water transport in onion (allium cepa 1.) roots' - plant physiology
TRANSCRIPT

Plant Physiol. (1993) 101: 1305-1315
Water Transport in Onion (Allium cepa 1.) Roots'
Changes of Axial and Radial Hydraulic Conductivities during Root Development
Walter Melchior and Ernst Steudle* Lehrstuhl für Pflanzenokologie, Universitat Bayreuth, Postfach 1 O1 251, D-8580 Bayreuth, Germany
The hydraulic architecture of developing onion (Allium cepa L. cv Calypso) roots grown hydroponically was determined by meas- uring axial and radial hydraulic conductivities (equal to inverse of specific hydraulic resistances). In the roots, Casparian bands and suberin lamellae develop in the endodermis and exodermis (equal to hypodermis). Using the root pressure probe, changes of hy- draulic conductivities along the developing roots were analyzed with high resolution. Axial hydraulic conductivity (L,) was also calculated from stained cross-sections according to Poiseuille's law. Near the base and the tip of the roots, measured and calculated 1, values were similar. However, at distances between 200 and 300 mm from the apex, measured values of L, were smaller by more than 1 order of magnitude than those calculated, probably because of remaining cross walls between xylem vessel members. During development of root xylem, L. increased by 3 orders of magnitude. In the apical 30 mm (tip region), axial resistance limited water transport, whereas in basal parts radial resistances (low radial hydraulic conductivity, Lp,) controlled the uptake. Because of the high axial hydraulic resistance in the tip region, this zone appeared to be "hydraulically isolated from the rest of the root. Changes of the Lp. of the roots were determined by measuring the hydraulic conductance of roots of different length and referring these data to unit surface area. At distances between 30 and 150 mm from the root tip, Lp, was fairly constant (1.4 X lO-' m s-' MPa-'). In more basal root zones, Lp, was considerably smaller and varied between roots. The low contribution of basal zones to the overall water uptake indicated an influence of the exodermal Casparian bands and/or suberin lamellae in the endodermis or exodermis, which develop at distances larger than 50 to 60 mm from the root tip.
Water transport through plants is governed by water po- tential differences and hydraulic conductances. Both the con- ductances and the driving forces may vary greatly in short or long time scales. For roots, it has been reported that different root zones contribute to different extents to the overall uptake of water (Rosene, 1937; Brouwer, 1954; Sanderson, 1983; Haussling et al., 1988; Frensch and Steudle, 1989; North and Nobel, 1991). On the other hand, the L, may strongly depend on environmental factors such as high salinity and drought of the soil or anoxia (Azaizeh and Steudle, 1991; Azaizeh et al., 1992; Cruz et al., 1992; Birner and Steudle, 1993). Fur- thermore, L, may be a function of the flow rate (Weatherley,
Supported by a grant from the Deutsche Forschungsgemein- schaft, Sonderforschungsbereich 137.
* Corresponding author; fax 49-921-552564. 1305
1982; Passioura, 1988; Steudle, 1992). Variable hydraulic root resistance is closely associated with changes of root structure and the pathways used for water movement in the root, both for axial transport in vessels and for the radial movement across the root cylinder.
During root development, L, increases due to the break- down of cross walls between vessel members. The main radial hydraulic resistance of the root often has been attributed to the endodermis or exodermis, where hydrophobic deposits block the transport of water and ions in the cell walls (Pas- sioura, 1988; Peterson, 1989). Therefore, a dependency on the state of development of endodermis or exodermis could be expected, and water uptake should occur mainly in the region near the root tip (Boyer, 1985). However, a substantial water uptake in zones with suberized endodermis (state 11) was reported for barley roots (Sanderson, 1983), which coq- tradicted the experiments of Robards et al. (1973). In the former study, water flow declined as the number of endo- dermal cells in state 111, i.e. cells exhibiting thick layers of cellulose, increased.
Recently, Peterson et al. (1993) demonstrated that the state I endodermis (with a Casparian band) was hardly a main barrier to radial water flow in maize roots. The authors dissected parts of the root cortex or punctured the endodermis with narrow tips of microcapillaries to characterize different resistances on the radial path of water. It was found that, different from solute permeability, the water permeability (hydraulic conductivity) was evenly distributed within the living tissue.
The recently developed root pressure probe has been em- ployed to measure both Lp, and L, of maize roots grown hydroponically (Frensch and Steudle, 1989). It allows the resolution of both parameters with higher precision along the root than other techniques available. Frensch and Steudle (1989) showed that the contribution of axial hydraulic resist- ance to the overall resistance was not important except for the apical 15 mm where early metaxylem was not mature. In more basal parts, Lp, was constant at distances between 20
Abbreviations: A,, root surface area (mm'); I , length of root end segment (mm); L,, axial hydraulic conductance (m3 s-' MPa-'); Lp,, radial hydraulic conductivity (m s-' MPa-I); L,, hydraulic conduct- ance (m3 s-' MPa-'); L,, axial hydraulic conductivity (m' s-' MPa-'); n, number of measurements; N, number of roots measured; P,, root pressure; P,,, stationary root pressure; z, distance from the root tip ("1.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

1306 Melchior and Steudle Plant Physiol. Vol. 101, 1993
and 120 mm from the root tip. Because of the development of laterals, the authors could not measure Lp, over larger distances where Casparian bands and suberin lamellae in the exodermis occurred (Perumalla and Peterson, 1986).
In this study, the pressure probe technique has been ap- plied to onion (Allium cepa L.) roots, which show an anatomy somewhat different from maize (St. Aubin et al., 1986; Frensch and Steudle, 1989). Onion roots are unbranched over long distances (up to 400 mm from the apex). When grown in hydroponics, this species develops Casparian bands in the exodermis and suberin lamellae in endodermis and exodermis at distances from the root tip of z > 50 to 60 mm, i.e. earlier than in roots of maize. Compared with maize, the large vessels of the late metaxylem mature closer to the root tip in a zone where no laterals have crossed the outer root tissue. Thus, onion roots should be better suited to follow changes of hydraulic properties of roots during development.
The dependency of both the Lp, and L, on the position along the root was studied. The measured L, was compared with that calculated from vessel dimensions obtained from cross-sections according to Poiseuille’s law. These data on changes of root hydraulics, i.e. the functions Lp,(z) and L,(z), are needed to create a detailed modeling of water transport across roots.
MATERIALS A N D METHODS
Plant Material
Bulbs of onion (Allium cepa L. cv Calypso) obtained from Vanderhave Seeds (NL-4410AA Rilland, The Netherlands), were kept in polyethylene bags for 9 d at 6OC. They were then grown for 3 d in vermiculite moistened with one-third Miller solution (Miller, 1980. Composition in mo1 m-3: K* 1.0, Ca2+ 0.5, Mg2+ 0.67, NH4+ 1.33, N03- 2.67, P043- 0.5, S042- 0.5. Micronutrients in mmol m-3: C1- 50, Fe3+ 36 and Na+ 36, added as FeNa-EDTA, B033- 25, Mn2+ 2, Zn2+ 2, Cu2+ 0.5, Mo0d2- 0.5. Osmotic concentration 7 mOsmol kg-’; pH 6.0, adjusted with sulfuric acid). After this time, each bulb was transferred to hydroculture in a nontransparent 15-L polyvinyl chloride container with aerated one-third Miller solution. Nutrient solution was exchanged every 7 d. A11 plants were grown in a climatic chamber at a lightldark rhythm of 14/10 h and a PAR of 500 to 650 pmol m-2 s-‘. The temperature was kept at 20 to 21OC and the RH was 70 to 80% (water vapor pressure deficit = 4-7 Pa kPa-I).
