infiltrated area with PINA-YC
+ YN-PINB
Infiltrated area with PINA∆HD-YC
+ YN-PINB∆HD

Chapter 4 Protein interaction
- 142 -
Figure 4.16 Western blot analysis to determine the interacting forms of wild type and mutant PIN proteins. Lane 1: YFP construct without insert (negative control); lanes 2 and 3: interacting form of recombinant PINA∆HD-YC+YN-PINB∆HD; lane 4: interacting form of recombinant YN-PINB∆HD+YC-PINB∆HD; lane 5 and 6: interacting form of recombinant PINA-YC+YN-PINB; lane 7: interacting form of recombinant YN-PINB+YC-PINB.
4.2.9 Cloning of PINA∆TRD into pNBV vectors
PCR was also conducted by Dr R. Alfred to delete the TRD at positions 34-46 in the
mature PINA protein (PINA∆TRD) in the yeast 2-hybrid system vectors pGBK (details
in section 2.15.1). The clone (kindly donated by Dr R. Alfred) was also used in the
present work as template for PCR (data not shown) to clone in pNBV. The primers
PIN-Forward and PIN-Reverse (Section 2.14.3) were used to insert restriction sites
(XbaI and XmaI) at the 5’ and 3’ ends of the PCR fragment. The resulting PCR
fragments were ~400 bp for PINA∆TRD was consistent with the gene sections encoding
putative mature PINA without the TRD (324 bp). The PCR fragments were digested
and cloned into pNBV as above to obtain the construct pNBV-PINA∆TRD-YC with the
translational fusion with the YC fragment of YFP (Figure 4.17). The BiFC cassette in
pNBV was released by NotI digestion, inserted into pMLBART transformed into E. coli
which cells and screened by colony PCR with insert-specific primers (Table 2.11),
followed by DNA sequencing. Alignment with Pina-D1a (DQ363911) showed 100%
identity (Figure 4.18). The pMLBART construct was then transformed into the
Agrobacterium strain GV3101.
1 2 3 4 5 6 7
50kDa
75kDa
37kDa
15kDa

Chapter 4 Protein interaction
- 143 -
Figure 4.17 The pNBV module construct generated encoding PINA∆TRD protein. 35S: promoter of the cauliflower mosaic virus; NosT: terminator of the Nos gene; YFP: yellow fluorescent protein (YC=C-terminal fragment 156 to 238 amino acid); pNBV-PINA∆TRD-YC.
Figure 4.18 Sequences of PINA∆TRD in pNBV construct. DNA sequences of the mutated Pina-D1a genes cloned in pNBV vector. The top line (PINA) is the deduced protein sequence of the reported WT Pina-D1a (Genbank ABD72477, mature protein), line 2 is the DNA sequence of the reported Pina-D1a gene (DQ363911), and line 3 is the DNA sequence of PINA∆TRD in pNBV (gene-YC) vector. The TRD (FPVTWRWWKWWKG) is underlined.
NosT 35S LacZ YFP-(YC)
NotI NotI XbaI XmaI
PINA∆TRD

Chapter 4 Protein interaction
- 144 -
4.2.10 Fluorescence detection for mutant PINA∆TRD protein-protein interactions
Infiltration of the relevant constructs into N. benthamiana leaves showed no
fluorescence or only background fluorescence for PINA∆TRD-YC+YN-PINB. Further,
no fluorescence signal was detected for interaction between PINA∆TRD-YC+YN-
PINB∆HD. These results suggest that the TRD deletion in PINA leads to significant
change in the interactions of PIN. The full set of PIN interactions tested by the BiFC
assay is shown in Table 4.1.
Table 4.1 BiFC constructs for PIN proteins (wild type and mutant) to detect fluorescence signal
PINA∆TRD=Deletion of the tryptophan-rich domain (TRD), location at 34-46 amino acid in PINA PINA∆HD=Deletion of a hydrophobic domain (HD), location at 75-85 amino acid in PINA PINB∆HD=Deletion of a hydrophobic domain (HD), location at 75-85 amino acid in PINA
No Construct 1 Construct 2 Heterodimerisation
wild type PIN proteins Fluorescence
signal 1 PINA-YC YN-PINB PINA+PINB Yes 2 PINA-YC PINB-YN PINA+PINB No 3 PINA-YN YC-PINB PINA+PINB No 4 PINA-YN PINB-YC PINA+PINB No 5 YC-PINA PINB-YN PINA+PINB No 6 YC-PINA YN-PINB PINA+PINB No 7 YN-PINA PINB-YC PINA+PINB No 8 YN-PINA YC-PINB PINA+PINB No
No Construct 1 Construct 2 Homodimerisation
wild type PINA proteins Fluorescence
signal 1 YN-PINA PINA-YC PINA+PINA No 2 YN-PINA YC-PINA PINA+PINA No 3 PINA-YN YC-PINA PINA+PINA No 4 PINA-YN PINA-YC PINA+PINA No
No Construct 1 Construct 2 Homodimerisation
wild type PINB proteins Fluorescence
signal 1 YN-PINB PINB-YC PINB+PINB No 2 YN-PINB YC-PINB PINB+PINB Yes 3 PINB-YN YC-PINB PINB+PINB No 4 PINB-YN PINB-YC PINB+PINB No
No Construct 1 Construct 2 Heterodimerisation
of mutant PIN proteins Fluorescence
signal 1 PINA∆HD-YC YN-PINB∆HD PINA∆HD+PINB∆HD No 2 PINA∆HD-YC YN-PINB PINA∆HD+PINB No 3 PINA-YC YN-PINB∆HD PINA+PINB∆HD No 4 PINA∆TRD-YC YN-PINB∆HD PINA∆TRD+PINB∆HD No 5 PINA∆TRD-YC YN-PINB PINA∆TRD+PINB No
No Construct 1 Construct 2 Homodimerisation
of mutant PINB proteins Fluorescence
signal 1 YN-PINB YC-PINB∆HD PINB+PINB∆HD No 2 YN-PINB∆HD YC-PINB∆HD PINB∆HD+PINB∆HD No

Chapter 4 Protein interaction
- 145 -
4.2.11 Expression of mutant PIN proteins in magnICON® system
To determine the roles of the HD in both PINA and PINB and the TRD in PINA on
expression of PIN proteins in planta, the magnICON® system was also used. The 3’
modules pICH11599 containing PINA∆HD, PINB∆HD and PINA∆TRD were
constructed as detailed in Chapter 2 (Figure 4.19). This was followed by transformation
of the plasmids into E. coli Mach1 cells and purification of plasmids. Sequencing
confirmed the correct deletions and 100% identity of the rest of the sequence with the
wild type alleles (Figures 4.20 and 4.21).
Figure 4.19 The 3’ module constructs encoding mutant PIN proteins with His-tag. A: pICH-PINA∆TRD-His; B: pICH-PINA∆HD-His C: pICH-PINB∆HD-His
His Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB EcoRI BamHI
PINA∆TRD
His Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB EcoRI BamHI
PINA∆HD
His PINB∆HD Tnos NPT Pnos Intron 3’NTR Tnos AttB
RB LB NcoI SacI
A
B
C

Chapter 4 Protein interaction
- 146 -
Figure 4.20 Sequences of PINA∆HD-His and PINA∆TRD-His in the 3’ module pICH11599. The top line (PINA) is the deduced protein sequence of the reported WT Pina-D1a (Genbank ABD72477, mature protein), line 2 is the DNA sequence of the reported Pina-D1a gene (DQ363911), line 3 is the DNA sequence of PINA∆HD-His, line 4 is the DNA sequence of PINA∆TRD-His. The HD (VIQGRLGGF) and TRD (FPVTWRWWKWWKG) are underlined. His-tag sequences are in box.

Chapter 4 Protein interaction
- 147 -
Figure 4.21 Sequences of PINB∆HD-His in 3’ module pICH11599. The top line (PINB) is the deduced protein sequence of the reported WT Pinb-D1a (Genbank ABD72479, mature protein), line 2 is the DNA sequence of the reported Pinb-D1a gene (DQ363913), line 3 is the DNA sequence of PINB∆HD-His. The HD (VIQGRLGGF) is underlined. His-tag sequence is in box. The plasmids were electroporated into Agrobacterium GV3101 and infiltrated in N.
benthamiana leaves side by side with wild type of PINA and PINB. Approximately
~40 µg of TSP was loaded in each lane, followed by western blot. Interestingly, bands
were detected at ~45 kDa for expressions of chloroplast targeted PINA∆HD and
PINB∆HD, suggesting aggregate forms (Figure 4.22; lanes 1 and 2) similar to wild
type PINA and PINB (lanes 3 and 4), but PINA∆TRD was not detected (lane 5). The
results suggest that PINA with deletion of TRD may be degraded, or the Durotest
antibody was not able to detect it. The cytosol targeted PINs indicated the same result
(data not shown).

Chapter 4 Protein interaction
- 148 -
Figure 4.22 Determination of wild type and mutant PIN proteins expressions in infiltrated leaves with chloroplast viral vector module by western blotting of TSP extract. Lane1: chloroplast targeted PINA∆HD; lane 2: chloroplast targeted PINB∆HD; lane 3: chloroplast targeted PINA; lane 4: chloroplast targeted PINB; lane 5: chloroplast targeted PINA∆TRD; lane 6: negative control (5’+integrase modules only).
1 2 3 4 5 6
50kDa
37kDa
15kDa

Chapter 4 Protein interaction
- 149 -
4.2.12 Bioinformatics analysis of PIN sequences for potential interaction domain(s)
In order to gain more insights into any particular residues/domains that could be
involved in the interaction of PIN proteins, predictive modelling was conducted using I-
TASSER (Section 2.10.5) and the outputs analysed in PyMOL for viewing the 3D
structure and predicted functional domains such as the TRD and HD of the PINA and
PINB. Modelling of wild type PINA (Figure 4.23) showed very similar structures to
that predicted by Lesage et al. (2011) (Figure 1.5). The model showed homology to the
2S albumin storage protein and exhibited four alpha helices, with the TRD located as an
extended loop between α-helices 1 and 2, as reported previously by Marion et al.
(1994). The five tryptophan residues (W) in PINA and three in PINB with their indole-
ring are shown in blue (Figures 4.23 and 4.24).
An earlier prediction by using Kyte-Doolittle Hydropathy plot had shown strong
hydrophobic properties for the sequence at position 75-80 in the mature PINA protein,
this distinct hydrophobic domain (HD) formed a separate loop between α-helices 3 and
4 (Alfred, 2013). These HDs for PINA and PINB are shown in purple (Figures 4.23 and
4.24) and the phenylalanine residues (F) are marked. The C-score is a confidence score
for estimating the quality of predicted models by I-TASSER (Roy et al., 2010; Zhang,
2008) (Section 2.10.5). The calculated C-score was 0.28 for PINA and 0.37 for PINB,
signifying a model with a high confidence.

Sequence
20 40 60 80 100 120
| | | | | |
DVAGGGGAQQCPVETKLNSCRNYLLDRCSTMKDFPVTWRWWKWWKGGCQELLGECCSRLGQMPPQCRCNIIQGSIQGDLGGIFGFQRDRASKVIQEAKNLPPRCNQGPPCNIPGTIGYYW
Prediction CCCCCCCCCCCCHHHHHCHHHHHHHHHCCCCCCCCCCCCCCCCCCCCHHHHHHHHHHHHHHCCHHHHHHHHHHHHHHHHHHHHCCCCCHHHHHHHHHHCCCHCCCCCCCCCCCCCCCCCC
Conf.Score 987766545883666650799999996743113653334511253416999999999888388877359999999999987750140079999999865621029998887788777779
Figure 4.23 Structure prediction using PyMOL and prediction secondary structure for PINA. Prediction line, elements are shown as H for Alpha helix and C for Coil) using I-TASSER for PINA. Structure displaying TRD (blue) and HD (purple) domains are enclosed.
W
W W
W
W
F

Sequence
20 40 60 80 100
| | | | |
EVGGGGGSQQCPQERPKLSSCKDYVMERCFTMKDFPVTWPTKWWKGGCEHEVREKCCKQLSQIAPQCRCDSIRRVIQGRLGGFLGIWRGEVFKQLQRAQSLPSKCNMGADCKFPSGYYW
Prediction CCCCCCCCCCCCHHCHCCCHHHHHHHHHCCCCCCCCCCHHHHHHHCCCHHHHHHHHHHHHHHCCHHHCHHHHHHHHHHHHHHHCCCCCHHHHHHHHHHHCCCCCCCCCCCCCCCCCCCC
Conf.Score 98776743248053010038999999977433357510347874152179999999999985766346299999999998735054303899999998647510299999988888779
Figure 4.24 Structure prediction using PyMOL and prediction secondary structure for PINB. Prediction line, elements are shown as H for Alpha helix and C for Coil) using I-TASSER for PINB. Structure displaying TRD (blue) and HD (purple) domains are enclosed.
W
W W
F

Chapter 4 Protein interaction
- 152 -
The COTH (Threading-recombination approach) software (Section 2.10.6) was also
used to predict the possible structure of PINA and PINB protein-protein complex. The
quality of the complex template was detected by TM-score, alignment coverage, and
complex root mean-square deviation (Mukherjee and Zhang, 2011). TM-score was
used for quality assessment of PIN structure predictions, since it has ability in
combining alignment accuracy and coverage. First rank of the template for PIN
proteins was selected when TM-score was determined as individual protein; templates
>0.5 but the complex TM-score was <0.4. The prediction complex model for wild type
PINA and PINB with coverage of alignment, 0.816 is given in Figure 4.25. These
models were determined also for PINA∆HD and PINB∆HD with coverage of
alignment, 0.795 (Figure 4.26). Interestingly, coverage of the alignment which is based
on dividing the length of the alignment by the total length of the query complex is
higher for interaction of wild type PINs than for mutant proteins. The top models
predicted by COTH for interaction of PINA∆HD+PINB∆HD showed change in protein
folding that can lead to aggregation. PIN proteins without HD were folded somehow
differently and the TRD locations in both PIN proteins were changed (Figure 4.26).
The complex structure when the HD was removed exhibited a change in position of the
two tryptophan residues (W40 and W41) (shown in blue) in PINA and proline and
valine residues (P36 and V37) (shown in blue) in PINB. The change in position of the
TRD in predicted PINA∆HD and PINB∆HD complexes may explain the results of lack
of protein-protein interactions in plant cells when the HD is removed. The result shows
that both the TRD and HD regions may have roles in the interactions of PIN proteins,
but may contribute in different ways to the stability of the protein complexes.

