12.adipogenic differentiation potential of rat
DESCRIPTION
adscTRANSCRIPT

Journal of Plastic, Reconstructive & Aesthetic Surgery (2014) 67, 1427e1435
Adipogenic differentiation potential of ratadipose tissue-derived subpopulations ofstromal cells*
M. Gierloff a,*, L. Petersen a, H.-H. Oberg b, E.S. Quabius b,c,J. Wiltfang a, Y. Acil a
a Department of Oral & Maxillofacial Surgery, Christian-Albrechts-University, Kiel, Germanyb Department of Immunology, Christian-Albrechts-University, Kiel, Germanyc Department of Othorhinolaryngology, Head and Neck Surgery, Christian-Albrechts-University, Kiel,Germany
Received 21 January 2014; accepted 20 May 2014
KEYWORDSAdipose-derivedstromal cells;ASCs;Subpopulations;Differentiationpotential;Fat regeneration
* The work should be attributed to: D16, 24105 Kiel, Germany.* Corresponding author. Department
Kiel, Germany. Tel.: þ49 15142319383E-mail address: matthias_gierloff@
http://dx.doi.org/10.1016/j.bjps.2014.01748-6815/ª 2014 British Association of P
Summary Adipose-derived stromal cells (ASCs) are mostly isolated by enzymatic digestion,centrifugation and adherent growth resulting in a very heterogeneous cell population. There-fore, other cell types in the cell culture can comprise the differentiation and proliferation po-tential of the ASC population. Recent studies indicated that an antibody-aided isolation ofdistinct ASC subpopulations provides advantages over the conventional method of ASC isola-tion. The aim of this study was to investigate the adipogenic differentiation potential ofCD29-, CD71-, CD73- and CD90-selected ASCs in vitro.
The stromal vascular fraction (SVF) was obtained from rat adipose tissue by enzymatic diges-tion and centrifugation. Subsequently, CD29þ-, CD71þ-, CD73þ- and CD90þ cells were isolatedby magnetic activated cell sorting (MACS), seeded into culture plates and differentiated intothe adipogenic lineage. ASCs isolated by adherent growth only served as controls. Adipogenicdifferentiation was assessed by Oil Red O staining and quantification of the adiponectin andleptin concentrations in the cell culture supernatants. Statistical analysis was carried out usingone-way analysis of variance (ANOVA) followed by the Scheffe’s post hoc procedure.
The results showed that different subpopulations with different adipogenic differentiationpotentials can be isolated by the MACS procedure. The highest adipogenic differentiation po-tential was determined in the CD29-selected ASC population followed by the unsorted ASC
epartment of Oral & Maxillofacial Surgery, Christian-Albrechts, University Kiel, Arnold-Heller-Strasse
of Oral & Maxillofacial Surgery, Christian-Albrechts-University Kiel, Arnold-Heller-Strasse 16, 24105; fax: þ49 4315972107.icloud.com (M. Gierloff).
5.042lastic, Reconstructive and Aesthetic Surgeons. Published by Elsevier Ltd. All rights reserved.

1428 M. Gierloff et al.
population. The CD71-, CD73- and CD90-selected cells exhibited significantly the lowest adipo-genic differentiation potential.
In conclusion, the CD29-selected ASCs and the unsorted ASCs exhibited a similar adipogenicdifferentiation potential. Therefore, we do not see a clear advantage in the application of ananti-CD29-based isolation of ASCs over the conventional technique using adherent growth.However, the research on isolation/purification methods of adipogenic ASCs should continuein order to make this stem cell source even more attractive for future adipose tissue engineer-ing applications.ª 2014 British Association of Plastic, Reconstructive and Aesthetic Surgeons. Published byElsevier Ltd. All rights reserved.
Introduction
Adipose-derived stromal cells (ASCs) are multipotentialmesenchymal progenitor cells1,2 and represent promisingcandidates for tissue engineering applications.3e6 ASCsreside in the so-called stromal vascular fraction (SVF) ofadipose tissue. The SVF is obtained by enzymatic digestionand centrifugation1,7,8 and consists of a heterogeneous cellpopulation containing haematopoietic cells, adipocytes,endothelial cells, vascular smooth muscle cells, fibroblasts,resident monocytes, macrophages, lymphocytes and theirparticular precursors.5,9e13 The frequency of ASCs in theSVF accounts for approximately 1e5%.14 Currently, themajor drawback in the clinical and experimental use ofASCs is to clearly separate ASCs from other cells and toisolate a pure ASC population which can be expanded anddifferentiated in culture. To date, most experimentalresearch groups isolate ASCs by tissue digestion, centrifu-gation and the capacity of ASCs to adhere to cell cultureplastic surfaces.1,2,4 This technique allows to exclude allnon-adherent growing cells of the SVF and results in arelatively homogeneous cell population9 that contains ahigh number of differentiationable ASCs. However, theadherent cell population also contains other cell typeswhich are not multipotential and comprise the proliferationand differentiation potential of the population. In order toovercome the problem of a “contamination”, Rada et al.used an antibody-aided technique to purify ASCs.15 Thetechnique is based on immunomagnetic beads coated withspecific antibodies against surface molecules of mesen-chymal precursors. The authors could show that CD105þ-and CD29þ-ASCs exhibit a higher chondrogenic differentia-tion potential16 compared to a cell population obtainedsolely by adherent growth. In a different study, theydemonstrated that Stro-1þ-ASCs represent ideal mesen-chymal precursors for bone tissue engineering applica-tions.17 To date, not many studies investigated theantibody-aided selection of ASCs with regard to their adi-pogenic differentiation potential. There is slight evidencethat CD105�-ASCs are more prone to differentiate intoadipocytes compared to CD105þ-ASCs.18 Yet, further nichesof ASC subpopulations and their adipogenic differentiationpotential are not known. The aim of the current study wasto investigate the adipogenic differentiation potential ofCD29-, CD71-, CD73- and CD90-selected cells of the SVF in arat model.
