2007 -- fabricius et al_octocorals palau - monogr
TRANSCRIPT

79
Chapter 7 Octocorallia in Palau, Micronesia: Effects of biogeography and coastal influences on local and regional biodiversity
K.E. Fabricius1, P. Alderslade2, G.C. Williams3, P.L. Colin4, and Y. Golbuu5
1) Australian Institute of Marine Science, PMB No. 3, Townsville MC, Qld 4810, Australia. [email protected] 2) Museum and Art Gallery of the Northern Territory, GPO Box 4646, Darwin NT 0801, Australia.
3) California Academy of Sciences, Department of Invertebrate Zoology & Geology, 875 Howard Street, San Francisco, CA 94103-3009
4) Coral Reef Research Foundation, P.O. Box 1765, Koror, Palau 96940 5) Palau International Coral Reef Center, Koror, Palau
In: Kayanne H, Omori M, Fabricius K, Verheij E, Colin P, Golbuu Y, Yurihira H (eds) Coral Reefs of Palau. Palau International Coral Reef Centre, Palau.
1. Introduction Octocorals (subclass Octocorallia; orders Alcyonacea - soft corals and sea fans, Pennatulacea - sea pens, and Helioporacea - blue corals) are an abundant and species-rich group of reef inhabiting organisms on Indo-Pacific coral reefs, including Palau. More than 100 genera in 23 families of Alcyonacea (e.g., Alderslade 2001, 2002; Fabricius and Alderslade 2001; van Ofwegen 2005), 9 genera in 5 families of Pennatulacea (Williams 1995), and 1 genus in 1 family of Helioporacea (Blainville 1830), have been described from shallow waters of the Indo-Pacific to date. Previous taxonomic inventories of octocorals from the tropical and subtropical Indo-Pacific include those from the Southern Red Sea (Benayahu et al. 2002), Southern Africa (Williams 1992), Taiwan (Benayahu et al. 2004), Southern Japan (Benayahu 1995), Guam (Benayahu, 1997), Hong Kong (Fabricius and McCorry 2006), the Great Barrier Reef and other parts of Australia (Verseveldt 1977), Fabricius and De’ath, in prep.), Hansa Bay in north New Guinea (Tursch and Tursch 1982), New Caledonia (Grasshoff 1999), and the Marianas Islands (Paulay et al. 2003). Based on these and numerous other publications and museums records, Fabricius and Alderslade (2001) have compiled the distribution ranges of many of the Indo-Pacific shallow warm water genera of Alcyonacea as they are presently known. However, there are still major gaps in the understanding of octocoral distribution ranges. Many of the past inventories had focused on subsets of the octocoral families or growth forms, namely ‘soft corals’ or ‘sea fans’, and most excluded the sea pens that tend to live in soft bottom environments. Under-represented are also systematic surveys from the global center of marine biodiversity, the ‘Coral Triangle’ containing the archipelagos of Indonesia (especially Sulawesi and West Papua), Papua New Guinea and the Philippines (Briggs 1995, Allen 2000, Veron 2000). The main archipelago of Palau is located 900-1200 km each to the east of the Philippines, and north of western New Guinea and other areas of Indonesia’s Sulawesi. Due to the open water barrier to the main coral triangle area, Palau’s octocoral
fauna is somewhat reduced from that found in the area of highest diversity. Oceanic areas near Palau in the western Pacific are dominated by eddy systems (Heron et al., in press) which would appear to favor transport of planktonic larvae from coral triangle regions towards Palau. However, the dispersal abilities of larval stages of soft corals are poorly known and many may not be able to utilize such eddies for dispersal. To the southwest of the main Palau are a group of widely spaced oceanic islands and one atoll (the 'Southwest Islands") which reach to near the western end of New Guinea. The open water gaps between these islands are no more than about 300 km, greatly reducing the absolute distances between shallow water areas, but these islands are only limited in size which may reduce their effectiveness as "stepping stones". Species richness of a given site within a larger area (such as for a single reef in the Palau Archipelago) is affected by three factors. First, the biogeographic location and colonization history of a region determines the regional species pool present (Ricklefs 2004). Second, severe environmental conditions may limit how much of the local species pool occurs at that locality. Octocorals can occur on many types of coral reefs (Benayahu and Loya 1981, Tursch and Tursch 1982, Dinesen 1983, Fabricius 1997, Fabricius and De'ath 2001), however the abundances of particular taxa are known to be strongly determined by the physical environment, especially light availability and water currents (Fabricius and De'ath 1997). Third, at any point in time local and regional species richness also depend on disturbance history, specifically the nature and intensity of the disturbance, and the time since past disturbances have removed colonies. For octocorals, disturbances include episodes of high water temperatures (causing coral bleaching), storms with high wave energy (dislodging or damaging colonies), chronically reduced water clarity (reducing photosynthesis) and sedimentation (smothering colonies or hampering larval settlement). After a disturbance, the speed and efficiency of recolonization of a taxon will determine whether the taxon will again be present or not: fast colonizers continuously re-establish if propagules from surviving colonies (locally or further upstream) are

80
Figure 1: About half of the octocoral genera contain endosymbiontic algae and need light for photosynthesis; they are generally brownish in colour and grow in shallow waters. Here a large, fleshy Sarcophyton colony (top left), and a colony of Blue Coral (Heliopora) which forms massive skeletons (top right).
available, whereas slow-colonizing or slow-growing taxa may take decades to re-establish previous abundances. Similarly, chronic disturbance such as water pollution reduces diversity, because only persistent species can survive and flourish. In order to understand regional and local biodiversity patterns, biogeographic settings, environmental requirements of taxa and consequences of disturbances need to be investigated simultaneously. Some knowledge of the biology of octocorals also helps explaining distribution patterns. For example, more than half of the warm shallow-water Indo-Pacific octocorals are phototrophic and depend on light for carbon gain. Colonies of these taxa contain endosymbiotic dinoflagellate algae (zooxanthellae) in their tissue which supply the colonies with energy. Phototrophic taxa Figure 2: Octocoral taxa without algal endosymbionts such as this Dendronephthya (top left) are generally brightly coloured. Without the need for light, these taxa can grow on steep walls and at greater depths. Some species such as this Siphonogorgia can grow to large sizes in areas of strong currents (left).

