a delay in consolidation is observed in a heterozygous...
TRANSCRIPT

Noggin and Chordin knockdown in Distraction Osteogenesis
Priscille Grenier-Vallée, MD
Department of Experimental Surgery
McGill University
Montreal, Québec Canada
April 2013
A Delay in Consolidation is Observed in a Heterozygous Conditional BMP2 Deficient Mouse Model of Distraction
Osteogenesis
Norine Alam
Department of Human Genetics McGill University Montreal, Québec
Canada
February 2009
A thesis submitted to McGill University, Faculty of Graduate and Postdoctoral Studies, in partial fulfillment of the requirements of the
degree of Masters of Science in Human Genetics
©Copyright Norine Alam, 2009.
1
A thesis submitted to McGill University, Faculty of Graduate and Postdoctoral Studies, in partial fulfillment of the requirements of the degree of Masters of Science in
Experimental Surgery

ABSTRACT
Introduction: Numerous reports have shown that recombinant BMPs have positive effects in several conditions associated with poor bone formation. We hypothesize that by inhibiting BMP antagonists Noggin and Chordin using RNA interference we may upregulate endogenous BMP expression and enhance osteogenesis.
Methods: MC3T3-E1 cells were transduced with lentiviruses expressing various shRNAs targeting the mouse Noggin (5 shRNAs) and Chordin (3 shRNAs) genes and 1 negative control. At various time points after infection, levels of RNA expression for Noggin and Chordin were monitored through RT-PCR. Western blotting were performed on the cell extracts and culture media to verify the expression and secretion of Noggin and Chordin proteins. Cell extracts were also analyzed on day 2 and 4 after transduction for alkaline phosphastase activity which is a marker of osteogenic differentiation.
DO was performed on the right tibia of 54 wild-type mice using a miniature Illizarov distraction device. Animals were randomized into two major groups according to the time of sacrifice: end of distraction (day 17) and mid-consolidation (day 34). Each major group was sub-divided in 3 subgroups representing the type of injection that was administered at the site of distraction on day 8. For each time point, 9 mice were injected with PBS, 9 with a lentivirus plasmid containing a Non-Target (NT) shRNA and 9 with lentivirus plasmids containing shRNAs targeting Noggin-Chordin. To evaluate the success of the infection a GFP marker was added to lentivirus plasmid targeting Noggin-Chordin. For each subgroup 6 collected samples were studied using Faxitron X-ray, µCT and immunohistochemistry and 3 were studied using Rt-PCR.
Results: On non-tranduced MC3T3-E1 cells, the levels of RNA expression of Noggin and Chordin was at its highest level on Day 7. At this timepoint, qRT-PCR analysis showed that the shRNAs were effective in knocking down Noggin and Chordin endogenous mRNA levels down to 10% and 17%, respectively, by the most potent of shRNAs tested compared to control. Western Blot analysis also corroborates that the respective shRNAs were effective in knocking down the Noggin and Chordin proteins. Specific activity of alkaline phosphatase was increased in MC3T3-E1 cells stably expressing Noggin and Chordin shRNA.
Concerning our in vivo work, all the surgeries and distraction were successful except for the death of 2 animals secondary to post-operative complications. However, at the time of the injection the majority of the injection content was extravasating out of the injection site. Once the samples collected, the Faxitron X-ray and µCT studies did not show a significant difference of bone volume or in the ratio of bone volume/tissue volume at the distraction site in between the 3 injection subgroups at both time points. The Rt-PCR studies did not show a significant difference in inhibiting Noggin or Chordin mRNA levels in between the 3 subgroups at both time points. Finally, the immunohistochemistry studies were stopped after the GFP marker was found in all the three subgroups.
2

Conclusion: In our in vitro study, western blot analysis, qRT-PCR and ALP assay were consistent with a successful knockdown of Noggin and Chordin.
During our in vivo study, the main issues faced during this project were technical problems during the injection. Unfortunately, our study did not show any significant difference in bone formation after using injection of lentivirus plasmid with shRNA targetting BMPs antagonists Noggin and Chordin.
3

RÉSUMÉ
Introduction: Plusieurs études ont demontré que l’utilisation des BMPs ont des effets positifs sur plusieurs conditions associées avec une mauvaise formation osseuse. Notre hypothèse est qu’en inhibant les antagonites des BMPs Noggin et Chordin en utilisant une technique d’interférence de l’ARN nous pourrions augmenter l’expression endogène des BMP et par le fait même augmenter l’osteogénèse.
Methodes: Les cellules MC3T3-E1 furent transfectées avec les lentivirus qui expriment différents shRNAs visant Noggin (5 shRNAs) et Chordin (3 shRNAS) chez la souris. Un contrôle négatif fut aussi sélectionné, NT (Non-Target shRNA). À différents temps après l’infection, les niveau d’expression d’ARN furent mesurés à l’aide d’un RT-PCR. Des études de Western blot furent performées sur les extraits cellulaires et sur les milieux de culture pour vérifier l’expression et la sécrétion des protéines Noggin et Chordin. Des extraits cellulaires furent aussi analysés au jour 2 et 4 après l’infection pour mesurer l’activité de l’alkaline phosphatase, qui est un marqueur de la différentiation osteogenique.
Par la suite, une distraction osseuse a été réalisée sur le tibia droit de 54 souris de type sauvage à l’aide d’un miniature fixateur externe Ilizarov. Les animaux étaient randomisés en 2 groupes selon leur temps de sacrifice : fin de la distraction (Jour 17) et mi-consolidation (Jour 34). Chaque groupe était sous-divisé en 3 sous-groupes représentant le type d’injection qui était administrée au site de distraction au jour 8. Pour chaque temps de sacrifice, 9 souris étaient injectées avec du PBS, 9 souris avec un Non-Target shRNA (NT) et 9 souris avec des lentivirus contenant Noggin et Chordin. Pour évaluer le succès notre infection, le shRNA qui visait Noggin fut transféré dans un plasmide contenant un marqueur GFP. Aussi, pour chaque sous-groupe, 6 échantillons étaient collectés pour une étude au Faxitron X-ray, µCT and immunohistochimie et 3 échantillons furent étudiés à l’aide du RT-PCR.
Resultats: Sur les cultures de MC3T3-E1 non transfectées, les niveaux d’expression d’ARN pour Noggin et Chordin étaient à leur plus haut niveau au Jour 7. Au Jour 7, les études au RT-PCR ont aussi démontré que les lentivirus contenant les shRNAs les plus efficaces avaient créé une diminution des niveaux endogènes d’expression d’ARN de Noggin et Chordin à 10 et 17% respectivement comparés au contrôle. Les analyses de Western Blot ont aussi confirmé que ces mêmes shRNAs avaient réussi une diminution significative des protéines de Noggin et Chordin. Aussi, l’activité de l’alkaline phosphatase était augmentée par les cultures exprimant de façon stable les shRNAs visant Noggin et Chordin.
Pour les études in vivo, toutes les chirurgies et les distraction furent réussies à l’exception de 2 morts suite à des complications chirurgicales. Toutefois, au moment de l’injection la majorité du contenu de l’injection However, at the time of the injection the majority of the injection ressortait du site d’injection. Une fois les échantillons collectés, les études Faxitron X-ray et µCT n’ont pas démontré de différence significative entre les 3 groupes dans le volume osseux ou le ration du volume osseux/volume tissulaire et ce, aux 2 temps de sacrifices. Finalement, les études en
4

immunohistochimie furent arrêtées après qu’un marquage au GFP fut retrouvé dans les 3 groupes.
Conclusion: Les analyses de Western blot analysis, qRT-PCR et les essais d’ ALP assay sont en accord avec un knockdown réussi de Noggin et Chordin lors de notre étude in vitro.
Lors de notre étude in vivo, les principaux problèmes rencontrés furent reliés à la technique de l’injection. Malheureuse, cette partie de notre étude n’a pas pu démontrer de différence significative dans la formation osseuse après l’utilisation d’injection de lentivirus contenant des shRNAs visant les antagonistes des BMPs Noggin et Chordin.
5

ACKNOWLEDGEMENTS
I would like to greatly thank my supervisors, Dr. Reggie Hamdy (Orthopedic Surgery) that took
me under his wing when I needed to take the next important step in my career and Dr Pierre
Moffatt ( Human Genetics ) for his constant support, guidance, and availability.
In addition, I would like to thank Dr. Abdallah Husseini for his help for the surgical procedures.
Thanks to the members of the McGill Bone Center for Micro-CT, Faxitron X-ray, and
biomechanical testing analysis.
My sincere gratitude to Bahar Kasaii with her help in coordinating all the experiments and
analyzing the results.
I would like to also thank the Shriners animal technicians, Mia, and Nathalie for their assistance
with distractions in the animal model.
Lastly, I would like to thank my parents, my brother and my fiance for their constant support and
help without whom I would never be able to reach all of my goals.
6

TABLE OF CONTENTS
ABSTRACT ................................................................................................................................. 2
RÉSUMÉ .......................................................................................................................................4
ACKNOWLEDGEMENTS ......................................................................................................... 6
TABLE OF CONTENTS .............................................................................................................. 7
LIST OF FIGURES ......................................................................................................................11
LIST of GRAPHICS......................................................................................................................13
LIST LIST OF TABLES .............................................................................................................. 14
LIST OF ABBREVIATIONS ...................................................................................................... 15
1-INTRODUCTION .................................................................................................................. 17
1.1-Distraction osteogenesis............................................................................................. 17
1.1.1-History..........................................................................................................19
1.1.2-Principle of tension-stress........................................................................... 19
1.1.3-Phases of distraction osteogenesis...............................................................20
1.1.4-Bone development during DO ................................................................... 22
1.1.5-Types of ossification in DO.........................................................................23
1.6-Complications and drawbacks of DO............................................................ 24
1.2-Attempts to accelerate................................................................................................ 25
1.2.1-Mechanical Stimulation ............................................................................. 26
1.2.2- Low-Intensity Pulse Ultrasound (LIPUS).................................................. 26
1.2.3-Transplantation of Bone Marrow Cells (BMCs)......................................... 28
1.2.4- Transplantation of Osteoblastlike cells ......................................................29
1.2.5- Demineralized Bone Matrix........................................................................31
7

1.2.6- Bone-graft substitutes..................................................................................31
1.2.7-Bisphosphonates ..........................................................................................33
1.2.8-Growth Factors............................................................................................ 34
1.2.8.1 - Growth Hormone Injection..........................................................34
1.2.9 - Cartilage Oligomeric Matrix Protein - angiopoietin-1 (COMP-Ang1)......34
1.2.10- Bone Morphogenetic Proteins(BMPS) .....................................................35
1.2.10.1 - BMPs Function......................................................................... 36
2.10.2- The BMP pathway........................................................................ 37
1.2.10.3 Use of exogenous BMPs during Distraction Osteogenesis..........40
1.2.10.3.1 Complications usage of exogenous BMPs....................41
1.2.11 -BMP antagonists....................................................................................... 42
1.2.11.1 Noggin.........................................................................................42
1.2.11.2 Chordin........................................................................................43
1.2.12 RNA interference (RNAi)...........................................................................44
1.2.12.1 shRNA.........................................................................................46
1.2.12.2 Delivery systems shRNA.............................................................48
1.2.12.2.1 Lentiviral vector............................................................48
2 - OBJECTIVE OF STUDY .................................................................................................... 50
3 -MATERIALS & METHODS IN VITRO STUDIES .......................................................... 51
3.1-shRNA selection .........................................................................................................51
3.2-Lentiviruses production.............................................................................................. 52
3.3- MC3T3-E1 transduction............................................................................................ 53
3.4-RT-PCR study............................................................................................................. 54
3.4.1 -RNA extraction from cell cultures............................................................. 54
8

3.4.2-RNA purification ........................................................................................54
3.4.3-RNA Quantification.................................................................................... 55
3.4.4-Reverse transcription of RNA......................................................................55
3.4.5- Real-Time PCR...........................................................................................55
3.5- Western Blot study.....................................................................................................56
3.5.1 - Proteins collection from cell culture and media........................................56
3.5.2 - Samples preparation..................................................................................57
3.5.3 - Western Blot..............................................................................................57
3.6 - Alkaline phosphatase assay......................................................................................58
4 - MATERIALS & METHODS IN VIVO STUDIES............................................................59
4.1 - Operative and distraction protocol...........................................................................60
4.2 - Injection....................................................................................................................63
4.2.1- Transfer of Noggin shRNA into lentivirus plasmid containing GFP..........63
4.2.2 - Injection protocol.......................................................................................64
4.3 - Faxitron X-ray and Micro-Computed Tomography..................................................64
4.4 - Immunohistochemistry..............................................................................................65
4.4.1- Paraffin embedding.....................................................................................65
4.4.2 - GFP marking..............................................................................................66
4.5- RT-PCR on tibia samples...........................................................................................66
4.5.1- Bone collection...........................................................................................66
4.5.2 - Tissue Homogenisation.............................................................................66
4.5.3 - RNA extraction, reverse transcription RNA and RT-PCR ........................67
4.5.4- RT-PCR : GFP and puromycin...................................................................67
9

5- IN VITRO RESULTS..............................................................................................................68
5.1 - RT-PCR results non-transfected MC3T3-E1.............................................................68
5.2 - RT-PCR transduced MC3T3-E1 cultures..................................................................69
5.3 - Western blot...............................................................................................................71
5.4 - Phosphatase alkaline assay........................................................................................74
6- IN VIVO RESULTS................................................................................................................76
6.1- Surgical and distraction protocol success rate...........................................................76
6.2 - Injection success rate................................................................................................76
6.3 - Faxitron X-Ray results.............................................................................................76
6.4 - Micro-Computed Tomography (µ-CT).....................................................................79
6.4.1 - Bone volume..............................................................................................79
6.4.2 - Bone volume/ Tissue volume ratio.............................................................81
6.5 - RT-PCR tibia samples...............................................................................................82
6.6-Immunohistochemistry - GFP marking.......................................................................83
7- DISCUSSION...........................................................................................................................85
8- REFERENCES ...................................................................................................................... 89
10

LIST OF FIGURES
Figure 1: Ilizarov external fixator on human patient
Figure 2: Phase of distraction osteogenesis.
Figure 3: BMP signaling pathway.
Figure 4: shRNA mediated RNA interference pathway
Figure 5: Position of shRNAs
Figure 7: Lentiviral plasmid
Figure 8: Mini Ilizarov external fixator
Figure 9: Mouse in DO
Figure 10: Final plasmid containing GFP after Noggin shRNA transfer
Figure 11 : Noggin - Western blot on secreted proteins in condition media of MC3T3-E1 stably expressing shRNAs targeting Noggin
Figure 12 : Noggin - Western blot on cellular proteins of MC3T3-E1 stably expressing shRNAs targeting Noggin
11

Figure 13: Chordin - Western blot secreted proteins in condition media of MC3T3-E1 stably expressing shRNAs targeting Chordin
Figure 14 : Puromycin and GFP signal in MC3T3-E1 cells stably expressing shRNAs targeting Noggin and Chordin
12

LIST OF GRAPHICS
Graph 1 : RT-PCR -MC3T3-E1 Noggin and Chordin mRNA level
Graph 2 : Noggin mRNA levels in stable pools MC3T3-E1 expressing individual shRNA through lentiviral infection
Graph 3 : Chordin mRNA levels in stable pools MC3T3-E1 expressing individual shRNA through lentiviral infection
Graph 4 : ALP activity in MC3T3-E1 stably expressing the shRNA targeting Noggin and Chordin
Graph 5 : Bone filling scores
Graph 6 : Bone volumes
Graph 7 : Bone volume / Tissue volume ratios
Graph 8 : RT-PCR tibia samples
13

LIST OF TABLES
Table 1 : Timeline animal procedures during in vivo studies
Table 2 : Bone filling scores
14

LIST OF ABBREVIATIONS
µCT Micro-computed tomography
ActR Activin receptor
ATF-4 Activating transcription factor-4
ALK Activin receptor-like kinase
ALP Alkaline phosphotase
BMC Bone marrow cell
BMP Bone morphogenetic protein
BMPR Bone morphogenetic protein receptor
BSA Bovine serum albumin
BV Bone volume
BV/TV Bone volume/tissue volume ratio
cDNA Complementary deoxyribonucleic acid
co-SMAD Common-partner mothers against decapentaplegic homolog
Col1a1 Collagen type I
DEPC Diethylpyrocarbonate
DO Distraction osteogenesis
DTT Dithiothreitol
EDTA Ethylenediaminetetraacetic acid
FGF Fibroblast growth factor
IGF Insulin-like growth factor
I-SMAD Inhibitory mothers against decapentaplegic homolog
LIPUS Low-intensity pulse ultrasound
mRNA Messenger ribonucleic acid NaCl Sodium chloride
15