DVAGGGGAQQCPVETKLNSCRNYLLDRCSTMKDFPVTWRWWKWWKGGCQELLGECCSRLGQMPPQCRCNIIQGSIQGDLGGIFGFQRDRASKVIQEAKNLPPRCNQGPPCNIPGTIGYYWEVGGGGGSQQCPQERPKLSSCKDYVMERCFTMKDFPVTWPTKWWKGGCEHEVREKCCKQLSQIAPQCRCDSIRRVIQGRLGGFLGIWRGEVFKQLQRAQSLPSKCNMGADCKFPSGYYW Figure 4.25 Modeling result of COTH for PINA and PINB Complex in horizontal and vertical direction. Wild type PINA (green) and PINB (red) complex; two residues of TRD domain for both proteins are positioned in blue; HD domains for both PINA and PINB proteins are bordered of protein sequences.
W
W
P
V

DVAGGGGAQQCPVETKLNSCRNYLLDRCSTMKDFPVTWRWWKWWKGGCQELLGECCSRLGQMPPQCRCNIIQGSQRDRASKVIQEAKNLPPRCNQGPPCNIPGTIGYYWEVGGGGGSQQCPQERPKLSSCKDYVMERCFTMKDFPVTWPTKWWKGGCEHEVREKCCKQLSQIAPQCRCDSIRRLGIWRGEVFKQLQRAQSLPSKCNMGADCKFPSGYYW Figure 4.26 Modeling result of COTH for PINA∆HD and PINB∆HD Complex. HD deletion mutation PINA∆HD (green) and PINB∆HD (red) complex; two residues of TRD domain are positioned in blue
W W
P
V

Chapter 4 Protein interaction
- 155 -
4.3 Discussion
Previously PIN proteins have been expressed successfully in maize with the 27 kDa γ-
Zein promoter and the 35S polyadenylation region terminator (Zhang et al., 2009).
RNA-mediated silencing (RNAi) of the Pin genes in transgenic wheat reported with
stable transformation via A. tumefaciens under control of 35S promoter (Gasparis et al.,
2011). Moreover recently overexpression of Pina gene in transgenic durum wheat was
conducted under the control of the maize Ubiquitin promoter and also under the control
of 35S promoter (Li et al., 2014). In the present work, the PIN proteins for BiFC assay
have been expressed successfully under the control of 35S promoter.
Protein-protein interaction is one of the most important considerations for the function
of many proteins. The present work is the first investigation of in vivo interaction for
PIN proteins based on the BiFC assay in a plant system. Defining protein interactions
can be assisted by BiFC assays in live plant cells such as N. benthamiana (Goodin et al.,
2008; Ohad et al., 2007; Waadt et al., 2008). This approach is based on the
complementation between two non-fluorescent fragments of the yellow fluorescent
protein (YFP) when they are brought together by interactions between proteins fused to
each fragment resulting in fluorescence (Hu and Kerppola, 2003; Kerppola, 2006).
Since the induction of the fluorescence signal is dependent on the physical association
between the two terminal parts of YFP, it is dependent not only on their presence but
also their orientation. Overall, when the structural basis of a protein complex is not
clear, as in the present case, the two interacting proteins should be fused N- and also C-
terminally to the C- and N- terminal parts of YFP and all eight combinations tested for
fluorescence complementation (Kerppola, 2006). Table 4.1, shows interactions results
in different directions for heterodimersations and homodimersations, for both PINA and
PINB. Strong interactions were observed only in one direction: PINA-YC+YN-PINB,
i.e., co-expression of the C-terminal half of YFP fused to PINA and the N-terminal half
of YFP fused to PINB. The finding suggests that the orientation of the PIN proteins is
important for their interactions. Previous work by Ziemann et al. (2008) using the yeast
two-hybrid system (Fields and Song, 1989) showed physical interactions between wild
type of PINA and PINB, and also PINB interacting with itself while PINA showed
weaker self interaction. Further, the important role of direction for PIN proteins in the

Chapter 4 Protein interaction
- 156 -
yeast two-hybrid system confirmed in studies of wild type as well as the mutated (TRD
and HD domain-deleted) proteins (Alfred, 2013; Alfred et al., 2014).
Using BiFC, leaf cells of N. benthamiana showed fluorescence signal due to PINB
homo-dimerisation (YN-PINB+YC-PINB). The presence of three tryptophan residues
in the TRD of PINB may explain the result. However, most interestingly, no
interactions were detected for PINA homo-dimerisation, suggesting the domain alone or
its tryptophans (five in PINA) may not be directly relevant to the interactions.
Fluorescence complementation can occur when the YFP fragments are fused to
positions that are separated by an average distance of greater than 100A°, which
provides enough flexibility to allow reconstitution of the split-YFP fragments (Hu et al.,
2002). The lack of fluorescence of PINA homo-dimerisation may result from
insufficient flexibility compared to that of PINB complexes.
The region or amino acid residues involved in the PIN proteins interactions still remains
elusive. The TRD is the most important region for lipid interactions (Kooijman et al.,
1997; Kooijman et al., 1998; Jing et al., 2003), therefore one strong candidate is the
TRD in both proteins, at residues 34-46 in mature PINA and 35-47 in mature PINB.
The other candidate is the little-studied hydrophobic domain (HD) at position of residue
75-85 of the mature PINA (Le Bihan et al., 1996), and the corresponding residues 75-83
of the mature PINB. Clones created previously (Alfred, 2013; Alfred et al., 2014) by
site-directed mutagenesis (SDM) PCR to delete the TRD within PINA and the HD
within PINA and PINB were therefore used in the present work. The PINA∆HD
interactions with PINB∆HD showed no fluorescence signal based on BiFC assays, in
contrast to interaction of a specific orientation of the wild type PINA and PINB. The
interactions for PINA and PINB without HD tested in the same orientation were
unsuccessful in inducing fluorescence. Furthermore, no fluorescence signal was
observed with itself or between PINB∆HD and wild type PINB. The results indicate
that the HD domains are important for co-expression and/or stability and/or interaction
between PINA and PINB and among PINB and PINB in N. benthamiana leaf cells.
There is one possible explanation for the non-interaction of PINA∆HD+PINB∆HD;
aggregation of mutant proteins, due to the change in the protein folding.

Chapter 4 Protein interaction
- 157 -
Deletion of the HD may also have an effect on the refolding of the PINs. Dimerisation
and/or oligomerisations can have different structural and functional advantages for
proteins, such as improved stability, control over accessibility and specificity of active
sites, and increased complexity (Marianayagam et al., 2004). Protein complexes are
stabilized by the burial of hydrophobic complementary occluding surfaces, covalent
bonds such as disulphide bridges, and hydrogen bonding (Lesk, 2010). The disulphide
bonds are indeed necessary to stabilize the α-helical structure and maintain the native
structure and solubility of PINA and PINB (Le Bihan et al, 1996). The formation of α-
helices at membrane surfaces is also an important point for binding to membrane
(Seelig, 2004). The preliminary protein-protein complex predictions based on the
COTH have shown refolding and a shift in the rotation of structure of the mutant PINs,
which may affect their functionality.
No protein-protein interactions were noted for PINA∆TRD with wild type PINB. Thus
the TRD of PINA may be important for expression in planta and protein-protein
interactions. Previous studies have emphasized that the TRD region has important role
in binding to the membranes polar lipids on the surface of starch (Blochet et al., 1993;
Kooijman et al., 1997; Giroux and Morris, 1997), direct binding to surface of starch
granules (Wall et al., 2010) and the activity of PINs (Capparelli et al., 2005; Dubreil et
al., 1998; Jing et al., 2003). The TRD has also been shown to be important for the
antibacterial and antifungal activities of PINs (Jing et al., 2003; Miao et al., 2012;
Phillips et al., 2011). The self-assembly of PINA micelle in solution has been shown by
hydrophobic forces that involve intermolecular interactions between the tryptophan
residues in the TRD (Clifton et al., 2011a). However no such self-assembly has been
observed for PINB, possibly due to fewer hydrophobic tryptophan residues in its TRD
(Clifton et al., 2011a; Pauly et al., 2013). There is evidence that PINA forms
aggregates, and TRD has been implicated in the formation of PINA aggregates (Le
Bihan et al., 1996). Thus, PINA-PINB interaction may explain the reported change to
hard grain when there is a mutation or a lack in either PIN protein (Giroux and Morris,
1997, 1998). Transgenic wheat work has provided further insights into the roles of
PINA and PINB together and individually in influencing grain texture (Giroux and
Morris, 1998; Morris et al., 2001; Wanjugi et al., 2007) (reviewed in Section 1.5.4).

Chapter 4 Protein interaction
- 158 -
Two tryptophan residues (W41 and W44) at the TRD have shown essential role for
interaction of PINA with the yeast plasma membranes, while the lysine in the TRD of
PINB seems to be more important (Evrard et al., 2008). In planta mutagenesis studies
had found that mutations within the TRD of PINA and PINB resulted significantly
increased the grain hardness, and PINB function was likely to be more critical to overall
Ha locus effects (Feiz et al., 2009a). The TRD has a role in stabilizing the structure of
both PIN proteins (Pauly et al., 2013).
The present work has shown that the PINA and PINB proteins do interact physically in
planta, and PINB can also self-associate, indicating that its function may more
considerable despite having a TRD with fewer tryptophan residues. A more detailed
understanding of the mechanisms for protein-protein interaction between the PIN
proteins is required to explain many of their co-operative and inter-dependant properties
involved in starch binding and weather these are also relevant to their bioactivity against
microbial pathogen.

- 159 -
CHAPTER 5
General discussion and future directions

Chapter 5 General discussion and future directions
- 160 -
5.1 General discussion
Wheat is the third most widely consumed cereals in the world after maize and rice. The
quality of wheat is critical in the food industry owing to its nutritional value and health
benefits. The hardness or texture of the wheat grain is a major determinant of the
quality, as it affects milling and end use properties.
The first chapter of this thesis presented a comprehensive review of literature on
structure of the wheat grain, grain hardness and its effects on the end-use of wheat and
the discoveries of friabilin and puroindoline proteins. Friabilin is a 15 kDa protein
complex that consists of two structurally similar proteins, puroindoline-a (PINA) and
puroindoline-b (PINB). It was originally suggested that friabilin act as a non-stick
surface between the starch granules and the protein matrix to determine wheat grain
hardness. The PINs are unique small proteins with a highly conserved 10 cysteine-
backbone and a tryptophan-rich domain (TRD), the latter most likely involved in the
binding to the phospholipids on the surface of starch granules. The soft grain texture in
wheat encoded by the wild type Pin alleles (Pina-D1a and Pinb-D1a), while the hard
texture occurs as a result of mutation in one or both of the Pin genes, and deletions of
both genes result in the ‘very hard’ texture of durum wheat. Chapter 1 also described
evidence of transgenic Pin genes altering grain texture, as well as plant-based
expression systems for production of valuable recombinant proteins such as anti-
microbial peptides (AMPs). The review also presented several predicted homology
models of PIN proteins due to the lack of an absolute three dimensional structure.
Another limitation in structural analysis of PINs is their strong aggregation properties at
high protein concentration. Several other gaps in literature were also identified, such as
the in vivo roles of PIN proteins within the wheat seed and the exact mode of actions of
PINs in grain hardness and their co-operative activity at membranes. Hence,
characterisation of PIN proteins is critical to exploit the potential benefits of PIN
proteins and PIN-based peptides in agricultural or medical applications. Therefore, the
major objectives of this research were as follows:

Chapter 5 General discussion and future directions
- 161 -
1. To clone the Pin genes encoding the putative mature PINA and PINB in magnICON®
viral vector system and A. tumefaciens infiltrations, to express and optimize their
transient expression for high yield in the leaves of Nicotiana benthamiana.
2. To test the antibacterial and antifungal activities of recombinant PIN proteins purified
with affinity purification using His-tag and hydrophobic interactions systems.
3. To analyse the protein-protein interactions among the PIN proteins in planta using
the BiFC system based on pNBV vectors and co-expression of the silencing suppressor
protein p19.
4. To test the significance of the tryptophan-rich domain (TRD) of PINA and the
hydrophobic domain (HD) of both PINs in any interactions between the PINs using the
BiFC system.
The above aims were addressed using a number of experimental approaches as set out in
Chapter 2. In brief, these included molecular and biochemical methodologies such as,
DNA extractions, PCRs, cloning into viral vectors, plasmid purifications, DNA
sequencing, electroporation of A. tumefaciens cells, agroinfiltration, in planta protein
expression, protein purification and quantitation, SDS-PAGE, western blot and ELISA
analyses, fluorescence microscopy, microbiological techniques and also bioinformatics
tools (e.g., DNA and protein sequence alignments, primer design, BLAST searches, 3D
modeling and protein-protein complex structure predications).
The aims 1 and 2 were addressed in Chapter 3. In brief, this chapter investigated the
plant expression system as a platform to produce recombinant PINA and PINB proteins
in sufficient yield for antimicrobial activities. The study focussed on the rapid transient
expression of PINs using the deconstructed tobacco mosaic virus-based ‘magnICON®’
for subcellular localisation of PINs in different compartments of N. benthamiana. The
results have shown: (i) the presence of recombinant PINA and PINB in both monomeric
and oligomeric forms when targeted to the chloroplast, cytosol and ER as shown by
western blot and ELISA analyses (ii) PIN A and PINB yields of up to 0.8% TSP or 51.7

Chapter 5 General discussion and future directions
- 162 -
µg/g of fresh weight for PINA, and 70 µg/g of fresh weight for PINB when proteins
targeted to the chloroplast (iii) the achievement of maximum yields of recombinant PIN
proteins at 10 dpi (iv) heat-sensitivity of recombinant PIN proteins (v) establishment of
the most efficient purification method for recombinant PINs based on hydrophobic
interaction (vi) antimicrobial activity for recombinant PINA and PINB against
Escherichia coli (a Gram negative bacterium) and Staphylococcus aureus (a Gram
positive bacterium) and four phytopathogenic fungal strains and low haemolytic activity
towards mammalian cells. Overall, the work has led to successful expression,
purification and characterisation methodology for obtaining PIN proteins with
appropriate quality that is essential for potential infection control and therapeutic
applications.
Aims 3 and 4 were addressed in Chapter 4. There is evidence that PINs may interact
co-operatively or interdependently to confer the soft grain texture. The present work
showed interactions between PINA and PINB proteins using BiFC system in N.
benthamiana leaves, just in one direction; PINA-YC+YN-PINB, based on the
fluorescence intensity visualised in living plant cells. The finding suggests that the
orientation of the PIN proteins is important for their interactions. In addition,
homodimerisation was noted for PINB (YN-PINB+YC-PINB) but not PINA. Moreover
the chapter investigated the key regions/residues essential for protein-protein
interaction. Site-directed mutagenesis (SDM) PCR had been conducted previously to
deletions in PINA at the TRD and in both PINs at the hydrophobic domain (HD). These
inserts were subcloned into the BiFC vectors in the present work. The results indicate
that no interaction occurred between PINA and PINB, or between PINB and PINB,
without their HD. The HD domain may affect the folding of PIN proteins and rotation
possibilities of the structure. Predictions of the 3D models of the possible structure for
PINs further corroborated this hypothesis. The deletion of the TRD in PINA also
showed no interaction with wild type PINB. Overall, the result shows that both the
TRD and HD regions may have roles in the in planta interactions of PIN proteins, but
the two domains may contribute in different ways to stability of the protein complexes.
The mutant PIN proteins were also expressed in planta based on magnICON® system to
test their expression. The result showed that PINs without the HD were successfully

Chapter 5 General discussion and future directions
- 163 -
expressed, while the PINA without TRD was not detected. The results suggest that
PINA with deletion of TRD may be degraded, or the Durotest antibody was not able to
detect this mutant protein, suggesting the TRD is among the epitopes for this antibody.
5.2 Future directions
The significance of the PIN proteins in the wheat industry has led to a considerable
amount of research focus on their structures and mechanisms. The worldwide research
has led to over 150 publications that outline their lipid-binding properties, the mutant
alleles as well as orthlogs in other monocot crop species, also the antimicrobial
properties of the proteins and the peptides based on these. However, the exact
mechanisms of action for PINs are not completely clear yet. The results of the present
study led to successful expression of bioactive proteins in a plant systems and also
establishe PINA-PINB and PINB-PINB associations in planta, as summarized above.
However further in-depth research is required on nature and mechanisms for their
functionality. The identity of the active protein can be established using peptide
mapping of the trypsinized protein, sizing using MALDI-TOF MS methods and identity
through LC–MS/MS (Klimyuk et al., 2014). Additionally, it would be important to use
circular dichroism (CD) tools (Greenfield, 2007) to determine the secondary structure
and folding properties of recombinant proteins to assess how the different mutations
affects their structure and stability.
Designing more strategies for increased production of soluble PIN proteins, such as
plant-codon optimisation would be important, as effected on increasing yield of protein
in plant system (Webster et al., 2009). The choice of expression systems such as cell-
free system (Endo and Sawasaki, 2006; Sawasaki et al., 2002), or vectors with fusion
tag, for instance, Halo-tag (Los et al., 2008) would be important. Furthermore, several
fusion strategies have been developed recently to significantly enhance the production
yield of plant-made recombinant proteins, as reviewed by Conley et al. (2011).
The exact mechanism of membrane activity for PIN-based peptides and proteins still is
unclear. Hence it would be useful to design chloroplast transformation vector with the
PIN-based peptide fused with GFP to confer stability to the peptide (Lee et al., 2011b).