Root Pressure Probe
After 6 to 22 d in hydroculture, roots were 140 to 510 mm in length. They were excised close to the bulb, transferred to a 1-L container with aerated nutrient solution, and fixed to a root pressure probe (Fig. 1). The roots were left on the probe for at least 24 h. Only those roots were taken that built up root pressures of larger than 0.2 MPa (maximum, 0.62 MPa). Apical parts of the roots that did not have visible laterals were used to avoid difficulties in determining root surface area. Experiments were carried out at a teinperature of 21.0 f 0.5OC. Only a few roots were used from each bulb (5-10 of about 50-100).
The root pressure probe connected the xylem of the cut
micrometer
Figure 1. Root pressure probe for measuring root pressure and hydraulic conductivity of roots. An excised root ora root segment open at both sides was connected tightly to the probe by a silicone seal to determine Lp, and L, of root segments. Root pressures were monitored continuously by a pressure transducer in the probe. Water flows were induced with the aid of a metal rod. The meniscus between water and silicone oil served as a reference to measure volume changes in the capillary (for further explanations, see text).
root to a pressure transducer (Fig. 1). Root pressure could be recorded continuously. The probe was filled with silicone oil and water so that a meniscus formed in the measuring capillary that was used as a point of reference. With the aid of a metal rod connected with a micrometer screw, volume changes were induced in the measuring system and responses of root pressure were monitored on a chart recorder. Volume changes were calculated from the i.d. of the measuring cap- illary (200 or 250 pm) and from the shift of the meniscus in the capillary. Severa1 step changes of pressure (volume) were produced in each direction to determine the elastic coefficient of the system (APr/AVs), i.e. the relation between pressure (APJ and volume changes (AVs). AP,/AVs ranged between 0.7 and 3.6 MPa mm-3 (7-36 bar/pL). Thus, changes of pressure with time measured by the root pressure probe (dP,/ dt) could be converted into water flows (dVJdt).
Measurement of 1,
To measure the L, of the root, a stationary root pressure was first established. Then the meniscus was shifted instan- taneously and held in the new position. This induced a volume (water) flow from the root xylem into the medium when P, was increased (“exosmotic”) or in the opposite direc- tion when P, was lowered (“endosmotic“). After the change, root pressure tended to reach a new stationary value follow- ing a response curve that was close to exponential with time (“pressure relaxation”). From the rate constant of the water- flow equilibration (kWr) and from the measured elasticity of the system (AP,/AVs), the L, in m3 s-’ MPa-’ was evaluated (Steudle and Jeschke, 1983; Steudle et al., 1987):
T,, = half time of water exchange of root. For details of the
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

Hydraulic Conductivity of Onion Roots 1307
calculation and for the background of the theory, see the references given. Although pressure relaxations were vir- tually exponential (f > 0.91, mean -+ SD = 0.986 & 0.011, n = 187, N = 28), the P,(t) curves appeared to incorporate other components. They could be, in fact, described as a sum of three exponential functions (Peterson and Steudle, 1993). The first phase, which has a very short time constant (4 s), has been interpreted to be due to an extensibility of the measuring system and/or to difficulties in rapidly adjusting the position of the meniscus. The last (third) phase was very slow. Because it did not appear in “osmotic experiments” when the water flow was much smaller, it was attributed to concentration polarization effects in the xylem (Peterson and Steudle, 1993). The very short and the very slow phases could be distinguished visually from the main body of the curves when digitizing the recorder strips with a digitizing tablet (DIGIKON, Kontron Registriertechnik, Eching, Ger- many) and transforming the P( t ) curves to semilog plots of In(P - P,) as a function of time, where P, is the end pressure reached after the relaxation.
In a preliminary experiment using a different onion culti- var, it could be shown that half-times of the second and the third phase were not correlated (f = 0.008, P = 0.58, n = 40, N = 10; half-times 3-16 s and 26-690 s, respectively). Fitting the sum of two exponential functions to the time courses, it was shown that the influence of the slower (third) phase on the determination of the half-time of the second phase could be neglected, because corrected and uncorrected values differed by only 1%. Thus, the evaluation of TrK after visual exclusion of the first and third phases was justified.
The absolute value of P, largely influenced the half-times calculated. Therefore, a fit program was used and the opti- mum P, for the second phase was calculated. Compared with the visual determination of P,, the fit resulted in changes of the rate constants (and L,) by factors of 0.72 to 2.42 (mean f SD = 1.43 f 0.26, n = 187, N = 28) and 0.35 to 2.00 (mean 2 SD = 1.15 f 0.19, n = 266, N = 31) in intact and open root segments, respectively.
Measurement of 1, Using the Root Pressure Probe
First Method
After excising the apical part (length > 45 mm) of a root fixed to the probe, root pressure fel1 to zero because of the opening of mature xylem. The L. was much larger than the radial so that the water flow induced across the remaining root segment still attached to the probe was dominated by the flow along the xylem and out of the cut vessels. Therefore, under these circumstances, the axial hydraulic conductivity per unit of root length ( L , = L, X e; where e= length of the remaining root segment) could be directly determined from pressure relaxations, neglecting the radial component of water flow. L, in m4 s-’ MPa-’ was calculated from:
In Equation 2, L, included the hydraulic resistance of the root segment in the seal that was determined separately (see
“Second Method”). Hence, the L, of the part of the root outside the seal could be also evaluated.
To eliminate effects of a reduced conductance at the seal during regular relaxations with intact roots (apical ends closed), it was ensured that, after cutting off the root at the seal, the half-time decreased by at least a factor of 10. The hydraulic resistances of the root segments remaining in the seal were subtracted from the total hydraulic resistances measured. It should be noted that L, represented the axial hydraulic conductivity per unit length of root in m4 s-’ MPa-’ (m3 m s-’ MPa-’), whereas L, was the hydraulic conductance in m3 s-’ MPa-I. Hence, the values of L, and L, cannot be compared directly.
Second Method
Using a different technique for determining L,, 18-mm root segments that were left in the seal of the equipment after cutting off the root right at the seal were measured (Frensch and Steudle, 1989). This technique avoided any radial com- ponent of water flow. It was used to determine L, as a function of the position along the root (L,[z]) since small segments of roots could be measured.