81
include the “true” soft corals (the Alcyoniina group, especially many genera within the abundant families Nephtheidae, Alcyoniidae and Xeniidae), but also a few members of most of the other large octocoral groupings, such as some from the Stolonifera group, the Gorgoniidae and the blue coral Heliopora coerulea (Figs. 1 and 3), to name just a few (review in van Oppen et al. 2005). In contrast, most sea fans (i.e., the Scleraxonia group, and most Indo-Pacific genera within the suborders Holaxonia and Calcaxonia, including most Plexauridae, Ellisellidae), and also several of the Alcyoniina (especially among the families Nephtheidae and Nidaliidae) do not contain dinoflagellate endosymbionts. These azooxanthellate genera are heterotrophic suspension feeders that do not depend on light but need currents to transport food particles towards the polyps. Heterotrophic taxa are easily visually distinguished from their phototrophic relatives by their bright coloration (generally yellow, orange, red, pink, purple or a bright white; Fig. 2). In this study we provide a first taxonomic inventory of the octocoral genera and families so far recorded from the reefs of Palau between 0 and 300 m depth. We also determine how the octocoral communities, their cover and taxonomic richness change along gradients in environmental conditions within some of the many and diverse types of shallow-water habitats found in the archipelago of Palau, and interpret the findings in the context of known biogeographic distribution ranges of some of the main octocoral genera.
2. Methods a) Taxonomic inventories Systematic and opportunistic taxonomic inventories were conducted in many habitats. The Coral Reef Research Foundation, as part of the US National Cancer Institute marine collection program in Palau, made a focused effort to sample all possible species of octocorals. Sampling was done by conventional (compressed air) scuba equipment from 1-75 m depth, mix-gas scuba (trimix) from 75-150 m and from 75-300 m using a small submersible (Deepworker 2000). Comparable collections were made to 60 m depth in the countries adjacent to Palau (Philippines, Papua New Guinea, Indonesia and Federated States of Micronesia) by the same collectors using air scuba equipment. This allowed them to become familiar with the field characters of many genera presently unknown from Palau and the inability of these collectors to find these genera from Palau imply these genera are not present. The Southwest Islands of Palau were sampled on two cruises (Fana, Sonsorol and Merir in 1996, Tobi and Helen Reef in 1997) of only one week each. Consequently the collections obtained there were fragmentary and a definitive sampling has not yet been made.
Figure 3: Clear-water environments with strong flow, such as found in the channels and on the barrier reefs support highly diverse octocoral communities, including Nephtheidae such as these Lemnalia. Material was generally hand-collected, fixed and preserved in 75% ethanol. Some dried specimens were obtained. Underwater photographs were taken at most sites. At present, many Indo-Pacific octocoral species are yet to be described, and some of the genera are presently awaiting taxonomic revision. Therefore, many groups were only identified to genus level. However, where an estimate of the numbers of species present in Palau could be made, we have provided such an estimate. These numbers will have to be revised either up or down in the future. Marine habitats sampled included outer barrier reefs including steep drop offs, reef slopes, reef flats and channels, sea grass beds on sand, sandy slopes, lagoonal reefs to the eastern and western sides of Babeldaob, muddy reef flats and slopes near Babeldaob, bays, channels and marine lakes within the Rock Islands, and the northern and southern atolls. Deeper habitats were sampled using mix-gas scuba (to 150 m) and a submersible (to 300 m). Specimens of Octocorallia from the NCI collections are housed in the California Academy of Science (San

82
Francisco), the Museum and Art Gallery of the Northern Territory (Darwin), the U.S. National Museum (Smithsonian) and the Coral Reef Research Foundation (Palau). A reference collection of common genera is maintained at the Palau International Coral Reef Center. Additional collections are present at the Leiden Museum (Naturalis), Leiden, Netherlands.
b) Ecological surveys Systematic octocoral surveys were conducted on 16 shallow reef sites (51 surveys of individual depth zones), using a rapid ecological assessment technique (REA), in September and October 2005. REA is based on ratings of estimated substratum cover in six standard ranked categories. It was initially developed for vegetation analyses (Braun-Blanquet 1964), and further refined for coral reef surveys by Done (1982), Devantier et al. (1998) and Fabricius and De'ath (2001). REA is superior over other survey techniques in terms of area surveyed, time requirements, and a more comprehensive representation of rare and/or heterogeneously distributed taxa (a majority of taxa are rare in highly diverse communities). The method used here is described in detail in (Fabricius and De'ath (2001). In brief, surveys were conducted by scuba diving over a strip typically 200 to 300 m long and 3 m wide, within each of up to 5 depth zones: 18 – 13 m (where available), 13 – 8 m, 8 – 3 m, 3 – 1 m, and reef flat (where available). During and after completion of the survey at each depth zone, the following data were recorded: 1) Relative abundances of genera, visually estimated on
the following rating scale for each depth zone: 0 = absent; 1 = one or few colonies; 2 = uncommon; 3 = common; 4 = abundant; and 5 = dominant.
2) Visual estimate of percent total cover of octocorals and hard corals. Cover was estimated in 2%-increments from 1 – 10%, in 5%-increments from 10 – 30%, and in 10%-increments for >30% cover.
3) Turbidity (visibility at the time of survey, in meters), estimated as the maximum visible horizontal distance of a bright object.
4) Sediment deposits on the substratum, rated on a 4-point scale (0 = none, 1 = thin layer, 2 = considerable amount, but completely resuspendible by fanning, and 3 = thick deep layer of sediment).
5) Flow speed (in cm s-1) during the survey, estimated by timing the traveling distance along a ruler of suspended particles 1 m above the reef substratum.
