xiaohua wang, yang zhou, ruoyun tan, mingxia xiong...
TRANSCRIPT

doi:10.1152/ajprenal.00216.2010 299:973-982, 2010. First published Sep 15, 2010;Am J Physiol Renal Physiol
Ping Wen, Lei Jiang and Junwei Yang Xiaohua Wang, Yang Zhou, Ruoyun Tan, Mingxia Xiong, Weichun He, Li Fang,
You might find this additional information useful...
37 articles, 19 of which you can access free at: This article cites http://ajprenal.physiology.org/cgi/content/full/299/5/F973#BIBL
including high-resolution figures, can be found at: Updated information and services http://ajprenal.physiology.org/cgi/content/full/299/5/F973
can be found at: AJP - Renal Physiologyabout Additional material and information http://www.the-aps.org/publications/ajprenal
This information is current as of November 7, 2010 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0363-6127, ESSN: 1522-1466. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2010 by therespective cells and vasculature, as well as to the control of body fluid volume and composition. It is published 12 times a year
publishes original manuscripts on a broad range of subjects relating to the kidney, urinary tract, and theirAJP - Renal Physiology
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

Mice lacking the matrix metalloproteinase-9 gene reduce renal interstitialfibrosis in obstructive nephropathy
Xiaohua Wang, Yang Zhou, Ruoyun Tan, Mingxia Xiong, Weichun He, Li Fang, Ping Wen, Lei Jiang,and Junwei YangCenter of Kidney Disease, 2nd Affiliated Hospital, Nanjing Medical University, Nanjing, Jiangsu Province, China
Submitted 21 April 2010; accepted in final form 31 August 2010
Wang X, Zhou Y, Tan R, Xiong M, He W, Fang L, Wen P,Jiang L, Yang J. Mice lacking the matrix metalloproteinase-9 genereduce renal interstitial fibrosis in obstructive nephropathy. Am JPhysiol Renal Physiol 299: F973–F982, 2010. First published Sep-tember 15, 2010; doi:10.1152/ajprenal.00216.2010.—Matrix metallo-proteinase-9 (MMP-9) is one of the major components of the matrixproteolytic network, and its role in the pathogenesis of renal intersti-tial fibrosis remains largely unknown. Here, we demonstrate thatablation of MMP-9 attenuated renal interstitial fibrotic lesions inobstructive nephropathy. Mice lacking MMP-9 were less likely todevelop morphological injury, which was characterized by a reduceddisruption of tubular basement membrane (TBM) and expression offibronectin as well as deposition of total tissue collagen in the kidneysafter sustained ureteral obstruction compared with their wild-typecounterparts. Deficiency of MMP-9 blocked tubular epithelial-to-myofibroblast transition (EMT) but did not alter the induction oftransforming growth factor (TGF)-�1 axis expression in the ob-structed kidneys. In vitro, TBM, which was digested by MMP-9instead of MMP-9 itself, induces EMT and enhances migration oftransformed cells. Thus increased MMP-9 is detrimental in renalinterstitial fibrogenesis through a cascade of events that leads to TBMdestruction and in turn to promotion of EMT. Our findings establisha crucial and definite importance of MMP-9 in the pathogenesis ofrenal interstitial fibrosis at the whole-animal level.
tubular basement membrane; epithelial-to-myofibroblast transition;collagen
REGARDLESS OF THE DIVERSE disease process, renal interstitialfibrosis is considered to be the common pathological pathwayand irreversible process that eventually leads to end-stage renaldisease. Both human disease and animal models have been ableto show that massive interstitial myofibroblast activation is thepredominant source of matrix proteins, such as collagen typesI, III, fibronectin, etc., which results in renal fibrosis. Numer-ous evidence suggested that the myofibroblasts may derivefrom tubular epithelial cells undergoing epithelial-to-myofi-broblast transition (EMT). EMT is an orchestrated, highlyregulated process consisting of four key steps, in which themechanism of destruction of the tubular basement membrane(TBM) resulting from upregulation of matrix proteinases ispoorly understood. Matrix metalloproteinases (MMPs) are afamily of more than 28 members that were initially identifiedas enzymes to cleave elements of the extracellular matrix(ECM) (20), which subsequently have been found to be up-regulated in tumors (18). As digestion of the ECM is essentialfor cell migration and invasion, MMPs have been studied for
their role in tumor metastasis. Alternation in regulation ofMMP activity is implicated in diseases such as cancer, fibrosis,arthritis, and atherosclerosis (2, 6, 11, 27). Among the MMPfamily, there is emerging evidence indicated that MMP-9 canstimulate processes associated with EMT, a developmentalprocess that is activated in renal fibrosis (15, 31). Gelatinzymographic analysis of both whole-kidney lysate of ureteral-obstructed mice and conditioned media of transforming growthfactor (TGF)-�1-treated tubular epithelial cells displayed no-ticeable induction of MMP-9, while a marked decrease inMMP-9 induction resulting from tissue-type plasminogen ac-tivator (tPA) knockout led to a dramatic preservation of thestructural and functional integrity of the TBM and preventionof EMT (32). Given the significance of MMP-9 for matrixhomeostasis in multicellular organisms, it is not difficult tocomprehend that its expression and activity are tightly regu-lated through a multitude of controlling mechanisms. Dysregu-lation of MMP-9 may contribute to the pathogenesis of a widevariety of diseases ranging from tumor metastasis to tissuefibrogenesis.
While the importance of the MMP-9 in matrix proteolyticsystems is well documented, its role in the pathogenesis ofrenal interstitial fibrosis, characterized by excess matrix accu-mulation and deposition, is poorly defined. Tubular interstitialfibrosis is often regarded as a final, common endpoint/outcomeof many forms of chronic renal disease (5, 14, 24, 30, 33, 36).One of the key events during interstitial fibrogenesis is the denovo activation of �-smooth muscle actin (�-SMA)-positivemyofibroblast cells (31). Because these cells are primarilyresponsible for interstitial matrix accumulation and depositionin chronically diseased states, and because they are essentiallyabsent in the interstitial compartment of normal kidney, eluci-dation of their origin(s) and the activation process is of funda-mental importance both for understanding the pathologicmechanism of renal interstitial fibrosis and for identifyingtherapeutic targets. Myofibroblasts are often presumed to orig-inate from residential fibroblasts (21, 25); however, numerouspieces of evidence suggests that they may also derive fromtubular epithelial cells via an epithelial-to-myofibroblast tran-sition (EMT) (14). It is well accepted that renal EMT is anorchestrated, highly regulated, stepwise process in which de-struction of the integrity of the tubular basement membrane(TBM) may be crucial. However, whether the molecules suchas MMP-9, an upstream regulator of collagen and laminin,which are the major components of TBM (4, 29), are involvedin the destruction of the TBM under pathological conditions atthe whole-animal level remains unknown.
The availability of genetically engineered mice in which theendogenous MMP-9 gene is inactivated provides an invalu-able, unprecedented model system for studying the function of
Address for reprint requests and other correspondence: J. Yang, Center ofKidney Disease, 2nd Affiliated Hospital, Nanjing Medical Univ., 262 Zhong-shan East Rd., Nanjing 210003, Jiangsu Province, China (e-mail: [email protected]).