Materials and methods
Animals
Use and care of the animals were approved by the Ministerof Nature, Environment, and Forestry of Schleswig-Holsteinand were in accordance with the local ethics committee(application no. V312-72241.121-14 (53-5/10)). The exper-iments were conducted with cells of two isogenic adultmale rats of the Lewis strain. The animals were kept understerile housing conditions with a fixed day/night cycle atthe central animal facility of the University of Kiel. Theanimals had free access to food and water.
Adipose tissue digestion, cell harvest and cellculture
Two male rats of the Lewis strain were euthanized by CO2
asphyxiation and adipose tissue was explanted from theintra-abdominal cavity and the inguinal area. ASCs wereisolated according to a modified protocol that has beenpreviously described.1 Briefly, the adipose tissue waswashed 3� with phosphate-buffered saline (Gibco�,Paisly, Scotland, UK). For digestion, the tissue was trans-ferred into a sterile 50-ml centrifuge tube containing0.075% collagenase Type II (Sigma Aldrich Co. LLC, Stein-heim, Germany) and was incubated for 30 min at 5% CO2 at37 �C under constant shaking. The enzyme activity wasneutralized by adding equal amounts of Dulbecco’s Modi-fied Eagle’s Medium (DMEM, Biochrom AG, Berlin, Ger-many) and 10% foetal calf serum (FCS; Biochrom AG,Berlin, Germany). The cell suspension was centrifuged at1200 rpm for 5 min and the supernatant was discarded.The cell pellet was resuspended carefully with 10 mlphosphate-buffered saline (PBS) and centrifuged a secondtime. After removing the supernatant, the pellet wasresuspended in 10 ml erythrocyte lysis buffer (Qiagen,Hilden, Germany) and incubated for 10 min at roomtemperature. Following the lysis of red blood cells, thecell suspension was filtered through a 100-mm nylon filter(VWR International GmbH, Damstadt, Germany) to removecellular debris. After another centrifugation, the cellpellet was resuspended in cell culture medium consistingof DMEM (Biochrom AG, Berlin, Germany) supplementedwith FCS 10% (Biochrom AG), 100 IU/ml penicillin
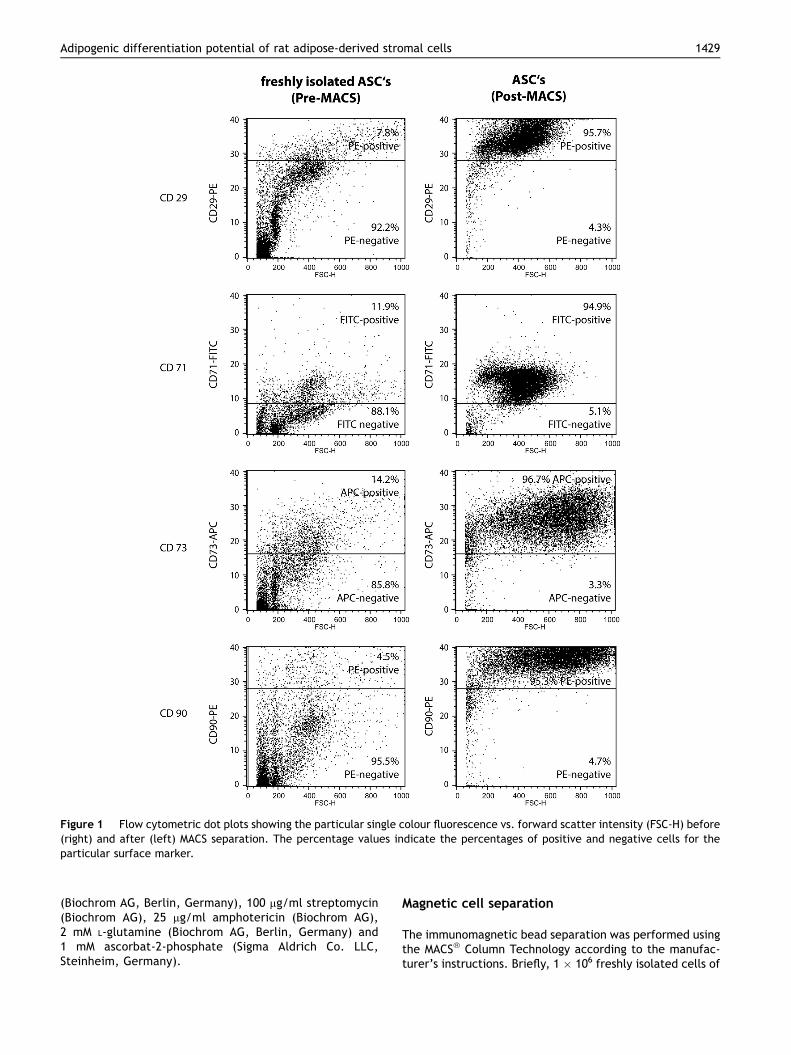
Figure 1 Flow cytometric dot plots showing the particular single colour fluorescence vs. forward scatter intensity (FSC-H) before(right) and after (left) MACS separation. The percentage values indicate the percentages of positive and negative cells for theparticular surface marker.
