a common pathway mediated through toll-like receptors - blood
TRANSCRIPT

IMMUNOBIOLOGY
A common pathway mediated through Toll-like receptors leads to T- and naturalkiller–cell immunosuppressionIlan Vaknin,1 Liora Blinder,1 Lynn Wang,1 Roi Gazit,1 Elena Shapira,1 Olga Genina,2 Mark Pines,2 Eli Pikarsky,3 andMichal Baniyash1
1Lautenberg Center for General and Tumor Immunology, Hebrew University Hadassah Medical School, Jerusalem; 2Institute of Animal Science, AgriculturalResearch Organization, Volcani Center; and 3Department of Pathology, Hebrew University Hadassah Medical School, Ein Kerem Campus, Kiryat Hadassah,Jerusalem, Israel
T- and natural killer (NK)–cell immunosup-pression associated with �-chain down-regulation has been described in cancer,autoimmune, and infectious diseases.However, the precise stimuli leading tothis bystander phenomenon in such differ-ent pathogen-dependent and sterile pa-thologies remained unresolved. Here, wedemonstrate that Toll-like receptors(TLRs) play a major role in the inductionof innate and adaptive immune systemsuppression; repetitive administration of
single TLR 2, 3, 4, or 9 agonists, which donot exhibit any virulent or immune inva-sive properties, was sufficient to induce abystander NK- and T-cell immunosuppres-sion associated with �-chain down-regulation mediated by myeloid suppres-sor cells, as observed in the course ofactive pathologies. We identified a 35-amino acid (aa) region within the �-chainas being responsible for its degradationunder TLR-mediated chronic inflamma-tion. Furthermore, we provide evidence
that �-chain levels could serve as a bi-omarker for chronic inflammation-dependent immunosuppression. Thus,although acute TLR-mediated activationcould be beneficial in clearing pathogensor may serve as an immune adjuvant,such activation could be detrimental un-der sustained conditions. (Blood. 2008;111:1437-1447)
© 2008 by The American Society of Hematology
Introduction
During a sustained response to inactivated pathogens, and in thecourse of cancer, infections, or autoimmune diseases, T and naturalkiller (NK) cells can become �-chain deficient and immunologi-cally nonfunctional (reviewed in1). In our previous studies, wesuggested that the causative link between these different patholo-gies is chronic inflammation and the ensuing accumulation ofmyeloid suppressor cells (MSCs), which lead to decreased �-chainexpression and impaired T-cell function.2,3 However, several criti-cal questions remain unresolved such as how is a similar immuno-suppressed state induced during sterile and pathogen-dependentdiseases, and what is the nature of the stimuli leading to thegeneration of massive inflammation resulting in such generalizedimmunosuppression?
We raised the hypothesis that in the course of the above-mentioned pathologies, chronic stimulation of the innate immunesystem via Toll-like receptors (TLRs) would lead to the inductionof immunosuppression. TLRs recognize conserved pathogen-associated molecular patterns, which serve as TLR agonists/ligands(TLRLs).4-9 Moreover, recent studies suggest that endogenous,host-derived components, including fibrinogen, heat shock pro-teins, �-defensins, RNA, and DNA could also serve as TLRLs.5,10
TLRs are primarily expressed on cells of the innate immune system(myeloid and NK cells) and by some cells of the adaptive immunesystem (regulatory and activated T cells). Upon activation of any ofthe TLR signaling pathways, a primarily Th1-inflammatory re-sponse ensues, as reflected by the production of proinflammatorycytokines, chemokines, nitric oxide (NO), and other factors.1,11
Thus, TLRs could be activated during infections as well as in the courseof sterile pathologies. It is well established that TLR-mediate acuteactivation is beneficial to the host because it leads to a critical responseresponsible for the clearance of pathogens and abnormal or damagedself-tissues. However, based on our observations, when TLR-mediatedstimulation is sustained and chronic inflammation ensues, the appear-ance of immunosuppression is expected.
In the current study, we tested whether chronic exposure to a singleTLRL, which does not exhibit any virulent or immune invasiveproperties, could induce immunologic nonresponsiveness characterizedby reduced �-chain expression. To this end, we developed an in vivoexperimental model system in which normal mice were repeatedlyexposed to a single TLRL (specific for TLRs 2, 3, 4, or 9), encapsulatedin multilamellar vesicles (MLVs). The effect of this treatment on theadaptive (T cell) and innate (NK cell) immune responses was assessed inconjunction with testing the expression levels of the �-chain. The�-chain, in both cell types, plays a crucial role in receptor expression andsignaling function,12-14 and its expression is affected by the immunosup-pressive conditions.
We demonstrate that chronic exposure to a single TLRL specific forTLR 2, 3, 4, or 9 is sufficient to induce T- and NK-cell immunosuppres-sion mediated by MSCs, which is associated with �-chain degradation.A 35-amino acid (aa) region within the �-chain was identified as beingresponsible for its targeting to degradation under TLR-mediated chronicinflammation. Furthermore, because �-chain expression levelsdiscriminate between acute and chronic inflammation character-ized by immunosuppression, we suggest that reduced �-chain
Submitted July 10, 2007; accepted November 5, 2007. Prepublished onlineas Blood First Edition paper, November 8, 2007; DOI 10.1182/blood-2007-07-100404.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2008 by The American Society of Hematology
1437BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom

expression levels could serve as a biomarker for chronicinflammation-induced immunosuppression.
Methods
Mice
Female BALB/c, C3H/HeN, C3H/HeJ, B10.A, and C57BL/6 mice, 6 to8 weeks of age, were bred at the Hebrew University specific pathogen-freefacility. Transgenic mice encoding full-length (FL) �-chain and �-chainvariants containing only a single distal immunoreceptor tyrosine-basedactivation motif (ITAM; distal) or no ITAMs (tail-less [TL]) were generatedon the background of ��/� as described previously.15,16 Animal use followedprotocols approved by the Hebrew University-Hadassah Medical SchoolInstitutional Animal Care and Use Committee.
Antigen, TLRLs, and immunization
Antigens and immune modulators were obtained as follows: ovalbumingrade 3, Zymosan A, poly(I:C), lipopolysaccharide (LPS) from Escherichiacoli strain 055:B5 (cat.L-4524), all purchased from Sigma-Aldrich (St Louis,MO). Endotoxin-free ( � 1 ng/mg DNA) phosphorothioate CpG-ODNs(5�-TGA CTGTGAACGTTCGAGATGA-3�), and the mutant (m)CpG-ODNs (5�-TGACTGTGAAGG TTAGAGATGA-3�) were kindly providedby Dr Eyal Raz from the University of California, San Diego. Mice wereinjected subcutaneously 3 times with specific TLR ligands (LPS [70 �g],Zymosan [200 �g], CpG-ODNs [70 �g], and Poly (I:C) [150 �g])encapsulated in MLVs as described previously.17
Immunostaining and fluorescence-activated cell sorteranalysis
Antibodies used for cell surface labeling were purchased from BDPharMingen (San Diego, CA) and included fluorescein isothiocyanate(FITC)–labeled anti-Thy-1.2, anti-Gr-1, and anti-bromodeoxyuridine (BrdU),phycoerythrin-labeled anti-CD4, anti-CD3, and anti-NK1.1. Biotinylatedanti-CD3ε, anti–T-cell receptor (TCR)��, anti-Thy-1.2, and anti-Mac-1(CD11b) were detected by streptavidin-Cy5 (Jackson Immunoresearch).Cells were precoated with anti-mouse CD16/CD32 (BD PharMingen),incubated for 30 minutes at 4°C with the specific labeled antibodies,washed, and then incubated with a second-step reagent. Intracellularstaining of � and CD3ε-chains was performed as described previously.2
Samples were analyzed in a FACSCalibur apparatus using Cell Questsoftware (BD PharMingen).
6-Carboxyfluorescein succinimidyl ester staining and ex vivoproliferation assay
Splenocytes (20 � 106/mL) were incubated in phosphate-buffered saline(PBS) without Ca2�/Mg2� containing 5 �M 6-carboxyfluorescein succin-imidyl ester (CFSE; Molecular Probes) for 8 minutes at 25°C. At the end ofthis incubation, fetal calf serum (FCS) was added and the cells were washedin RPMI/8% FCS serum. CFSE-labeled splenocytes were activated withanti-CD3ε and anti-CD28 or phorbol-12-myristate-13-acetate (PMA) andCa2� ionophore, and the number of cell divisions of Thy1.2� cells wasdetermined by fluorescence activated cell sorter (FACS).