Evaluation of f, from the Anatomy of Root Xylem
Two staining procedures were used to identify mature, conducting xylem vessels on free-hand cross-sections. (a) The fluorescent staining technique of Brundrett et al. (1988) with berberine sulfate (1 g/L, exposure, 1 h) and aniline blue (5 g/L, 1 h) resulted in a bright staining of the lignified xylem vessels. With this method, Casparian bands in the endoder- mis and exodermis could be detected. Photographs were taken using a fluorescence microscope ‘(Zeiss, Oberkochen, Germany), exciting wavelengths 390 to 420 nm, dichroitic mirror FT 425, barrier filter LP 450 nm. (b) In a quicker procedure, toluidine blue was used for staining (0.3 g/L, 1 min). With the latter technique, cross-sections were photo- graphed under white light.
To determine the dimensions of mature vessels, the largest diameter and the diameter perpendicular to it of the cross- sections of stained xylem elements were digitized using a digitizing tablet. From the data, the L, per unit of length was calculated according to Poiseuille’s law:
(3)
for cylindrical vessels (ri = radius of the vessel; 1 = viscosity of water [9.7 X 10-” MPa s at 2l0C]). Because cross-sections of vessels were not circular, a mean radius (ri = m; a = largest radius and b = smallest radius) was used in Equation 3. Cross-sections could be better described as ellipses. There- fore, a different equation was used (Nonweiler, 1975):
1 Ai x m: L,=-Z-
D ki (4)
(A, cross-sectional area of vessel; mi, ”mean hydraulic radius”
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

1308 Melchior and Steudle Plant Physiol. Vol. 101, 1993
r
Figure 2. Freehand cross-sections of onion (A. cepa L.) roots grown hydroponically. Mature xylem was stained withtoluidine blue O (A) or with berberine sulfate and aniline blue (B-D). The latter technique also stained Casparian bandsin endodermis (B, arrows) and exodermis (C). A, Section taken at a z = 300 mm from the apex. Xylem vessels showed aradial arrangement with the late metaxylem (LMX) in the center. Except for the "passage cells" (PC, arrow), the endodermishad thickened cell walls in this state (Co, cortex). B, Mature xylem vessels and Casparian bands (CB) in the endodermis(En; z = 230 mm). C, Rhizodermis (Rh) and exodermis (Ex) 300 mm from the apex. In part, two parallel Casparian bandsare visible in the exodermis. D, Typical example of a cross-section in which some of the vessels (marked by "?") couldnot be classified as either mature or immature (z = 155 mm).
of vessel, j, which is given by Ai divided by the circumference;k{, coefficient depending on the geometry of the vessel.) Foran elliptical cross-section of the eccentricity t'i (= Va2 — b2/a),ki is given by (Nonweiler, 1975):
1 + (5)
In some cases, the staining did not clearly indicate whethera vessel was conducting or not. These vessels were digitizedseparately. This resulted in a certain error of the calculatedconductivity. Other errors (e.g. digitizing errors) could beneglected compared with this uncertainty.
RESULTS
Anatomy of Onion Roots
To evaluate Lx of the roots, stained cross-sections weretaken (see below). Staining procedures were used to identifylignified xylem vessels as well as thickenings of the endo-dermal cell walls (toluidine blue, Fig. 2A) and Casparianbands in the endodermis and exodermis (berberine sulfateand aniline blue, Fig. 2, B and C). It is known from theliterature that Casparian bands develop very early in theendodermis of onion roots even when plants are grownhydroponically (Perumalla and Peterson, 1986). In this study,
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

Hydraulic Conductivity of Onion Roots 1309
Casparian bands were already found in the endodermis 10 mm from the apex. Casparian bands in the exodennis (equal to the hypodermis) appeared at larger distances from the root tip (about 50 mm). The development of the endodermis and exodermis was not investigated systematically. However, it was found that Casparian bands did not appear simultane- ously in a11 exodermal cells. Rather, they first formed "patches." Therefore, on cross-sections, interruptions in the exodermal ring of Casparian bands were observed. With the two staining methods, it was not always possible to identify all xylem vessels as mature (i.e. as lignified) or not. An example of such a transition in the staining is shown in Figure 2D. This uncertainty resulted in the most important error in the determination of the L, from the cross-sections (see "Ma- terials and Methods" and below).
L ,
The hydraulic conductance measured on intact root seg- ments (apical ends closed) was composed of axial and radial conductances in series and in parallel, respectively. Water taken up radially will be continuously moved away in the
E 4 10-10 i o
1 o-"
í O - " U 3 2 n
3 I I O - ~
(d a 2 10-'O
.-I
o 10-" I
*E 10-12 U
I I I I
cross sections / berberine and aniline
C
I I I
vessels. Thus, the system should exhibit properties compa- rable to those of a leaky electric cable. It has been treated theoretically by Landsberg and Fowkes (1978) and extended by Frensch and Steudle (1989). At least for the very tip of the root where the xylem is yet immature, the axial resistance should contribute substantially to the overall hydraulic re- sistance or even should be the limiting component. On the other hand, the axial conductance for water flow across short root segments open at both ends is quite large if the xylem is mature so that it will dominate the total conductance. That is why the L, could be determined separately from the radial and why it is treated here first. From L, and from the total conductance of roots, the Lp, was then evaluated.
L,
L, per unit of root length (in m4 s-l MPa-') as calculated from the xylem vessel dimensions according to Poiseuille's law increased with increasing z in the apical 100 to 200 mm. L, ranged over more than 3 orders of magnitude (Fig. 3, A and B). At distances of about 200 mm from the apex, all xylem vessels including the central late metaxylem element
I I t I T
B
toluidine
D 1
a
root segments in seal
I I I I
1 o-0
10-'
1 0 - ' O
10-"
10-
10-'
10-'O
1 o-"
10-12
1 0-13 0 100 200 300 400 0 100 200 300 400
z ["I z ["I Figure 3. L, per unit of root length of onion roots as a function of z. Values were calculated from cross-sections according to Poiseuille's law (A and 6) or measured using the root pressure probe (C and D). L, values are presented in a logarithmic scale because they ranged over more than 3 orders of magnitude. The axial hydraulic resistances (logarithmic data) were fitted using the function R, = 1/L, = a X exp (-b X z) + c; a, b, and c > O (see Table I). A, Mature xylem stained with berberine sulfate/aniline blue (n = 101, N = 11). B, Sections stained with toluidine blue O (n = 89, N = 14). C, Short open root segments outside the seal were measured excluding the effect of the part of the root inside the seal (see D). D, Only the parts of roots inside the seal were measured, avoiding radial water flow, after cutting the root right at the seal. As in C, roots had been connected to the pressure probe for at least 24 h and root pressures were >0.2 MPa. Error bars in C and D represent the SD (n = 3-5 and 2-11, respectively). In A and B, the main error resulted from the uncertainty of identifying mature and immature vessels (see "Materials and Methods").