Ten sites were surveyed in the western lagoon off Babeldaob, near the Ngerdmau river, in duplicates at five stations along a gradient of at increasing distance from the river mouth. Furthermore, two outer barrier reefs (Peliliu-West, “Big Dropoff”), two wave-protect outer barrier reef reef sites in channels (Toachel Mlengui and German
Channel), and two inner Rock Islands sites (an unnamed channel, and “Soft Coral Arc”) were surveyed. Patterns in the octocoral communities were analyzed at the level of sites (i.e., depth-averaging environmental, abundance and cover data, and aggregating data over depths for richness). Lowess smoothers were used to characterize the relationships between biotic and environmental variables to visibility and to distance from the Ngerdmau River. A redundancy analysis, with visibility, sedimentation, flow and distance from river as environmental variables, was used to assess patterns in the octocoral community data (depth-averaged abundance ratings were square-root transformed to reduce heteroscedascity). The statistical software S-Plus was used for all data analyses (Statistical Sciences 1999). Figure 4 (next page): Some of the major reef habitat types along environmental gradients in Palau. The first plate of photographs shows habitats that support rich octocoral communities, while the second plate shows a range of habitat types that are marginal or unsuitable for octocorals. The locations of the various habitat types can be found in the maps of Chapters I-1 and II-2 in this monograph. Photos a to c: Outer barrier reef walls in clear water, with heterotrophic communities dominated by large fans of Annella (a), colonies of Dendronephthya (b) and fans of Melithaeidae (c). Photos d and e: Outer barrier reef slopes dominated by phototrophic nephtheid communities, especially Nephthea (d) and Lemnalia (e). Photos f to h: Flow-swept channel through the outer barrier reef, here with communities of Junceella (f); Clavularia (g) and Clavularia and Dendronephthya (h). Photo i: Flow-swept reef patch in the eastern lagoon off Babeldaob, with communities of Sinularia flexibilis, Sarcophyton, Nephthea and Dendronephthya. Photos j and k: Sheltered and shaded embayment with low flow in the Rock Islands tend to feature high and diverse communies of hard corals (especially Faviidae, Fungiidae, Mussiidae and Poritidae) and sponges, but are void of most octocorals. Photos l to n: Western lagoon patch reef off Babeldaob, with low-diversity communities of Porites and the green calcareous alga Halimeda tend to feature only the alcyoniids Sinularia (l) and Sarcophyton (m and n); Photos o and q: very muddy nearshore reef sites that are exposed to terrestrial runoff from rivers are often void of octocorals, with the exception of few small colonies of white or dark red Chironephthya. Photo p: Reef flats are also unsuitable for many groups of octocorals, however some large colonies of Alcyoniidae (e.g., Sinularia, Lobophytum and Sarcophyton), the blue coral Heliopora and occasionally some Xenia or Heteroxenia can be found here.

83

84

85
3. Results a) The Octocoral Fauna A total of 63 genera of octocorals, belonging to 24 families and 3 orders, were recorded in the shallow waters of the Palau archipelago (Table 1). Thirteen additional genera that are limited to deeper waters (below 60-90 m depth) were also recorded, resulting in a total of 76 genera. Our present estimate is approximately 150 species present in Palau (Table 1). In clear water, arborescent Nephtheidae were the dominant octocoral feature of many upper reef slopes. Most conspicuous and abundant was Lemnalia (Fig. 3) but Nephthea and Stereonephthea were also common on some of the outer slopes. As in many parts of Indo-Pacific, the encrusting or massive Alcyoniidae genera Sinularia, Sarcophyton (Fig. 1) and Lobophytum were also wide-spread throughout the shallow waters and in places quite abundant. The blue coral Heliopora coerulea that forms massive skeletons was found commonly (Fig. 1), often growing in patches where abundances could be high; the size of the largest individual colony observed exceeded 4 m in diameter. Xeniidae, that dominate many Western and Central Indo-Pacific reefs, were rare throughout the area. Some soft corals, such as Solenocaulon and Studeriotes, were restricted to sandy and muddy soft bottom habitats in lagoonal areas where narrowing of reef openings produced moderate currents. Pennatulaceans had low diversity (only 4 species in 3 genera and 3 families) and generally appeared to be relatively rare in Palau; large beds such as found in some other regions, have not yet been located. In current-swept channels and deeper down (below about 45 m), gorgonians dominate the octocoral fauna, with large numbers present along the current swept near-vertical reef faces at 60-100 m depth. The many types of reef habitats in Palau contrasted strongly in their octocoral communities. A few examples are listed here and illustrated in Fig. 4. Steep slopes were occupied by diverse communities of predominantly heterotrophic soft corals and gorgonians, including the conspicuous Stereonephthya, Dendronephthya, Melithaea and Annella (Figs. 4a - c). Well-lit shallow clear-water sites on the outer slopes of the barrier reefs were dominated by Nephtheidae (especially Lemnalia, but also Steronephthya, Nephthea, Dendronephthya, Paralemnalia and Scleronephthya (Figs. 4d - e). Highest octocoral densities and highest diversity were found in flow-exposed channels traversing through the chain of outer barrier reefs, often with high abundances of Junceella and other Ellisellidae, and also some Acanthogorgiidae and Plexauridae (Figs. 4f - h). Lagoonal areas had generally quite poor octocoral communities, with low richness and low cover especially in sheltered areas, but higher densities and richness in flow-swept patches (Fig. 4i, and Figs. 4l – n). Here, the eurytopic alcyoniid genera Sarcophyton and Sinularia,
and also some Nephtheidae and Briareum were encountered. Inner sheltered Rock Island bays with little flow and little water exchange and one of the marine lakes that houses a coral reef (“Heliofungia lake”) were void of any types of octocorals (Figs 4j and k). Similarly, very muddy reefs within a few hundred meters of river mouths had almost no octocorals, with the exception of a few colonies of the highly mud-tolerant azooxanthellate Chironephthya and Carijoa (Figs. 4o - q).