Intracellular BrdU staining
Splenocytes were activated as described in the previous section. They werethen surface labeled with antiThy1.2, fixed as described previously,2 andstained with FITC-conjugated anti-BrdU antibodies. The cells were washedand analyzed by FACS. Nonpulsed activated cells were used as a control forthe anti-BrdU staining.
Cell isolation and separation
For magnetic column separation, splenocytes were first labeled (30 minutesat 4°C) with biotin-conjugated antibodies. Negative selection using anti-
CD11b and anti-B220 antibodies was performed for T-cell isolation( 90% purity), and positive selection was performed for the separation ofGr-1�Mac-1� cells using anti-Gr-1 antibodies ( 98% purity). The cellswere then washed, labeled (45 minutes at 4°C) with anti-biotin antibodiesconjugated to magnetic microbeads (Miltenyi Biotec, Auburn, CA), washed,and loaded onto a column placed in a magnetic field (Miltenyi Biotec).
Coincubation experiments
Hybridoma T cells or T cells isolated from the spleen were suspended incomplete growing medium and coincubated for 16 hours at 37°C with aGr1�Mac-1�–enriched cell population obtained from the spleen of LPS-treated mice. The cells were then harvested and the level of �-chainexpression within the T-cell population was determined by FACS.
Cytotoxicity assays
For the in vitro assay, splenocytes were harvested from control orLPS-treated mice 18 hours on intraperitoneal injection of 200 �g poly(I:C)or PBS, and their killing activity was examined against target cells (YAC-1)that were pre-labeled with [35S]methionine. Various effector/target ratioswere used in a cytotoxicity assay of 5-hour incubation at 37°C. Specifickilling was calculated as described18 and normalized according to NK1.1�
cell percentage. For the in vivo assay, a published fluorescence labelingmethod19 was used. Briefly, splenocytes from BALB/C and C57BL/6 micewere stained with CFSE (Invitrogen, Carlsbad, CA) at a final concentrationof 0.5 �M (CFSElow) and 5 �M (CFSEhigh), respectively. Cells (5 � 106) ofeach type were mixed and injected intravenously into recipient C57BL/6mice (control, LPS-treated, and NK1.1-depleted mice). Peripheral bloodlymphocytes (PBLs), lymph node, and spleen cells were harvested 18 hoursafter injection, and the ratio between the CFSEhigh and CFSElow populationswas determined by FACS. The index of specific allogeneic cell clearancewas calculated according to the formula: Index of specific allogeneic cellclearance 1� BALB/C splenocytes (CFSElow) cells of total CFSE�
cells/ B6 splenocytes (CFSEhigh) cells of total CFSE� cells.
Influenza virus infection and mice survival
Mice were infected by intranasal inoculation with the LD50 dose ofinfluenza virus (A/PR/8/34) 2 days before the last LPS injection and micesurvival was followed. LPS treatment itself did not affect mice survival.
Spleen immunohistochemistry
Five-micrometer, formalin-fixed, and paraffin-embedded sections from thespleens of control and LPS-treated mice were deparaffinized and rehy-drated. After endogenous peroxidase quenching (H2O2), antigens wereretrieved by boiling the sections in 10 mM citrate buffer (pH 6.0) or afterpronase digestion. Immunostaining was then performed using the specificanti-Gr1 (clone RB6-8C5) or anti-CD3 antibodies, and the appropriatesecond reagents were applied (Zymed Laboratories, South San Francisco,CA) to visualize the labeled cells. Images of immunohistochemical stainingwere viewed with a Zeiss Axioplan 2 microscope and acquired with a ZeissAxioCam digital camera (Zeiss, Oberkochen, Germany), using a 10�/0.30NA Plan neofluar (Figures 4D,5Ci-iv) or a 20�/0.50 NA Plan neofluar(Figure 5Cv-vi) objective lens. Digitized images were processed usingAdobe Photoshop 7 image processing and manipulation software (AdobeSystems, San Jose, CA).
Measurement of serum cytokines
Serum was obtained from control and LPS-treated (1 day after the third LPSinjection) mice. Interleukin-4 (IL-4), IL-6, interferon-� (IFN-�), and tumornecrosis factor (TNF) concentrations in the serum were measured byenzyme linked immunosorbent assay (ELISA) according to manufacturerinstructions (BD Biosciences PharMingen, San Diego, CA).
Constructs and transfection
FL and proximal �-chain constructs were generated by polymerase chainreaction and cloned into the expression vector pcDNA3.1. �-Deficient
1438 VAKNIN et al BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom
T-cell hybridoma MA5.8 cells16 were stably transfected with the FL andproximal �-chain constructs using the Bio-Rad Gene Pulser (Bio-Rad,Hercules, CA) at a setting of 250 V and 950 microfarad with 10 �g ofplasmid/1 � 107 cells. G418 resistant clones were screened for �-chainexpression by FACS and Western blot analysis.
Results
Repeated LPS treatment induces TLR4-dependent impairmentof T-cell function associated with �-chain down-regulation
We first analyzed the effect of sustained in vivo TLR4 activationon the adaptive immune system (T cells). This TLR is com-monly activated because of its ability to recognize bacterialcompounds, such as LPS, as well as endogenous proteins that
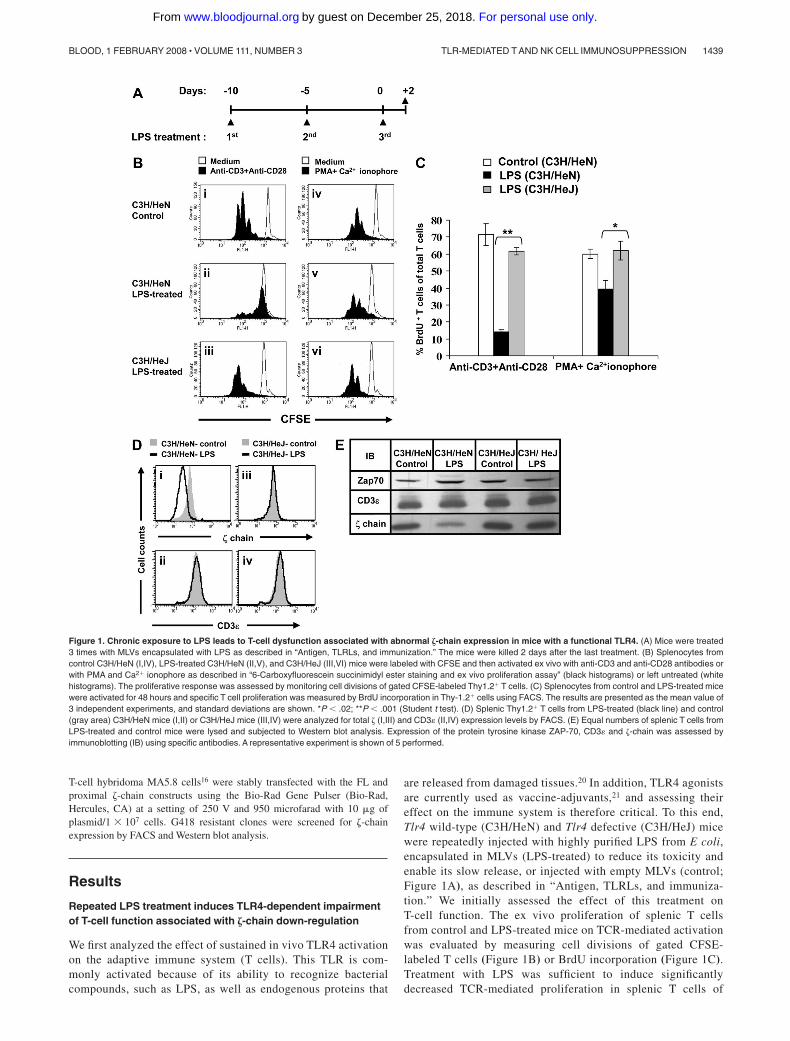
are released from damaged tissues.20 In addition, TLR4 agonistsare currently used as vaccine-adjuvants,21 and assessing theireffect on the immune system is therefore critical. To this end,Tlr4 wild-type (C3H/HeN) and Tlr4 defective (C3H/HeJ) micewere repeatedly injected with highly purified LPS from E coli,encapsulated in MLVs (LPS-treated) to reduce its toxicity andenable its slow release, or injected with empty MLVs (control;Figure 1A), as described in “Antigen, TLRLs, and immuniza-tion.” We initially assessed the effect of this treatment onT-cell function. The ex vivo proliferation of splenic T cellsfrom control and LPS-treated mice on TCR-mediated activationwas evaluated by measuring cell divisions of gated CFSE-labeled T cells (Figure 1B) or BrdU incorporation (Figure 1C).Treatment with LPS was sufficient to induce significantlydecreased TCR-mediated proliferation in splenic T cells of
Figure 1. Chronic exposure to LPS leads to T-cell dysfunction associated with abnormal �-chain expression in mice with a functional TLR4. (A) Mice were treated3 times with MLVs encapsulated with LPS as described in “Antigen, TLRLs, and immunization.” The mice were killed 2 days after the last treatment. (B) Splenocytes fromcontrol C3H/HeN (I,IV), LPS-treated C3H/HeN (II,V), and C3H/HeJ (III,VI) mice were labeled with CFSE and then activated ex vivo with anti-CD3 and anti-CD28 antibodies orwith PMA and Ca2� ionophore as described in “6-Carboxyfluorescein succinimidyl ester staining and ex vivo proliferation assay” (black histograms) or left untreated (whitehistograms). The proliferative response was assessed by monitoring cell divisions of gated CFSE-labeled Thy1.2� T cells. (C) Splenocytes from control and LPS-treated micewere activated for 48 hours and specific T cell proliferation was measured by BrdU incorporation in Thy-1.2� cells using FACS. The results are presented as the mean value of3 independent experiments, and standard deviations are shown. *P � .02; **P � .001 (Student t test). (D) Splenic Thy1.2� T cells from LPS-treated (black line) and control(gray area) C3H/HeN mice (I,II) or C3H/HeJ mice (III,IV) were analyzed for total � (I,III) and CD3 (II,IV) expression levels by FACS. (E) Equal numbers of splenic T cells fromLPS-treated and control mice were lysed and subjected to Western blot analysis. Expression of the protein tyrosine kinase ZAP-70, CD3 and �-chain was assessed byimmunoblotting (IB) using specific antibodies. A representative experiment is shown of 5 performed.