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022
1310 Melchior and Steudle Plant Physiol. Vol. 101, 1993
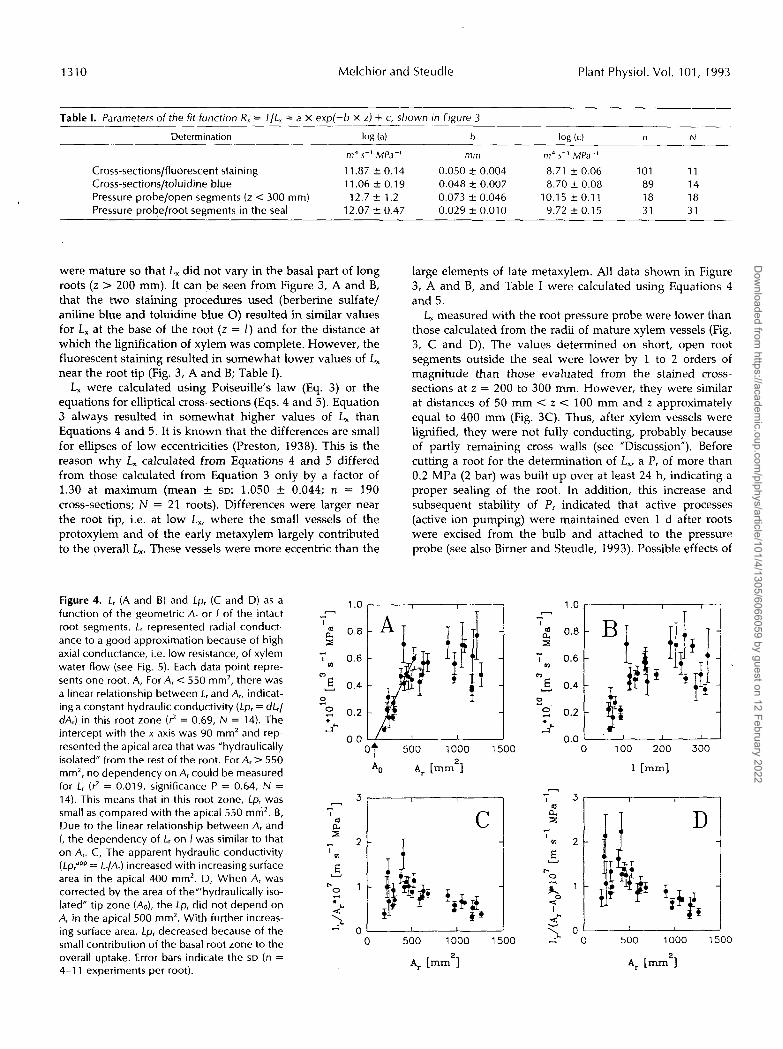
Table 1. Parameters of the fit function R, = I / L , = a x exp(-b X z) + c, shown in Figure 3
Determination log (a) I log (4 n N
m4 5-l MPa-’ mm m4 5-’ MPa-’
Cross-sections/fluorescent staining 11.87-+_0.14 0.050 rt 0.004 8.71 f 0.06 1 o1 11 Cross-sections/toluidine blue 11.06f0.19 0.048 2 0.007 8.70 k 0.08 89 14 Pressure probe/open segments ( L < 300 mm) 12.7 2 1.2 0.073 k 0.046 10.15 k 0.1 1 18 18 Pressure probelroot segments in the seal 12.07 -+_ 0.47 0.029 2 0.01 O 9.72 2 0.1 5 31 31
were mature so that L, did not vary in the basal part of long roots ( z > 200 mm). It can be seen from Figure 3, A and B, that the two staining procedures used (berberine sulfate/ aniline blue and toluidine blue O) resulted in similar values for L, at the base of the root (z = I ) and for the distance at which the lignification of xylem was complete. However, the fluorescent staining resulted in somewhat lower values of L, near the root tip (Fig. 3, A and B; Table I). L, were calculated using Poiseuille’s law (Eq. 3) or the
equations for elliptical cross-sections (Eqs. 4 and 5). Equation 3 always resulted in somewhat higher values of L, than Equations 4 and 5. It is known that the differences are small for ellipses of low eccentricities (Preston, 1938). This is the reason why L, calculated from Equations 4 and 5 differed from those calculated from Equation 3 only by a factor of 1.30 at maximum (mean f SD: 1.050 f 0.044; n = 190 cross-sections; N = 21 roots). Differences were larger near the root tip, i.e. at low L,, where the small vessels of the protoxylem and of the early metaxylem largely contributed to the overall L,. These vessels were more eccentric than the
Figure 4. L, (A and B) and Lp, (C and D) as a function of the geometric A, or I of the intact root segments. L, represented radial conduct- ance to a good approximation because of high axial conductance, i.e. low resistance, of xylem water flow (see Fig. 5). Each data point repre- sents one root. A, For A, < 550 mm’, there was a linear relationship between L, and A,, indicat- ing a constant hydraulic conductivity (Lp, = dL,/ dA,) in this root zone (? = 0.69, N = 14). The intercept with the x axis was 90 mmz and rep- resented the apical area that was ”hydraulically isolated“ from the rest of t h e root. For A, > 550 mm2, no dependency on A, could be measured for L, (? = 0.019, significance P = 0.64, N = 14). This means that in this root zone, Lp, was small as compared with the apical 550 mmz. B, Due to the linear relationship between A, and I, the dependency of L, on I was similar to that on A,. C, The apparent hydraulic conductivity (LppPP = L,/A,) increased with increasing surface area in the apical 400 mm2. D, When A, was corrected by the area of the”’hydraulically iso- lated” tip zone (Ao), the Lp, did not depend on A, in the apical 500 mm2, With further increas- ing surface area, Lp, decreased because of the small contribution of the basal root zone to the overall uptake. Error bars indicate the SD ( n = 4-1 1 experiments per root).
large elements of late metaxylem. A11 data shown in Figure 3, A and B, and TabIe I were calculated using Equations 4 and 5.
L, measured with the root pressure probe were lower than those calculated from the radii of mature xylem vessels (Fig. 3, C and D). The values determined on short, open root segments outside the seal were lower by 1 to 2 orders of magnitude than those evaluated from the stained cross- sections at z = 200 to 300 mm. However, they were similar at distances of 50 mm < z < 100 mm and z approximately equal to 400 mm (Fig. 3C). Thus, after xylem vessels were lignified, they were not fully conducting, probably because of partly remaining cross walls (see “Discussion”). Before cutting a root for the determination of L,, a P, of more than 0.2 MPa (2 bar) was built up over at least 24 h, indicating a proper sealing of the root. In addition, this increase and subsequent stability of P, indicated that active processes (active ion pumping) were maintained even 1 d after roots were excised from the bulb and attached to the pressure probe (see also Birner and Steudle, 1993). Possible effects of
O? 500 1000 1500
*o A, [“‘I
0 500 1000 1500
[“‘I
0.4
0.2 I:i I l i 7
I
# I 0.0 1 I I , I
O 100 200 300
.+ I sl a z
d
I
E u
r- O -4
h
a 8 Y
I
0- O 500 1000 1500
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

Hydraulic Conductivity of Onion Roots 1311
the hydraulic resistance of those parts of the roots that were in the seal could be excluded because the data were corrected for this, i.e. the data obtained using the second method (Fig. 3D) were used to correct the data of Figure 3C.