Adipogenic differentiation potential of rat adipose-derived stromal cells 1429
(Biochrom AG, Berlin, Germany), 100 mg/ml streptomycin(Biochrom AG), 25 mg/ml amphotericin (Biochrom AG),2 mM L-glutamine (Biochrom AG, Berlin, Germany) and1 mM ascorbat-2-phosphate (Sigma Aldrich Co. LLC,Steinheim, Germany).
Magnetic cell separation
The immunomagnetic bead separation was performed usingthe MACS� Column Technology according to the manufac-turer’s instructions. Briefly, 1 � 106 freshly isolated cells of

Figure 2 Photomicrographs (�20) showing the Oil Red O staining of the ASC populations stimulated with adipogenic culturemedium (right) and control medium (left) for 14 days. Significant higher percentages of Oil Red O-positive cells were determined inthe CD29-enriched and the unsorted ASC populations compared to the CD90-enriched, the CD73-enriched or the CD71 enriched ASCpopulations (p < 0.001).
1430 M. Gierloff et al.
the SVF were labelled with a primary antibody againstCD90, CD73, CD71 or CD29, respectively. Subsequently, theprimary antibodies were coupled with a magnetic second-ary antibody (MicroBeads; Miltenyi Biotec GmbH, Bergisch
Gladbach, Germany). The following primary antibodieswere used: IgG1-Anti-Rat-CD90-PE (clone OX-7) (BD Bio-sciences, Heidelberg, Germany), IgG1-Anti-Rat-CD73 (clone5F/B9) (BD Biosciences), IgG2-Anti-Rat-CD71-FITC (clone

Adipogenic differentiation potential of rat adipose-derived stromal cells 1431
OX-26) (BD Biosciences) and IgM-Anti-Rat-CD29-PE (cloneHa2/5) (BD Biosciences). All primary antibodies were usedat a concentration of 0.5 mg/ml. A total of 1 � 106 cells wasstained with 5.0 mg of each particular antibody. Afterwashing the cells in buffer (PBS containing 0.5% bovineserum albumin (BSA) and 2 mM EDTA), the cells werecentrifuged at 300� g for 10 min. The supernatant wasdiscarded and the cell pellet was resuspended in 80 mlbuffer þ either 20 ml Anti-IgG1 MicroBeads or 20 ml Anti-PEMicroBeads according to the manufacturer’s protocol. Forstaining of the CD71-positive cells, 90 ml buffer þ 10 ml Anti-FITC MicroBeads were used. After incubation at 7 �C for15 min the cells were, again, washed in buffer and centri-fuged at 300� g for 10 min. The supernatant was discardedand the cells were resuspended in 500 ml buffer for themagnetic separation process. For this purpose, the MACS�
MS Columns and MACS� Separators (Miltenyi Biotec GmbH,Germany) have been used according to the manufacturer’sinstructions. After the magnetic separation, the number ofviable cells of each cell lineage was determined in a hae-mocytometer after staining with trypan blue. Samples ofeach cell population were measured prior to and after themagnetic separation process on a FACSCalibur flow cytom-eter (Becton Dickinson, Heidelberg, Germany). The datawere analysed using the CellQuest Pro� software (BectonDickinson).