PBS Phosphate buffer saline
PCR Polymerase chain reaction
PDGF Platelet-derived growth factor
PFA Paraformaldehyde
PRP Platelet-rich plasma
R-SMAD Receptor-regulated mothers against decapentaplegic homolog
RT-qPCR Real Time quantitative polymerase chain reaction
rhBMP Recombinant human bone morphogenetic protein
RNA Ribonucleic acid
SD Standard deviation
SMAD Mothers against decapentaplegic homolog
SMURF Smad ubiqutin regulatory factor
TBS-T Tri-phosphate buffer saline-tween TCA Trichloroacetic acid
TGF-ß Transforming growth factor-ß
TRIS Hydroxymethylaminomethane
µCT Micro-computed tomography
16

1- INTRODUCTION
1.1-Distraction osteogenesis
Distraction osteogenesis (DO) is a surgical procedure and treatment for lengthening of bone
adressing bony deficiencies secondary to congenital deformities, traumatic bone loss, infection,
malignancy and congenital limb discrepancies. Distraction osteogenesis is performed with a
transverse osteotomy. The fracture bone is then kept in position with an external fixator. During
the process the fractured sites are slowly pulled apart for a specific amount of time. The patients
bone regenerative properties will form new remodeled bone in the distraction gap. [1-2]
17

Figure 1 : Ilizarov external fixator on human patient
Schematic presentation of Ilizarov apparatus for human tibial distraction lengthening.
18

1.1.1-History
Long bone lengthening has been undertaken for more than 100 years, dating back to Codvilla
who performed femoral lengthening using traction after femoral osteotomy, using a pin placed in
calcaneus. [3]
Eventually, bone distraction evolved into different modern techniques. The Illizarov technique is
certainly one of the most well-known. In 1951, Dr. Gavriil Abramovitch Ilizarov developed the
surgical technique of distraction osteogenesis. (Figure 1) Ilizarov first had his idea of DO when a
patient mistakenly started turning the rods of his circular external fixator. Ilizarov noticed that
progressively, new bone had developed within the fractured gap. Intrigued by this biological
finding and its potential use in clinical treatment, Ilizarov began researching the technique using
animal models. In 1980, Ilizarov used DO to treated Carlo Mauri’s, an Italian explorer who was
suffering of pseudoarthrosis Mauri was very impressed with the results of his surgery and he set
out with Ilizarov to introduce this novel method of fracture treatment to the orthopedic
community. A few years after, DO gained public recognition and was commonly practiced. [4]
1.1.2-The Principle of Tension-Stress
The Principle of Tension-Stress was developed by Ilizarov and his team. This principle explains
the bone regeneration during DO. According to them, the slow and steady traction of a tissue
metabolically activates biosynthetic and proliferative pathways involved in bone regeneration
within the distracted zone. Many factors are involved in the bone quality that is developed at the
19

distraction site : the stability of the external fixator, the quality of tissues around the distraction
site and the rate and the frequency of distraction. [2-4]
1.1.3-Phases of Distraction Osteogenesis
Distraction osteogenesis is divided into three major phases: the latency phase, the distraction
phase and the consolidation phase. (Figure 2) The latency phase starts the first day post surgery
and lasts 5 to 7 days. Then the distraction is debuted and usually performed at a rate of 1mm/day
in humans . The act of distraction enhances angiogenic activity, activating intracellular pathways
at the distraction site. As distraction progresses, fibroblast-like cells grow parallel to the direction
of the tension-stress vector in the lengthened gap. The fibroblast-like cells produce collagen
fibers that condense into bundles throughout the callus. In addition, osteoblasts deposit osteoid
along the collagen fibers, beginning the consolidation phase of DO. During the consolidation
phase, the fixator is left in place until the bone acquires enough strength to withstand mechanical
stress of weight bearing. The patient is usually asked to keep on the Ilizarov device on for a
month for every centimeter distracted. During consolidation, the process of bone remodeling
continues developing trabecular and cortical bone within the callus. The remodeled bone within
the distracted gap usually contains the same properties as the host’s original bone. [2-4]
20

21
Figure 2 : Phases of Distraction osteogenesis
Radiological images of a wild-type mouse model of DO demonstrate the process of bone regeneration within the distracted gap. Distracted tibial samples were collected from the different phases of DO.
21

1.1.4-Bone Development during DO
There are two types of ossification that are essential for osteogenesis: endochondral and
intramembranous ossification. Intramembranous ossification is a one-stage process in which
mesenchymal cell-derived osteoblasts produce bone. Endochondral ossification is a two-step
process that uses cartilaginous intermediate to develop new bones. [5] Both methods of
ossification are initiated by mesenchymal stem cell condensation. The condensed mesenchymal
cells that take part in intramembranous ossification develop into preosteoblast cells. [6] These
preosteoblast cells further differentiate into mature osteoblasts to take part in the bone
remodeling process, forming woven bone. [7] Contrarily, the multistage process of endochondral
ossification is initiated with mesenchymal condensation and the production of type II collagen.
Mesenchymal cell-derived chondrocytes undergo proliferation and perichondrial cells start
expressing type I collagen. [8] The type II collagen-expressing, proliferating chondrocytes form
a cartilaginous template. Eventually, the chondrocytes stop proliferating and become pre-
hypertrophic. The pre- hypertrophic chondrocytes mature into hypertrophic chondrocytes that
express type X collagen and participate in matrix mineralization. Following maturation, the
hypertrophic chondrocytes undergo cell death by apoptosis [9]. Periosteal-derived osteoblasts
and newly formed blood vessels invade the cartilaginous template for trabecular bone formation
eventually forming the primary spongiosa [10]. With continual bone remodeling by osteoblasts
and osteoclasts, the center of the primary spongiosa is split into two epiphyseal growth plates
that are pushed apart in opposite directions.[9]
22

1.1.5 Types of ossification in DO
Kojimoto and colleagues completed DO on young Japanese rabbits, stabilized by a unilateral
dynamic fixator. Their results concluded that the principal bone development process was
endochondral ossification. Their histologic studies detected the presence of elongated
cartilaginous cells and hypertrophic chondrocytes in the callus [11].
Delloye and colleagues performed DO on adult mongrel dogs using a circular external fixation
system. Intramembranous ossification was identified in their samples analysis. [12] Similarly,
the same conclusion was found by the group of Arnonson and colleagues. They performed DO
on a canine model using two types of fixators, the Ilizarov fixator and the Wagner fixator, and
also noticed intramembranous ossification was the only mechanism used for bone formation
during DO.[13]
However, Rauch and colleagues performed DO on New Zealand rabbits. The osteotomy was
completed on the right tibia of the rabbits and stabilized using Orthofix uniplanar fixators. They
found both types of ossification in their model.[14]
A group of Japanese researchers detected the presence of three types of ossification used within a
rat model of DO: endochondral, intramembranous and transchondroid ossification. Distraction
osteogenesis was performed on adult rats, in which bony fragments were supported by a
monolaternal external fixator. Histological findings showed the formation of a fibrocartilaginous-
based callus that became hypertrophic during early distraction, indicating endochondral
ossification was used during this time point of DO. By 10 to 20 days of distraction, histological
images revealed the presence of preosteoblasts, osteoblasts and fibroblast-like cells within the
23

cartilaginous callus. Eventually, the cartilaginous callus was resorbed and replaced with new
bone, demonstrating that intramembranous ossification was used during the late distraction phase
of DO. In between these two phases of bone development, transchondroid ossification was
observed during the mid-distraction phase of DO. During transchondroid bone formation,
chondrocyte-like cells produced chondroid bone, an intermediate type of tissue that contained
properties of both cartilage and bone. The chondroid matrix resembled bone matrix more than a
cartilaginous matrix. Furthermore, round chondrocyte-like cells and smaller osteocyte-like cells
were present in the histological sections of transchondroid bone. [15]
Finally, if we come back to the Principle of Tension and Stress, Ilizarov observed that type of
bone formation that occurred within a distracted gap depends on the stability of the external
fixation system. A stable fixation system ensures that the surrounding bone marrow and
periosteal tissues around the fracture site are intact, allowing bone regeneration to occur without
the development of cartilaginous intermediate. Thus, Ilizarov concluded that intramembranous
ossification is the main method of bone development used within a healthy callus of DO [2-4].
Contrarily, a cartilaginous callus may form within the distracted site via endochondral
ossification when there is an unstable fixation system. A cartilaginous callus is usually formed to
stabilize and support the fractured segments within the distracted gap.[16]
1.1.6-Complications and Drawbacks of DO
Like any surgery, DO comes with risks for different complications. The regular complications
related to the anesthesia can always apply. Then during the surgery itself, risks of bleeding,
injury to major vessels of nerves are always possible. In the post-operative period, the risk of
24

infections is always possible and complications related to the use of hardware occasionally
happened. Post-consolidation, there may be a possibility that the bone can refracture in the
distracted site or a different site during the removal of the external fixator. [17]
The main drawback associated with DO is the long period of time patients are required to keep
on the external fixation system on the affected site. [17] For every centimeter the bone is
lengthened, the external fixator has to been kept on for one month (i.e. if a patient requires ten
centimeters of lengthening, the fixator must kept on for ten months). This drawback can cause
social, psychological and medical problems for the patient and also lead to problems with patient
compliance. Thus, researchers are studying methods to accelerate the consolidation phase of DO.
1.2-Attempts to Accelerate DO
Many studies have been conducted using animal models to analyze the effects of physical and
molecular methods that can potentially accelerate the consolidation phase of DO. [17] Physical
methods have included the use of mechanical stimulation [18-19] and the application of low-
intensity pulse ultrasound (LIPUS) [20 to 24]. Whereas, biological methods have included the
transplantation of bone marrow [25] and osteoblastlike cells into the fracture site [26-27], the
local injection of bisphosphonates [28-29], and peptide growth factors.[30]
25

1.2.1-Mechanical Stimulation
There have been few attempts using mechanical stimulation, such as compressive stimulation, to
increase the consolidation phase of DO. The application of compressive stimulation during the
distraction phase of mandibular DO demonstrated no significant changes in bone regeneration
[31]. Kim et al. studied the effects of compressive stimulation during early consolidation in a rat
model of mandibular DO. DO was performed on the mandible of rats followed by a 3 day
latency period, a 4 day distraction period (a distraction rate of 0.25 mm/day twice daily) and a 7
day consolidation period. A compressive force was applied in the direction opposite to distraction
at the compression rate of 0.25 mm/day twice daily for the first four days of consolidation.
Therefore, after a total of 4.0 mm of distraction and 2.0 mm of compression, the total distraction
length was 2.0 mm. Controls were subjected to the same conditions of distraction in the absence
of compression. Histological and radiological data demonstrated the presence of a softer callus
that had primarily developed via intramembranous ossification in the experimental rat groups
compared to the control group during early consolidation. By late consolidation, the compressive
force helped decrease the size of the distracted gap, forming a thicker volume compact bone
regenerate. Thus, the use of compressive stimulation during the consolidation phase of
mandibular DO possibly contributed to the formation of more mature and dense bone by
accelerating the differentiation of osteoprogenitor cells to osteogenic cells.[19]
1.2.2- Low-Intensity Pulse Ultrasound (LIPUS)
Low-intensity pulsed ultrasound or (LIPUS) has also been studied for the acceleration of bone
healing during DO. LIPUS is a form of mechanical energy that travels to the site of bone repair
26

as pressure waves. At the fracture site, LIPUS enhances the process of angiogenesis, facilitating
the exchange of nutrients and waste removal among cells and stimulating the differentiation and
proliferation of fibroblasts, chondroblasts and osteoblasts. However, the exact mechanism of
LIPUS interaction with the surrounding soft tissue environment of the fracture site still remains
poorly understood. [20]
Sakurakichi and colleagues studied the application of LIPUS during the distraction phase of DO.
Distraction osteogenesis was performed on the right tibia of white Japanese rabbits using
unilateral external fixators (7 day latency period, 7 day distraction period using a distraction rate
of 1.5 mm/day and 7 day consolidation period). LIPUS was applied to the fracture site of rabbits
using an ultrasound transducer at 200µs burst of 1.5 MHz sine waves at a frequency of 1.0 kHz
for 20 minutes post- distraction each day (the same conditions are used in humans). The data
showed that application of LIPUS during the distraction phase of DO resulted in early bone
mineralization with high trabecular bone formation and increased mechanical strength of the
distracted rabbit tibia. [21] Similar results were observed in another rat model with unilateral
femoral lengthening and LIPUS application during the consolidation phase by Eberson group.
[22]
The application of LIPUS was also studied during the consolidation period. The Shimazaki’s
group performed DO using a unilateral fixator on the right tibia of rabbits (white Japanese). The
project was separated in this fashion : latency period (7 days), distraction (10 days) and
consolidation (21 days). The LIPUS was applied using the same conditions used in humans. The
27

results showed that the LIPUS treatment group compared to the controls had an increase in bone
mineral density and hard callus formation. [23] Another study also examining the effect of
LIPUS on consolidation came to similar conclusion. Claes and colleagues also analyzed the
outcomes of LIPUS stimulation on a sheep model of DO. Using a custom-made fixation system,
DO was performed on the metatarsus of the animals in this fashion : 4 days of latency, 16 days of
distraction and 64 days of consolidation. The sheep were stimulated with LIPUS for 20 minutes
each day starting the first day of consolidation. Results demonstrated a greater axial stiffness and
early callus formation in the LIPUS-treated sheep. In addition, a 32% increase in bone formation
was observed in the LIPUS-treated sheep compared to the controls that was predominantly
formed via endochondral ossification. This process of callus development in the LIPUS-treated
sheep is quite different from the LIPUS untreated controls that primarily form bone via
intramembranous bone formation. Thus, LIPUS treatment may enhance endochondral bone
formation within the distracted gap. [24]
1.2.3 Transplantation of Bone Marrow Cells (BMCs)
Transplantation of bone marrow cells (BMCs) has been studied in the recent years. They were
mainly used to shorten the consolidation period. The animal studies were created around the idea
that the transplantation of multipotent BMCs during DO would develop into osteogenic
progenitors may enhance bone formation within the fracture site. The advantages of BMC
transplantation include the easy isolation of these cells from bone marrow sections and it is a safe
treatment with relatively few side effects. [25] The biggest challenge with the use of BMC
28

transplantation is to find an appropriate delivery system at the distraction site. Thus, the choice of
osteoinductive factors and type of scaffold are important for BMC transplantation during DO.
Kitoh and colleagues discovered that the combination of culture expanded BMCs in platelet-rich
plasma (PRP) stimulated bone regeneration in patients that had undergone tibial or femoral
lengthening. PRP supplied osteoinductive factors, such as platelet-derived growth factor (PDGF),
insulin-like growth factor-1(IGF-1) and transforming growth factors (TGF-ßs) that are necessary
for bone regeneration. Furthermore, the use of a biodegradable scaffold helped prevent infection
and inflammation of the fracture site.[32]
Kitoh’s study measured a standardized healing index for each patient. This healing index was
calculated by the division of the time required for complete consolidation by the total distracted
length observed by x-ray analysis. BMC-PRP-treated patients had a smaller healing index than
untreated patients, demonstrating a decrease in the consolidation time for DO. Furthermore, a
delay in consolidation was observed in 45% of untreated BMC-PRP patients compared to the
BMC-PRP-treated patients that exhibited normal bone healing patterns. [32]
1.2.4- Transplantation of Osteoblast-like cells
Transplantation of periosteum-derived osteoblast-like cells were also studied during DO. Tsubota
and colleagues studied the effects this technique in a white Japanese rabbits using a hemilateral
external fixator. The treatment groups were divided in 3 : group 1- a control group that
underwent distraction without the transplantation of osteoblastlike cells, group 2- a control group
of rabbits that were only injected with physiological saline and group 3-test group of rabbits that
29

were injected with periosteum-derived osteoblast-like cells dissolved in physiogical saline. The
rabbits underwent 7 day latency period, 20 day distraction period with a distraction rate of 1mm/
day and sacrifices were made every 2 weeks during consolidation. Results showed that group 3
rabbits contained larger and stronger callus formation than the controls as measured by a high
transaxial area ratio and three point-bending analysis. Furthermore, the bone mineral density of
the treatment group was significantly higher than the control groups at all time points. [26]
Tamakine et al. studied in 2002 the use of bone marrow-derived osteoblast-like cells using a
collagen-based scaffold in a rat model. DO was performed on the left femur of rats and stabilized
using a unilateral fixator. Post-surgery, rats were subjected to a 7 day latency period and a 10 day
distraction period with a distraction rate of 0.25 mm every 12 hours. The treatment group was
receiving an injection of osteoblast-like cells in collagen gel in the distraction zone. The control
groups injected with : physiological saline, collagen gel or bone marrow derived osteoblast-like
cells without the gel vector. The results showed that the rats from the treatment group had larger
callus formation compared to the controls. The group of rats that were treated with bone marrow-
derived osteoblast-like cells in gel also contained an increase in fracture strength two, four and
six weeks post- transplantation as determined by three-point bending analysis. Therefore, both
studies demonstrated that the transplantation of osteoblast-like cells enhanced bone formation in
the rabbit and rat models of DO. It is possible that the transplantation of the osteoblast-like cells
population into the distracted zone increased the number of osteogenic cells and recruited
osteoinductive factors to the fracture site that are necessary for bone regeneration, that may
account for the large callus formation [27]
30