TLR-MEDIATED T AND NK CELL IMMUNOSUPPRESSION 1439BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom

C3H/HeN-treated mice compared with that of cells derived fromC3H/HeJ-treated mice or untreated control mice (Figure 1Bi-iii,C). However, when splenocytes derived from LPS-treatedC3H/HeN mice were stimulated with a combination of PMA andCa�2 ionophore, which bypasses TCR signaling, the T-cellproliferative response was partially rescued (up to 70%) relativeto those isolated from the LPS-treated C3H/HeJ or control mice(Figure 1Biv-vi,C). Although both TCR-dependent and -indepen-dent signaling events are impaired in T cells derived fromLPS-treated C3H/HeN mice, the effect on the TCR-mediatedactivation was by far more pronounced.
We next assessed whether the impaired TCR-mediated signal-ing function is associated with an abnormal TCR structure.Intracellular staining of splenic T cells (Figure 1D) and immuno-blot analysis (Figure 1E) showed a reduced �-chain expressioninduced by LPS in C3H/HeN (Figure 1Di,E) but not in C3H/HeJ(Figure 1Diii,E) mice. Despite the loss of �-chain in splenic T cellsfrom LPS-treated C3H/HeN mice, these cells expressed normaltotal levels of the CD3ε-chain (Figure 1Dii) as well as of thesurface CD3ε and TCR �� subunits (data not shown). Theexpression level of the protein tyrosine kinase, �-associatedprotein-70 (ZAP-70), a T-cell protein that is unrelated to the TCRsubunits, was unaffected under these conditions (Figure 1E). Thus,persistent activation via a single TLR can induce the entireimmunosuppressive milieu affecting all T cells similarly to thatinduced by an intact inactivated pathogen (Figure S1, available onthe Blood website; see the Supplemental Materials link at the top ofthe online article). The bystander T-cell immunosuppression wasobserved in both CD4� and CD8� subpopulations (data notshown). The results showing that both TCR-dependent and-independent signaling pathways are affected on chronic LPStreatment suggest that the abnormal TCR structure is not the solecause for the observed T-cell dysfunction.
Reduced antiviral response and impaired NK-cell function inLPS-treated mice
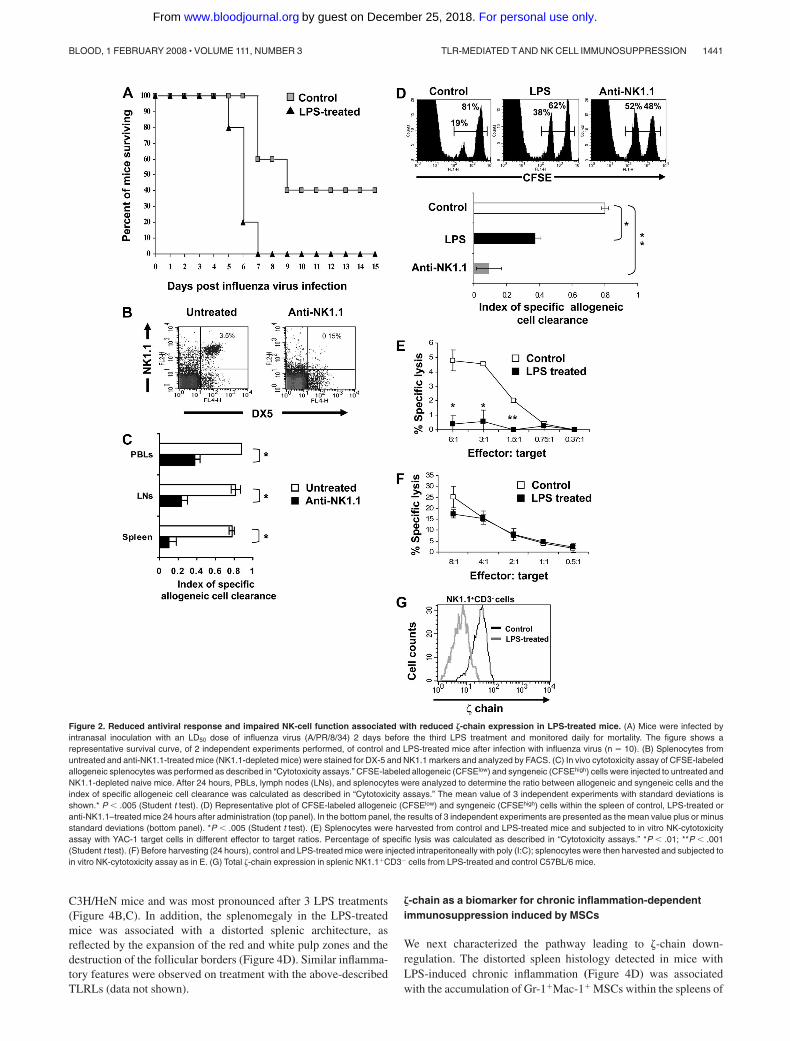
Because the immune response to viral infections involves acomplex coordination between the innate and adaptive systems,we next examined the in vivo antiviral immunity of LPS-treatedmice relative to controls. Mice were infected with an LD50 doseof influenza virus and monitored daily for mortality. The resultsrevealed a reduced survival rate of LPS-treated mice relative tocontrols (Figure 2A). Because of the kinetics of an antiviralimmune response in which NK-cell activity precedes the T-cellresponse, the expedited death of LPS-treated mice, 5 to 7 daysafter the viral infection, suggests that mortality is primarilyattributable to an impaired NK-mediated innate immune re-sponse, although the initial involvement of T cells could not beexcluded. These results demonstrate the suppressed antiviralactivity of the innate and adaptive immune systems on repeatedtreatment of mice with LPS.