Cell seeding and cell cultureThe CD90-enriched, CD73-enriched, CD71-enriched, CD29-enriched cell populations and the unsorted ASC populationwere seeded at a density of 1250 cells/cm2 into 6-well-plates containing adipogenic induction medium consistingof DMEM (Biochrom AG) supplemented with FCS 10% (Bio-chrom AG), 100 IU/ml penicillin (Biochrom AG), 100 mg/mlstreptomycin (Biochrom AG), 25 mg/ml amphotericine(Biochrom AG), 2 mM L-glutamine (Biochrom AG), 1 mMascorbat-2-phosphate (Sigma Aldrich Co. LLC), 0.5 mMdexamethasone, 0.5 mM isobutylmethylxanthine, 50 mMindomethacin and 10 mg/ml insulin (Biochrom AG). ASCpopulations cultured in “normal” culture medium withoutadipogenic supplements served as control. The cells werecultured under standard conditions in a humidified atmo-sphere with 5% CO2 at 37
�C for 14 days. Medium change wasperformed every 3 days.
Oil Red O stainingAt day 14 of the culture period, the Oil Red O staining wasperformed as previously described.19 Briefly, the Oil Red Oworking solution was prepared using three parts of stocksolution in 0.5% isopropanol (all from Sigma Aldrich Co.LLC, Steinheim, Germany) and two parts of water andsubsequently filtered through a filter funnel. Afterwashing the cell culture, 60% isopropanol was added for4 min prior to staining with the Oil Red O working solutionfor 5 min. Undifferentiated cells, cultured without adi-pogenic supplements, served as controls. In four differentwells of the 6-well-plate, two fields of view(0.72 � 1.08 mm in size) were randomly picked and thetotal number of cells and the number of Oil Red O-positivecells was assessed using a haemocytometer. Numbers ofOil Red O-positive cells were calculated as a percentageof total cells.
Enzyme-linked immunosorbent assay
LeptinLeptin is a secreted proteohormone which is mainly pro-duced by adipocytes. At day 14 of the culture period, thecell culture supernatants were collected for determinationof the leptin concentration by an enzyme-linked immuno-sorbent assay (ELISA) using the Leptin Rat ELISA Kit (Abcamplc, Cambridge, UK). The test is based on a polyclonalantibody against rat leptin that is coated on a 96-well plate.The test was performed according to the manufacturer’sinstructions. Briefly, standards and samples were preparedand pipetted into the wells. After washing the wells, thebiotinylated Anti-Rat Leptin antibody was added. Unboundbiotinylated antibody was removed by another washingstep and horseradish peroxidase (HRP)-conjugated strep-tavidin was pipetted into the wells. The wells were washedagain, and in order to initiate the colour development, the3,30,5,50-tetramethylbenzidine (TMB) substrate solutionwas applied to the wells. At last, the stop solution waspipetted into the wells and the extinction was measured at450 nm using the Roche Hitachi Modular P (Roche DiagnosticGmbH, Mannheim, Germany). Three independent testswere performed.
AdiponectinAdiponectin is a secreted protein expressed exclusively bydifferentiated adipocytes. At day 14 of the culture period,the cell culture supernatants were collected for determi-nation of the adiponectin concentration by an ELISA usingthe Adiponectin Rat ELISA Kit (Abcam plc, Cambridge, UK).The test is based on a polyclonal antibody, specific for ratadiponectin, that has been pre-coated onto a 96-wellmicroplate. Adiponectin in standards and samples is sand-wiched by an immobilized polyclonal antibody and a bio-tinylated polyclonal antibody against rat adiponectin,which is recognized by a streptavidineperoxidase conju-gate. The test was performed as described above (seeLeptin) according to the manufacturer’s instructions. Threeindependent tests were performed.
Statistics
Normal distribution was proved by using the Kolmogor-oveSmirnoff test and the ShapiroeWilk test. Statisticalanalysis was carried out using one-way analysis of variance(ANOVA) followed by the Scheffe’s post hoc procedure. Thelevel of statistical significance was set at p � 0.05.
