a multi-joint task d. - library and archives...
TRANSCRIPT

Learning and Retention Adaptations of Myoelectric Activity
During a Novel Multi-Joint Task
George D. V. Sarantinos
Department of Physical Education
McGill University, Montreal
Canada
May, 1999
A Thesis submitted to the Faculty of Graduate Studies and Research
In partial fulfillment of the requirements for the degree of
Master of Arts
0 George D. V. Sarantinos 1999

National Library 1*1 of Canada BiMithèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services senrices bibliographiques
395 Wellington Street 395. rue WeWWtCm OnawaON KlAON4 OlWwaON K 1 A W CaMda cana&
The author bas granted a non- exclusive licence alIowing the National Library of Canada to reproduce, loan, distribute or seil copies of this thesis in rnicroform, paper or electronic formats.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial exbacts fkom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

TABLE OF CONTENTS
ACKNOWLEDGEMENTS ......................................................................... v
.......................................................................................... ABSTRACT vi . . ............................................................................................. RESUME VII
... ................................................................................ LIST OF FIGURES v i i ~
LIST OF TABLES ................................................................................... x
GLOSSARY OF DEFINITIONS AND ABBREVIATIONS ................................ .xi
CHAPTER I
INTRODUCTION ................................................................................ - 1
1 . 1 General Objective ........................................................................... 1
......................................................... 1.2 Nature and Scope of the Problem 3
................................................................. 1.3 Statement of the Problem -5
1.3.1 Assumptions and limitations ...................................................... 6
1.4 Hypotheses .................................................................................. -7
1.5 Rationale .................................................................................... -7
CHAPTER 11
METHODOLOGY ................................................................................ -9
................................................................................ 2.1 Participants -9
2.2 Apparatus ................................................................................. -10
2.3 Experimental Task and Protocol ........................................................ I0
2.4 Data Acquisition and Analysis .......................................................... 16
....................................................................... 2.5 Statistical Analysis -24
CHAPTER In
.......................................................................................... RESULTS 25
............................................................ 3.1 Participant Characteristics - 2 5
3.2 Adaptations in Performance Outcome with Learning and Retention ........... - 2 7
3.3 Adaptations in Performance Production during both Learning and Retenion .. -29
............................................. 3.3.1 Qualitative EMG Data Analysis 29


APPENDUC 7 ANOVA Table for the Performance Time Differences Between
the Experimental and Control Groups.. ................................. -123
APPENDIX 8 ANOVA Table for the Composite Score Data of W2 for the
.............................................. ...... Experimental Group.. .. -1 24
APPENDIX 9 ANOVA Table for the Composite Score Differences of W2
Between the Experimental and Control Groups.. ..................... .125
APPENDIX 10 Within Muscle Group Representations of W3 for the
Experimental Group - Day 1 to Day 5.. ................................ 127
APPENDUC 1 1 Within Muscle Group Representations of W3 for the
Experimental Group - Ret 1 to Ret 2.. ................................. -1 29
APPENDIX 12 Within Muscle Group Representations of W3 for the
Experimental Group - Ret 2 to Ret 3.. .................................. 13 1
APP ENDIX 1 3 Within Muscle Group Representations of W 3 for the
Experimental Group - Ret 3 to Ret 5 . .................................. 1 33
APPENDIX 14 Within Muscle Group Representations of W3 for the Control
Group - Day 1, Day 5 and Ret 5.. ....................................... 135

ACKNOWLEDGEMENTS
1 would like to express my appreciation for the support provided by my thesis
advisor Dr. René Turcotte, who encouraged me to persevere through the myriad of
challenges offered by the undertaking of this research.
1 also extend my thankfulness to the chairman of the Physicai Education
Department Dr. Greg Reid, for his assistance in the finalization of this project.
Special prayers are offered to Dr. Vassilios G. Vardaxis for his expertise and
association in this work and to his wife Panayiota and their chikiren George, Petros and
Theodore.
Extra admiration is held for the participants of this study without whom the
current investigation would not have been realized!

ABSTRACT
The learning and retention adaptations of muscle activity were studied during a
novel multi-joint task. Electrornyographic (EMG) signals were recorded fiom the
posterior deltoid, long and lateral heads of the triceps, pectoralis major, biceps and
brachioradialis muscles. These data were assayed in a pattern recognition analysis (SVD)
to ascertain the minimum number of 'common features7 or waveforms (W's) required to
describe the set of input EMG patterns (IP).
Fifteen participants perforrned targeted arm rnovements, which incorporated the
shoulder and elbow articulations, as fast and as accurately as possible in the horizontal
plane. Both experimental (E) and control (C) groups were employed. The E group was
trained (Day 1 to Day 4) and tested both pre- and post-learning. They were further re-
tested during a retention period (ET) consisting of 1, 2, 4, 6 and 8 week post-learning
sessions. The C group was tested before and after leaming and at the end of the RET
period.
The SVD analysis revealed three W's among the six IP7s. The first W(1)
represented a "running average" of the iP's with a generally higher load for the lateral
head of the triceps across both learning (LRN) and RET conditions. The second W(2)
demonstrated an 'out-of-phase' relationship that defined an increase in reciproca1
inhibition between the agonist (AG) and antagonist (ANT) muscle groups within each of
the shoulder and elbow joints with LRN. The third W(3) described the relationships
among the muscles within each of the AG and ANT groups. The AG group revealed an
enhanced proximal to distal activation of the muscles with LRN while the ANT group
exhibited a decreased representation and/or intensity of the muscles involved in the motor
task for the same period.
The adaptations in both the intensities of the out-of-phase relationship (W2) and
the proximal to dista1 activation of the AG group (W3) demonstrated a persistence of the
motor memory program consolidated with motor skill acquisition at one-week post-
learning. At two weeks post-learning, however, a decrement in the aforementioned
adaptations was revealed by each of W2 and W3. This demonstrated a motor memory
program that was indeed impressionable to an interruption in specific motor task training.
v i

Les adaptations de l'activité musculaire concernant l'apprentissage et le maintien
d'une habilité motrice ont été éxaminés dans cet étude a propos d'une tâche multi-
segmentaire. Les donneés électromyographiques (EMG) ont été enregistreés pour le
deltoïde postérieure, le grand pectorale, les portions longues et vaste externes du triceps.
le biceps et le brachioradiale. Ces enregistrements ont été analysés dans une méthode de
reconnaissance de motifs principaux, nécessaires à décrire la série des traces EMG.
Quinze participants ont éxécuté des movernents du bras à une cible, incorporant
les articulations de l'épaule et du coude, aussi rapidement que possible dans le plan
horizontale. Les membres d'une groupe expérimentale (E) et de contrôle (C) ont été
employés. Le groupe E a suivi un entraînement de quatre jours et a été évalué a chacune
des sessions avant et après l'apprentissage (APPR) ainsi qu'à 1, 2, 4. 6, et 8 semaines
apres I'APPR [les étapes de maintien (MAINT)]. Le groupe C a été évalué avant et apres
1' APPR ainsi qu'à la fin de la période de MAINT.
L'analyse a révélé trois traits communs (TC) parmi la série de traces EMG. Le
premier trait commun (TC1) consistait de la moyenne des traces EMG avec un
coefficient préférentiel pour la portion vaste externe du triceps a travers toutes les
conditions de I'APPR et de MAINT. Le deuxième trait commun (TC2) a demontré une
élévation de l'inhibition réciproque entre les muscles agonists (AG) et antagonistes
(ANT) à Ia suite de I'APPR. Le troisième trait commun (TC3) a décrit les relations panni
les muscles dans chacun des groupes AG et ANT, révélant une activation rehauseé du
séquence proximale au distal des AG et une décroissance du nombre et/ou de l'intensité
concernant l'activation des ANT.
Les adaptations demontrés par TC2 et TC3 ont révelé une persistance du
programme moteur en mémoire qui a été consolidé avec I'APPR jusqu'a une semaine
apres la fin de l'entrainment. Cependent, à deux semaines apres l'entrainement, les
adaptations de TC2 et TC3 ont subi des détériorations qui ont indiqués que le programme
moteur était susceptible à affaiblir suite à l'inactivité.


Figure 18. Sample singular value decomposition analysis on EMG patterns for
a single participant, four weeks pst - leamhg (Ret 3). ......................... .42
Figure 19. Sample singular value decomposition analysis on EMG pattems for
.......................... a single participant, six weeks post-learning (Ret 4). -43
Figure 20. Sample singular value decomposition analysis on EMG pattems for
...................... a single participant, eight weeks post-leaming (Ret 5). ..44
Figure 21. Eigenvalue fluctuations and prominent muscle characteristics for W 1
of the experimental group.. ....................................................... .46
Figure 22. Eigenvalue fluctuations and prominent muscle characteristics for W 1
................................................................ of the control group.. 48
Figure 23. Eigenvalue fluctuations and intensity of the 'out of phase' relationship
............................... as revealed by W2 for the experimental group.. .50
Figure 24. Eigenvalue fluctuations and intensity of the 'out of phase' relationship
as revealed by W2 for the control group.. .................................... ...52

TABLE 1. Anthropometric characteristics for the participants of the
experimental and control groups-. .............................................. . .26
TABLE 2. Eigenvectors of each of the waveforms W1, W2 and W3, for al1 levels
of leaniing and retention for the motor task of participant P7.. ................ 57
TABLE 3. Eigenvalues, percent variabi iity accounted for by each wave form
and the sum of al1 three W's within both learning and retention levels
................................. for the motor task of the experimental group.. ..6 1
TABLE 4. Eigenvalues, percent variability accounted for by each waveform
and the surn of d l three W's across post-training and post-retention
periods of the control group.. ....................................................... 63

GLOSSARY OF DEFINITIONS AND ABBREVIATIONS
Basis Functions (BF's): Basis functions are also referred to as "common features" or
waveforms (W's) that are representative of the original input pattems to a singular
value decomposition algorithm.
Bic: Short head of biceps brachii.
BrR: Brachioradialis.
Composite Score: The difference between the highest and the Iowest points on an
eigenvector (W2) which provides an indication of the degree of latency between the
agonists and antagonists under investigation.
Coordination: Collectives of muscles and joints that are involved in the control of a
speci fic action (Bernstein, 1967).
EMG: Electromyogram.
Cross Motor Skiil: An action or task that is charactenzed by the utilization of large
musculature and smooth coordination of movement where the precision of
movement is not as important to its execution as it is for a fine motor ski11 (Magill.
1993).
LA: Lateral head of the triceps brachii.
LO: Long head of the triceps brachii.
Pec: Pectoralis major.
PD: Posterior deltoideus.
PT: Performance time.
Ret: Retention condition.
SVD: Singular value decomposition analysis that involves a reformulation of a set of
input patterns into a smaller set of basis functions which are derived fiorn the
original data. It was employed herein to evaluate the eigenvectors and eigenvaiues of
the input EMG pattems.
Whipping Movement: A movement that involves the rotation of al1 joints in a similar
direction.

INTRODUCTION
1.1 Gen eral Objective
Motor behavior is an integral part of our daily lives. Many of o w movement
capabilities are enacted to realize particular movement objectives within various
movement contexts, including walking, running, throwing, kicking, etc. These skills have
been acquired throughout our motor development by active practice. They have been
learned both as practical skills and through participation in recreational and/or sporting
endeavors.
Motor skill learning entails the selection and timing of many muscles across
multiple joints and thus the coordination of those joints (Brooks, 1986). An interesting
phenornenon of inquiry would not only concem the nature of motor skill acquisition but
also, and eq uall y as signi ficant, the consequences of terrninating or abstaining fiom
movement practice. In other words, would the acquisition of multi-joint coordination also
possess with it some element of "use it or lose it"? Just how resistant would complex
motor ski11 learning be to forgetting?
Bernstein's (1967) theory of motor coordination relates the problern of the
multitude of degrees of freedom encountered by the leamer in the acquisition of a new
motor skill. Specifically, the minimization of those degrees of fkeedom and the
development of efficient movement are what characterize motor coordination. It is the
purpose of this study to inquire into the reduction of degrees of freedom with leaming
and investigate the consequences of terminating movement practice on multi-joint
coordination.
Compound (rnulti-articular) limb movements, that are learned and adapted to a
particular activity or sport, reside within the central nervous system [CNS (i.e. brain and
spinal cord)] in the form of overall plans, narnely complex motor prograrns (Brooks,
1986). The neuromuscular transmission of these motor behaviors is mediated by the

motor unit, the final common pathway of neuromotor k c t i o n (Brooks, 1986).
Ultimately, both centrally and peripherall y processed commands are translated into force
by the effector organ, skeletal muscle (Vandervoort, 1 WZ), enabling the muscles
involved in a motor action to operate in a smooth and expedient manner. A decrement in
the motor performance of a previously acquired complex skiII, (due to the tennination of
practice or training), would be difficult to measure if changes first manifested themselves
at the neuromuscular level. Consequently, inquiries regarding the acquisition and
subsequent retention of multi-joint movement patterns would necessitate a
comprehensive analysis of neuromuscular function; that is, an analysis of performance
production rneasures (Magill, 1993).
Motor skill learning research has often utilized electromyography (EMG). as a
performance production measure, to study the neural activation of human skeletal muscle
within the context of single (Persons, 1958; Finley, Wirta and Cody, 1968: Payton and
Kelley, 1972) and multi-joint coordination (Normand, Rouillard and Tremblay, 1982;
Hasan and Karst, 1989; Karst and Hasan, 1991). The intermuscular latencies and
amplitude of EMG activity in the agonist and/or antagonist muscles under investigation
have been used to infer specific modifications in neuromuscular coordination consequent
to learning (Kamon and Gormley, 1968; Hobart, Kelley and Bradley, 1975; Vorro and
Hobart, 1981; Engelhom, 1983). Indeed, EMG measures are considered to be the
"gateway" or "window" to the various motor programs within the CNS of the individual
(Ange1 and Garland, 1971; Angel, 1975; Strick and Waters, 198 1 ; Jemings and Sanes.
1984).
Earlier work by Persons (1958), for instance, reported a reciprocal activation of
agonist/antagonist involvement in the development of a motor habit, which supplanted a
previously unorganized pattern of simultaneous activation concerning opposing muscles.
Later, in a pilot study, Payton and Kelley (1972) observed what appeared to be a
differentiation of two muscles involved in a motor skill, into both auxiliary (supporti~te)
and prime mover roles; and that the reduction in total muscle activation and duration of
the movement agonist responsible for the successful performance of the motor task
indicated a more efficient use of that muscle.

Vorro and Hobart (1981) who extended the work of Hobart et al. (1975). using a
unilateral ball tossing task, confirmed that with practice an increase in the total electricat
output of a rnovement agonist served to increase limb velocity at ball release while an
increase in the activity of the antagonist functioned to decelerate the limb, producing a
braking e ffect. The latency of both muscles had also decreased substantiall y following
practice underscoring as Ludwig (1982) himself has suggested, the importance of
intemuscular timing in motor ski11 learning.
A detailed electromyographical analysis of the retention aspects of a newly
acquired complex motor ski11 is still not entirely evident in the literature. Within the
context of the present study, the EMG changes in the agonist and antagonist muscles of a
previously l emed bi-articular motor task will be measured to assess the persistence of
neurornuscular multi-joint coordination.
1.2 Nature and Scope of the Problem
In motor learning research, the concept of 'motor memory' is used to underscore
the importance of a person's sense of effort and its memory in both the planning and
execution of motor action (Brooks, 1986). Specifically, motor prograrnrning functions as
an indelible neuromuscular foundation that is set in the progressive, systematic
performance of any motor act. What remains to be clarified, entails the characterization of
the tenability of the resultant motor schema as a fùnction of motor skill leaming.
in even the simplest motor behavior, mediated by the vertebrate CNS, the spinal
stretch reflex (SSR), or tendon jerk in the biceps or triceps brachii, has shown an adaptil-e
plasticity in primates wherein SSR amplitude can be changed without modifications in
initial muscle length or background EMG activity (Wolpaw, 1983; Wolpaw et al., 1983;
Wolpaw et al., 1986; Wolpaw, 1994).
Following a 60 day control mode, whereby the initial amplitudes of EMG activity
(in volts) were determined in the primates, Wolpaw et al. (1986) proceeded to either
increase (SSR-up mode) or decrease (SSR-dom mode) the magnitude of the response via
operant conditioning (for 35-274 days) of the SSR pathway [Le. the Ia afferent fiber from

the muscle spindle, its synapse on the alpha-motor neuron and the alpha-motor neuron
itself (Wolpaw, l986)J.
Specifically, a two phase learning process of both experimental modes was
identified, composed of an early precipitous, although small, modification in EMG
response followed by a more gradual, incremental change that ultimately accounted for
80-90% of the final change. Furthemore, the average SSR amplitude in the control mode
as well as in the SSR-down mode were shown to persist during a senes of gaps in
performance of 10-38 days suggesting a persistent aiteration at the segmentai level. An
increase in SSR amplitude as a result of training was found to decay slowly over weeks,
however. with a half-life of about 17 days.
Motor learning paradigms of simple, single-joint, voluntary Iimb movements have
consistently demonstrated a characteristic decrease in task error variance d u h g practice
leading to improved motor performance (Persons, 1958; Payton and Kelley, 1972;
Hobart, Kelley and Bradley, 1975; Hobart, Vorro and Dotson, 1978; Vorro and Hobart,
1981; Ludwig, 1982; Engelhorn, 1983; Corcos et al., 1993; Vardaxis, 1996). This
improvement, which according to Newell (1991) "is due to the acquisition of
prescriptions for action that speciS the movement dynamics in relation to the task
demands" also follows a curvilinear fhction in which rnovement stability is realized
early on in motor practice (Corcos et al., 1993; Vardaxis, 1996).
Information regarding the underlying myoelectric activity patterns comrnensurate
with enhanced performance due to motor training, however, has been equivocal in view
of the task and procedure specific nature of the separate investigations (Persons, 1958;
Payton and Kelley, 1972; Vorro and Hobart, 198 1 ; Engelhorn, 1983).
Motor ski11 practice has exhibited an increase, decrease, or no change in the
amplitude and/or duration of EMG activity in opposing muscles kom pre-learning to
post-learning trials (Payton and Kelley, 1972; Finley, Wirta and Codey, 1968; Payton,
Sue and Meydrech, 1976; Jeagers, 1989). More important, however, have been the
modifications in intennuscular latencies and electromechanical characteristics associated
with ski11 acquisition (Person, 1958; Hobart et al., 1975; Hobart, Vorro and Dotson, 1978;
Vorro and Hobart, 198 1 ; Ludwig, 1982; Engelhom, 1983; Corcos et al., 1993; Vardaxis,
4

1996). Additionally, there has also been evidence conceming the use of mental practice
along with physical practice to increase the rate of motor skill learning (Maring, 1990).
Until recently, the study of the enhanced performance of single- or rnulti-degree
of fieedom movement due to practice has provided data inferring the effects of abstention
fiom motor training on ski11 acquisition. Using such performance production measures as:
(a) angular or linear displacement, velocity and acceleration (kinematics), (b) forces,
torques and powers (kinetics) and (c) muscle activation patterns (electromyography) in
their analyses, researchers observed improvements in motor behavior during one
experirnental session that were partially retained and demonstrated at the start of the next
session (Corcos et al., 1993; Vardaxis, 1996).
Now, CO-extensive with investigations into motor learning are demonstrations of
motor skill retention, including the lasting temporal characteristics of intemal mode1 (IM)
consolidations of the dynarnics of an acquired motor task (Shadmer and Brashers-hg.
1996; Shadrner and Holcomb, 1997).
Furthemore, researchers' tests of critical time periods following practice reveal
an ongoing development of such M ' s from fragile to more stable representations in
motor rnemory (Shadrner and Holcomb, 1997; Brashers-Krug et al., 1996). There is also
evidence of the dynamic neural mapping associated with motor skill Iearning and
retention using functional magnetic resonance imaging (MN) and positron emission
tomography (PET) (Kami et al., 1995; Shadmer and Holcomb, 1996).
The present study investigated the adaptations in neuromuscuiar coordination
consequent to both learning and retention using electromyography to further the scope of
Bernstein's problem. If the CNS adopts strategies a d o r plans to minimize the number of
degrees of freedorn available to the learner mastenng a motor skill, then what would the
rarnmi fications to motor coordination be if movement practice were terminated?
A pattern recognition analysis of singular value decomposition (SVD) was used to
ascertain the minimum number of "common features" or waveforms required to describe
a set of six myoelectric signals of the left upper limb. This type of analysis will permit the 5

evaluation of the muscle activation patterns in their entirety and will entai1 the
interpretation of the temporal characteristics andlor the phasic information of the latter
whic h are O ften overlooked in discrete time-characteristic analyses.
1.3.1 Assumptions and Limitations
The limitations associated with the collection of surface electromyographic signais
include:
1. "Cross-talk" artifacts fiom adjacent muscles. in the present study, carefûl
exarnination for the detection sites was effected as well as outlining the same
locations for use across experimental sessions in order to minimize such artifacts.
7 . S kidelectrode irnpedance. Electrodekkin interface impedance was minimized
with thorough cleansing procedures involving shaving and mild abrasion of the
selected recording sites followed by alcohol swabbing.
3. Signal contamination consisting of prevailing noise from power line hum and
movement artifacts. Appropriate filtering and processing techniques were used to
control such influences.
Assumptions asociated with the experimental design and the motor task.
1. The EMG data will be asssumed to provide a measure of neurornuscular function
and a 'window' to the motor learning program(s) of the motor task.
3 -. It was assumed that the EMG data collected throughout the learning and retention
conditions of the experimental period represent the resulting adaptations of each
participant at the given interval as a function of the level of practice experienced
up to that period.
3. Each trial was assumed to have been accomplished as quickly as possible,
according to the outlined accuracy requirements of the motor task.