b) Patterns in the distribution of octocorals At the Ngerdmau gradient, both environmental and ecological values strongly changed with increasing distance to the river: visibility almost tripled (from 4 to ~11 m) and the amount of sediment deposited on the ground greatly decreased from the river mouth to sites 2 – 3 km away from the river (Fig. 5). Along the same gradient, octocoral cover greatly increased (from ~0% to ~10%), and hard coral cover tended to decrease (from >20% to <10%). The strongest changes however occurred in octocoral richness: the number of both zooxanthellate and azooxanthellate octocoral genera increased about six-fold along the water quality gradient, from 2 to 12 and 1 to 6 genera per site, respectively. Over all study sites including the outer barrier reefs, octocoral richness averaged 11.1 genera per site (range 2 - 23), with the mean richness of genera with algal endosymbionts being ~30% higher than that of genera without algal endosymbionts: the mean richness of the former genera was 6.3 (range: 0 – 12), versus 4.8 for the latter (range: 1 – 13). Total octocoral richness was highest in the two channels (Toachel Mlengui and German Channel), with 23 and 22 genera per site, respectively. The two outer barrier reef sites were also rich, housing 15 and 18 genera per site. The two inner Rock Island sites yielded a low richness with 3 and 8 genera per site. The redundancy analysis confirmed that structures in octocoral communities strongly changed along gradients in the physical environment. The greatest differences were observed along a gradient of visibility and sedimentation, however flow and distance from river also influenced community structure (Fig. 6). The four environmental variables explained 55.3% of the variation in the community data, with most variation occurring along axis 1 (strongly related to the visibility –sedimentation gradient). Communities of sites belonging to the Ngerdmau gradient were not clearly different to those with similar environmental conditions but located elsewhere. Most species vectors were associated with sites of medium to high visibility and medium to low sedimentation. Chironephthya, Briareum and Sarcophyton were among the few taxa that additionally occurred also at moderately low levels of visibility and sedimentation (Figs. 1, 4 and 6).

86
Figure 5: Changes in visibility, sedimentation, octocoral and hard coral cover, and octocoral richness (mean number of zooxanthellate and azooxanthellate taxa per depth zone) at 10 sites along a gradient of increasing distance away from the mouth of the Ngerdmau River (western Babeldaob). Red lines characterise means (based on lowess smoothers), thin green lines are 90% confidence intervals.
4. Discussion Our inventories and surveys showed that the octocoral fauna of the archipelago of Palau is quite diverse, containing at least 76 genera in 24 families. Species numbers can not be estimated reliably due to the fact that many species still await taxonomic descriptions or revisions, but the number could be in the vicinity of around 150 species. Visually, Nephtheidae were the dominating feature in clear-water habitats, with tall colonies and high abundances in several genera (Fig. 3). As in other Indo-Pacific areas, a diverse range of alcyoniid species belonging to the genera Sinularia spp., Sarcophyton spp. and Lobophytum spp. were also conspicuous in many places. All other octocoral genera were far less commonly encountered, and a majority of genera were classified as rare. At any particular site, the occurrence and abundance of a species is defined by the regional species pool (based on biogeographic distribution ranges), by local and regional environmental conditions, and by the disturbance history. In Palau, the relatively high number of taxa recorded is most certainly related to the proximity of Palau to the ‘coral triangle’, the global centre of biodiversity for hard corals (Veron 1995), fishes (Allen 2000) and most probably octocorals. More systematic data on the octocoral fauna within the coral triangle and surrounding regions are urgently needed to confirm this suggestion and determine relationships. At least 14 shallow-water genera occurring in the coral triangle are not presently known from Palau; these were Anthelia, Cespitularia, Efflatounaria, Sansibia, Sympodium, Pacifiphyton, Alertigorgia, Iciligorgia, Isis, Ifalukella, Plumigorgia, Ctenocella, Pseudopterogorgia, and Plumigorgia - although some doubtful visual records exist for some of these in earlier literature. This would equal a reduction in diversity of about 15% compared to the generic richness of ~86 shallow-water genera expected in the Coral Triangle. Sampling intensity for octocorals has been high in Palau compared with many other Indo-Pacific regions, especially due to numerous collections conducted to investigate marine natural products. Faulkner et al. (2004) have summarized such collections from Palau and provide references to published papers describing new compounds from octocorals from Palau. Some of the genera not yet recorded may eventually be found in Palau, but we are reasonably certain that most of them appear to have their distribution limits located to the west and south of Palau. In contrast, it is likely that more taxa will be eventually described from reefs and soft bottom areas in the far less intensely studied Coral Triangle. To the east of Palau, the octocoral fauna of Guam, the only other area of Micronesia which has been thoroughly sampled, is more reduced than Palau. Paulay et al. (2003) found only 79 species of octocorals in 32 genera from Guam (methods comparable to our surveys in Palau), and
0.0 1.0 2.0 3.0
0
5
10
0.0 1.0 2.0 3.0
% O
ctoc
oral
Cov
er
0
10
20
30
40
% H
ard
cora
l Cov
er
0.0 1.0 2.0 3.0
4
6
8
10
12
Visi
bilit
y (m
)R
ichn
ess
(zoo
x. g
ener
a/si
te)
Ric
hnes
s (a
zoox
. gen
era/
site
)
Distance to river (km)
0.0 1.0 2.0 3.0 0.0 1.0 2.0 3.0
1.0
1.5
2.0
2.5
3.0
Sedi
men
tatio
n (ra
ting)
0.0 1.0 2.0 3.0
0
2
4
6
8
10
12
1
2
3
4
5
6
0.0 1.0 2.0 3.0
0
5
10
0.0 1.0 2.0 3.0
% O
ctoc
oral
Cov
er
0
10
20
30
40
% H
ard
cora
l Cov
er
0.0 1.0 2.0 3.00.0 1.0 2.0 3.0
4
6
8
10
12
Visi
bilit
y (m
)R
ichn
ess
(zoo
x. g
ener
a/si
te)
Ric
hnes
s (a
zoox
. gen
era/
site
)
Distance to river (km)
0.0 1.0 2.0 3.0 0.0 1.0 2.0 3.0
1.0
1.5
2.0
2.5
3.0
Sedi
men
tatio
n (ra
ting)
0.0 1.0 2.0 3.00.0 1.0 2.0 3.0
0
2
4
6
8
10
12
1
2
3
4
5
6

87
Figure 6: Redundancy analysis biplot of octocoral communities on 16 reef sites in Palau. Superimposed onto the plot are the environmental variables visibility, distance to river, sedimentation, and flow. The groups surrounded by convex hulls characterises the reefs by visibility at the time of surveys. Square symbols mark sites from the Ngerdmau gradient, round symbols mark sites from other locations. The first 5 to 7 letters of the genera (listed in Table 1) were used as abbreviations; only names of the more abundant genera are printed for clarity. Most variation in the community data occurred along axis 1, which is strongly related to the visibility – sedimentation gradient but also influenced by water flow. The 4 environmental variables together explain 55.3% of the variation in the community data.