Am J Physiol Renal Physiol 299: F973–F982, 2010.First published September 15, 2010; doi:10.1152/ajprenal.00216.2010.
0363-6127/10 Copyright © 2010 the American Physiological Societyhttp://www.ajprenal.org F973
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

MMP-9 in the pathogenesis of renal interstitial fibrosis in vivo.In this study, we investigated the progression of renal intersti-tial fibrosis induced by unilateral ureteral obstruction (UUO) inMMP-9�/� and MMP-9�/� mice. We found that the nullmutation of the MMP-9 gene protected the kidney from devel-oping interstitial fibrotic lesions. The detrimental effect ofendogenous MMP-9 was apparently not mediated by alter-ations in the expression and activation of profibrogenicTGF-�1 in the kidney. Instead, it was likely mediated bydestruction of the TBM, which in turn resulted in myofibro-blastic transition of tubular epithelial cells via EMT. Ourresults validate the hypothesis that TBM integrity plays anessential role in preventing tubular EMT, thereby protectingnormal renal morphology against development of fibrotic le-sions.
MATERIALS AND METHODS
Animal models. Colonies of homozygous MMP-9 knockout (MMP-9�/�) and wild-type (MMP-9�/�) mice were raised from originalbreeding pairs obtained from the Jackson Laboratory. They werehoused in the animal facilities at the National Model Animal Center(NMAC; Nanjing, Jiangsu Province, China) with free access to foodand water. Animals were treated humanely in accordance with NMACguidelines and approved procedures of the Institutional Animal Useand Care Committee at the Nanjing Medical University. Genotypewas confirmed using PCR amplification of genomic DNA from thetail. The experiments were performed using male MMP-9�/� andMMP-9�/� mice with identical genetic profiles and with an averageage of 10 wk. There was no statistical difference in the body weightof MMP-9�/� and MMP-9�/� mice at the time of the experiments.Both knockout (MMP-9�/�) and wild-type (MMP-9�/�) animalswere randomly assigned to four groups with five mice per group:sham for 3 days, UUO for 3 days, sham for 7 days, and UUO for 7days. UUO was performed using an established procedure (31).Briefly, under general anesthesia, complete ureteral obstruction wasperformed by double-ligating the left ureter using 4-0 suture after amidline abdominal incision. Sham-operated mice had their uretersexposed and manipulated but not ligated. Mice were killed at differenttime points as indicated after surgery, and the obstructed kidneys wereremoved. One part of the kidney was fixed in 10% phosphate-bufferedformalin followed by paraffin embedding for histological and immu-nohistochemical studies. The remaining kidneys were snap-frozen inliquid nitrogen and stored at �80°C for protein and total RNAextractions.
Cell culture and treatment. Rat proximal tubular epithelial cells(NRK-52E) were obtained from American Type Culture Collection(Manassas, VA). Cells were cultured in Dulbecco’s modified Eagle’smedium-F12 medium supplemented with 5% fetal bovine serum(Sigma, Sigma-Aldrich, St. Louis, MO). For MMP-9 treatment, NRK-52E cells were seeded at �80% confluence in complete mediumcontaining 5% fetal bovine serum. Twenty-four hours later, the cellswere changed to serum-free medium, and recombinant MMP-9 (R&DSystems, Minneapolis, MN) was added at a final concentration of 10ng/ml except where otherwise indicated. The cells and conditionedmedia were collected at different time points for further characteriza-tion. GM6001 was purchased from Sigma (Sigma-Aldrich).
RNA isolation and RT-PCR. Total RNA was extracted from thekidney tissue using an Ultraspec RNA isolation system according tothe instructions specified by the manufacturer (Biotecx Laboratories,Houston, TX). RT-PCR was performed as described elsewhere (26).Briefly, the first-strand of cDNA synthesis was carried out by using aReverse Transcription System kit according to the instructions of themanufacturer (Promega, Madison, WI). PCR amplification was per-formed with a HotStar TaqMaster Mix Kit (Qiagen, Valencia, CA).The primer sequences were as follows: MMP-9 (forward) 5=-CCCA-
CATTTGACGTCCAGAGAAGAA and (reverse) 5=-GTTTTTGAT-GCTATTGCTGAGATCC, and �-actin (forward) 5=-CTCTTCCAGC-CTTCCTTCCTG and (reverse) 5=-GAAGCATT TGCGGTGGACGAT.The PCR products were size fractionated on a 1.0% agarose gel anddetected by ethidium bromide staining. No detectable signal was found ina parallel control tube without reverse transcriptase (data not shown).Expression levels of mRNA were calculated after normalizing withhousekeeping gene �-actin.
Western blot analysis. Cells were lysed with SDS sample buffer(62.5 mmol/l Tris·HCl, pH 6.8, 2% SDS, 10% glycerol, 50 mmol/ldithiothreitol, and 0.1% bromophenol blue). Kidney tissue was ho-mogenized by a polytron homogenizer (Brinkmann Instruments,Westbury, NY), and the supernatant was collected after centrifugationat 13,000 g at 4°C for 20 min, as described previously (31). Afterprotein concentration was determined using a bicinchoninic acid(BCA) protein assay kit (Sigma), the tissue lysate was mixed with anequal amount of 2� SDS sample buffer. Samples were heated at100°C for 5–10 min before loading and separated on precasted 10%SDS-polyacrylamide gels (Bio-Rad, Hercules, CA). The proteins wereelectrotransferred to a nitrocellulose membrane (Amersham, Arling-ton Heights, IL) in transfer buffer containing 48 mmol/l Tris·HCl, 39mmol/l glycine, 0.037% SDS, and 20% methanol at 4°C for 1 h.Nonspecific binding to the membrane was blocked for 1 h at roomtemperature with 5% Carnation nonfat milk in TBS buffer (20 mmol/lTris·HCl, 150 mmol/l NaCl, and 0.1% Tween 20). The membraneswere incubated for 16 h at 4°C with various primary antibodies inTBS buffer containing 5% milk at the dilutions specified by themanufacturers. The mouse monoclonal anti-�-SMA antibody (clone1A4) was purchased from Sigma. The anti-TGF-�1 type I receptor(sc-398) and anti-actin (sc-1616) antibodies were purchased fromSanta Cruz Biotechnology (Santa Cruz, CA). Binding of primaryantibodies was followed by incubation for 1 h at room temperaturewith the secondary horseradish peroxidase-conjugated IgG in 1%nonfat milk. The signals were visualized with the enhanced chemilu-minescence system (ECL, Amersham), as described previously.