Hypoth eses
The SVD analysis will provide more than one significant waveform that will be used
to explain the six input muscle activation patterns of the experimental group across
both learning and retention conditions.
The waveform analysis will reveal more than a simple amplitude modulation of
muscle activity patterns within both learning and retention levels. It will characterize
the phasic adaptations between the agonist and antagonist muscle groups to define a
degree of reciprocal inhibition within the joints involved in the rnotor task.
The waveform andysis will characterize the phasic adaptations among the musctes
within each of the agonist and antagonist groups fiom pre- to post-learning
conditions. It wilI outline an increase in the proximal to distal relationship of the
agonists as well as an increased synergistic activation of the antagonists with
learning.
The waveform analysis will characterize the phasic adaptations between the agonist
and antagonist muscle groups to define a decrease in the reciprocal inhibition within
the joints involved in the motor task with a termination in training.
The waveform analysis will characterize the phasic adaptations among the muscles
wititin each of the agonist and antagonist groups after training has stopped. It will
demontsrate a diminution in the power of the proximal to distal pattern of the
agonists as well as a decrease in the synergistic activation of the antagonists.
Rationale
The study of multi-joint rnovement is warranted on the basis that many of our
everyday rnotor activities, recreational and sporting preserves, involve the coordination of
various muscles across many joints. Our actions are inherently complex, including the
arrangement of serial single-joint movements in the production of motor skills. The
investigation of singular muscles or joints, although informative, is not entirely practical.
Furthemore, the fact that motor ski11 learning occurs is an attestation to the
possibility that "what c m be gained may also be lost". Consequently, it was the focus of
7

the present research to inquire into the retention phenomenon as it applied to the
production of a complex rnotor task. In order to study motor ski11 retention, the issue of
learning must first be addressed; the function of which would be to establish a
comparative baseline performance level for the subsequent analysis of post-learning
adaptations. In this study, an experimental paradigm was employed to facilitate the
acquisition of a fast complex whipping movement. The neuromuscular correlates of
motor memory (i.e. the motor prograrns) were indirectly assayed via the
eIectromyographic activity of the various muscles subserving the movement to provide a
'gateway' into the underlying processes of the CNS.
The effects of an interruption or cessation in motor ski11 training have received
substantial attention among researchers in a variety of disciplines. These studies have
included explorations in pursuit rotor and tracking tasks of experimental psychology
(Bell, 1 950; Jahnke, 1 958; Hammerton, 1 963), strength training/detraining paradigms in
exercise physiology (Hakkinen and Komi, 1983; Hakkinen et aI., f 985) and
reachinglpointing movements of motor control (Shadmehr and Brashers-Kmg, 1 996).
However, there has been a limited focus, if any, on the retention of 'maximum effort'
type movements of curvilinear trajectories that are inherent in many complex athletic
skills such as throwing, kicking and striking with and/or without an implement. Although
these skills possess significant force distribution characteristics. they are also defined by
salient timing features of segmenta1 rotations which allow the practitioner of a motor ski11
to perform in an efficient manner that is distinguished by an absence of superfluous
actions-
The impressionability of the aforementioned timing features to imposed no-
training intervafs, is very important if the goal is to maintain a certain level of task
proficiency or to return to a previously acquired level of performance. Knowledge of the
time course of pertinent pauses in training would permit an informed decision on the
restructuring and implementation of training programs to realize resumption in a prior
motor leaming status.

CHAPTER II
METHODOLOGY
Each of the experimental sessions, including measurement of anthropometric data,
task proficiency testing and re-testing following the non-performance penods took place
in the exercise physiology laboratory of McGill University's Seagram Spon Science
Centre. The methods and procedures of this study will be delineated within the ensuing
sections: (2.1) Participants; (2.2) Apparatus; (2.3) Expenmental task and protocol; (2.1)
Data acquisition and analysis; and (2.5) Statistical analysis.
In this study, fifteen male volunteen between the ages of 18 and 30 years, with no
history of neurological or physical disabilities, were randomly assigned to either an
experimental (10) or control (5) group. Each person's age. height and body mass were
recorded for descriptive purposes.
A pre-requisite met by al1 participants was the determination of their right
arm/hand preference in normal daily as well as sporting activities, the function of which
was simply to provide each individual the goal of having to perform and subsequently
l e m a unilateral bi-segmental motor task with the non-preferred arm (i-e. the Iefi). For
this purpose a questionnaire (Appendix 2) was submitted to each prospective subject to
better establish strongly lateralized (Le. right-handers) individuals and ovemile
participant "ambidexterity".
A description of the expenmental task and protocol (Appendix 3) was provided to
each prospective participant before an infonned consent form was read, understood and
signed by the participants pnor to the start of the study (Appendix 4). Finally, a statement
of ethics approval was received prior to the start of subject data collection (Appendix 5 ) .

2.2 Apparatus
All training and testing of the study's participants occurred via a specially
designed experimental table. Each individuaI was seated in a straight-backed, armless
chair with the trunk of the body secured to the back of the chair using velcro straps so as
to minimize any extraneous movement of the shoulder joint. The chair was located on the
same floor space for each person and was adjustable, allowing each subject to be in the
same relative position when performing the required skill. More specifically, the surface
of the table met the torso of the individual below the axilla and allowed resting of the
subject's upper limb on it in between the trials and blocks of practice, see Figure 1A.
A plastic splint, extending the length of the forearm and pronated hand and
terminating distally (beyond the fingerç) in a circular lOcm - diameter plate, was h l y
secured to both the forearm and the hand via velcro straps. This procedure immobilized
the wrist, restricted movement to the elbow and shoulder joints only, and defined the
circular plate as the end-point target of the upper extremity.
Two photodiodes were affixed to the experimental table designating the starting
and stopping points of the task and will otherwise be referred to herein as the "home" and
"target" areas respectively. Each photodiode consisted of an emitter and a receiver which
established a narrow uniform beam when set, and depending on which beam was
intempted, the output of the photodiodes consisted of either O or 5 volts. A chronometer
to which the photodiodes were wired provided a ruming time count of the limb's
excursion fiom the "home" to the "target" positions.
2.3 Experimental Task and Protocol
Originated by Vardaxis (1996), the motor task that was performed consisted of a
two-joint "a11-out" unidirectional whipping movernent of the lefl arm involving
horizontal abduction at the shoulder and extension of the elbow, both occuning in the
transverse plane. The task was chosen on the b a i s of it being a novel, gross motor skill;
although not too difficult to perform, it required practice to master. Moreover, the task
was neither akin to any normal everyday movement nor to a specific sporting activity or
10

Figure 1 : A schematic representation of the experimental set-up. (A) side view (B) top view.

skill. It did possess similar characteristics to such movements as striking and kicking with
the limbs and/or with implements, for instance, in that both c m be executed with relative
maximum effort, and to an extemal target. However, these movements would involve
different muscle groups, alternative planes and ranges of motion as well as contact with
objects as targets.
To begin with, a 'ready' or 'still' position was defined with the subject fitted to
both the chair and forearmhand splint, his arm in 90 degrees of shoulder flexion and 90
degrees of elbow flexion, with the circular end-point plate intersecting the beam of the
"home" photodiode. The subject was instmcted to move his hand fiom the "home" to the
"target" photodiode position, which could only be accomplished by a 45 degree
horizontal shoulder abduction and 180 degree elbow extension, as quickly as possible, see
Figure 1 B. The two photodiodes were adjusted to accommodate the variable segmental
lengths of each of the participants and ensure the same angular displacement for the two
joints during task execution.
The movement did not consist of a reaction time task. Prior to each trial the
experimenter prepared each subject with the phrase "Whenever you are ready ...".
whereupon the latter moved his am, to accomplish the task, on his own initiative and
without undue hesitation. A 'final' position was defined by the end-point plates' complete
stop at and intersection of the "target" photodiode beam.
With the arm segments (Le. brachiurn and fore-) operating in the horizontal
plane and rotating around the longitudinal axes of each of the glenohumeral and elbow
articulations, the following single and double joint agonist muscles were assayed: the
posterior deltoideus (single-joint horizontal shoulder abductor) and the long and Iateral
heads of the triceps brachii (double-joint horizontal shoulder abductor/elbow extensor and
single-joint, elbow extensor respectively).
The single and double-joint antagonist rnuscies consisted of: the clavicular
pectoralis major (single-joint, horizontal shoulder adductor), short head of the biceps
brachii (double-joint, horizontal shoulder adductor and elbow flexor), and brachioradiafis
(single-joint, elbow flexor).

Since motor skill learning has been demonstrated to occur even after many
hundreds of repetitions (Nomand et al., 1981; Gottlieb et al., 1988; Corcos et al., 1993)
the protocol selected was similar to that used by Vardaxis, (1996) which was shown to
facilitate complex motor skill acquisition. The experimental group performed three (3)
sets of twenty-five (25) trials, for a total of seventy-five (75) trials per day on four (4)
consecutive days. There was a 24 h o u period between testing sessions and each session
was undertaken at the same time of day for each individual to minimize any diurnal
change in performance (Corcos et al.. 1993).
Each set was intempted by a 5-min. rest period in an effort to minimize fatigue.
They were also allowed to rest between trials whilst receiving performance feedback
from the experimenter. The specific feedback was verbal, relating to both the movement
time and accuracy demands of the task as observed by the experimenter on a cornputer
monitor at the end of each trial. Performance time (PT) consisted of the elapsed time
behveen the circular end-point plate's departure from the "home" position (movement
onset) to its arriva1 at the "target" point (movement end). The accuracy demands of a
successful trial entailed the single intersection of each of the "home" and "target"
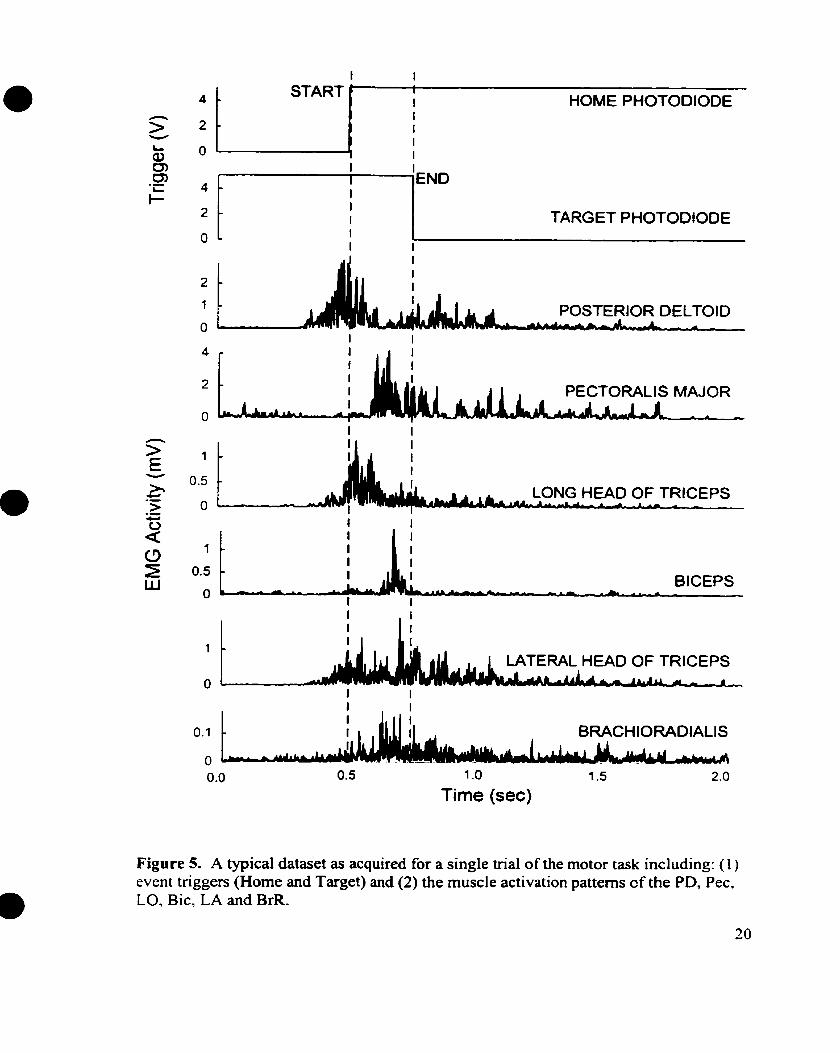
photodiode beams by the circular end-plate, see Figures 1 B and Figure 5.
Eac h individual of the experimental group was accorded ten (1 0) familiarization
trials without feedback on the first testing day. This process acquainted him with the
procedures of the task as well as with the experimental apparatus; these trials were
discarded at the time of the analysis. Subsequently, ten (10) preparatory, or "wm-up".
trials without feedback were permitted prior to the actual practice trials of the first and at
the beginning of each of the remaining practice sessions on succeeding days.
In addition to being used to prepare the individual for an optimal level of
performance, the latter five of the ten wann-up trials for each of the four practice sessions
constituted the learned trials or the 'stable' adaptations of the task (Le. the aspects of the
motor task which had been retained). Evidently, on the first day of practice, these five
stable adaptation trials consisted of an initial performance level in the motor task. The last
five of the practice trials of each of the aforementioned training days comprised transient

learning, which featured the level of task proficiency attained as a result of training for
that particular day.
Also included was a fif3.h day incorporating ten (10) successful trials of which the
latter five trials were used to determine the final stable adaptations of the motor task or
the resultant leaming level as a function of the four-day training protocol. Only the two
sets of five stable adaptation trials f?om Day 1 and Day 5 fiom each individual were used
in the ensuing analysis, to determine the learned aspects of the rnotor task. Transient
learning will not be addressed in this study. A complete representation of the
experimental protoc01 is offered in Figure 2.
It should be noted, however, that the greater the number of testing trials required
before actual practice in the task, the greater the conceivable index or gauge of learning
which can be procured fiom the ensuing training mals (Corcos et al., 1993). The risk that
is run, though, is one that exacts fûrther pressure on the patience of the subjects
cornmitted to the study, who may already be performing to the limits of their discretion.
Consequently, the nurnber of warm-up trials used in this study was determined according
to the above contention as well as in view of the familiarization and pre-training
procedures used by Vardaxis, ( 1996).
Motor skill learning or task proficiency was determined by the following
performance outcome criterion: a decrease in the performance time of each person. to
within a stabilized time period. Following the training protocol, there was a 'time-course'
method used to evaluate the effects of 'no-training' or 'no-practice', consisting of
increasing retention intervals. It was systematized in a manner that required the
expenmental group to abstain fiom the task for an initial one week period (starting fiom
the end of Day 5 to the following seventh day - Ret 1) at the end of which al1 returned for
a re-testing session of 10 successfûl trials. Once again, the first five attempts constituted
warm-up trials while the latter five stable adaptation trials were used in the analysis. The
participants desisted tiom practicing the task for another week followed by the same re-
testing protocol at this, the second week post-learning (Ret 2). They underwent a similar
re-testing procedure at 4, 6 and 8 weeks post-leaming (Ret 3, Ret 4 and Ret 5
respectively). Only trials 6-10 of each of the five retention intervals (i.e. the 5 stable
14

[I F A M I L I A R I Z A T T PRA CTICE I TRANSIENT 1
II Trials: 1
5 1 5 Il
Figure 2: The experimental protocol for leaming of the motor task. The farniliarization, preparation and stable adaptation trials did not include feedback. Stable learning represents the adaptations consolidated in memory. Transient learning which included feedback reflects the practice effect within experimental sessions. Transient learning was not assessed in this study.
15

adaptation trials at 1, 2, 4, 6 and 8 weeks post-leaming) were analyzed. These p ~ i c u l a r
time penods were chosen to represent two short-term (Ret 1 and Ret 2), and three long-
t e m (Ret 3, Ret 4 and Ret 5) 'non-performance' possibilities, in order. An overview of
the retention protocol is available in Figure 3.
The control group performed 10 familiarization trials without feedback followed
by 10 additional 'successful' trials on the fint day o f the learning period of which the
latter 5 of the 'correct' trials were analyzed. As well, each control subject performed 10
successhil tnals of the task on the fifth day of the learning penod and at the completion of
the expenment; on the day of the fifth retention interval. see Figure 4. Only the latter 5
tnals of each of the three conditions were subjected to the ensuing analysis.
Al1 subjects were requested to abstain from either performing the experimental
task privately (Le. on their own) or engaging in any progressive resistance type training of
the upper limbs and trunk during the entire experimental period. Furthemore. they were
asked to keep a log of their physical activities and sports participation (recreational or
othewise) during each of the retention periods for the expenmenter's information.
2.4 Data Acquisition and Analysis
During the experimental penod the electrical output of the muscles under
investigation was recorded using disposable, uni-patch dual-element silver/silver chloride
surface EMG electrodes. These bi-polar electrodes were placed near the enervation points
and onented longitudinally according to the muscle fiber direction of each of the six
muscles (Warfel, 1993; Kearney, 1994). In order to ensure the same electrode placement
for al1 seven testing sessions (Le. Day 1, Day 5 and Ret's 1 - 5 ) , the specific sites were
marked with a non-toxic permanent ink marker. A ground electrode was located on the
stemal extremity of the nght clavicle.
To minimize electrode impedance and decrease the skin potentials at the
elec trode/skin interface sites, the surface areas used to monitor muscle activi ty were
shaven of epidermal hair, abraded of dead skin cells and cleansed by alcohol swabbing
(Kearney, 1994). Care was also taken regarding electrode placement to avoid the

PROTOCOL WTFJVTION)
P R E ~
1 PREPARA TION
Trials: 5
PREPARA TION Trials: 5
II Trials: 5 II
5
STABLE 5
11 P R E P A R A T ' N 1 STABLE 1 11 Trials:
L
5 I 5 II
Figure 3: The experimental protocol for retention of the motor task. The preparation and stable adaptation trials did not include feedback. Stable learning represents the adaptations consolidated in mernory.

PREPARA TION STABLE Trials: 5 5
c
F A M ~ ~ STABLE I>
PREPAM TION STABLE Trials: 5
Trials: 15
Figure 4: The control protocol including matching intervals for Day 1 (before learning), Day 5 (after learning) and Ret 5 (8 weeks post-learning).
5

potential of "cross-talk" or interference in the signal of one muscle from adjacent muscles
(Kearney, 1994).
Al1 six 'raw' EMG signals were recorded via an eight channel GRASS High
Performance AC Preamplifier (mode1 p5 1 I ). Powered by a regulated power supply it
included a 60 Hz notch filter, 20 Megaohm input impedance amplifiers. 4 microvolt
sensitivity and a Cornmon Mode Rejection Ratio (CMRR) adjustable to 10 000:l at 60
Hz. Individual signals were differentially arnplified with a gain of 1000, and band-pass
filtered with the high and low cut-off fiequencies set at 10 and 1000 Hz respectively.
The EMG data along witb the two photodiode signals were then passed through
eight, gain and phase-matched, pole Bessel analog low-pass active anti-aliasing filters
with a cut-off frequency of 250 Hz (Frequency Devices). The conditioned signals were
digitized at a sampling rate of 1000 Hz by an analog to digital board. They were
represented as both raw and full-wave rectified EMG waveforms on a computer analog
display using Labview: a prograrn development application that uses graphical
programming language, (G), to make programs in block diagram form. The program. or
virtual instrument (VI), that was accessed employed a conditional retrieval mechanisrn to
speciQ a software trigger, which started the acquisition and made the data availabie while
the VI continued to operate. By design, a11 six of the muscle activation waveforms fiom
each trial, including both photodiode signals, were observed in a two second acquisition
-window' including a 500 ms pre-trigger time, prior to being saved on computer
diskettes, see Figure 5 .
Further analysis of the stored EMG records entailed the smoothing of individual
EMG patterns using a fourth order Butterworth low-pass digital filter applied in both
directions to avoid any phase shift (Vardaxis, 1996). Additionally, the EMG data were
normalized for performance time (PT) in such a way as to include an 80% PT period
before movement onset as well as a 20% PT period afler the temination of the
movernent.
The time base of each resulting EMG trace was then resarnpled in MATLAB, a
rnatrix laboratory computer software prograrn (version 4.0), to 100 points in total, taking
I I 4 - 2 - PECTORALIS MAJOR
O * * -
f I
s 1 - I I E -
0.5 - Y *-
LONG HEAD OF TRICEPS > A -- O
4 - > 2 - w
.- Y
2 I
C3 1 - 0.5 -
W BICEPS
1 * LATERAL HEAD OF TRICEPS
O I
- START
0.5 1 .O 1.5
Time (sec)
i 1
1 HOME PHOTODIODE I f
O 5
Figure 5. A typical dataset as acquired for a single trial of the motor task including: ( 1 ) event triggers (Home and Target) and (2) the muscle activation patterns cf the PD, Pec, LO, Bic, LA and BrR.
I I
CT) 1 I END
TARGET PHOTODIODE
.- al 4 : t=
2
1 I
I I
- 1 I
O - I

into account both of the pre- and post-performance time provisions resulting in a data set
consisting of:
Motor task = 80% PT +- PT + 20% PT
100 points = 40 pts. + 50 pts. + 10 pts.
(Vardaxis, 1996). The resarnpling procedure did not change the nature of the individual
EMG traces, rather, it enabled the temporal evaluation of al1 the EMG pattems on an
equal time line. Every processed signal was also nomalized to peak ( 1 00%) amplitude,
see Figure 6. Further processing of the stored EMG records entailed averaging the five
trials for each of the six different muscles, fiorn each of the learning (Day 1 and Day 5)
and retention (Ret's 1, 2, 3, 4 and 5) levels, for each subject of the experimental group.
The five trials fiom each of the three testing sessions of the control group. as deheated
above, were prepared in a manner consistent with that of the expenmental group.
The averaged and smoothed data that were performance time nomalized were
then quantitatively assessed using Singular Value Decomposition ( S m ) analysis, also
known as Principal Component (PC) analysis. SVD is a mathematical technique that can
facilitate the quantification of specific component contributions to myoelectric patterns.
among other electrophysiological phenornena (Flanders, 199 1 ). Specifically. the analysis
involves a reformulation of a set of input patterns into a smaller set of basis functions
(BF's) which are derived fiom the original data. In this manner, the patterns of different
experimental conditions c m be compared or contrasted.
Each bais fùnction, also referred to as a "common feature" or simply a waveform
(W). is orthogonal in relation to other BF's in that the dot product or covariance of any
two W's is equivalent to zero. The W's of the SVD analysis are not sinusoidal however;
insteâd this analysis is more like Fourier analysis since each of the original input patterns
c m be reconstnicted as a weighted sum of the W's. The construction of an auto-
correlation matrix enabled the calculation of the W's. The eigenvectors of the
syrnmetncal matrix consisted of the W's among the input patterns while the eigenvalues
signified the energy contained in each of the waveforms. Consequently, a waveform will
be descnbed herein as a signal that is representative of al1 of the input pattems analyzed,
which possesses a rneasure of variance accounted for by (Le. the eigenvalue). If, then, al1
2 1

of the waveforms are used with respect to an equal number of input pattems, al1 of the
variance cm be accounted for (Le. 100%).
The aim of the SVD analysis was to determine the minimum number of
waveforms required to describe the EMG data for each of the leaniing and retention
conditions of the experiment. That is, how the eigenvalues and eigenvecton of the
analysis changed with learning and, more importantly, following periods O f abstinence
fiom motor task training. An evaluation of the complete myoelectric signais focussed on
the phasic characteristics of the data to elucidate the temporal patterns of muscle
activation.
Numerous researchers have employed singular value decomposition analysis, also
known as principle component analysis, as a technique to quantify the pattems of muscle
activation associated with movements of the upper and lower extremities (Patla, 1985;
Soechting and Lacquanity, 1989; Flanders, 199 1 ; Flanders and Hermann, 1992).
Furthemore, investigators have recently used SVD in the evaluation of movement
synergies employing kinematic data in hurnan locomotion (Mah et al., 1994). the anaiysis
of cerebral activity fiom a multi-channel electroencephalograrn (Lagerlund. Sharbrough
and Busacker, 1997), and in the extraction of repeating pattems in cyclic biomechanical
data (Stokes, Lanshamrner and Thorstensson, 1999).
The singular value decomposition analysis was executed using the SVD algorithm
contained in MATLAB (version 4.0). The analysis was initially performed for each of the
learning levels in this study, using the myoelectric patterns of the six different muscles,
for each participant of the experimental group. That is, a set of six input pattems from
each of Days 1 and 5 per individual. tt should be noted that each EMG trace represented
an average of the corresponding five trials for each of the two learning levels that was
normalized to both percent performance time (100%) and peak amplitude (lOOOh), (see
Figure 6). The analysis was used to determine any changes due to learning after the shape
of the waveforms, the eigenvalues and eigenvectors were evaluated.
SVD was also performed for each of the five (5) post-trainingketention levels by.
once again, using the six different muscle input pattems for every subject of the
experimental group. In this manner, any changes within the 'non-performance' intervals
22

100 -
1 O0
40 Pectoralis Major 20
O
80 60 40 20
100
Percent Performance Time
- - Posterior Deltoid - -
80 60 40 20
Figure 6. Sample averaged and smoothed EMG waveforms for al1 six muscles that have been normalized to both penormance time (100%) and peak ampiinide (1 00%) before input to the singuiar value decomposition analysis.
O =
- - - Lateral Head of Triceps -
O

would be demonstrated. Fwtherrnore, cornparisons between the analyses of both the
learning and retention levels conceming the changes in the activation patterns of al1 the
muscles at once was also possible; specificaliy in terms of the timing characteristics of
the myoelectric signals.
Equally, SVD was employed on the EMG signals of the six muscles of each of the
subjects of the control group on the first and f i fth days of the learning period as well as on
the day of the fifth retention period.
2.5 Statistical A nakysis
A two-way repeated measures univariate analysis of variance (ANOVA) was used
to compare the expenmental and control groups, at three points in time of the expenmetal
period: (1 ) the first day of practice, Day 1, (2) the final day of testing in the fearning
protocol, Day 5, and (3) on day of the final retention interval, RET 5. The single
dependent variable that was analyzed included the performance outcome measure of
performance time.
Additionally, a repeated measures one-way analysis of variance was conducted on
the experimental group to identiw any changes across the five levels of practice or
training and five retention intervals using, once again, performance time as the dependent
\ x i able.
SVD analysis will be presented qualitatively in this study.