Chapter 5 General discussion and future directions
- 164 -
PIN-based peptides can be obtained with the tag to help their purification for further
analysis of direct interactions with biological membranes. Expression and
characterisation of antimicrobial peptides in chloroplasts to control viral and bacterial
infections has been successfully utilised (Daniell et al., 2009a; Lee et al., 2011b; Nadai
et al., 2009; Ruhlman et al., 2007; Sanz-Barrio et al., 2011). Further work is needed to
explore these applications for PINs, especially for activity against pathogens of
relevance to the food and agriculture industries.
This study has shown protein-protein interaction of PIN proteins in planta. The
knowledge gained will be invaluable for future work for more detailed understanding of
the mechanisms interactions, which is required to explain their co-operative or
interdependent properties in starch binding. Site-directed mutagenesis of residues that
may have a role in the hardness phenotype would be important for exploring their
effects on protein interaction. Further, the GSP-1 and PINB-2v1 proteins may be
additional factors that interact with the PINs, contribute to further variation in wheat
grain hardness, and therefore would be important for exploring their effects on protein
interaction. Flow cytometry techniques (Berendzen et al., 2012; Morell et al., 2008) and
multicolour BiFC analysis may allows simultaneous visualisation of multiple protein
complexes in the same cell (Hu and Kerppola, 2003; Kerppola, 2008) and enable
subcellular localisation of the interaction (Kerppola, 2013). Furthermore, quantification
of BiFC fluorescence can be measured by analytical flow cytometry (Berendzen et al.,
2012).
The PIN proteins were discovered more than twenty years ago and numerous studies so
far have already revealed a number of their fundamental biochemical and functional
aspects. Further investigations building on this wealth of knowledge will one day
explain the unique activities and effects of these small proteins.

References
- 165 -
References
Adam Z, Clarke AK (2002) Cutting edge of chloroplast proteolysis. Trends in Plant Science, 7: 451-456.
Alfred RL, Palombo EA, Panozzo JF, Bhave M (2014) The co-operative interaction of puroindolines in wheat grain texture may involve the hydrophobic domain. Journal of Cereal Science, In Press.
Alfred RL (2013) Biochemical analysis of the functionality of puroindoline peptides and proteins. PhD Thesis, Swinburne University of Technology, http://hdl.handle.net/1959.3/368064.
Alfred RL, Palombo EA, Panozzo JF, Bariana H, Bhave M (2013a) Stability of puroindoline peptides and effects on wheat rust. World Journal of Microbiology and Biotechnology, 29: 1409-1419.
Alfred RL, Palombo EA, Panozzo JF, Bhave M (2013b) The antimicrobial domains of wheat puroindolines are cell-penetrating peptides with possible intracellular mechanisms of action. PLoS ONE, 8, e75488.
Amoroso MG, Longobardo L, Capparelli R (2004) Real Time RT-PCR and flow cytometry to investigate wheat kernel hardness: Role of puroindoline genes and proteins. Biotechnology Letters, 26: 1731-1737.
Arlen PA, Falconer R, Cherukumilli S, Cole A, Cole AM, Oishi KK, Daniell H (2007) Field production and functional evaluation of chloroplast-derived interferon-α2b. Plant Biotechnology Journal, 5: 511-525.
Arnau J, Lauritzen C, Petersen GE, Pedersen J (2006) Current strategies for the use of affinity tags and tag removal for the purification of recombinant proteins. Protein Expression and Purification, 48: 1-13.
Bakó A, Gárdonyi M, Tamás L (2007) In vitro starch binding experiments: Study of the proteins related to grain hardness of Wheat. In: Buck HT, Nisi JE and Salomón N (eds.) Wheat Production in Stressed Environments. Springer Netherlands. pp 685-691.

References
- 166 -
Bally J, Nadai M, Vitel M, Rolland A, Dumain R, Dubald M (2009) Plant physiological adaptations to the massive foreign protein synthesis occurring in recombinant chloroplasts. Plant Physiology, 150: 1474-1481.
Barampuram S, Zhang ZJ (2011) Recent advances in plant transformation. Methods in Molecular Biology (Clifton, N.J.), 701: 1-35.
Barlow K, Buttrose M, Simmonds D, Vesk M (1973) Nature of starch-protein interface in wheat endosperm. Cereal Chemistry, 50: 443-454.
Basaran P, Rodriguez-Cerezo E (2008) Plant molecular farming: Opportunities and challenges. Critical Reviews in Biotechnology, 28: 153-172.
Beecher B, Bettge A, Smidansky E, Giroux MJ (2002a) Expression of wild-type pinB sequence in transgenic wheat complements a hard phenotype. Theoretical and Applied Genetics, 105: 870-877.
Beecher B, Bowman J, Martin JM, Bettge AD, Morris CF, Blake TK, Giroux MJ (2002b) Hordoindolines are associated with a major endosperm-texture QTL in Barley (Hordeum vulgare). Genome, 45: 584-591.
Benchabane M, Goulet C, Rivard D, Faye L, Gomord V, Michaud D (2008) Preventing unintended proteolysis in plant protein biofactories. Plant Biotechnology Journal, 6: 633-648.
Benko-Iseppon AM, Galdino SL, Calsa Jr T, Kido EA, Tossi A, Belarmino LC, Crovella S (2010) Overview on plant antimicrobial peptides. Current Protein and Peptide Science, 11: 181-188.
Berendzen KW, Böhmer M, Wallmeroth N, Peter S, Vesić M, Zhou Y, Tiesler FKE, Schleifenbaum F, Harter K (2012) Screening for in planta protein-protein interactions combining bimolecular fluorescence complementation with flow cytometry. Plant Methods, 8: 25-40.
Bettge AD, Morris CF, Greenblatt GA (1995) Assessing genotypic softness in single wheat kernels using starch granule-associated friabilin as a biochemical marker. Euphytica, 86: 65-72.
Bhave M, Morris CF (2008a) Molecular genetics of puroindolines and related genes: Allelic diversity in wheat and other grasses. Plant Molecular Biology, 66: 205-219.

References
- 167 -
Bhave M, Morris CF (2008b) Molecular genetics of puroindolines and related genes: Regulation of expression, membrane binding properties and applications. Plant Molecular Biology, 66: 221-231.
Bi X, Wang C, Ma L, Sun Y, Shang D (2013) Investigation of the role of tryptophan residues in cationic antimicrobial peptides to determine the mechanism of antimicrobial action. Journal of Applied Microbiology, 115: 663-672.
Blochet JE, Chevalier C, Forest E, Pebay-Peyroula E, Gautier MF, Joudrier P, Pezolet M, Marion D (1993) Complete amino acid sequence of puroindoline, a new basic and cystine-rich protein with a unique tryptophan-rich domain, isolated from wheat endosperm by Triton x-114 phase partitioning. FEBS Letters, 329: 336-340.
Bracha-Drori K, Shichrur K, Katz A, Oliva M, Angelovici R, Yalovsky S, Ohad N (2004) Detection of protein-protein interactions in plants using bimolecular fluorescence complementation. The Plant Journal, 40: 419-427.
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72: 248-254.
Branlard G, Amiour N, Igrejas G, Gaborit T, Herbette S, Dardevet M, Marion D (2003) Diversity of puroindolines as revealed by two-dimensional electrophoresis. Proteomics, 3: 168-174.
Broekaert WF, Cammue BPA, De Bolle MFC, Thevissen K, De Samblanx GW, Osborn RW (1997) Antimicrobial peptides from plants. Critical Reviews in Plant Sciences, 16: 297-323.
Broekaert WF, Terras FRG, Cammue BPA, Vanderleyden J (1990) An automated quantitative assay for fungal growth inhibition. FEMS Microbiology Letters, 69: 55-60.
Bruce BD (2000) Chloroplast transit peptides: Structure, function and evolution. Trends in Cell Biology, 10: 440-447.
Capparelli R, Amoroso MG, Palumbo D, Iannaccone M, Faleri C, Cresti M (2005) Two plant puroindolines colocalize in wheat seed and in vitro synergistically fight against pathogens. Plant Molecular Biology, 58: 857-867.

References
- 168 -
Capparelli R, Borriello G, Giroux MJ, Amoroso MG (2003) Puroindoline A-gene expression is involved in association of puroindolines to starch. Theoretical and Applied Genetics, 107: 1463-1468.
Capparelli R, Palumbo D, Iannaccone M, Ventimiglia I, Di Salle E, Capuano F, Salvatore P, Amoroso M (2006) Cloning and expression of two plant proteins: similar antimicrobial activity of native and recombinant form. Biotechnology Letters, 28: 943-949.
Capparelli R, Ventimiglia I, Palumbo D, Nicodemo D, Salvatore P, Amoroso MG, Iannaccone M (2007) Expression of recombinant puroindolines for the treatment of staphylococcal skin infections (acne vulgaris). Journal of Biotechnology, 128: 606-614.
Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC (1994) Green fluorescent protein as a marker for gene expression. Science, 263: 802-805.
Chan DI, Prenner EJ, Vogel HJ (2006) Tryptophan- and arginine-rich antimicrobial peptides: Structures and mechanisms of action. Biochimica et Biophysica Acta - Biomembranes, 1758: 1184-1202.
Chang C, Zhang H, Xu J, Li W, Liu G, You M, Li B (2006) Identification of allelic variations of puroindoline genes controlling grain hardness in wheat using a modified denaturing PAGE. Euphytica, 152: 225-234.
Chantret N, Cenci A, Sabot F, Anderson O, Dubcovsky J (2004) Sequencing of the Triticum monococcum Hardness locus reveals good microcolinearity with rice. Molecular Genetics and Genomics, 271: 377-386.
Chantret N, Salse J, Sabot F, Rahman S, Bellec A, Laubin B, Dubois I, Dossat C, Sourdille P, Joudrier P, Gautier MF, Cattolico L, Beckert M, Aubourg S, Weissenbach J, Caboche M, Bernard M, Leroy P, Chalhoub B (2005) Molecular basis of evolutionary events that shaped the Hardness locus in diploid and polyploid wheat species (Triticum and Aegilops). The Plant Cell, 17: 1033-1045.
Charnet P, Molle G, Marion D, Rousset M, Lullien-Pellerin V (2003) Puroindolines form ion channels in biological membranes. Biophysical Journal, 84: 2416-2426.
Chase MW, Knapp S, Cox AV, Clarkson JJ, Butsko Y, Joseph J, Savolainen V, Parokonny AS (2003) Molecular systematics, GISH and the origin of hybrid taxa in Nicotiana (Solanaceae). Annals of Botany, 92: 107-127.

References
- 169 -
Chen F, He Z, Xia X, Lillemo M, Morris C (2005) A new puroindoline b mutation present in Chinese winter wheat cultivar Jingdong 11. Journal of Cereal Science, 42: 267-269.
Chen F, He ZH, Xia XC, Xia LQ, Zhang XY, Lillemo M, Morris CF (2006) Molecular and biochemical characterization of puroindoline a and b alleles in Chinese landraces and historical cultivars. Theoretical and Applied Genetics, 112: 400-409.
Chen F, Yu Y, Xia X, He Z (2007) Prevalence of a novel puroindoline b allele in Yunnan endemic wheats (Triticum aestivum ssp. yunnanense King). Euphytica, 156: 39-46.
Chen F, Beecher B, Morris C (2010a) Physical mapping and a new variant of Puroindoline b-2 genes in wheat. Theoretical and Applied Genetics, 120: 745-751.
Chen F, Xu HX, Zhang FY, Xia XC, He ZH, Wang DW, Dong ZD, Zhan KH, Cheng XY, Cui DQ (2010b) Physical mapping of puroindoline b-2 genes and molecular characterization of a novel variant in durum wheat (Triticum turgidum L.). Molecular Breeding, 28: 153-161.
Chen F, Zhang F, Cheng X, Morris C, Xu H, Dong Z, Zhan K, Cui D (2010c) Association of Puroindoline b-B2 variants with grain traits, yield components and flag leaf size in bread wheat (Triticum aestivum L.) varieties of the Yellow and Huai Valleys of China. Journal of Cereal Science, 52: 247-253.
Chevreau E, Dupuis F, Ortolan C, Parisi L (2001) Transformation of apple for durable scab resistance: Expression of a puroindoline gene in susceptible and resistant (Vf) genotypes. Acta Horticulturae, 560: 323-327.
Christou P, Stoger E, Twyman RM (2004) Monocot expression systems for molecular farming. In: Fischer R and Schillberg S (eds.) Molecular Farming. Wiley-VCH Verlag GmbH & Co. KGaA,Weinheim. pp 55-67.
Clifton LA, Green RJ, Frazier RA (2007a) Puroindoline-b mutations control the lipid binding interactions in mixed puroindoline-a:puroindoline-b systems. Biochemistry, 46: 13929-13937.
Clifton LA, Green RJ, Hughes AV, Frazier RA (2008) Interfacial structure of wild-type and mutant forms of puroindoline-b bound to DPPG monolayers. Journal of Physical Chemistry B, 112: 15907-15913.

References
- 170 -
Clifton LA, Lad MD, Green RJ, Frazier RA (2007b) Single amino acid substitutions in puroindoline-b mutants influence lipid binding properties. Biochemistry, 46: 2260-2266.
Clifton LA, Sanders MR, Castelletto V, Rogers SE, Heenan RK, Neylon C, Frazier RA, Green RJ (2011a) Puroindoline-a, a lipid binding protein from common wheat, spontaneously forms prolate protein micelles in solution. Physical Chemistry Chemical Physics, 13: 8881-8888.
Clifton LA, Sanders MR, Hughes AV, Neylon C, Frazier RA, Green RJ (2011b) Lipid binding interactions of antimicrobial plant seed defence proteins: Puroindoline-a and β-purothionin. Physical Chemistry Chemical Physics, 13: 17153-17162.
Conley AJ, Joensuu JJ, Richman A, Menassa R (2011) Protein body-inducing fusions for high-level production and purification of recombinant proteins in plants. Plant Biotechnology Journal, 9: 419-433.
Conrad U, Fiedler U (1998) Compartment-specific accumulation of recombinant immunoglobulins in plant cells: An essential tool for antibody production and immunomodulation of physiological functions and pathogen activity. Plant Molecular Biology, 38: 101-109.
Cornell H (2003) The chemistry and biochemistry of wheat. In: Cauvain S (ed.) Bread Making: Improving Quality. Woodhead Publishing. Cambridge, United Kingdom. pp 31-70.
Corona V, Gazza L, Boggini G, Pogna NE (2001) Variation in friabilin composition as determined by A-PAGE fractionation and PCR amplification, and its relationship to grain hardness in bread wheat. Journal of Cereal Science, 34: 243-250.
Cremazy FGE, Manders EMM, Bastiaens PIH, Kramer G, Hager GL, Van Munster EB, Verschure PJ, Gadella Jr TWJ, Van Driel R (2005) Imaging in situ protein-DNA interactions in the cell nucleus using FRET-FLIM. Experimental Cell Research, 309: 390-396.
Crone DE, Huang Y-M, Pitman DJ, Schenkelberg C, Fraser K, Macari S, Bystroff C (2013) GFP-Based Biosensors. In: Rinken T (ed.) State of the Art in Biosensors - General Aspects. Rijeka, croatia: InTech. pp 3-36.
Daniell H (2006) Production of biopharmaceuticals and vaccines in plants via the chloroplast genome. Biotechnology Journal, 1: 1071-1079.