1.2.5-Demineralized Bone Matrix
Demineralized bone matrix has been used in the past for the treatment of non-unions [33], spinal
fusions [34], craniomaxillofacial defects[35-36], and segmental bone defects [37-39]. The first
ones to investigate the effect of demineralized bone matrix during DO were Hagino et al. in 1999
[40]. Their study was constructed around the use of grafting demineralized bone matrix during
lengthening of the tibia in young Japanese White rabbits. The demineralized bone matrix was
made from frozen cortical bone harvested from other rabbits. The control group was not
receiving any graft during the process. Their study was able to show a significant difference in
the formation of a radiographic callus and in the rate of non-union favoring the grafted group.
They also tried to accelerate their distraction rate up to 2-3 mm/day and still got acceptable
results. Thus, they demonstrated that the use of demineralized bone matrix can allow for
satisfactory bone formation at faster rates than normal.
1.2.6- Bone-graft substitutes
Synthetic or natural biomaterials have been used as bone-graft substitutes to autologous or
allogeneic bone grafts. Although they lack osteoinductive or osteogenic properties, synthetic
bone substitutes and biomaterials are already widely used in clinical practice for
osteoconduction. They consist of scaffolds that promote the migration, proliferation and
differentiation of bone cells for bone regeneration. A wide range of synthetic bone substitutes
are currently used. The main ones currently used being collagen, hydroxyapatite (HA), β-
tricalcium phosphate (β-TCP), calcium-phosphate cements, and glass ceramics [41,42].
31

Especially for reconstruction of large bone defects, for which there is a need for a substantial
structural scaffold, an alternative to massive cortical auto or allografts is the use of cylindrical
metallic or titanium mesh cages as a scaffold combined with cancellous bone allograft,
demineralized bone matrix or autologous bone [43,44]. Furthermore, there are also non-
biological osteoconductive substrates, such as fabricated biocompatible metals (for example,
porous tantalum) that offer the potential for absolute control of the final structure without any
immunogenicity [41].
Wang and al. showed in 2008 in a study comparing 3 groups of rabbits with a 1.0 cm bone defect
on their left tibia. Group A, the defect gap was reduced with the tibia shortened for 1.0-cm and
received lengthening for 10 days. Group B, the defect gap was filled with 1.0-cm restorable
porous hydroxyapatite and Tri-calcium phosphates cylindrical block (HA/TCP block, diameter is
0.5-cm) and did not receive lengthening. Group C, The 1.0-cm defect gap was reduced 0.5 cm
and the remaining 0.5-cm defect gap was filled with the 0.5-cm HA/TCP block and received
lengthening for 5 days. The bone specimens were subject to microcomputed tomography,
mechanical testing, and histological examinations. Their results showed that a combined use of
biomaterials and DO technique can reduce the treatment time and enhance bone consolidation in
bone defect management. [45]
Research is ongoing to improve the mechanical properties and biocompatibility of scaffolds, to
promote osteoblast adhesion, growth and differentiation, and finally to allow vascular ingrowth
and bone-tissue formation. Improved biodegradable and bioactive three-dimensional porous
scaffolds [46] are being investigated, as well as new technologies using nanotechnology, such as
magnetic biohybrid porous scaffolds acting as a crosslinking agent for collagen for bone
32

regeneration guided by an external magnetic field [47], or injectable scaffolds which provide
easier application [48].
1.2.7-Bisphosphonates
In theory the biphosphonates act by decreasing the catabolic activity during DO. Takahashi and
colleagues studied the effects of administering nitrogen-containing bisphosphonates on a rabbit
model of DO. Rabbits underwent tibial lengthening with a 5 day latency period, 21 day
distraction period (distraction rate of 0.35mm every 12 hours) and a consolidation phase of four
weeks. Rabbits were injected with 0.4 mg/kg of nitrogen-containing bisphosphonates for a period
of 6 weeks post-osteotomy. Controls were injected with vehicle only. Results showed that the
bisphosphonate-treated rabbits contained new bone with greater mechanical strength in the
osteopenic zones including 5.6x greater bone volume and 3.3x greater mechanical resistance than
the controls.[29]
33

1.2.8-Growth Factors
The use of different growth factors may potentially be the most resourceful technique for
accelerated bone healing during DO. Different growth factors have been analyzed in fracture
healing studies including Growth Hormone (GH) Transforming growth factor ß (TGF- ß),
Platelet-derived growth factor (PDGF), Insulin-like growth factors (IGF), Fibroblast growth
factor (FGF) and Bone morphogenetic proteins (BMP). [30]
1.2.8.1-Growth Hormone Injection
Growth hormone injections was studied by Bail et al. in 2002. They applied this treatment to the
process of mandibular DO. They demonstrated that systemic GH administration significantly
increases the torsional stability of the regenerate in comparison to the contralateral side with no
exogenous GH administration and showed a larger callus formation. [49]
1.2.9-Cartilage Oligomeric Matrix Protein - angiopoietin-1
(COMP-Ang1)
COMP-Ang1 is an angiogenic factor that has been studied in DO. Park et al. in 2010 investigated
the effect of local COMP-Ang1 administration in the center of the distraction zone [50].Control
rats underwent tibial distraction without treatment. In the two remaining groups, BSA (100 ug) or
COMP-Ang1 (100 ug) were injected transcutaneously into the center of the distraction zone.
34

They demonstrated that bone formation in the distraction gap progressed more quickly in the
COMP–Ang1-treated group than in the BSA-treated group. Also, histological findings and
immunostaining of endothelial cells for factor VIII revealed that the COMP–Ang1 group of
animals exhibited higher levels of vascularity. NanoCT and dual-energy X-ray absorptiometry
analysis also revealed increased new bone formation along capillaries in the COMP–Ang1 group
compared with the BSA group show an increase. Runt-related transcription factor 2 and its target
genes, including bone sialoprotein, type 1 collagen, osteopontin, and osterix were significantly
upregulated in the COMP–Ang1 group. Their results suggested that COMP–Ang1 could be
potentially used as a therapeutic agent for treatment of distracted limbs by enhancing
angiogenesis [50].
1.2.10-Bone Morphogenetic Proteins (BMPs)
BMPs had been involved in the regulation of cell proliferation, survival, differentiation and
apoptosis. They play a pivotal role in inducing bone, cartilage, ligament, and tendon formation at
both heterotopic and orthotopic sites. 91 BMPs were first identified by Dr. Marshall Urist in
1965 when he noticed at implantation of demineralized bone matrix in different sites of a rodent
model induced ectopic bone formation.[50] Twenty BMPs have been identified to date and
except for BMP1, all BMPs are a part of the transforming growth factor ß (TGF- ß) superfamily.
[51]
BMPs are known to be potent inducers of osteogenesis during embryological bone formation,
and fracture healing. These proteins act through serine-threonine kinase transmembrane
35

receptors, type I and type II [52-53]. Their activities are mediated through Smads signaling
pathways.
1.2.10.1-BMPs Function
BMPs have been implicated in a variety of functions including bone and cartilage generation and
a number of non-osteogenic roles.
BMPs have also been shown to be important regulators of chondrogenesis and skeletogenesis
during normal embryological development.
BMP-2, -4, -6, -7, and -9 have the greatest osteogenic capacity. BMP-2 has been demonstrated to
be expressed strongly around cartilage condensations, periosteal and osteogenic zones. BMP-4
on the other hand is expressed in the perichondrium and BMP-6 is expressed in hypertrophic
chondrocytes. BMP-7 has been observed to be expressed highly in the perichondrium with no
expression in the zones of joint formation [51].
Both BMP-2 and -7 have been implicated to have active critical roles in the expression of major
transcription factors during osteogenesis such as Runt-related transcription factor 2 (Runx2 ) and
Osterix (Osx) in mesenchymal stem cells. BMP-7 knock-outs exhibit skeletal alterations in the
rib cage, hind limbs and skull. [54]
BMPs appear to play an important role in regulating stem cells differentiation. In embryonic
stem cells, BMP signaling appears to be required for self-renewal. In mesenchymal stem cells,
36

BMP signaling stimulates osteoblastic differentiation. However, it has been suggested that due to
the pleiotropic functions of BMPs, it is possible for the signaling to be mutagenic or oncogenic.
In particular, BMP-7 located at 20q13, has been demonstrated to play a key role in osteoblast
differentiation. [51]
1.2.10.2-BMP Pathway
BMP ligands bind to two distinct receptors: type I and type II serine/threonine kinase receptors.
Both of these receptors are required for signaling [119]. There are three type I receptors: type IA
and IB BMP receptors (BMPIA or ALK3 and BMPIB or ALK6), type IA activin receptor
(ActRIA or ALK2). Type II receptors also exist in three forms: type II BMP receptor (BMPR-II),
type II and IIB activin receptors (ActR-II and ActR- IIB). [55]
BMP ligands bind to type II receptors. The serine/threonine kinase domains of type II receptors
are constitutively active and phosphorylate Gly-Ser [50] domains of type I receptors upon ligand
binding, leading to activation of type I receptor kinases [55]. This leads to the recruitment of
pathway restricted Smads (R-Smads, Smads1, 5 or 8) and later recruitment of the common
mediator Smad (Co-Smad, Smad 4) into the complex. This complex migrates into the nucleus
and activates the transcription of specific target genes [57].
The signaling of BMP is modulated at different steps along the pathway. In the extracellular
environment, antagonists such as Cerbarus, Dan, Gremlin, Noggin, and Inhibin which are known
to bind specifically to BMP receptors can regulate initiation of the cascade. Additionally at the
37

cell surface, the oligomerization mode of the receptors can activate the signaling pathway
specifically [51].
Once the signal is transduced into the intracellular compartment, the signal can be modulated by
the activation of inhibitory Smad proteins (I-Smad, Smad 6, 7) or Smurf 1. In the nucleus, there
are a number of co-activators needed for the activation of specific target genes and their
transcription can be inhibited by co-repressors [51, 58].
38

nal induces osteoblastic di!erentiation through Bmpr1b butinhibits osteoblastic di!erentiation through Bmpr1a [8]; inDrosophila germline stem cells (GSCs), Dpp (homolog ofBMP2/4) is essential for the maintenance of stem cells [9].
BMPs may have pleiotropic functions including mutagene-sis or oncogenesis, thus, extensive studies have beenreported in the field of musculoskeletal oncology, especiallyosteosaroma [10]. Urist et al. first reported that certain via-ble and devitalized human osteosarcomas induced ectopicbone formation in athymic nude mice, suggesting thathuman osteosarcomas may produce BMP-like proteins. Insubsequently studies, BMP-2/4 proteins or genes in a vari-ety of bone tumors have been studied by immunohisto-chemistry, in situ hybridization, or both [11]. BMPscontrol many key steps in the formation and di!erentiationof the vertebrate nervous system, which act at di!erentstages of neural development and in di!erent regions ofthe CNS to regulate cell fate, proliferation and di!erentia-tion [12]. BMP-2, BMP-4, and BMP-6 have been localizedto areas of vascular calcification. Subsequently, studiesshowed that BMP-2 inhibiting vascular smooth muscle cellsproliferation when stimulated with serum or growth factorsin vitro [13]. During embryonic development, BMPs medi-ate programmed cell death, or apoptosis, the process thatremoves unnecessary tissues, thereby ensuring proper mor-phogenesis. BMP-4 was first demonstrated to induce apop-tosis in rhombomeric neural crest cells [14]. The function ofBMPs family members is shown in Table 1.
Receptors and molecular signaling
Members of the BMP family bind to two distinct type IIand type I serine/threonine kinase receptors, both of whichare required for signal transduction [15]. There are threetype II receptors have been shown to bind BMP ligands:type II BMP receptor (BMPR-II), type II and IIB activinreceptors (ActR-II and ActR-IIB), and three type I recep-tors for BMPs have also been characterized: type IA andIB BMP receptors (BMPIA or ALK3 and BMPIB orALK6), type IA activin receptor (ActRIA or ALK2)
Table 1The list of BMP family members
BMP Gene locus Function Receptors and inhibitors
BMP-1 8p21 It is a metalloprotease that acts on procollagen I, II, and III. It isinvolved in cartilage development
Type I receptors
BMP-2 20pl2 Acts as a disulfide-linked homodimer and induces bone and cartilageformation. It is a candidate as a retinoid mediator. Plays a key role inosteoblast di!erentiation
ACVRIA, ACVRIB, ACVRIC,ACVRLI-BMPR (BMPRIA–BMPRIB)
BMP-3 14p22 Induces bone formation Type II receptorsBMP-4 14q22–q23 Regulates the formation of teeth, limbs and bone from mesoderm. It
also plays a role in fracture repairACVR2A, ACVR2B, AMHR2,BMPR2
BMP-5 6p12.1 Performs functions in cartilage developmentBMP-6 6p12.1 Plays a key role in joint integrity in adults Type III receptorsBMP-7 20q13 Plays a key role in osteoblast di!erentiation. It also induces the
production of SMADI. Also key in renal development and repairBetaglycan
BMP-8 1p35–p32 Involved in bone and cartilage developmentBMP-9 Promoting chondrogenic di!erentiation of human multipotential
mesenchymal cellsLigand inhibitors
BMP-10 2p14 Play a role in the trabeculation of the embryonic heart Cerberus, Chordin, DAN, Decorin,Follistatin, Gremlin, Lefty, LTBPI
BMP-15 Xp11.2 May play a role in oocyte and follicular development Noggin, THBSI
In this table, it shows the function (if known), gene locus, receptors and ligand inhibitors.
Fig. 2. Bone morphogenetic protein (BMP) receptors mediate BMPsignaling through activating Smad. BBMP ligands bind to the BMPreceptors BMPRI and BMPRII. After BMPRII phosphorylated, follow-ing activates BMPRI. Phosphorylated BMPRI subsequently phosphory-lates receptor-activated Smad proteins (R-Smads), which associate withcommon mediator-Smad (co-Smad) and enter the nucleus, where theyregulate gene expression. The Smad proteins regulate promoter activity byinteracting with transcriptional co-activators or co-repressors to positivelyor negatively control gene expression. The BMP signal can be blocked byextracellular antagonists, such as noggin, which bind BMP ligands andprevent their association with the BMP receptors, as well as byintracellular proteins, such as inhibitory Smads (I-Smads), which preventthe association between R-Smads and co-Smad and Smurf 1. Smurf1 is aHect domain E3 ubiquitin ligase. It interacts with Smad1 and 5 andmediates the degradation of these and also forms a complex with Smad6,is exported from the nucleus and targeted to the type I BMP receptors fortheir degradation [21]. Modified from Niswander et al.
552 Y.-T. Xiao et al. / Biochemical and Biophysical Research Communications 362 (2007) 550–553
Figure 3 : BMP pathway
Bone morphogenetic protein (BMP) receptors mediate BMP signaling through activating Smad. BBMP ligands bind to the BMP receptors BMPRI and BMPRII. After BMPRII phosphorylated, following activates BMPRI. Phosphorylated BMPRI (phosphorylation expressed has p-bodies in figure) subsequently phosphorylates receptor-activated Smad proteins (R-Smads), which associate with common mediator-Smad (co-Smad) and enter the nucleus, where they regulate gene expression. The Smad proteins regulate promoter activity by interacting with transcriptional co-activators or co-repressors to positively or negatively control gene expression. The BMP signal can be blocked by extracellular antagonists, such as noggin, which bind BMP ligands and prevent their association with the BMP receptors, as well as by intracellular proteins, such as inhibitory Smads (I-Smads), which prevent the association between R-Smads and co-Smad and Smurf 1. Smurf1 is a Hect domain E3 ubiquitin ligase. It interacts with Smad1 and 5 and mediates the degradation of these and also forms a complex with Smad6, is exported from the nucleus and targeted to the type I BMP receptors for their degradation [62] . Modified from Niswander et al. [51]
39