RESULTS
The present study investigated the neuromuscular adaptations consequent to both
Ieaming and retention conditions of a novel muhi-joint task. The linear envelope detected
EMG profiles (waveforrns) were used in an SVD analysis, as an indirect measure of
neuro-motor function, which aIluded to the motor prograrns of the C N S . The aim was to
elucidate the changes occurring with motor task practice and more specifically, to
determine the effects of abstaining fiom such exercise. A time course evaluation of motor
ski11 retention was employed to test the stability of the resultant motor coordination as a
function of motor ski11 leaming. The topics conceming this chapter wiIl be subdivided
into the following sections: (3.1) Participant characteristics, (3.2) Adaptations in
performance outcome with learning and retention and (3.3) Adaptations in performance
production during both learning and retention.
3. I Participant Characteristics
Fi fteen male volunteers agreed to participate in the present study, without
remuneration, following an explanation of the experimentai task and protocol and the
signing of an informed consent form. Ten individuals were randomly assigned to an
expenmental group while five others were likewise allocated to a control group. They
reported no persona1 injuries or prior history of neurological, motor system, disorders. All
participants identified themselves as right arm preferred for the motor tasks listed within
the questionnaire. Their ages ranged from 23 to 34 years, with a mean age of 26.6 years.
The heights and body masses of the participants ranged fiom 1.65 to 1.85 m and 67.3 to
106.8 kgs respectively. Table 1A displays the age, height and body mass for each
participant of the experimental group. Table 1B shows the same data for the members of
the control group.

TABLE 1
A. An thropometric charactenstics for the participants of the experimental group.
Participant Age (Years) Heigbt (m) Body Mass (kg)
Mean (* SD) 26.6 (* 2.91) 1.76 (* 0.07) 76.7 (& 12.00)
B. Anthropometric characteristics for the participants of the control group.
Participant Age (Years) Height (m) Body Mass (kg)
- - - - - - - - - -
Mean (* SD) 26.4 (I3.29) 1.77 (kO.08) 78.3 (*4.27)

3.2 Adaptations in Performance Outcome with Learning and Retention
Performance tirne, (PT), was the single dependent variable used to evaluate
performance outcome on the motor task by the participants of the experimental and
control groups, within their respective learning and retention levels. Al1 participants
executed each trial with maximum effort and according to the accuracy demands of the
motor task. A one-way repeated measures ANOVA with tests of within-subjects contrasts
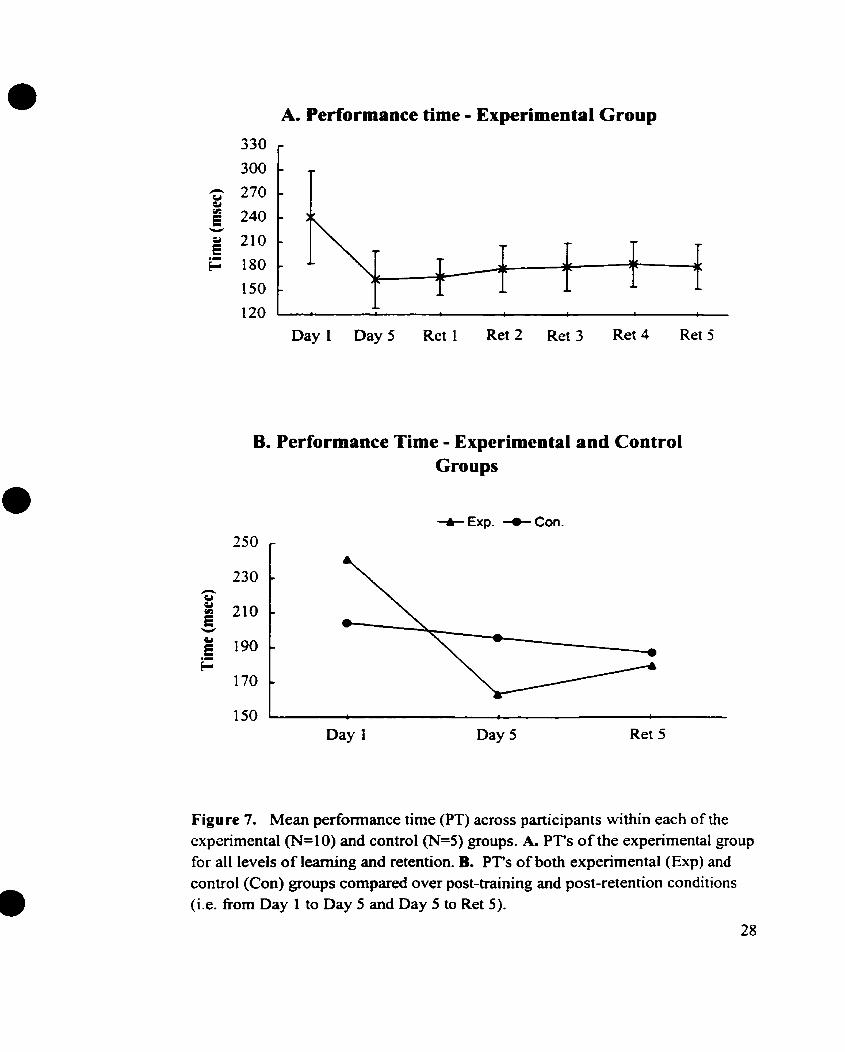
applied to the data of the experimental group revealed a significant decrease in
performance time (i-e. 47%) across participants, from pre- to post-learning trials (Le. Day
1 to Day 5) . F (1,9) = 29.3, p<0.0001, as shown in Figure 7A. That is. with learning. each
member OJ=l O) was able to perform the movement faster without sacrificing the end-
point accuracy conditions of the motor task. The slight increase in performance time from
Day 5 to one-week post-learning (Ret 1) was not significant, F (1,9) = 0.2, p<0.649,
indicating that the performance tirne of the experimental group had attained a plateau at
this retention interval. However, the change in performance time from Ret 1 to Ret 2 was
significant, F (1,9) = 10.4, pc0.011, and attested to the fact that the members were
slowing d o m in their movements to the target with two-weeks of no-practice. There
were no other significant differences in the performance times of the group beyond Ret 2,
as demonstrated by the pair-wise cornparisons of Ret's 2-3, 3-4 and 4-5, see Appendix 6.
A two-way repeated measures ANOVA, applied to the 'pooled' data of both the
experimental and control groups demonstrated significant changes in performance time
across post-training and post-retention conditions, F (2,26) = 13.4, pc0.0001, as well as
significant interactions of the data over the same time periods, F (2'26) = 7.3, p4.003
(see Figure 7B). The tests of within-subjects contrasts revealed significant decreases in
PT for both groups fiom Day 1 to Day 5, F (1,13) = 18.2, p<0.001 and a significant
increase and decrease in PT for the experimental and controI groups respectivsly, from
Day 5 to Ret 5, F (1,13) = 8.625, pc0.012. The rate of the decrease in PT fiom pre- to
post-learning conditions for each group was found to be significantly different, F (1.13) =
5.8, ~ ~ 0 . 0 3 2 , with the experimental group showing a much greater index of change (Le. a
47% decrease) than the control group (Le. a 4% decrease). Additionally, the rate of the
A. Performance time - Experimental Group
r
Day 1 Day 5 Ret 1 Ret 2 Ret 3 Ret 4 Ret 5
B. Performaoce Time - Experimental and Control Groups
- -- Day 1 Day 5 Ret 5
Figure 7. Mean performance time (PT) across participants within each of the experimental (N=10) and control (N=5) groups. k PT's of the expenmental group for al1 levels of leaming and retention. B. PT's of both experimental (Exp) and control (Con) groups compared over post-training and post-retention conditions (i.e. fiom Day 1 to Day 5 and Day 5 to Ret 5).
28

changes for each of the groups fiom post-training to post-retention conditions was also
significant, F (1 ,l3) = 8.8, p<0.011. This dernonstrated that the experimental group had
slowed down by the end of the retention period whereas the control group exhibited an
increase in movement speed over the sarne interval, (see Figure 7B and Appendix 7).
3.3 Adaptations in Performance Producrion during both Learning and Retention
EMG measures were acquired fiom the study's participants to provide a
neuromuscular performance index, in ternis of the specific coordination of the muscles
selected as producers of the motor task. The resultant adaptations in motor coordination
due to both leaming and retention conditions would thenceforth provide some recourse to
the rnotor programs of the CNS. An examination of the EMG data in this section will
include a preliminary qualitative interpretation of the muscle activation wavefoms. This
wilI be followed by an in-depth report of the changes in the EMG patterns of the agonist
and antagonist muscle groups within each leaming and retention level using a method of
SVD.
3.3.1 Qualitative EMG Data Analysis
The EMG records employed in the qualitative data analysis consisted of the
ensemble averaging of five trials nonnalized to time base, that is 100 data elements (note:
for the qualitative interpretation, whose focus was purely descriptive, the EMG signals
were presented at each of their non-normalized amplitudes). Of those, elements 40 to 90
relate to the initiation (Onset) and termination (End) points of the motor task respectfully.
Figure 8 represents the linear envelopes corresponding to the agonist (PD, LO, LA) and
antagonist (Pec, Bk, BrR) muscles as exemplified by one participant, P7, before leaming,
on Day 1. The latter muscles are presented as inverted curves in the figure and paired
with their respective agonist counterparts for purposes of clarity as well as to underscore
their antagonistic or reciprocal nature in the motor task. The trials represented within the
figure, proceeded without feedback and followed both the farniliarization and preparation
(i.e. wam-up) penods. The solid line of each plot represents the mean signal whiie the

Motor Task for Participant P7
A. Before Learning (Day 1
END
B. After Learning @ay 5)
Posterior Deltoid r
Pectoralis Major (f ec)
Long Head
Lateral Head of Triceps
I ONSET END - -
Brachioradialis -1 ,b 1
1 1
61 91 1 31 61 91
Percent Performance Time
Figure 8. A sample plot of ensemble averaged (5) muscle activity patterns for the motor task of participant P7. The solid line line represents the mean pattern while the thin vertical lines demonstrate (*) 1 SD. A. Before learning, Day 1. B. After learning, Day 5 .

thin vertical bars demonstrate *1 SD. Also included in the figure are the EMG wavefoms
for the same muscles on the fiAh day @ay 5), following the learning schedule. These
trials were likewise cornpleted without performance feedback by P7. They clearly reveal
that learning was consistent with EMG wavefoms of greater amplitude with more phasic
and distinctive bursts. in comparison to the patterns manifested before learning, the well-
practiced movements were characterized by steeper slopes of the tising EMG signals
concomitant with initial b m t peaks that were located or displaced, earlier in time with
respect to movement onset.
Figures 9-1 3 inclusiveIy, compare the rectified and fiItered EMG data of both the
agonist and antagonist muscles across the post-learninghetention periods for the same
participant. An evaluation of these data demonstrated that at one week post-leaming (Ret
1) there was a decrease in the amplitude of the muscle signals, unlike the greater
amplitude charactenstics of the same EMG patterns observed on Day 5. However, an
additional phase shift of each of the agonist muscles, earlier in time, with respect to
rnovement onset, indicated that there were adaptations consequent to the learning
paradigm which continued to evolve for this individual.
At two weeks post-learning (Ret 2) the agonist muscles for P7 appeared to gain in
amplitude, except for the LA. Nevertheless, these muscles also demonstrated a shift in
phase. later in time, with respect to movement onset, which suggested that they were
being activated at a later time in comparison to Day 5. These changes alluded to a
decrement in the adaptations seen with learning and which had persisted to Ret 1. The
antagonist muscle activation patterns in tum appeared to lose some amplitude in
cornparison to the waveforms of Ret 1. By four weeks post-learning (Ret 3) the PD was
similar in character if not of slightly greater amplitude than at Ret S. Meanwhile. the
triceps muscles were observed to shifi still later in tirne, with the LO decreasing in
amplitude, unlike the LA, which demonstrated an increase in the sarne EMG burst
parameter. The antagonist muscies appeared to be similar in nature as in Ret 2; that is
they had not changed much in terrns of intensity or displacement.
At six weeks post-learning (Ret 4), the agonist muscles showed a slight nse in
amplitude with little shift in phase. The antagonist patterns of the Pec and BrR may seem
3 1

Motor Task for Participant Pl
-4. After Learniog (Day 5) B. One Week Post-Learoing (Ret 1 )
Posterior Deltoid
3
Pectoralis Major
(Pet)
Long Head of Triceps
@O)
Biceps (Bic)
1 Lateral Head
61 91 1 31 61 91
Percent Performance Time
Figure 9. A sample plot of ensemble averaged (5) muscle activity patterns for the motor task of participant P7. The solid line represents the mean signal while the vertical lines demonstrate (*) 1 SD. A. AAer leaming, Day 5. B. One week post-learning, Ret 1 .

Motor Task for Participant PI
A. One Week Post-Learning (Ret 1) B. Two Weeks Post-Learning (Ret 2)
-0.30 1 ONSET
END
Posterior Deltoid
Z / Pectoral is
Major (Pet)
Long Head of Triceps y Lateral Head o f Triceps (LA)
ONSET END
61 91 1 31 61 91
Percent Performance Time
Figure 10. A sample plot of ensemble averaged (5) muscle activity patterns for the motor task o f participant P7. The solid line line represents the mean pattern while the thin vertical lines demonstrate (*) 1 SD. A. One week post-learning, Ret 1. B. Two weeks post-learning, Ret 2.

Motor Task for Participant P7
A. Two Weeks Post-Learning (Ret 2)
-0.30 1 ONSET
1 31
END
B. Four Weeks Post-Learning (Ret 3)
Long Head of Triceps
2 L Biceps
(Bit)
Lateral Head of Triceps
Posterior Deltoid
Pectoralis Major (Pet)
61 91 1 31 61 91
Percent Performance Time
Figure 11. A sample plot of ensemble averaged (5) muscle activity patterns for the motor task of participant P7. The solid line line represents the mean pattern while the thin vertical lines demonstrate (*) 1 SD. A. Two weeks post-learning, Ret 2. B. Four weeks post-learning, Ret 3.

Motor Task for Participant P7
A. Four Weeks Post-Learning (Ret 3)
ONSET
B. Six Weeks Post-Learniog (Ret 4)
Posterior Deltoid
Pectoralis Major
(Pet)
Long Head
(Bic)
1 Lateral Head
I
t-'
ENDI / ONSET END
61 91 1 31 61 91
Percent Performance Time
Figure 12. A sample plot of ensemble averaged (5) muscle activity patterns for the motor task of participant P7. The solid line line represents the mean pattern while the thin vertical lines demonstrate (*) 1 SD. A. Four weeks post-learning, Ret 3. B. Six weeks post-learning, Ret 4.

Motor Task for Participant P7
-4. Six Weeks Post-Learaing (Ret 4) B. Eight Weeks f ost-Learning (Ret 5)
-0.30 1 ONSET
END
Posterior Deltoid (PD)
P - Pectoralir Major
(Pet)
Long Head of Triceps
Biceps (Bic)
Lateral Head of Triceps
/' ONSET END -É&zzzizG\ - (BrR) I I
61 91 1 31 61 91
Percent Performance Time
Figure 13. A sample plot of ensemble averaged (5) muscle activity patterns for the motor task of participant P7. The solid line line represents the mean pattern while the thin vertical lines demonstrate (+) 1 SD. A. Six weeks post-learning, Ret 4. B. Eight weeks post-leaming, Ret 5.

to decrease whereas the Bic increases in amplitude but these signais too demonstrate
limited displacement. As a result, the limited changes expressed by the muscles at Ret 4
would imply a stabilization or persistence of their features with respect to Ret 3. Finally.
at eight weeks post-leaming (Ret 3, the agonists are al1 of lower amplitude, as compared
to Ret 4, with the LA exhibiting a shifi in phase ahead or earlier in tirne with respect to
the onset of movement. Among the antagonists, the Pec displays a srna11 increase in
amplitude while the Bic and BrR are each observed to decrease in intensity. These
muscles did not reveal any shifts in phase.
3.3.2 EMG Patterns Across Muscles Wifhin Euch Learning and Reîention Level-
Singular Value Decomposition Andysis (SVD)
The singular value decomposition analysis employed on the six processed ELMG
signals (normalized to both 100% performance time and peak amplitude) of each of the
learning and retention periods revealed three wavefoms (W's) that may be used to
explain the six original input patterns. The results of such an anaIysis as exemplified by
one participant, P7, are presented in Figures 14-20. As seen in these figures.
superirnposed upon the six original input EMG patterns are the reconstnicted traces using
only these three W's. Minor deviations fiom the original EMG signais and, in various
instances, close to identical plots are concordant with the error of estimation in using
these waveforms to describe the changes in the levels of both the leaming and retention
conditions.
Each of the W's possesses an eigenvalue, in tenns of a percentage score,
s ipiQing the total variability accounted for by that waveform. ï h e eigenvectors or
muscle coefficients in turn reveal the relative significance of those muscles as represented
by the W. Both the eigenvalues and eigenvectors are relative scores, meaning that each
can be considered as a multiple of the other scores in its category making it possible to
refer to one value as being two or three times another. Plotted in parts B, C and D of
Figures 14-20 are W's 1, 2 and 3 for participant P7 of the before and after learning
conditions, along with each of the five retention periods.

Motor Task for Participant P7: Day 1 Thin Lines: Original Data Thick Lines: Reconstructed Data
BrR: 0.37
LA: 0.54
Pec: -0.48 Bit: -0.38
Pec: 0.28 - w3: PD: -0.32
6.04% C
Percent Performance Time
Figure 14. Sample singular value decomposition analysis on the EMG patterns for the motor task of participant P7 before learning @ay 1). A. Original input activation pattems (thin lines) and reconstructed data (thick lines) using al1 three Waveforms (W's). W's I to 3 including their eigenvalues and eigenvectors are presented in parts B to D. B. WI. C. W2. D. W3.

Motor Task for Participant P7: Day 5 Thin Lines: Original Data Thick Lines: Reconstructed Data - PO
0 100 - ---- LO --- 8 0 - LA
W Pec 60 - --- Bic
A. w
cc - BrR
2 40 -
20 oi O
BrR: 0.40 LA: 0.56
PD: 0.69
B R : 0.17
1 12 23 34 45 56 67 78 89 100
Percent Performance Time
Figure 15. Sample singular value decomposition analysis on the EMG patterns for the motor task of participant P7 aRer learning @ay 5). A. Original input activation patterns (thin lines) and reconstmcted data (thick lines) using al1 three Waveforms (W's). W's 1 to 3 including their eigenvalues and eigenvectors are presented in parts B to D. B. W1. C. W2. D. W3.

Motor Task for Participant P7: Ret 1 Thin Lines: Original Data Thick Lines: Reconstructed Data
w1: Pec: 0.32 \ Bk: 0.35 BrR: 0.39
LA: 0.55
0.3 - PD: 0.54 Pec: 0.21 Bic: 0.28
0.0
Percent Performance Time
Figure 16. Sample singular value decomposition analysis on the EMG patterns for the rnotor task of participant P7 at one week post-leaming (Ret 1). A. Original input activation patterns (thin lines) and reconstructed data (thick lines) using dl three Waveforms (W's). W's 1 to 3 including their eigenvalues and eigenvectors are presented inpartsBtoD.B.WLC.W2.D.W3.

Motor Task for Participant P7: Ret 2 Thin Lines: Original Data Thick Lines: Reconstmcted Data - PD
Pet: 0.30
LO: 0.40 Bic: 0.35
LA: 0.57 BrR: 0.41
Pec: -0.51 Bic: -0.44
O 3 r Pec: 0.16
PD: 0.20 Bic: 0.12
0.0 w3: LO: 0.55 BrR: 0.14
Percent Performance Time
Figure 17. Sample singular value decomposition analysis on the EMG patterns for the motor task of participant P7 nt two weeks post-learning (Ret 2). A. Original input activation patterns (thin lines) and reconstructed data (thick lines) using al1 three Waveforms (W's). W's 1 to 3 including their eigenvalues and eigenvectors are prescnted inpartsB toD. B. W1.C. W2. D. W3.

Motor Task for Participant P7: Ret 3 Thin Lines: Original Data Thick Lines: Reconstructed Data
Pec: -0.50 Bic: -0.34
PD: 0.69 Pec: 0.18 Bic: 0.28
1 12 23 34 45 56 67 78 89 100
Percent Performance Time
Figure 18. Sample singular value decomposition analysis on the EMG patterns for the motor task of participant P7 at four weeks post-learning (Ftet 3). A. Original input activation pattems (thin lines) and reconstnicted data (thick lines) using al1 three Waveforms (W's). W's 1 to 3 including their eigenvalues and eigenvectors are presented inpartsB toD. B. W1.C. W2. D. W3.