The lowest values of L, were obtained when roots were cut right at the seal after being fixed to the probe for at least 24 h (Fig. 3D; before the cut P, > 0.2 MPa). This indicated a compression of some of the root xylem vessels due to sealing. It should be noted that significant contributions of the seal (leading to extremely low L, values) could be easily excluded. As in the determination of the overall hydraulic conductance, only those experiments were taken into account in which half-times of water exchange of the intact (closed) root seg- ment were by at least 1 order of magnitude larger compared with the half-times measured after the cut right at the seal. Because the data of Figure 3C were corrected for effects of compressing root xylem in the seals, they refer to the same state as in the intact root.
LPr
The overall L, increased with increasing A, and 1 of the root segment (Fig. 4, A and B). There was a linear relationship of L, as a function of A, for values of A, < 550 mm2, as one would expect (Fig. 4A, ? = 0.686, significance P < 0.001, N = 14). The slope of this regression line represented the radial hydraulic conductivity Lp, = 1.37 X 10-7 m s-' MPa-I. However, for values of A, > 550 mm2, the L,(A,) curve reached a saturation value, and there was no significant correlation between L, and A, (r' = 0.019, P = 0.64, N = 14). This means that due to root development, apical parts of the root con- tributed to a higher extent to the overall water uptake than basal. Lp, of basal root zones was reduced substantially.
10-'
O 100 200 300
["I Figure 5. Comparison between the axial conductance of a root segment from the root base to a given position z from the root apex (L , lines) and the total hydraulic conductance of a closed root segment distal to that position (Lr, symbols). The values for L, were calculated assuming a root length of 300 m m and using the fit functions of the L, (Fig. 3 and Table I ) . In the range where L , could be measured (z > 63 mm), the L, between t h e position z and the root base was larger than the hydraulic conductance of the apical root part by a factor of at least 5, if the measured L, values were taken (measured on open root segments or on the part of the roots inside the seal). I f L, was calculated o n t h e base of the L, values from the cross-sections (stained with berberine/aniline or toluidine), it was even larger (2 or more orders of magnitude difference to L,).
However, the apical 90 mm2 (Ao, corresponding to a root length of about 27 mm) were considered as "hydraulically isolated," i.e. they did not contribute much to water uptake of the root because of immature xylem. The effect is shown as a positive intercept of the regression line with the A, axis (Ao, Fig. 4A). Ao was subtracted from the geometric surface area of the root to give the "effective surface area." The effective surface area of the roots was then used to recalculate Lp, (Fig. 4D). The correction had a large effect for short roots as compared with the "apparent hydraulic conductivity," which refers to the total (geometric) surface area (Fig. 4C). Because of the linear relationship between A, and I (A, = 4.21 mm X I - 66 mm2, ? = 0.966; N = 28; for I > 63 mm), the critica1 value of A, of 550 mm2 up to which L, was linearly related to A, corresponded to z = 146 mm. The regression line of A,(I) did not pass through the origin because of the decrease of root diameter toward the root tip.
L, versus Radial Hydraulic Conductance
The contribution of the L, ata given position from the apex (z) was evaluated by comparing the total hydraulic conduct- ance of the part of the root distal from that position (LJ with the axial conductance between that position and the base of the root, L, (Fig. 5). That value was evaluated by integrating the specific axial hydraulic resistance (R, = l/L,) over the distance between the given position and the base of the root ( 1 = 300 mm was taken) and calculating the inverse of this integrated value. For z > 60 mm, the L, calculated from the cross-sections following Equations 4 and 5 (or Poiseuille's law) was larger than the total conductance of the apical part by at least 2 orders of magnitude. If the L, values measured with the root pressure probe were taken (Fig. 3, C and D), L, was larger than L, by a factor of at least 5 for a total root length of 300 mm. However, no measurements of L, were made for short roots ( I < 63 mm), and determination of L, using the pressure probe was restricted to z > 51 mm. Therefore, no comparison between measured L, and L, could be made for the root zone close to the apex.
DlSCUSSlON
The results show that in onion roots, both the L, and Lp, change strongly from the tip toward the base. L, increased over more than 3 orders of magnitude with increasing z, whereas the Lp, was lower by a factor of at least 5 in basal relative to apical root zones, although the small contribution to radial water uptake in the basal parts was quite variable and, therefore, difficult to quantify. Using the root pressure probe technique, L,(z) and Lp,(z) could be determined at a rather high resolution that, in principle, could be increased further. This is hardly possible with other available techniques.
L X
The results of the staining procedures involve some uncer- tainty because the decision whether a vessel was conducting or not was difficult. The technique could be improved by making longitudinal sections in addition to cross-sections to
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

1312 Melchior and Steudle Plant Physiol. Vol. 101, 1993
examine at which position in the root and to what extent cross walls in the vessels are dissolved. Another possibility to improve the method would be to suck dye across root segments that would stain only conducting vessels, the di- mensions of which are then used in the calculations (Peterson and Steudle, 1993).
At I = 200 to 300 mm, L, values calculated according to Poiseuille's law were larger than those measured with the root pressure probe by more than 1 order of magnitude. This difference is in accordance with Zimmermann and Brown (1971); Greacen et al. (1976); Giordano et al. (1978); and Frensch and Steudle (1989). The results support the view that the application of Poiseuille's law to xylem transport should be done with caution. Deviations from the theory seem to be due mainly to remaining cross walls. On the other hand, the elliptical shape of vessel cross-sections and the roughness of walls play a minor role if any (Nonweiler, 1975).
In pressure probe experiments, a proper sealing was indi- cated by root pressures of more than 0.2 MPa that were built up over time periods of at least 24 h. Preliminary experiments showed that extremely high apparent Lp, (Lp:PP = LJA,) of up to 5 x 10-6 m s-' MPa-' indicated a bypass of water at the seal. They were measured only at P,, below 0.15 MPa. For P, > 0.2 MPa, there was no dependency of Lp$PP on P,. The method of Frensch and Steudle (1989), which involved a cutting of root segments attached to the pressure probe right at the seal and measuring the remaining parts of the roots inside the seal, resulted, in part, in extremely low values of L, (not shown in Fig. 3D). Even when only those experi- ments were chosen where the half-times of water flow were lower by a factor of at least 10 after the cut at the seal than in intact root segments, low L, values were calculated for z > 300 mm. These cases indicated a substantial compression of the xylem by the seal, although this did not interfere with the determination of the overall hydraulic conductance of the root. Without considering these rare cases, a good corre- spondence was found between this method and the deter- mination of L, on short, open root segments outside the seal, where the values were corrected for the effect of the seal (compare Fig. 3, C and D). Thus, in the pressure probe experiments presented, "sealing effects" were not important. This is readily shown by comparing the two methods to determine L,.
Clarkson et al. (1984) perfused onion root segments of a length of 65 mm. In agreement with the data found in this study, these authors measured an increase of L, with increas-
I ing z even for z > 250 mm. They explained the increase by the existence of cross walls between members of lignified xylem vessels. Similarly, in barley roots, the maximal axial conductivity was reached at distances of 150 to 250 mm from the root tip. This was correlated with the complete break- down of cross walls between vessel members (Sanderson et al., 1982).