References
- 171 -
Daniell H, Cohill PR, Kumar S, Dufourmantel N (2004) Chloroplast genetic engineering. In: Daniell H and Chase CD (eds.) Molecular Biology and Biotechnology of Plant Organelles. Springer, Netherlands. pp 443-490.
Daniell H, Ruiz G, Denes B, Sandberg L, Langridge W (2009a) Optimization of codon composition and regulatory elements for expression of human insulin like growth factor-1 in transgenic chloroplasts and evaluation of structural identity and function. BMC Biotechnology, 9: 33-48.
Daniell H, Singh ND, Mason H, Streatfield SJ (2009b) Plant-made vaccine antigens and biopharmaceuticals. Trends in Plant Science, 14: 669-679.
Daniell H, Streatfield SJ, Wycoff K (2001) Medical molecular farming: Production of antibodies, biopharmaceuticals and edible vaccines in plants. Trends in Plant Science, 6: 219-226.
Dathe M, Schumann M, Wieprecht T, Winkler A, Beyermann M, Krause E, Matsuzaki K, Murase O, Bienert M (1996) Peptide helicity and membrane surface charge modulate the balance of electrostatic and hydrophobic interactions with lipid bilayers and biological membranes. Biochemistry, 35: 12612-12622.
Davies HM (2010) Review article: Commercialization of whole-plant systems for biomanufacturing of protein products: evolution and prospects. Plant Biotechnology Journal, 8: 845-861.
Day L, Bhandari DG, Greenwell P, Leonard SA, Schofield JD (2006) Characterization of wheat puroindoline proteins. FEBS Journal, 273: 5358-5373.
De Bolle MFC, Osborn RW, Goderis IJ, Noe L, Acland D, Hart CA, Torrekens S, Van Leuven F, Broekaert WF (1996) Antimicrobial peptides from Mirabilis jalapa and Amaranthus caudatus: Expression, processing, localization and biological activity in transgenic tobacco. Plant Molecular Biology, 31: 993-1008.
De Jaeger G, Buys E, Eeckhout D, De Wilde C, Jacobs A, Kapila J, Angenon G, Van Montagu M, Gerats T, Depicker A (1999) High level accumulation of single-chain variable fragments in the cytosol of transgenic Petunia hybrida. European Journal of Biochemistry, 259: 426-434.
Dean R, Van Kan JaL, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, Rudd JJ, Dickman M, Kahmann R, Ellis J, Foster GD (2012) The Top 10 fungal pathogens in molecular plant pathology. Molecular Plant Pathology, 13: 414-430.

References
- 172 -
Delano WL (2002) PyMOL: An Open-Source Molecular Graphics Tool. DeLano Scientific, San Carlos, CA, USA, 40.
Delcour JA, Hoseney RC (2010) Principles of cereal science and technology authors provide insight into the current state of cereal processing. Cereal Foods World, 55: 21-22.
Desai PN, Shrivastava N, Padh H (2010) Production of heterologous proteins in plants: Strategies for optimal expression. Biotechnology Advances, 28: 427-435.
Digeon JF, Guiderdoni E, Alary R, Michaux-Ferrière N, Joudrier P, Gautier MF (1999) Cloning of a wheat puroindoline gene promoter by IPCR and analysis of promoter regions required for tissue-specific expression in transgenic rice seeds. Plant Molecular Biology, 39: 1101-1112.
Doekes GJ, Belderok B (1976) Kernel hardness and baking quality of wheat - A genetic analysis using chromosome substitution lines. Euphytica, 25: 565-576.
Doran PM (2006) Foreign protein degradation and instability in plants and plant tissue cultures. Trends in Biotechnology, 24: 426-432.
Doran PM (2013) Therapeutically important proteins from in vitro plant tissue culture systems. Current Medicinal Chemistry, 20: 1047-1055.
Douliez JP, Michon T, Elmorjani K, Marion D (2000) Structure, biological and technological functions of lipid transfer proteins and indolines, the major lipid binding proteins from cereal kernels. Journal of Cereal Science, 32: 1-20.
Dubcovsky J, Dvorak J (2007) Genome plasticity a key factor in the success of polyploid wheat under domestication. Science, 316: 1862-1866.
Dubovskii PV, Vassilevski AA, Slavokhotova AA, Odintsova TI, Grishin EV, Egorov TA, Arseniev AS (2011) Solution structure of a defense peptide from wheat with a 10-cysteine motif. Biochemical and Biophysical Research Communications, 411: 14-18.
Dubreil L, Compoint JP, Marion D (1997) Interaction of puroindolines with wheat flour polar lipids determines their foaming properties. Journal of Agricultural and Food Chemistry, 45: 108-116.

References
- 173 -
Dubreil L, Gaborit T, Bouchet B, Gallant DJ, Broekaert WF, Quillien L, Marion D (1998) Spatial and temporal distribution of the major isoforms of puroindolines (puroindoline-a and puroindoline-b) and non specific lipid transfer protein (ns-LTPle1) of Triticum aestivum seeds. Relationships with their in vitro antifungal properties. Plant Science, 138: 121-135.
Eckardt NA (2001) A sense of self: the role of DNA sequence elimination in allopolyploidization. The Plant Cell, 13: 1699-1704.
Egelkrout E, Rajan V, Howard JA (2012) Overproduction of recombinant proteins in plants. Plant Science, 184: 83-101.
Egorov TA, Odintsova TI, Pukhalsky VA, Grishin EV (2005) Diversity of wheat anti-microbial peptides. Peptides, 26: 2064-2073.
Elmorjani K, Geneix N, Dalgalarrondo M, Branlard G, Marion D (2013) Wheat grain softness protein (Gsp1) is a puroindoline-like protein that displays a specific post-translational maturation and does not interact with lipids. Journal of Cereal Science, 58: 117-122.
Emanuelsson O, Nielsen H, Von Heijne G (1999) ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Science, 8: 978-984.
Endo Y, Sawasaki T (2006) Cell-free expression systems for eukaryotic protein production. Current Opinion in Biotechnology, 17: 373-380.
Espinel-Ingroff A, Cantón E (2007) Antifungal susceptibility testing of filamentous fungi. In: Schwalbe R, Steele-Moore L and Goodwin AC (eds.) Antimicrobial Susceptibility Testing Protocols. Hoboken: CRC Press. pp 209-241.
Evrard A, Lagarde V, Joudrier P, Gautier MF (2008) Puroindoline-a and puroindoline-b interact with the Saccharomyces cerevisiae plasma membrane through different amino acids present in their tryptophan-rich domain. Journal of Cereal Science, 48: 379-386.
Faize M, Sourice S, Dupuis F, Parisi L, Gautier MF, Chevreau E (2004) Expression of wheat puroindoline-b reduces scab susceptibility in transgenic apple (Malus × domestica Borkh). Plant Science, 167: 347-354.

References
- 174 -
Faye L, Boulaflous A, Benchabane M, Gomord V, Michaud D (2005) Protein modifications in the plant secretory pathway: Current status and practical implications in molecular pharming. Vaccine, 23: 1770-1778.
Feiz L, Beecher BS, Martin JM, Giroux MJ (2009a) In planta mutagenesis determines the functional regions of the wheat puroindoline proteins. Genetics, 183: 853-860.
Feiz L, Martin JM, Giroux MJ (2009b) Creation and functional analysis of new puroindoline alleles in Triticum aestivum. Theoretical and Applied Genetics, 118: 247-257.
Feiz L, Wanjugi HW, Melnyk CW, Altosaar I, Martin JM, Giroux MJ (2009c) Puroindolines co-localize to the starch granule surface and increase seed bound polar lipid content. Journal of Cereal Science, 50: 91-98.
Feldman M (1995) Wheats. In: Smartt. J and Simmonds. NW (eds.) Evolution of Crop Plants. Longman Scientific and Technical. Harlow, UK. pp 184-192.
Feldman M (2001) Origin of cultivated wheat. In: Bonjean AP and Angus WJ (eds.) The world wheat book: a history of wheat breeding. Lavoisier Publishing. France. pp 3-56.
Fischer R, Emans N (2000) Molecular farming of pharmaceutical proteins. Transgenic Research, 9: 279-299.
Fischer R, Schillberg S, Buyel JF, Twyman RM (2013) Commercial aspects of pharmaceutical protein production in plants. Current Pharmaceutical Design, 19: 5471-5477.
Fischer R, Stoger E, Schillberg S, Christou P, Twyman RM (2004) Plant-based production of biopharmaceuticals. Current Opinion in Plant Biology, 7: 152-158.
Fischer R, Twyman RM, Schillberg S (2003) Production of antibodies in plants and their use for global health. Vaccine, 21: 820-825.
Fujiwara M, Suzuki G, Kudo D, Oba H, Wada Y, Wada H, Wada N, Rahman S, Fukui K, Mukai Y (2014) Localization of transgene-derived friabilins in rice endosperm cells. Plant Biotechnology, 31: 67-70.

References
- 175 -
Gaberc-Porekar V, Menart V (2001) Perspectives of immobilized-metal affinity chromatography. Journal of Biochemical and Biophysical Methods, 49: 335-360.
Gao AG, Hakimi SM, Mittanck CA, Wu Y, Woerner BM, Stark DM, Shah DM, Liang J, Rommens CMT (2000) Fungal pathogen protection in potato by expression of a plant defensin peptide. Nature Biotechnology, 18: 1307-1310.
García-Olmedo F, Molina A, Alamillo JM, Rodríguez-Palenzuéla P (1998) Plant defense peptides. Peptide Science, 47: 479-491.
Gasparis S, Orczyk W, Nadolska-Orczyk A (2013) Sina and Sinb genes in triticale do not determine grain hardness contrary to their orthologs Pina and Pinb in wheat. BMC Plant Biology, 13: 190-201.
Gasparis S, Orczyk W, Zalewski W, Nadolska-Orczyk A (2011) The RNA-mediated silencing of one of the Pin genes in allohexaploid wheat simultaneously decreases the expression of the other, and increases grain hardness. Journal of Experimental Botany, 62: 4025-4036.
Gautier MF, Aleman ME, Guirao A, Marion D, Joudrier P (1994) Triticum aestivum puroindolines, two basic cystine-rich seed proteins: cDNA sequence analysis and developmental gene expression. Plant Molecular Biology, 25: 43-57.
Gautier MF, Cosson P, Guirao A, Alary R, Joudrier P (2000) Puroindoline genes are highly conserved in diploid ancestor wheats and related species but absent in tetraploid Triticum species. Plant Science, 153: 81-91.
Gazza L, Nocente F, Ng PKW, Pogna NE (2005) Genetic and biochemical analysis of common wheat cultivars lacking puroindoline a. Theoretical and Applied Genetics, 110: 470-478.
Gazza L, Zanella L, Pogna N (2008) Development of durum wheat (Triticum turgidum ssp durum) lines with soft kernel texture by chromosome engineering In: Appels R, Eastwood R, Lagudah E, Langridge P, Mackay M, Mcintyre L and Sharp P (eds.) Proceedings of the 11th International, Wheat Genetics Symposium. Sydney, Australia: Sydney University. pp 339-41.
Gell DA, Grant RP, Mackay JP (2012) The detection and quantitation of protein oligomerzation In: Matthews JM (ed.) Protein Dimerization and Oligomerization in Biology. Springer Science+Business Media, LLC. New York, USA. pp 19-37.

References
- 176 -
Gelvin SB (2003) Agrobacterium-mediated plant transformation: The biology behind the "gene-jockeying" tool. Microbiology and Molecular Biology Reviews, 67: 16-37.
Giddings G, Allison G, Brooks D, Carter A (2000) Transgenic plants as factories for biopharmaceuticals. Nature Biotechnology, 18: 1151-1155.
Gill BS, Appels R, Botha-Oberholster AM, Buell CR, Bennetzen JL, Chalhoub B, Chumley F, Dvořák J, Iwanaga M, Keller B, Li W, Mccombie WR, Ogihara Y, Quetier F, Sasaki T (2004) A workshop report on wheat genome sequencing: International genome research on wheat consortium. Genetics, 168: 1087-1096.
Gillies SA, Futardo A, Henry RJ (2012) Gene expression in the developing aleurone and starchy endosperm of wheat. Plant Biotechnology Journal, 10: 668-679.
Gils M, Kandzia R, Marillonnet S, Klimyuk V, Gleba Y (2005) High-yield production of authentic human growth hormone using a plant virus-based expression system. Plant Biotechnology Journal, 3: 613-620.
Giritch A, Marillonnet S, Engler C, Van Eldik G, Botterman J, Klimyuk V, Gleba Y (2006) Rapid high-yield expression of full-size IgG antibodies in plants coinfected with noncompeting viral vectros. Proceedings of the National Academy of Sciences of the United States of America, 103: 14701-14706.
Giroux MJ, Kim KH, Hogg AC, Martin JM, Beecher B (2013) The Puroindoline b-2 variants are expressed at low levels relative to the Puroindoline D1 genes in wheat seeds. Crop Science, 53: 833-841.
Giroux MJ, Morris CF (1997) A glycine to serine change in puroindoline b is associated with wheat grain hardness and low levels of starch-surface friabilin. Theoretical and Applied Genetics, 95: 857-864.
Giroux MJ, Morris CF (1998) Wheat grain hardness results from highly conserved mutations in the friabilin components puroindoline a and b. Proceedings of the National Academy of Sciences of the United States of America, 95: 6262-6266.
Gleba Y, Giritch A (2012) Vaccines, antibodies, and pharmaceutical proteins. In: Altman A and Hasegawa PM (eds.) Plant Biotechnology and Agriculture. Elsevier Inc, USA. pp 465-479.

References
- 177 -
Gleba Y, Klimyuk V, Marillonnet S (2005) Magnifection - A new platform for expressing recombinant vaccines in plants. Vaccine, 23: 2042-2048.
Gleba Y, Klimyuk V, Marillonnet S (2007) Viral vectors for the expression of proteins in plants. Current Opinion in Biotechnology, 18: 134-141.
Gleba Y, Marillonnet S, Klimyuk V (2004) Engineering viral expression vectors for plants: The 'full virus' and the 'deconstructed virus' strategies. Current Opinion in Plant Biology, 7: 182-188.
Gleba Y, Tusé D, Giritch A (2014) Plant viral vectors for delivery by Agrobacterium. In: Palmer K and Gleba Y (eds.) Current Topics in Microbiology and Immunology; Plant Viral Vectors. Springer, Berlin Heidelberg, Germany. pp 155-192.
Gomord V, Faye L (2004) Posttranslational modification of therapeutic proteins in plants. Current Opinion in Plant Biology, 7: 171-181.
Goodin MM, Zaitlin D, Naidu RA, Lommel SA (2008) Nicotiana benthamiana: Its history and future as a model for plant-pathogen interactions. Molecular Plant-Microbe Interactions, 21: 1015-1026.
Goodspeed TH (1954) The genus Nicotiana. Journal of the American Pharmaceutical Association, 45: 193-193.
Greenblatt GA, Bettge AD, Morris CF (1995) Relationship between endosperm texture and the occurrence of friabilin and bound polar lipids on wheat starch. Cereal Chemistry, 72: 172-176.
Greenfield NJ (2007) Using circular dichroism spectra to estimate protein secondary structure. Nature Protocols, 1: 2876-2890.
Greenwell P, Schofield J (1986) A starch Granule protein associated with endosperm softness in wheat. Cereal Chemistry, 63: 379-380.
Greenwell P, Stimson W, Mackay E (1992) Durotest-rapid purity control for pasta semolina. Chorleywood Dig, 121: 115-116.

References
- 178 -
Grefen C, Städele K, Růžička K, Obrdlik P, Harter K, Horák J (2008) Subcellular localization and in vivo interactions of the Arabidopsis thaliana ethylene receptor family members. Molecular Plant, 1: 308-320.
Haider N (2013) The origin of the B-genome of bread wheat (Triticum aestivum L.). Russian Journal of Genetics, 49: 263-274.
Hamada MS, Yin Y, Chen H, Ma Z (2011) The escalating threat of Rhizoctonia cerealis, the causal agent of sharp eyespot in wheat. Pest Management Science, 67: 1411-1419.
Hancock REW, Chapple DS (1999) Peptide antibiotics. Antimicrobial Agents and Chemotherapy, 43: 1317-1323.
Haney EF, Petersen AP, Lau CK, Jing W, Storey DG, Vogel HJ (2013) Mechanism of action of puroindoline derived tryptophan-rich antimicrobial peptides. Biochimica et Biophysica Acta - Biomembranes, 1828: 1802-1813.
Hearn MTW, Acosta D (2001) Applications of novel affinity cassette methods: Use of peptide fusion handles for the purification of recombinant proteins. Journal of Molecular Recognition, 14: 323-369.
Hellens R, Allan A, Friel E, Bolitho K, Grafton K, Templeton M, Karunairetnam S, Gleave A, Laing W (2005) Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods, 1: 13-27.
Hogg AC, Sripo T, Beecher B, Martin JM, Giroux MJ (2004) Wheat puroindolines interact to form friabilin and control wheat grain hardness. Theoretical and Applied Genetics, 108: 1089-1097.
Horn ME, Woodard SL, Howard JA (2004) Plant molecular farming: Systems and products. Plant Cell Reports, 22: 711-720.
Horton P, Park KJ, Obayashi T, Fujita N, Harada H, Adams-Collier CJ, Nakai K (2007) WoLF PSORT: Protein localization predictor. Nucleic Acids Research, 35: W585-W587.
Hoseney R, Seib P (1973) Structural differences in hard and soft wheat. Bakers Dig, 47 (6): 26.