Motor Task for Participant P7: Ret 4 Thin Lines: Original Data Thick Lines: Recoastmcted Data
Bic: 0.36 BrR: 0.40
LA: 0.57
3 0.2 - .I
e: 2 Pec: -0.47
Bic: -0.39 c* 0.0 E
C 5 26.17% 4 -0.2 -
PD: 0.70 Pec: 0.26 Bk: 0.27
Percent Performance Time
Figure 19. Sarnple singular value decomposition analysis on the EMG patterns for the motor task of participant P7 at six weeks post-leaming (Ret 4). A. Original input activation patterns (thin lines) and reconstmcted data (thick lines) using al1 three Waveforms (W's). W's 1 to 3 including their eigenvalues and eigenvectors are presented in parts B to D. B. W1. C. W2. D. W3.

Motor Task for Participant Pf: Ret 5 Thin Lines: Original Data Thick Lines: Recoashvcted Data
-- Bic - BrR
55.46% r u
LO LA. ,.,,
4 0.38 . n A* Bic: 0.35
BrR: 0.40
PD: 0.59
Bic: 0.21 BrR: -0.24
1 1 1 1 I 1 I I
Percent Performance Time
Figure 20. Sample singular value decomposition anaiysis on the EMG patterns for the motor task of participant P7 at eight weeks post-learning (Ret 5). A. Original input activation patterns (thin lines) and reconstnicted data (thick lines) using al1 three Waveforms (W's). W's 1 to 3 including their eigenvalues and eigenvectors are presented in parts B to D. B. W1. C. W2. D. W3.

The first waveform, WI, for the above participant accounted for 62.99Y0 and
61.75%, of the total variability before and aller leaming respectfblly. At the end of one
week without practice (Ret 1), the variabitity acounted for decreased slightly to 60.48%
and further to 59.00% by Ret 2. At Ret 3 this value was augmented to 60.79% before
reducing to 56.74% at Ret 4 and settling to a lowest percentage of 55.46% by the end of
the retention period (Ret 5), see Figure 2 1 A.
Unlike the slight decrease in the eigenvalue following learning for P7 above, the
total variability accounted for by W1 increased in 8 out of 10 or 80% o f experimental
members. as a group, Erom Days 1 to 5. The opposite appeared to be tme fiom Day 5 to
Ret 1 with 7 out of 10 people exhibiting eigenvalues for W1 that decreased. Across the
levels of the retention condition there was a tendency for this percentage to rise as 40%.
?O%, 60% and 70% of individuals experienced an increase in the eigenvalue fiorn R 1 -W.
R2-R3. R3-R4 and R4-R5 respectfully, (see figure ZA), in comparison the trend of
declining values demonstrated by P7.
W 1, in this analysis, represented a sort of ' m i n g average' of al1 the input
patterns accounting for muscle coefficients that were al1 positive. W1 is an eigenvector
that is also related to amplitude, representing the strength of i ts pattern within each of the
input signals. A higher load for the LA was evident for P7 on Day 1, before learning.
which was dissimilar to the other muscle activation patterns (0.54 vs 0.34, 0.49, 0.35,
0.30 and 0.37 for the LA vs. the PD, LO, Pec, Bic and BrR respectfülly). The same higher
order eigenvector loading for the LA continued to predominate after learning (Day 5 ) and
throughout the entire retention period, (see Figure 21 B). This shows that for P7, the first
common feature strongly represented the pattern for the LA and highiighted the
importance of this muscle within each of the learning and retention levels of the
experimental period.
Six out of ten or 60% of experimental participants displayed the trend for a higher
load of the LA on Day 1. Likewise, 7 out of 10 or 70% demonstrated a higher weightins
for the sarne muscle on Day 5 (see Figure 218). The predominance of the LA above al1
other muscle coefficients, across the five levels of the retention condition was found in
90%, 70%, 60°h, 60% and 60% of participants in Ret 1, Ret 2, Ret 3, Ret 4 and Ret 5
45

A. Eigenvalue fluctuations for W1 - Experimental group
Day 1 Day 5 Ret 1 Ret 3 Ret 3 Ret 4 Ret 5
B. Prominent muscle across participants for W1 - Experimental group
Participant: P 1
PZ P3 P4 P5 P6 P7 Pl3 P9
P l 0
w BrR
LA LA
BrR LA
LO LA LA
LO LA
LA LO LO
LA LA LA LA LOUA
L O LA
Figure 2 1 . (A) Eigenvalue fluctuations and (B) Prominent muscle characteristics for W1 of the experimental group. The bold features highlight participant P7.

respectively (see Figure 21B). Therefore, just as for P7, the uniqueness of the LA pattern,
as revealed by W1, was also a general attribute across experimental participants.
Moreover, for those individuals not displaying a preferential loading for the LA, it was
the LO which predominated arnong the muscle coefficients. The single exception to the
preceding statement was that for Ret 4, the BrR constituted a different muscle, which,
other than the usual appearance of the two triceps muscles in every other experimental
Ievel, dernonstrated its supenority among the agonists and antagonists of two participants.
Regarding the members in the conboi group, the total variability accounted for by
W1 decreased in 3 out of 5 people, increased for one and remained approximateiy the
sarne for another from Day 1 to Day 5 (see Figure 22A). From Day 5 to Ret 5 three
participants displayed an increase in the eigenvalue, as two others showed a reduction in
the latter. In addition, three of five people in the sarne group demonstrated higher muscle
coefficients for the LA on Day 1 while members Cl and C2 each possessed higher LO
loadings for the sarne day (see Figure 22B). On Day 5, C l and C3 displayed greater
coefficients for the LO, whereas C2 and C5 exhibited supenor weightings for the LA.
The PD predorninated arnong muscles in C4 only. At Ret 5 two people possessed higher
muscle coefficients for the LA, two in turn for the LO and one individual displayed a
greater loading for the BrR above the other five muscles (see Figure 22B). The variability
in the eigenvalue fluctuations as well as in the predominance of individual muscle
coefficients within the control group evidently represented nul1 results.
An examination of the eigenvector associated with the second waveform (W2) of
P7. before learning, illuminated an out of phase relationship between the PD (0.62. high
positive loading) and LO (0.32, moderate positive loading) versus the Pec, Bic and BrR
(-0.38, -0.38 and -0.36, moderately high to moderate negative loadings respectively), see
figure 14C. That is, W2 represented the timing of the power of activation between the
agonist and antagonist muscle groups. It was evident that this out of phase relationship
was getting stronger with leaming as the eigenvector coefficients for Day 5 are obsewed
closely (see Figure 15C). Before leaming, the structure of the eigenvector was 0.62 and
0.32 versus -0.48, -0.38 and -0.36 for PD and LO vs. Pec, Bic and BrR respectively.
accounting for 24.04% of the total variability. AAer leaming, the eigenvector coefficients
47

A. Eigenvalue fluctuations for W1 - Coatrol group
Day 1 Ret 5
B. Prominent muscle across participants for W1- Control group
Figure 22. (A) Eigenvalue fluctuations and (B) Prominent musde charactenstics for W1 of the control group.
LhLl IhL5 IkCi Participant:
Cl C2 C3 C4 CS
LO LO LA LA LA
LO LA LO PD LA
BrR LO LO LA LA

were 0.55 and 0.40 versus -0.48, -0.42 and -0.36 for the same muscles as above
accounting for higher variability, 28.3 1%. An increase in the intensity of the out-of-phase
relationship for P7 revealed a greater antagonistic inhibition with learning that was
di fferent from a more prevalent CO-contraction before learning.
At one week beyond practice, the power of the relationship appeared to have
plateaued as revealed by the eigenvector for Ret 1 of 0.55 and 0.37 for PD and LO in
sequence versus the Pec (-0.48), Bic (-0.41) and BrR (-0.38) accounting for 29.3S0/0 af the
total variability. At two weeks post-learning the muscle coefficients dernonstrated a small
increase in the intensity of the out of phase relationship at 0.5 1 and 0.39 vs. -0.5 1. -0.34
and -0.34 concomitant with a diminution in the eigenvalue to 28.12% From 29.38% at
Ret 1. From Ret 2 to Ret 4 the eigenvector loadings did not offer a concrete case for
either the progression or regression of the out of phase pattern. The intensity of the
pattern was more or less maintained, supported by the stabilization of the eigenvalues
frorn 28.12% to 26.37% and 26.17% for the three retention intervals respectively (see
Figures 16C - 19C).
Finally, at eight weeks post-learning the eigenvector was characterized by the
values of 0.50 and 0.25 for the agonist muscles of the PD and LO versus -0.50, -0.44 and
-0.39 for the antagonists of the Pec, Bic and BrR respecthlly (see Figure ZOC). When
compared to the muscle coefficients for Ret 4, an increase in the pattern between the two
muscle groups was evident in spite of an eigenvalue that had not changed measurably
frorn Ret 3. In summary, the eigenvector coef'fïcients, reIating the intensity of the
aforementioned out of phase pattern between muscle groups for P7, clearly divulged an
enhanced antagonistic inhibition with learning that continued to manifest itself at one
week post-learning. However, other than a small increase in the strength of the
relationship at Ret 2, the same muscle coefficients did not reveal any categorical changes
to the relationship with retention. For a report on the eigenvalue fluctuations for W2
arnong members of the experimental group see figure 23A.
Another method of quantifjwg the out of phase relationship between the agonist
and antagonist muscle groups was the evaluation of the delay between the peak and the
valIey of the eigenvector for W2. If a line is vertically drawn fiom both the highest point
49

A. Eigenvalue fluctuations for W Z - Experimental group
7 r - r r -
Day 1 Day 5 Ret 1 Ret 2 Ret 3 Ret 4 Ret 5
B. Composite score fluctuations for W2 - Experimental group
-
Day 1 Day 5 Ret 1 Ret 2 Ret 3 Ret 4 Ret 5
Figure 23. (A) Eigenvaiue fluctuations and (B) Intensity of the 'Out of Phase' relationship as revealed by W2 for the experimental group. The solid line represents the data for participant P7 while the faint lines descnbe the data for the other members.

(peak) and the lowest point (valley) of the eigenvector to the abscissa or x-mis, which
represents percent performance time, a percentage score is obtained. Subtracting one from
the other provides a composite score that can be used to demonstrate either an increase or
a decrease in the aforementioned relationship. For example, it can be determined that for
Day 1 . the composite score is 28% (76%, the percentage value associated with the lowest
point of the eigenvector, minus 48%, or the value paired to the highest point of the sarne
eigenvector), For Day 5 it is 35%, which therefore represents an increase in the intensity
of the out of phase relationship with learning. For P7 the post-learning level was
maintained at Ret 1 but showed a decrease in the composite score and hence a diminution
in the power of the relationship at Ret 2. Thereafier, the values were not observed to have
changed considerably, see Figure 23 B.
Looking at the entire pool of expenmental participants, the group was found to
display a significant increase in the strength of the out of phase relationship with learning,
F (1.9) = 35.1, p<0.001, and thus an increase in reciprocal inhibition. Furthemore,
fluctuations in the level of intensity of the above pattern during the retention period were
only statistically significant fiom Ret I to Ret 2, F (1,9) = 10.8, pc0.01. At two weeks
post-leaming then, there was a decrease in the values of the composite scores that
promoted a diminution in the level of reciprocal inhibition and an increase in the
antagonistic CO-contraction between the two muscle groups, see figure 23B. There were
no other significant differences in the composite scores of this group beyond Ret 2 as
dernonstrated by the pair-wise cornparisons of Ret's 2-3, 3-4, and 4-5, see Appendix 8.
Changes in the eigenvalues for W2 within the control group can be seen in figure
24A. The percentages did not reveal any significant trends. Altematively, a two-way
repeated measures ANOVA that was conducted on the composite score data that was
pooled over both the experimental and control groups revealed significant adaptations in
the intensity of the 'out of phase' relationship across each of the post-training and post-
retention conditions, F (2,261 = 9.4, p<0.001. The pooled data also demonstrated
significant interactions between the two groups, F (2,26) = 4.3, pc0.025. The tests of
within subjects contrasts exhibited significant increases in the power of the relationship
for both groups fiom pre- to post-learning trials, F (1,13) = 12.2, p<0.004. The tests also
5 1

A. Eigenvalue fluctuations for W2 - Control group
B. Composite score fluctuations for W2 - Experimental and
15
Control groups
-
Day 1
12 I
Day 1 Day 5 Ret 5
Day 5 Ret 5
Figure 24. (A) Eigenvalue fluctuations for the control group and (B) Intensity of the 'Out of Phase' relationship for both expenmental (Exp) and control (Con) groups as revealed by W2.

divulged a significant decrease and increase in the intensity of the pattern for the
experimental and control groups respectively, from post-learning to post-retention
periods, F (1,13) = 5.5, pc0.036. The rates of the increases seen in the out of phase
relationships fiom Day 1 to Day 5 for each of the groups were not statistically different, F
(1,13) = 0.2, pc0.634. However, the rates of the changes describing a reduction in the
intensity of the pattern for the experimental group by the end of the retention protocol. in
contrast to the continuing increase in the power of the relationship for the control group.
was found to be significant, F (1,13) = 10, p<0.007 (see Figure 24B and Appendix 9).
The third waveform, W3, for P7 accounted for 6.04% and 4.39% of the variability
before and after learning in sequence. Thereafier the eigenvalue rose to 5.96% at Ret 1
and increased fiirther to 8.30% at two weeks post-learning before falling to 6.98% four
weeks afier training. At Ret 4 there was an increase in the variabiiity accounted for to
8.74% and 9.49% by Ret 5.
W3 explained the remaining variability of the three wavefoms used to interpret
the EMG data. In other words, after the variability for both W 1 (a running average of the
muscle activation patterns with a stronger representation of the LA) and W2 (out-of-
phase relationship between agonist and antagonist groups) is removed, what remains is
the variability associated with W3. Even though the eigenvalues for W3 are low, as seen
above for P7, the fact that there exist opposing polarities of the eigenvector coefficients
within each of the agonist and antagonist groups made it evident that this waveform
represen ted the complex contrasts among the individual muscles within each group.
An observation of the eigenvector of P7 for Day 1, before learning, divulged the
LA (0.56) with a high positive loading in contrast to the PD (-0.32) which had a
moderately low negative weighting coefficient among the agonists; the LO (-0.07) was
essentially, not represented. The Bic (-0.69) with a very high negative coefficient differed
in polarity from the Pec (0.28), whose low positive weighting, along with the BrR's
(-0.14) very low weighting, made each of these muscles within the antagonist group
negligible.
An examination of the shape of the waveform for W3, on Day 1, dispiayed a more
local pattern of the motor task, whose amplitude was concentrated within the latter half of
53

the movement (Le. between 50 and 100 points of percent performance time). The local
pattern seemed to portray the high positive loading of the LA and more importantly, the
very high negative loading of the Bic (-0.69), see figure 14D. Consequently, before
learning, P7 demonstrated an antagonist representation of the motor task with a relatively
weaker contribution of the agonist muscles.
After learning, on Day 5, each of the PD (0.69) and LA (-0.61) had increased
remarkably in magnitude and maintained their contrasting polarity, attesting to the strong
latency between them. The LO (-0.22), although demonstrating an increase in its
coefficient as compared to Day 1, was still at a relatively low and negtigible weighting. In
contrast, there was little, if any, presence of the forearm flexors (0.24, 0.17 and 0.13 for
the Bic, BrR and Pec respectfully). The waveform on Day 5 was evidently sinusoidal and
eshibited a more global pattern of the task that was distributed over the entire
performance time. At this point the amplitude of the waveform appeared to be
concentrated within the initial half of the task (Le. between 1 and 56 points of percent
performance time). As a result, following learning, P7 demonstrated an essentially
agonist representation of the motor task owing to the negligible presence of the
antagonists (see figure 15D). An examination of the agonist pattern revealed a proximal
to distal relationship defined by the very high positive loading of the PD versus the high
negative loading of the LA. This interpretation of the latter muscles on Day 5, wouid
seem to quali@, in retrospect, the agonist representation on Day 1 as a weaker or
underdeveloped proximal to distal relationship since the LA was shown to be the
predominant muscle in contrast to the PD before practice.
With one week of no-practice the PD maintained its contrasting polarity with the
LA but was lower in magnitude at 0.54, with respect to the latter muscle which had
become much more prevalent at -0.76. The small gain realized by the LO afier learning,
had effectively been compromised at Ret 1 (LO: 0.1 1). Among the antagonists, there was
no representation for the BrR (0.01), as the Bic and Pec possess low weighting
coefficients (0.28 and 0.21 respectively). The global, sinusoidal shape of the waveform
was maintained at Ret 1 that continued to depict an agonist characterization of the
'strategy' employed by P7 to produce the motor task. However, the lower eigenvector
54

loading of the PD in contrast to the higher weighting of the LA revealed what appeared to
be a decrement in the strength of the proximal to distal pattern-
At two-weeks post-leaniing the agonist portrayal changed with the LO (0.55)
corning into prominence at a high positive loading in opposition to the LA at -0.78, a very
high negative coefficient. The PD (0.20) was conspicuously reduced in its presence
among the extensor muscles at this time which was readily observed by a leveling out of
the eigenvector for Ret 2 (see figure 17D). Consequently, the LO muscle seerned to take
on the role of the PD in describing the proximal to distal timing of the activations
between the two triceps muscles. All of the antagonist muscle coefficients remained
insignificant here.
At four weeks beyond the completion of training (Ret 3) the flexor group was still
conspicuously absent. Within the extensor group however, the PD had increased
substantially in its loading and was positive at 0.69, in relation to the LA and LO each at
a high (-0.56) and moderate (-0.32) negative weighting, respectively. It appeared that at
this time the proximal to distal pattern had become re-energized as evidenced by a greater
latency between the PD and the combined activation of the LA and LO muscles which
demonstrated an increase in synergistic activation. The aforementioned pattern also
appeared to persist through to Ret 4, except for the LO, whose eigenvector coefficient
decreased to the point of being negligible (see figures 18-1 9D).
At the end of the retention period, the PD decreased in weight to 0.59 as the LO
displayed itself at a very high negative loading of -0.69. There was no representation for
the LA however, which had consistently demonstrated a strong latency with respect to the
proximal joint agonist in each of the preceding experimental sessions. It would appear
that the LO muscle supplanted the LA in characterizing the proximal to distal pattem
upith the PD. Each of the Pec (0.27), BrR (-0.24) and Bic (0.21) remained, as throughout
the retention penod, at low weighting coefficients.
In sumrnary, for this participant, W3 remained global and unchanged for the
majority of the retention penod; except for Ret 2, which demonstrated a virtual absence
of the PD. Moreover it was dominated particularly by the extensor group and depicted the
contrast of the firing pattem between the PD versus the LA and LO. According to the
55

eigenvector coefficients, this pattern deteriorateci at two-weeks pst-training after which it
appeared to make a corne back at four and six weeks of no-practice. It seemed to dirninish
in strength again by Ret 5 (eight weeks post-leaming). The eigenvectors of al1 three
waveforms (Le. W1, W2 and W3) for participant P7, within each of the learning and
retention conditions, are presented in Table 2.
A complete depiction of the within muscle group contrasts of al1 experirnental
members, is provided in Appendices 10 - 13. Appendix 10A shows the agonist ancüor
antagonist representations of the rnotor task fiom Day 1 to Day 5, before and after
learning respectively. The actual data (Le. the eigenvector coefficients) o f t he
a forernent ioned muscles depicted in the preceding Appendix are avai lable in Appendix
10B. Appendices 1 1 - 13 include the within muscle group characterizations of the rnotor
task, as well as the eigenvector coefficients of the muscles for both the short- (Day 5 to
Ret 2) and long-term (Ret 3 to Ret 5) retention intervals. Within each of the these figures
the muscles are presented in order of decreasing magnitude according to their respective
coefficients. This was done in order to highlight those muscles contributing most to the
production of the motor task. They do not necessarily imply an order of muscle firing
unless otherwise stated. instead they cornrnunicate the relative importance or significance
of each muscle within the agonist and/or antagonist pattern of the particular learning or
retention condition (see page 1 26).
Across experimental participants, learning also resulted in significant phasic
adaptations to the sequencing of the muscle activity patterns within the agonist muscle
group in particular. It revealed a general increase in the pattern of the proximal t o distal
activation of these muscles, in eight out of ten people (80%: P2-P9), which contributed to
an improvement in motor performance. At the sarne time, the level of antagonist
involvement was observed to decrease for the majority of experimental members (70%:
Pl-P2, P4, and P6-P9). The latter was demonstrated by a reduction in either the
magnitude or the nurnber of eigenvector coefficients for the muscles highlighted as
significantly contributing to the motor task on Day 1. The muscles which remained
representative within the task after learning consisted of either the Bic or BrR.
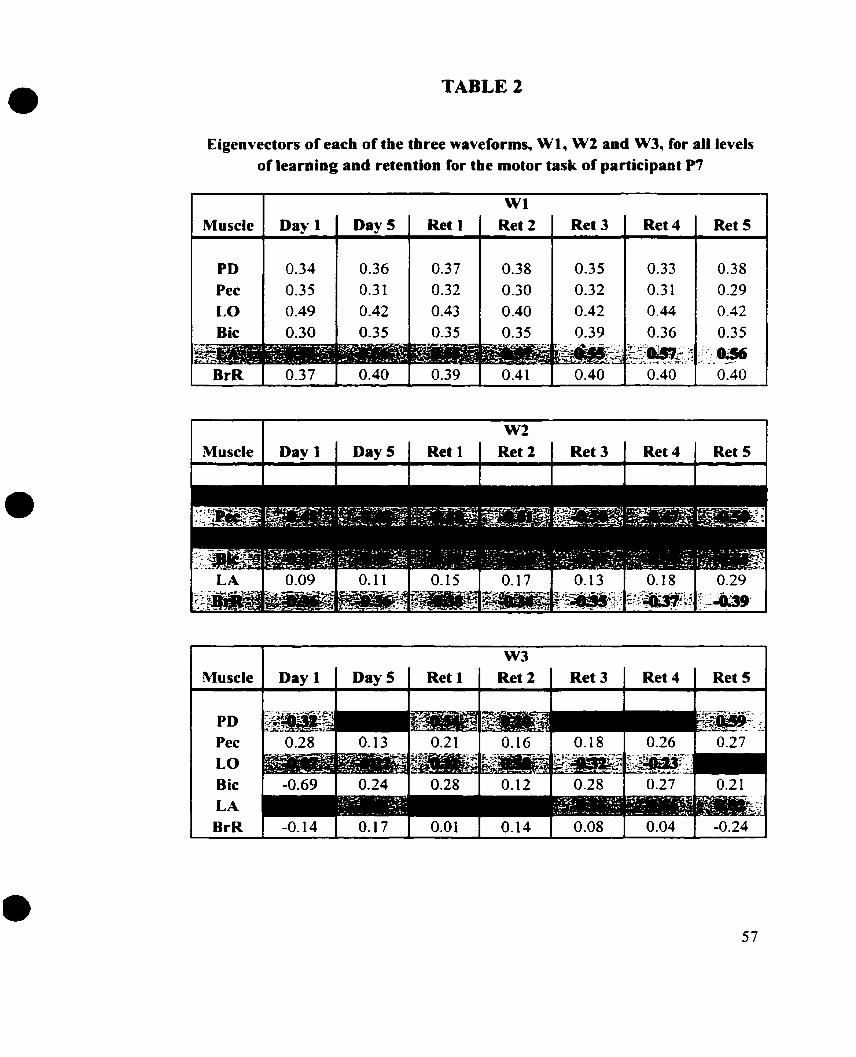
TABLE 2
Eigenvectors of each of the three waveforms, W1, W2 and W3, for al1 levels of learning and retention for the motor task of participant P7
1 Muscle 1 Day 1
PD 0.34 Pec 0.35 L O 0.49 Bic 0.30
1 BrR 1 0.37
Day 5 Retl 1 Retz 1 Ret3 1 Ret4 1 Rets
Muscle
1 Pec 1 0 . 2 8 1 0 . 1 3 1 0 . 2 1 1 0 . 1 6 1 0 . 1 8 1 0 . 2 6 1 0 . 2 7 1
Muscle l
w2 Day t 1 Day 5
I
Day 1
Ret4 1 Ret 5
1 1
Ret 1
Day 5 Ret4
Ret 2
Ret 1 Rets
Ret 3
Ret 2 Ret 3