Zymographic analysis. Gelatin zymographic analysis of MMPproteolytic activity in kidney tissue homogenates was performedaccording to the method described previously (31). Kidney homoge-nates were prepared essentially according to the methods described byKim et al. (12). Protein concentration was determined using a BCAprotein assay kit with BSA as a standard (Sigma-Aldrich). A constantamount of protein from the kidney tissue homogenates (30 �g) wasloaded onto a 10% SDS-polyacrylamide gel containing 1 mg/mlgelatin (Bio-Rad Laboratories). After electrophoresis, SDS was re-moved from the gel by incubation in 2.5% Triton X-100 at roomtemperature for 45 min with gentle shaking. The gel was washedthoroughly with distilled water to remove detergent and incubated at37°C for 16 h in a developing buffer containing 50 mM Tris·HCl atpH 7.6, 0.2 M NaCl, 5 mM CaCl2, and 0.02% Brij 35 (Sigma-Aldrich). The gel was then stained with a solution of 30% methanol,10% glacial acetic acid, and 0.5% Coomassie blue G250, followed bydestaining in the same solution without dye. Proteinase activity wasdetected as unstained bands on a blue background representing areasof gelatin digestion.
Histology and immunostaining. Paraffin-embedded kidney sectionswere prepared at 4-�m thickness by a routine procedure. Sectionswere stained with hematoxylin and eosin for general histology. Indi-rect immunofluorescence staining was performed on kidney cryosec-tions using an established procedure. Cryosections were prepared at5-�m thickness and fixed for 5 min in PBS containing 3% parafor-maldehyde. After being blocked with 1% normal donkey serum inPBS for 30 min, the sections were incubated with primary antibodiesagainst �-SMA (Sigma-Aldrich) and collagen (Southern Biotechnol-ogy Associates, Birmingham, AL), respectively, in PBS containing1% BSA overnight at 4°C. Sections were then incubated for 1 h withfluorescein isothiocyanate-conjugated secondary antibodies (Sigma-Aldrich) at a dilution of 1:100 in PBS containing 1% BSA, before
F974 LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

being washed extensively with PBS. As a negative control, theprimary antibody was replaced with nonimmune IgG, and no stainingoccurred. Some slides were then stained with proximal tubularmarker, fluorescein-conjugated lectin from Tetragonolobus purpureas(Sigma-Aldrich) for localizing the proximal tubules. Slides weremounted with Vectashield antifade mounting media (Vector Labora-tories, Burlingame, CA) and viewed with an Eclipse E600 epifluo-rescence microscope equipped with a digital camera (Nikon, Melville,NY). Immunohistochemical staining was performed by use of theVector M.O.M. immunodetection kit, as described above (VectorLaboratories). The primary antibody used was anti-fibronectin(Sigma, Sigma-Aldrich). As a negative control, the primary antibodywas replaced with nonimmune normal IgG, and no staining occurred.
For quantitation of TBM lesions in the obstructed kidneys ofMMP-9�/� and MMP-9�/� mice, sections from kidney cortex ofeach mouse were double-stained with collagen/tubular marker. Thepercentages of renal tubules that had one or more breaks in their TBMwere counted in a blinded fashion in randomly chosen, nonoverlap-ping high-power (�400) fields, and the data were calculated based onindividual values that were determined in 10 fields/mouse (n � 5mice/group).
Biochemical measurement of total kidney collagen content. Forquantitative measurement of collagen accumulation and deposition inthe kidney, total tissue collagen content was determined by biochem-ical analysis of the hydroxyproline in the hydrolysates extracted from
kidney samples. This assay is based on the observation that essentiallyall the hydroxyproline in animal tissues is found in collagen. Briefly,accurately weighed portions of the obstructed kidneys were homog-enized in distilled H2O. The homogenates were hydrolyzed in 10 NHCl by incubation at 110°C for 18 h. The hydrolysates were dried byspeed vacuum centrifugation over 3–5 h and redissolved in a buffercontaining 0.2 M citric acid, 0.2 M glacial acetic acid, 0.4 M sodiumacetate, and 0.85 M sodium hydroxide, pH 6.0. Hydroxyprolineconcentrations in the hydrolysates were chemically measured accord-ing to the techniques previously described (17). Total collagen wascalculated on the assumption that collagen contains 12.7% hydroxypro-line by weight. The results of total tissue collagen content were expressedas micrograms of collagen per milligram of kidney weight.
Determination of tissue TGF-�1 levels by ELISA. For measurementof active TGF-�1 protein levels in tissue, kidneys were homogenizedin extraction buffer containing 20 mM Tris·HCl, pH 7.5, 2 M NaCl,0.1% Tween 80, 1 mM ethylenediamine tetraacetate, and 1 mMphenylmethylsulfonyl fluoride, and then the supernatant was recov-ered after centrifugation at 19,000 g for 20 min at 4°C. Kidney tissueTGF-�1 level was determined by using a commercial QuantikineTGF-�1 ELISA kit in accordance with the protocol specified by themanufacturer (R&D Systems). This kit measures the abundance ofactive TGF-�1 protein that binds to its soluble type II receptor, whichis precoated on a microplate. The concentration of TGF-�1 in kidneyswas expressed as picograms per milligram total protein.
Fig. 1. Tubular basement membrane (TBM) integrity is largely preserved in the obstructed kidneys in matrix metalloproteinase-9 MMP-9 gene knockout mice(MMP-9 �/�). A and B: induction of MMP-9 is largely blunted after ureteral obstruction in MMP-9 �/� mice. A: RT-PCR analysis of whole-tissue lysate ofsham and obstructed kidneys at 7 days after unilateral ureteral obstruction (UUO) in MMP-9�/� and MMP-9�/� mice. Representative pictures show the resultsof 2 animals/group. B: zymographic analysis of whole-tissue lysate of the obstructed kidneys at 3 and 7 days after UUO in MMP-9�/� and MMP-9�/� mice.C: Western blot shows collagen IV expression in obstructed kidney at day 7 after UUO in MMP-9�/� and MMP-9�/� mice. D–G: representative micrographsshow collagen IV (major component of TBM; red) and tubular cells (green) in obstructed kidneys at 7 days after UUO in MMP-9�/� (D and E) and MMP-9�/�(F and G) mice. Renal proximal tubules were stained with lectin from Tetragonolobus purpureas (green). The TBM in the kidney of MMP-9�/� mice isdisplayed as severely impaired, broken, and interrupted, whereas TBM structural integrity was largely preserved in MMP-9�/� mice. H: graphic presentationof the percentages of TBM lesions in the obstructed kidneys from MMP-9�/� and MMP-9�/� mice at 7 days after UUO. TBM integrity was examined byimmunofluorescence staining of collagen IV. Results in various groups are expressed as fluorescent integrity of collagen IV and presented as means SE (n � 5).**P 0.01, MMP-9�/� vs. MMP-9�/� mice. †P 0.01 vs. MMP-9�/� sham mice.
F975LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

Boyden chamber coated with Matrigel for motility and invasionassay. Cell motility and migration were evaluated using a Boydenchamber motogenicity assay with tissue culture-treated Transwellfilters (Costar). Matrigel (1.43 mg/cm2) was added onto the Transwellfilters (8-�m pore size, 0.33-cm2 growth area) of the Boyden chamberto form Matrigels at 1.0-mm depth. NRK-52E cells (1 � 104) wereseeded onto the filters (8-�m pore size, 0.33-cm2 growth area) in thetop compartment of the chamber. After 3 days of incubation with orwithout MMP-9 plus GM6001 in the lower compartment at 37°C,filters were fixed with 3% paraformaldehyde in PBS and stained with0.1% Coomassie blue in 10% methanol and 10% actic acid, and theupper surface of the filters was carefully wiped with a cotton-tippedapplicator. Cells that invaded and migrated across the Matrigel andpassed the Transwell filter pores toward the lower surface of the filterswere counted in 5 nonoverlapping �10 fields and photographed witha Nikon microscope. The experiments were performed in triplicate.