In the present study, remains of cross walls between vessels were found in basal root parts and should be studied system- atically in the future using seria1 cross- and longitudinal sections. A narrowing by remains of cross walls may result in turbulent flow of water and in a reduction of L,. This may also occur as a consequence of circular stiffenings of vessel walls that cannot be seen on cross-sections. Giordano et al.
'
(1978) proved that measured axial conductivities can differ greatly from calculated ones according to Poiseuille's law if the vessels were periodically narrower, leading to turbu- lences. Thus, small Reynolds numbers by themselves do not indicate that water flow would be laminar. The absolute values of L, calculated from the data given by Clarkson et al. (1984) range from 2.3 X 10-" m4 s-' MPa-' ( z = 70-135 mm) to 7.2 X 10-l' m4 s-' MPa-' (z = 300-365 mm) and correspond with the values measured in this study (10-l2 to 10-9 m4 s-' MPa-', Fig. 3, C and D). However, in the present study, the changes during root development were recorded at a resolution that was higher by a factor of 3 to 4.
lnfluence of L, on Determination of Lp,
Over the whole range of length of the root segments measured in this study (60-320 mm), L, to a hypothetical root base at z = 300 mm was larger by a factor of at least 5 than the total L, of the segment. The linear relationship between L, and the A, indicated a low influence of L, on L, (for z > 63 mm). This result shows that for onion roots as well as for roots of other species, a two-compartment model (which neglects the longitudinal component of LI ) may be used to evaluate Lp, to a good approximation (Steudle and Jeschke, 1983; Steudle et al., 1987). However, it has to be taken into account that in the tip region, immature xylem would cause a high axial resistance that would hydraulically isolate this apical zone from the basal root parts. The effect is shown as a positive intercept with the A, axis (AO, Fig. 4A). Due to the large variation between roots, the intercept had a large error because the slope of the regression line and intercept with the L, axis had errors of 20 and 80%, respec- tively. The surface area of the hydraulically isolated tip zone could be higher than Ao = 90 mm2 (corresponding to z = 27 mm) because the inverse relationship (A, plotted as a function of L,) had an intercept with the A, axis of 171 f 39 mm2 (corresponding to z = 51 f 12 mm). In the experiments of Clarkson et al. (1984), the apical 60 mm had high axial hydraulic resistances that contributed substantially to the overall resistance.
LPr
Using segments of primary roots of maize of a length of 18 to 140 mm, which were shorter than those used in this paper, Frensch and Steudle (1989) reported an increase of the "apparent Lp," (measured with the root pressure probe) with increasing root length. However, when it was taken into account that the apical 15 mm of the segments were "hy- draulically isolated" because of immature metaxylem, the Lp, Treferred to the effective surface area) was constant over the entire root length. The authors state that a decrease of Lp, in more basal root zones (z > 140 mm, as measured in this study) could, however, not be excluded. In maize roots, Casparian bands and suberin lamellae in the exodermis were detected at distances from the root tip of z = 120 to 130 mm, whereas in onion roots they develop at z = 50 to 60 mm (Perumalla and Peterson, 1986). In addition, the low contri- bution of basal zones of onion roots could be explained by a minor contribution of apbplasmic bypasses in those areas
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

Hydraulic Conductivity of Onion Roots 1313
where secondary root initials penetrate the endodermis. In part, Frensch and Steudle (1989) interpreted the high contri- bution of the apoplasmic pathway to radial water transport by a flow across the latter structures.
An increased water uptake at the site of secondary root formation was also indicated by experiments with apoplasmic tracers (Peterson et al., 1981) and was discussed for Norway spruce (Haussling et al., 1988). If the exodermis would effi- ciently block off the apoplasmic water movement in onion roots, as indicated by the work of Peterson and coworkers (see the refs. given by Peterson, 1989), the penetration of root primordia across the cortex (consisting of 10-15 cell layers) would only slightly affect the water taken up through the cell walls unless the laterals wound the exodermis. Al- though it was found in this study that laterals develop much earlier than they emerge from the rhizodermis (similar to maize), the position where the exodermis was penetrated and wounded was not examined. However, the differences in the formation of laterals as well as the differences in the forma- tion of Casparian bands and suberin lamellae in endodermis and exodermis in both species seem to be important for the difference of the Lp,(z) characteristics observed for the two species.
North and Nobel (1991) measured the apparent Lp, of Agave deserti by sucking solution across root segments about 80 mm in length. They found that the apparent Lp, of apical parts of nodal roots ("young roots") was larger by a factor of 5 compared with that of basal parts ("older roots"). Using the model of Landsberg and Fowkes (1978), North and Nobel evaluated Lp , to be 1.8 X 10-6 m s-' MPa-' and 3.8 X 10-8 m s-' MPa-' for apical and basal parts of the roots, respec- tively, i.e. there was a decrease in Lp, by a factor of about 50 during root development. The result was mainly explained by the suberization of endodermal and exodermal cell walls.
In a study on roots of water-stressed Sorghum bicolor, Cruz et al. (1992) applied the same technique to determine L, and L,. However, these authors did not correct for the apical zone with low axial conductance. Rather, they gave an ap- parent Lp, of 2.2 X 10-7 m s-' MPa-' and 0.6 X 10-7 m s-' MPa-' for controls and water-stressed plants, respectively. L, declined by 2 orders of magnitude due to the drought treat- ment (1.8 X 10-l' m4 s-' MPa-' and 1.7 X 10-l2 m4 s-' MPa-' in control and stressed plants, respectively). Compared with hydroponically grown onion roots, the low L, of water- stressed Sorghum roots was similar to L, close to the root tip, where only the small protoxylem vessels were mature. The large decrease of L, of Sorghum roots during drought was correlated with smaller vessel diameters and a lower propor- tion of lignified xylem elements (Cruz et al., 1992). In stressed roots, exodermis and endodermis cell walls were lignified and suberized earlier. It should be noted that, different from the root pressure probe technique, in the experiments of North and Nobel(l991) and Cruz et al. (1992), there was no criterion that roots were well sealed and not wounded before measuring the conductances. A bypass of water at the seal could possibly be detected by an overestimation of axial and radial conductivities. However, the results of North and Nobel(l991) and Cruz et al. (1992) show that even with the more simplifying techniques, there were indications of large changes of L, and Lp, caused by environmental factors such
as drought stress. With these techniques, however, hydraulic parameters could not be resolved in finer detail.