nepht
stere
scler
dendr
lemna
parale
sipho
chiro
sinul
sarco
lobop
klyxu
parami
helioclavu
carij
briar
suberg
melit
acant
euplx
echgomenel
rumph
ellis
junce
4 - 88 - 11
11 - 40
Visibility [m]:
Dim 1 66.12% : [ 0.955 ]
Dim
2
24.2
1% :
[ 0.9
41 ]
%Variance explained by x-variables = 55.3%
Distance to River
Visibility
Flow
Sediment
nepht
stere
scler
dendr
lemna
parale
sipho
chiro
sinul
sarco
lobop
klyxu
parami
helioclavu
carij
briar
suberg
melit
acant
euplx
echgomenel
rumph
ellis
junce
4 - 88 - 11
11 - 40
Visibility [m]:
Dim 1 66.12% : [ 0.955 ]
Dim
2
24.2
1% :
[ 0.9
41 ]
%Variance explained by x-variables = 55.3%
Distance to River
Visibility
Flow
Sediment

88
Pennatulaceans are unknown from that island. The limited suite of habitats in Guam, its distance from more diverse western and southern sources of additional species and a paucity of information on an apparently rich deeper water octocoral fauna may account for genus numbers just over one half of those recorded from Palau. Xeniidae were only represented by 3 genera, Xenia, Heteroxenia and Asterospicularia, with the former two having their eastern distribution limit in Palau and the latter in Guam (Paulay et al. 2003). These Xeniidae were all recorded as rare or uncommon during the taxonomic inventories in shallow waters. The xeniid genera Cespitularia, Efflatounaria, Anthelia, Sansibia and Sympodium that are common on many clear-water reefs of the Indo-Pacific, were not encountered in the surveys; these genera appear to be beyond the eastern margin of their distribution in Palau. Xeniidae appear very poor in dispersal over longer distances, probably because their larvae are large and after developing on the surface of colonies, they drift or crawl a few meters to a few kilometers away from the parental colony, restricting dispersal to short distances. Once established, some Xeniidae (especially Heteroxenia and Xenia) have been observed to rapidly establish extensive patches of colonies through asexual reproduction or the settlement of brooded larvae near the parent colony (Benayahu and Loya 1985), but such colonization is restricted to areas nearby. These life history data indicate that such brooding taxa do not easily disperse over longer distances. It is interesting to note that Xeniidae have been found to be particularly sensitive to bleaching mortality from high water temperatures (Fabricius 1999). For the Xeniidae, the existence of a very small gene pool in Palau, combined with high bleaching mortality rates and poor larval dispersal, suggests that some members of this family are at particular risk of being lost from the Palauan reefs if sea surface temperatures continue to rise from global warming. Palau is renown for its marine habitat diversity, which includes outer barrier reef walls with steep drop-offs and oceanic water clarity, channels, sheltered and often shaded fringing reefs around the calcareous Rock Islands, and lagoonal inshore patch reefs and estuarine coral reefs off the volcanic Babeldaob Island (Fig. 4). Clear-water channels and outer barrier reefs contained highly diverse octocoral communities including, among many others, high abundances of Nephtheidae, as well as large colonies of genera in Subergorgiidae, Ellisellidae, and Gorgoniidae. Gorgonians, principally members of Muricella, Echinogorgia and Annella, some of which are home to small "pygmy" seahorses (e.g., Hippocampus denise on Muricella, Lourie and Randall 2003), occur predominantly on outer reef slopes. In contrast, lagoonal (both Rock Islands and Babeldaob) and estuarine habitats contained small subsets of the genera found in the clear-water habitats, i.e., these environments did not create
habitats for many octocoral genera that were not found elsewhere. No genus was preferentially associated with low-visibility areas on the runoff-exposed coral reefs near rivers. This suggests that for octocorals, communities are taxonomically and numerically dominated by clear-water reefs, and depleted towards the disturbed and muddy estuarine reefs and sheltered sites. As sheltered and estuarine areas did not provide suitable habitat for many new octocoral species, the existence of high habitat diversity in Palau did not contribute to explain the relatively high diversity in octocorals. For many species of hard corals, the reefs around the sheltered Rock Islands represented a refuge during the 1998 mass bleaching event, but as the Rock Islands are unsuitable habitats for most octocorals, such refuge does not exist for this group. Of course, lagoonal and estuarine areas are important habitats for other groups of organisms including seagrasses, macroalgae and probably also a range of other invertebrates and vertebrates. The ecological surveys showed that octocoral richness was more strongly related to water clarity and amounts of sediments deposited than were total hard coral and octocoral cover, which were poorly explained by these variables. Changes in taxonomic richness of octocorals have previously been suggested to be suitable as indicators of reef “quality” and disturbance by poor water quality on the Great Barrier Reef and in Hong Kong (Fabricius and De'ath 2001, Fabricius and McCorry 2006). On the Great Barrier Reef, at visibility <10 m the richness of zooxanthellate octocorals declined by one genus for each meter visibility lost in otherwise comparable locations; richness of azooxanthellate taxa was unrelated to visibility (Fabricius and De'ath 2001). In Hong Kong, no zooxanthellate taxa are found in the most polluted and river-influenced subtropical conditions, whereas several zooxanthellate taxa are found in the region least influenced by the river; richness in azooxanthellate taxa is again unrelated to water quality (Fabricius and McCorry 2006). Interestingly, in Palau, declines in richness along the water quality gradient and with visibility were stronger than shown in the previous studies, suggesting that the extent of disturbance by river runoff is severe close to the river mouth but spatially limited to sites within a few kilometers from the river mouth. Interestingly, in Palau, the richness of both zooxanthellate and azooxanthellate genera decreased at similar rates with increasing turbidity and sedimentation. The reasons for the decline in azooxanthellate taxa in turbid and riverine regions deserve further study. Despite a number of unanswered questions, our study suggests that in Palau, ecological monitoring programs that discriminate taxa at least to genus level will better indicate disturbances by terrestrial runoff than records that are restricted to total coral cover.