The above mentioned recession of antagonist activation within the motor task
reinforced the significance of the agonist representation and suggested the apparent
'strategy' of the neuro-rnotor system: to program the initial propulsive characteristics of
the motor task so as to engage only the appropriate or sufficient arnount and/or timing of
antagonist activity. Furthemore, the degree to which the magnitude of the relationship
between the PD versus the LO and LA is expressed, is not the same for everyone but
specific to the individual performing the task. In other words people will leam to petfom
the motor task with varying extents of muscle activations owing to simple individual
di fferences.
Certain exceptions among the members of the experimentai group were
noteworthy, however, such as for P8, who was singular in his portrayal of a global pattem
of the third waveform (W3) before practice. The pattern highlighted a rather distinguished
proximal to distal relationship of the PD (-0.66) versus the LA (0.41) and LO (0.38) on
Day 1, apparently superseding the involvement of the Bic (-0.42). Members P4 and P l0
also displayed a similarly conspicuous proximal to distal agonist pattern on Day 1, before
practice, but whose wavefonns were characteristic of the local pattern described
previousIy (see Appendix 10A and B). With learning, the agonist representation did not
change for P4 and P8, instead the power of the relationship between the muscles becarne
stronger as evidenced by the eigenvector loadings for both participants (P4: 0.61 vs. -0.5 1
on Day 1 and 0.65 vs. 4-71 on Day 5 for the PD versus the LA respectively and f8:
-0.66 vs. 0.41 and 0.38 on Day 1 with respect to 0.61 vs. -0.47 and -0.43 for the PD
versus the LA and LO respectively). In addition, P l0 was the only person to convey a
decrease in the intensity of the agonist representation with learning in favor of a Full
complement of antagonist muscles involved in the task, see Appendix 10A and B.
Other interesting findings included the antagonist representations of participants
P6 and P9 before training, which exposed a dissociation of the Pec (a proximal joint
horizontal adductor) versus the Bic and/or BrR (muscles primarily associated with elbow
flexion). The differentiation of these muscles on Day 1 would suggest a proximal to distal
activation concerned with arresting the initial propulsion characteristics of the brachium
by the PD and the resulting extension of the fore- to the target as effected by the LA
58

andior LO muscles. It was observed that following training, these same individuals
simpli fied the manner in which they stopped the whipping movement by mostly enlisting
the action of the Bic, a two-joint muscle, which may have acted at both points of the two-
joint system.
Across the short-tenn retention intervals, the data of the third waveform for the
experimental group provided results that corroborated the findings of the second
waveform which, as previously noted, described a persistence of the adaptations between
muscle groups at Ret 1, and a decrement in the latter at two weeks post-learning. The
third waveform revealed a majority (i.e. 60%) of experimental members (P3. P5. and
P7-P IO) who also exhibited a persistence or even an improvement in the within muscle
group adaptations fiom Day 5 to one-week post-learning. These included a maintenance
or enhancement of the power of the proximal to distal relationship and a continuing
abatement in the significance of the antagonists during the motor task (Le. P3, P5, P7-
P 10. see Appendix 1 1A and B). Some exceptions to the preceding declarations included
P9 who showed an irnprovement in performance based on a substantial decrease in
performance time, from 255.4 msec on Day 5 to 208.8 msec at Ret 1. Underlying the
decrease in performance time was an agonist representation of the LA (-0.55) versus the
LO (0.50) muscle that introduced the latter as a significant proximal joint agonist in the
motor task. Participant P l 0 was also exceptional within the experimental group at Ret 1
in that he demonstrated a fortified antagonist sequence of the BrR (-0.77) versus the Bic
(0.54) and Pec (0.31) cornmensurate with a total dissolution of the agonist muscle
presence (see Appendix 1 1 A and B).
The four rernaining individuals, Pl-P2, P4 and P6, exhibited decrements in motor
performance at one-week post-learning that were characterized by either a weakening of
the proximal to distal pattern of the agonists and/or a resurgence of a significant
antagonist presence in the motor task. It is important to note, however, that the
diminut ion in coordination expressed by three of the aforementioned members did not
necessarily demonstrate regression to pre-training levels. The within muscle group
representations of these people (N=3) still promoted certain adaptations acquired with
learning. Participant Pl for instance, showed that the LA muscle, which solely
59

distinguished the agonist representation on Day 5, continued to present itself as a
preeminent muscle within the agonist group at Ret 1. P2, who exhibited a reduction in the
degree of the proximal to distal pattern of the agonists at Ret 1, nonetheless continued to
show an absence in the antagonist presence that was specified with learning on Day 5 . in
addition. P6 retained a degree of the proximal to distal character of the agonists. after
training, in the form of the LO versus the LA, in spite of a reappearance of the antagonist
muscles which were observed to characterize the motor task on Day 1. In fact, P4 was the
only person to display a complete reversa1 of adaptations at the end of one-week without
training, see Appendices 10 - 11, A and B.
At two-weeks post-learning, however, 60% of individuals (Pz. PS, and P7-P10)
demonstrated decrements in the within muscle group characterizations which, like the
results in the second waveform above, provided fùrther evidence of a limit to which
adaptations in motor ski11 acquisition remained unaffected by a termination in training.
For P2, P5 and P7-P9 the decline in performance entailed a decrement in the status of the
proximal to distal nature of the agonists and a reaffirmation of the antagonist presence in
the motor task with respect to the within muscle group characterizations of the previous
retention interval (see Appendix 1 1A and B). Participant P l0 on the other hand, reverted
to a greater representation of the agonist muscles in a proximal to distal sequence of
activation, which was similar to that exhibited on Day 1.
The data of the long-term retention penods, however. did not necessarïly reveal
that the diminution in neuromuscular coordination observed at Ret 2, marked the
beginning of a progressive decrease in performance over Ret's 3 - 5. Rather, in each of
Ret7s 3, 4, and 5 there were 70% of experimental participants who exhibited either
persistent or slightly augmented levels of motor performance in comparison to the degree
of coordination displayed at Ret 2 (see Appendices I l - 13, A and B). The improvements
in movement production were usually accompanied by decreases in performance time but
these values, which were sometimes better than the counterpart times at Ret 2, were never
lower than the PT'S demonstrated at Ret 1. In other words, the instances of an
enhancement in performance within experimental members across the long-tenn retention
penods were indicative of individual residual improvements in motor coordination that
60

TABLE 3
Eigenvalues, percent variability accounted for (* 1 SD) by eacb waveform and the sum of al1 three W's within both learning and retention levels
for the motor task of the experimental group (N=10)
Da' 1
Day 5
Ret 1
Ret 2
Ret 3
Ret 4
Ret 5
SUM W1-3

were restricted to these intervals. They did not represent superior performances in relation
to those observed at Ret 1 , the last indication of a persistence or improvement in the
neuromuscular adaptations acquired with leming.
Table 3 displays the average variability terms for each of the wavefonns of the
SVD analysis (Wl, W2 and W3) as welt as the surn of a11 three, for the entire
experimental group, within each Iearning and retention level. Although the changes
observed for the individual waveforms were not conspicuous, the sum of al1 the
wavefoms put together demonstrated an increase in the variability accounted for with
leaming and a decrease in the variability accounted for at the end of the retention period.
These data confirmed the tower variabiiity in the EMG waveforms as a result of the
training regimen and an increase in the variability of the muscle activation patterns after
eight weeks of detraining.
Appendix 14A presents the within muscle goup characterizations of the control
group fiom Day 1 to Day 5 . The actual data for the muscles depicted in the figure is
available in Appendix 14B. Four out of five individuals clearly dernonstrated a local
pattern of the waveform for W3 on Day 1, while a fiflh person was singular in the
exhibition of a global pattern of the waveform for the sarne day. Similar to the findings of
the expenmental group above, before practice, the local pattem of the four control group
members (N= 4) portrayed a significant presence of the antagonists in deference to a
weaker contribution of the agonist muscles. The magnitudes of the antagonist coefficients
ranged fiom -0.31 (C4: BrR) to 0.85 (Cl: Pec) see Appendix 14B. Among the five
rnembers of this group, C 1 , C4 and CS show the presence of one antagonist muscle, as C2
and C3 demonstrate two and three antagonists respectively.
On Day 5, four out of five rnembers displayed a global pattem of the wavefonn
for CF3. Only one person, C l , continued to show a local pattem characterizing the motor
task whose within muscle group representation was identical to Day 1. Therefore,
equivalent to the changes expenenced by the majority of the expenmental group, the
aforementioned subgroup (N= 4) of control participants conveyed an increase in the
power of the proximal to distal relationship cornmensurate with a diminution in the roles
of the antagonist muscles.

At Ret 5, al1 of the control group members portrayed a global shape of the
waveform for W3, which communicated an ongoing manifestation of the intensity
conceming the proximal to distal pattern of the agonists developed by Day 5. A
reconstitution of the antagonist presence was also apparent however, indicating a retum
to the significant involvement of these muscles in the motor task, as was previously noted
on Day 1, and hence a small but notable regression in petformance. The fact that the
control group displayed a persistence of some of the acquired adaptations expressed on
Day 5 further attested to the learning effect in this group in spite of a minimal amount of
exposure to the motor task (see Appendix 11B).
The averaged variability tenns for each of the three waveforms and the surn of al1
three together for the five members of the control group are presented in Table 4. The
data showed an increase in the variability accounted for fiom Day 5 to Ret 5. which
demonstrated a higher contribution of the second waveforrn (W2).
TABLE 4
Eigenvalues, percent variability accounted for (* 1 SD) by each waveform and the sum of al1 three W's across post-training and post-retention
periods of the control group (N=5)
L
Day 1 W l
62.2 1 (12.63)
W2 22.78 (13.79)
W3 8.37
(* 1.76)
SUM W1-3 93 -3 5 (* 1 -23)

CHAPTER IV
The current study empioyed electromyographical techniques to assess the neural
activations of six muscles of the left upper limb during both leaming and retention
protocols. These EMG signals constituted mesures of performance production that were
analyzed by a method of singular value decomposition. SVD analysis permits the
examination of entire wavefoms as opposed to discrete evaluations conceming the time
series of the data, which observe only portions of the actual muscle activity patterns, (e-g.
onsets and offsets, threshold levels etc.). Consequently, it is a useful method in describing
the phasic information of the muscle activity patterns being anaiyzed. Moreover, the
identification of a minimum number of common patterns or wavefoms underiying the
EMG data is consistent with the reduction of degrees of fieedom theory in motor control
as proposed by Bernstein (1967). That is, similar to Bernstein's theory, the SVD analysis
seeks to generate a minimum number of control pattems whose changes across both
learning and retention levels of the current experiment c m elucidate the modifications in
the neural input to the muscles being analyzed.
Of the six original input patterns to the SVD process, three wavefonns were
extracted fiom the data to describe the neuromuscular adaptations to motor ski11 learning
and retention. As alluded to above, the aim was to illuminate the CNS strategies
goveming the reduction of degrees of freedom with learning as theorized by Bernstein
( 1 967) and more importantly, to determine how these strategies change with increasing
penods of no-practice.
The results of this investigation are discussed within sections delineating the
learning adaptations of the experimental group as a fùnction of the practice protocol first.
This will serve as a preface to the subsequent evaluation of the effects of both short- and
long-term penods of no-practice respectively, on the coordination of the muscles
goveming the task. The changes associated with the control group will be dealt with
separately, at the end of the chapter.

The expenmental participants in the current study achieved the principle criterion
of leaming, namely a significant decrease in performance time. Even though the motor
task consisted of a relatively simple whipping movement, the volunteers were able to
improve their execution of an as fast as possible action to an extemal target over
successive days of the learning paradigm. It is important to note that although the
mechanical aspects of the motion are easily reproducible, the requirement of perfonning
to the limits of one's maximum efforts entails a vigorous challenge for the neuromuscular
system. Consequently, investigations into apparently simple movements are valuable
when inquiring into the underlying processes and products of movement (Gottlieb et al.,
1988; Corcos et al., 1993; Vardaxis, 1996).
When studying multi-joint movement in particular there is always a question
regarding the representation by the sample of muscles selected for analysis. In this
investigation six muscles were chosen according to each of their primary and/or
secondary îùnction as an agonist (PD, LO and LA) or antagonist (Pec, Bic and BrR) to
the motor task. A simple way of looking at this sarnple is to consider pairs of reciprocal
muscles in the same anatomical location that possess antagonistic roles. For instance. the
PD and the Pec can both be observed as the proximal, single-joint muscles involved in
horizontal abductiodadduction. The former contracts concentrically first. to decrease the
posterior angle of the shoulder while the latter follows with an eccentnc contraction, to
control the increasing anterior angle of the same shoulder. Likewise the LO and Bic are
both two-joint muscles that cross each of the shoulder and elbow articulations. In order,
they fiinction as a secondary agonist and antagonist to the proximal joint respectively. as
well as to the distal joint, the elbow. Finally, each of the LA and BrR share in their
unique actions at the distal joint, as a primary elbow extensor and flexor respectively. The
above muscles were chosen as being representative of the motor program used to produce
the task. There are other muscles of the upper limb, shoulder and postenor back regions,
which contribute to the movement, however these muscles are located deep within the
lirnb and shoulder complex (e.g. media1 head of the triceps, brachialis, coracobrachialis
and pronater teres) which would preclude the use of surface electrodes.

4.1 Adaptations of the agonist and antagonist muscle groups with learning
The leaming protocol employed in this study was successful in determining
changes conceming the neural activations of the muscles composing each of the agonist
and antagonist groups. The SVD analysis appropnately revealed those changes, as
observed by the three wavefoms used to explain the muscle activation patterns
contributing to the production of the motor task. It is meaninghl to discuss the 'number'
of wavefoms involved since they are orthogonal and independent one from the other.
sharing no covariance between them. The first waveform (Wl) had, associated with it.
eigenvector coeficients that were al1 positive across both pre- and post-learning
conditions. This communicated the fact that W1 constituted a 'nuuiing average' of the
original EMG pattems, representing the strength of the W 1 waveform within each of the
original input pattems. The shape of the waveform before leaming appeared to possess
two 'humps', of which the second was larger than the fint. They seemed to reflect the
temporal locations of the initial agonist EMG bursts followed by those of the antagonist
muscles (see Figure 14A and B). At the end of training, the waveform had a more
'rounded-out' look (see Figure 15B). It clearly attested to EMG bursts that had become
more phasic, with earlier and faster rise times in relation to the onset of movement
(particularly among the extensor muscles), whose areas were considerably larger than
before training (see Figures 8 and 15B). Although there was a general increase in the
eigen~ralues for W 1 across experimental participants, From pre- to post-learning settings,
the magnitude of the changes with leaming were not pronounced (see Figure 2 1 A, Day 1
to Day 5). The inconspicuous nature of these results was interesting since qualitative
observations of each of the six muscles for an example participant (P7) categoncally
attested to the effects of training on the muscle activation pattems, as referred to above
(see Figure 8).
Among the muscles represented by W l , the LA was one that demonstrated a
higher eigenvector coefficient above al1 other muscle coefficients from pre- to post-
learning trials for the majority of experimental participants (see Figure 21B). The
prominence of this muscle probably indicated its importance in extending the forearm to
the target of the motor task following the initial propulsion of the Srachium by the PD

and/or the LO. This could be explained by the fact that the angular displacement required
at the elbow (90 degrees of extension from the initial starting position) to reach the target
was greater than that of the shoulder (45 degrees of horizontal abduction from the initial
holding position). These results concur with the findings of Wadman et al., (1980) who
demonstrated that for the muscles having a real contribution to the movement under
investigation, the duration of the agonist excitation was longer for a larger movement.
Thus the LA, as the single-joint elbow extensor, wouId be central to the task.
Furthemore, it is possible that a stretch reflex was elicited from the LA muscle, during
the initial acceleration of the brachium by the proximal joint agonists, that could have
placed the former muscle on stretch. It is well known that a muscle which is piaced on
stretch, to a certain degree accorJing to the length tension relationship of the specific
muscle, responds with a more forcefil contraction (Kreighbaurn & Barthels, 1985).
It is interesting to note that the LO exhibited the second highest eigenvector
coefficient arnong the muscles studied from Day 1 to Day 5 and was often quite close to
the magnitude of the value for the LA. This may reflect the LO's dual nature in the motor
task as horizontal shoulder abductor and elbow extensor, such that it could have been
assisting in each of the aforementioned actions during the performance. Once again.
however, in view of the greater amplitude of movement required at the elbow in the task,
i t would be plausible to assume that, like the LA, the LO was being recruited as an elbow
extensor rather than as a shoulder abductor. As a result, it couId have also experienced a
stretch reflex during the motor task that would help explain its comparable muscle
coefficients, with respect to the values of the LA. Any comments related to the actual
strength of the above muscles, as a contributing factor to their superior manifestation
within the eigenvector, would be speculative since no pre- or post-learning strength
measures were taken.
Although the amplitude of a muscle's EMG activity can be equated with the
muscle's force characteristics (Woods & Bigland-Ritchie, 1983), there are a number of
factors associated with surface EMG recording that should be taken into account when
interpreting such a parameter. These include the variability in the fiber-type composition
of different muscles, the low-pass filtering effects of larger skinfolds as well as the inter-
electrode distance of bi-polar configurations (Bilodeau et al., 1990). In this investigation.

the EMG patterns were normalized to peak amplitude before the SVD analysis, a process
that would preclude an evaluation of the amplitude characteristics of the signals and
hence a measure of the force characteristics of the muscles. As a result, only the phasic
effects of the muscle activation pattems assayed during both the learning and retention
protocols were capable of being analyzed. An increase in the strength of the muscles
following the practice penod is possible however, considering that strength
training/detraining paradigms have alluded to the prospect of increasing neural
adaptations prior to actual muscle fiber hypertrophy (Hakkinen & Komi, 1983; Narici et
al.. 1989).
Moreover, researchers have found a preferential hypertrophy and an increase in
the integrated EMG activity of individual muscles within the same muscle group
following a strength training regimen that alluded to a different fraction of the total force
of the muscle group being exerted by these muscles (Narici et al., 1989). This knowledge
emphasizes the importance of ascertaining the strength levels of the muscles being
investigated in movement studies, throughout the experimental period, in order to clearly
assess their precise roles in producing the motor task in question (van Bolhuis & Gielen.
1997).
The appearance of the second waveform, (W2), was sinusoidal with an initial
positive peak located in the fint half of the task followed by a negative peak present in
the second half of the motor task. The eigenvector coefficients for W2 were divided into
positive scores for each of the PD, LO and LA muscles within the agonist group and
negative scores for the Pec, Bic and BrR muscles of the antagonist group. Both the shape
of the waveform and the division of the coefficients along sign lines underscored a
synergistic activation of the agonist and antagonist muscles with an activation lag
between the two groups. Specifically, this cornmon feature highlighted the out-of-phase
relationship regarding the PD and LO versus the Pec, Bic and BrR which exhibited a
higher CO-contraction between the two muscle groups before learning (see Fisure 14C).
-4cross experimental participants the eigenvector coefficient for the LA muscle
possessed a minimal weighting which revçaled that it did not share in the synergy
expressed by the other two agonist muscles. The proximal origins of the PD and LO
would apparently explain the increased covariation between these muscles and dictate a

greater latency of their EMG bursts with respect to the patterns and, in particular, the nse
times of the antagonists. It is not that the LA burst or its rise time was not distinctively
separated from each of the shoulder and f o r e m flexors, but rather to a slightly lesser
degree as compared to the other two agonist muscles. In fact for one particular individual
(participant P l ) the rise time and presence of the LA burst was actually in the same
temporal location (Le. in the second half of the motor task) as each o f the antagonist
bursts. on Day 1. This was substantiated by an eigenvector loading for the elbow extensor
that was negative and approxirnately of the sarne magnitude as the other shoulder and
elbow flexor muscles. Furthemore, it is interesting to note that the LO was not
represented equally with the PD in terms of its eigenvector loading and. for al1
experimental participants, was always o f a lower weighting with respect to the latter
muscle. Once again, this is probably due to its position within the proximal to distal
hierarchy of the agonists; that is, within the motor task it was the secondary shoulder
extensor after the PD.
With leaming, the phasic effect, as revealed by the shape of the wavefonn,
became very clear among the experimental participants, demonstrating a decrease in
antagonistic co-contraction in light of an increase in reciprocal inhibition between the
agonist and antagonist muscle groups (see Figure 15C). This was also observed by a
general increase in the eigenvalues and eigenvectors of the SVD analysis for most
members. Some participants did not show significant gains in either o f the two SVD
parameters but inspection of the original data wavefoms and the eigenvectors for CF2
unequivocally revealed the changes brought about by motor task practice. As described
within the Results chapter, an evaluation of the delay between the 'peak' and the 'valley'
of this waveform and therefore of the activation lag between the agonist and antagonist
muscles provided a more meaningfùl portrait of the intensity of the out of phase
relationship. In particular, al1 members displayed an increase in the strength of the
relationship with practice that was statistically significant ( p c O.OS), see Figure 23B and
Appendix 8.
The third waveform demonstrated a local pattem of the motor task before
practice, in al1 but one of the experimental participants (P8). This pattem was defined by
a greater amplitude of the waveform towards the latter half of the task (i-e. tiom 50% to