We next assessed the effect of sustained activation of TLR4 byLPS on the innate immune system, focusing on NK cells, given thatthe �-chain also plays a crucial role in the function of NK-cellactivating receptors. In this set of experiments, we used C57B/6mice because NK cells could be detected by anti-NK1.1 antibodies.To study the effect of LPS treatment on in vivo NK-cell function,we followed their capacity to clear fluorescently labeled allogeneiccells.19 To verify that allogeneic cell clearance depends on NK-cellactivity, naive C57B/6 mice were first depleted of NK cells usinganti-NK1.1 antibody or nondepleted (Figure 2B) and then wereintravenously injected with a mixture of labeled allogeneic (CF-
SElow) and syngeneic (CFSEhigh) cells. The index of allogeneic cellclearance within the cell population of different lymphatic organswas then calculated. The results confirm that rejection of allogeneicsplenocytes is mediated primarily by NK cells because theclearance index of the allogeneic cells was significantly reduced inmice that were depleted of NK1.1-expressing cells. Based on theresults showing that the most pronounced effect was observed inthe spleen (Figure 2C), we then analyzed the function of NK cellsin the spleens of LPS-treated mice. The results revealed signifi-cantly impaired NK-cell function within LPS-treated mice, mani-fested by reduced clearance of allogeneic cells (Figure 2D, top andbottom panels). Furthermore, the dysfunction of NK cells isolatedfrom LPS-treated mice was also apparent by their reduced in vitrocytotoxic activity (Figure 2E). Poly(I:C) treatment of mice 24 hoursbefore cell harvest induced recovery of the NK-cell function(Figure 2F). As in the case of T cells, the impaired NK-cell function inthe LPS-treated mice was associated with a dramatic loss of �-chainexpression (Figure 2G). The approximate 5-fold increase in specificlysis activity of splenic NK cells derived from control or LPS-treatedmice observed on treatment with poly(I:C) is mostly attributable to thein vivo priming of NK cells by type 1 IFNs22 and could result fromactivation of NK-cell receptors that do not require �-chain expression.Thus, chronic exposure to LPS immunosuppressed the innate immunesystem, as reflected by the impaired killing function of NK cells and thereduction in �-chain expression levels.
Repetitive treatment with TLR 2, 3, or 9 ligands affect T-cellfunction and �-chain expression
To evaluate whether TLRLs other than the TLR4L, LPS, could inducean immunosuppressive effect in our in vivo model, we repeatedlyexposed normal mice to TLR2, 3, or 9 ligands encapsulated in MLVs.To evaluate the function of splenic T cells derived from these mice, exvivo proliferation assays were performed using intracellular staining forBrdU. We found that the ex vivo T-cell proliferation in the TLRL-treatedmice after TCR-mediated activation was impaired, whereas activationwith PMA and Ca2� ionophore partially overcame this defect (Figure3A). Impaired NK-cell function within the TLRL-treated mice was alsoobserved (data not shown). The immunosuppression was associatedwith �-chain down-regulation. In all cases, whereas �-chain expressionwas reduced, the expression of other TCR subunits (CD3ε) or signalingmolecules (ZAP-70) was unaffected (Figure 3B). These results showthat treatment with each of the TLRLs induced T-cell immunosuppres-sion associated with reduced �-chain expression, similar to that gener-ated by LPS (Figure 1) or by entire inactivated pathogens (Figure S1).
LPS treatment leads to an inflammatory response associatedwith abnormal spleen architecture
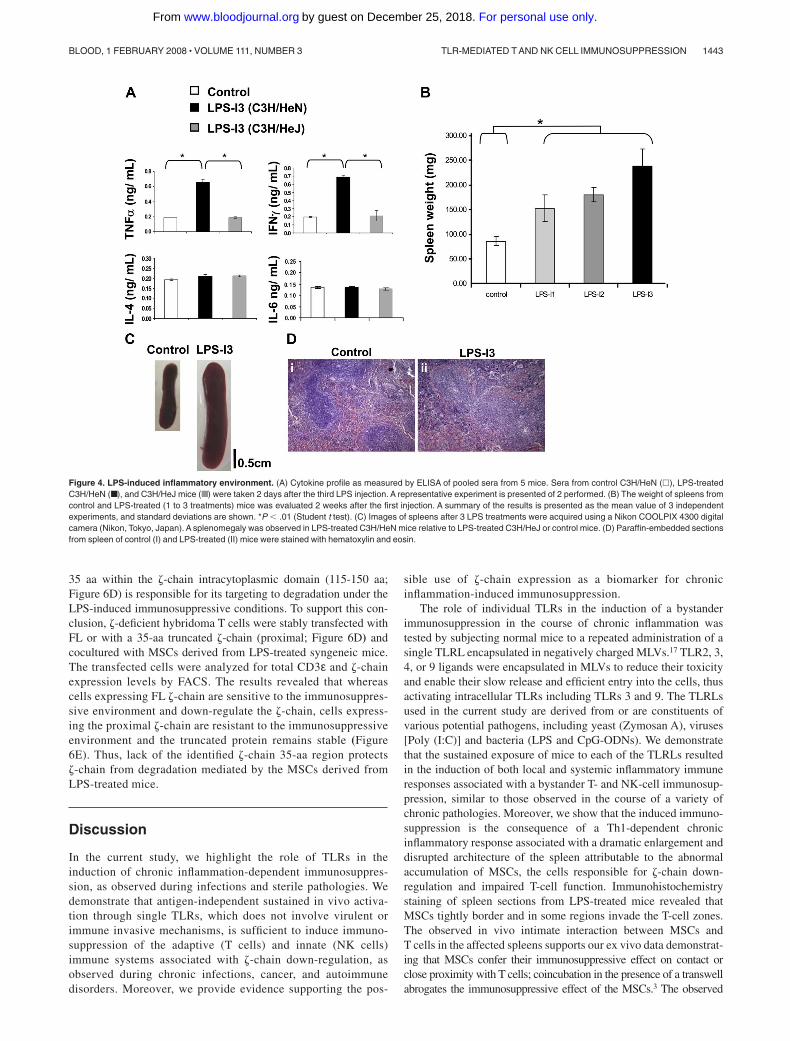
LPS treatment is known to induce an inflammatory response asreflected by the secretion of high levels of proinflammatorycytokines such as TNF-�, IL-1�, IL-8, and the induction of thesynthesis of other proinflammatory proteins, such as inducible NOsynthase.23 However, LPS was not previously shown to have anyimmunosuppressive effect on T- and NK-cell function or on �-chainexpression. We therefore evaluated whether, under our experimen-tal conditions, LPS treatment induces an inflammatory responseand how this inflammation affects secondary lymphatic organs.The first indication for an inflammatory response was the elevatedlevel of IFN-� and TNF-� within LPS-treated C3H/HeN mice; nosuch elevation was seen in LPS-treated C3H/HeJ or control mice(Figure 4A). Moreover, splenomegaly was observed in LPS-treated
1440 VAKNIN et al BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom
C3H/HeN mice and was most pronounced after 3 LPS treatments(Figure 4B,C). In addition, the splenomegaly in the LPS-treatedmice was associated with a distorted splenic architecture, asreflected by the expansion of the red and white pulp zones and thedestruction of the follicular borders (Figure 4D). Similar inflamma-tory features were observed on treatment with the above-describedTLRLs (data not shown).
�-chain as a biomarker for chronic inflammation-dependentimmunosuppression induced by MSCs
We next characterized the pathway leading to �-chain down-regulation. The distorted spleen histology detected in mice withLPS-induced chronic inflammation (Figure 4D) was associatedwith the accumulation of Gr-1�Mac-1� MSCs within the spleens of
Figure 2. Reduced antiviral response and impaired NK-cell function associated with reduced �-chain expression in LPS-treated mice. (A) Mice were infected byintranasal inoculation with an LD50 dose of influenza virus (A/PR/8/34) 2 days before the third LPS treatment and monitored daily for mortality. The figure shows arepresentative survival curve, of 2 independent experiments performed, of control and LPS-treated mice after infection with influenza virus (n 10). (B) Splenocytes fromuntreated and anti-NK1.1-treated mice (NK1.1-depleted mice) were stained for DX-5 and NK1.1 markers and analyzed by FACS. (C) In vivo cytotoxicity assay of CFSE-labeledallogeneic splenocytes was performed as described in “Cytotoxicity assays.” CFSE-labeled allogeneic (CFSElow) and syngeneic (CFSEhigh) cells were injected to untreated andNK1.1-depleted naive mice. After 24 hours, PBLs, lymph nodes (LNs), and splenocytes were analyzed to determine the ratio between allogeneic and syngeneic cells and theindex of specific allogeneic cell clearance was calculated as described in “Cytotoxicity assays.” The mean value of 3 independent experiments with standard deviations isshown.* P � .005 (Student t test). (D) Representative plot of CFSE-labeled allogeneic (CFSElow) and syngeneic (CFSEhigh) cells within the spleen of control, LPS-treated oranti-NK1.1–treated mice 24 hours after administration (top panel). In the bottom panel, the results of 3 independent experiments are presented as the mean value plus or minusstandard deviations (bottom panel). *P � .005 (Student t test). (E) Splenocytes were harvested from control and LPS-treated mice and subjected to in vitro NK-cytotoxicityassay with YAC-1 target cells in different effector to target ratios. Percentage of specific lysis was calculated as described in “Cytotoxicity assays.” *P � .01; **P � .001(Student t test). (F) Before harvesting (24 hours), control and LPS-treated mice were injected intraperitoneally with poly (I:C); splenocytes were then harvested and subjected toin vitro NK-cytotoxicity assay as in E. (G) Total �-chain expression in splenic NK1.1�CD3� cells from LPS-treated and control C57BL/6 mice.