Results
MACS
The sedimented cell pellet of the SVF contained 7.8% CD29-positive cells, 11.9% CD71-positive cells, 14.2% CD73-positive cells and 4.5% CD90-positive cells (Figure 1).After the magnetic separation a 12.3-fold enrichment ofthe CD29-positive cells (95.7%), an 8.0-fold enrichment ofthe CD71-positive cells (94.9%), a 6.8-fold enrichment of
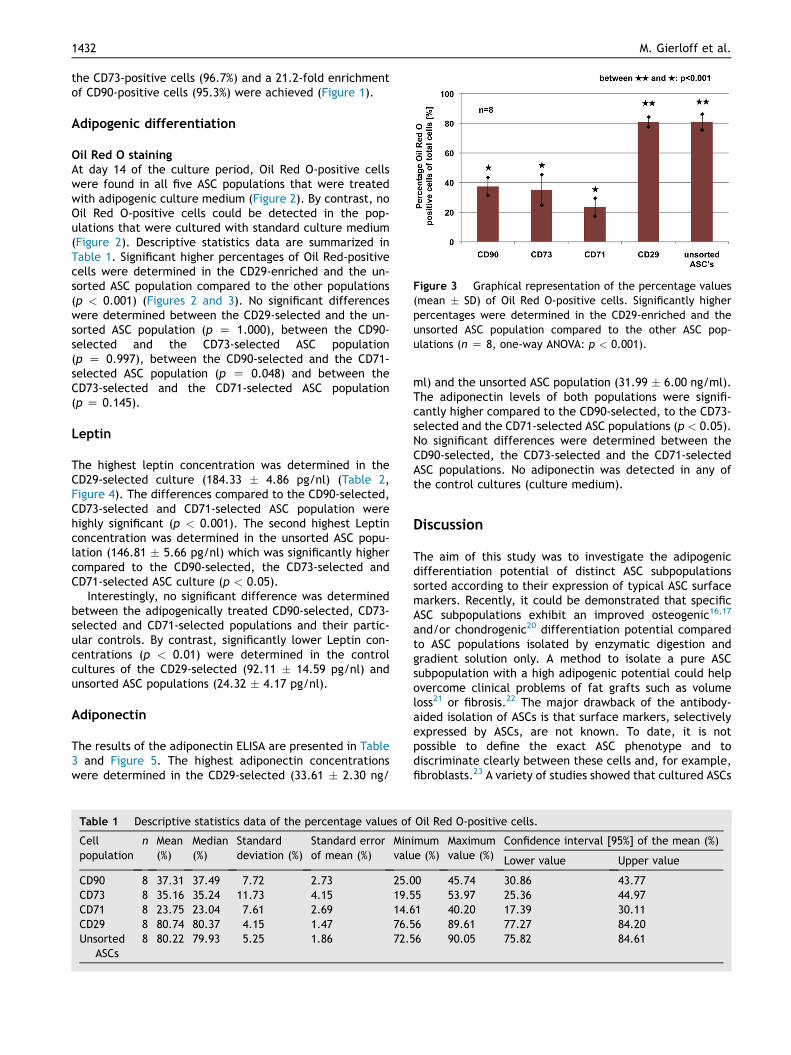
Figure 3 Graphical representation of the percentage values(mean � SD) of Oil Red O-positive cells. Significantly higherpercentages were determined in the CD29-enriched and theunsorted ASC population compared to the other ASC pop-ulations (n Z 8, one-way ANOVA: p < 0.001).
1432 M. Gierloff et al.
the CD73-positive cells (96.7%) and a 21.2-fold enrichmentof CD90-positive cells (95.3%) were achieved (Figure 1).
Adipogenic differentiation
Oil Red O stainingAt day 14 of the culture period, Oil Red O-positive cellswere found in all five ASC populations that were treatedwith adipogenic culture medium (Figure 2). By contrast, noOil Red O-positive cells could be detected in the pop-ulations that were cultured with standard culture medium(Figure 2). Descriptive statistics data are summarized inTable 1. Significant higher percentages of Oil Red-positivecells were determined in the CD29-enriched and the un-sorted ASC population compared to the other populations(p < 0.001) (Figures 2 and 3). No significant differenceswere determined between the CD29-selected and the un-sorted ASC population (p Z 1.000), between the CD90-selected and the CD73-selected ASC population(p Z 0.997), between the CD90-selected and the CD71-selected ASC population (p Z 0.048) and between theCD73-selected and the CD71-selected ASC population(p Z 0.145).
Leptin
The highest leptin concentration was determined in theCD29-selected culture (184.33 � 4.86 pg/nl) (Table 2,Figure 4). The differences compared to the CD90-selected,CD73-selected and CD71-selected ASC population werehighly significant (p < 0.001). The second highest Leptinconcentration was determined in the unsorted ASC popu-lation (146.81 � 5.66 pg/nl) which was significantly highercompared to the CD90-selected, the CD73-selected andCD71-selected ASC culture (p < 0.05).
Interestingly, no significant difference was determinedbetween the adipogenically treated CD90-selected, CD73-selected and CD71-selected populations and their partic-ular controls. By contrast, significantly lower Leptin con-centrations (p < 0.01) were determined in the controlcultures of the CD29-selected (92.11 � 14.59 pg/nl) andunsorted ASC populations (24.32 � 4.17 pg/nl).
Adiponectin
The results of the adiponectin ELISA are presented in Table3 and Figure 5. The highest adiponectin concentrationswere determined in the CD29-selected (33.61 � 2.30 ng/
Table 1 Descriptive statistics data of the percentage values of
Cellpopulation
n Mean(%)
Median(%)
Standarddeviation (%)
Standard errorof mean (%)
Minvalu
CD90 8 37.31 37.49 7.72 2.73 25.0CD73 8 35.16 35.24 11.73 4.15 19.5CD71 8 23.75 23.04 7.61 2.69 14.6CD29 8 80.74 80.37 4.15 1.47 76.5Unsorted
ASCs8 80.22 79.93 5.25 1.86 72.5
ml) and the unsorted ASC population (31.99 � 6.00 ng/ml).The adiponectin levels of both populations were signifi-cantly higher compared to the CD90-selected, to the CD73-selected and the CD71-selected ASC populations (p < 0.05).No significant differences were determined between theCD90-selected, the CD73-selected and the CD71-selectedASC populations. No adiponectin was detected in any ofthe control cultures (culture medium).