Rosene (1937) carefully examined the contribution of dif- ferent root zones to the overall water absorption of intact onion roots (still attached to the bulbs). She used micropo- tometers and found that a11 parts of the root from the tip to the base (the root cap itself was not measured) absorbed water but at different rates. In longer (older) roots ( I > 80 mm), the water uptake rate was maximal in the tip zone between z = O and z = 40 mm and declined strongly at z = 60 to 70 mm. Water absorption ranged from 3 X 10-9 to 8 X
10-* m s-', with a large variation between individual plants. There was good evidence that intact and excised roots of corresponding age and length showed similar behavior with respect to absolute rates and axial gradients of water uptake (Rosene, 1941a). Rosene (1941b) determined the driving force for water uptake by applying osmotic solutions to different root zones at concentrations just sufficient to stop the water flow. Taking into account a reflection coefficient of about 0.65 for the solute used (KNO,; our unpublished result), the driving force in the absorption experiment could be calculated to be about 0.4 MPa. Thus, Lp, would have ranged from 7 X 10-9 to 2 X 10-7 m s-' MPa-' for intact roots measured in an atmosphere saturated with water.
These values are smaller than the Lp, measured in the present study (pressure relaxations driven by gradients of hydrostatic pressure) and larger than Lp , measured in osmot- ically driven relaxations (our unpublished results). However, it is doubtful whether RH really was 100% as stated by Rosene (1941b), so that even at near water saturation, tran- spiration (pressure)-driven water uptake could have substan- tially contributed to total water absorption. Therefore, the driving force in the experiments of Rosene (1937) may have been a mixed hydrostatic and osmotic gradient and the absolute value of Lp, would be, thus, understandable.
Recently, Zimmermann et al. (1992) criticized the root pressure probe technique as being inadequate for the deter- mination of water and solute relations parameters for severa1 reasons. (a) The assumption of a high rigidity of the xylem would not hold because (from their own measurements) the xylem would have an elastic modulus of a few tenths of 1 MPa. This would mean that this structure would double its volume when subjected to pressure increases of a few bar. (b) Zimmermann et al. (1992) are concerned about effects of unstirred layers during the measurement of water flow. How- ever, they overlooked that a rigorous analysis has proven that results of pressure probe experiments (eg. low reflection coefficients of roots) could not be completely explained by unstirred layers (Steudle and Frensch, 1989). (c) Zimmer- mann et al. (1992) are also concerned by the application of the theory of irreversible thermodynamics to complex com- posite structures such as roots. However, this is possible as shown by basic theoretical work ( e g Kedem and Katchalsky, 1963a, 1963b) as well as by papers where it has been applied to roots (e.g. Fiscus, 1975; Pitman et al., 1981; Steudle and Jeschke, 1983; Dainty, 1985; Miller, 1985; Steudle et al., 1987; Schambil and Woermann, 1989). Application of irreversible thermodynamics to roots has led to the "composite transport model of the root" (Steudle, 1992).
The problems of Zimmermann et al. (1992) arise from their
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

1314 Melchior and Steudle Plant Physiol. Vol. 101, 1993
own cell pressure probe measurements on root cortex cells of Aster tripolium (measurements at the root level could not be performed). The results indicated that in young Aster roots there was a steep inwardly directed gradient of turgor and osmotic pressure. Astonishingly, this resulted in a constant water potential of close to zero across the entire cortex, even when the roots were subjected to 300 m NaCl (osmotic pressure approximates 1.4 MPa). Zimmermann et al. (1992) claim that this was due to complete exclusion of NaCl from the apoplast of the young roots. Even more surprising, a zero water potential in the cortex was found despite a high hy- draulic conductivity of cortical cells that equilibrated with half-times of about 14 s with their surroundings. When roots were excised from the plants, gradients of turgor vanished within 15 to 30 min. The authors claim that this indicates that the use of excised roots for measuring water relations should be treated with caution. However, they do not resolve the inconsistencies in their own data that refer (a) to the exclusion of salts from the apoplast of young roots; (b) to the lack of any tendency of an equilibration of water potential of the roots with the medium of high salinity; and (c) to the fact that if excision would alter turgor at a given water potential; the ionic composition of cortical cells would also change. This means that the ionic concentration of root cells should rapidly change upon excision. This has not yet been observed. Cell pressure probe work indicated that there were no gradients of cell turgor in the cortex of intact roots of wheat and maize (Pritchard et al., 1989; Spollen and Sharp, 1991). Thus, the results of Zimmermann et al. (1992) would need some veri- fication. Contradictions need to be resolved.
In conclusion, the data presented in this paper show that L, and L p , may both vary substantially along a developing root. The deposition of Casparian bands in the exodermis and the formation of suberin lamellae in the endodermis and exodermis appear to be very important. For a detailed mod- eling of the water uptake of roots, i.e. of their efficiency, the pattern of changes of hydraulic conductivities along the roots is crucial. Root pressure probes can be used to evaluate the governing parameters at the resolution necessary to work out the hydraulic architecture of root systems. The technique would also allow measurement of. interactions between water and solute (nutrient) flows. Using the cell pressure probe, the analysis could be extended further from the tissue or organ level to that of individual cells (Steudle et al., 1987; Zhu and Steudle, 1991). It has been demonstrated that this is impor- tant since the hydraulic conductivity of cell membranes may change in response to environmental changes (Azaizeh et al. 1992; Bimer and Steudle, 1993).
ACKNOWLEDCMENTS
The authors are indebted to Dr. C.A. Peterson and Ms. D.E. Enstone for the introduction to the fluorescent staining technique. We wish to thank A. Elend and M. Scherer, who were involved in part of the anatomical work, and M. Lauerer and Dr. M. Paul for carefully reading the manuscript. The expert technical assistance of B. Stumpf is gratefully acknowledged.
Received September 8, 1992; accepted December 14, 1992. Copyright Clearance Center: 0032-0889/93/101/1305/11.