References
- 179 -
Howard JA, Hood E (2005) Bioindustrial and biopharmaceutical products produced in plants. Advances in Agronomy, 85: 91-124.
Hu CD, Chinenov Y, Kerppola TK (2002) Visualization of interactions among bZIP and Rel family proteins in living cells using bimolecular fluorescence complementation. Molecular Cell, 9: 789-798.
Hu CD, Kerppola TK (2003) Simultaneous visualization of multiple protein interactions in living cells using multicolor fluorescence complementation analysis. Nature Biotechnology, 21: 539-545.
Huang FC, Studart-Witkowski C, Schwab W (2010) Overexpression of hydroperoxide lyase gene in Nicotiana benthamiana using a viral vector system. Plant Biotechnology Journal, 8: 783-795.
Hyunjong B, Lee DS, Hwang I (2006) Dual targeting of xylanase to chloroplasts and peroxisomes as a means to increase protein accumulation in plant cells. Journal of Experimental Botany, 57: 161-169.
Ikeda TM, Cong H, Suzuki T, Takata K (2010) Identification of new Pina null mutations among Asian common wheat cultivars. Journal of Cereal Science, 51: 235-237.
Ikeda TM, Ohnishi N, Nagamine T, Oda S, Hisatomi T, Yano H (2005) Identification of new puroindoline genotypes and their relationship to flour texture among wheat cultivars. Journal of Cereal Science, 41: 1-6.
Inoue H, Nojima H, Okayama H (1990) High efficiency transformation of Escherichia coli with plasmids. Gene, 96: 23-28.
Issaly N, Solsona O, Joudrier P, Gautier MF, Moulin G, Boze H (2001) Optimization of the wheat puroindoline-a production in Pichia pastoris. Journal of Applied Microbiology, 90: 397-406.
Jacob L, Zasloff M (1994) Potential therapeutic applications of magainins and other antimicrobial agents of animal origin. Ciba Foundation symposium, 186: 197-216.
Jasiewicz B, Mrówczyńska L, Malczewska-Jaskóła K (2014) Synthesis and haemolytic activity of novel salts made of nicotine alkaloids and bile acids. Bioorganic and Medicinal Chemistry Letters, 24: 1104-1107.

References
- 180 -
Jenssen H, Hamill P, Hancock REW (2006) Peptide antimicrobial agents. Clinical Microbiology Reviews, 19: 491-511.
Jerkovic A, Kriegel AM, Bradner JR, Atwell BJ, Roberts TH, Willows RD (2010) Strategic distribution of protective proteins within bran layers of wheat protects the nutrient-rich endosperm. Plant Physiology, 152: 1459-1470.
Jing W, Demcoe AR, Vogel HJ (2003) Conformation of a bactericidal domain of puroindoline a: Structure and mechanism of action of a 13-residue antimicrobial peptide. Journal of Bacteriology, 185: 4938-4947.
Jolly CJ, Glenn GM, Rahman S (1996) GSP-1 genes are linked to the grain hardness locus (Ha) on wheat chromosome 5D. Proceedings of the National Academy of Sciences of the United States of America, 93: 2408-2413.
Jolly CJ, Rahman S, Kortt AA, Higgins TJV (1993) Characterisation of the wheat Mr 15000 "grain-softness protein" and analysis of the relationship between its accumulation in the whole seed and grain softness. Theoretical and Applied Genetics, 86: 589-597.
José-Estanyol M, Gomis-Rüth FX, Puigdomènech P (2004) The eight-cysteine motif, a versatile structure in plant proteins. Plant Physiology and Biochemistry, 42: 355-365.
Kerppola TK (2006) Design and implementation of bimolecular fluorescence complementation (BiFC) assays for the visualization of protein interactions in living cells. Nature Protocols, 1: 1278-1286.
Kerppola TK (2008) Bimolecular fluorescence complementation (BiFC) analysis as a probe of protein interactions in living cells. Annual Review of Biophysics, 37: 465-487.
Kerppola TK (2013) Multicolor bimolecular fluorescence complementation (BiFC) analysis of protein interactions with alternative partners. Cold Spring Harbor Protocols, 798-803.
Kim KH, Feiz L, Dyer AT, Grey W, Hogg AC, Martin JM, Giroux MJ (2012a) Increased resistance to Penicillium seed rot in transgenic wheat over-expressing puroindolines. Journal of Phytopathology, 160: 243-247.

References
- 181 -
Kim KH, Feiz L, Martin JM, Giroux MJ (2012b) Puroindolines are associated with decreased polar lipid breakdown during wheat seed development. Journal of Cereal Science, 56: 142-146.
Kitamura S, Inoue M, Shikazono N, Tanaka A (2001) Relationships among Nicotiana species revealed by the 5S rDNA spacer sequence and fluorescence in situ hybridization. Theoretical and Applied Genetics, 103: 678-686.
Klimyuk V, Marillonnet S, Knäblein J, Mccaman M, Gleba Y (2008) Production of recombinant proteins in plants. In: Knaeblein J (eds.) Modern Biopharmaceuticals, Wiley-VCH Verlag GmbH & Co, Weinheim, Germany. 3: pp 893-917.
Klimyuk V, Pogue G, Herz S, Butler J, Haydon H (2014) Production of recombinant antigens and antibodies in Nicotiana benthamiana using ‘Magnifection’ technology: GMP-compliant facilities for small- and large-scale manufacturing. In: Palmer K and Gleba Y (eds.) Current Topics in Microbiology and Immunology; Plant Viral Vectors. Springer, Berlin Heidelberg, Germany. pp 127-154.
Knapp S, Chase MW, Clarkson JJ (2004) Nomenclatural changes and a new sectional classification in Nicotiana (Solanaceae). Taxon, 53: 73-82.
Konopka I, Kozirok W, Tańska M (2005) Wheat endosperm hardness. Part I. Relationships to colour of kernel cross-section. European Food Research and Technology, 220: 11-19.
Kooijman M, Orsel R, Hamer RJ, Bekkers ACaPA (1998) The insertion behaviour of wheat puroindoline-a into diacylgalactosylglycerol films. Journal of Cereal Science, 28: 43-51.
Kooijman M, Orsel R, Hessing M, Hamer RJ, Bekkers ACaPA (1997) Spectroscopic characterisation of the lipid-binding properties of wheat puroindolines. Journal of Cereal Science, 26: 145-159.
Krishnamurthy K, Balconi C, Sherwood JE, Giroux MJ (2001) Wheat puroindolines enhance fungal disease resistance in transgenic rice. Molecular Plant-Microbe Interactions, 14: 1255-1260.
Krishnamurthy K, Giroux MJ (2001) Expression of wheat puroindoline genes in transgenic rice enhances grain softness. Nature Biotechnology, 19: 162-166.

References
- 182 -
Krishnan HB, Natarajan SS (2009) A rapid method for depletion of Rubisco from soybean (Glycine max) leaf for proteomic analysis of lower abundance proteins. Phytochemistry, 70: 1958-1964.
Lalonde S, Ehrhardt DW, Frommer WB (2005) Shining light on signaling and metabolic networks by genetically encoded biosensors. Current Opinion in Plant Biology, 8: 574-581.
Law C, Young C, Brown J, Snape J, Worland A. (1978) The study of grain protein control in wheat using whole chromosome substitution lines. Seed protein improvement by nuclear techniques, Conference Vienna, Austria. International Atomic Energy Agency. pp 483-502.
Le Bihan T, Blochet JÉ, Désormeaux A, Marion D, Pézolet M (1996) Determination of the secondary structure and conformation of puroindolines by infrared and Raman spectroscopy. Biochemistry, 35: 12712-12722.
Le Guernevé C, Seigneuret M, Marion D (1998) Interaction of the wheat endosperm lipid-binding protein puroindoline-a with phospholipids. Archives of Biochemistry and Biophysics, 360: 179-186.
Leckie BM, Stewart J, C. N. (2011) Agroinfiltration as a technique for rapid assays for evaluating candidate insect resistance transgenes in plants. Plant Cell Reports, 30: 325-334.
Lee LY, Gelvin SB (2008) T-DNA binary vectors and systems. Plant Physiology, 146: 325-332.
Lee S-H, Kim S-J, Lee Y-S, Song M-D, Kim I-H, Won H-S (2011a) De novo generation of short antimicrobial peptides with simple amino acid composition. Regulatory Peptides, 166: 36-41.
Lee S, Li B, Jin S, Daniell H (2011b) Expression and characterization of antimicrobial peptides Retrocyclin-101 and Protegrin-1 in chloroplasts to control viral and bacterial infections. Plant Biotechnology Journal, 9: 100-115.
Lesage VS, Bouchet B, Rhazi L, Elmorjani K, Branlard G, Marion D (2011) New insight into puroindoline function inferred from their subcellular localization in developing hard and soft near-isogenic endosperm and their relationship with polymer size of storage proteins. Journal of Cereal Science, 53: 231-238.

References
- 183 -
Lesk AM (2010) Introduction to Protein Science: Architecture, Function, and Genomics., Oxford University Press New York, ISBN: 9780199541300.
Leuzinger K, Dent M, Hurtado J, Stahnke J, Lai H, Zhou X, Chen Q (2013) Efficient agroinfiltration of plants for high-level transient expression of recombinant proteins. Journal of Visualized Experiments, 77: 50521.
Lewis R (2011) Nicotiana. In: Kole C (ed.) Wild Crop Relatives: Genomic and Breeding Resources. Springer Berlin Heidelberg, Germany. pp 185-208.
Li G, He Z, Lillemo M, Sun Q, Xia X (2008) Molecular characterization of allelic variations at Pina and Pinb loci in Shandong wheat landraces, historical and current cultivars. Journal of Cereal Science, 47: 510-517.
Li G, He Z, Peña RJ, Xia X, Lillemo M, Sun Q (2006) Identification of novel secaloindoline-a and secaloindoline-b alleles in CIMMYT hexaploid triticale lines. Journal of Cereal Science, 43: 378-386.
Li Y, Geng Y, Song H, Zheng G, Huan L, Qiu B (2004) Expression of a human lactoferrin N-lobe in Nicotiana benthmiana with potato virus X-based agroinfection. Biotechnology letters, 26: 953-957.
Li Y, Mao X, Wang Q, Zhang J, Li X, Ma F, Sun F, Chang J, Chen M, Wang Y, Li K, Yang G, He G (2014) Overexpression of Puroindoline a gene in transgenic durum wheat (Triticum turgidum ssp. durum) leads to a medium-hard kernel texture. Molecular Breeding, 33: 545-554.
Li Y, Wang Q, Li X, Xiao X, Sun F, Wang C, Hu W, Feng Z, Chang J, Chen M, Wang Y, Li K, Yang G, He G (2012a) Coexpression of the high molecular weight glutenin subunit 1Ax1 and puroindoline improves dough mixing properties in Durum Wheat (Triticum turgidum L. ssp. durum). PLoS ONE, 7.
Li Y, Xiang Q, Zhang Q, Huang Y, Su Z (2012b) Overview on the recent study of antimicrobial peptides: Origins, functions, relative mechanisms and application. Peptides, 37: 207-215.
Lillemo M, Chen F, Xia X, William M, Peña RJ, Trethowan R, He Z (2006) Puroindoline grain hardness alleles in CIMMYT bread wheat germplasm. Journal of Cereal Science, 44: 86-92.

References
- 184 -
Lillemo M, Morris CF (2000) A leucine to proline mutation in puroindoline B is frequently present in hard wheats from Northern Europe. Theoretical and Applied Genetics, 100: 1100-1107.
Ling HY, Pelosi A, Walmsley AM (2010) Current status of plant-made vaccines for veterinary purposes. Expert Review of Vaccines, 9: 971-982.
Listwan P, Martin JL, Kobe B, Cowieson NP (2004) Methods for high-throughput protein expression, purification and structure determination adapted for structural genomics. Australian Biochemist, 35: 43-46.
Los GV, Encell LP, Mcdougall MG, Hartzell DD, Karassina N, Zimprich C, Wood MG, Learish R, Ohana RF, Urh M, Simpson D, Mendez J, Zimmerman K, Otto P, Vidugiris G, Zhu J, Darzins A, Klaubert DH, Bulleit RF, Wood KV (2008) HaloTag: A novel protein labeling technology for cell imaging and protein analysis. ACS Chemical Biology, 3: 373-382.
Ludevid D, Torrent M, Lasserre-Ramassamy S (2005) Production of peptides and proteins by accumulation in plant endoplasmic reticulumderived protein bodies. Patent EP1523558.
Luo L, Zhang J, Yang G, Li Y, Li K, He G (2008) Expression of puroindoline a enhances leaf rust resistance in transgenic tetraploid wheat. Molecular Biology Reports, 35: 195-200.
Luo Y, Zhang DD, Dong XW, Zhao PB, Chen LL, Song XY, Wang XJ, Chen XL, Shi M, Zhang YZ (2010) Antimicrobial peptaibols induce defense responses and systemic resistance in tobacco against tobacco mosaic virus. FEMS Microbiology Letters, 313: 120-126.
Ma JKC, Barros E, Bock R, Christou P, Dale PJ, Dix PJ, Fischer R, Irwin J, Mahoney R, Pezzotti M, Schillberg S, Sparrow P, Stoger E, Twyman RM (2005) Molecular farming for new drugs and vaccines. Current perspectives on the production of pharmaceuticals in transgenic plants. EMBO Reports, 6: 593-599.
Ma JKC, Drake PMW, Christou P (2003) The production of recombinant pharmaceutical proteins in plants. Nature Reviews Genetics, 4: 794-805.
Ma JKC, Hiatt A, Hein M, Vine ND, Wang F, Stabila P, Van Dolleweerd C, Mostov K, Lehner T (1995) Generation and assembly of secretory antibodies in plants. Science, 268: 716-719.

References
- 185 -
Ma JKC, Hikmat BY, Wycoff K, Vine ND, Chargelegue D, Yu L, Hein MB, Lehner T (1998) Characterization of a recombinant plant monoclonal secretory antibody and preventive immunotherapy in humans. Nature Medicine, 4: 601-606.
Magliery TJ, Regan L (2006) Reassembled GFP: Detecting protein-protein interactions and protein expression patterns. In: Chalfie M and Kain S (eds.) Green Fluorescent Protein: Properties, Applications, and Protocols. John Wiley & Sons, Inc. Toronto, Canada. pp 391-405.
Mainieri D, Rossi M, Archinti M, Bellucci M, De Marchis F, Vavassori S, Pompa A, Arcioni S, Vitale A (2004) Zeolin. A new recombinant storage protein constructed using maize γ-zein and bean phaseolin. Plant Physiology, 136: 3447-3456.
Marianayagam NJ, Sunde M, Matthews JM (2004) The power of two: Protein dimerization in biology. Trends in Biochemical Sciences, 29: 618-625.
Marillonnet S, Giritch A, Gils M, Kandzia R, Klimyuk V, Gleba Y (2004) In planta engineering of viral RNA replicons: Efficient assembly by recombination of DNA modules delivered by Agrobacterium. Proceedings of the National Academy of Sciences of the United States of America, 101: 6852-6857.
Marillonnet S, Thoeringer C, Kandzia R, Klimyuk V, Gleba Y (2005) Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nature Biotechnology, 23: 718-723.
Marion D, Bakan B, Elmorjani K (2007) Plant lipid binding proteins: Properties and applications. Biotechnology Advances, 25: 195-197.
Marion D, Gautier MF, Joudrier P, Ptak M, Pezolet M, Forest E, Clark DC, Broekaert W (1994) Structure and function of wheat lipid binding proteins. In: Wheat kernel proteins: molecular and functional aspects. Universitá Degli Studi Della. Tuscia - Consiglio Nazionale Delle Ricerche, Viterbo. pp 175-180.
Martin JM, Frohberg RC, Morris CF, Talbert LE, Giroux MJ (2001) Milling and bread baking traits associated with puroindoline sequence type in hard red spring wheat. Crop Science, 41: 228-234.
Martin JM, Meyer FD, Smidansky ED, Wanjugi H, Blechl AE, Giroux MJ (2006) Complementation of the pina (null) allele with the wild type Pina sequence restores a soft phenotype in transgenic wheat. Theoretical and Applied Genetics, 113: 1563-1570.