TLR-MEDIATED T AND NK CELL IMMUNOSUPPRESSION 1441BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom

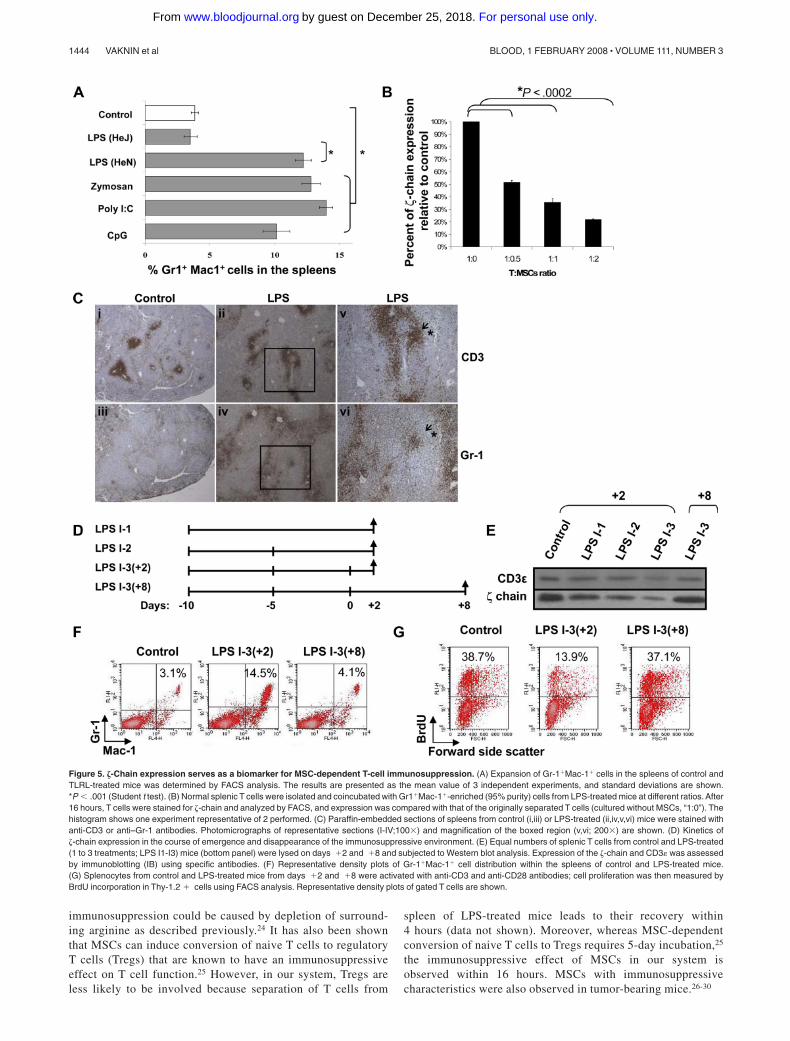
the TLR2, 3, 4, and 9 ligand-treated mice (Figure 5A). Ex vivocoincubation experiments revealed that Gr-1�Mac-1� cells derivedfrom LPS-treated mice induced �-chain down-regulation in normalT cells (Figure 5B) and ex vivo T-cell dysfunction (data not shown)in a ratio-dependent manner. MSCs isolated from mice treated withvarious TLRLs exhibited similar inhibitory effects on T cells (datanot shown). To determine whether there is a correlation betweenthe immunosuppressive function of MSCs and their in vivolocalization relative to T cells in the spleen of LPS-treated mice, weperformed immunohistochemistry on spleen sections using anti-CD3 and anti–Gr-1 antibodies (Figure 5C). The results revealedthat relative to control mice, in spleens of LPS-treated mice, a morediffused CD3 staining (Figure 5Ci,ii) and an increased number ofGr-1–positive cells surrounding the T-cell zone (Figure 5Ciii,iv)were observed. Moreover, in some regions, an invasion of Gr-1–positive cells into the T cell zones was observed (Figure 5Cv,vi).
To evaluate whether the �-chain expression levels could serve asa biomarker for sensing changes in chronic inflammation-dependent immunosuppression, its emergence and disappearance,we performed kinetic experiments comparing the time frame ofboth processes (Figure 5D). Gradual �-chain down-regulation wasobserved as the inflammatory response progressed during thedifferent intervals (1 to 3 LPS injections), with the most pro-nounced effect after the third LPS injection (Figure 5E). Interest-ingly, 8 days after the third LPS injection, a recovery of �-chainexpression was observed relative to day � 2 (Figure 5E), correlat-ing with normal percentages of MSCs (Figure 5F) and recovered invitro TCR-mediated T-cell proliferation (Figure 5G). Thus, �-chainexpression levels are reversible and are modified according to theinflammatory environment, suggesting its possible use as a biomar-ker to monitor chronic inflammation-dependent immunosuppression.
The mechanism for �-chain down-regulation under chronicinflammatory conditions
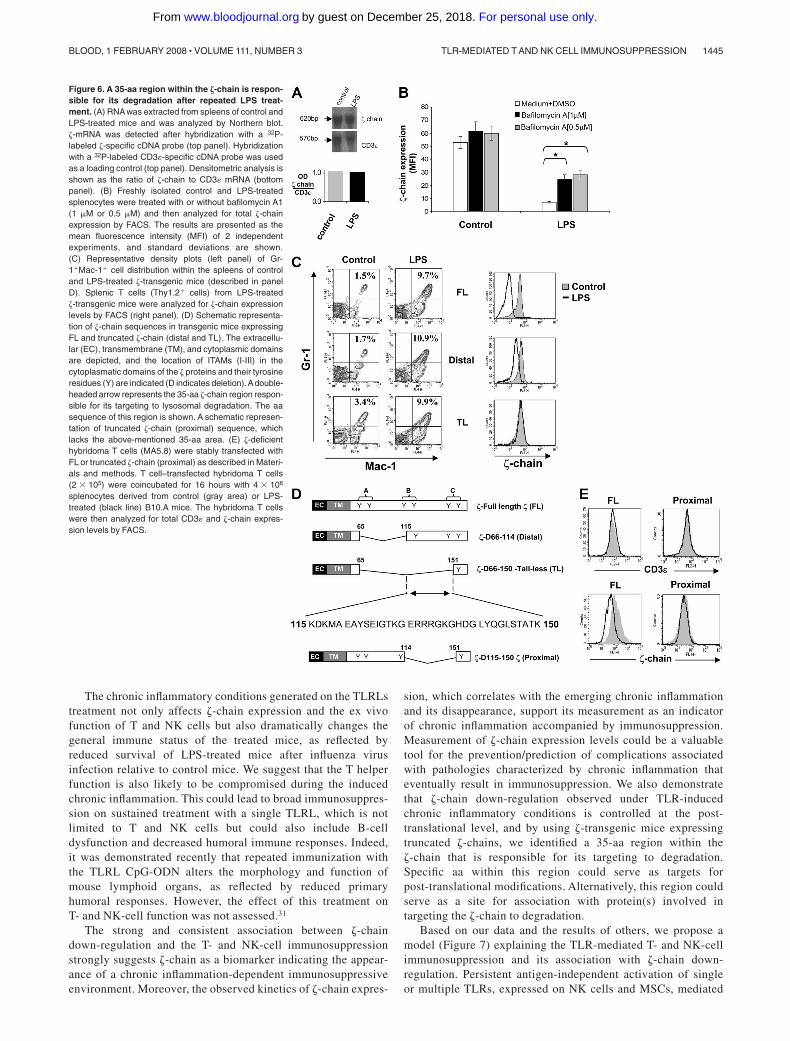
We next assessed at which level �-chain expression is regulatedunder chronic inflammatory conditions. Northern blot analysis ofsplenocytes revealed that �-chain mRNA levels of LPS-treatedmice were similar to those of control mice (Figure 6A), suggestingthat �-chain expression is regulated at the translational or post-translational level. Treatment of splenocytes isolated from LPS-treated mice with the lysosome inhibitor bafilomycin A increased�-chain expression up to 3-fold relative to �-chain down-regulatedcells that were untreated and to approximately 60% of �-chainexpression levels in control cells (Figure 6B). Furthermore, protea-somal inhibitors did not affect � expression under these conditions(data not shown). These results suggest a post-translational mecha-nism controlling �-chain expression under chronic inflammatoryconditions.