LITERATURE ClTED
Azaizeh H, Gunse B, Steudle E (1992) Effects of NaCl and CaCI2 on water transport across root cells of maize (Zea mays L.) seedlings. Plant Physiol99 886-894
Azaizeh H, Steudle E (1991) Effects of salinity on water transport of excised maize (Zea mays L.) roots. Plant Physiol97: 1136-1145
Birner T, Steudle E (1993) Effects of anaerobic conditions on water and solute relations and active transport in root of maize (Zea mays L). Planta (in press)
Boyer JS (1985) Water transport. Annu Rev Plant Physiol 3 6
Brouwer R (1954) The regulating influence of transpiration and suction tension on the water and salt uptake by the roots of intact Vicia faba plants. Acta Bot Neerl 3 264-312
Brundrett MC, Enstone DE, Peterson CA (1988) A berberine-aniline blue fluorescent staining procedure for suberin, lignin, and callose in plant tissue. Protoplasma 146 133-142
Clarkson DT, Williams L, Hanson JB (1984) Perfusion of onion root xylem vessels: a method and some evidence of control of the pH of the xylem sap. Planta 162 361-369
Cruz RT, Jordan WR, Drew MC (1992) Structural changes and associated reduction of hydraulic conductance in roots of Sorghum bicolor L. following exposure to water deficit. Plant Physiol 9 9
Dainty J (1985) Water transport through the root. Acta Hortic 171:
Fiscus EL (1975) The interaction between osmotic- and pressure- induced water flow in plant roots. Plant Physiol55 917-922
Frensch J, Steudle E (1989) Axial and radial hydraulic resistance to roots of maize (Zea mays L.). Plant Physiol91: 719-726
Giordano R, Salleo A, Salleo S, Wanderlingh F (1978) Flow in xylem vessels and Poiseuille’s law. Can J Bot 5 6 333-338
Greacen EL, Ponsana P, Barley KP (1976) Resistance to water flow in the roots of cereals. In OL Lange, L Kappen, E-D Schulze, eds, Ecological Studies, Vol 19: Water and Plant Life. Springer Verlag, New York, pp 86-100
Haussling M, Jorns CA, Lehmbecker G, Hecht-Buchholz C, Mar- schner H (1988) Ion and water uptake in relation to root devel- opment in Norway spruce (Picea abies (L.) Karst.). J Plant Physiol
Kedem O, Katchalsky A (1963a) Permeability of composite mem- branes. Part 2: Parallel elements. Transactions of the Faraday Society 5 9 1931-1940
Kedem O, Katchalsky A (1963b) Permeability of composite mem- branes. Part 3: Series array of elements. Transactions of the Fara- day Society 59 1941-1953
Landsberg JJ, Fowkes ND (1978) Water movement through plant roots. Ann Bot 42: 493-508
Miller DM (1980) Studies of root function in Zea mays. I. Apparatus and methods. Can J Bot 5 8 351-360
Miller DM (1985) Studies of root function in Zea mays. 111. Xylem sap composition at maximum root pressure provides evidence of active transport into the xylem and a measurement of the reflection coefficient of the root. Plant Physiol77: 162-167
Nonweiler TRF (1975) Flow of biological fluids through non-ideal capillaries. In MH Zimmermann, JA Milbum, eds, Encyclopedia of Plant Physiology, New Series, Vol 1: Transport in Plants, I: Phloem Transport, Appendix I. Springer Verlag, New York, pp 474-477
North GB, Nobel PS (1991) Changes in hydraulic conductivity and anatomy caused by drylng and rewetting roots of Agave deserti (Agavaceae). Am J Bot 78: 906-915
Passioura JB (1988) Water transport in and to roots. Annu Rev Plant Physiol Plant Mo1 Biol39 245-265
Perumalla CJ, Peterson CA (1986) Deposition of Casparian bands and suberin lamellae in the exodermis and endodermis of young com and onion roots. Can J Bot 6 4 1873-1878
Peterson CA (1989) Significance of the exodermis in root function. In BC Loughman, O Gasparikova, J Kolek, eds, Structural and Functional Aspects of Transport in Roots. Kluwer Academic Pub- lishers, Dordrecht, The Netherlands, pp 35-40
Peterson CA, Emanuel ME, Humphreys GB (1981) Pathway of movement of apoplastic fluorescent dye tracers through the en-
473-516
203-212
21-31
133: 486-491
-
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022

Hydraulic Conductivity of Onion Roots 1315
dodennis at the site of secondary root formation in com (Zea mays) and broad bean (Vicia faba). Can J Bot 5 9 618-625
Peterson CA, Murrmann M, Steudle E (1993) Location of the major bamers to water and ion movement in young roots of Zea mays L. Planta (in press)
Peterson CA, Steudle E (1993) Lateral hydraulic conductivity of early metaxylem vessels in Zea mays L. roots. Planta 189 288-297
Pitman MG, Wellfare D, Carter C (1981) Reduction of hydraulic conductivity during inhibition of exudation from excised maize and barley roots. Plant Physiol 67: 802-808
Preston RD (1938) The contents of the vessels of Fraxinus americana L., with respect to the ascent of sap. Ann Bot 2: 1-21
Pritchard J, Williams G, Wyn Jones RG, Tomos AD (1989) Radial turgor pressure profiles in growing and mature zones of wheat roots-a modification of the pressure probe. J Exp Bot 4 0
Robards AW, Jackson SM, Clarkson DT, Sanderson J (1973) The structure of barley roots in relation to the transport of ions into the stele. Protoplasma 77: 291-311
Rosene HF (1937) Distribution of the velocities of absorption of water in the onion root. Plant Physiol12: 1-19
Rosene HF (1941a) Comparison of rates of water intake in contiguous regions of intact and isolated roots. Plant Physioll6 19-38
Rosene HF (1941b) Control of water transport in local root regions of attached and isolated roots by means of the osmotic pressure of the extemal solution. Am J Bot 28: 402-410
Sanderson J (1983) Water uptake by different regions of the barley root. Pathways of radial flow in relation to development of the endodermis. J Exp Bot 3 4 240-253
Sanderson J, Whitbread FC, Fielding WJ (1982) Axial conductivity in the xylem vessels of the seminal root of barley. In Agricultura1 Research Council Letcombe Lab Annual Report 1981, pp 70-72
Schambil F, Woermann D (1989) Radial transport of water across cortical sleeves of excised roots of Zea mays L. Planta 178:
Spollen WG, Sharp RE (1991) Spatial distribution of turgor and root
567-571
488-494
growth at low water potentials. Plant Physiol96 438-443
St Aubin G, Canny MJ, McCully ME (1986) Living vessel elements in the late metaxylem of sheathed maize roots. Ann Bot 5 8
Steudle E (1992) The biophysics of plant water: compartmentation, coupling with metabolic processes, and flow of water in plant roots. In GN Somero, CB Osmond, CL Bolis, eds, Water and Life: Comparative Analysis of Water Relationships at the Organ- ismic, Cellular, and Molecular Levels. Springer Verlag, Berlin,
Steudle E, Frensch J (1989) Osmotic responses of maize roots. Water and solute relations. Planta 177: 281-295
Steudle E, Jeschke WD (1983) Water transport in barley roots. Measurements of root pressure and hydraulic conductivity of roots in parallel with turgor and hydraulic conductivity of root cells. Planta 158: 237-248
Steudle E, Oren R, Schulze E-D (1987) Water transport in maize roots. Measurement of hydraulic conductivity, solute permeability, and of reflection coefficients of excised roots using the root pres- sure probe. Plant Physiol84 1220-1232
Weatherley PE (1982) Water uptake and flow in roots. In OL Lange, I'S Nobel, CB Osmond, H Ziegler, eds, Encyclopedia of Plant Physiology, New Series, Vol 128: Physiological Plant Ecology, 11: Water Relations and Carbon Assimilation. Springer Verlag,. New York, pp 79-109
Zhu GL, Steudle E (1991) Water transport across maize roots. Simultaneous measurement of flows at the cell and root leve1 by double pressure probe technique. Plant Physiol95 305-315
Zimmermann MH, Brown CL (1971) Transport in the xylem. In Trees. Structure and Function. Springer Verlag, New York, pp
Zimmermann U, Rygol J, Balling A, Klock G, Metzler A, Haase A (1992) Radial turgor and osmotic pressure profiles in intact and excised roots of Aster tripolium. Pressure probe measurements and nuclear magnetic resonance-imaging analysis. Plant Physiol 99:
577-588
pp 173-204
169-220
186-196
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/4/1305/6066059 by guest on 12 February 2022