References
- 186 -
Marusic C, Nuttall J, Buriani G, Lico C, Lombardi R, Baschieri S, Benvenuto E, Frigerio L (2007) Expression, intracellular targeting and purification of HIV Nef variants in tobacco cells. BMC Biotechnology, 7: 1-12.
Massa AN, Morris CF, Gill BS (2004) Sequence diversity of Puroindoline-a, Puroindoline-b, and the grain softness protein genes in Aegilops tauschii Coss. Crop Science, 44: 1808-1816.
Matsuoka Y (2011) Evolution of polyploid Triticum Wheats under cultivation: the role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant and Cell Physiology, 52: 750-64.
Mattanovich D, Rüker F, Da Cämara Machado A, Laimer M, Regner F, Steinkeliner H, Himmler G, Katinger H (1989) Efficient transformation of Agrobacterium spp. by eletroporation. Nucleic Acids Research, 17: 6747.
Mattern P, Morris R, Schmidt J, Johnson V. (1973) Location of genes for kernel properties in the wheat variety ‘Cheyenne’ using chromosome substitution lines. the 4th International Wheat Genetics Symposium, Conference Columbia. pp 703-708.
Melnik S, Stoger E (2013) Green factories for biopharmaceuticals. Current Medicinal Chemistry, 20: 1038-1046.
Melo MN, Ferre R, Castanho MaRB (2009) Antimicrobial peptides: Linking partition, activity and high membrane-bound concentrations. Nature Reviews Microbiology, 7: 245-250.
Mett V, Farrance CE, Green BJ, Yusibov V (2008) Plants as biofactories. Biologicals, 36: 354-358.
Miao Y, Chen L, Wang C, Wang Y, Zheng Q, Gao C, Yang G, He G (2012) Expression, purification and antimicrobial activity of puroindoline A protein and its mutants. Amino Acids, 43: 1689-1696.
Michaud D, Vrain T, Gomord V, Faye L (1998) Stability of recombinant proteins in plants. In: Cunningham C and Porter AR (eds.) Recombinant Proteins from Plants. Humana Press. New York, USA. pp 177-188.
Mikulíková D (2007) The effect of friabilin on wheat grain hardness. Czech Journal of Genetics and Plant Breeding, 43: 35-43.

References
- 187 -
Mohammadi M, Zaidi MA, Ochalski A, Tanchak M, Altosaar I (2007) Immunodetection and immunolocalization of tryptophanins in oat (Avena sativa L.) seeds. Plant Science, 172: 579-587.
Morell M, Espargaro A, Aviles FX, Ventura S (2008) Study and selection of in vivo protein interactions by coupling bimolecular fluorescence complementation and flow cytometry. Nature Protocols, 3: 22-33.
Morell M, Espargaró A, Avilés FX, Ventura S (2007) Detection of transient protein-protein interactions by bimolecular fluorescence complementation: The Abl-SH3 case. Proteomics, 7: 1023-1036.
Morris CF (2002) Puroindolines: the molecular genetic basis of wheat grain hardness. Plant Molecular Biology, 48: 633-647.
Morris CF, Greenblatt GA, Bettge AD, Malkawi HI (1994) Isolation and characterization of multiple forms of friabilin. Journal of Cereal Science, 20: 167-174.
Morris CF, Lillemo M, Simeone MC, Giroux MJ, Babb SL, Kidwell KK (2001) Prevalence of puroindoline grain hardness genotypes among historically significant North American spring and winter wheats. Crop Science, 41: 218-228.
Morris CF, Simeone MC, King GE, Lafiandra D (2011) Transfer of soft kernel texture from Triticum aestivum to durum wheat, Triticum turgidum ssp. durum. Crop Science, 51: 114-122.
Morrison WR, Greenwell P, Law CN, Sulaiman BD (1992) Occurrence of friabilin, a low molecular weight protein associated with grain softness, on starch granules isolated from some wheats and related species. Journal of Cereal Science, 15: 143-149.
Mukherjee S, Zhang Y (2011) Protein-protein complex structure predictions by multimeric threading and template recombination. Structure, 19: 955-966.
Munro S, Pelham HRB (1987) A C-terminal signal prevents secretion of luminal ER proteins. Cell, 48: 899-907.
Nadai M, Bally J, Vitel M, Job C, Tissot G, Botterman J, Dubald M (2009) High-level expression of active human alpha1-antitrypsin in transgenic tobacco chloroplasts. Transgenic Research, 18: 173-183.

References
- 188 -
Nadolska-Orczyk A, Gasparis S, Orczyk W (2009) The determinants of grain texture in cereals. Journal of Applied Genetics, 50: 185-197.
Nagpal S, Gupta V, Kaur KJ, Salunke DM (1999) Structure-function analysis of tritrypticin, an antibacterial peptide of innate immune origin. Journal of Biological Chemistry, 274: 23296-23304.
Naureen F, Humaira B, Viqar S, Ara J, Ehteshamul-Haque S (2009) Prevalance of post-harvest rot of vegetables and fruits in Karachi, Pakistan. Pakistan Journal of Botany, 41: 3185-3190.
Nawrot R, Barylski J, Nowicki G, Broniarczyk J, Buchwald W, Goździcka-Józefiak A (2014) Plant antimicrobial peptides. Folia Microbiologica, 59: 181-196.
Nebenfuhr A, Frohlick JA, Staehelin LA (2000) Redistribution of Golgi stacks and other organelles during mitosis and cytokinesis in plant cells. Plant Physiology, 124: 135-151.
Oda S, Schofield JD (1997) Characterisation of friabilin polypeptides. Journal of Cereal Science, 26: 29-36.
Odintsova TI, Vassilevski AA, Slavokhotova AA, Musolyamov AK, Finkina EI, Khadeeva NV, Rogozhin EA, Korostyleva TV, Pukhalsky VA, Grishin EV, Egorov TA (2009) A novel antifungal hevein-type peptide from Triticum kiharae seeds with a unique 10-cysteine motif. FEBS Journal, 276: 4266-4275.
Oey M, Lohse M, Scharff LB, Kreikemeyer B, Bock R (2009) Plastid production of protein antibiotics against pneumonia via a new strategy for high-level expression of antimicrobial proteins. Proceedings of the National Academy of Sciences of the United States of America, 106: 6579-6584.
Ohad N, Shichrur K, Yalovsky S (2007) The analysis of protein-protein interactions in plants by bimolecular fluorescence complementation. Plant Physiology, 145: 1090-1099.
Ormö M, Cubitt AB, Kallio K, Gross LA, Tsien RY, Remington SJ (1996) Crystal structure of the Aequorea victoria green fluorescent protein. Science, 273: 1392-1395.
Painter J, Rah JH, Lee YK (2002) Comparison of international food guide pictorial representations. Journal of the American Dietetic Association, 102: 483-489.

References
- 189 -
Palumbo D, Iannaccone M, Porta A, Capparelli R (2010) Experimental antibacterial therapy with puroindolines, lactoferrin and lysozyme in Listeria monocytogenes-infected mice. Microbes and Infection, 12: 538-545.
Pan Z, Song W, Meng F (2004) Characterization of genes encoding wheat grain hardness from Chinese cultivar Gaocheng 8901. Cereal Chemistry, 82: 38-43.
Parachin NS, Mulder KC, Viana AaB, Dias SC, Franco OL (2012) Expression systems for heterologous production of antimicrobial peptides. Peptides, 38: 446-456.
Patiño-Rodríguez O, Ortega-Berlanga B, Llamas-González YY, Flores-Valdez MA, Herrera-Díaz A, Montes-De-Oca-Luna R, Korban SS, Alpuche-Solís ÁG (2013) Transient expression and characterization of the antimicrobial peptide protegrin-1 in Nicotiana tabacum for control of bacterial and fungal mammalian pathogens. Plant Cell, Tissue and Organ Culture, 115: 99-106.
Pauly A, Pareyt B, Fierens E, Delcour JA (2013) Wheat (Triticum aestivum L. and T. turgidum L. ssp. durum) Kernel Hardness: I. Current view on the role of puroindolines and polar lipids. Comprehensive Reviews in Food Science and Food Safety, 12: 413-426.
Peña RJ, Trethowan R, Pfeiffer WH, Van Ginkel M (2002) Quality (end-use) improvement in wheat: Compositional, genetic, and environmental factors. Journal of Crop Production, 5: 1-37.
Phillips RL, Palombo EA, Panozzo JF, Bhave M (2011) Puroindolines, Pin alleles, hordoindolines and grain softness proteins are sources of bactericidal and fungicidal peptides. Journal of Cereal Science, 53: 112-117.
Pickering PA, Bhave M (2007) Comprehensive analysis of Australian hard wheat cultivars shows limited puroindoline allele diversity. Plant Science, 172: 371-379.
Pogue GP, Lindbo JA, Garger SJ, Fitzmaurice WP (2002) Making an ally from an enemy: Plant virology and the new agriculture. Annual Review of Phytopatology, 40: 45-74.
Portieles R, Ayra C, Gonzalez E, Gallo A, Rodriguez R, Chacón O, López Y, Rodriguez M, Castillo J, Pujol M, Enriquez G, Borroto C, Trujillo L, Thomma BPHJ, Borrás-Hidalgo O (2010) NmDef02, a novel antimicrobial gene isolated from Nicotiana megalosiphon confers high-level pathogen resistance under greenhouse and field conditions. Plant Biotechnology Journal, 8: 678-690.

References
- 190 -
Prasher DC, Eckenrode VK, Ward WW, Prendergast FG, Cormier MJ (1992) Primary structure of the Aequorea victoria green-fluorescent protein. Gene, 111: 229-233.
Ragupathy R, Cloutier S (2008) Genome organisation and retrotransposon driven molecular evolution of the endosperm Hardness (Ha) locus in Triticum aestivum cv Glenlea. Molecular Genetics and Genomics, 280: 467-481.
Rahman S, Jolly CJ, Skerritt JH, Wallosheck A (1994) Cloning of a wheat 15-kDa grain softness protein (GSP). GSP is a mixture of puroindoline-like polypeptides. European Journal of Biochemistry, 223: 917-925.
Rakszegi M, Wilkinson MD, Tosi P, Lovegrove A, Kovacs G, Bedo Z, Shewry PR (2009) Puroindoline genes and proteins in tetraploid and hexaploid species of Triticum. Journal of Cereal Science, 49: 202-211.
Ram S, Jain N, Shoran J, Singh R (2005) New frame shift mutation in puroindoline B in Indian wheat cultivars Hyb65 and Nl5439. Journal of Plant Biochemistry and Biotechnology, 14: 45-48.
Ramalingam A, Palombo EA, Bhave M (2012) The Pinb-2 genes in wheat comprise a multigene family with great sequence diversity and important variants. Journal of Cereal Science, 56: 171-180.
Rao AQ, Bakhsh A, Kiani S, Shahzad K, Shahid AA, Husnain T, Riazuddin S (2009) The myth of plant transformation. Biotechnology Advances, 27: 753-763.
Rochelle JC (2001) Bread for the wilderness: Baking as spiritual craft, Xulon Press, ISBN: 9781931232524.
Roy A, Kucukural A, Zhang Y (2010) I-TASSER: a unified platform for automated protein structure and function prediction. Nature Protocols, 5: 725-738.
Rubio-Infante N, Govea-Alonso DO, Alpuche-Solís AG, García-Hernández AL, Soria-Guerra RE, Paz-Maldonado LMT, Ilhuicatzi-Alvarado D, Varona-Santos JT, Verdín-Terán L, Korban SS, Moreno-Fierros L, Rosales-Mendoza S (2012) A chloroplast-derived C4V3 polypeptide from the human immunodeficiency virus (HIV) is orally immunogenic in mice. Plant Molecular Biology, 78: 337-349.

References
- 191 -
Ruhlman T, Ahangari R, Devine A, Samsam M, Daniell H (2007) Expression of cholera toxin B-proinsulin fusion protein in lettuce and tobacco chloroplasts - Oral administration protects against development of insulitis in non-obese diabetic mice. Plant Biotechnology Journal, 5: 495-510.
Rybicki EP (2009) Plant-produced vaccines: promise and reality. Drug Discovery Today, 14: 16-24.
Saitoh H, Kiba A, Nishihara M, Yamamura S, Suzuki K, Terauchi R (2001) Production of antimicrobial defensin in Nicotiana benthamiana with a potato virus X vector. Molecular Plant-Microbe Interactions, 14: 111-115.
Samarkina ON, Popova AG, Gvozdik EY, Chkalina AV, Zvyagin IV, Rylova YV, Rudenko NV, Lusta KA, Kelmanson IV, Gorokhovatsky AY, Vinokurov LM (2009) Universal and rapid method for purification of GFP-like proteins by the ethanol extraction. Protein Expression and Purification, 65: 108-113.
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory manual (3rd ed) Cold Spring Harbor Laboratory Press N.Y, ISBN: 10 0-87969-577-3.
Sanders MR, Clifton LA, Neylon C, Frazier RA, Green RJ (2013) Selected wheat seed defense proteins exhibit competitive binding to model microbial lipid interfaces. Journal of Agricultural and Food Chemistry, 61: 6890-6900.
Santi L, Batchelor L, Huang Z, Hjelm B, Kilbourne J, Arntzen CJ, Chen Q, Mason HS (2008) An efficient plant viral expression system generating orally immunogenic Norwalk virus-like particles. Vaccine, 26: 1846-1854.
Santi L, Giritch A, Roy CJ, Marillonnet S, Klimyuk V, Gleba Y, Webb R, Arntzen CJ, Mason HS (2006) Protection conferred by recombinant Yersinia pestis antigens produced by a rapid and highly scalable plant expression system. Proceedings of the National Academy of Sciences of the United States of America, 103: 861-866.
Sanz-Barrio R, Millán AFS, Corral-Martínez P, Seguí-Simarro JM, Farran I (2011) Tobacco plastidial thioredoxins as modulators of recombinant protein production in transgenic chloroplasts. Plant Biotechnology Journal, 9: 639-650.
Sathish K, Sriraman R, Subramanian BM, Rao NH, Balaji K, Narasu ML, Srinivasan VA (2011) Plant expressed EtMIC2 is an effective immunogen in conferring protection against chicken coccidiosis. Vaccine, 29: 9201-9208.

References
- 192 -
Sawasaki T, Ogasawara T, Morishita R, Endo Y (2002) A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America, 99: 14652-14657.
Scheller J, Conrad U (2004) Production of spider silk proteins in transgenic tobacco and potato. In: Fischer R and Schillberg S (eds.) Molecular Farming. Wiley-VCH Verlag GmbH & Co. KGaA,Weinheim, Germany. pp 171-181.
Schibli DJ, Epand RF, Vogel HJ, Epand RM (2002) Tryptophan-rich antimicrobial peptides: Comparative properties and membrane interactions. Biochemistry and Cell Biology, 80: 667-677.
Schiermeyer A, Dorfmüller S, Schinke H (2004) Production of pharmaceutical proteins in plants and plant cell suspension cultures. In: Fischer R and Stefan S (eds.) Molecular Farming. Wiley-VCH Verlag GmbH & Co. KGaA,Weinheim, Germany. pp 91-112.
Schiffer M, Chang CH, Stevens FJ (1992) The functions of tryptophan residues in membrane proteins. Protein Engineering, 5: 213-214.
Schmidt FR (2004) Recombinant expression systems in the pharmaceutical industry. Applied Microbiology and Biotechnology, 65: 363-372.
Scholthof HB, Scholthof KBG, Jackson AO (1996) Plant virus gene vectors for transient expression of foreign proteins in plants. Annual Review of Phytopatology, 34: 299-323.
Schütze K, Harter K, Chaban C (2009) Bimolecular fluorescence complementation (BiFC) to study protein-protein interactions in living plant cells. Methods in Molecular Biology, 479: 189-202.
Scopes RK (1993) Protein Purification: Principles and Practice, Springer New York, ISBN: 978-1-4419-2833-7.
Seelig J (2004) Thermodynamics of lipid-peptide interactions. Biochimica et Biophysica Acta - Biomembranes, 1666: 40-50.
Shai Y (2002) Mode of action of membrane active antimicrobial peptides. Biopolymers - Peptide Science Section, 66: 236-248.