89
Acknowledgments
The marine collection contracts (1992-2006) of the US National Cancer Institute Natural Products Research Branch to the Coral Reef Research Foundation (CRRF) has supported most recent collections of soft corals from Palau (deposited at the California Academy of Sciences, Museum and Art Gallery of the Northern Territory, and Coral Reef Research Foundation) and their taxonomic determination. Staff members of CRRF, including L. Bell, M. Mesubed, E. Basilius, L. Martin, M. Dawson and L. Sharron, have placed special emphasis on obtaining specimens and their efforts are much appreciated. Primnoid and other deepwater gorgonians were identified by Dr. Steve Cairns, USNM, while Dr. Leen van Ofwegen identified most Nephtheidae. Many thanks also to Lori Colins for comments on the manuscript, and to the Palau International Coral Reef Center for providing support, facilities and field access for this study. Further support was provided by the Australian Institute of Marine Science.
References
Alderslade P (2001) Six new genera and six new species of soft coral, and some proposed familial and subfamilial changes within the Alcyonacea (Coelenterata: Octocorallia). Bulletin of the Biological Society of Washington 10: 15-65
Benayahu Y (1995) Species composition of soft corals (Octocorallia, Alcyonacea) on the coral reefs of Sesoko Island, Ryukyu Archipelago, Japan. Galaxea 12: 103-124
Benayahu Y, Jeng M-S, Perkol-Finkel S, Dai C-F (2004) Soft Corals (Octocorallia: Alcyonacea) from Southern Taiwan. II. Species Diversity and Distributional Patterns. Zoological Studies 43: 548-560
Benayahu Y, Loya Y (1981) Competition for space among coral-reef sessile organisms at Eilat, Red Sea. Bulletin of Marine Science 31: 514-522
Benayahu Y, Loya Y (1985) Settlement and recruitment of a soft coral: Why is Xenia macrospiculata a successful colonizer? Bulletin of Marine Science 36: 177-188
Benayahu Y, Yosief T, Schleyer M (2002) Soft corals (Octocorallia, Alcyonacea) of the southern Red Sea. Israel Journal of Zoology 48: 273-283
Braun-Blanquet JJ (1964) Pflanzensoziologie, Grundzüge der Vegetationskunde. 3rd Edition. Springer Press, Vienna, New York
Devantier LM, De'ath G, Done TJ, Turak E (1998) Ecological assessment of a complex natural system: A case study from the Great Barrier Reef. Ecological Applications 8: 480-496
Dinesen ZD (1983) Patterns in the distribution of soft corals across the central Great Barrier Reef. Coral Reefs 1: 229-236
Done TJ (1982) Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1: 95-107
Fabricius K (1999) Tissue loss and mortality in soft corals following mass-bleaching. Coral Reefs 18: 54
Fabricius K, Alderslade P (2001) Soft Corals and Sea Fans: A comprehensive guide to the tropical shallow water genera of the central-west Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville (264)
Fabricius K, De'ath G (1997) The effects of flow, depth and slope on cover of soft coral taxa and growth forms on Davies Reef, Great Barrier Reef. 8th International Coral Reef Symposium 2: 1071-1076.
Fabricius K, McCorry D (2006) Changes in octocoral communities and benthic cover along a water quality gradient in the reefs of Hong Kong. Marine Pollution Bulletin: 22-33