Statistical analysis. All the computed data were expressed as means SE. Each of the experiments was repeated three times independently. ForWestern blot analysis, quantitation was performed by scanning andanalyzing the intensity of the hybridization signals using NIH Imagesoftware. Statistical analysis of the data was performed by the Student-Newman-Keuls test using SigmaStat software (Jandel Scientific Soft-ware, San Rafael, CA). Comparisons between groups were made usingone-way ANOVA, followed by the Student-Newman-Keuls test. P 0.05 was considered significant.
RESULTS
TBM integrity is preserved in obstructed kidneys of MMP-9�/�mutant mice. Age- and sex-matched wild-type (MMP-9�/�) andknockout (MMP-9�/�) mice were induced to develop chronic renalinterstitial fibrosis by UUO. The expression and activity of intrarenal
Fig. 2. Reduced �-smooth muscle actin (SMA) induction occurs at day 7 after UUO in mice lacking MMP-9. A: Western blot shows �-SMA expression in theobstructed kidney homogenates probed with antibodies against �-SMA at day 7 after ureteral obstruction in MMP-9�/� and MMP-9�/� mice. The sampleswere reprobed with actin to confirm equal loading of each lane. Representative pictures show the results of 2 animals/group. B: graphic presentation of relative�-SMA abundance normalized to actin in MMP-9�/� and MMP-9�/� mice. **P 0.05 vs. sham mice. †P 0.01, MMP-9�/� vs. MMP-9�/� mice, n � 5.
Fig. 3. Renal morphological injury and interstitial matrix deposition are attenuated in obstructive nephropathy mice lacking MMP-9. A–D: representativemicrographs show kidney morphology with hematoxylin and eosin staining at 7 days after ureteral obstruction in MMP-9�/� (A and B) and MMP-9�/� (Cand D) mice. No appreciable difference in renal morphology and matrix deposition was observed in the kidneys of sham-operated mice between these 2genotypes. However, a significant reduction in morphological injury and interstitial matrix deposition was found at 7 days after UUO in the obstructed kidneysof MMP-9�/� mice. E and F: semiquantitative analysis of injury (E) and collagen score (F) in obstructive kidneys presented as means SE (n � 5). **P 0.01, MMP-9�/� vs. MMP-9�/� mice. †P 0.01, 7 days after UUO vs. MMP-9�/� sham mice.
F976 LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

MMP-9 in MMP-9�/� and MMP-9�/� mice were examined byboth RT-PCR analysis and gelatin zymography. As shown in Fig. 1A,ureteral obstruction for 7 days induced a dramatic increase inMMP-9 mRNA expression in the kidneys of wild-type mice. Asexpected, no MMP-9 mRNA was detected in the kidneys of eithersham-operated or obstructed MMP-9�/� mice. Similarly, noMMP-9 activity was found in the kidney homogenates ofMMP-9 �/� mice whereas a clear band at �92 kDa was evidentin the samples derived from wild-type counterparts after 3 and 7days of obstruction (Fig. 1B).
Because one of the specific substrates for MMP-9 is collagenIV, which is also a major component of TBM, induction ofMMP-9 could lead to destruction of renal TBM integrity,which is one of the key events during tubular EMT. We furtherdirectly examined the collagen IV expression of the obstructedkidneys in MMP-9�/� and MMP-9�/� mice. Western blot-
ting of whole-kidney lysates revealed that the level of collagenIV protein in the kidneys at 7 days after UUO in MMP-9�/�mice was significantly preserved compared with their wild-type controls (Fig. 1C). Immunofluorescence staining withcollagen IV revealed a broken, discontinuous TBM in signifi-cant numbers of renal tubules from the kidneys of MMP-9�/� miceat 7 days after UUO (Fig. 1E). Through the interrupted TBM,tubular epithelial cells tended to migrate toward the interstitialcompartment. However, under identical conditions, TBM in-tegrity was largely preserved with uninterrupted staining ofTBM in MMP-9�/� mice (Fig. 1G). Quantitative determina-tion revealed that �30% of renal tubules in the cortical regionsdisplayed TBM lesions manifested by one or more breaks inthe kidneys of MMP-9�/� mice at 7 days after UUO, whereas15% of renal tubules had similar TBM lesions in MMP-9�/� kidneys (Fig. 1H). These results suggest that deficiencyof MMP-9 expression preserves TBM from destruction underpathological conditions.
Blockade of tubular EMT in MMP-9�/� mice after ureteralobstruction. Because �-SMA-positive myofibroblasts are theprincipal cells responsible for interstitial matrix accumulationand deposition, we next investigated the magnitude of myofi-broblast activation after ureteral obstruction in MMP-9�/�and MMP-9�/� mice. As shown in Fig. 2A, the level of�-SMA protein in the kidneys at 7 days after ureteral obstruc-tion in MMP-9�/� mice was significantly lower than that intheir wild-type controls. Quantitative determination of theWestern blotting of whole-kidney lysates revealed �70% in-hibition of �-SMA protein in MMP-9�/� mice compared withthe levels in MMP-9�/� mice (Fig. 2B). A similar level of�-SMA protein was detected in the sham-operated kidneys inMMP-9�/� and MMP-9�/� mice, indicating that MMP-9null mutation did not affect �-SMA protein expression underbasal conditions. These results suggest that null mutation of theMMP-9 gene attenuates myofibroblast activation by blockingtubular EMT in the diseased kidneys.
Fig. 4. Disruption of MMP-9 gene reduces total collagen accumulated in theobstructed kidney. Total kidney collagen content in the obstructed kidneys insham-operated mice and 7 days after UUO in MMP-9�/� and MMP-9�/�mice was determined by biochemical assay. Results in various groups areexpressed as micrograms per milligram kidney and presented as means SE(n � 5). **P 0.01 vs. all other groups. †P 0.01 vs. MMP-9�/� mice.
Fig. 5. Reduced fibronectin (FN) induction occurs at day 7 after UUO in mice lacking MMP-9. A: Western blot shows FN expression in the obstructed kidneyhomogenates probed with antibodies against fibronectin at day 7 after ureteral obstruction in MMP-9�/� and MMP-9�/� mice. The samples were reprobedwith actin to confirm equal loading of each lane. Representative pictures show the results of 2 animals/group. B: graphic presentation of relative FN abundancenormalized to actin in MMP-9�/� and MMP-9�/� mice. **P 0.05 vs. sham mice. †P 0.01, MMP-9�/� vs. MMP-9�/� mice, n � 5.C–F: immunohistochemical staining of FN protein in the obstructed kidneys at 7 days after UUO in MMP-9�/� (C and D) and MMP-9�/� (F and G) mice (brown).