References
- 193 -
Sharma KK, Bhatnagar-Mathur P, Thorpe TA (2005) Genetic transformation technology: Status and problems. In Vitro Cellular and Developmental Biology - Plant, 41: 102-112.
Shevchenko A, Tomas H, Havliš J, Olsen JV, Mann M (2007) In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nature Protocols, 1: 2856-2860.
Shewry PR (2009) Wheat. Journal of Experimental Botany, 60: 1537-1553.
Shewry PR (2013) Improving grain quality: Wheat. In: Becraft PW (ed.) Seed Genomics. John Wiley & Sons, Inc. Toronto, Canada. pp 159-178.
Shimomura O, Johnson FH, Saiga Y (1962) Extraction, purification and properties of Aequorin, a bioluminescent protein from the Luminous Hydromedusan, Aequorea. Journal of Cellular and Comparative Physiology, 59: 223-239.
Silva ON, Mulder KCL, Barbosa AEaD, Otero-Gonzalez AJ, Lopez-Abarrategui C, Rezende TMB, Dias SC, Franco OL (2011) Exploring the pharmacological potential of promiscuous host-defense peptides: From natural screenings to biotechnological applications. Frontiers in Microbiology, 2: 1-14.
Simeone MC, Lafiandra D (2005) Isolation and characterisation of friabilin genes in rye. Journal of Cereal Science, 41: 115-122.
Simmonds D, Barlow K, Wrigley C (1973) Biochemical basis of grain hardness in wheat. Cereal Chemistry, 50: 553-562.
Skosyrev VS, Rudenko NV, Yakhnin AV, Zagranichny VE, Popova LI, Zakharov MV, Gorokhovatsky AY, Vinokurov LM (2003) EGFP as a fusion partner for the expression and organic extraction of small polypeptides. Protein Expression and Purification, 27: 55-62.
Slavin J (2003) Why whole grains are protective: Biological mechanisms. Proceedings of the Nutrition Society, 62: 129-134.
Smyth DR, Mrozkiewicz MK, Mcgrath WJ, Listwan P, Kobe B (2003) Crystal structures of fusion proteins with large-affinity tags. Protein Science, 12: 1313-1322.

References
- 194 -
Sorrentino A, Iannaccone M, Palumbo D, Capparelli R, Porta R, Mariniello L (2009) Tobacco (Nicotiana tabacum cv. Bright Yellow 2) cells as an effective bioreactor for the production of puroindolines. Biotechnology and Applied Biochemistry, 53: 193-199.
Stoger E, Sack M, Nicholson L, Fischer R, Christou P (2005) Recent progress in plantibody technology. Current Pharmaceutical Design, 11: 2439-2457.
Stulemeijer IJE, Joosten MHJ (2008) Post-translational modification of host proteins in pathogen-triggered defence signalling in plants. Molecular Plant Pathology, 9: 545-560.
Swan CG, Bowman JGP, Martin JM, Giroux MJ (2006a) Increased puroindoline levels slow ruminal digestion of wheat (Triticum aestivum L.) starch by cattle. Journal of Animal Science, 84: 641-650.
Swan CG, Meyer FD, Hogg AC, Martin JM, Giroux MJ (2006b) Puroindoline B limits binding of puroindoline A to starch and grain softness. Crop Science, 46: 1656-1665.
Symes KJ (1965) The inheritance of grain hardness in wheat as measured by the particle size index. Australian Journal of Agricultural Research, 16: 113-123.
Symes KJ (1969) Influence of a gene causing hardness on the milling and baking quality of two wheats. Australian Journal of Agricultural Research, 20: 971-979.
Taheri P, Tarighi S (2011) Cytomolecular aspects of rice sheath blight caused by Rhizoctonia solani. European Journal of Plant Pathology, 129: 511-528.
Tanaka H, Morris CF, Haruna M, Tsujimoto H (2008) Prevalence of puroindoline alleles in wheat varieties from eastern Asia including the discovery of a new SNP in puroindoline b. Plant Genetic Resources: Characterisation and Utilisation, 6: 142-152.
Terpe K (2003) Overview of tag protein fusions: From molecular and biochemical fundamentals to commercial systems. Applied Microbiology and Biotechnology, 60: 523-533.

References
- 195 -
Terras FRG, Eggermont K, Kovaleva V, Raikhel NV, Osborn RW, Kester A, Rees SB, Torrekens S, Van Leuven F, Vanderleyden J, Cammue BPA, Broekaert WF (1995) Small cysteine-rich antifungal proteins from radish: Their role in host defense. The Plant Cell, 7: 573-588.
Thomas DR, Penney CA, Majumder A, Walmsley AM (2011) Evolution of plant-made pharmaceuticals. International Journal of Molecular Sciences, 12: 3220-3236.
Tranquilli G, Heaton J, Chicaiza O, Dubcovsky J (2002) Substitutions and deletions of genes related to grain hardness in wheat and their effect on grain texture. Crop Science, 42: 1812-1817.
Tremblay R, Feng M, Menassa R, Huner NPA, Jevnikar AM, Ma S (2011) High-yield expression of recombinant soybean agglutinin in plants using transient and stable systems. Transgenic Research, 20: 345-356.
Turnbull KM, Rahman S (2002) Endosperm texture in wheat. Journal of Cereal Science, 36: 327-337.
Turnbull KM, Turner M, Mukai Y, Yamamoto M, Morell MK, Rahman S, Appels R (2003) The organization of genes tightly linked to the Ha locus in Aegilops tauschii, the D-genome donor to wheat. Genome, 46: 330-338.
Twyman RM (2004) Host plants, systems and expression strategies for molecular farming. In: Fischer R and Schillberg S (eds.) Molecular Farming. Wiley-VCH Verlag GmbH & Co. KGaA,Weinheim, Germany. pp 191-216.
Twyman RM, Stoger E, Schillberg S, Christou P, Fischer R (2003) Molecular farming in plants: Host systems and expression technology. Trends in Biotechnology, 21: 570-578.
Vaghchhipawala Z, Rojas CM, Senthil-Kumar M, Mysore KS (2011) Agroinoculation and agroinfiltration: simple tools for complex gene function analyses. Methods in Molecular Biology. 678: 65-76.
Ventura S (2011) Bimolecular fluorescence complementation: illuminating cellular protein interactions. Current Molecular Medicine, 11: 582-598.
Vitale A, Pedrazzini E (2005) Recombinant pharmaceuticals from plants: The plant endomembrane system as bioreactor. Molecular Interventions, 5: 216-225.

References
- 196 -
Voinnet O, Rivas S, Mestre P, Baulcombe D (2003) An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. The Plant Journal, 33: 949-956.
Waadt R, Schlücking K, Schroeder JI, Kudla J (2014) Protein fragment bimolecular fluorescence complementation analyses for the in vivo study of protein-protein interactions and cellular protein complex localizations. In: Jose J. Sanchez-Serrano and Julio Salinas (eds.) Arabidopsis Protocols, Methods in Molecular Biology. Springer Science, New York, USA. pp 629-658.
Waadt R, Schmidt LK, Lohse M, Hashimoto K, Bock R, Kudla J (2008) Multicolor bimolecular fluorescence complementation reveals simultaneous formation of alternative CBL/CIPK complexes in planta. The Plant Journal, 56: 505-516.
Wada N, Kajiyama S, Cartagena JA, Lin L, Akiyama Y, Otani M, Suzuki G, Mukai Y, Aoki N, Fukui K (2010) The effects of puroindoline b on the ultrastructure of endosperm cells and physicochemical properties of transgenic rice plant. Journal of Cereal Science, 51: 182-188.
Wall ML, Wheeler HL, Huebsch MP, Smith JC, Figeys D, Altosaar I (2010) The tryptophan-rich domain of puroindoline is directly associated with the starch granule surface as judged by tryptic shaving and mass spectrometry. Journal of Cereal Science, 52: 115-120.
Walter M, Chaban C, Schütze K, Batistic O, Weckermann K, Näke C, Blazevic D, Grafen C, Schumacher K, Oecking C, Harter K, Kudla J (2004) Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. The Plant Journal, 40: 428-438.
Wandelt CI, Khan MRI, Craig S, Schroeder HE, Spencer D, Higgins TJV (1992) Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants. The Plant Journal, 2: 181-192.
Wang L, Webster DE, Campbell AE, Dry IB, Wesselingh SL, Coppel RL (2008) Immunogenicity of Plasmodium yoelii merozoite surface protein 4/5 produced in transgenic plants. International Journal for Parasitology, 38: 103-110.
Wang S, Hazelrigg T (1994) Implications for bcd mRNA localization from spatial distribution of exu protein in Drosophila oogenesis. Nature, 369: 400-403.

References
- 197 -
Wanjugi HW, Hogg AC, Martin JM, Giroux MJ (2007) The role of puroindoline A and B individually and in combination on grain hardness and starch association. Crop Science, 47: 67-76.
Ward TH (2006) The uses of green fluorescent protein in mammalian cells. In: Chalfie M and Kain SR (eds.) Green Fluorescent Protein: Properties, Applications, and Protocols. Second ed. John Wiley & Sons, Inc. Toronto, Canada. pp 305-337.
Waugh DS (2005) Making the most of affinity tags. Trends in Biotechnology, 23: 316-320.
Webster DE, Thomas MC (2012) Post-translational modification of plant-made foreign proteins; glycosylation and beyond. Biotechnology Advances, 30: 410-418.
Webster DE, Wang L, Mulcair M, Ma C, Santi L, Mason HS, Wesselingh SL, Coppel RL (2009) Production and characterization of an orally immunogenic Plasmodium antigen in plants using a virus-based expression system. Plant Biotechnology Journal, 7: 846-855.
Werner S, Breus O, Symonenko Y, Marillonnet S, Gleba Y (2011) High-level recombinant protein expression in transgenic plants by using a double-inducible viral vector. Proceedings of the National Academy of Sciences of the United States of America, 108: 14061-14066.
Wiegand I, Hilpert K, Hancock REW (2008) Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature Protocols, 3: 163-175.
Wiley PR, Tosi P, Evrard A, Lovegrove A, Jones HD, Shewry PR (2007) Promoter analysis and immunolocalisation show that puroindoline genes are exclusively expressed in starchy endosperm cells of wheat grain. Plant Molecular Biology, 64: 125-136.
Wilken LR, Nikolov ZL (2012) Recovery and purification of plant-made recombinant proteins. Biotechnology Advances, 30: 419-433.
Wilkinson M, Wan Y, Tosi P, Leverington M, Snape J, Mitchell RaC, Shewry PR (2008) Identification and genetic mapping of variant forms of puroindoline b expressed in developing wheat grain. Journal of Cereal Science, 48: 722-728.

References
- 198 -
Wimley WC (2010) Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chemical Biology, 5: 905-917.
Witzel K, Shahzad M, Matros A, Mock HP, Mühling KH (2011) Comparative evaluation of extraction methods for apoplastic proteins from maize leaves. Plant Methods, 7: 48-58.
Wrigley C, Corke H, C.E. W (2004) Encyclopedia of grain science, Elsevier Academic Press, Massachusetts, USA. ISBN: 978-0-12-765490-4.
Wrigley CW (2009) Wheat: A unique grain for the world. In: Khan K and Shewry PR (eds.) Wheat Chemistry and Technology. 4th ed. AACC International, Inc, St. Paul, Minnesota, USA. pp 1-17.
Wroblewski T, Tomczak A, Michelmore R (2005) Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnology Journal, 3: 259-273.
Xia L, Chen F, He Z, Chen X, Morris CF (2005) Occurrence of puroindoline alleles in Chinese winter wheats. Cereal Chemistry, 82: 38-43.
Xia L, Geng H, Chen X, He Z, Lillemo M, Morris CF (2008) Silencing of puroindoline a alters the kernel texture in transgenic bread wheat. Journal of Cereal Science, 47: 331-338.
Yang F, Moss LG, Phillips Jr GN (1996) The molecular structure of green fluorescent protein. Nature Biotechnology, 14: 1246-1251.
Yang SJ, Carter SA, Cole AB, Cheng NH, Nelson RS (2004) A natural variant of a host RNA-dependent RNA polymerase is associated with increased susceptibility to viruses by Nicotiana benthamiana. Proceedings of the National Academy of Sciences of the United States of America, 101: 6297-6302.
Yevtushenko DP, Misra S (2007) Comparison of pathogen-induced expression and efficacy of two amphibian antimicrobial peptides, MsrA2 and temporin A, for engineering wide-spectrum disease resistance in tobacco. Plant Biotechnology Journal, 5: 720-734.
Zhang J, Martin JM, Balint-Kurti P, Huang L, Giroux MJ (2011) The wheat puroindoline genes confer fungal resistance in transgenic corn. Journal of Phytopathology, 159: 188-190.

References
- 199 -
Zhang J, Martin JM, Beecher B, Lu C, Hannah LC, Wall ML, Altosaar I, Giroux MJ (2010a) The ectopic expression of the wheat Puroindoline genes increase germ size and seed oil content in transgenic corn. Plant Molecular Biology, 74: 353-365.
Zhang J, Martin JM, Beecher B, Morris CF, Curtis Hannah L, Giroux MJ (2009) Seed-specific expression of the wheat puroindoline genes improves maize wet milling yields. Plant Biotechnology Journal, 7: 733-743.
Zhang R, Cao Y, Wang X, Feng Y, Chen P (2010b) Development and characterization of a Triticum aestivum-H. villosa T5VS-5DL translocation line with soft grain texture. Journal of Cereal Science, 51: 220-225.
Zhang Y (2008) I-TASSER server for protein 3D structure prediction. BMC Bioinformatics, 9: 40-47.
Ziemann M, Ramalingam A, Bhave M (2008) Evidence of physical interactions of puroindoline proteins using the yeast two-hybrid system. Plant Science, 175: 307-311.