Fabricius KE (1997) Soft coral abundance on the central Great Barrier Reef: Effects of Acanthaster planci, space availability, and aspects of the physical environment. Coral Reefs 16: 159-167
Fabricius KE, De'ath G (2001) Biodiversity on the Great Barrier Reef: Large-scale patterns and turbidity-related local loss of soft coral taxa. In: Wolanski E (ed.) Oceanographic processes of coral reefs: physical and biological links in the Great Barrier Reef. CRC Press, London (pp 127 - 144)
Faulkner DJ, Newman DJ, Cragg GM (2004) Investigations of the marine flora and fauna of the Islands of Palau. Natural Products Rep. 21: 50-76
Grasshoff M (1999) The shallow water gorgonians of New Caledonia and adjacent islands (Coelenterata, Octocorallia). Senckenbergiana Biologica 78: 1-121
Lourie SA, Randall JE (2003) A new pygmy seahorse, Hippocampus denise (Teleostei: Syngnathidae) from the Indo-Pacific. Zoological Studies 42: 284-291
Paulay G, Puglisi MP, Starmer JA (2003) The non-scleractinian Anthozoa (Cnidaria) of the Mariana Islands. Micronesica 35-36: 138-155
Ricklefs RE (2004) A comprehensive framework for global patterns in biodiversity. Ecology Letters 7: 1-15
Tursch B, Tursch A (1982) The soft coral community on a sheltered reef quadrat at Laing Island (Papua New Guinea). Marine Biology 68: 321-332
van Oppen MJH, Mieog JC, Sánchez CA, Fabricius KE (2005) Diversity of algal endosymbionts (zooxanthellae) in octocorals: the roles of geography and host relationships. Molecular Ecology 14: 2403-2417
van Ofwegen LP (2005) A new genus of nephtheid soft corals (Octocorallia: Alcyonacea: Nephtheidae) from the Indo-Pacific. Zoologische Mededelingen 79-4: 3-236
Veron J (2000) Corals of the World. Australian Institute of Marine Science, Townsville
Verseveldt J (1977) Australian Octocorallia (Coelenterata). Aust. J. Mar. Freshwater Res 28: 171-240
Williams G (1992) Biogeography of the octocorallian coelenterate fauna of southern Africa. Biological Journal of the Linnean Society 46: 351-401
Williams GC (1995) Living genera of sea pens (Coelenterata: Pennatulacea): illustrated key and synopses. Zoological Journal of the Linnean Society 113: 93-140
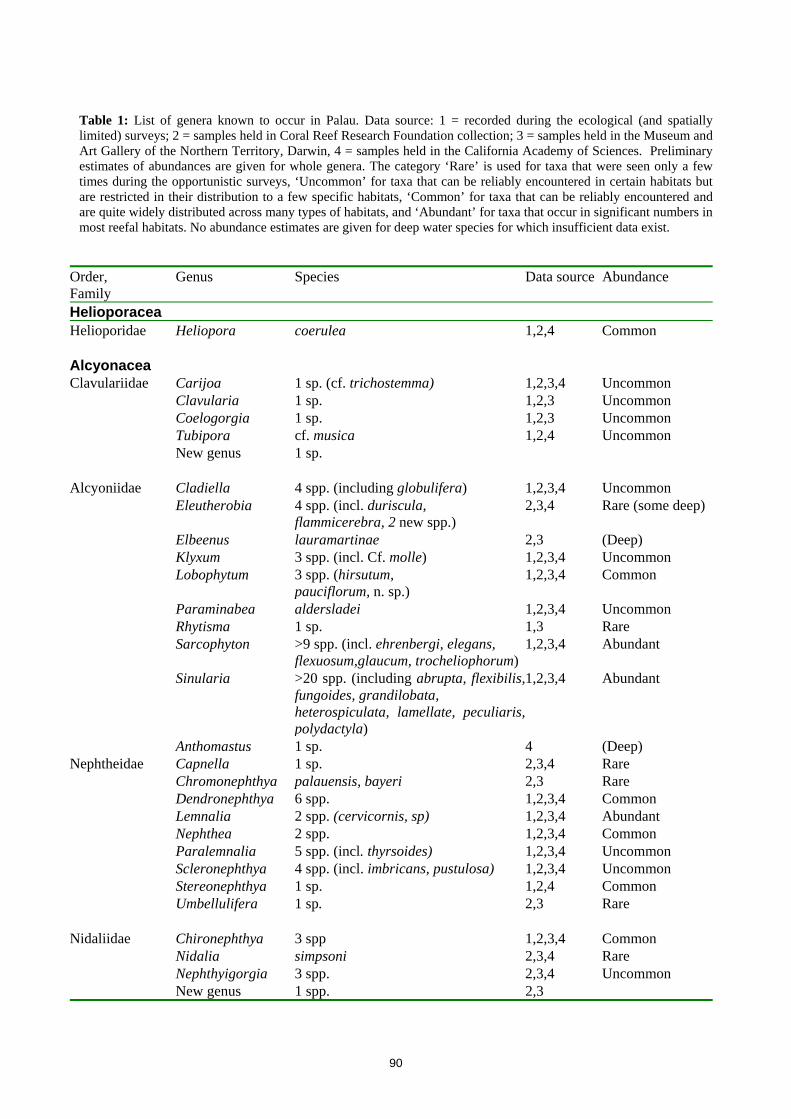
90
Table 1: List of genera known to occur in Palau. Data source: 1 = recorded during the ecological (and spatially limited) surveys; 2 = samples held in Coral Reef Research Foundation collection; 3 = samples held in the Museum and Art Gallery of the Northern Territory, Darwin, 4 = samples held in the California Academy of Sciences. Preliminary estimates of abundances are given for whole genera. The category ‘Rare’ is used for taxa that were seen only a few times during the opportunistic surveys, ‘Uncommon’ for taxa that can be reliably encountered in certain habitats but are restricted in their distribution to a few specific habitats, ‘Common’ for taxa that can be reliably encountered and are quite widely distributed across many types of habitats, and ‘Abundant’ for taxa that occur in significant numbers in most reefal habitats. No abundance estimates are given for deep water species for which insufficient data exist.
Order, Family
Genus Species Data source Abundance