F977LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

Attenuation of renal interstitial fibrosis in obstructive ne-phropathy in MMP-9�/� mice. Figure 3 shows representativemicrographs of the obstructed kidneys in MMP-9�/� andMMP-9�/� mice at 7 days after surgery. As expected, dis-ruption of the MMP-9 gene resulted in less morphologicalinjury to the obstructed kidneys than those seen in the wild-type. In MMP-9�/� mice with sustained, complete ureteralobstruction for 7 days, kidneys developed severe morpholog-ical lesions characterized by tubular dilation with epithelialatrophy, interstitial expansion with hypercellularity resultingfrom myofibroblast activation, and extracellular matrix accu-mulation. Under the same conditions, renal pathology wassignificantly improved in mice lacking MMP-9 (Fig. 3), sug-gesting that endogenous MMP-9 induces damage by promotingrenal fibrotic lesions after chronic injury.
To quantitatively measure matrix accumulation, we deter-mined the total tissue collagen content in the obstructed kid-neys by a biochemical assay. Figure 4 shows the total kidneycollagen content in the obstructed kidneys at day 7 aftersurgery in MMP-9�/� and MMP-9�/� mice. About sixfold-higher levels of collagen deposition were observed in theobstructed kidneys compared with sham-operated controls atday 7 in MMP-9�/� mice. However, deficiency of MMP-9dramatically inhibited collagen accumulation in the kidneys afterthe same period (7 days) of ureteral obstruction. Thus disruptionof the MMP-9 gene protects the kidney from developing intersti-tial fibrotic lesions after chronic, persistent injury.
To demonstrate ECM protein expression, the magnitude offibronectin after ureteral obstruction in MMP-9�/� and MMP-9�/� mice was investigated. As shown in Fig. 5A, the level offibronectin protein in the kidneys at 7 days after ureteralobstruction in MMP-9�/� mice was significantly lower thanthat in their wild-type controls. Quantitative determination ofthe Western blotting of whole-kidney lysates revealed �80%inhibition of fibronectin protein in MMP-9�/� mice comparedwith the levels in MMP-9�/� mice (Fig. 5B). A similar levelof fibronectin protein was detected in the sham-operated kid-neys in MMP-9�/� and MMP-9�/� mice, suggesting thatMMP-9 deficiency did not alter fibronectin expression under basalconditions. Furthermore, fibronectin deposition in the obstructedkidneys in MMP-9�/� and MMP-9�/� mice was examined byimmunohistochemical staining. As shown in Fig. 5, C–F, at 7days after UUO, marked deposition of fibronectin protein wasobserved in the MMP-9�/� mice (Fig. 5D). In contrast, signifi-cantly less fibronectin was stained in the obstructed kidney ofMMP-9�/� mice at 7 days after surgery, and this was predom-inantly confined to the interstitial compartment (Fig. 5F). Theseresults suggest that null mutation of the MMP-9 gene attenuatesrenal interstitial fibrosis in obstructive nephropathy.
Induction of TGF-�1 axis expression is not altered in ob-structed kidneys in tPA�/� mice. Since TGF-�1 is believed toplay a critical role in initiating and promoting myofibroblastactivation and fibrotic lesions in vivo, we tested the hypothesisthat altered TGF-�1 expression may account for the differencein interstitial fibrosis after chronic injury in MMP-9�/� andMMP-9�/� mice. TGF-�1 protein levels were determined byELISA of whole-kidney lysates. As demonstrated in Fig. 6A,ureteral obstruction caused a marked increase in renal TGF-�1expression in the kidneys in a time-dependent manner. How-ever, no significant difference in TGF-�1 protein level wasfound in the obstructed kidneys in MMP-9�/� and MMP-9�/�
mice at 7 days after UUO, suggesting that the renal protectionseen in MMP-9�/� mice is not attributable to altered TGF-�1induction following persistent injury. Levels of TGF-�1 type Ireceptor were also examined by Western blot analysis of whole-kidney lysates in MMP-9�/� and MMP-9�/� mice after ure-teral obstruction. As shown in Fig. 6B, the protein level of TGF-�1receptor type I was virtually identical between MMP-9�/� andMMP-9�/� mice in the obstructed kidneys at 7 days after UUO.Hence, expression of the TGF-�1 axis does not account for therenal protection elicited by MMP-9 deficiency in obstructivenephropathy.
Decomposition product from MMP-9-digested TBM but notMMP-9 itself induces EMT. We next examined whetherMMP-9 directly induced EMT in vitro. As shown in Fig. 7A,
Fig. 6. Expression of transforming growth factor (TGF)-�1 axis is induced ina similar manner in the obstructed kidneys of MMP-9�/� and MMP-9�/�mice. A: TGF-�1 protein levels in the obstructed kidneys were determined byspecific ELISAs at different time points in MMP-9�/� and MMP-9�/� mice.No significant difference in TGF-�1 induction after ureteral obstruction wasobserved between MMP-9�/� and MMP-9�/� mice. Data are presented asmeans SE (n � 5, P � 0.05). B: Western blot demonstrates TGF-�1 typeI receptor (T�R1) in the obstructed kidneys at day 7 after UUO fromMMP-9�/� and MMP-9�/� mice. The same blot was reprobed with actin toconfirm equal loading of each lane. Representative pictures show the results of2 animals/group. C: graphic presentation of relative T�R1 abundance normal-ized to actin in MMP-9�/� and MMP-9�/� mice. Virtually identical levelsof T�R1 protein were found in MMP-9�/� and MMP-9�/� mice afterquantitative determination.