Discussion
The aim of this study was to investigate the adipogenicdifferentiation potential of distinct ASC subpopulationssorted according to their expression of typical ASC surfacemarkers. Recently, it could be demonstrated that specificASC subpopulations exhibit an improved osteogenic16,17
and/or chondrogenic20 differentiation potential comparedto ASC populations isolated by enzymatic digestion andgradient solution only. A method to isolate a pure ASCsubpopulation with a high adipogenic potential could helpovercome clinical problems of fat grafts such as volumeloss21 or fibrosis.22 The major drawback of the antibody-aided isolation of ASCs is that surface markers, selectivelyexpressed by ASCs, are not known. To date, it is notpossible to define the exact ASC phenotype and todiscriminate clearly between these cells and, for example,fibroblasts.23 A variety of studies showed that cultured ASCs
Oil Red O-positive cells.
imume (%)
Maximumvalue (%)
Confidence interval [95%] of the mean (%)
Lower value Upper value
0 45.74 30.86 43.775 53.97 25.36 44.971 40.20 17.39 30.116 89.61 77.27 84.206 90.05 75.82 84.61

Table 2 Descriptive statistics data of the leptin concentrations in the cell culture supernatants of the adipogenically inducedASCs determined by ELISA.
Cellpopulation
n Mean(pg/ml)
Median Standarddeviation
Standard errorof mean
Minimumvalue
Maximumvalue
Confidence interval [95%] of the mean
Lower value Upper value
CD90 3 43.38 39.22 9.51 5.49 36.66 54.26 19.76 67.00CD73 3 59.27 59.67 11.27 6.51 47.80 70.33 31.27 87.26CD71 3 92.47 94.97 21.48 12.40 69.85 112.58 39.12 145.81CD29 3 184.33 184.04 4.86 2.80 179.63 189.33 172.27 196.40Unsorted
ASCs3 146.81 147.56 5.66 3.27 140.81 152.05 132.75 160.86
Figure 4 Graphical representation of the leptin concentra-tions (mean � SD) in the cell culture supernatants of the adi-pogenically induced ASCs (red) and the ASCs cultured in“normal” culture medium (white) on day 14 of the cultureperiod (n Z 3, one-way ANOVA).
Adipogenic differentiation potential of rat adipose-derived stromal cells 1433
are positive for CD166, CD151 CD105, CD90, CD73, CD63CD71, CD49d, CD44, CD34, CD29, CD13 and negative forCD146, CD45, CD31.4,5,9,10,24,25 However, none of thesemarkers allow the exact discrimination of ASCs from othercell types. Another drawback is presented by the fact thatthe expression of known ASC surface markers changesdramatically with plating and successive passaging. Forexample, freshly isolated cells of the SVF barely express thestromal cell-associated markers CD13, CD29, CD44, CD73 orCD90. Yet, during culture the expression of these markersincreases.9 The results obtained by the Oil Red O staining
Table 3 Descriptive statistics data of the adiponectin concentrinduced ASCs determined by ELISA.
Cellpopulation
n Mean(ng/ml)
Median Standarddeviation
Standard errorof mean
Mva
CD90 3 12.17 11.41 2.19 1.27 10CD73 3 17.75 17.60 0.72 0.42 17CD71 3 20.82 19.25 3.52 2.03 18CD29 3 33.61 34.84 2.30 1.33 30Unsorted
ASCs3 31.99 29.46 6.00 3.46 27
and the quantification of adiponectin and leptin in the cellculture supernatants demonstrated the highest adipogenicdifferentiation potential for the CD29-selected ASC popu-lation. The differences compared to the CD71-selected, theCD73-selected and the CD90-selected cell populations weresignificant for all tests (p < 0.05). Interestingly, the un-sorted ASCs showed a similar or only slightly lower adipo-genic differentiation potential compared to the CD29-selected ASCs. The reasons why the CD90-, CD73- andCD71-selected ASCs exhibited a significantly lower adipo-genic differentiation potential compared to the other twopopulations remain unclear. One reason could be thepresence of fibroblasts which tend to overgrow other cellsin a culture. The transferrin receptor 1 (CD71) is highlyexpressed not only on erythroid precursors26 and ASCs2,27
but also to a large amount on fibroblasts.28 It is possiblethat the growth of CD71-selected ASCs was limited by CD71-selected fibroblasts. Likewise, the surface molecules CD73and CD90, which are known to be mesenchymal stromal cellmarkers,29 are also expressed on specific fractions offibroblasts.30e32 A second explanation could be that theselected CD71þ, CD73þ and CD90þ cells of the stromalvascular fraction were in an earlier developmental stage ofadipocyte differentiation compared to CD29þ cells. Incontrast to CD29, the other markers are not constitutivelyexpressed on all adipocyte precursors during the differen-tiation towards a mature adipocyte.27 Furthermore, it ispossible that specific cells, which are required for efficientadipocyte differentiation, were not included in the CD90-,CD73- and CD71-selected populations. It is known that thepreadipocyte growth and differentiation is controllednotably by multiple cellecell interactions33 as, forexample, between preadipocytes and adipocytes.34 Yet,adipocytes do not express regularly CD71, CD73 or CD90
ations in the cell culture supernatants of the adipogenically
inimumlue
Maximumvalue
Confidence interval [95%] of the mean
Lower value Upper value
.37 14.57 6.68 17.55
.12 18.54 15.96 19.55
.35 24.85 12.07 29.57
.96 35.03 27.90 39.32
.68 38.84 17.10 46.89

Figure 5 Graphical representation of the adiponectin con-centrations (mean � SD) in the cell culture supernatants of theadipogenically induced ASCs on day 14 of the culture period.No adiponectin was detected in any of the control cultures(culture medium). The error bars indicate the standard error ofmean (n Z 3, one-way ANOVA).