1.2.10.3 Use of exogenous BMPs during Distraction Osteogenesis
Usage of exogenous BMPs during distraction osteogenesis have been tested as a treatment to
accelerate the process of DO. Mizumoto et al. have studied in 2003 the use of recombinant
human BMP-7 during DO in a rat model. The rats were divided in 2 groups : one group was
treated with rhBMP-7 in an aquaeous solvent and the other group just received the solvent
(control) through an injection in the osteotomy site. Their results showed an increase in bone-
mineral density and biomechanical testing results in the treatment group.[60] Yang and al studied
in 2012 the use of BMP-2 during DO through a tri-calcium phosphate (TCP)/hydroxyapatite
(HA) delivery system in a rat model. The animals were randomized into three groups: group I
served as a control, group II was treated with only TCP/HA, and group III was treated with
recombinant human (rh) BMP-2-coated TCP/HA. Materials were inserted into the medullary
canal at the femoral osteotomy site at the end of the lengthening period. Their results showed that
application of rhBMP-2, at the end of the rather rapid distraction period, as a single bolus
significantly increased the osteogenic process, while beta-TCP/HA behaved effectively as a
sustained delivery system for this osteoinductive protein. [61]
Studies investigating the clinical effects of BMP administration in human long bone fractures
have also been numerous during the past decade. A Cochrane review was published in 2010
studying eleven randomized control trials. Their analysis showed that apart from one study, the
times to fracture healing were comparable between the BMP and control groups. There was some
evidence for increased healing rates, without requiring a secondary procedure, of BMP compared
40

with usual care control in acute, mainly open, tibial fractures (risk ratio (RR) 1.19, 95% CI 0.99
to 1.43). The pooled RR for achieving union for nonunited fractures was 1.02 (95% CI 0.90 to
1.15). One study found no difference in union for patients who had corrective osteotomy for
radial malunions. Data from three RCTs indicated that fewer secondary procedures were required
for acute fracture patients treated with BMP versus controls (RR 0.65, 95% CI 0.50 to 0.83).
[63]
1.2.10.3.1 Complications usage of exogenous BMPs
Some controversies linger around the usage of BMPs. Even if some studies have demonstrated
that therapy with rhBMP-7 in non-unions is significantly more cost-effective than traditional
treatment modalities [65], most of the recent systematic reviews come with the opposite
conclusion. [63-68] Even if multiple studies have also demonstrated that BMP-2 and -7 are
generally safe, effective, and at times superior alternatives to autologous bone grafting during
interbody spinal fusion procedures [65-66], some animal studies have reported some
complications. Recently Zara and Al. studied the use of high doses of BMPs-2 in a rat model.
Their results showed that they consistently reproduced BMP2 side effects of cyst-like bone and
soft tissue swelling using high BMP2 concentration approaching 1500 µg/mL. [67]
41

1.2.11 BMP Antagonists
BMP antagonists have been shown to modulate BMP activities in diverse and critical ways in a
myriad of tissues and systems during normal vertebrate development. [69]
There are three main types of BMP antagonists: pro- region inhibitors, ligand antagonists, and
receptor antagonists [70].
Pro-region inhibitors interact with the mature peptide [70]. Recent studies with BMP-2 and -9
have shown that the pro-region inhibitors are inefficient inhibitors of osteogenesis as they can
easily get degraded by proteolytic cleavage [71-72].
The second group of inhibitors is the ligand antagonists such as Noggin, Chordin, Follistatin,
Gremlin, and Sclerosin that bind directly to the BMP ligand and prevent the interaction of the
complex with the receptor.
Lastly, the receptor antagonists are the group of inhibitors that interact with the BMP receptors
and prevent the BMP-receptor interaction. Additionally, inhibitory Smads (Smad 6 and Smad 7)
are the BMP antagonists that interfere with BMP activity at intracellular level [70].
1.2.11.1 Noggin
Noggin is a potent inhibitor of BMP activity that directly binds BMP in the extracellular
environment. Noggin binds with differing affinities to BMP-2, -4, -5, -6, and -7, with the highest
affinities observed between noggin and BMP-2 and -4 and the relatively weakest affinity
42

between noggin and BMP-7 [73-74]. Noggin is essential for normal skeletal and joint
development [75-76]. A single dose of noggin in rats significantly decreases the rate of
membranous ossification as measured by a bone in-growth chamber technique. Interestingly, the
total amount of tissue in-growth was similar between controls and the noggin groups, but
ossification was significantly reduced in the noggin group [73].
Noggin expression is clearly upregulated by BMP as a protective feedback mechanism to prevent
bone overgrowth in BMP stimulated osteoblasts [77].
The laboratory of Wan et al. [78] realized in vitro and in vivo studies using siRNA to silence
noggin activity . Pre-osteoblasts were infected with specific siRNA constructs that were shown
to effectively silence noggin expression. In these infected cells, expression of Smad 1 and 5, and
rate of osteoblast differentiation were all significantly increased compared to sham infected
controls. Expression of osteoblastic differentiation markers Runx2, osteocalcin, and osteopontin
all peaked approximately three days earlier in the siRNA infected cells compared to sham
infected controls. For their in vivo studies, defects were created in mouse calvaria and the
cortical defects were seeded with either siRNA infected osteoblasts or sham infected osteoblasts.
At two and four weeks, the defects seeded with siRNA infected osteoblasts demonstrated
significantly more bone formation. Interestingly, at eight weeks there were similar levels of bone
formation with no appreciable difference between siRNA infected and sham infected control
groups.
Their results were suggesting that it may be possible to manipulate noggin modulation of BMP
activity in such a way to enhance and speed bone regeneration and, potentially, fracture healing.
43

Studies examining the effects of noggin silencing on long bone fracture healing have yet to be
performed, but are necessary to further the possibility of therapeutic algorithms utilizing the
manipulation of noggin. Noggin expression peaks in the callus 10 days post fracture [79].
1.2.11.2 Chordin
Chordin appears to bind BMP-2, BMP-4, and BMP-7 in the extracellular environment similar to
noggin and thus prevents BMP activity by preventing it from binding to receptors [80-83].
Chordin is very important in embryological development, especially skeletogenesis [81-83].
Zhang et al found in 2002 that chordin expression during development was shown to be
inversely related to the stages of chondrocyte maturation, indicating that chordin is likely an
inhibitor in this process [81]. Then Nilsson et al. found that chordin expression is significantly
lower in the proliferative and hypertrophic zones versus the resting zone in 7-day old rat tibial
physes. [84] Despite conclusive evidence that chordin is an inhibitor of BMP during normal
development, few studies to date have specifically investigated whether chordin plays any role
during fracture healing. One investigation demonstrated a peak of chordin expression within
callus 10 days after fracture in normally healing fractures and a consistent level of chordin
expression in experimentally produced non-unions [79].
44

1.2.12 RNA interference (RNAi)
RNA interference (RNAi), an accurate and potent gene-silencing method, was first
experimentally documented in 1998 in Caenorhabditis elegans by Fire et al., who subsequently
were awarded the 2006 Nobel Prize in Physiology/Medicine. [85]
RNAi is a natural process in which the expression of a targeted gene can be knocked down with
a good specificity and selectivity. Methods of mediating the RNAi effect involve small
interfering RNA (siRNA), short hairpin RNA (shRNA) and bi-functional shRNA. The simplicity
of siRNA manufacturing and transient nature of the effect per dose are optimally suited for
certain medical disorders (i.e. viral injections). However, using the endogenous processing
machinery, optimized shRNA constructs allow for high potency and sustainable effects using low
copy numbers resulting in less off-target effects. Bi-functional design may further enhance
potency and safety of RNAi-based therapeutics. [86]
45

1.2.12.1 shRNA
shRNAs are synthesized in the nucleus of cells, further processed and transported to the
cytoplasm, and then incorporated into the RISC for activity [87]. To be effective, the shRNA are
designed to follow the rules predicated by the specifics of the cellular machinery.
shRNA can be transcribed by either RNA polymerase II or III through RNA polymerase II or III
promoters on the expression cassette. The primary transcript generated from RNA polymerase II
promoter contains a hairpin like stem-loop structure that is processed in the nucleus by a
complex containing the RNase III enzyme Drosha and the double-stranded RNA-binding domain
protein DGCR8 [88]. The complex measures the hairpin and allows precise processing of the
long primary transcripts into individual shRNAs with a 2 nt 3′ overhang [89]. The processed
primary transcript is the pre-shRNA molecule. It is transported to the cytoplasm by exportin 5, a
Ran-GTP- dependent mechanism [90-91]. In the cytoplasm the pre-shRNA is loaded onto
another RNase III complex containing the RNase III enzyme Dicer and TRBP/PACT where the
loop of the hairpin is processed off to form a double-stranded siRNA with 2 nt 3′ overhangs
[92-94]. The Dicer containing complex then coordinates loading onto the Ago2 protein
containing RISC as described earlier for siRNA. Pre- shRNA has been found to be part of the
RLC; thus, pre-shRNA may potentially directly associate with RLC rather than through a two
steps process via a different Dicer/TRBP/PACT complex [95].
46
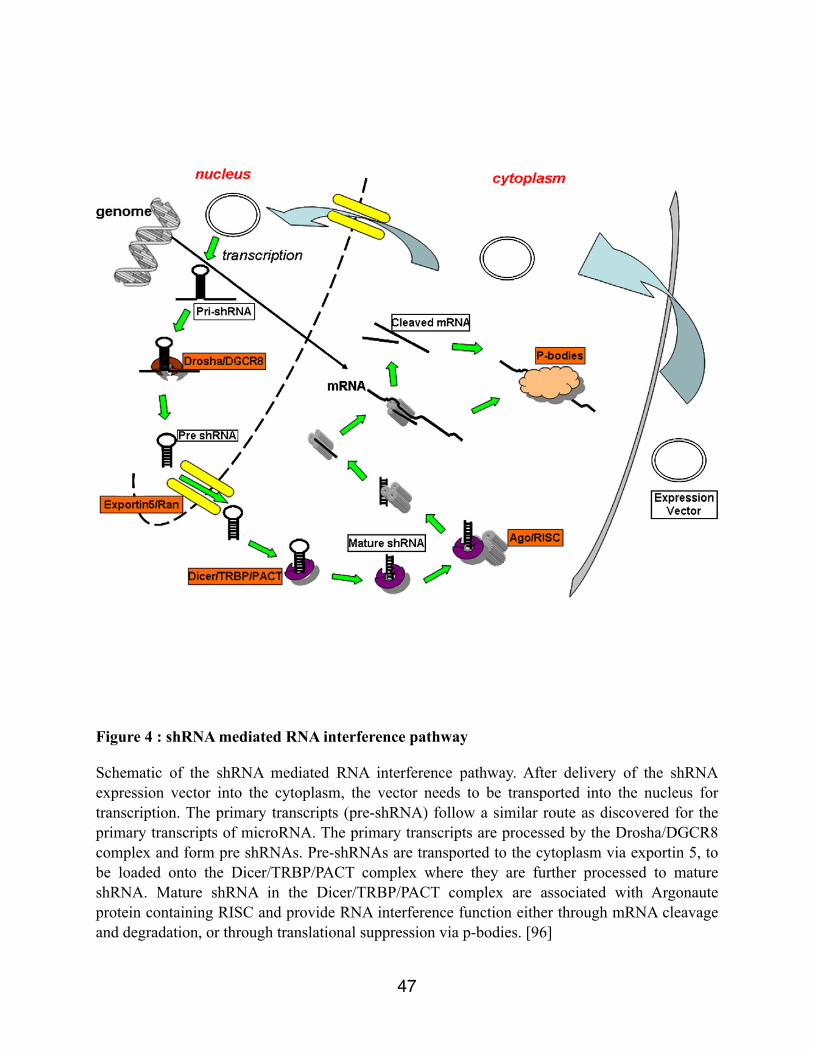
Figure 4 : shRNA mediated RNA interference pathway
Schematic of the shRNA mediated RNA interference pathway. After delivery of the shRNA expression vector into the cytoplasm, the vector needs to be transported into the nucleus for transcription. The primary transcripts (pre-shRNA) follow a similar route as discovered for the primary transcripts of microRNA. The primary transcripts are processed by the Drosha/DGCR8 complex and form pre shRNAs. Pre-shRNAs are transported to the cytoplasm via exportin 5, to be loaded onto the Dicer/TRBP/PACT complex where they are further processed to mature shRNA. Mature shRNA in the Dicer/TRBP/PACT complex are associated with Argonaute protein containing RISC and provide RNA interference function either through mRNA cleavage and degradation, or through translational suppression via p-bodies. [96]
47

1.2.12.2 Delivery systems shRNA
Efficacy of an RNAi therapeutic is limited by the quantity of the oligomer that effectively enters
the cells. In the clinical setting this is primarily dependent on the method of delivery.
Viral vectors are popular for laboratory delivery of shRNA because of their high transfection
efficiency and effective integration of exogenous DNA, but they have been losing support in
recent years because of concerns over safety and immunogenicity [96-97]. Non-viral polymeric
delivery systems, in particular those with biodegradable components, have much better safety
profiles than their viral counterparts though their transfection efficiency is generally lower.[98 to
100].
There are three major classes of non-viral delivery vehicle systems: synthetic polymers, natural/
biodegradable polymers, and lipids; many of the vehicles that are showing promise are actually
hybrids of these classes.
1.2.12.2.1 Lentiviral vector
Lentiviral vectors is an attractive gene delivery system for stable gene transduction in relatively
quiescent cells. This is why, lentiviral vectors have also been used for direct in situ infection
within a living organism. [101-103]
48

Unlike other retroviruses, lentivectors do not necessarily require cell division for proviral
integration and productive infection [104] and so offer a gene delivery method that does not need
growth factor administration to induce cell proliferation in cells transduced ex vivo. Additionally,
lentivectors are sufficiently robust for in vivo administration. Dong Sung An and Al. reported in
2003 the successful construction of lentiviral vectors to express shRNA stably in human cells.
They demonstrated that lentiviral vectors expressing siRNA directed to the reporter gene
luciferase, when stably transduced into human cells without drug selection, were capable of
protecting the cells from infection by a lentiviral vector encoding humanized firefly luciferase as
a reporter gene. [102]
49

2- OBJECTIVES OF STUDY
The objectives of the study if a successful knockdown of Noggin and Chordin would promote or
accelerate osteogenesis. The first objective or our study was to do a successful knockdown of
Noggin and Chordin in an in vitro setting using shRNAs through a lentiviral delivery system.
Then the second objective was to evaluate if an injection of those shRNAs through a lentiviral
delivery system would increase osteogenesis in an animal model of DO (wild-type mice).
50