F978 LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from
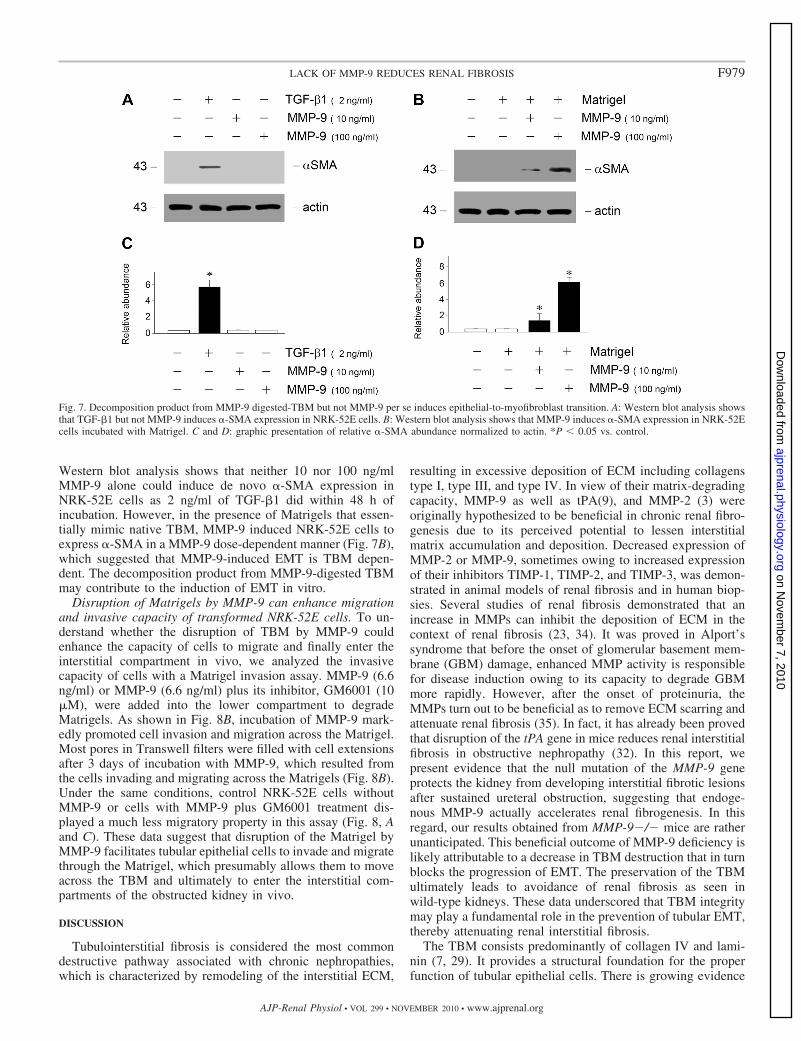
Western blot analysis shows that neither 10 nor 100 ng/mlMMP-9 alone could induce de novo �-SMA expression inNRK-52E cells as 2 ng/ml of TGF-�1 did within 48 h ofincubation. However, in the presence of Matrigels that essen-tially mimic native TBM, MMP-9 induced NRK-52E cells toexpress �-SMA in a MMP-9 dose-dependent manner (Fig. 7B),which suggested that MMP-9-induced EMT is TBM depen-dent. The decomposition product from MMP-9-digested TBMmay contribute to the induction of EMT in vitro.
Disruption of Matrigels by MMP-9 can enhance migrationand invasive capacity of transformed NRK-52E cells. To un-derstand whether the disruption of TBM by MMP-9 couldenhance the capacity of cells to migrate and finally enter theinterstitial compartment in vivo, we analyzed the invasivecapacity of cells with a Matrigel invasion assay. MMP-9 (6.6ng/ml) or MMP-9 (6.6 ng/ml) plus its inhibitor, GM6001 (10�M), were added into the lower compartment to degradeMatrigels. As shown in Fig. 8B, incubation of MMP-9 mark-edly promoted cell invasion and migration across the Matrigel.Most pores in Transwell filters were filled with cell extensionsafter 3 days of incubation with MMP-9, which resulted fromthe cells invading and migrating across the Matrigels (Fig. 8B).Under the same conditions, control NRK-52E cells withoutMMP-9 or cells with MMP-9 plus GM6001 treatment dis-played a much less migratory property in this assay (Fig. 8, Aand C). These data suggest that disruption of the Matrigel byMMP-9 facilitates tubular epithelial cells to invade and migratethrough the Matrigel, which presumably allows them to moveacross the TBM and ultimately to enter the interstitial com-partments of the obstructed kidney in vivo.
DISCUSSION
Tubulointerstitial fibrosis is considered the most commondestructive pathway associated with chronic nephropathies,which is characterized by remodeling of the interstitial ECM,
resulting in excessive deposition of ECM including collagenstype I, type III, and type IV. In view of their matrix-degradingcapacity, MMP-9 as well as tPA(9), and MMP-2 (3) wereoriginally hypothesized to be beneficial in chronic renal fibro-genesis due to its perceived potential to lessen interstitialmatrix accumulation and deposition. Decreased expression ofMMP-2 or MMP-9, sometimes owing to increased expressionof their inhibitors TIMP-1, TIMP-2, and TIMP-3, was demon-strated in animal models of renal fibrosis and in human biop-sies. Several studies of renal fibrosis demonstrated that anincrease in MMPs can inhibit the deposition of ECM in thecontext of renal fibrosis (23, 34). It was proved in Alport’ssyndrome that before the onset of glomerular basement mem-brane (GBM) damage, enhanced MMP activity is responsiblefor disease induction owing to its capacity to degrade GBMmore rapidly. However, after the onset of proteinuria, theMMPs turn out to be beneficial as to remove ECM scarring andattenuate renal fibrosis (35). In fact, it has already been provedthat disruption of the tPA gene in mice reduces renal interstitialfibrosis in obstructive nephropathy (32). In this report, wepresent evidence that the null mutation of the MMP-9 geneprotects the kidney from developing interstitial fibrotic lesionsafter sustained ureteral obstruction, suggesting that endoge-nous MMP-9 actually accelerates renal fibrogenesis. In thisregard, our results obtained from MMP-9�/� mice are ratherunanticipated. This beneficial outcome of MMP-9 deficiency islikely attributable to a decrease in TBM destruction that in turnblocks the progression of EMT. The preservation of the TBMultimately leads to avoidance of renal fibrosis as seen inwild-type kidneys. These data underscored that TBM integritymay play a fundamental role in the prevention of tubular EMT,thereby attenuating renal interstitial fibrosis.
The TBM consists predominantly of collagen IV and lami-nin (7, 29). It provides a structural foundation for the properfunction of tubular epithelial cells. There is growing evidence
Fig. 7. Decomposition product from MMP-9 digested-TBM but not MMP-9 per se induces epithelial-to-myofibroblast transition. A: Western blot analysis showsthat TGF-�1 but not MMP-9 induces �-SMA expression in NRK-52E cells. B: Western blot analysis shows that MMP-9 induces �-SMA expression in NRK-52Ecells incubated with Matrigel. C and D: graphic presentation of relative �-SMA abundance normalized to actin. *P 0.05 vs. control.
F979LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

suggesting that the TBM is not only a static physical barrierthat separates tubular epithelial and interstitial compartmentsbut also a regulator of tubular epithelial cell phenotype andphysiology in the kidney (28). For example, a report shows thatalterations in TBM composition regulate tubular epithelial celltransdifferentiation in vitro (7, 13, 18). It is also reported thatinterstitial collagen type I tends to promote EMT, whereas thecollagen type IV is likely to inhibit this phenotypic transition.It was previously demonstrated that EMT is an orchestrated,highly regulated process consisting of four key steps, in whichdestruction of the TBM plays a crucial role by clearing the pathfor transformed cells to ultimately migrate into the interstitialcompartment of the kidney (31). The present study usingMMP-9�/� mice implies that TBM integrity may play agreater role in EMT than we anticipated before. Preservation ofTBM integrity in MMP-9�/� mice leads to fewer myofibro-blast cells in renal interstitium, which is probably due torestraint of cell migration by the TBM barrier, meanwhilereducing overall �-SMA expression and myofibroblast activa-tion in the obstructed kidneys. This suggests that TBM integ-rity per se may have the ability to block myofibroblast trans-differentiation from tubular epithelia in vivo by preventing theaccess of tubular epithelial cells to the interstitial matrixmicroenvironment after destruction of TBM. Hence, TBMintegrity is likely to be one of the pivotal determinant factors inregulating tubular epithelial cell phenotypic transition underpathological conditions.