Our next goal was to locate the region within the �-chain that isresponsible for its targeting to degradation. To this end, we usedtransgenic mice expressing either FL �-chain (FL-�) or �-chainvariants truncated in the intracytoplasmic domain, one containingonly a single distal ITAM (distal) and the second containing noITAMs (TL).15,16 These mice were treated with LPS as described inMaterials and methods. Chronic inflammation was generated in allLPS-treated transgenic mice as shown by the elevation of MSCswithin their spleen relative to the nontreated mice (Figure 6C leftpanel). Interestingly, analysis of �-chain expression in these micerevealed that although down-regulation was induced in the FL-�and distal transgenic mice after LPS treatment, �-chain expressionlevels in the TL mice were not altered in response to LPS (Figure6C right panel). These results indicate that the region spanning
Figure 3. Treatment with TLR 2, 3, or 9 ligands inducesimpaired T-cell proliferation and �-chain down-regulation.Mice were injected subcutaneously 3 times with MLVs andencapsulated with one of the following TLRLs: Zymosan(TLR 2), poly (I:C) (TLR 3), or CpG (TLR 9), as described in“Antigen, TLRLs, and immunization.” (A) Proliferative re-sponse as determined by the percentage of BrdU�Thy1.2�
of total Thy1.2� cells isolated from control mice (�) relativeto TLRL-treated mice (■) after stimulation with anti-CD3 andanti-CD28 antibodies or with PMA and Ca2� ionophore. Theresults are presented as the mean values of 3 independentexperiments, and standard deviations are shown. *P � .005(Student t test). (B) Equal numbers of splenic T cells fromZymosan, poly (I:C), and CpG-treated and control micewere lysed and subjected to Western blot analysis. Expres-sion of the protein kinase ZAP-70, CD3 , and �-chain wasassessed by immunoblotting (IB) using specific antibodies.
1442 VAKNIN et al BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom
35 aa within the �-chain intracytoplasmic domain (115-150 aa;Figure 6D) is responsible for its targeting to degradation under theLPS-induced immunosuppressive conditions. To support this con-clusion, �-deficient hybridoma T cells were stably transfected withFL or with a 35-aa truncated �-chain (proximal; Figure 6D) andcocultured with MSCs derived from LPS-treated syngeneic mice.The transfected cells were analyzed for total CD3ε and �-chainexpression levels by FACS. The results revealed that whereascells expressing FL �-chain are sensitive to the immunosuppres-sive environment and down-regulate the �-chain, cells express-ing the proximal �-chain are resistant to the immunosuppressiveenvironment and the truncated protein remains stable (Figure6E). Thus, lack of the identified �-chain 35-aa region protects�-chain from degradation mediated by the MSCs derived fromLPS-treated mice.
Discussion
In the current study, we highlight the role of TLRs in theinduction of chronic inflammation-dependent immunosuppres-sion, as observed during infections and sterile pathologies. Wedemonstrate that antigen-independent sustained in vivo activa-tion through single TLRs, which does not involve virulent orimmune invasive mechanisms, is sufficient to induce immuno-suppression of the adaptive (T cells) and innate (NK cells)immune systems associated with �-chain down-regulation, asobserved during chronic infections, cancer, and autoimmunedisorders. Moreover, we provide evidence supporting the pos-
sible use of �-chain expression as a biomarker for chronicinflammation-induced immunosuppression.
The role of individual TLRs in the induction of a bystanderimmunosuppression in the course of chronic inflammation wastested by subjecting normal mice to a repeated administration of asingle TLRL encapsulated in negatively charged MLVs.17 TLR2, 3,4, or 9 ligands were encapsulated in MLVs to reduce their toxicityand enable their slow release and efficient entry into the cells, thusactivating intracellular TLRs including TLRs 3 and 9. The TLRLsused in the current study are derived from or are constituents ofvarious potential pathogens, including yeast (Zymosan A), viruses[Poly (I:C)] and bacteria (LPS and CpG-ODNs). We demonstratethat the sustained exposure of mice to each of the TLRLs resultedin the induction of both local and systemic inflammatory immuneresponses associated with a bystander T- and NK-cell immunosup-pression, similar to those observed in the course of a variety ofchronic pathologies. Moreover, we show that the induced immuno-suppression is the consequence of a Th1-dependent chronicinflammatory response associated with a dramatic enlargement anddisrupted architecture of the spleen attributable to the abnormalaccumulation of MSCs, the cells responsible for �-chain down-regulation and impaired T-cell function. Immunohistochemistrystaining of spleen sections from LPS-treated mice revealed thatMSCs tightly border and in some regions invade the T-cell zones.The observed in vivo intimate interaction between MSCs andT cells in the affected spleens supports our ex vivo data demonstrat-ing that MSCs confer their immunosuppressive effect on contact orclose proximity with T cells; coincubation in the presence of a transwellabrogates the immunosuppressive effect of the MSCs.3 The observed
Figure 4. LPS-induced inflammatory environment. (A) Cytokine profile as measured by ELISA of pooled sera from 5 mice. Sera from control C3H/HeN (�), LPS-treatedC3H/HeN (■), and C3H/HeJ mice (■) were taken 2 days after the third LPS injection. A representative experiment is presented of 2 performed. (B) The weight of spleens fromcontrol and LPS-treated (1 to 3 treatments) mice was evaluated 2 weeks after the first injection. A summary of the results is presented as the mean value of 3 independentexperiments, and standard deviations are shown. *P � .01 (Student t test). (C) Images of spleens after 3 LPS treatments were acquired using a Nikon COOLPIX 4300 digitalcamera (Nikon, Tokyo, Japan). A splenomegaly was observed in LPS-treated C3H/HeN mice relative to LPS-treated C3H/HeJ or control mice. (D) Paraffin-embedded sectionsfrom spleen of control (I) and LPS-treated (II) mice were stained with hematoxylin and eosin.
TLR-MEDIATED T AND NK CELL IMMUNOSUPPRESSION 1443BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom
immunosuppression could be caused by depletion of surround-ing arginine as described previously.24 It has also been shownthat MSCs can induce conversion of naive T cells to regulatoryT cells (Tregs) that are known to have an immunosuppressiveeffect on T cell function.25 However, in our system, Tregs areless likely to be involved because separation of T cells from
spleen of LPS-treated mice leads to their recovery within4 hours (data not shown). Moreover, whereas MSC-dependentconversion of naive T cells to Tregs requires 5-day incubation,25
the immunosuppressive effect of MSCs in our system isobserved within 16 hours. MSCs with immunosuppressivecharacteristics were also observed in tumor-bearing mice.26-30
Figure 5. �-Chain expression serves as a biomarker for MSC-dependent T-cell immunosuppression. (A) Expansion of Gr-1�Mac-1� cells in the spleens of control andTLRL-treated mice was determined by FACS analysis. The results are presented as the mean value of 3 independent experiments, and standard deviations are shown.*P � .001 (Student t test). (B) Normal splenic T cells were isolated and coincubated with Gr1�Mac-1�-enriched (95% purity) cells from LPS-treated mice at different ratios. After16 hours, T cells were stained for �-chain and analyzed by FACS, and expression was compared with that of the originally separated T cells (cultured without MSCs, “1:0”). Thehistogram shows one experiment representative of 2 performed. (C) Paraffin-embedded sections of spleens from control (i,iii) or LPS-treated (ii,iv,v,vi) mice were stained withanti-CD3 or anti–Gr-1 antibodies. Photomicrographs of representative sections (I-IV;100�) and magnification of the boxed region (v,vi; 200�) are shown. (D) Kinetics of�-chain expression in the course of emergence and disappearance of the immunosuppressive environment. (E) Equal numbers of splenic T cells from control and LPS-treated(1 to 3 treatments; LPS I1-I3) mice (bottom panel) were lysed on days �2 and �8 and subjected to Western blot analysis. Expression of the �-chain and CD3 was assessedby immunoblotting (IB) using specific antibodies. (F) Representative density plots of Gr-1�Mac-1� cell distribution within the spleens of control and LPS-treated mice.(G) Splenocytes from control and LPS-treated mice from days �2 and �8 were activated with anti-CD3 and anti-CD28 antibodies; cell proliferation was then measured byBrdU incorporation in Thy-1.2 � cells using FACS analysis. Representative density plots of gated T cells are shown.