3- MATERIALS & METHODS IN VITRO STUDIES
First RT-PCR were realized on non-transfected culture of MC3T3-E1 to measure the mRNA
level of Noggin and Chordin expression at various timepoints. The purpose was to identify the
timepoints of highest expression.
MC3T3-E1 cells were transduced with lentiviruses expressing various shRNAs targeting the
mouse Noggin (5 shRNAs) and Chordin (3 shRNAs) genes and 1 negative control.(Sigma-
Aldrich). On day 7 after infection, levels of RNA expression for Noggin and Chordin were
monitored through RT-PCR. Western blotting were performed on the cell extracts and culture
media to verify the expression and secretion of Noggin and Chordin proteins. Cell extracts were
also analyzed on day 2 and 4 after transduction for alkaline phosphastase activity which is a
marker of osteogenic differentiation.
3.1- shRNA selection
Using the Sigma-Aldrich company selection, 5 shRNAs targeting Noggin, 3 shRNA targeting
Chordin were selected. A control was also chosen : NT which is a non-target shRNA.
51
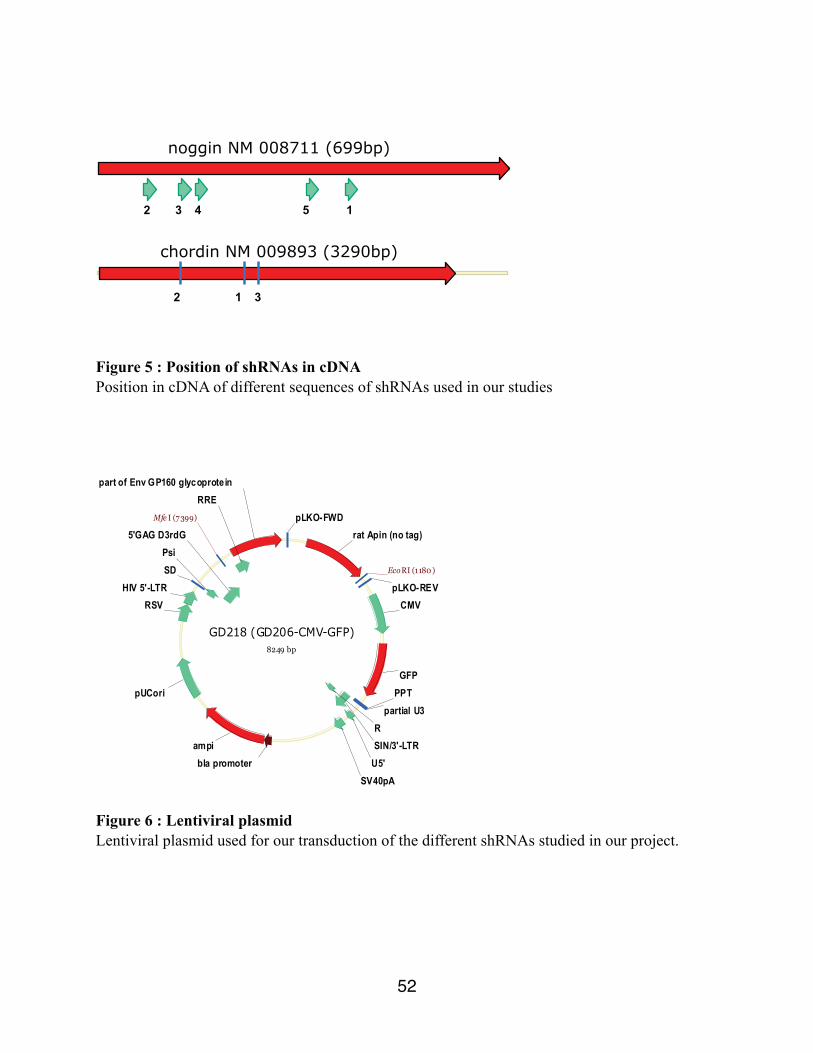
noggin NM 008711 (699bp)
3 4 5 1 2
chordin NM 009893 (3290bp)
1
3 2
Figure 5 : Position of shRNAs in cDNA Position in cDNA of different sequences of shRNAs used in our studies
GD218 (GD206-CMV-GFP)8249 bp
ampi
part of Env GP160 glycoprotein
rat Apin (no tag)
GFPPPT
partial U3
SIN/3'-LTRR
U5'SV40pA
pUCori
HIV 5'-LTRSDPsi
5'GAG D3rdG
RRE
pLKO-REV
pLKO-FWD
RSV CMV
bla promoter
EcoRI (1180 )
Mfe I (7399)
Figure 6 : Lentiviral plasmid Lentiviral plasmid used for our transduction of the different shRNAs studied in our project.
52

3.2 - Lentiviruses production
HEK293 were used for the first transfection. HEK 293 is a cell line originally derived from
human embryonic kidney cells. They have been used a lot in cellular biology because for their
easy growth, transfection and production of proteins and viruses. On day 1, the HEK293 were
seeded. The next day, they were transfected with the lentiviral shRNA plasmids using FuGENE 6
(Roche Applied Science) reagent following a ration of 3:1 (volume FuGENE : ug of DNA).
FuGENE 6 reagent was added to to media without serum and then vortex for 1 second. The mix
was then incubated at room temperature for 5 minutes. The plasmids containing the different
shRNAs where added to the mix of FuGENE and media and incubated at room temperature for
15 minutes. The plamids FuGENE mix was transfected into HEK293 cells. Culture were grown
for a period of 96 hours and the media containing the lentiviruses were collected and filtered
through 0.45 um filters. The lentiviruses were concentrated by ultracentrifugation at 25 000 rpm
for 2.5 hours. The viral pellet were resuspended in 100 ul of PBS, aliquoted, and frozen at -80 C.
[106]
3.3 - MC3T3-E1 transduction
MC3T3-E1-E1 cell line is a cell line that behave as immature, committed osteoblast cells. [107]
The lentiviruses produced in the earlier step were added to MC3T3-E1 cells in the presence of
polybrene (8ug/ ml) in 6 well-plates. After 24 h, the media was changed and the cells were
grown to confluence. The cells were then passage into 10 cm dishes and Puromycin (8ug/ml)
53

was added to select stable pools of stably expressing clones. As a control wild-type MC3T3-E1
were also treated with puromycin to monitor for selection efficacy.
3.4 - RT-PCR study
Then we decided to see if a successful knockdown of Chordin and Noggin had been produced by
our transduction. We first went ahead with PCR studies.
3.4.1 - RNA extraction from cell cultures
Every cultures were first washed 2-3 times using PBS. Then TRIzol (Invitrogen) was added to
the cells (at least 1ml/10cm2). Multiple up and down were realized using the pipet to assure a
good lysation of the cells. The cellular lysate were then stored at -20C.
3.4.2 - RNA purification
For each cellular lysate, 200ul of chlorophorm per ml of TRIzol was added. After obtaining a
good homogenization and being incubated for 3 minutes at room temperature, the samples were
centrifuged at 10 000 rpm for 15 mins at 4C. The centrifugation permitted to recuperate the
supernatant that was collected in new eppendorf tubes. A volume of 500 µl of isopropanol was
added to the samples to precipitate the RNA. The samples were mixed by inversion and allowed
to stand at room temperature for 10 minutes. The samples were centrifuged at 12 000 rpm for 10
minutes at 4C. The RNA pellets were washed with 1 ml of cold 75% ethanol per ml of TRIzol.
(Invitrogen) Again the mix was spun down at 12 000 rpm for 5 minutes at 4C. After removal of
ethanol, the samples were allowed to dry at room temperature for 30 minutes. The RNA pellets
were resuspended in DEPC water.
54

3.4.3 - RNA Quantification
We quantify the RNA using UV (OD260/ OD280). The RNA was diluted in sterile H20 and a
calculation of the quantification was made using this equation : µg/µl = (OD260 x Dilution factor
x 40)/1000.
3.4.4 - Reverse transcription of RNA
We used the applied Biosystems High-Capacity cDNA Reverse transcription kit. First the RNA
was diluted to a concentration of 2ug in 10 uL of DEPC H2O. Then a Master Mix of 2uL of 10x
RT buffer, 0.8uL of 25X dNTP Mix (100 mM), 2 uL of 10X RT Random primers, 1.0L of
MultiScribe Reverse Transcriptase and 4.2 uL of nuclease-free H20 were mixed. After, 10 uL of
diluted RNA and 10 uL of the Master Mix were mixed together in a well late and used for RT-
PCR. The GeneAmp PCR System 9700 thermal cycler (Applied Systems) was programmed as
followed : 10 mins at25C, 120 mins at 37C, 5 mins at 85C and then 4C until recuperated from
the machine.
3.4.5 - Real-Time PCR
Reverse transcribed mRNA collected were loaded into a 96-well plate and mixed with Universal
Master Mix (Applied Biosystems), specific TaqMan probe for Noggin and Chordin (TaqMan®
Gene Expression-Applied Biosystems) and RNAse free water. The 96-well plate was placed into
55

a 7500 Real Time PCR system (Applied Biosystems) for RT-qPCR analysis. Actine was used as
an endogenous control to normalize all samples used in the RT-qPCR reaction. Relative
quantification of the target RNA was performed using Applied Biosystems’ comparative Ct
method (ABI Prism 7700 Sequence Detection System User Bulletin #2, 2001).
3.5 - Western Blot study
3.5.1 - Protein collection from cell cultures and media
The stably expressing pools of MC3T3-E1 cells were seeded and grown to confluence in 6 well-
plates. The media was then switched to serum-free media and the cells were further grown for
48h. Then, the media and the cells were collected for downstream analysis.
The media was collected in eppendorf tubes and centrifugated for 5 mins. The supernatant was
then collected and Thricholoroacetic acid was added in a final concentration of 10% (v/v). The
mix was putted on ice for 30 mins. Then we proceeded with centrifugation again at 13 000 rpm.
The pellets were recuperated and washed with -20C acetone and centrifugated again at 13 000
rpm. The pellets were let to dry and then resuspended in Laemli 1x.
The cells were washed with PBS. Then a small amount of lysis buffer ( 50 mM Tris-HCl pH 7.5,
150 mM NaCl, 1% NP-40, 2 mM EDTA) was added to the cells. The cells were removed from
the wells and transferred to an Eppendorf . The samples were then incubated on ice for 30
minutes. Then they were centrifuged for 5 minutes at 13 000 rpm at 4C. After centrifugation, the
supernatant from each sample was collected in eppendorf tubes. The pellets were recuperated
56

and washed with -20C acetone and centrifugated again at 13 000 rpm. The pellets were let to dry
and then resuspended in Laemli 1x.
3.5.2 - Samples preparation
Then the protein sample was mixed to the Laemmli buffer in a ration 1 : 0,5. The volume was
then completed to 40 ul of H2O. The complete mix was then boiled for 5 minutes and then
cooled at room temperature for 5 minutes.
3.5.3 - Western Blot
Protein collected from cells culture were separated using gel electrophoresis on a 10%
polyacrylamide gel. The separated proteins were transferred onto nitrocellulose membranes and
blocked in 5 % Bovine Serum Albumin (BSA)-0.1% Phosphate Buffer Saline-Tween (PBS-T)
for an hour. The membranes were probed with anti-Noggin (ORIGENE co.- 1:2000 dilution with
5 % BSA-0.1% TBS-T) for an hour at room temperature. Subsequently, the membranes were
washed twice with 0.1% PBS-T and incubated with Horseradish peroxidase (HRP) conjugated
anti-Mouse IgG (1:25000 dilution in 1 % BSA-0.1% TBS-T) for an hour at room temperature.
Following incubation with the secondary antibody, the membranes were washed three times
using 0.1% TBS-T. Signal was detected by chemiluminescence after incubation of the blots with
the ECL+ reagent (Amersham) exposure to film. The same process was performed using anti-
Chordin (R and D systems),1:2000 dilution with 5 % BSA-0.1% TBS-T) but this time using an
HRP-conjugated anti-goat IgG 1: 25000.
57

3.6 - Alkaline phosphatase assay
Finally we wanted to see if our knockdown would be related to an increase osteogenic
differentiation. The alkaline phosphatase was then answering our question.
First the cell cultures medium was removed and then the cultures were rinsed with Tris Buffered
Saline (TBS) three times. Then for each well, lysis buffer 62.5 µl ( 20 mM Tris ph 10 + 0.5 mM
MgCl2 + 0.1% Triton X-100). Then a freeze-thaw (-20C) was performed to extract the ALP from
the cells. Then, 25 ul of the cells extracts was added to 75 ul of ALP reaction buffer (100 mM
Tris+ 100 mM NaCl, pH 9.5) and 100 ul of p-Nitrophenyl phosphate liquid substrate (SIGMA).
We incubated our samples at 37C. The activity of the alakaline phosphatase enzyme was
measured by the appearance of the p-Nitrophenol (pNP) product, which gives a yellow color,
every 5 mins for a period of 45 mins. Quantification of pNP was analyzed through a standard
curve. The protein content was quantified using the Bradford assay. The data are expressed as the
amount of pNP produced per min/ug of protein.
58

4 - MATERIALS & METHODS IN VIVO STUDIES
The second part of the project was to see if we would be able to transfer our findings from our in
vitro studies to an in vivo model. We decided to study the 2 most potent shRNAs, one targeting
Noggin and the second targeting Chordin and to use them in a wild-type mice model. Once
again, we would use them through a lentiviral vector to inject them at the distraction site for a
localized increase osteogenesis.
DO was performed on the right tibia of 54 wild-type mice using a miniature Illizarov distraction
device. Animals were randomized into two major groups according to the time of sacrifice: end
of distraction (day 17) and mid-consolidation (day 34). Each major group was sub-divided in 3
subgroups representing the type of injection that was administered at the site of distraction on
day 8. For each time point, 9 mice were injected with PBS, 9 with a lentivirus plasmid containing
a Non-Target (NT) shRNA and 9 with lentivirus plasmids containing shRNAs targeting Noggin-
Chordin combined together. To evaluate the success of the infection a GFP marker was added to
lentivirus plasmid targeting Noggin. For each subgroup 6 collected samples were studied using
Faxitron X-ray, µCT and immunohistochemistry and 3 were studied using Rt-PCR.
59

Day Procedure
0 Surgery day
5 Start distraction
8 Injection
11 End of distraction / 1st sacrifice
14 Mid-consolidation / 2nd sacrifice
Table 1 : Timeline animal procedures during in vivo studiesThis timeline explain the different steps and procedures that were effectuated on the animals during the in vivo studies
4.1 - Operative and distraction protocol
The surgical procedure was approved by the McGill University Animal Care Committee. DO
was performed on strain C57Bl/6 male mice, aged 2-3 months obtained from Charles River
using a miniature Ilizarov fixator adapted from Jill Helms study (Tay et al., 1998). Mice were
subcutaneously injected with 0.1 ml of (1mg/kg-Sigma) of buprenorphine and then anesthetized
with isoflurane using a Fortec pump (Cyprane Keighley, England) throughout surgery
(approximately 30 minutes). After shaving the right leg, two 0.25 mm insect pins (Austerlitz,
Marlborough, MA) were drilled 90° apart into the proximal metaphysis of the right tibia. Both
pins remained perpendicular to the tibia. The pins were secured into position using a ring and
four hexagonal nuts. A second set of pins was drilled into the distal metaphysis of the right tibia
approximately 12 mm away from the proximal set of pins. Three rods were inserted through the
60

first ring of the mini-Ilizarov fixator (Paolo Alto, CA) (Figure 7 and Figure 8) and secured into
position. A second ring was placed through the three rods and secured onto the distal set of pins
using hexagonal nuts. Once the mini-fixator was installed into place, a small incision was made
along the diaphysis of the right tibia using a no.11 surgical scalpel (Fisher Scientific, Osaka,
Japan). A small hole was drilled into the middle of the diaphysis using an insect pin to weaken
the bone and facilitate the osteotomy. Using the scalpel, the bone was cut along the hole to
complete the osteotomy. The fractured ends of the bone were aligned and the incision was closed
using coated vicryl sutures (J385H-Johnson & Johnson, North Ryde, NSW). Animals were
subcutaneously injected with 0.1 ml of buprenorphine and checked regularly throughout a 5 day
latency period for pain management before initiating distraction. Mice were allowed to weight
bear immediately. Mice were distracted for 12 days using a distraction rate of 0.2 mm every 12
hours. Mice were euthanized by using carbon dioxide. Distracted tibial samples were collected at
2 time points: 17 days (end of distraction phase) and 34 days (mid-consolidation phase) for
further analysis.
61

Figure 8: Mini Ilizarov external fixator
Small scale Ilizarov external fixator developped by Dr. Jill Helms in Stanford University, Palo Alto, CA
Figure 9 : Mouse in DO
Mouse with mini Ilizarov external fixator applied on a wild-type mice
62

4.2 - Injection
4.2.1 - Transfer of noggin shRNA into lentivirus plasmid containing GFP
The plasmid containing the active Noggin shRNA was changed to replace the puromycin cassette
with GFP. To do so, a plasmid containing a GFP coding sequence was digested with EcoRI and
the fragment purified on a minelute column. This fragment was then inserted into the Noggin
shRNA plasmid that was previously restricted with EcoRI, thus removing the puromycin
sequence. The resulting plasmid was confirmed by digestion with HinDIII.
N420 noggin shRNA lentiviral plasmid7397 bp
GFP
ampi
part of Env GP160 glycoprotein
TRCN0000066294 (N420)
extra C in more recent Sigma sequence5'GAG D3rdG
PsiSD
HIV 5'-LTR
pUCoriSV40pAU5'
RSIN/3'-LTR
partial U3PPT
RRE
pLKO-REVpLKO-FWD
primer REV GFP
primer FWD GFP
CMV
RSV
U6
bla promoter
Figure 10: Final plasmid containing GFP after Noggin shRNA transfer
Position of Noggin shRNA in plasmid containing GFP after transfer
63