Renal EMT and interstitial fibrosis are regulated by severalgrowth factors (1, 8, 10, 16, 37). Profibrogenic TGF-�1 ini-tiates and completes the entire EMT course in cultured tubularepithelial cells in vitro. Renal tubular cells incubated withTGF-�1 lose the epithelial marker E-cadherin, induce de novoexpression of �-SMA, and reorganize actin to form stressfibers. These cells also express abundant MMPs, which canspecifically degrade TBM, and acquire an enhanced motilityand invasive capacity. It is logical to speculate that alteredTGF-�1 expression and activation may account for ameliora-tion of obstructive nephropathy in MMP-9�/� mice. Uponexamination, TGF-�1 induction and activity in the obstructedkidneys were found to be similar in mice either retaining orlacking MMP-9, suggesting that any role for MMP-9 inTGF-�1 activation is compensated for by another mechanismin MMP-9�/� mice. Furthermore, it is suggested that theattenuation of renal myofibroblast activation and interstitialfibrosis in MMP-9�/� mice appears to be independent fromTGF-�1. Thus preservation of TBM integrity alone has majorimplications in preventing EMT and renal interstitial fibrosis atthe whole-animal level.
Our finding on the destruction of TBM integrity by MMP-9suggests that this protease may have much broader functions inthe diseased kidney than we originally thought. Although nottested, we would not be surprised if the expression of othergenes besides collagen could also be regulated by MMP-9.Consistent with this view, there are numerous studies demon-
Fig. 8. Disruption of Matrigel by MMP-9enhances migration of transformed cells.NRK-52E cells were seeded on the Matrigel-coated Transwell filters of a Boyden cham-ber (pore size, 8 um) in different conditionsfor 2 days. The cells or cell extensions thatpassed through the pores of filters werecounted after staining. A–C: representativemicrographs show the pores of the Transwellfilters filled with cell extensions, and thetransformed cells migrated through the poresto the opposite side of filters without MMP-9(A), with MMP-9 (B), or with MMP-9 plusGM6001 (C) for 2 days. D: graphic pre-sentation of the numbers of cells or cellextensions migrated through the pores ofthe filters after incubation for 2 days. *P 0.01 vs. control. †P 0.01 vs. MMP-9without GM6001, n � 3.
F980 LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

strating that components of the MMP systems, such as MMP-2and TIMP-1, elicit a wide range of cellular activities byinitiating specific gene expression in ways other than theirconventional function in the proteolytic cascade (3, 18, 22). Inparticular, MMP-9 has been reported to impact cellular signal-ing pathways that stimulate cell growth at early stages of tumorprogression (19). MMP-9 has also been found to cleave intra-cellular targets and induce mitotic abnormalities and genomicinstability. The Harris group (38) has recently demonstratedthat MMP-9 directly mediated Slug-dependent EMT in NRK-52E cells downstream of TGF-�1. However, in our study, weapplied a condition of lower dose (10 or 100 ng/ml) for ashorter time duration (48 h) of incubation, in which MMP-9alone couldn’t induce de novo �-SMA expression in NRK-52Ecells as TGF-�1 did. However, in the same condition with thepresence of Matrigel, MMP-9 induced NRK-52E cells toexpress �-SMA in a MMP-9 dose-dependent manner. In viewof its ability to digest TBM, although the exact decompositionproduct from MMP-9-digested TBM is unknown; it is plausi-ble to speculate that MMP-9 may function to trigger a cascadeof intracellular signal transduction, leading to expression ofspecific genes.
In summary, the studies presented in this paper demonstratethat deficiency of MMP-9 protects the kidney from developingfibrotic lesions in obstructive nephropathy. This beneficialeffect is likely mediated by a cascade of events that includereduction of extracellular matrix expression, preservation ofTBM integrity, and blockade of tubular EMT. In addition, wehave uncovered a link between TBM destruction and epithelial-to-myofibroblast transition in which the MMP-9 digest TBMand destruct its integrity. We show here that without significantalteration of major pro-fibrogenic TGF-�1, blockade of tubularEMT alone in MMP-9�/� kidneys dramatically amelioraterenal interstitial fibrosis after obstructive injury. Our resultsestablish a vital role and definite contribution of MMP-9 inEMT process of renal interstitial fibrogenesis at the whole-animal level. Hence, blockage of any one of the key steps inTBM destruction will offer unique opportunities for initializingmyofibroblast activation from tubular epithelial cells and forultimately mitigating renal interstitial fibrosis.
ACKNOWLEDGMENTS
Part of this work was presented as an abstract at the annual meeting of theAmerican Society of Nephrology, November 14–19, 2006, San Diego, CA.
GRANTS
This work was supported by National Science Foundation of China Grants30470800/ 30771010/30871201, the “973” Science Program of the Ministry ofScience and Technology, China (2006CB503909), and Jiangsu Province’sOutstanding Medical Academic Leader Program (SWKJ 2006-50) to J. W.Yang.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
REFERENCES
1. Bottinger EP, Bitzer M. TGF-beta signaling in renal disease. J Am SocNephrol 13: 2600–2610, 2002.
2. Catania JM, Chen G, Parrish AR. Role of matrix metalloproteinases inrenal pathophysiologies. Am J Physiol Renal Physiol 292: F905–F911,2007.
3. Cheng S, Pollock AS, Mahimkar R, Olson JL, Lovett DH. Matrixmetalloproteinase 2 and basement membrane integrity: a unifying mech-anism for progressive renal injury. FASEB J 20: 1898–1900, 2006.
4. Daley WP, Peters SB, Larsen M. Extracellular matrix dynamics indevelopment and regenerative medicine. J Cell Sci 121: 255–264, 2008.
5. Eddy AA. Progression in chronic kidney disease. Adv Chronic Kidney Dis12: 353–365, 2005.
6. Hader C, Marlier A, Cantley L. Mesenchymal-epithelial transition inepithelial response to injury: the role of Foxc2. Oncogene 29: 1031–1040,2009.
7. Houard X, Germain S, Gervais M, Michaud A, van den Brule F,Foidart JM, Noel A, Monnot C, Corvol P. Migration-stimulating factordisplays HEXXH-dependent catalytic activity important for promotingtumor cell migration. Int J Cancer 116: 378–384, 2005.
8. Hu K, Lin L, Tan X, Yang J, Bu G, Mars WM, Liu Y. tPA protectsrenal interstitial fibroblasts and myofibroblasts from apoptosis. J Am SocNephrol 19: 503–514, 2008.
9. Hu K, Mars WM, Liu Y. Novel actions of tissue-type plasminogenactivator in chronic kidney disease. Front Biosci 13: 5174–5186, 2008.
10. Kie JH, Kapturczak MH, Traylor A, Agarwal A, Hill-Kapturczak N.Heme oxygenase-1 deficiency promotes epithelial-mesenchymal transitionand renal fibrosis. J Am Soc Nephrol 19: 1681–1691, 2008.