1434 M. Gierloff et al.
and, therefore, were mainly excluded by the MACS proce-dure. In this case, the selection of a distinct ASC subpop-ulation would constitute a clear disadvantage compared tochoosing the heterogeneous SVF for adipogenic cultures.
In conclusion, the highest adipogenic potential wasdetermined in the CD29-selected population. The surfacemolecule CD29 (integrin b1) is expressed by numerousprecursor cells during differentiation towards mature adi-pocytes35 including ASCs,4 early-stage adipocyte precursorsand committed adipocytes.24 However, all these differentcell types are also included in the SVF, which is reflected bythe comparably high adipogenic potential of the unsortedpopulation. Therefore, according to the results of thecurrent study, we do not see a clear advantage in theapplication of the antibody-aided isolation of ASCs by usingthe chosen surface molecules. However, as long as nospecific ASC surface markers are available, the search forideal surface molecules for a positive selection of ASCs or,otherwise, for a negative selection in order to exclude non-differentiationable cells such as fibroblasts from a pure ASCculture, should continue.
Conflict of interest
None.
Funding
None.
Acknowledgements
The authors thank the medical faculty of the Christian-Albrechts University Kiel (grant number: F342972) for the
financial support of this study. They also thank Mrs. Refrathand Mrs. Neßenius for their kind support and technicalassistance. The authors declare that they did not makedemand on any writing assistance.
References
1. Zuk PA, Zhu M, Mizuno H, et al. Multilineage cells from humanadipose tissue: implications for cell-based therapies. TissueEng 2001;7:211e28.
2. Zuk PA, Zhu M, Ashjian P, et al. Human adipose tissue is asource of multipotent stem cells. Mol Biol Cell 2002;13:4279e95.
3. Uysal CA, Tobita M, Hyakusoku H, Mizuno H. Adipose-derivedstem cells enhance primary tendon repair: biomechanical andimmunohistochemical evaluation. J Plast Reconstr AesthetSurg 2012;65:1712e9.
4. Yoshimura K, Shigeura T, Matsumoto D, et al. Characterizationof freshly isolated and cultured cells derived from the fattyand fluid portions of liposuction aspirates. J Cell Physiol 2006;208:65e76.
5. Yoshimura K, Suga H, Eto H. Adipose-derived stem/progenitorcells: roles in adipose tissue remodeling and potential use forsoft tissue augmentation. Regen Med 2009;4:265e73.
6. Zografou A, Tsigris C, Papadopoulos O, et al. Improvement ofskin-graft survival after autologous transplantation of adipose-derived stem cells in rats. J Plast Reconstr Aesthet Surg 2011;64:1647e56.
7. Van RL, Bayliss CE, Roncari DA. Cytological and enzymologicalcharacterization of adult human adipocyte precursors in cul-ture. J Clin Invest 1976;58:699e704.
8. Van RL, Roncari DA. Isolation of fat cell precursors from adultrat adipose tissue. Cell Tissue Res 1977;181:197e203.
9. Mitchell JB, McIntosh K, Zvonic S, et al. Immunophenotype ofhuman adipose-derived cells: temporal changes in stromal-associated and stem cell-associated markers. Stem Cells2006;24:376e85.
10. Mizuno H, Tobita M, Uysal AC. Concise review: adipose-derivedstem cells as a novel tool for future regenerative medicine.Stem Cells 2012;30:804e10.
11. Tholpady SS, Liull R, Ogle RC, Rubin JP, Futrell JW, Katz AJ.Adipose tissue: stem cells and beyond. Clin Plast Surg 2006;33:55e62.
12. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL,Ferrante Jr AW. Obesity is associated with macrophage accu-mulation in adipose tissue. J Clin Invest 2003;112:1796e808.