4.2.2 - Injection protocol
For the injection 3 different groups were formed. The first group received an injection of shRNA
lentiviruses targetting Chordin and Noggin together , the second group received NT shRNAS
lentiviruses end the third group received PBS. The injection was realized as follow : On day 8
post-surgery, a 10 ul percutaneous bolus injection was realized using a 100 ul insulin syringe.
4.3 - Faxitron X-ray and Micro-Computed Tomography
In preparation for micro-computed tomography (µCT) and Faxitron x-ray, distracted bones
collected from wild-type and heterozygous mice (n=6 per time point) were fixed in 4%
paraformaldehyde (PFA) 4g of paraformaldehyde in 100 ml 1x PBS) overnight at 4°C. Samples
were washed in 1x PBS for 30 minutes and consecutively dehydrated in graded solutions of 50%
and 70% ethanol twice for one hour intervals at room temperature. Samples stored in 70%
ethanol were taken to the Centre of Bone and Periodontal Research of McGill University for
analysis.
µCT analysis was completed using the SkyScan 1072 (Aartselaar, Belgium) that has a 20-100
KeV/0-250 mA sealed, air-cooled, microfocus X-ray source with a polychromatic beam. The
SkyScan 1072 was also equipped with a detector that contained a 12-bit, cooled CCD camera
(1024 by 1024 pixels) coupled by a fiber optics taper to the scintillator. The distracted tibiae were
scanned at 45 KeV/222 µA with 25X magnification (11.25 µm pixel size). Image reconstruction
was performed using NRecon (1.4.4, SkyScan). Static histomorphometry parameters such as,
tissue volume (mm3), bone volume (mm3), bone volume/tissue volume (BV/TV %) were
64

measured using the CT Analyser (1.8.0.2, SkyScan). Furthermore, the Faxitron MX-20 was used
to produce radiographs of the distracted tibiae.
4.4 - Immunohistochemistry
4.4.1 - Paraffin embedding
Collected tibial samples from 17 days (end of distraction) and 34 days (mid-consolidation phase)
were suspended in 4% PFA overnight at 4°C. The following day, samples were washed in 1x
PBS for 30 minutes. Samples were then submerged in a solution of 0.85% saline/30% ethanol for
15 minutes. The samples underwent a gradual dehydration process by immersing them in
solutions of 50%, 70%, 90% and 100% ethanol for 15 minute each. The dehydrated tibial
samples were resuspended in solutions of 1:1 ethanol:xylene (Fisher Scientific) and xylene for
15 minutes each. Moreover, the samples were washed three times with molten paraplast (Oxford
Labware, St. Louis, MO) and then kept in paraplast overnight at 55°C under vacuum. The
samples were positioned in molten paraplast-containing cassettes and left to solidify at 4°C
forseveral hours. After paraffinization, the blocks were trimmed at 0.7 µm and sectioned
longitudinally at 5 µm using a Lecia RM 2255 microtome (Leica Microsystems, Richmond Hill,
ON). Sections were warmed in a 30°C water bath for 30 seconds and then placed on Superfrost
Plus slides (Scientific Device Laboratory, Des Plaines, IL).
65

4.4.2 - GFP marking
Distracted tibial samples (n=3 per time point) were deparaffinized and blocked with 10%
hydrogen peroxide for 10 minutes to inhibit endogenous peroxidase activity. The sections were
washed with distilled water and 1x PBS (phosphate buffered saline). Following the washings, the
samples were incubated in phosphate-buffered saline containing 10% normal goat serum (Vector
Labs, Burlingame, CA) in a humidified chamber for 20 minutes to block nonspecific binding.
Sections were rinsed with 1x PBS and blocked once more using the avidin-biotin blocking kit
(Vector, SP-2001). For immunostaining, commercially available GFP antibodies were used to
detect GFP marking in a dilution 1/50.
4.5 - RT-PCR on tibia sample
4.5.1 - Bone collection
First the tibia bone specimens were collected from the mice. As much soft tissue and skin were
removed. They the specimen was place in a 15 ml tube in an RNA later solution. Then the tube
was placed at -20oC for future steps, until all specimens are ready for analysis.
4.5.2 - Tissue Homogenisation
First the bone was place in a fresh Petri dish and only the distracted zone was cutted in small
pieces. The crushed tissue was suspended in 2 ml of Trizol (Invitrogen) and further homogenized
using a Polytron PT-MR 3000 (Kinematica) for 20 seconds at 20 000 rpm.
66

4.5.3 - RNA extraction, reverse transcription RNA and RT-PCR Noggin and Chordin
The RNA extraction, the reverse transcription of RNA and the RT-PCR for Noggin and Chordin
followed the same protocols as described earlier for the extract from the cells cultures.
4.5.4 - RT - PCR : GFP and puromycin
The PCR reactions were carried out in a final volume of 25 ul containing 2.5 µl of 10x PCR
Buffer- MgCl2 (200 mM Tris-HCl pH 8.4, 500 mM KCl, 1.25 ml MgCl2), 0.5 µl of 10 mM
dNTP, 0.5 µl of each oligonucleotides primers 25 uM (GFP and puromycin) , 0.25 µl of 5U/µl
Taq DNA polymerase (Invitrogen, Carlsbad, CA) and 0.1 ug of cDNA. The primer sequences
for GFP and puromycin are as followed : GFP-FWD (CTCGTGACCACCCTGACC) GFP-REV
(TCTGCTGGTAGTGGTCGG), PURO-FWD (GTACAAGCCCACGGTGCG) PURO-REV
(TCGGCGGTGACGGTGAAG). The PCR reactions were amplified using a GeneAMP PCR
System 9700 thermal cycler (Applied Biosystems, Singapore). PCR conditions included heating
for 5 minutes, 35 cycles of denaturation at 94°C for 30 seconds, annealing at 58°C for 30
seconds and extension at 72°C for 45 seconds, with further extension at 72°C for 7 minutes. The
expected size for puromycin and GFP fragments were 534 bp and 376 bp respectively. The PCR
products were separated via electrophoresis using a 1.3% UltraPure Agarose (Invitrogen) gel.
The PCR products were visualized by Ethidum Bromide staining.
67

5- RESULTS IN VITRO
5.1 - RT-PCR results non-transfected MC3T3-E1
MC3T3-E1 cells were seeded and grown to confluence (designated as day 0). Cells were then
differentiated in a-MEM with ascorbic acid and beta-glycerophosphate. Then we proceeded with
RT-PCR at different timepoints to be able to evaluate the time when the RNA expression for both
Noggin and Chordin was at its highest level. In both cases, the levels of RNA expression of
Noggin and Chordin was at its highest on Day 7.
Graph 1 : RT-PCR -MC3T3-E1 Noggin and Chordin mRNA level
MC3T3-E1 cells were seeded and grown to confluence (designated as day 0). Cells were then differentiated in a-MEM with ascorbic acid and beta-glycerophosphate. Then we proceeded with RT-PCR at different timepoints to be able to evaluate the time when the RNA expression for both Noggin and Chordin was at its highest level. In both cases, the levels of RNA expression of Noggin and Chordin was at its highest on Day 7.
68
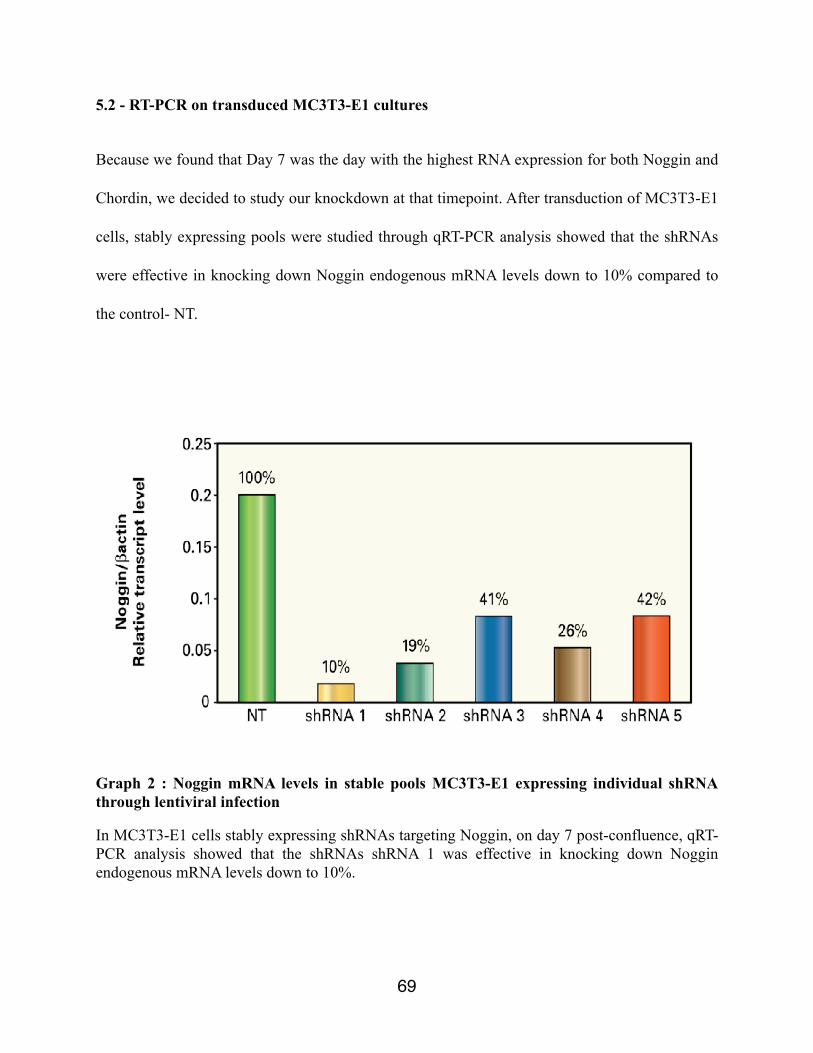
5.2 - RT-PCR on transduced MC3T3-E1 cultures
Because we found that Day 7 was the day with the highest RNA expression for both Noggin and
Chordin, we decided to study our knockdown at that timepoint. After transduction of MC3T3-E1
cells, stably expressing pools were studied through qRT-PCR analysis showed that the shRNAs
were effective in knocking down Noggin endogenous mRNA levels down to 10% compared to
the control- NT.
Graph 2 : Noggin mRNA levels in stable pools MC3T3-E1 expressing individual shRNA through lentiviral infection
In MC3T3-E1 cells stably expressing shRNAs targeting Noggin, on day 7 post-confluence, qRT-PCR analysis showed that the shRNAs shRNA 1 was effective in knocking down Noggin endogenous mRNA levels down to 10%.
69

Also, after transduction of MC3T3-E1 cells, qRT-PCR analysis showed that the shRNAs were
effective in knocking down Chordin endogenous mRNA levels down to 17% compared to the
control-NT.
Graph 3 : Chordin mRNA levels in stable pools MC3T3-E1 expressing individual shRNA through lentiviral infection
In MC3T3-E1 cells stably expressing shRNAs targeting Noggin, on day 7 post-confluence, qRT-PCR analysis showed that the shRNAs 1 was effective in knocking down Chordin endogenous mRNA levels down to 17%.
Those results are consistent with a successful knockdown of Noggin and Chordin mRNA level in
MC3T3-E1 cells after our transduction.
70

5.3 - Western Blot
Culture of MC3T3-E1 stably expressing shRNAs targetting Noggin were grown and the proteins
from the media and the cells were collected and studied through Western Blot analysis. The
results showed a successful knockdown of the Noggin protein by shRNA 1 for both the proteins
secreted in the condition media and the proteins from the cells. Only the 2 most successful
shRNA have been shown in the following images.
Figure 11 : Noggin - Western blot on secreted proteins in condition media of MC3T3-E1 stably expressing shRNAs targeting Noggin1-Noggin recombinant proteins, 2-NT, 3-shRNA 2, 4-shRNA 1Culture of MC3T3-E1 stably expressing shRNAs targeting Noggin were grown and the proteins from the media and the cells were collected and studied through Western Blot analysis. The results showed a successful knockdown of the Noggin protein by shRNA 1 for the proteins secreted in the condition media
71

Figure 12 : Noggin - Western blot on cellular proteins of MC3T3-E1 stably expressing shRNAs targeting Noggin 1-Noggin recombinant proteins, 2-NT, 3-shRNA 2, 4-shRNA 1Culture of MC3T3-E1 stably expressing shRNAs targeting Noggin were grown and the proteins from the media and the cells were collected and studied through Western Blot analysis. The results showed a successful knockdown of the Noggin protein by shRNA 1 for the cellular proteins
The same study was performed for the shRNA targetting Chordin on an MC3T3-E1 culture
stably expressing shRNAs targeting Chordin. The results showed a successful knockdown of
Chordin in the proteins secreted in the condition media. Unfortunately our experiment was not
able to provide clear results that could be interpreted for the cellular proteins.
72

Figure 13 : Chordin Western blot secreted proteins in condition media of MC3T3-E1 stably expressing shRNAs targeting Chordin1- Noggin recombinant proteins, 2-NT, 3-shRNA 2, 4-shRNA 1Culture of MC3T3-E1 stably expressing shRNAs targeting Chordin were grown and the proteins from the media and the cells were collected and studied through Western Blot analysis. The results showed a successful knockdown of the Chordin protein by shRNA 1 for the proteins secreted in the condition media
73

5.4 - Phosphatase alkaline assay
The phosphatase alkaline activity is a marker of the osteogenic differentiation. The test was
performed on cell cultures stably expressing the most potent shRNA from our previous test (RT-
PCR and Western Blot) for Noggin and Chordin. The ALP assays were done on day 2 and day 4
post confluence. They were normalized via the Bradford Assay.
They were compared to the control NT.
The results showed an increase of the phosphatase alkaline activity that was significant for
Noggin at both time points. For Chordin the results also showed an increase activity on day 2,
the difference compared to the control (NT) was not significant on day 4.
74

Graph 4 : ALP activity in MC3T3-E1 stably expressing the shRNA targeting Noggin and ChordinThe test was performed on cell cultures stably expressing the most potent shRNA from our previous test (RT-PCR and Western Blot) for Noggin and Chordin. The ALP assays were done on day 2 and day 4 post confluence. They were normalized via the Bradford Assay.The alkaline phosphatase activity is significantly increase in cells stably expressing shRNAs targeting Noggin and Chordin on Day 2.
75

6 - IN VIVO RESULTS
6.1 - Surgical and distraction protocol success rate
Distraction osteogenesis was performed on a total of 56 2-3 months old wild-type mice. Of the
56 mice, 54 successfully survived DO, accounting for a 96.4 % success rate post-surgery. The 2
mice died post-surgery due to the development of a necrotic foot.
6.2 - Injection success rate
An injection of either PBS, lentiviral plasmids including NT shRNA or lentiviral plasmids
including shRNAs targetting Noggin and Chordin was done on day 8 post surgery.
Unfortunately, at the time of the injection a significant volume of the injection was coming out of
injection site. This complication was observed for most of the injections realized.
6.3 - Faxitron x-ray
Distracted tibial samples were collected from wild-type mice at 17 and 34 days post-surgery
( end of distraction and mid-consolidation). For each group (Noggin-Chordin, NT, PBS) 6
samples were studied for each timepoints. Bone fill scores were utilized to evaluate the
consolidation rate between the distracted segments.
Three independent and blinded reviewers evaluated the Faxitron x-ray results of mice belonging
to different dosing groups sacrificed at end of distraction (day 17) and mid-consolidation (day
34).
76

75
Chapter 4.0-Results
4.1-Mid-Consolidation Phase
4.1.1-Faxitron x-ray
Distracted tibial samples were collected from C57Bl/6 wild type male mice at 34 and 51 days
post-surgery (mid-consolidation and end of consolidation respectively) (Figure 6). Bone fill
scores were utilized to evaluate the consolidation rate between the distracted segments (Table 1).
Table 1. Bone-fill scores definition
Bone-Fill Scores Radiographic Evaluation
0 No visible bone-fill (0%)
1 Visible bone-fill, but less than 50%
2 More than 50% visible bone-fill, but less than 100% visible bone-fill
3 100% visible bone-fill
Three independent and blinded reviewers evaluated the Faxitron x-ray results of mice belonging
to different dosing groups sacrificed at mid-consolidation (day 34) and end of consolidation
(51d).
Table : Bone filling scoresSubjective Bone-Fill scores assign to the Faxitron images obtained
Unfortunately, there were no visible bone-filling score difference that could be noted in between
the treatment group and the two control groups at both timepoints.
77