Helioporacea Helioporidae Heliopora coerulea 1,2,4 Common Alcyonacea Clavulariidae Carijoa 1 sp. (cf. trichostemma) 1,2,3,4 Uncommon Clavularia 1 sp. 1,2,3 Uncommon Coelogorgia 1 sp. 1,2,3 Uncommon Tubipora cf. musica 1,2,4 Uncommon New genus 1 sp. Alcyoniidae Cladiella 4 spp. (including globulifera) 1,2,3,4 Uncommon Eleutherobia 4 spp. (incl. duriscula,
flammicerebra, 2 new spp.) 2,3,4 Rare (some deep)
Elbeenus lauramartinae 2,3 (Deep) Klyxum 3 spp. (incl. Cf. molle) 1,2,3,4 Uncommon Lobophytum 3 spp. (hirsutum,
pauciflorum, n. sp.) 1,2,3,4 Common
Paraminabea aldersladei 1,2,3,4 Uncommon Rhytisma 1 sp. 1,3 Rare Sarcophyton >9 spp. (incl. ehrenbergi, elegans,
flexuosum,glaucum, trocheliophorum)1,2,3,4 Abundant
Sinularia >20 spp. (including abrupta, flexibilis,fungoides, grandilobata, heterospiculata, lamellate, peculiaris, polydactyla)
1,2,3,4 Abundant
Anthomastus 1 sp. 4 (Deep) Nephtheidae Capnella 1 sp. 2,3,4 Rare Chromonephthya palauensis, bayeri 2,3 Rare Dendronephthya 6 spp. 1,2,3,4 Common Lemnalia 2 spp. (cervicornis, sp) 1,2,3,4 Abundant Nephthea 2 spp. 1,2,3,4 Common Paralemnalia 5 spp. (incl. thyrsoides) 1,2,3,4 Uncommon Scleronephthya 4 spp. (incl. imbricans, pustulosa) 1,2,3,4 Uncommon Stereonephthya 1 sp. 1,2,4 Common Umbellulifera 1 sp. 2,3 Rare Nidaliidae Chironephthya 3 spp 1,2,3,4 Common Nidalia simpsoni 2,3,4 Rare Nephthyigorgia 3 spp. 2,3,4 Uncommon New genus 1 spp. 2,3
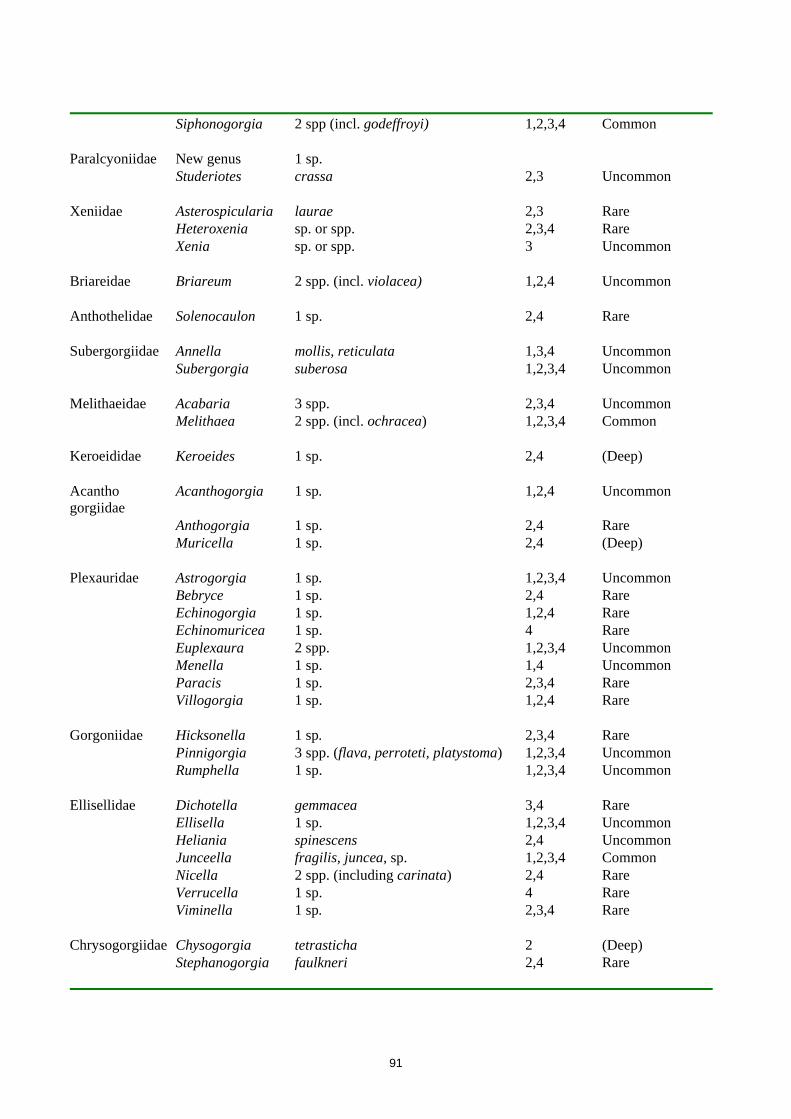
91
Siphonogorgia 2 spp (incl. godeffroyi) 1,2,3,4 Common Paralcyoniidae New genus 1 sp. Studeriotes crassa 2,3 Uncommon Xeniidae Asterospicularia laurae 2,3 Rare Heteroxenia sp. or spp. 2,3,4 Rare Xenia sp. or spp. 3 Uncommon Briareidae Briareum 2 spp. (incl. violacea) 1,2,4 Uncommon Anthothelidae Solenocaulon 1 sp. 2,4 Rare Subergorgiidae Annella mollis, reticulata 1,3,4 Uncommon Subergorgia suberosa 1,2,3,4 Uncommon Melithaeidae Acabaria 3 spp. 2,3,4 Uncommon Melithaea 2 spp. (incl. ochracea) 1,2,3,4 Common Keroeididae Keroeides 1 sp. 2,4 (Deep) Acantho gorgiidae
Acanthogorgia 1 sp. 1,2,4 Uncommon
Anthogorgia 1 sp. 2,4 Rare Muricella 1 sp. 2,4 (Deep) Plexauridae Astrogorgia 1 sp. 1,2,3,4 Uncommon Bebryce 1 sp. 2,4 Rare Echinogorgia 1 sp. 1,2,4 Rare Echinomuricea 1 sp. 4 Rare Euplexaura 2 spp. 1,2,3,4 Uncommon Menella 1 sp. 1,4 Uncommon Paracis 1 sp. 2,3,4 Rare Villogorgia 1 sp. 1,2,4 Rare Gorgoniidae Hicksonella 1 sp. 2,3,4 Rare Pinnigorgia 3 spp. (flava, perroteti, platystoma) 1,2,3,4 Uncommon Rumphella 1 sp. 1,2,3,4 Uncommon Ellisellidae Dichotella gemmacea 3,4 Rare Ellisella 1 sp. 1,2,3,4 Uncommon Heliania spinescens 2,4 Uncommon Junceella fragilis, juncea, sp. 1,2,3,4 Common Nicella 2 spp. (including carinata) 2,4 Rare Verrucella 1 sp. 4 Rare Viminella 1 sp. 2,3,4 Rare Chrysogorgiidae Chysogorgia tetrasticha 2 (Deep) Stephanogorgia faulkneri 2,4 Rare

92
Isididae Lepidisis 2 spp. 2,4 (Deep) Primnoidae Callogorgia elegans, versluysi, formosa 2,5 (Deep) Calyptrophora 1 sp. cf. japonica 5,5 (Deep) Narella leilae, megalepis 5,5 (Deep) Pennatulacea Veretillidae Cavernulina cylindrica 2,4 Uncommon Veretillum 1 sp. 2 (Deep) Funiculinidae Funiculina quadrangularis 2,4 (Deep) Anthoptilidae Anthoptilum grandiflorum 2,4 (Deep) Virgulariidae Virgularia gustaviana 2,4 Uncommon Pennatulidae Pennatula murrayi and n.sp. 2,4 (Deep) Pteroeides 1 or 2 spp. 2,4 Uncommon 24 Families 76 Genera approximately 150 species