1444 VAKNIN et al BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom
The chronic inflammatory conditions generated on the TLRLstreatment not only affects �-chain expression and the ex vivofunction of T and NK cells but also dramatically changes thegeneral immune status of the treated mice, as reflected byreduced survival of LPS-treated mice after influenza virusinfection relative to control mice. We suggest that the T helperfunction is also likely to be compromised during the inducedchronic inflammation. This could lead to broad immunosuppres-sion on sustained treatment with a single TLRL, which is notlimited to T and NK cells but could also include B-celldysfunction and decreased humoral immune responses. Indeed,it was demonstrated recently that repeated immunization withthe TLRL CpG-ODN alters the morphology and function ofmouse lymphoid organs, as reflected by reduced primaryhumoral responses. However, the effect of this treatment onT- and NK-cell function was not assessed.31
The strong and consistent association between �-chaindown-regulation and the T- and NK-cell immunosuppressionstrongly suggests �-chain as a biomarker indicating the appear-ance of a chronic inflammation-dependent immunosuppressiveenvironment. Moreover, the observed kinetics of �-chain expres-
sion, which correlates with the emerging chronic inflammationand its disappearance, support its measurement as an indicatorof chronic inflammation accompanied by immunosuppression.Measurement of �-chain expression levels could be a valuabletool for the prevention/prediction of complications associatedwith pathologies characterized by chronic inflammation thateventually result in immunosuppression. We also demonstratethat �-chain down-regulation observed under TLR-inducedchronic inflammatory conditions is controlled at the post-translational level, and by using �-transgenic mice expressingtruncated �-chains, we identified a 35-aa region within the�-chain that is responsible for its targeting to degradation.Specific aa within this region could serve as targets forpost-translational modifications. Alternatively, this region couldserve as a site for association with protein(s) involved intargeting the �-chain to degradation.
Based on our data and the results of others, we propose amodel (Figure 7) explaining the TLR-mediated T- and NK-cellimmunosuppression and its association with �-chain down-regulation. Persistent antigen-independent activation of singleor multiple TLRs, expressed on NK cells and MSCs, mediated
Figure 6. A 35-aa region within the �-chain is respon-sible for its degradation after repeated LPS treat-ment. (A) RNA was extracted from spleens of control andLPS-treated mice and was analyzed by Northern blot.�-mRNA was detected after hybridization with a 32P-labeled �-specific cDNA probe (top panel). Hybridizationwith a 32P-labeled CD3 -specific cDNA probe was usedas a loading control (top panel). Densitometric analysis isshown as the ratio of �-chain to CD3 mRNA (bottompanel). (B) Freshly isolated control and LPS-treatedsplenocytes were treated with or without bafilomycin A1(1 �M or 0.5 �M) and then analyzed for total �-chainexpression by FACS. The results are presented as themean fluorescence intensity (MFI) of 2 independentexperiments, and standard deviations are shown.(C) Representative density plots (left panel) of Gr-1�Mac-1� cell distribution within the spleens of controland LPS-treated �-transgenic mice (described in panelD). Splenic T cells (Thy1.2� cells) from LPS-treated�-transgenic mice were analyzed for �-chain expressionlevels by FACS (right panel). (D) Schematic representa-tion of �-chain sequences in transgenic mice expressingFL and truncated �-chain (distal and TL). The extracellu-lar (EC), transmembrane (TM), and cytoplasmic domainsare depicted, and the location of ITAMs (I-III) in thecytoplasmatic domains of the � proteins and their tyrosineresidues (Y) are indicated (D indicates deletion).Adouble-headed arrow represents the 35-aa �-chain region respon-sible for its targeting to lysosomal degradation. The aasequence of this region is shown. A schematic represen-tation of truncated �-chain (proximal) sequence, whichlacks the above-mentioned 35-aa area. (E) �-deficienthybridoma T cells (MA5.8) were stably transfected withFL or truncated �-chain (proximal) as described in Materi-als and methods. T cell–transfected hybridoma T cells(2 � 105) were coincubated for 16 hours with 4 � 106
splenocytes derived from control (gray area) or LPS-treated (black line) B10.A mice. The hybridoma T cellswere then analyzed for total CD3 and �-chain expres-sion levels by FACS.
TLR-MEDIATED T AND NK CELL IMMUNOSUPPRESSION 1445BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom

via TLR agonists (pathogenic or endogenous), in conjunction orseparately of an antigen-dependent T-cell activation mediatedvia the TCR, when stimulated, lead to an inflammatory responsecharacterized by the production of IFN-�. These activationpathways are characteristic of both sterile and infectiouspathologies. Elevated levels of IFN-�, together with TLRLpersistence, lead to the recruitment/expansion and activation ofMSCs within the affected tissue and lymphatic organs. Thecontinuous presence of activated MSCs affects the surroundingT and NK cells, leading to their hyporesponsiveness, associatedwith the loss of �-chain expression. Thus, �-chain expressionlevels could serve as a biomarker “sensing” changes in thechronic inflammatory immunosuppressive environment and in-dicative of immunosuppression. Our results lead us to proposethat the nature of the generated TLR-mediated inflammatoryresponse, whether acute or chronic, will dictate whether itsimmunologic consequence is beneficial or harmful. Whereas anacute inflammatory response is vital for the immediate immunedefense against pathogens and for the clearance of abnormal self-cellsand molecules, a chronic inflammatory response could be detrimental tothe host under conditions in which the host is unable to clear thepathogen because of the developing immunosuppression. In fact, TLRshave been shown to play a key role in the development of widespreadpathologies.32 Because TLRs, and particularly TLR4, recognize endoge-nous molecules,5 TLRLs might have an important role in the inductionof the local and systemic inflammation characterizing “sterile” patholo-gies, such as autoimmune diseases, cancer, and other pathologiescharacterized by chronic inflammation, in which the cause is unknown.We suggest that immunosuppression resulting from TLR-mediatedchronic inflammation evolved to limit immunopathology in the contextof chronic stimulation of the innate immune system and avoid tissuedamage, sepsis, organ failure, and death. Thus, the delicate balancebetween inflammatory response and its extent is critical to a normal orimmunosuppressed immune system.
Because of the key role of TLRLs in activation of the innate immunesystem leading mostly to a Th1-dependent inflammatory response, theseagents are currently being tested as adjuvants for antimicrobial, antialler-gic, and anticancer immunotherapy.5,33 However, based on our results,the feasibility of using TLRLs in a given therapy must be carefullyinvestigated; the duration of such a treatment will dictate whether itseffect is beneficial or harmful to the patient. Our description of theimmunosuppressive process of the adaptive and innate immune systemstriggered upon sustained activation of individual TLRs may allow arange of challenges in immunology and medicine to be addressed, withthe goal of developing modalities that will block the inflammatoryenvironment specifically induced by TLRLs. Identification of specific
TLRs involved in the induction of chronic inflammation in a givenpathology (eg, TLR9 in Lupus, TLR4 in LPS-mediated septic shock)could enable the development of controlled and disease-specific immu-notherapy. Moreover, the mouse model system that we used in thecurrent study might serve as a platform for testing new or currentlyused drugs for their ability to reduce chronic inflammation and to avoidcomplications resulting from the associated immunosuppression.
Acknowledgments
We gratefully acknowledge the support of the Society of ResearchAssociates of the Lautenberg Center, the Concern Foundation ofLos Angeles, and the Harold B. Abramson Chair in Immunology.Thanks to Dr Elizabeth Shores from the FDA, National Institutes ofHealth, Bethesda, MD, for providing us with �-transgenic and��/� mice. We thank Dr Eyal Raz from the University ofCalifornia, San Diego, for providing the CpG-ODNs and (m)CpG-ODNs. Thanks also to Prof Eli Kedar and Dr Aviva Joseph forhelping us with MLV preparation and to Prof Zichria Zakay-Ronesand Roi Gazit for providing the influenza virus. Thanks to Dr TaliNave for helping with Northern blot analysis. Thanks also to EfratManaster, Dr Shelley Shwarzbaum, Dr Steve Caplan, and ProfRachel Ehrlich for critical reading of the manuscript.
This study was supported by the Israel Academy of Sciencesand Humanities, the Israeli Ministry of Health, the Israel CancerAssociation, the Joint German-Israeli Research Program (DKFZ),the Israel Cancer Research Fund (ICRF), and by the Joseph andMatilda Melnick Funds.
Authorship
Contribution: I.V. designed and performed research and wrote thepaper; L.B., R.G., and E.S. performed research; L.W. contributedvital new reagents; E.P. contributed vital new reagents or analyticaltools and analyzed data; M.P. contributed vital new reagents oranalytical tools; and M.B. designed research and wrote the paper.
Conflict-of-interest disclosure: The authors declare no compet-ing financial interests.