Graph 5 : Bone filling scoresBone filing scores were given for each of the samples. After analysis of the results, no difference could be noted in the bone filling scores between the treatment group Noggin-Chordin and the 2 controls
78

6.4 - Micro-Computed Tomography (µ-CT)
Distracted tibial samples were collected from the wild type mice at 17 and 34 days
corresponding to end of distraction and mid-consolidation respectively. Samples (6 for each
group at each timepoints), a total of 36, were analyzed using µ-CT.
6.4.1 - Bone volume
Bone volume was calculated by the same blinded technician at McGill Bone Center.
Unfortunately for all three groups, at both timepoints no statistical difference could be measure
in terms of bone volume.
79

0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
Noggin/Chordin NT PBS
BV
(mm
^3)
Treatment group
Bone Volume (BV)
17 days
34 days
Graph 6 : Bone volumesBone volume was measured using the uCT images for the samples (n=6) of each groups. After analysis, no statistically significant difference could be noted in between the Noggin/Chordin group and the two controls.
80

6.4.2 - Bone volume/Tissue volume ratio
The same results could be observe for the calculation of the ration of Bone Volume / Tissue
volume. No statistically significant difference could be observe between the treatment 3 groups.
0.00
5.00
10.00
15.00
20.00
25.00
30.00
Noggin/Chordin NT PBS
BV/
TV
Treatment group
Bone Volume/Tissue Volume (BV/TV)
17 days
34 days
Graph 7 : Bone volume / Tissue volume ratiosThe ratio of Bone volume/ Tissue volume was measured using the uCT images for the samples (n=6) of each group. After analysis, no statistically significant difference could be noted in between the Noggin/Chordin group and the two controls.
81

6.5 - RT-PCR
RT-PCR studies were realized on the distracted zone of the tibia for all the groups. At each
timepoints a for each group 3 samples were collected. After analysis, at both 17 and 34 days, the
results did not show any significant knockdown of Noggin or Chordin.
Also, there were no detectable and specific amplification products for both and GFP and
Puromycin.
Chordin mRNA levels in stable pools MC3T3-E1 expressing individual shRNA through lentiviral infection
Graph 8 : RT-PCR tibia samplesRT-PCR were realized on distracted tibia samples (n=3) for each group. No statistically significant knockdown of Noggin and Chordin could be observed in the treatment group compared to the controls at both timepoints (17 and 34 days).
82

6.6 - Immunohistochemistry - GFP marking
For the in vivo studies, the lentiviral plasmid containing the shRNA targeting Noggin, the
puromycin segment was changed for GFP segment. As we were proceeding with our
immunohistochemistry studies, we were expecting to found signal in the samples that had
received the injection of lentiviruses for Noggin and Chordin. However, as the experiment
started, to be able to find any GFP marking we had to greatly increase the sensitivity of our test.
When we finally obtained results, we found GFP marking in all 3 groups : Noggin-Chordin, NT
and PBS. Most probably we had increased the sensitivity of our test so much to get detection that
we had lost the specificity of our test.
83

Puromycin and GFP signal in MC3T3-E1 cells stably expressing shRNAs targeting Noggin and Chordin
Figure 14 : Puromycin and GFP signal in MC3T3-E1 cells stably expressing shRNAs targeting Noggin and ChordinThere were no detectable and specific amplification products for both and GFP and Puromycin in the Noggin and Chordin group.
84

7- DISCUSSION
Multiple technique and treatment have been investigated to accelerate bone formation during
DO. The use of BMPs have been investigated in the past and seems to be presenting promising
results and participate to an accelerated bone formation in DO [108].
However, one of the main concerns with BMPs are the large concentrations of these proteins
needed to be administered at the distraction site with unknown side effects [109] and high costs.
Thus, an alternative method is manipulation of the intrinsic pathways to accelerate bone
formation during DO. This could be achieved by manipulating the endogenous BMP pathways in
order to up regulate their expression. So our hypothesis was that by inhibiting the BMP
antagonists Noggin and Chordin we would increase osteogenesis. So far no study had been made
of knockdown of Noggin and Chordin in an in vivo model of DO.
For this reason, we first tested our hypothesis in an in vitro setting. A knockdown of Noggin and
Chordin had been tried using a shRNAs targeting both of them though a lentiviral plasmid
delivery system. The cells culture chosen was the MC3T3-E1 because of their preosteoblastic
nature. First our RT-PCR studies confirmed that we achieved an efficient knockdown of the
mRNA levels for both Chordin and Noggin by the most potents shRNAs. Following those
results, the decision to evaluate if this knockdown would translate in a protein knockdown.
Western blot analysis were made for both Noggin and Chordin. A successful knockdown of
Noggin for both cellular proteins and culture media proteins was found. For Chordin, also a
successful knockdown was proven for the cellular proteins. The next step was then to see if those
85

successful knockdown would translate in an increase osteogenic activity. The ALP assay showed
an increased activity for Noggin at both 2 and 4 days post-confluence and for Chordin at day 2.
Encouraged by those results we decided to transport our study to an in vivo model. We decided
to use our most efficient shRNAs targetting Noggin and Chordin (one for each) and use them in
our in vivo studies. We used an animal model of wild-type mice that underwent distraction
osteogenesis using a small scale Ilizarov external fixator. The treatment group was receiving an
injection on day 8 post-surgery with lentiviruses containing each 1 shRNA targeting Noggin or
Chordin. The control groups were receiving either an injection of lentiviruses containing a Non-
Target (NT) shRNA or a PBS injection. I operated on a total of 56 mice and 54 survived the
surgery and post-operative period. Unfortunately, during the experimentation our main issue was
related to the injection. Most of the injected solution was extravasating from the injection site. It
was also very difficult to assess the volume of injection loss because the liquid formula was
getting loss on the animal skin/hair.
Faxitron and uCT were realized to evaluation the osteogenesis at both time points (17 and 34
days). A total of 6 samples for each group at each timepoints were sent to the bone center for
analysis. Sadly, none of the results showed a significant increase in osteogenesis in the treatment
group compared to the controls. Also, our RT-PCR study (performed on 3 samples/group/
timepoint) was concordant with those results as it did not show any significant knockdown of
Noggin and Chordin in the treatment group compared to the controls. Finally, it is interesting to
see that GFP marking was found in all 3 groups while we were only expecting it for the Noggin-
86

Chordin group. Also, no GFP RNA could be identify in our PCR study performed on the
distracted tibia samples from the treatment group Noggin and Chordin. At the light of those
results, we decided to stop our immunohistochemistry studies.
At this point, I think the main issue with our results are related to the injection technique. The
proper way of injection, the volume of injection and the consistency of the injection should be
eventually reevaluated. But it is certain that if most of the injection does not reach the site that
we wish to treat, it is very difficult to imagine that we would get any significant difference in
between the groups. Possible solutions would be to use a systemic delivery system that would
target specifically to the bone-formation surfaces. Zhang et al. were recently able to develop a
targeted delivery system for RNA interference-based anabolic therapy using (AspSerSer)6-
liposome. [110]
However, our disappointing in vivo results could also be related to other physiologic cascades
that could be activated by our knockdown. It is easy to think that other BMPs antagonists could
be upregulated in response to a knockdown of Noggin and Chordin. This would then open the
hypothesis that knockdown of Noggin and Chordin might not be enough to achieve an increased
osteogenesis. Although this idea goes in contradiction with previous successful studies
performed for example by Wan and Al [78] where they obtained an accelerated bone formation
with Noggin knockdown.
Finally it is important to think that this in vivo study was mainly constructed as a proof of
concept since there are still concerns related to the usage of lentiviral delivery system.
Theorically, because thee site of integration is unpredictable, the provirus can disturb the
87

function of cellular genes and lead to activation of oncogenes promoting the development of
cancer.However, studies have shown that lentivirus vectors have a lower tendency to integrate in
places that potentially cause cancer than gamma-retroviral vectors.[111] More specifically, one
study found that lentiviral vectors did not cause either an increase in tumor incidence or an
earlier onset of tumors in a mouse strain with a much higher incidence of tumors.[112]
This is why, the next step would be to find another efficient delivery system. Different options
are available to us, but so far the nanoparticles delivery system has been the one that have
interested us the most.
In conclusion, we were able to show a successful knockdown of Noggin and Chordin in an in
vitro model. Our ALP assay was also consistent with and increased osteogenic differentiation
activity. Our in vivo study was interesting by being one of the first one to study Noggin and
Chordin knockdown in a DO animal model. Unfortunately, our results were not able to confirm
either a successful knockdown or an increased osteogenesis.
88

8-References
1. Ilizarov, G.A. and A.A. Deviatov, [Surgical elongation of the leg]. Ortop Travmatol Protez,
1971. 32(8): p. 20-5.
2. Ilizarov G. The tension-stress effect on the genesis and growth of tissues: Part II. The
influence of the rate and frequency of distraction. Clinical orthopaedics and related research.
1989;239: 263-85.
3. Urist, M.R., Bone: formation by autoinduction. Science, 1965. 150(698): p. 893-9.
4. Ilizarov G. Operative Principles of Ilizarov. Italy: A.S.A.M.I. Group; 1990. 3-8.
5. Ducy P, Karsenty G. Genetic control of cell differentiation in the skeleton. Current Opinion
in Cell Biology. 1998;10(5): 614-19.
6. Vaes BLT, Ducy P, Sijbers AM, Hendriks JMA, van Someren EP, de Jong NG, et al.
Microarray analysis on Runx2-deficient mouse embryos reveals novel Runx2 functions and
target genes during intramembranous and endochondral bone formation. Bone. 2006;39(4):
724-38.
7. Tsiridis E, Upadhyay N, Giannoudis P. Molecular aspects of fracture healing: Which are the
important molecules? Injury. 2007;38(1S): 11-25.
8. Kosher RA. Collagen gene expression during limb cartilage differentiation. The Journal of
Cell Biology. 1986;102(4): 1151-56.
89

9. Provot S, Schipani E. Molecular mechanisms of endochondral bone development.
Biochemical and Biophysical Research Communications. 2005;328(3): 658-65.
10. Hartmann C. Skeletal Development-Wnts Are in Control. Molecules and Cells. 2007;24(2):
177.
11. Kojimoto H, Yasui N, Goto T, Matsuda S, Shimomura Y. Bone Lengthening In Rabbits By
Callus Distraction. Journal of Bone & Joint Surgery, British Volume. 1988(70-B): 543-9.
12. Delloye C, Delefortrie G, Coutelier L, Vincent A. Bone regenerate formation in cortical bone
during distraction lengthening. An experimental study. Clin Orthop Relat Res. 1990;250:
34-42.
13. Arnonson J, Harrison BH, Stewart CL, Harp Jr JH. The histology of distraction osteogenesis
using different external fixators. Clin Orthop Relat Res. 1989;241: 106-16.
14. Rauch F, Lauzier D, Croteau S, Travers R, Glorieux FH, Hamdy R. Temporal and spatial
expression of bone morphogenetic protein-2,-4, and-7 during distraction osteogenesis in
rabbits. Bone. 2000;27(3): 453-59.
15. Yasui N, Sato M, Ochi T, Kimura T, Kawahata H, Kitamura Y, et al. Three Modes of
Ossification During Distraction Osteogenesis in The Rat. Journal of Bone & Joint Surgery,
British Volume. 1997;79(5): 824-30.
16. Yasui N, Sato M, Ochi T, Kimura T, Kawahata H, Kitamura Y, et al. Three Modes of
Ossification During Distraction Osteogenesis in The Rat. Journal of Bone & Joint Surgery,
British Volume. 1997;79(5): 824-30.
17. Paley D. Problems, obstacles, and complications of limb lengthening by the Ilizarov
technique. Clin Orthop Relat Res. 1990;250: 81-104.
90

18. Greenwald JA, Luchs JS, Mehrara BJ, Spector JA, Mackool RJ, McCarthy JG, et al.
Pumping the regenerate: An evaluation of oscillating distraction osteogenesis in the rodent
mandible. Annals of plastic surgery. 2000;44(5): 516-21.
19. Kim UK, Chung IK, Lee KH, Swift JQ, Seong WJ, Ko CC. Bone Regeneration in
Mandibular Distraction Osteogenesis Combined With Compression Stimulation. Journal of
Oral and Maxillofacial Surgery. 2006;64(10): 1498-505.
20. Malizos KN, Hantes ME, Protopappas V, Papachristos A. Low-intensity pulsed ultrasound
for bone healing: An overview. Injury. 2006;37(1S): 56- 62.
21. Sakurakichi K, Tsuchiya H, Uehara K, Yamashiro T, Tomita K, Azuma Y. Effects of timing of
low-intensity pulsed ultrasound on distraction osteogenesis. Journal of Orthopaedic
Research. 2004;22(2): 395-403.
22. Eberson CP, Hogan KA, Moore DC, Ehrlich MG. Effect of low-intensity ultrasound
stimulation on consolidation of the regenerate zone in a rat model of distraction osteogenesis.
Journal of pediatric orthopedics. 2003;23(1): 46-51.
23. Shimazaki A, Inui K, Azuma Y, Nishimura N, Yamano Y. Low-intensity pulsed ultrasound
accelerates bone maturation in distraction osteogenesis in rabbits. Journal of Bone & Joint
Surgery, British Volume. 2000;82(7): 1077-82.
24. Claes L, Rüter A, Mayr E. Low-intensity ultrasound enhances maturation of callus after
segmental transport. Clinical orthopaedics and related research. 2005(430): 189-94.
25. Kitoh H, Kitakoji T, Tsuchiya H, Mitsuyama H, Nakamura H, Katoh M, et al.
Transplantation of marrow-derived mesenchymal stem cells and platelet- rich plasma during
distraction osteogenesis—a preliminary result of three cases. Bone. 2004;35(4): 892-98.
91

26. Tsubota S, Tsuchiya H, Shinokawa Y, Tomita K, Minato H. Transplantation of osteoblast-like
cells to the distracted callus in rabbits. Journal of Bone & Joint Surgery, British Volume.
1999;81(1): 125-29.
27. Takamine Y, Tsuchiya H, Kitakoji T, Kurita K, Ono Y, Ohshima Y, et al. Distraction
osteogenesis enhanced by osteoblastlike cells and collagen gel. Clinical orthopaedics and
related research. 2002(399): 240-46.
28. Riggs BL, Parfitt AM. Drugs used to treat osteoporosis: the critical need for a uniform
nomenclature based on their action on bone remodeling. J Bone Miner Res. 2005;20(2):
177-84.
29. Takahashi M, Yukata K, Matsui Y, Abbaspour A, Takata S, Yasui N. Bisphosphonate
modulates morphological and mechanical properties in distraction osteogenesis through
inhibition of bone resorption. Bone. 2006;39(3): 573-81.
30. Yeung H, Lee S, Fung K, Leung K. Expression of basic fibroblast growth factor during
distraction osteogenesis. Clin Orthop Relat Res. 2001;385: 219-29.
31. Greenwald JA, Luchs JS, Mehrara BJ, Spector JA, Mackool RJ, McCarthy JG, et al.
Pumping the regenerate: An evaluation of oscillating distraction osteogenesis in the rodent
mandible. Annals of plastic surgery. 2000;44(5): 516-21.
32. Kitoh H, Kitakoji T, Tsuchiya H, Katoh M, Ishiguro N. Transplantation of culture expanded
bone marrow cells and platelet rich plasma in distraction osteogenesis of the long bones.
Bone. 2007;40(2): 522-28.
33. Tiedeman, J.J., et al., Treatment of nonunion by percutaneous injection of bone marrow and
demineralized bone matrix. An experimental study in dogs. Clin Orthop Relat Res,
1991(268): p. 294-302.
92