Appendices
- 200 -
Appendices
Appendix I
Table I.1 Primer used for deletion of HD in PINA and PINB; TRD in PINA
* F: Forward primer; R: Reverse primer
Primer* Sequence (5’ to 3’) Application PINA∆TRD-F
PINA∆TRD-R
5’-atgaaggatGGTTGTCAAGAGCTCCTTGG-3’ Deletion of TRD in PINA (position 34-46) 5’-ttgacaaccATCCTTCATCGTTGAGCATC-3’
PINA∆HD-F
PINA∆HD-R
5’-caggggtcaCAGCGTGATCGGGCAAGCAA-3’ Deletion of hydrophobic domain (HD) in PINA (position 75-85) 5’-atcacgctgTGACCCCTGGATGATGTTGC-3’
PINB∆HD-F PINB∆HD-R
5’-atccggcgaTTGGGCATTTGGCGAGGTGA-3’ Deletion of hydrophobic domain (HD) in PINB (position 75-83) 5’-aatgcccaaTCGCCGGATAGAATCACAGC-3’

Appendices
- 201 -
Appendix II
Figure II.1 pICH-PINA expressions constructs. DNA sequence of mature putative Pina-D1a (DQ363911, line1) aligned with the sequence of pICH-PINA-His clone (line 2) and with the sequence of pICH-PINA-His-SEKDEL clone (line 3). His-tag and SEKDEL site underlined.
Figure II.2 pICH-PINA expressions construct. Amino acid sequence of mature putative PINA (line1) aligned with the sequence of pICH-PINA-His clone (line 2) with sequence of pICH-PINA-His-SEKDEL clone (line 3).
10 20 30 40 50 60. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature ~ ~ ~ ~ ~ ~ ~ ~ ~ G A T G T T G C T G G C G G G G G T G G T G C T C A A C A A T G C C C T G T A G A G A C A A A G C T ApICH-PINA-His A A T T C G A T G G A T G T T G C T G G C G G G G G T G G T G C T C A A C A A T G C C C T G T A G A G A C A A A G C T ApICH-PINA-His-SEKDEL A A T T C G A T G G A T G T T G C T G G C G G G G G T G G T G C T C A A C A A T G C C C T G T A G A G A C A A A G C T A
70 80 90 100 110 120. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature A A T T C A T G C A G G A A T T A C C T G C T A G A T C G A T G C T C A A C G A T G A A G G A T T T C C C G G T C A C CpICH-PINA-His A A T T C A T G C A G G A A T T A C C T G C T A G A T C G A T G C T C A A C G A T G A A G G A T T T C C C G G T C A C CpICH-PINA-His-SEKDEL A A T T C A T G C A G G A A T T A C C T G C T A G A T C G A T G C T C A A C G A T G A A G G A T T T C C C G G T C A C C
130 140 150 160 170 180. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature T G G C G T T G G T G G A A A T G G T G G A A G G G A G G T T G T C A A G A G C T C C T T G G G G A G T G T T G C A G TpICH-PINA-His T G G C G T T G G T G G A A A T G G T G G A A G G G A G G T T G T C A A G A G C T C C T T G G G G A G T G T T G C A G TpICH-PINA-His-SEKDEL T G G C G T T G G T G G A A A T G G T G G A A G G G A G G T T G T C A A G A G C T C C T T G G G G A G T G T T G C A G T
190 200 210 220 230 240. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature C G G C T C G G C C A A A T G C C A C C G C A A T G C C G C T G C A A C A T C A T C C A G G G G T C A A T C C A A G G CpICH-PINA-His C G G C T C G G C C A A A T G C C A C C G C A A T G C C G C T G C A A C A T C A T C C A G G G G T C A A T C C A A G G CpICH-PINA-His-SEKDEL C G G C T C G G C C A A A T G C C A C C G C A A T G C C G C T G C A A C A T C A T C C A G G G G T C A A T C C A A G G C
250 260 270 280 290 300. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature G A T C T C G G T G G C A T C T T C G G A T T T C A G C G T G A T C G G G C A A G C A A A G T G A T A C A A G A A G C CpICH-PINA-His G A T C T C G G T G G C A T C T T C G G A T T T C A G C G T G A T C G G G C A A G C A A A G T G A T A C A A G A A G C CpICH-PINA-His-SEKDEL G A T C T C G G T G G C A T C T T C G G A T T T C A G C G T G A T C G G G C A A G C A A A G T G A T A C A A G A A G C C
310 320 330 340 350 360. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature A A G A A C C T G C C G C C C A G G T G C A A C C A G G G C C C T C C C T G C A A C A T C C C C G G C A C T A T T G G CpICH-PINA-His A A G A A C C T G C C G C C C A G G T G C A A C C A G G G C C C T C C C T G C A A C A T C C C C G G C A C T A T T G G CpICH-PINA-His-SEKDEL A A G A A C C T G C C G C C C A G G T G C A A C C A G G G C C C T C C C T G C A A C A T C C C C G G C A C T A T T G G C
370 380 390 400 410. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | .
Pina-Mature T A T T A C T G G T G A pICH-PINA-His T A T T A C T G G C A C C A T C A C C A T C A C C A T T G A G G A T C C pICH-PINA-His-SEKDEL T A T T A C T G G C A C C A T C A C C A T C A C C A T T C T G A G A A A G A T G A G C T A T G A G G A
10 20 30 40 50 60. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature ~ ~ ~ D V A G G G G A Q Q C P V E T K L N S C R N Y L L D R C S T M K D F P V T W R WW K WW K G G C Q E L L G E C C SpICH-PINA-His N S M D V A G G G G A Q Q C P V E T K L N S C R N Y L L D R C S T M K D F P V T W R WW K WW K G G C Q E L L G E C C SpICH-PINA-His-SEKDEL N S M D V A G G G G A Q Q C P V E T K L N S C R N Y L L D R C S T M K D F P V T W R WW K WW K G G C Q E L L G E C C S
70 80 90 100 110 120. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Pina-Mature R L G Q M P P Q C R C N I I Q G S I Q G D L G G I F G F Q R D R A S K V I Q E A K N L P P R C N Q G P P C N I P G T I GpICH-PINA-His R L G Q M P P Q C R C N I I Q G S I Q G D L G G I F G F Q R D R A S K V I Q E A K N L P P R C N Q G P P C N I P G T I GpICH-PINA-His-SEKDEL R L G Q M P P Q C R C N I I Q G S I Q G D L G G I F G F Q R D R A S K V I Q E A K N L P P R C N Q G P P C N I P G T I G
130. . . . | . . . . | . . . . | . .
Pina-Mature Y YW * pICH-PINA-His Y YW H H H H H H * G S pICH-PINA-His-SEKDEL Y YW H H H H H H S E K D E L * G

Appendices
- 202 -
Figure II.3 pICH-PINB expressions construct. DNA sequence of mature putative PINB aligned with the sequence of pICH-PINB-His clone (line 2) and with sequence of pICH-PINB-His-SEKDEL clone (line 3). His-tag and SEKDEL site underlined.
Figure II.4 pICH-PINB-His expressions construct. Amino acid sequence of mature putative PINB (line1) aligned with the sequence of pICH-PINB-His clone (line 2) with sequence of pICH-PINB-His-SEKDEL clone (line 3).
10 20 30 40 50 60. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature ~ ~ ~ ~ ~ ~ G A A G T T G G C G G A G G A G G T G G T T C T C A A C A A T G T C C G C A G G A G C G G C C G A A G C T ApICH-PINB-His A C C A T G G A A G T T G G C G G A G G A G G T G G T T C T C A A C A A T G T C C G C A G G A G C G G C C G A A G C T ApICH-PINB-His-SEKDEL T C G A T G G A A G T T G G C G G A G G A G G T G G T T C T C A A C A A T G T C C G C A G G A G C G G C C G A A G C T A
70 80 90 100 110 120. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature A G C T C T T G C A A G G A T T A C G T G A T G G A G C G A T G T T T C A C A A T G A A G G A T T T T C C A G T C A C CpICH-PINB-His A G C T C T T G C A A G G A T T A C G T G A T G G A G C G A T G T T T C A C A A T G A A G G A T T T T C C A G T C A C CpICH-PINB-His-SEKDEL A G C T C T T G C A A G G A T T A C G T G A T G G A G C G A T G T T T C A C A A T G A A G G A T T T T C C A G T C A C C
130 140 150 160 170 180. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature T G G C C C A C A A A A T G G T G G A A G G G C G G C T G T G A G C A T G A G G T T C G G G A G A A G T G C T G C A A GpICH-PINB-His T G G C C C A C A A A A T G G T G G A A G G G C G G C T G T G A G C A T G A G G T T C G G G A G A A G T G C T G C A A GpICH-PINB-His-SEKDEL T G G C C C A C A A A A T G G T G G A A G G G C G G C T G T G A G C A T G A G G T T C G G G A G A A G T G C T G C A A G
190 200 210 220 230 240. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature C A G C T G A G C C A G A T A G C A C C A C A A T G T C G C T G T G A T T C T A T C C G G C G A G T G A T C C A A G G CpICH-PINB-His C A G C T G A G C C A G A T A G C A C C A C A A T G T C G C T G T G A T T C T A T C C G G C G A G T G A T C C A A G G CpICH-PINB-His-SEKDEL C A G C T G A G C C A G A T A G C A C C A C A A T G T C G C T G T G A T T C T A T C C G G C G A G T G A T C C A A G G C
250 260 270 280 290 300. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature A G G C T C G G T G G C T T C T T G G G C A T T T G G C G A G G T G A G G T A T T C A A A C A A C T T C A G A G G G C CpICH-PINB-His A G G C T C G G T G G C T T C T T G G G C A T T T G G C G A G G T G A G G T A T T C A A A C A A C T T C A G A G G G C CpICH-PINB-His-SEKDEL A G G C T C G G T G G C T T C T T G G G C A T T T G G C G A G G T G A G G T A T T C A A A C A A C T T C A G A G G G C C
310 320 330 340 350 360. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature C A G A G C C T C C C C T C A A A G T G C A A C A T G G G C G C C G A C T G C A A G T T C C C T A G T G G C T A T T A CpICH-PINB-His C A G A G C C T C C C C T C A A A G T G C A A C A T G G G C G C C G A C T G C A A G T T C C C T A G T G G C T A T T A CpICH-PINB-His-SEKDEL C A G A G C C T C C C C T C A A A G T G C A A C A T G G G C G C C G A C T G C A A G T T C C C T A G T G G C T A T T A C
370 380 390 400. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . .
PINB-Mature T G G T G A pICH-PINB-His T G G C A C C A T C A C C A T C A C C A T T G A C T C G A G G C pICH-PINB-His-SEKDEL T G G C A C C A T C A C C A T C A C C A T T C T G A G A A A G A T G A G C T A T G A G G A T C C
10 20 30 40 50 60. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature ~ ~ E V G G G G G S Q Q C P Q E R P K L S S C K D Y V M E R C F T M K D F P V T W P T K WW K G G C E H E V R E K C C KpICH-PINB-His T M E V G G G G G S Q Q C P Q E R P K L S S C K D Y V M E R C F T M K D F P V T W P T K WW K G G C E H E V R E K C C KpICH-PINB-His-SEKDEL S M E V G G G G G S Q Q C P Q E R P K L S S C K D Y V M E R C F T M K D F P V T W P T K WW K G G C E H E V R E K C C K
70 80 90 100 110 120. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
PINB-Mature Q L S Q I A P Q C R C D S I R R V I Q G R L G G F L G I W R G E V F K Q L Q R A Q S L P S K C N M G A D C K F P S G Y YpICH-PINB-His Q L S Q I A P Q C R C D S I R R V I Q G R L G G F L G I W R G E V F K Q L Q R A Q S L P S K C N M G A D C K F P S G Y YpICH-PINB-His-SEKDEL Q L S Q I A P Q C R C D S I R R V I Q G R L G G F L G I W R G E V F K Q L Q R A Q S L P S K C N M G A D C K F P S G Y Y
130. . . . | . . . . | . . . . | .
PINB-Mature W * pICH-PINB-His W H H H H H H * L E pICH-PINB-His-SEKDEL W H H H H H H S E K D E L * G S

Appendices
- 203 -
Appendix III Protein identification using MALDI-TOF
To obtain an accurate mass for recombinant PINs, and further investigate multimer
formation, MALDI-TOF analysis was done. The result was inconclusive as data
provide unknown peptides sequence.
The protocol for MALDI-TOF was based on Shevchenko et al. (2007) with minor
modification and was applied to SDS-PAGE bands of interest at various stages of
purification. Coommassie stained gels were used for protein identification. Protein
bands of PINs resolved on SDS-PAGE were excised from the gel using a sterile scalpel
and transferred into microcentrifuge tubes. The gel plugs were first added in 500 µL of
neat acetonitrile by incubating at 37°C for 10 minutes until the gel pieces shrank. The
gel plug solution was removed after centrifugation at 10,000×g, then 100 µL DTT was
added to cover the gel pieces and incubated for 30 minutes at 56°C. The tubes were
allowed to cool down to room temperature for 10 minutes and solution was removed.
50 µL of iodoacetamide solution added to tubes and incubated for 20 minutes at RT in
dark. Then 500 µL of acetonitrile were added and incubated for 10 minutes. Then 100
µL of solution (50% v/v acetonitrile and 50% v/v 100 mM ammonium bicarbonate) was
added and tubes incubated at RT with occasional vortex for 2 hours. 50 µL of the
trypsin solution (Biolabs, Australia) was added to each sample then added 20 µL
ammonium bicarbonate buffer (100mM) and incubate at 37°C for overnight. The next
day tubes were incubated at room temperature and the gel pieces spun down. Then 100
µL extraction buffer (5% formic acid: acetonitrile 1:1 (v/v)) was added to each tube and
incubated for 15 minutes at 37°C on a shaker. The supernatant was collected into new
tube and dried down in vacuum centrifuge for 2 hours at 50°C. Samples were
resuspended in trifluoroacetic acid and then with equal volume of saturated α-Cyano-4-
hydroxycinnamic acid (CHCA) matrix was spotted onto target plate of Axima MALDI-
TOF mass spectrometer (Shimadzu-Biotech, Japan). Samples were analysed using the
Shimadzu MALDI-MS software. A total of four sample spots (1 μL each) were
analysed for each aliquot taken.

Appendices
- 204 -
Appendix IV
Figure III.1 Partial sequence of pNBV constructs. A: pNBV-gene-YC; B: pNBV-gene-YN 35S promoter highlighted in gray. The two NotI (GCGGCCGC) site highlighted in red; XbaI site [TCTAGA] and XmaI [CCCGGG] highlighted; DNA sequence of YC and YN highlighted in blue. Sequence of NosT and LacZ highlighted.
A
B

Appendices
- 205 -
Figure III.2 Partial Sequence of pNBV constructs. A: pNBV-YC-gene; B: pNBV-YN-gene 35S promoter highlighted in gary. The two NotI (GCGGCCGC) site highlighted in red; EcoRI site [GAATTC] and BamHI [GGATCC] highlighted; DNA sequence of YC and YN highlighted in blue. Sequence of NosT and LacZ highlighted.
A
B

Appendices
- 206 -
Appendix V Record of Proposed Notifiable Low Risk Dealing(s) (NLRDs):
(Regulation 13A (1) (a) - Gene Technology Regulations 2001 (as amended))
INSTRUCTIONS: Under r.13A(1)(c) of the Gene Technology Regulations 2001 (as amended), the Institutional Biosafety Committee (IBC) must provide a copy of this record to (i) the person or accredited organisation that requested the IBC to assess the proposed dealing; and (ii) the project supervisor for the proposed dealing. Under r.13A (2)(a)(i) an accredited organisation that proposed the dealing must include a copy of the IBC's record with the Accredited Organisation Annual Report to the Gene Technology Regulator. Under r.13A (2)(a)(ii) non-accredited organisations must provide a copy of this record to the Regulator at the end of the financial year.
NOTE: This form also captures information required by the Gene Technology Regulator to meet the requirements of r. 39 for NLRDs in maintaining a record of GMO and GM product dealings.
* References to NLRD types are the paragraph numbers relating to the kinds of dealing detailed in Parts 1 and 2 in Schedule 3 of the Gene Technology Regulations 2001 (as amended). These Regulations are available on the OGTR website <www.ogtr.gov.au>.
IBC NLRD Identifier (number) PC2-N26/10
Date of IBC Assessment 18 November 2010
Name of IBC Monash University IBC #309
Name of Organisation Conducting Dealing
Monash University
Project Title Production, characterisation and evaluation of recombinant puroindolines in plants
PC1 NLRD TYPE (a) – (c)*
PC2 NLRD TYPE (a) – (i)* (b) (ba) (c) (d)
Name(s) of Project Supervisor(s) Dr Diane Webster, Prof Mrinal Bhave
Has the IBC assessed the proposed dealing to be an NLRD?
Yes

Appendices
- 207 -
Publications arising from this work
Poster presentations:
Azadeh Niknejad, Diane Webster, Mrinal Bhave. (10th-14th February 2013);
Recombinant plant-made puroindolines exhibit anti-microbial activity and interaction
in plant system. 38th Lorne conference on protein structure and function, Lorne,
Victoria, Australia.
Azadeh Niknejad, Diane Webster, Mrinal Bhave. (11th July 2013); Investigating PIN
proteins interactions in a plant system using Bimolecular Fluorescence
Complementation (BiFC). 12th Melbourne protein group student symposium, La Trobe
University, Melbourne, Victoria, Australia.
Manuscripts:
Azadeh Niknejad, Diane Webster, Mrinal Bhave. Transient expression and
characterization of puroindoline proteins in Nicotiana benthamiana using a virus-
based expression system. (in preparation)
Azadeh Niknejad, Diane Webster, Mrinal Bhave. Evidence of physical interactions of
puroindoline proteins in living plant cells using bimolecular fluorescence
complementation (BiFC) analysis. (in preparation)