Correspondence: Dr Michal Baniyash, Lautenberg Center forGeneral and Tumor Immunology, Hebrew University-HadassahMedical School, PO Box 12272, Jerusalem 91120, Israel; e-mail:[email protected].
Figure 7. Model depicting TLR-mediated T- and NK-cell immunosup-pression. (A) Persistent Ag-independent activation of NK cells and MSCsby single or multiple TLRLs, typical of pathogens (infections) or endoge-nous compounds (cancer and autoimmune diseases), leads to aninflammatory response characterized by the production of IFN-�. This canoccur in conjunction with Ag-dependent T cell activation, which alsocontributes to the increased concentrations of IFN-�. (B) Elevated levelsof IFN-�, together with TLRL persistence, lead to the recruitment/expansion and activation of MSCs within the affected tissue and lym-phatic organs. (C) The continuous presence of activated MSCs affectsthe surrounding T and NK cells, leading to their hyporesponsiveness andloss of �-chain expression, which is a biomarker “sensing” changes in thechronic inflammatory immunosuppressive environment.
1446 VAKNIN et al BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom

References
1. Baniyash M. TCR zeta-chain downregulation:curtailing an excessive inflammatory immune re-sponse. Nat Rev Immunol. 2004;4:675-687.
2. Bronstein-Sitton N, Cohen-Daniel L, Vaknin I, etal. Sustained exposure to bacterial antigen in-duces interferon-gamma-dependent T cell recep-tor zeta down-regulation and impaired T cell func-tion. Nat Immunol. 2003;4:957-964.
3. Ezernitchi AV, Vaknin I, Cohen-Daniel L, et al.TCR zeta down-regulation under chronic inflam-mation is mediated by myeloid suppressor cellsdifferentially distributed between various lym-phatic organs. J Immunol. 2006;177:4763-4772.
4. Akira S, Takeda K. Toll-like receptor signalling.Nat Rev Immunol. 2004;4:499-511.
5. Beutler B. Inferences, questions and possibilitiesin Toll-like receptor signalling. Nature. 2004;430:257-263.
6. Alexopoulou L, Holt AC, Medzhitov R, et al. Rec-ognition of double-stranded RNA and activation ofNF-kappaB by Toll-like receptor 3. Nature. 2001;413:732-738.
7. Hoshino K, Takeuchi O, Kawai T, et al. Cuttingedge: Toll-like receptor 4 (TLR4)-deficient miceare hyporesponsive to lipopolysaccharide: evi-dence for TLR4 as the Lps gene product. J Immu-nol. 1999;162:3749-3752.
8. Hemmi H, Takeuchi O, Kawai T, et al. A Toll-likereceptor recognizes bacterial DNA. Nature. 2000;408:740-745.
9. Sato M, Sano H, Iwaki D, et al. Direct binding ofToll-like receptor 2 to zymosan, and zymosan-induced NF-kappa B activation and TNF-alphasecretion are down-regulated by lung collectinsurfactant protein A. J Immunol. 2003;171:417-425.
10. Wagner H. Endogenous TLR ligands and autoim-munity. Adv Immunol. 2006;91:159-173.
11. Aderem A, Ulevitch RJ. Toll-like receptors in theinduction of the innate immune response. Nature.2000;406:782-787.
12. Alberola-Ila J, Takaki S, Kerner JD, et al. Differen-tial signaling by lymphocyte antigen receptors.Annu Rev Immunol. 1997;15:125-154.
13. Moretta A, Bottino C, Vitale M, et al. Activatingreceptors and coreceptors involved in humannatural killer cell-mediated cytolysis. Annu RevImmunol. 2001;19:197-223.
14. Lanier LL, Yu G, Phillips JH. Coassociation ofCD3 zeta with a receptor (CD16) for IgG Fc onhuman natural killer cells. Nature. 1989;342:803-805.
15. Shores EW, Huang K, Tran T, et al. Role of TCRzeta chain in T cell development and selection.Science. 1994;266:1047-1050.
16. Love PE, Shores EW, Johnson MD, et al. T celldevelopment in mice that lack the zeta chain ofthe T cell antigen receptor complex. Science.1993;261:918-921.
17. Joseph A, Louria-Hayon I, Plis-Finarov A, et al.Liposomal immunostimulatory DNA sequence(ISS-ODN): an efficient parenteral and mucosaladjuvant for influenza and hepatitis B vaccines.Vaccine. 2002;20:3342-3354.
18. Gazit R, Gruda R, Elboim M, et al. Lethal influ-enza infection in the absence of the natural killercell receptor gene Ncr1. Nat Immunol. 2006;7:517-523.
19. Oberg L, Johansson S, Michaelsson J, et al. Lossor mismatch of MHC class I is sufficient to triggerNK cell-mediated rejection of resting lymphocytesin vivo–role of KARAP/DAP12-dependent and-independent pathways. Eur J Immunol. 2004;34:1646-1653.
20. Mollen KP, Anand RJ, Tsung A, et al. Emergingparadigm: toll-like receptor 4-sentinel for the de-tection of tissue damage. Shock. 2006;26:430-437.
21. Baldridge JR, McGowan P, Evans JT, et al. Takinga Toll on human disease: Toll-like receptor 4 ago-nists as vaccine adjuvants and monotherapeuticagents. Expert Opin Biol Ther. 2004;4:1129-1138.
22. Lee CK, Rao DT, Gertner R, et al. Distinct re-quirements for IFNs and STAT1 in NK cell func-tion. J Immunol. 2000;165:3571-3577.
23. Palsson-McDermott EM, O’Neill LA. Signal trans-duction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology. 2004;113:153-162.
24. Makarenkova VP, Bansal V, Matta BM, et al.CD11b�/Gr-1� myeloid suppressor cells causeT cell dysfunction after traumatic stress. J Immu-nol. 2006;176:2085-2094.
25. Huang B, Pan PY, Li Q, et al. Gr-1�CD115� im-mature myeloid suppressor cells mediate the de-velopment of tumor-induced T regulatory cellsand T-cell anergy in tumor-bearing host. CancerRes. 2006;66:1123-1131.
26. Otsuji M, Kimura Y, Aoe T, et al. Oxidative stressby tumor-derived macrophages suppresses theexpression of CD3 zeta chain of T-cell receptorcomplex and antigen-specific T-cell responses.Proc Natl Acad Sci U S A. 1996;93:13119-13124.
27. Bronte V, Apolloni E, Cabrelle A, et al. Identifica-tion of a CD11b(�)/Gr-1(�)/CD31(�) myeloidprogenitor capable of activating or suppressingCD8(�) T cells. Blood. 2000;96:3838-3846.
28. Gabrilovich DI, Velders MP, Sotomayor EM, et al.Mechanism of immune dysfunction in cancer me-diated by immature Gr-1� myeloid cells. J Immu-nol. 2001;166:5398-5406.
29. Kusmartsev S, Nefedova Y, Yoder D, et al. Anti-gen-specific inhibition of CD8� T cell responseby immature myeloid cells in cancer is mediatedby reactive oxygen species. J Immunol. 2004;172:989-999.
30. Gallina G, Dolcetti L, Serafini P, et al. Tumors in-duce a subset of inflammatory monocytes withimmunosuppressive activity on CD8� T cells.J Clin Invest. 2006;116:2777-2790.
31. Heikenwalder M, Polymenidou M, Junt T, et al.Lymphoid follicle destruction and immunosup-pression after repeated CpG oligodeoxynucle-otide administration. Nat Med. 2004;10:187-192.
32. Cristofaro P, Opal SM. Role of Toll-like receptorsin infection and immunity: clinical implications.Drugs. 2006;66:15-29.
33. Weeratna RD, Makinen SR, McCluskie MJ, et al.TLR agonists as vaccine adjuvants: comparisonof CpG ODN and Resiquimod (R-848). Vaccine.2005;23:5263-5270.
TLR-MEDIATED T AND NK CELL IMMUNOSUPPRESSION 1447BLOOD, 1 FEBRUARY 2008 � VOLUME 111, NUMBER 3
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom
online November 8, 2007 originally publisheddoi:10.1182/blood-2007-07-100404
2008 111: 1437-1447
Pikarsky and Michal BaniyashIlan Vaknin, Liora Blinder, Lynn Wang, Roi Gazit, Elena Shapira, Olga Genina, Mark Pines, Eli
cell immunosuppression−natural killerA common pathway mediated through Toll-like receptors leads to T- and
http://www.bloodjournal.org/content/111/3/1437.full.htmlUpdated information and services can be found at:
(5644 articles)Immunobiology and Immunotherapy Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on December 25, 2018. by guest www.bloodjournal.orgFrom