100% performance time) as was seen in Figure 14D for participant (P7). The eigenvector
coefficients of W3 displayed values of contrasting poluities among the muscles within
each of the agonist and antagonist muscle groups. This evidently revealed that the nature
of W3 was to illuminate the individual roles, or more specifically, the activation patterns
of the within group muscles with respect to the production of the motor task.
The initial local concentration of the waveform before practice appeared to
convey two things: (1) the slowly rising slopes of the agonist EMG signals cornmensurate
with burst peaks that were closer to the onset of movement (see Figures 8 and 14A) and
(2) the preerninence of either one or a combination of antagonist muscles that were
involved in the task (see Appendix 1OA and B). On Day 1, six out of ten (PLP3, P5, P7,
and P9) or 60% of experimental members exhibited a dissociation between the PD versus
the LO and/or LA in which the latter muscles possessed a greater eigenvector loading. It
would appear that before practice, these participants subscribed to a proximal to distal
strategy that entailed a greater emphasis of the triceps muscles in the production of the
task (except for P4 and P l 0 who possessed a higher Pdel coefficient in relation to the LA
and the LA and LO coefficients respectively, see Appendix 10A and B). Furthemore. the
lesser amplitude of the first half of the waveform, that is associated with the agonist
muscles and which is temporally located at about the onset of movernent (Le. at 40% PT),
suggested that, for this penod, the proximal to distal characterization was essentially
underdeveloped.
For the same members above (N=6), either the Bic or BrR (i.e. the antagonist
muscles operating at the elbow joint, including the distal aspect of the biceps, a two-joint
muscle) was shown to be central in slowing down and/or stopping the limb's excursion to
the target (for P4 it was the Pec). This finding would be an appropriate complement to the
greater representation of the triceps muscles within the proximal to distal relationship of
the agonists described above. That is, the increased action of extending the elbow to
accelerate the forearm to the target would require an enhanced activation of the
antagonists subserving the elbow joint, to decelerate the distal limb. Concerning the result
of participant P4, a representation of the Pec (-0.51) would entai1 a logical selection for
this person who demonstrated a higher magnitude of the PD coefficient (0.61) in
opposition to the LA muscle (-0.5 1) within the agonist muscle sequence. An activation of

the proximal joint antagonist would seem necessary against the more forcehl action of
the abducting brachiurn produced by the leading role of the PD. This, however. was not
the case with P8 and P l 0 who also exhibited a predominating PD in their proximal to
distal pattern- Instead each of them demonstrated the Bic muscle as the preferential
antagonist in the motor task.
In two out of ten memben (P6 and P9) the local pattern of the waveform
characterized an even greater involvement of the antagonist muscie group. P6 displayed
the Pec versus the Bic, which outnurnbered the single LA of the agonist group, whereas
the Pec versus the Bic and BrR outnurnbered the PD versus the LO for P9. In fact, the
dissociation of the Pec fkom the Bic (a two-joint muscle) and/or the BrR, according to the
polarities of the eigenvector loadings for these muscles, actually demonstrated a proximal
to distal relationship of the antagonists. The greater magnitude of the Bic within the
aforementioned pattern of each of these individuals still highlighted the increased role of
the muscle among the antagonists in the motor task, as it had for the other participants
above. These findings are both unique and meaningful, in that they support the proximal
to distal programming of the agonist muscles discussed previously. For instance. the
sequential firing of the Pec versus the Bic and/or BrR would be necessary to decelerate
and stop the initial propulsion characteristics of the brachium, to get the two-joint system
moving, followed by the angular velocity of the forearm in that order. Moreover, the
greater angular velocity of the forearm. as dictated by the increased actions of the LA and
LO muscles for P6 and P9 respectively, explained the superior manifestation of the Bic to
counteract and brake the increasing angle at the elbow. For these volunteers then, the
presentation and/or coordination of the antagonists in arresting the movement of the limb
revealed a strategy that was equally as important if not more so than the initial commands
to get the ann moving to the target.
Finally, the facility with which P8 performed the motor task on Day 1 afler oniy
15 farniliarization trials was remarkable (PT: 185 I 1 msec). The outnght expression of
an unequivocally strong PD (-0.66) in contrast to the synergistic activation of the LA
(0.4 1 ) and LO (0.3 8) muscles before practice signalled the neurornotor systems capacity
to appropriately identiQ the prescriptions for action relating to a supenor rnotor
performance with minimal practice. In fact McGrain (1986) found similar evidence for

the application of an apparently suitable motor program to a novel motor task when
investigating a knee extension movement. In this study participants were required to push
a four wheel carriage through a timing gate located eight feet away at a criterion speed of
5 mph. There were no significant changes observed in either total movement time or
timing of EMG activity for two muscles across twenty practice trials of the task.
According to the author, the simplicity of the motor task may have resulted in the
seIection of a movement strategy which had established the necessary timing of EMG
activity for both muscles from the very beginning. The implication was that the
participants were able to sense the timing of muscle activity needed to perform the task
before they had executed the first trial. Therefore. even though P8 had no pnor
experience in the motor task of the current study, the participant was able to 'get a good
idea' of the timing characteristics required for a good motor performance after the first
few trials of the task.
With learning the shape of the waveform for W3 changed considerably for al1
experimental participants, into a global pattern whose augrnented magnitude spanned
across the entire task. It clearly revealed the development of the contrast in the firing
between the PD versus one or both of the triceps muscles, confinning the proximal to
distal strategy selected before learning. Other researchers have shown similar evidence
concerning the sequential progressions of initial agonist activations from axial joints to
peripheral ones (Wadman et al., 1980; Karst & Hasan, 1991; Vardaxis, 1996). This
sequencing of segmental rotations is more commonly known as the kinetic link principle
and is used for such patterns of movement as throwing, kicking and striking wherein the
action of the moving system is likened to the motion of a whip (Kreighbaum & Barthels,
1985). It involves an initial rotation of a stable base segment that is followed by a
fonvard rotation of the next distal segment. The latter only comes forward afier the
proximal segment has reached its peak angular velocity. Consequently, this scenano
creates an effect on the distal segment that is referred to as a 'lag'. In athletic situations.
the 'lagging back' of the distal segment@) is produced by a preliminary reversal in the
motion of a system most commonly observed as a 'wind-up'. Within the context of this
study's experimental task, which did not include a wind-up, a comparable lagging of the
foream may have been engaged by a progression of the brachium out and ahead of the

distal segment, a process called inertial lag (Kreighbaum & Barthels, 1985). As was
discussed earlier on in this chapter, it was the inertial lag of the forearm that may have
evoked a stretch reflex in each of the triceps muscles.
There were eight members who demonstrated the kinetic link principle via a
fortification of their agonist muscle sequences after the practice protocol. Five of these
people led with a hlgher muscle coefficient of the P D (P3, P5, P7, P8 and P9) whereas
three others possessed greater eigenvector loadings of one or both of the triceps muscles
(Pz, P4 and P6). In the latter three cases, the P D also showed an increase in its role in the
task with respect to Day 1, by virtue of a cornmensurate augmentation of its coefficient.
Alternatively, two individuals were unique in their portrayal of either a single highly
weighted LA or a prominent dissociation concerning the Pec versus the Bic and BrR.
respectively. These results communicated the effects of individual differences in the
performance of motor skills, such that the sequence of muscle activations for a particular
movernent are not necessanly the same and that participants will seek to enhance
performance in different ways (Corcos et al., 1993; Magill, 1993). ïhese findings are not
surprising since the members in this study were given no instructions about any particular
moLrement strategy that would assist them in their planning and/or execution of the motor
task. They were free to proceed at their own discretion.
Motor task practice also had the effect of diminishing the roles of the antagonists
in eight out of ten (80%) volunteers fiom pre- to post-leaming conditions, for which the
remnants of antagonist activity still featured the Bic or BrR acting at the elbow joint
(except for P i who displayed the Pec). Two others, P3 and P5, on the other hand.
revealed an increase in antagonist presence (see Figures 18 and 19). The general decrease
in the importance of the antagonist muscles following learning may reveal the CNS's
operation of economizing ancilor reducing the problem of the complexities of movement.
In other words, confronted with this study's novel whipping task, the above people were
initially more occupied with arresting the primary propulsion characteristics of the
agonists. Following the practice protocol, however, they learned to engage the
appropriate amount of whipping action to perfom the motor task more quickly and
efficiently without enlisting any greater a complement of antagonist muscles than was
necessary, In this manner, a simple decrease in the number of antagonist muscles

exploited during the execution of the motor task would indicate that the available
numbers of degrees of fieedom had been reduced by the CNS.
Additionally, an analysis of the cumulative variability accounted for, by the
eigenvalues of al1 three wavefoms (W's 1,2 and 3) put together, revealed that the value
for the expenmental group was higher on Day 5 than it was on Day 1 (Table 3). This
suggested a decrease in the variability of the muscle activation patterns with learning and
reinforced the fact that the participants had achieved a level of mastery in the motor task
which allowed them to perform in both an efficient and expedient manner.
4.2 Motor Task Retention Adaptations of the Agoni* and Antagonist Muscle
Groups: Functional Stages of S h o ~ - und Long-Term Motor Memory
In this study, a time-course evaluation of motor task retention was undertaken in
order to reveal the relative stability of the motor memory programs associated with
complex motor skill acquisition. The strength trainingdetraining literature of exercise
physiology research contains significant information regarding the impressionability of
strength-training induced adaptations to increasing periods of detraining (Hakkinen &
Komi, 1983 and 1985; Narici et al., 1989; Ishida et al., 1990; Staron et al., 1991 ).
However, these results have also alluded to certain selective, complex, post-training
adaptations which referred to a persistent alteration of the motor memory prograrn
subserving the particular motor skill. In fact 'periodization' of training schedules.
involving a reduction in one or more strength training variables such as volume of
training, has even shown improvements in motor skill performance to subsequent testing
or cornpetition (Hakkinen et al., 199 1 ).
Furthemore, more recent investigations in motor control have documented the
lasting effects of motor skill learning (e.g. over several months). These studies have
focussed on the neural mapping of motor memory correlates using functional M N and
Pet scans of rapid finger sequencing (Kami et. al., 1995) as well as of adjusted pointing
movernents to the torque pulses of a robotic manipulandum (Shadmer & Holcomb, 1997;
Shadmer et al., 1997). The above research will be used to help put the results of the
curent study into context.

4.2. I Short-term Changes l n Motor Memory
With one and two-weeks of no-practice respectively, neither did the shape of the
waveform for W1 nor the representations of the eigenvector coefficients conceming the
agonist and antagonist muscle groups. change measurably with respect to Day 5. Al1 of
the scores remained positive with unequivocal fluctuations in their magnitudes across the
two retention conditions for al1 memben of the expenmental group (for example. see
Figures 16 and 17B of participant P7). At the same time it was curious to note that 90%
of the people displayed a higher eigenvector loading for the LA at one-week post-
learning as opposed to 70% seven days later at Ret 2, see Figure 21B. The fact that there
were no changes conceming the eigenvalues, eigenvectors and waveforms of W1 for the
short-term memory conditions communicated the need to go beyond this waveform in
order to delineate how these retention intervals affected the memory of the motor task.
Changes in the 'out of phase' pattern between the agonist and antagonist groups
for the participants as a group, over Ret's 1 and 2, were difficult to ascertain using the
eigenvalue fluctuations and the structure of the eigenvectors across experimental group
members. However, statistics performed on the composite scores of the second
waveform, which revealed no significant differences in the tests of within subjects
contrats regarding the means fiom Day 5 (38.70%) to Ret 1 (38.10%), highlighted the
sisnikant decrease in the means From Ret 1 to Ret 2 (36.40%), p<0.009. These results
demonstrated a persistence of the intensity of the out of phase relationship with one-week
of no-practice. Therefore, the increase in antagonistic inhibition experienced with
learning was not lost or otherwise compromised seven days following the interruption of
practice. In this study, though, fourteen days rnay have constituted too long a time
without practice before the enhanced reciprocat inhibition between muscle groups was
influenced. indicating regression toward antagonistic CO-contraction.
The apparent extension of the increased latency between the two muscle groups at
Ret 1 begs the clarification of whether the ten retention trials (i-e. 5 preparation and 5
stable adaptation trials) were suffkient to comprise a retraining stimulus for the
volunteers. It is meaningful to consider since each of the participants did not correctly
achieve the first five stable adaptation trials sequentially and required more trials to attain

the criterion of tive. In particular, for each of the short-term retention periods as well as
the pre- and post-leaming trials of the control group, al1 of which occurred within a one-
week interval, the opportunity for additional trials could have constituted a factor limiting
the e ffec tiveness of the study 's treatment, namely abstinence from practice. Although the
above resdts concerning W2 may confirm that these 'extra' trials did not re-engage the
Iearning process or affect the retention intervals differently, the possibility that they could
have remains a question to be addressed in this investigation.
The third waveforrn (W3) was also sensitive in exposing changes to the Ieamed
adaptations of the motor task during the short-term periods of no-practice. To begin with,
the eigenvecton of the third waveform disclosed alterations of the within muscle group
relationships suggesting a lapse in motor coordination in only four people (PLP2, P4.
and P6) after the first retention interval. When cornpared to Day 5, the characterizations
of the agonist and antagonist muscles for these members were similar, although not
identical, to those exhibited on Day 1, before practice. They either manifested a
weakening of the proximal to distal relationship of the agonist muscles alone (Pl. P2), or
together with the reappearance of a significant antagonist presence (P4 and P6), (see
Appendices 10 and 11A and B). Researchers have previously alluded to the fragile
nature of short-terrn motor memory as revealed by the incomplete retention of both
simple and complex motor skills from one expenmental session to the next (Le. 24 to 48
hour intervais; Corcos et. al., 1993; Vardaxis, 19%).
It is important to note that although in the latter studies, the participants' next day
performance had been slightly beiow the peak attained at the end of the preceding day. it
was nonetheless above the level of performance exhibited at the b e g i ~ i n g of the
previous day. A between learning levels analysis was not performed in this investigation;
even so, similar findings of short-term motor memory lability should not be precluded in
this case. By extension, it would seem appropriate for the results of these participants
(N= 4), at one week post-learning, to reflect a cumulative effect of several days without
practice and hence a decrement in the coordination of the muscles producing the task.
Nevertheless, in three out of the four people for whom the latter situation was tme, rnotor
performance had not necessarily regressed to pre-training levels (e.g. P1-P2, and P6).
Wolpaw et. al., (1986), in their spinal stretch reflex studies have indicated that a major

function of supra-segmental control is to suppress spinal reflexes. This would
understandably allow for the perseverance of acquired skillful movements (Goode & Van
Hoeven, 1982). It may also underlie the observations of why the three participants still
exhibited some retention (however limited) of the learned adaptations of the motor task at
Ret 1. In other words, the CNS facilitates the acquisition and subsequent retention of the
motor commands having to do with skiIlfiil movements. This has a1so been demonstrated
within strength training/detraining paradigms conceming the selective persistence of
absolute measures of fast force production and maximal dynamic strength. These values
had decreased but were still greater than pre-training levels at 12 and 30-32 weeks post-
detrainhg respectively (Hakkinen & Komi, 1985; Staron et al., 1991). P4 was the only
individual of the above subgroup (N= 4) who displayed a complete reversa1 in learning
adaptations, experiencing what investigators of the detraining phenomenon would define
quite simply as " an effect comparable to the opposite effects of training" (Hakkinen &
Komi, 1983 and 1985).
The fact that the majority of experimental members (i.e. 60%: P3, P5 and
P7 - P10) showed a persistence or even an improvement in motor task performance at
Ret 1, further illuminated the capacity with which the CNS strives to replace the initial
pre-leaming rnotor comrnands with new ones borne out of deliberate and specific
practice. As a result, this group (N=6) confirmed that one week without training was not
deIeterious to the neuromuscuIar coordination acquired with training. Moreover, these
findings corroborated the results of the second waveforrn, which also revealed a
persistence and/or improvement in the learned adaptations for the experimental group at
Ret 1. Indeed as Brashers- h g et. al., (1996) and Shadmer & Brashers-Krug (1997)
have found with adjusted pointing movements, an interna1 mode1 (IM), which associates
a particular pattern o f muscle torques with the desired trajectory of a motor skill, is not
only stabilized but consolidated in rnotor memory at approximately 4 hours afler practice.
Thus people who learned the motor task were able to perfonn better on recall tests at 24
hours and up to three-weeks beyond training.
In accompanying experiments, the sarne authors tested the stability of the
acquired IM by intervening with a second, different, motor task following learning of the
initial one (i.e. retrograde interference). It was revealed that 5.5 hours between the two

tasks was sufficient time to allow for a significant recall of the first IM at one-week post-
training. When 24 hours had elapsed before the second task was introduced, recall of the
first IM approached the performance index of controls who had only practiced the first
task. Ishida et. al., (1990) have also suggested that short-term strength training may
actually lead to rather significant post-training adaptations (i-e. consolidation), regarding
an increase in the maximal rate of force development in the human triceps surae muscle
at four and eight weeks post-detraining. Thus, it would appear that in the present
investigation, the majority of experimental participants had consolidated the prescriptions
for action associated with the motor task in motor memory and that the motor program
itself was seemingly resistant to forgetting for up to a one week period without practice.
At two weeks post-leaming, however, the experimental group featured six
participants (P2, P5 and P7- P10) who expenenced decrements in their motor
coordination, with respect to the leamhg adaptations engendered with practice and
maintained over a one week penod, as described above. These individuals (N=6)
eshibited a deterioration in the proximal to distal relationship of the agonist muscles
including an increased representation of the antagonists [e-g. for P2 (BrR: -0.52) and P9
(Pec: 0.68 vs. the BrR: -0.47), see Appendix 1 1A and BI. The attenuation in the proximal
to distal sequencing of the agonist muscles together with the greater involvement of the
antagonists probably alluded to the decreasing acceteration and increasing deceleration
profiles of the movement. Consequently, the PT for these individuals was observed to
increase, indicating that they were slowing down.
These generahzed results of a decrease in motor performance with no-practice
would appear to agree, in principle, with the findings of strength trainingdetraining
paradigms, concerning the reversible nature of training-induced adaptations. In the
aforernentioned investigations, common decreases in: maximal dynamic or isometnc
contraction forces, associated neural activations (Le. maximum integrated EMG' s) and
size andlor percentages of fast twitch fibers (type IIa) are usually observed when the
training stimulus has been removed for up to 12 weeks. The changes however, were not
to pre-training levels (Hakkinen & Komi, 1983 and 1985; Narici et al., 1989; Staron et
al., 1991).

A reversa1 in the participants' adaptations within the current study at Ret 2
suggested a decay in the motor prograrn subserving the task. Whereas an upper limit in
the stabiiity of the motor program consolidated in rnemory was estabfished at one week
post-learning, two weeks without training was apparently too long a time to keep the
program from deteriorating. Moreover, the data of the third waveform afforded hrther
evidence against the argument that the re-testing trials of the retention conditions
provided an additional stimulus to the participants, which may have contributed to a
further improvement in their performance. If these trials were re-engaging the learning
process, then the trials executed within the first retention session (Ret 1) should have
reinforced the temporal sequencing features of the muscles involved in the task to curtail
any decrements in the motor prograrn at Ret 2; this was not the case.
4.2.2 Long-term Changes in Motor Memory
Across each of the long-term retention conditions, fiom four to eight weeks
beyond the completion o f training, the general shape of the first waveform (WI) did not
reveal any significant changes. Also, the characteristic preeminence of the LA muscle
within the majority o f the eigenvectors among the experimental participants was
maintained.
An analysis of the composite scores conceming the second waveform did not
revea1 any significant effects among the intervals of the long-term motor memory period
(see Appendices 12 - 13, A and B). In fact the fluctuations in the values fiom nvo- io
eight weeks post-learning were very small and did not provide any notable increase or
decrease in the out of phase relationship between the two muscle groups. Consequently.
the status of the last meaningful alteration in the out of phase pattern, regarding the
decrement in reciprocal inhibition expenenced from Ret 1 to Ret 2, remained the modus
operandi for each of Ret's 3 ,4 and 5 .
Modifications, in the phasic adaptations of the muscles within each of the agonist
and antagonist groups, as delineated by the eigenvectors of the third waveform, did not
necessarily expose any progressive deterioration in motor coordination across
participants. Lnstead, in each of Ret's 3, 4 and 5 there was a majority of expenmental
members who demonstrated either a persistence o r even an improvernent in motor
79

performance with respect to the preceding long-term retention period. Wi th few
exceptions the observed irnprovements in performance were accompanied by decreases in
performance time; however, the reductions in PT were never greater than the PT'S
exhibited on Day 5 or at Ret 1. This clearly indicated that the improvements in movernent
production were probably the result of compensatory adj ustments to the prevailing
coordination of the muscles involved in the motor task. In other words, in view of the
findings concerning a probable decay in the motor program at Ret 2, the participants who
returned for testing during the long-term retention penods were likely trying to make up
for the decrement in performance that they were experiencing. To that end, they used
whatever rnernory of the skill was still available to hem, in addition to any extra 'cues'
they could get from both the preparatory and stable adaptation trials at the time of the re-
testing sessions to execute the task as quickly and as accurately as possible.
The changes observed in the cumulative variability accounted for by the surn of
the three waveforms across the retention conditions were not very large but a decrease in
the values by the end of the retention period indicated an increased variability among the
EMG patterns (see Table 3). This reinforced the finding that the experimental participants
were no longer perfonning with the sarne certainty in the motor task as they had been on
Day 5 and Ret 1, periods which characterized a greater automaticity in their motor
responses.
4.3 Ch anges in the Performance Production and Outcome Measures
of the Control Group
The effects of the control group protocol in producing adaptations among its
members over the leaming condition of the experimental penod (i.e. Day 1 to Day 5) that
were sirnilar to the changes observed in the experimental group were unexpected but not
totally surprising. The changes in the adaptations for each of the groups over the retention
period (i-e. Day 5 to Ret 5) , however, were different and alluded to a superficial leaming
effect in the control group. To begin with, the twenty scheduled trials afforded to the
control members on Day 1 were evidently sufficient to stimulate the organization of a
motor program that accommodated the temporal features requirements of the motor skill.
An analysis of the composite scores fiom the second waveform (W2) for both
80

experirnental and control groups demonstrated that the difference in the rate of increase
in the intensity of the out of phase relationship observed from Day 1 to Day 5 was
statistically insignificant. Nevertheless, the experimental group sti 11 exhi bited a much
greater index of change (Le. a 20.6% increase) with training as opposed to the control
group (i-e. a 5.6%) which did not receive specific training in the task. Similarly, the
control group succeeded in exploiting the proximal to distal strategy of muscle activation
among the agonists and reduced the activation of the antagonists, to decrease their
performance time. However, inspection of the percent changes in PT measures over both
groups from Day 1 to Day 5 showed that the experimental group realized a 47% decrease
in PT as compared to the 4.3% reduction attained by the control group. Therefore, even
though the control group displayed adaptations in both performance production and
outcome measures fiom Day 1 to Day 5, these changes were substantially smaller than
the adaptations experienced by the experimental group as a result of the training protocol.
From Day 5 to Ret 5, the trends in the changes observed for each of the two
groups were found to be significantly different. For instance, the experimental group
experienced a reduction in the intensity of the out of phase relationship between the
agonist and antagonist muscle groups that indicated an increase in antagonistic co-
contraction. The majority of participants within this group also exhibited a regression in
the adaptations of the within muscle group relationships which showed that the proximal
to distal pattern of the agonists was weakened and that there was an increase in the
activation of the antagonists in the motor task. As a result, the overall performance time
of the experimental group was observed to increase fiom post-training to post-retention
trials. In contrast, the control group demonstrated an additional increase in the out of
phase pattern for the second waveforrn and a decrease in performance time over the same
interval. Moreover, an increase in the cumulative variability accounted for, by the sum of
al1 three waveforms for this group, indicated that the muscle activity patterns for the Ret
5 condition had becorne less variable than on Day 5 (see Table 4), which supported the
ongoing improvement in motor performance.
Consequently, the differences in the results of each group were significant in
highlighting a functional level of learning for the expenmental group and a superficial
level of acquisition for the control group. That is, participants in the former group had

achieved a degree of mastery in the motor task for which the consequences of terrninating
practice entailed a decrement in motor performance. In contrat, the members of the
control group had procured a comparativeiy limited degree of task proficiency that
defined a substantial performance reserve. It would explain their inclination to improve
upon a performance level on Day 5 that was not very high to begin with.