11. Kim SC, Singh M, Huang J, Prestigiacomo CJ, Winfree CJ, SolomonRA, Connolly ES Jr. Matrix metalloproteinase-9 in cerebral aneurysms.Neurosurgery 41: 642–666, 1997.
12. Kim TH, Mars WM, Stolz DB, Michalopoulos GK. Expression andactivation of pro-MMP-2 and pro-MMP-9 during rat liver regeneration.Hepatology 31: 75–82, 2000.
13. Koshikawa N, Giannelli G, Cirulli V, Miyazaki K, Quaranta V. Roleof cell surface metalloprotease MT1-MMP in epithelial cell migration overlaminin-5. J Cell Biol 148: 615–624, 2000.
14. Liu Y. Epithelial to mesenchymal transition in renal fibrogenesis: patho-logic significance, molecular mechanism, and therapeutic intervention. JAm Soc Nephrol 15: 1–12, 2004.
15. Liu Y. New insights into epithelial-mesenchymal transition in kidneyfibrosis. J Am Soc Nephrol 21: 212–222, 2010.
16. Mariappan MM, Shetty M, Sataranatarajan K, Ghosh Choudhury G,Kasinath BS. Glycogen synthase kinase 3beta is a novel regulator of highglucose- and high insulin-induced extracellular matrix protein synthesis inrenal proximal tubular epithelial cells. J Biol Chem 283: 30566–30575,2008.
17. Oda T, Jung YO, Kim HS, Cai X, Lopez-Guisa JM, Ikeda Y, EddyAA. PAI-1 deficiency attenuates the fibrogenic response to ureteral ob-struction. Kidney Int 60: 587–596, 2001.
18. Orlichenko LS, Radisky DC. Matrix metalloproteinases stimulate epi-thelial-mesenchymal transition during tumor development. Clin Exp Me-tastasis 25: 593–600, 2008.
19. Overall CM, Lopez-Otin C. Strategies for MMP inhibition in cancer:innovations for the post-trial era. Nat Rev 2: 657–672, 2002.
20. Page-McCaw A, Ewald AJ, Werb Z. Matrix metalloproteinases and theregulation of tissue remodelling. Nat Rev Mol Cell Biol 8: 221–233, 2007.
21. Picard N, Baum O, Vogetseder A, Kaissling B, Le Hir M. Origin ofrenal myofibroblasts in the model of unilateral ureter obstruction in the rat.Histochem Cell Biol 130: 141–155, 2008.
22. Ronco P, Chatziantoniou C. Matrix metalloproteinases and matrix re-ceptors in progression and reversal of kidney disease: therapeutic perspec-tives. Kidney Int 74: 873–878, 2008.
23. Samuel CS, Zhao C, Bond CP, Hewitson TD, Amento EP, SummersRJ. Relaxin-1-deficient mice develop an age-related progression of renalfibrosis. Kidney Int 65: 2054–2064, 2004.
24. Strutz FM. EMT and proteinuria as progression factors. Kidney Int 75:475–481, 2009.
25. Swetha G, Chandra V, Phadnis S, Bhonde R. Glomerular parietalepithelial cells of adult murine kidney undergo EMT to generate cells withtraits of renal progenitors. J Cell Mol Med. [Epub ahead of print].
26. Tan R, Zhang J, Tan X, Zhang X, Yang J, Liu Y. Downregulation ofSnoN expression in obstructive nephropathy is mediated by an enhancedubiquitin-dependent degradation. J Am Soc Nephrol 17: 2781–2791, 2006.
27. Thrailkill KM, Clay Bunn R, Fowlkes JL. Matrix metalloproteinases:their potential role in the pathogenesis of diabetic nephropathy. Endocrine35: 1–10, 2009.
28. Wiig H, Gyenge C, Iversen PO, Gullberg D, Tenstad O. The role of theextracellular matrix in tissue distribution of macromolecules in normal and
F981LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from

pathological tissues: potential therapeutic consequences. Microcirculation15: 283–296, 2008.
29. Xu J, Rodriguez D, Petitclerc E, Kim JJ, Hangai M, Moon YS, DavisGE, Brooks PC. Proteolytic exposure of a cryptic site within collagentype IV is required for angiogenesis and tumor growth in vivo. J Cell Biol154: 1069–1079, 2001.
30. Yang J, Liu Y. Blockage of tubular epithelial to myofibroblast transitionby hepatocyte growth factor prevents renal interstitial fibrosis. J Am SocNephrol 13: 96–107, 2002.
31. Yang J, Liu Y. Dissection of key events in tubular epithelial to myofi-broblast transition and its implications in renal interstitial fibrosis. Am JPathol 159: 1465–1475, 2001.
32. Yang J, Shultz RW, Mars WM, Wegner RE, Li Y, Dai C, Nejak K,Liu Y. Disruption of tissue-type plasminogen activator gene in micereduces renal interstitial fibrosis in obstructive nephropathy. J Clin Invest110: 1525–1538, 2002.
33. Zeisberg M, Bonner G, Maeshima Y, Colorado P, Muller GA, StrutzF, Kalluri R. Renal fibrosis: collagen composition and assembly regulatesepithelial-mesenchymal transdifferentiation. Am J Pathol 159: 1313–1321, 2001.
34. Zeisberg M, Bottiglio C, Kumar N, Maeshima Y, Strutz F, Muller GA,Kalluri R. Bone morphogenic protein-7 inhibits progression of chronicrenal fibrosis associated with two genetic mouse models. Am J PhysiolRenal Physiol 285: F1060–F1067, 2003.
35. Zeisberg M, Khurana M, Rao VH, Cosgrove D, Rougier JP, WernerMC, Shield CF 3rd, Werb Z, Kalluri R. Stage-specific action of matrixmetalloproteinases influences progressive hereditary kidney disease. PLoSMed 3: e100, 2006.
36. Zeisberg M, Maeshima Y, Mosterman B, Kalluri R. Renal fibrosis.Extracellular matrix microenvironment regulates migratory behavior ofactivated tubular epithelial cells. Am J Pathol 160: 2001–2008, 2002.
37. Zhang G, Kernan KA, Collins SJ, Cai X, Lopez-Guisa JM, Degen JL,Shvil Y, Eddy AA. Plasmin(ogen) promotes renal interstitial fibrosis bypromoting epithelial-to-mesenchymal transition: role of plasmin-activatedsignals. J Am Soc Nephrol 18: 846–859, 2007.
38. Zheng G, Lyons JG, Tan TK, Wang Y, Hsu TT, Min D, Succar L,Rangan GK, Hu M, Henderson BR, Alexander SI, Harris DC. Dis-ruption of E-cadherin by matrix metalloproteinase directly mediates epi-thelial-mesenchymal transition downstream of transforming growth fac-tor-beta1 in renal tubular epithelial cells. Am J Pathol 175: 580–591,2009.
F982 LACK OF MMP-9 REDUCES RENAL FIBROSIS
AJP-Renal Physiol • VOL 299 • NOVEMBER 2010 • www.ajprenal.org
on Novem
ber 7, 2010 ajprenal.physiology.org
Dow
nloaded from