34. Oikarinen, J., Experimental spinal fusion with decalcified bone matrix and deep-frozen
allogeneic bone in rabbits. Clin Orthop Relat Res, 1982(162): p. 210-8.
35. Glowacki, J., D. Altobelli, and J.B. Mulliken, Fate of mineralized and demineralized osseous
implants in cranial defects. Calcif Tissue Int, 1981. 33(1): p. 71-6.
36. Fowler, E.B., L.G. Breault, and G. Rebitski, Ridge preservation utilizing an acellular dermal
allograft and demineralized freeze-dried bone allograft: Part II. Immediate endosseous
implant placement. J Periodontol, 2000. 71(8): p. 1360-4.
37. Bolander, M.E., Balian G. The use of demineralized bone matrix in the repair of segmental
defects. Augmentation with extracted matrix proteins and a comparison with autologous
grafts.J Bone Joint Surg Am,1986. 68(8) : p. 1264-74
38. Einhorn, T.A., et al., The healing of segmental bone defects induced by demineralized bone
matrix. A radiographic and biomechanical study. J Bone Joint Surg Am, 1984. 66(2): p.
274-9.
39. Gepstein, R., R.E. Weiss, and T. Hallel, Bridging large defects in bone by demineralized bone
matrix in the form of a powder. A radiographic, histological, and radioisotope- uptake study
in rats. J Bone Joint Surg Am, 1987. 69(7): p. 984-92.
40. Hagino, T. and Y. Hamada, Accelerating bone formation and earlier healing after using
demineralized bone matrix for limb lengthening in rabbits. J Orthop Res, 1999. 17(2): p.
232-7.
41. Giannoudis PV, Dinopoulos H, Tsiridis E, Bone substitutes: an update. Injury 2005, 36(Suppl
3):S20-27.
42. Finkemeier CG, Bone-grafting and bone-graft substitutes.J Bone Joint Surg Am 2002, 84(3):
454-464.
93

43. Bullens PH, Bart Schreuder HW, de Waal Malefijt MC, Verdonschot N, Buma P: Is an
impacted morselized graft in a cage an alternative for reconstructing segmental diaphyseal
defects? Clin Orthop Relat Res 2009, 467(3):783-791.
44. Ostermann PA, Haase N, Rübberdt A, Wich M, Ekkernkamp A: Management of a long
segmental defect at the proximal meta-diaphyseal junction of the tibia using a cylindrical
titanium mesh cage. J Orthop Trauma 2002, 16(8):597-601.
45. Wang, Y., Ni M., Tang PF., Li G.,Novel application of HA-TCP biomaterials in distraction
osteogenesis shortened the lengthening time and promoted bone consolidation., J Orthop
Res. 2009 Apr;27(4):477-82
46. Akkouch A, Zhang Z, Rouabhia M: A novel collagen/hydroxyapatite/poly(lactide-co-ε-
caprolactone) biodegradable and bioactive 3D porous scaffold for bone regeneration. J
Biomed Mater Res A 2011, 96A:693-704.
47. Tampieri A, Landi E, Valentini F, Sandri M, D'Alessandro T, Dediu V, Marcacci M: A
conceptually new type of bio-hybrid scaffold for bone regeneration. Nanotechnology 2011,
22(1):015104.
48. Laschke MW, Witt K, Pohlemann T, Menger MD: Injectable nanocrystalline hydroxyapatite
paste for bone substitution: in vivo analysis of biocompatibility and vascularization.
J Biomed Mater Res B Appl Biomater 2007, 82(2):494-505.
49. Bail, H.J., et al., Systemic application of growth hormone for enhancement of secondary and
intramembranous fracture healing. Horm Res, 2002. 58 Suppl 3: p. 39-42.
50. Byung-Hyun Park, S.J.Y., Kyu Yun Jang, Mi-Ran Kim, Hyung-Seok Lee, Ki-Bum Kim, and
S.Y.L. Hyuk Park, Ho Sung Park, Seok Tae Lim, Kyung-Jin Song, Jung Ryul Kim, COMP–
94

angiopoietin-1 accelerates bone formation during distraction osteogenesis. Bone, 2010. 46: p.
6.
51. Xiao, Y.T., L.X. Xiang, and J.Z. Shao, Bone morphogenetic protein. Biochem Biophys Res
Commun, 2007. 362(3): p. 550-3.
52. Issack, P.S. and P.E. DiCesare, Recent advances toward the clinical application of bone
morphogenetic proteins in bone and cartilage repair. Am J Orthop, 2003. 32(9): p. 429- 36.
53. Yoon,B.S. and K.M. Lyons, Multiple functions of BMPs in chondrogenesis. J Cell Biochem,
2004. 93(1): p. 93-103.
54. Luyten, F.P., and al., Purification and partial amino acid sequence of osteogenin, a protein
initiating bone differentiation. J Biol Chem, 1989. 264(23): p. 13377-80.
55. Attisano, L., and al., Identification of human activin and TGF beta type I receptors that form
heteromeric kinase complexes with type II receptors. Cell, 1993. 75(4): p. 671-80.
56. Riggs, B.L. and A.M. Parfitt, Drugs used to treat osteoporosis: the critical need for a uniform
nomenclature based on their action on bone remodeling. Journal of Bone & Mineral
Research, 2005. 20(2): p. 177-84.
57. Heldin, C.H., K. Miyazono, and P. ten Dijke, TGF-beta signalling from cell membrane to
nucleus through SMAD proteins. Nature, 1997. 390(6659): p. 465-71.
58. Kawabata, M., T. Imamura, and K. Miyazono, Signal transduction by bone morphogenetic
proteins. Cytokine Growth Factor Rev, 1998. 9(1): p. 49-61.
59. Urist, M.R., Bone: formation by autoinduction. Science, 1965. 150(698): p. 893-9.
95

60. Mizumoto Y, Moseley T, Drews M, CooperIii VN, Reddi AH. Acceleration of Regenerate
Ossification During Distraction Osteogenesis with Recombinant Human Bone
Morphogenetic Protein-7. Journal of Bone & Joint Surgery. 2003;85(90003): 124-30.
61. Yang JH, Kim HJ, Kim SE, Yun YP, Bae JH, Kim SJ, Choi KH, Song HR,The effect of bone
morphogenic protein-2-coated tri-calcium phosphate/hydroxyapatite on new bone formation
in a rat model of femoral distraction osteogenesis, Cytotherapy. 2012 Mar;14(3):315-26.
Epub 2011 Nov 28.
62. Ebisawa T, Fukuchi M, Murakami G, Chiba T, Tanaka K, Imamura T, Miyazono K, Smurf1
interacts with transforming growth factor-beta type I receptor through Smad7 and induces
receptor degradation, J. Biol. Chem. 276 (2001) 12477–12480.
63. Garrison KR, Shemilt I, Donell S, Ryder JJ, Mugford M, Harvey I, Song F, Alt V Bone
morphogenetic protein (BMP) for fracture healing in adults. Cochrane Database Syst Rev.
2010 Jun 16;(6):CD006950.
64. Ziad Dahabreh, Rozalia Dimitriou, Peter Giannoudis: Health economics: A cost analysis of
treatment of persistent fracture non-unions using bone morphogenetic protein-7. Injury 38,
371-377 (2007)
65. Elliot Carlisle, Jeffrey Fischgrund: Bone morphogenetic proteins for spinal fusion. Spine J
5(6 Suppl), 240S-249S (2005)
66. Thomas Kleeman, Uri Michael Ahn, Anne Talbot- Kleeman: A Laparoscopic anterior lumbar
interbody fusion with rhBMP-2. Spine 26(24), 2751-2756 (2001)
67. Zara JN, Siu RK, Zhang X, Shen J, Ngo R, Lee M, Li W, Chiang M, Chung J, Kwak J, Wu
BM, Ting K, Soo C.High doses of bone morphogenetic protein 2 induce structurally
abnormal bone and inflammation in vivo. Tissue Eng Part A. 2011 May;17(9-10):1389-99.
Epub 2011 Mar 3.
96

68. Garrison KR, Donell S, Ryder J, Shemilt I, Mugford M, Harvey I, Song F. Clinical
effectiveness and cost-effectiveness of bone morphogenetic proteins in the non-healing of
fractures and spinal fusion: a systematic review. Health Technol Assess. 2007 Aug;11(30):
1-150, iii-iv.
69. Dean BJ, Watson T, Moed BR, Zhang Z, Role of bone morphogenetic proteins and their
antagonists in healing of bone fracture, Frontiers in Bioscience 14 2009 p. 2878-2888,
70. Rosen V., BMP and BMP inhibitors in bone. Ann N Y Acad Sci, 2006. 1068: p. 19-25.
71. Brown, M.A., et al., Crystal structure of BMP-9 and functional interactions with pro-region
and receptors. J Biol Chem, 2005. 280(26): p. 25111-8.
72. Hillger, F., et al., Biophysical comparison of BMP-2, ProBMP-2, and the free pro- peptide
reveals stabilization of the pro-peptide by the mature growth factor. J Biol Chem, 2005.
280(15): p. 14974-80.
73. Aspenberg P, Charlotte Jeppsson, Aris Economide: The bone morphogenetic proteins
antagonist noggin inhibits membranous ossification. J Bone Miner Res 2001 16(3), 497-500
74. Gazzerro E, Canalis E: Bone morphogenetic proteins and their antagonists. Rev Endo Meta
Disorders 2006 7(1-2),51-65
75. Yanagita M: BMP antagonists: their roles in development and involvement in
pathophysiology. Cytokine and Growth Factor Re 2005 16, 309-317
76. Wijerde M, Karp S, McMahon J, McMahon A: Noggin antagonism of BMP4 signaling
controls development of the axial skeleton in the mouse. Dev Biol 2005 286, 149-157
77. Gazzerro E, Gangji V, Canalis E: Bone morphogenetic proteins induce the expression of
noggin, which limits their activity in cultured rat osteoblasts. J Clin Invest 1998 102(12),
2106–2114
97

78. Wan D, Pomerantz J, Brunet L, Kim JB, Chou YF, Wu B, Harland R, Blau H, Longaker M:
Noggin suppression enhances in Vitro osteogenesis and accelerates in Vivo bone formation. J
Bio Chem 2007 282(36), 26450-26459
79. Niikura T, Hak D, Reddi AH: Global gene profiling reveals a downregulation of BMP gene
expression in experimental atrophic nonunions compared to standard healing fractures. J
Ortho Res 2006 24(7), 1463-1471
80. Piccolo S, Sasai Y, Lu B, De Robertis E: Dorsoventral patterning in Xenopus: inhibition of
ventral signals by direct binding of chordin to BMP-4. Cell 1996 86, 589 -598.
81. Zhang D, Ferguson CM, O’Keefe R, Puzas JE, Rosier R, Reynolds P: A role for the BMP
antagonist chordin in endochondral ossification. J Bone Miner Res 2002 17(2), 293-300
82. Balemans W, Van Hul W: Extracellular regulation of BMP signaling in vertebrates: A
cocktail of modulators. Dev Biol 2002 250, 231-250
83. Iemura S, Yamamoto T, Takagi C, Uchiyama H, Natsume T, Shimasaki S, Sugino H, Ueno N:
Direct binding of follistatin to a complex of bone-morphogenetic protein and its receptor
inhibits ventral and epidermal cell fates in early Xenopus embryo. Proc Natl Acad Sci USA
1998 95, 9337-9342
84. Nilsson O, Parker E, Hegde A, Cha M, Barnes K, Baron J: Gradients in bone morphogenetic
protein-related gene expression across the growth plate. J Endocrinology 2007 193, 75-84
85. Pushparaj PN, Aarthi JJ, Manikandan J, Kumar SD, siRNA, miRNA, and shRNA: in vivo
Applications J Dent Res 87(11) 2008 p.992-1003
86. Rao DD, Vorhies JS, Senzer N, Nemunaitis J. siRNA vs. shRNA: similarities and
differences.Adv Drug Deliv Rev. 2009 Jul 25;61(9):746-59. Epub 2009 Apr 20.
87. Cullen BR, RNAi the natural way, Nat. Genet. 37 (2005) 1163–1165.
98

88. Lee Y, C. Han AJ, Choi H, Kim J, Yim, J. Lee J, Provost P, Radmark O, Kim S, Kim VN,
The nuclear RNase III Drosha initiates microRNA processing, Nature 425 (2003) 415–419.
89. Zhang H, Kolb FA, Brondani V, Billy E, Filipowicz W, Human Dicer preferentially cleaves
dsRNAs at their termini without a requirement for ATP, EMBO J. 21 (2002) 5875–5885.
90. Lee Y, Jeon K, Lee JT, Kim S, Kim VN, MicroRNA maturation: stepwise processing and
subcellular localization, EMBO J. 21 (2002) 4663–4670.
91. Cullen BR, Transcription and processing of human microRNA precursors, Mol. Cell 16
(2004) 861–865.
92. Yi R, Qin Y, Macara IG, Cullen BR, Exportin-5 mediates the nuclear export of pre-
microRNAs and short hairpin RNAs, Genes Dev. 17 (2003) 3011–3016.
93. Lee YS, Nakahara K, Pham JW, Kim K, He Z, Sontheimer EJ, Carthew RW, Distinct roles
for Drosophila Dicer-1 and Dicer-2 in the siRNA/miRNA silencing pathways, Cell 117
(2004) 69–81.
94. Lund E, Guttinger S, Calado A, Dahlberg JE, Kutay U, Nuclear export of microRNA
precursors, Science 303 (2004) 95–98.
95. Landthaler M, Gaidatzis D, Rothballer A, Chen PY, Soll SJ, Dinic L, Ojo T, Hafner M,
Zavolan M, Tuschl T, Molecular characterization of human Argonaute-containing
ribonucleoprotein complexes and their bound target mRNAs, RNA 14 (2008) 2580–2596.
96. Check E, A tragic setback, Nature 420 (2002) 116–118.
97. Nguyen T, Menocal EM, Harborth J, Fruehauf JH, RNAi therapeutics: an update on delivery,
Curr. Opin. Mol. Ther. 10 (2008) 158–167.
99

98. Luten J, Van Nostrum CF, De Smedt SC, Hennink WE, Biodegradable polymers as non-viral
carriers for plasmid DNA delivery, J. Control. Release 126 (2008) 97–110.
99. Zhang S, Zhao B, Jiang H, Wang B, Ma B, Cationic lipids and polymers mediated vectors for
delivery of siRNA, J. Control. Release 123 (2007) 1–10.
100. Aigner A, Nonviral in vivo delivery of therapeutic small interfering RNAs, Curr. Opin. Mol.
Ther. 9 (2007) 345–352.
101. Naldini L et al. In vivo delivery and stable transduction of nondividing cells by a lentiviral
vector. Science 1996; 272: 263-267.
102. An DS, Xie Y, Mao SH, Morizono K, Kung SK, Chen IS, Lentiviral vectors for short
hairpin RNA delivery into human cells. Hum Gene Ther. 2003 Aug p.1207-12.
103. Kafri T., Blomer U., Peterson D.A., Gage F.H.,Verma I.M. Sustained expression of genes
delivered di- rectly into liver and muscle by lentiviral vectors. Nat. Genet. (1997) 17, 314–
317.
104. Miller DG, Adam MA, Miller AD. Gene transfer by retrovirus vectors occurs only in cells
that are actively replicating at the time of infection. Mol Cell Biol 1990; 10: 4239-4242.
105. Connolly JB, Lentiviruses in gene therapy clinical research, Gene Therapy (2002) 9, 1730–
1734
106. ShRNA Lentiviral Particles Transduction, Santa Cruz protocol
107. Bilezikian John P., Gideon Raisz L. Gideon A. R., Principles of bone biology : volume 1
(2002)
108. Rengachary, S.S., Bone morphogenetic proteins: basic concepts. Neurosurg Focus (2002) p.
e2.
100

109. Haque, T., et al., Characterizing the BMP pathway in a wild type mouse model of
distraction osteogenesis. Bone (2008), 1144-53.
110. Zhang G.; Guo B; Wu H.; Tang T. et al., A delivery system targeting bone formation
surfaces to facilitate RNAi-based anabolic therapy, Nature (2012) 18 (2) : 307-315
111. Cattoglio, C.; Facchini, G.; Sartori, D.; Antonelli, A.; Miccio, A.; Cassani, B.; Schmidt, M.;
Von Kalle, C. et al., Hot spots of retroviral integration in human CD34+ hematopoietic cells,
Blood (2007) 110 (6): 1770–1778
112. Montini, E.; Cesana, D.; Schmidt, M.; Sanvito, F.; Ponzoni, M.; Bartholomae, C.; Sergi, L.
S.; Benedicenti, F. et al., Hematopoietic stem cell gene transfer in a tumor-prone mouse
model uncovers low genotoxicity of lentiviral vector integration, Nature Biotechnology
(2006) 24 (6): 687–696
101