CHAPTER V
SUMMARY
The current research investigated the retention charactenstics of the acquired
neuromuscular adaptations in a fast complex motor task. Specifically, the aim of this
study was to elucidate the stability of the temporal sequencing of muscle activations
subserving the movement in motor memory, when specific training in the task has been
terrninated. Furthemore, how Bernstein's degree of freedom theory applied to the
changes in rnotor coordination during retention of the motor task, was aiso assessed. ft
was presumed that eiectromyographic activity provided a 'gateway' to the central
processing of the neural commands to the muscles involved in the motor task. It was
h)~othesized that the between and within muscle group adaptations with leaming would
be compromised during the retention penod.
In this study an experimental group cornprising ten participants was trained over a
four day learning protocol to execute targeted arm movements in the horizontal plane at a
maximal speed. They were re-tested at 1, 2, 4, 6, and 8 weeks following the end of the
leaming period which constituted the retention protocol. The first two weeks post-
leaming were categorized as a 'short-terni retention period' while a 'long-term retention
penod' was defined by four to eight weeks post-learning. A control group was also tested
in the motor task at pre- and post-learning conditions as well as at the end of the retention
penod for comparative purposes. Surface electromyography was used to monitor the
electrical activity of six muscles of the iefi upper limb and chest. EMG electrodes of a
bipolar configuration were placed directly over the bellies of the posterior deltoid,
clavicular pectoralis major, long and lateral heads of the triceps brachii, long head o f the
biceps brachii and brachioradialis. A reference electrode was placed over the right
clavicle. A method of singular value decomposition (SVD) was employed to determine
the minimum number of waveforms required to describe the set of six muscle activation
patterns.

5.1 Concluswns
Experimenfal Group
( 1 ) The learning protocol resulted in significant decreases in performance time for the
motor task, among the participants in the expenmental group. This performance
outcome rneasure appropnately defined motor skill acquisition in this
investigation.
(2) The learning protocol produced an increase in reciprocal inhibition between the
agonist and antagonist muscle groups in the motor task that detennined a decrease
in antagonistic CO-contraction within the shoulder and elbow joints involved in the
movement.
( 3 ) Learning enhanced the proximal to distal activation of the agonist muscles across
the shoulder and elbow joints and reduced the number and/or intensity of
antagonist muscles that were significantly exploited in the motor task. A decrease
in the representation of the antagonists with learning fulfilled Bernstein's theory
conceming a reduction in the degrees of freedom of movement with motor skill
acquisition.
(4) The re;ention protocol resulted in significant increases in performance time for
the motor task beginning at two weeks post-learning of the short-term retention
period. This performance outcome rneasure provided the first index of a
decrement in the motor performance arnong participants.
( 5 ) The retention protocol revealed a diminution in the reciprocal inhibition between
muscle groups observed with learning at two weeks beyond the completion of
training, in light of an increase in antagonistic CO-contraction within the shoulder
and elbow joints subserving the movement.
(6) The retention protocol also demonstrated that the learning adaptations of the
within muscle group relationships were compromised at two weeks post-learning.
The decrements included a weakening of the proximal to distal pattern of

activation arnong the agonist muscles and a relative resurgence in the number
and/or intensity of the antagonist muscles in the motor task. An increase in the
representation of the antagonists indicated a reversal in the reduction of the
nurnber of degrees of freedom with learning.
(7) The motor task was simple enough to engage significant improvements in both
the performance outcome and production measures of the control g r ~ u p .
Nevertheless, these adaptations characterized a superficial leaming of the motor
task arnong these members and reinforced the efficacy of the learning protocol for
the experimental group, which resulted in the achievement of a functional levei of
mastery in the motor task for the latter.
5.1 Recommendations for Future Research
The current study has raised some pertinent issues for tuture investigations into
motor skill learning and retention that include: (1) the nature and complexity of the motor
task employed in the research, (2) the performance index required of the participants and
(3) the skill level of the individuals taking part in the study.
First, the nature of the motor task used in the research must be explicitly specified
and categorized in order to allow for appropriate comparisons with the findings of both
existing and tiiture studies using similar tasks. For instance, the complex g r o s motor
whipping movement involved in the present study did not Iend itself entirely to
comparison with the majority of pointing andhr reaching movements of motor control
research. Therefore, more study is required on the ballistic tasks that are inherent in many
athletic skills such as throwing, kicking and striking with or without an implement-
Second, the practice or training used within a learning protocol must realize a
performance plateau in the participants involved, to eliminate any question of a
performance resenre among the individuals. This would allow for a clear delineation of
the performance level in a motor task and facilitate the comparison to other experimental
sessions a d o r members. For instance, in the current study, although the experimental
group members exhibited substantial improvements in performance time, there was no 85

guarantee that these changes necessarily characterized the performance peak within each
of these individuals.
Finally, an account needs to be made conceming the ski11 level andor prior
experience of a prospective participant so that the results of an investigation are
appropriate to the treatment adrninistered and not confounded by the particular abilities of
the individuals comprising the experimental group.

Accomero, N., Berardelli, A., Argenta, M. & Manfiedi, M. (1984). Two-joints ballistic arm movements. N e w o s c k e L e m , 4 , 9 1 -95.
Ammons, R. B., F m , R. G., Bloch, E., Neuman, E., Dey, M., Marion, R., & Ammons, C. H. (1958). Long-term retention of percephial-motor skills. Journal of M, II, 318-328.
Angel, R.W. (1974). Electromyography during voluntary movement: the two-burst . *
pattern- Electro- and- Neur-, 16,493-498.
Angel, R. W. (1975). Electrornyographic patterns during ballistic movement of normal and spastic limbs. Reseatçh, B, 387-392.
Angel, R. W. (1975). Myoelectric patterns associated with ballistic movement: effect of unexpected changes in load. Journal of Movem-, 1,96- 103.
Angel, R. W. (1977). Antagonist muscle activity during rapid arm movements: central versus proprioceptive influences. New-v and -, 4,683-686.
Bell. H. M. (1950). Retention o f pursuit rotor ski11 after one year. Jounial of 1 P s v c m , a, 648-649.
Berardelli, A., Rothwell, J. D., Day, B. L., Kachi, T., & Marsden, C . D. (1984). Duration of the first agonist EMG burst in ballistic a m movements. Drain Research, 304, 183-1 87.
Berardelli, A., Accornero, N., Argenta, M., Meco, G. & Manfiedi, M. (1986). Fast complex arm movements in Parkinson's disease. Journal of Neuro-, a, 1146- 1149.
. . Bernstein, N. A. (1967). Tb-on a d r-on of m o v m . London:
Pergammon Press.
Brashers-Kmg, T., Shadmehr, R., & Bizzi, E. (1996). Consolidation in hurnan motor memory. P m , m, 252-254.
Brooks, B.V. (1986). & N e u m O f m t o r C o M . New York: Oxford University Press.
Brown, S. S. C, & Cooke, J. D. (1981). Amplitude- and instruction- dependent modulation of movement-related electromyogram activity in humans. Journal of -, u, 97-107.

Brown, S. H. & Cooke, J.D. (1984). Initiai agonist burst duration depends on movement amplitade. m, Research,, 523-527.
Brown, J. M. M. & Gilleard, W. (1991). Transition fiom slow to ballistic movement: development of triphasic electrornyogram patterns. of AD-
Caminiti, R., Johnson, P. B. & Urbano, A. (199O).Making arrn movements within different parts of space: dynarnic aspects in the primary motor cortex. Journal of
e u e , J.Q, 2039-2058.
Corcos, D. M., Jaric, S., Agarwal, G.C., Gottlieb, G.L. (1993). Principles for leaming single-joint movements. 1. Enhanced performance by practice. E w -, B, 499-5 13.
Engelhom, R. (1983). Agonist and antagonist muscle EMG activity pattern changes with ski11 acquisition. -ch -, 54, 3 15-323.
Engelhom, R. (1987). Effects of ski11 practice on electromyographic activity pattems and fiequency spectra. Human S c h , 6, 1 1 7-1 3 1.
Finley, F., Wirta, R., & Cody, L. (1968). Muscle Synergies in motor performance. rchives of Ph ,*, 655-660.
Flanders, M. (1 99 1). Temporal patterns of muscle activation for arm movements in three- dimensional space. Journal of Neur-, 11,2680-2693-
Fianders, M. & Hermann, U. (1992). Two components of muscle activation: scaling with the speed of arm movement. J o u n i a l s i o m , 67,93 1 -943.
Flanders, M., Pellegrini, J. J., & Geisler, S. D. (1996). Basic features of phasic activation for reaching in vertical planes. R e s e d , m, 67-79.
Fleishman, E. A. (1962). Factors in the retention and releaming of perceptual-motor skiIl. Journal of E- Ps-, 64,2 15-226.
Gabriel, D. A. (1995). Muscle activation pattems for goal-directed multi-joint arm movements. Unpublished thesis.
Garland, H. & Angel, R.W. (1971). Spinal and supraspinal factors in voIuntax-y movement. Exbenmental Neuroloa, 23,343-350.
Garland, H., Angel, R. W. & Moore, W. E. (1972). Activity of triceps brachii during voluntary elbow extension: effect of lidocaine blockade of elbow flexors.
loey, x, 23 1-235.

Goode, D. J. & Van Hoeven, J. (1982). Loss of patellar and achilles tendon reflexes in classical ballet dancers. Archives of Neurology, 39, 323.
Gottlieb, G. L., Corcos, D. M., Jaric, S., & Agarwal, G. C. (1988). Practice improves even the sirnplest movements. m, Research,, 436-440.
Graves, J. E., Pollock, M. L., Leggett, S. H., Braith, R. W., Carpenter, D. M., & Bishop, L. E. (1988). Effect of reduced training fiequency on muscular strength. . .
e m m of Spprts M-, 9, 3 16-3 19.
Hakkinen, K. & Komi, P. V. (1983). Electromyographic changes during strength training . . and detraining. Me- Science in v, Exercise,, 455-460.
Hakkinen, K., Alen, M. & Korni, P.V. (1985). Changes in isometric force and relaxation- time, electrornyographic and muscle fiber characteristics of human skeletal muscle during strength training and detraining. Bçta Physiologjca Scandinavicg, 125,573-585.
Hakkinen, K., Kallinen, M., Komi, P. V., & Kauhanen, H. (1991). Neuromuscular adaptations during short-term "normal" and reduced training penods in strength . . athletes. Electr- C h c a l N e v s i o I o g y , 2, 3 5-42.
Hallett, M., Shahani, B. T., & Young, R. R. (1975). EMG analysis of stereotyped voluntary movements in man. Journal of Neurology. New-. and PSVC-, 3, 1 154- 1 162.
Hallett, M. & Marsden, C. D. (1979). Ballistic flexion movements of the human thurnb. j o u d of-, m , 3 3 - 5 0 .
Hammerton, M. (1963). Retention of learning in a difficult tracking task. m a i of PsychQ1Qgy, 66,108- 1 10.
Hasan, Z., & Karst, G. M. (1989). Muscle activity for initiation of planar two-joint arm movements ion different directions. ExDerimental R r b R e s e d , x,65 1-655.
Hobart, D. J., Vorro, J. R. & Dotson, C. 0. (1978). Synchronized myoelectric and cinematographic analysis of ski11 acquisition. Journal of H w MovemeN u, 4, 155-166.
Hobart, D. J., Kelley, D. L., & Bradley, L. S. (1975). Modifications occumng during . - acquisition of a novel throwing task. A- J o d of Ph-al Medicine, 54, 1-24.
Hortobagyi, T., Hournard, J. A., Stevenson, J. R., Fraser, D. D., Johns, R. A., & Israel, R. G. (1993). The effects of detraining on power athtetes. Medecine Science b
,x, 929-935.

Ishida, K., Moritani, T, & Itoh, K. (1 990). Changes in voluntary and electrically induced contractions during strength training and detraining. J o u d of A~plied v, a, 244-248.
Jaegers, S.M.H., Peterson, R.F., Dantuma, R., Hillen, B., Geuze, R., & Schellekens, J. (1989). Kinesiologic aspects of motor learning in dart throwing. -, Studies,, 161-172.
Jahnke, J. C. (1958). Retention in motor learning as a function of arnount and practice and rest. *, Psvcholoev,, 270-273.
Jaric, S., Corcos, D. M., Agarwal, G. C., & Gottlieb, G. L. (1993). Principles for learning single-joint movements. II. Generalizing a learned behavior. ExDenmental Br& R e s e b , 9_4,5 14-52 1.
Kamon, E., & Gormly, J. (1968). Muscular activity patterns for skilled performance and during learning of a horizontal bar exercise. -, u, 345-357.
Kami, A., Meyer, G., Jezzard, P., Adams, M. M., Turner, R., & Ungerleider, L. G. (1995). Functional MRI evidence for adult motor cortex plasticity during motor skill learning. m, m, 155-1 58.
Karst. G. M. & Hasan, 2. (1991a). Initiation rules for planar two-joint arm rnovements: Agonist selection for movements throughout the workspace. Journal of
euro-, 66, 1579-1 593.
Karst, G. M. & Hasan, 2. (199 1 b). Timing and magnitude of electromyographic activity for two-joint a m movements in different directions. J-l of NeurophvsiolQgy, 66, 1594- 1604.
Kearney, R. E. (1994). Electromyography. fimedical I m m e n t a t i ~ and Measurement Techniaues, 1 - 10.
Kreighbaum, E. & Barthels, K. M. (1985). -CS: A aualitati . . *. movement.imeapoiis, MN: Burgess publishing Company.
Lacquaniti, F., Soecthing, J. F. & Terzuolo, S. A. (1986). Path constraints on point-to- point a m movements in three dimensional space. Weuroscience, 1Z,3 13-324.
Lagerlund, T. D., Sharbraugh, F. W. & Busacker, N. E. (1997). Spatial filtering of multi- channel electroencephalographic recordings through pnnciple cornponent analysis . . by singular value decomposition. J o u d of C W a l Neurçghysiol~ey, 14, 73-82.
Lestienne, F. (1979). Effsts of inertial load and velocity on the braking process of voluntary lirnb movements. -, 31,407-418.

Ludwig, D. A. (1982). EMG changes during acquisition of a motor skill. Atnerica M a , 61,229-243.
. . Magill, R. A. (1993). M o f o r t s and (41h ed.). Dubuque, IA:
Wm. C. Brown.
Mah, C. D., Hulliger, M., Lee, R. G. & 07CalIaghan, 1. S. (1994). Quantitative analysis of human movement synergies: Constructive pattern analysis for gait. J o u d of
otor B*, 26, 83-1 02.
Manng, J. R. (1990). Effects of mental practice on the rate of skill acquisition. msiçial T h e r w , Zn, 165-1 72.
Mchdle, D. W., Katch, F. I., Katch, V. L. (1991). u i s e P ology: E- . . ~ u m t i o n ( 3 ~ 4.). Philadelphia: Lea g e b i g e r .
McGrain, P. (1986). Myotemporal changes associated with learning a simple motor task using two different strategies of movement.
Melnick, M. J. (1971). Effects of overleaming on the retention of a gross motor skill. The -ch Q m , 42,6 1-69.
Narici, M. V., Roi, G. S., Landoni, L., Minetti, A. E., & Cerretelli, P. (1989). Changes in force, cross-sectional area and neural activation during strength training and detraining of the human quadriceps. -an Jouml of A~plied Ph-v and Occ-, s, 3 10-3 19.
Newell, K. M. (1991). Motor skill acquisition. -view of Ps , 42, 213- 237.
Normand, M. C., Lagasse, P. P., Rouillard, C. A. & Tremblay, L. E. (1982). Modifications occurring in motor programs during learning of a complex task in man. -, m,87-93 .
Patton, N. J. & Mortenson, O. A. (1970). An electromyographic study of reciprocal activity of muscles. Anatomiçal records, m, 255-268.
Payton, O. D., Su, S., Meydrech, E. F. (1976). Abductor digiti quinti shumeboard: a . . . . . study in motor leaming. & & i v e s of Ph- M e d i c i n e _ a n d , 57, 169-1 73.
Payton, O. D., Kelley, D. L. (1972). Electromyographic evidence of the acquisition of a motor skill. -, 2 , 2 6 1 -266.
Pellegrini, J. J., & Flanders, M. (1 996). Force path curvature and conserved features of muscle activation. m, UQ, 80-90.

Person, R. S. (1958). An electromyographic investigation on coordination of the activity of antagonist muscles in man during the development of a motor habit. PavIovim
tivity, 8, 13-23.
Sanes, 1. N. & Jennings, V. A. (1984). Centrally prograrnrned patterns of muscle activity in voluntary motor behavior of humans. E- Br-, %,23-32.
Shadmehr, R., & Brashers-hg, T. (1996). Functional stages in the formation of human long-term motor memory. -Neuroscience, U , 409-4 19.
Shadmehr, R., & Holcomb, H. (1997). Neural correlates of motor memory consolidation. Sçience, m, 821-825.
Smeets, J. B. J, Erkelens, C. J. & van der Gon, J. J. D. (1993). Adjusmients of fast goal- directed movements in response to an unexpected inertial load. Experimental -, a, 303-3 12.
Soecthing, J. F. (1988). Effect of load perturbations on EMG activity and trajectories of pointing movements. Rese-, a, 390-396.
Staron, R. S., Leonardi, M. J., Karapondo, D. L., Malicky, E. S., Falkel, J. E., Hagerrnan, F. C., & Hikida, R. S. (1 991). Strength and skeletal musclz adaptations in heavy- resistance-trained women after detraining and retraining. m a l of AD^
s r o u , B, 63 1 -640.
Stokes, V. P., Lanshammar, H. & Thorstensson, A. (1 999). Dominant pattern extraction from 3-D kinematic data. IEEE on J3io&cal E-, 46, 100- 106.
Turner, R. S., Owens, W. M., & Anderson, M. E. (1995). Directional variation of spatial and temporal characteristics o f limb movernents made by monkeys in a two- dimensional work space. Journaleurqphysiology, 74,684-697.
Van Bolhuis, B. M., & Gielen, C. C. A. M. (1997). The relative activation of elbow- flexor muscles in isometric flexion and in flexion/extension movements. j o u d of Riomec-, x, 803-8 1 1.
Vandervoort, A. A. (1992). Effects of aeging on hurnan neurornuscular function: implications for exercise. J o u d of Sport Sdence~ , E 7 18-1 84.
Vardaxis, V. (1996). Learning adaptations in performance production measures of novel multi-joint tasks. Unpublished Thesis.

Vorro, J. & Hobart, D. (198la). Kinematic and myoelectrical analysis of skitl acquisition: * . . . .
1. 90 cm subject group. Archives of m a l Mec, 62, 575-582.
Vorro, J. & Hobart, D. (198 1 b). Kinematic and myoelectrical analysis of skill acquisition: - . . . . 1. 150 cm subject group. Archives of P w i n e and, 62, 582-589.
Wacholder K., & Altenburger, H. (1 926). Beitrage mr physiologie der willkurlichen bewegung. X. Mitteilung. Enzelbewegungen. Pflùeers fur die usamte Phvsiolo- der der, m, 642-66 1.
Wadman, W. J., van der Gon, J. J. D. & Derksen, R. J. A. (1980). Muscle activation patterns for fàst goal-directed arm movements. Jourtliil of M o v m s tuh, 6, 19-37.
Warfel, J. H. (1993). THE: MU-IJ&. Philadelphia: Lea and Febiger.
Waters, P. & Strick, P. L. (1981)- Influence of 'strategy' on muscle activity during ballistic movements. -eu&, m, 189- 194.
Wolpaw, J. R., Braitrnan, D. J. & Segal, R. F. (1983). Adaptive plasticity in the primate stretch reflex: initial development. J o u n i a l s i o l o e y , s, 1296-1 3 1 1.
Wolpaw, J. R., Segal, R. F. & O'Keefe, J. A. (1983). Adaptive plasticity in pnmate spinal stretch reflex: behavior of synergist and antagonist muscles. Jounial of Neuro-, ig, 1312-1319.
Wolpaw, J. R. (1983). Adaptive plasticity in the pnmate spinal stretch reflex: reversal and redevelopment. R e s d , 278, 299-304.
Wolpaw, J. R., O'Keefe, J. A., Noonan, P. A. & Sanders, M. G. (1986). Adaptive plasticity in primate spinal stretch reflex: persistence. Journal of
eur-, S, 272-279.
Wolpaw, J. R. (1994). Acquisition and maintenance of the simplest motor skill: . - invetsigation of CNS mechanisms. Mediçine and Science in Sports & Exercise, 2, 1475- 1479.
Woods, J. J. & Bigland-Ritchie, B. (1983). Linear and non-linear surface EMGlforce relationships in hurnan muscles. Amencan Medeck , 62.287- 299.


Appendix 1
LITERATURE REVIEW
A review of the related literature to the major topics of the present study will be
categorized as follows: firstly, a preliminary discussion of the electromyographic (EMG)
activity involved in motor skill learning will ensue as it pertains to single- and multi-joint
actions respectfully. Considering that true expressions of single joint movements are rare
in 'real', everyday activity, the focus on such laboratory based tasks does provide a
fundameatal assessrnent of the operation of the central nervous system (CNS) during
movement production. As a resutt, they are a necessary starting point to building a
progressive understanding of more complex ideas, such as when enquiring into the
electrophysiological dynamics of multi-segmental systems.
Secondly, a presentation of the information pertaining to motor skill retention, as
i t applies to the context of this investigation, will conclude the review.