13. Xu H, Barnes GT, Yang Q, et al. Chronic inflammation in fatplays a crucial role in the development of obesity-related in-sulin resistance. J Clin Invest 2003;112:1821e30.
14. Jurgens WJ, Oedayrajsingh-Varma MJ, Helder MN, et al. Effectof tissue-harvesting site on yield of stem cells derived fromadipose tissue: implications for cell-based therapies. Cell Tis-sue Res 2008;332:415e26.
15. Rada T, Reis RL, Gomes ME. Novel method for the isolation ofadipose stem cells (ASCs). J Tissue Eng Regen Med 2009;3:153e9.
16. Rada T, Reis RL, Gomes ME. Distinct stem cells subpopulationsisolated from human adipose tissue exhibit different chon-drogenic and osteogenic differentiation potential. Stem CellRev 2011;7:64e76.
17. Rada T, Gomes ME, Reis RL. A novel method for the isolation ofsubpopulations of rat adipose stem cells with different prolif-eration and osteogenic differentiation potentials. J Tissue EngRegen Med 2011;5:655e64.
18. Anderson P, Carillo-Galvez AB, Garcıa-Perez A, Cobo M,Martın F. CD105 (endoglin)-negative murine mesenchymal

Adipogenic differentiation potential of rat adipose-derived stromal cells 1435
stromal cells define a new multipotent subpopulation withdistinct differentiation and immunomodulatory capacities.PLoS One 2013;8:e76979.
19. Gierloff M, Nitsche T, Adam-Klages S, et al. In vitro comparisonof different carrier materials with rat bone marrow MSC’s. ClinOral Investig 2014;18:247e59.
20. Jiang T, Liu W, Lv X, et al. Potent in vitro chondrogenesis ofCD105 enriched human adipose-derived stem cells. Bio-materials 2010;31:3564e71.
21. Chung MT, Hyun JS, Lo DD, et al. Micro-computed tomographyevaluation of human fat grafts in nude mice. Tissue Eng Part CMethods 2013;19:227e32.
22. Zhu M, Zhou Z, Chen Y, et al. Supplementation of fat graftswith adipose-derived regenerative cells improves long-termgraft retention. Ann Plast Surg 2010;64:222e8.
23. Mizuno H, Nambu M. Adipose-derived stem cells for skinregeneration. Methods Mol Biol 2011;702:453e9.
24. Rodeheffer MS, Birsoy K, Friedman JM. Identification of whiteadipocyte progenitor cells in vivo. Cell 2008;135:240e9.
25. Zannettino AC, Paton S, Arthur A, et al. Multipotential humanadipose-derived stromal cells exhibit a perivascular phenotypein vitro and in vivo. J Cell Physiol 2008;214:413e21.
26. Marsee DK, Pinkus GS, Yu H. CD71 (transferrin receptor): aneffective marker for erythroid precursors in bone marrow bi-opsy specimens. Am J Clin Pathol 2010;134:429e35.
27. Cawthorn WP, Scheller EL, MacDougald OA. Adipose tissue stemcells meet preadipocyte commitment: going back to thefuture. J Lipid Res 2012;53:227e46.
28. Ekblom P, Thesleff I, Lehto VP, Virtanen I. Distribution of thetransferrin receptor in normal human fibroblasts and fibrosar-coma cells. Int J Cancer 1983;15:111e7.
29. Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria fordefining multipotent mesenchymal stromal cells. The interna-tional society for cellular therapy position statement. Cyto-therapy 2006;8:315e7.
30. Alt E, Yan Y, Gehmert S, et al. Fibroblasts share mesenchymalphenotypes with stem cells, but lack their differentiation andcolony-forming potential. Biol Cell 2011;103:197e208.
31. Choi HS, Ryu CJ, Choi HM, et al. Effects of the pro-inflammatory milieu on the dedifferentiation of culturedfibroblast-like synoviocytes. Mol Med Rep 2012;5:1023e6.
32. Saalbach A, Kraft R, Herrmann K, Haustein UF, Anderegg U. Themonoclonal antibody AS02 recognizes a protein on human fi-broblasts being highly homologous to Thy-1. Arch Dermatol Res1998;290:360e6.
33. Gregoire FM, Smas CM, Sul HS. Understanding adipocyte dif-ferentiation. Physiol Rev 1998;78:783e809.
34. Lai N, Sims JK, Jeon NL, Lee K. Adipocyte induction of pre-adipocyte differentiation in a gradient chamber. Tissue EngPart C Methods 2012;18:958e67.
35. Staquet MJ, Dezutter-Dambuyant C, Schmitt D, Amiot M,Boumsell L, Thivolet J. A surface glycoprotein complex relatedto the adhesive receptors of the VLA family, shared byepidermal Langerhans cells and basal keratinocytes. J InvestDermatol 1989;92:739e45.