EMG Adaptations of Single- and Multi-Joint Movements with Practice
Simple Movements
A biphasic or triphasic muscle activation pattem has been consistently
documented in the literature with respect to simple voluntary goal-directed movement
that is rapidly executed. As early as 1926 (Wacholder ar,d Altenberger), an initial agonist
volley (AGI) was observed to be followed by a 'quiet period' of reduced electncal
activity during which time the antagonist (ANT) woutd fire. A second agonist burst
(AG2) terminated the electrical event.
If fast, high speed or skilled ballistic trajectories are characterized by the above
motor output pattern (Hallet & Marsden, 1979; Lestienne, 1979; Brown and Cooke.
1980; Mustard & Lee, 1987; Brown & Gilleard, 1991), slow movements display a more
continuous agonist excitation in light of a diminished or complete pause of antagonist
activity (Patton & Mortenson, 1970; Hallet et al., 1975; Lestienne, 1979). Even slower
speed rarnp movements are manifested by agonist/antagonist CO-contraction.
The role of each of the EMG components as they relate to the kinematics of a fast
movement can be described as follows: AGI constitutes the propulsive force initiating
targeted limb movement, while the subsequent 'silent period' is apparently reciprocally
matched by ANT which attenuates the limbs acceleration in a deceleratory phase. AG2 as
the tertiary burst is also present during limb deceleration and may assist in stabilizing the
joint at its terminal end-point position.
The etiology and functional significance of the triphasic pattern within the CNS
has heretofore received comprehensive attention. Numerous investigators have proposed
a central origin for AG1 and ANT whereas the strong variability accrued in the assay of
AG2 has attested to its penpheral mediation (Garland & Angel, 1971; Garland et al.,
1972; Angel, 1974; 1975; 1977; Hallet et al., 1975). Regardless of the apparent pre-
construction of the first two EMG bursts however, researchers employing various
perturbations and paradigms of movernent have exploited the impressionability of these
vo1leys to segmenta1 manipulation (Hallet & Marsden, 1979; Brown & Cooke, 1980;
Waters & Strick, 1981; Berardelli et al., 1984; Sanes & J e ~ i n g s , 1984). Thus central

commands issued as starting points to a movement could still be modified at the
periphery according to the later properties of an action.
Using electromyography, researchers such as Person (1956) and Kamon &
Gormely (1968) studied the changes in muscle activity that accompanied the skilful
performances of sorne 'real-life' and athletic tasks. For instance, during early training of
filing and cutting with a chisel, Person observed the simultaneous activations of triceps
(agonist) and biceps (antagonist) muscles that were central to the tasks, arnong
participants.
Specifically, early movements were affected by an increased activation of the
agonist when the antagonist was excited a? the same time, resulting in erratic,
unrhythmical actions. After training, rhythmical manifestations of the movements were
CO-ordinated by reciprocally prograrnmed onsets and offsets of agonist and antagonist
volleys. Hence, the principle forcehl nature of the triceps and the ulterior corrective role
of the biceps (in braking the movement at specific times) were clearly delineated within
the tasks respectively.
In novices executing a gymnastic single-knee circle mount on the horizontal bar,
Kamon and Gomley witnessed the EMG bursts of several muscles investigated in the
hip, tmnk and amis which were characterized by extended durations contributing to
frequent overlapping or CO-contraction before training. As the panicipants performed the
exercise more fluently in the terminal stages of practice. however, there was a greater
incidence of sequential activations arnong muscles (Le. decreased CO-contractions) which
were ful filling more speci fic roles in the motor task.
Payton and Kelley (1972) devised a pilot study to determine the EMG
charactenstics o f the anterior deltoid and biceps brachii with the learning of a novel ball-
tossing task. Expenmental participants, who were fitted to a straight-backed chair, had a
Ieather cuff secured upon the forearm o f the dominant arm, which was placed in the
anatomical position. Attached to the cuff was a plastic cup containing a standard ping-
pong ball. The aim was to propel the ball to a target bucket whose rim was 76 cm away.
Both the shoulder and elbow joints were fiee to move. A decrease in the total electrical
output and in the rise time for the biceps, regarded as the focal muscle in the task, was

found with learning in cornparison to a non-significant increase in the activity of the
proximal joint agonist.
Experiments by Hobart, Kelley and Bradley (1975) used a similar method of an
underhand ball-tossing task to describe the movement changes with learning and funher
elucidate the electrophysiological manifestations subserving the action. To that end they
restricted the movement to the shoulder joint and an assay of the anterior deltoid and
pectoralis major as agonists versus the posterior deltoid and triceps brachii as antagonists
in the task. The target consisted of a 72 square inch board of concentnc circles with a
common centre. It was 150 cm away fiom the base of a throwing chair and at an
inclination of 22 degrees fiom the floor. Following 150 trials of the task, experimental
participants had decreased the limb angle at bal1 release while perfoming at higher
velocities early on in the movement to reduce both their throwing error and movernent
tirne. A decrease in the total electrical activity of the anterior deltoid with no change in
the pectoralis was observed concomitant with a decrease in the time to reach peak activity
by both agonists. These changes figured prominently in providing a greater impulsive
force to augment the velocity profile of the limb early on in the movement. Altematively.
increased activity in each of the antagonist muscles and a decreased latency of the
posterior deltoid, with respect to the onset of its agonist counterpart, contnbuted in
reducing the higher initial velocity of the limb and to release the bal1 earlier at an
increased velocity.
Payton, Su and Meydrech (1976) elaborated on the significance of the apparent
equivocal activity conceming the prime movers of a motor task fiom pre- to post-leming
conditions. To that end they restricted their focus on only the abductor digiti quinti of the
fifih digit in a shuffleboard type task. They found that sets of 100 trials used to reach a
learning cntenon provided no statistical difference between the E-MG data of the pre-
practice and post-practice trials. They concluded that the results reinforced a hypothesis
originated with the findings of Payton and Kelley (1972); narnely that the agonists
pertaining to a task resolve fiom a previously 'undifferentiated field of muscles' into the
prime movers and auxiliaries of a movement. The former, remain at a comparable level
of electrical activity as the latter show a decrease in activity with motor ski11 leaming.

In cornparison to previous results, however, specifically those upon which the
hypothesis was originally predicated, the postdates do not necessarily represent the
findings. For instance, in the pilot study by Payton & Kelley, it was the biceps, a two-
joint muscle, which was singled out as the probable prime mover in propelling the ball to
the target. Although no data was collected regarding the onset times of either the anterior
deltoid or the biceps, it would be plausible to suggest that it was the proximal joint
agonist that fired first providing the initial displacement of the brachium. As a result, it
likely possessed an assisting role in the task to guide the proximal limb to the target. In
that case, the biceps, as the principle mover. would be inclined to demonstrate no
particular change in activity, according to the hypothesis, while the EMG activity of the
anterior deltoid or auxiliary muscle, would decrease. The reverse was actually m e ; it was
the biceps, which decreased in activity, as the anterior deltoid experienced no significant
change. It would appear that the unrestrained elbow in the task precluded a clear
delineation of which muscle possessed the prime mover role. Indeed, in the investigation
by Hobart et al., above, a forearm splint restricting movernent to the shoulder, which
clearly exposed the anterior deltoid as the prime mover, also demonstrated a decrease in
totaI electncal output of the latter muscle. The pectoralis, another agonist, which actually
played a supporting role, displayed no change in its activity.
Additional research by Hobart, Vorro and Dotson (1978) reproduced the method
of the bail-throwing task used by Hobart et al., (1975), to investigate the dynarnics of
motor ski11 acquisition. The results of various kinematic and EMG variables were
analyzed at five separate intervals of a practice period totalling 104 trials of the task (Le.
trials 1-4, 26-29, 5 1-54, 76-79 and 10 1- 104). Statistically significant decreases in
throwing enor, movement time, iimb angle at ball release and in the time to peak velocity
of the limb, were found to occur within the first 25 tosses. Not surprisingly. these ieamed
adaptations were sirnilar in nature to those observed in the aforementioned study since
the same task was employed. In contrat to those reports though, there was no change in
the total integrated activity of either the anterior or postenor deltoid muscles, which had
previously shown a decrease and an increase in total electrical output respectively.
Nevertheless, a decrease in the latency of the posterior deltoid, an antagonist, was still

demonstrated which continued to support the importance of muscle timing (or the
sequencing of muscle activation) in de fining the changes associated with learning.
Vorro and Hobart (1981a-b) extended their research with the ball-tossing task to
two separate distances, 90 and 150 cm and analyzed a total of eight muscles, 6 agonists
and 2 antagonists.The practice of 103 trials was distributed into seven blocks (block 1:
throw 1, block 2: throws 2-3, block 3: 13-15, block 4: 26-29, block 5: 5 1-53, biock 6: 76-
79, block 7: 101-103). Moreover, they employed a regression equation to determine the
relative importance of a nurnber of kinematic and EMG variables, which best described
the acquisition of the motor ski11 at each of the designated distances. The investigators
noted that the greatest decreases in uirowing error also occurred early on in the practice
period, this time afier only the first two training blocks (Le. 15 trials).
The first equation underscored a large initial adjustment in the total electrical
output of both agonists and antagonists to increse the limbs veIocity at ball release and
contribute to its deceleration respectively, within the first three trials of the task. The
second equation, related to the changes between the first and second training blocks,
profferred significant increases in the initial velocity of the limb as well as a decrease (90
cm goup) and an increase (150 cm group) in the limb angle at ball release. Subserving
these aforementioned kinematic variables were pnmary increases in the pre-movernent
times of the agonists including an augmented synergistic activation of these muscles to
propel the limb toward the target. These were followed by a decreased latency of the
antagonists to decelerate the limb. Further modifications in the above variables, fiom
blocks 3 to 7, were considerably smaller than the adaptations reported between the first
two; they retlected only minor alterations in rnyo-temporal sequencing.
Ludwig (1 982) investigated the triceps and biceps muscles in horizontal extension
movements of the non-dominant ann dunng learning of a shuffleboard task by
participants whose aim was to propel a puck at a 4,s cm target zone located 89 cm away.
The elbow angle, which started at 90°, was required to end at an approximate angle of
120" in order for the target to be hit. Average changes in both myo-electric and myo-
temporal characteristics were calculated in blocks of four trials starting from the first
block (trials 1-4) to the second block (trials 8-1 1) etc., for a total of 100 trials in the motor
task. Significant decreases in target error as well as in puck speed (the time elapsed

between puck release and the attainrnent of the target) and movement-time (the time
between onset of movement to puck release), were observed across training blocks.
Ludwig (1952) confirmed the participants' predisposition to a greater use of force
in accomplishing the motor task within the first few trials (i.e. block 1 ) of the leaming
paradigm, as previously seen with Vorro and Hobart (l98la-b). This was outlined by the
elevated values of total integrated and performance integrated (the total iEMG of the
triceps fiom the onset of limb movement to puck release) EMG activity during execution
of the task. Following this initial period, however. the participants learned to distribute
the forces appropriately as they decreased both the time to peak amplitude of the mucsles
and the latency o f the triceps (an agonist) with respect to the onset of movement. As a
result, there were no meaningfül alterations in the magnitude of the triceps or biceps from
block 2 and on.
Engelhorn (1983)' who suggested that the prevailing kinematic features of the
motor tasks used in the preceding studies had an effect of camouflaging the actual
adaptations in the EMG responses proposed a different methodology. He employed
relatively simple tasks whose kinematic patterns would be rninimized with practice. Each
of these entailed an elbow flexion through a 60' angle fiom an initial joint position of
140". One task consisted of stopping at a point of coincidence with a separate stimulus
visible on an oscilloscope (i.e. a positioning task) while the other involved moving
through the coincidence point within a specified time (coincidence task). Both tasks were
executed at 40" and 200" per second. The EMG data were integrated over six time
periods, two before and four following the onset of movement, in 64 msec bins.
Significant reductions in both movernent time and angular displacernent errors
were found arnong participants after 120 trials of each of the motor skills. An analysis of
the discrete periods of both of the muscle activity patterns revealed that antagonistic
activity in the triceps increased in those time periods wherein adaptive decreases in the
agonist activity of the biceps did not occur, for both the slow and fast movements.
Therefore an augmented CO-contraction in the specific phases of the motor responses was
found to characterize learning.
Contrary to the reports already discussed, McGrain (1986) found no significant
alterations in either the total movement tirne or the myo-temporal variables of a pair of

agonist muscles selected for the analysis of two different strategies of a novel rnotor task.
Seated in a height-adjustable chair, participants used a knee extension rnovement
including the right foot to propel a four-wheel carriage (2,5 kgs) through a timing gate,
located 8 feet away, at 5 mph. Each of two expenmental groups was appropriated either a
back-swing or no back-swing condition of movernent. Although there were changes
between groups, with the back-swing group possessing greater movement times and
increased latencies of the vastus medialis and lateralis muscles by virtue of the reversa1
involved in the task, there were no differences in any of the variables across twenty
practice t d s within each group. These results provided evidence of an experimental task
that was easily acquired without changes in total movement time or timing of the musdes
involved in the activity. According to the investigator, the participants were able to begin
practice of the movements with a motor program, which already met the fundamental
myo-temporal requirements of the task. It was suggested that some other variable such as
the amplitude of EMG activity was manipulated to achieve the criterion velocity of the
motor task.
It is quite evident however, as shown by Gottlieb et al., (1988) that an entire range
of movement compiexity c m elicit significant improvements in critenon learning
variables, including the most seemingly farniliar single-joint tasks. For instance.
participants in the aforementioned study practised both fast and accurate eIbow flexions
of 54" to a target 3" wide within ten separate experimental sessions, conducted on
following days, at 200 trials per session. Researchers observed more than simple
réductions in movement time and increases in the total integrated activity of the biceps
and triceps muscles responsible for accelerating and decelerating the forearm
respectively. They also found that training at one distance increased the propensity with
which they could perform both speedily and accurately at other distances (e-g. a t 36" and
72" movements).
Maring (1990) also found that the mental rehearsai of a simple ball tossing routine
was effective in increasing the rate at which the task was acquired. According to the
elbow flexion movement involved in propelling a ping-pong ball to a horizontal 'bull's
eye' target at a distance of 150 cm, members of an expenmental group were directed to
visualize and 'feel' the movement before they actually performed 10 testing trials.

Following five separate periods of the aforementioned procedure, they had reduced their
tossing error to a significantly greater extent at each stage of the learning schedule as
compared to a control group that did not rehearse. Furthemore, in contrat to the control
group who demonstrated no change in selected myo-temporal variables, expenmen t al
participants had decreased the time to peak activity in both agonist (biceps brachii) and
antagonist (long and lateral heads of the triceps brachii) muscles. An increased latency
between the opposing muscles also chracterized learning that was enhanced by mental
practice.
More recent research by Corcos et al., (1993) extended the work of Gottlieb et al..
(1988) by investigating both within and between session adaptations of 1400 targeted
elbow flexion movements (identical to those described in the latter study) over seven
expenmental periods of 200 trials each. With one day intervening behveen sessions. the
investigators could compare changes in myo-electnc and mechanical variables as a
function of practice (at the end of the B n t session) and learning (across the remaining
sessions). Significant increases in peak velocity, peak acceleration and deceleration
profiles, as well as decreases in the terminal position errors of the movement were found
within the first session. These changes were smaller but similar to those observed and
augmented across sessions. EMG adaptations to the learning paradigrn were consistent
with increases in the intensity of agonist muscle activation in response to an increase in
movernent speed. However, the activation of the antagonist was contingent upon
individual performances as some participants demonstrated a decreased latency of the
muscle (with respect to the onset of the agonist) with learning while others exhibited an
opposite effect. In one particular case, the learner activated the antagonist earlier over
bIocks of the first session and later, across sessions on succeeding days.
Another salient feature of the experiment was the demonstration of retention in
the performance enhancement changes observed with practice fiom one experimental
session to the next. Specifically, performance in the first block of trials conceming the
second session had begun close to but just below the peak attained in the preceding
session and above the level to which performance had declined at the completion of
practice on that first day. Therefore, it was the contention of the authors that the
irnprovements in motor behavior seen with practice, the retention of those improvements

across sessions of the practice paradigm and the reduction in the variability of the
participants responses, which unequivocally defined learning.
A final testament to the above view, on what constitutes learning, was revealed in
the data of the transfer expenments of Jaric et al., (1993). These testing penods were
incorporated into the learning schedule of Corcos et al., (1993), the results of which
expanded on the premise of generalizing the learning at one distance to enhanced
performance at several distances, originally presented in Gottlieb et al., (1988). Pre-test
trials consisted of 11 movements at 18", 34", 54", 72" and 90°, followed by the 1400
trials of the practice schedule and a post-test condition that was identicai to the pte-test
session. Of the five participants studied, three exhibited positive transfer at al1 distances
whereas a fourth only demonstrated transfer to the two longer distances (72" and 90").
Complex Movements
The appearance of a triphasic pattern in fast complex arm movements has been
observed for antagonistic muscles pain at both the shoulder and elbow joints (Wadman.
Van der Gon & Derkson, 1980; Smeets, Erkelens & Van der Gon, 1990). Wadman et al.,
( 1980) found that the triple-burst pattem characterized the activities of opposing muscles
involved at an articulation which exhibited most of the action in accelerated reachinç
movements. When present in both pairs of muscles at each of the joints. the patterns
developed more or less synchronously. Smeets et al., (1990) showed similar results with
flexion movements in which the moments (torques) occumng at the shoulder and elbow
joints of a force in the direction of motion were equivalent. The €MG patterns
demonstrated approximately the sarne timing and intensity for the reciprocal pain of
shoulder and elbow muscles studied.
Altematively, in the execution of fast straight lines and geometric figures with an
electnc pen on an interactive tablet, Accornero et al., (1984) observed a biphasic EMG
pattern in antagonistic muscles of the shoulder and elbow. Although, at times. the
electrical activity in the agonist muscles presented two bursts, the researchers suggested
that a basic building block of one volley in the agonist and one in the antagonist
subserved the various patterns of two-joint ballistic a m movements. These results were
corroborated in an ensuing study by Berardelli et al., (1986) for normal participants
1 O4

perfoming the same drawings of geometric figures in comparison to patients with
Parkinson's disease. The latter group tended to show compensatory multiple bursts in the
agonist for the same actions.
In order to investigate the effects of leaming on both the timing and sequence of
muscle activations underlying a bi-articular motor task, Normand et al., (1982) employed
an experimental protocol of eight separate training sessions, at 100 trials per session, of a
maximum speed horizontal arm adduction, forearm flexion movement. A control group
participated in the pre-test and post-test sessions of the experimental penod each of
which consisted o f five trials. Learners succeeded in reducing their total movement time
for the task as weIl as the movernent times of each of the proximal and distal segment
motions, evaluated separately, in comparison to control group members who were only
found to decrease the time of elbow flexion. Similar to the control group however.
participants trained in the task did not dispiay modifications in the duration o f any of the
agonist or antagonist bursts studied, prompting the researchers to intimate a pre-
programming of the task's myo-temporal features. Instead the individuals learned to
increase the latency between the antagonistic muscles at each joint to allow for the
unimpeded progress o f each of the two limbs during the motor task, for maximum speed.
In a rnotor learning study by Vardaxis (1996), to which the current research is an
addition, a four-day training period of 60 trials per day was utilized to exploit both within
and between session adaptations in the kinematic and myo-eIectric characteristics of two
tasks. They included a uni-directional whipping movement, also used in the present study
(simple task), as well as a bi-directional movement that entailed a reversal at the target of
the latter task, back to the original starting position (complex task).
Significant decreases in performance time were observed for both tasks, within
and between sessions of the experimental protocol commensurate with srnoother and
more consistent end-point, curvilinear, trajectories. Increases in the shoulder and elbow
joint angular velocities were also seen across sessions along with a compensatory
synchronization between the two joints in the complex task. Both amplitude and phasic
modulations in EMG activity were found in the first and second to third wavefoms of the
SVD analysis respectively. They defined a greater activation of agonist and antagonist
muscies as well as an incrrase in the synergistic activation within muscle groups

concomitant with a decrease in antagonistic CO-contraction to enhance the control of both
elbow and shoulder joints, for the two tasks. Furthemore, a partial retention of the
adaptations acquired with motor task practice was also noted for each of the motor skills,
from one expenmental session to the next, which reinforced the efficacy o f the learning
protocol.
In addition to the works presented above, there are numerous investigations
pertaining to the EMG activity associated with multi-joint arm movements primarily
involving reaching andior pointing tasks. Some of these have studied the relationships
between the magnitude and timing characteristics of neuronal activity in the motor cortex
and muscle activations of arm movements in a two-dimensional work space in monkeys
(Caminiti. Johnson & Urbano, 1990; Turner, Owens & Anderson, 1995). Other
researchers investigated path constraints and load perturbations on EMG activity in both
point to point and targeted human arm movements (Lacquanti, Soechting & Terzuolo,
1986; Soechting 1988). Recent work has included a delineation of the basic features
inherent in the EMG pattern for two-joint reaching movements and dynamic isometric
forces in various directions (Flanders, 199 1 ; Flanders & Hermann, 1992; Flanders.
Pellegrin & Geisler, 1996; Pellegrin & Flanders, 1996). Still other studies have sought to
promulgate certain rules or strategies of the CNS conceming the initiation, timing and
magnitude of EMG activity for planar two-joint arm movements (Karst & Hasan. 199 1 a-
b; Gabriel. 1995).
-4tthough al1 of the above studies are revealing in their inquiries into the
phenomenological neurobiology of complex movements, which are central towards an
understanding of how the CNS programs human motor behavior, the motor tasks
involved are fundamentally different from the whipping rnovement being explored in this
investigation. To reiterate, they promote pointing and reaching actions which create more
straight line paths of movement as opposed to the curved trajectory inherent in the motor
task of the present study. Also, a maximum speed condition, employed to elicit the
greatest possible learning adaptations in the currently investigated movement, was not a
necessary circumstance in the tasks of the above research.

Motor Ski11 Retention
The phenornenon of motor ski11 retention has been a focus of inquiry among
investigators across various scientific disciplines. Early studies in ~xperimenral
psychology, which provide an historical perspective on the topic, ernployed pursuit rotor
skills (Bell, 1950; Jahnke, 1958) and other compensatory tracking tasks (Arnmons et al..
1958; Fleishman & Parker, 1962; Harnrnerton, 1963) to assess the effects of interpolated
pauses. or rest periods of no-practice, on motor ski11 training and/or acquisition.
Bell (1 950) utilized a relatively brief practice paradigm of 20 one-minute trials on
a simple pursuit rotor task. Compared to the participants' level of learning on their last
pre-pause test trial, he found that within the first training trial of a retention test one year
Iater. they had only experienced a 29% decrease in their performance scores. Moreover.
any decrements in their motor responses were completely recovered at the end of eight
post-test trials of the motor task.
Jahnke (1958) trained his volunteers at 1, 2.5, 5 or 10 minutes on a similar rotary
pursuit task and re-evaluated their performance at 10 min., 1 day or 1 week post-training.
He uncovered a positive correlation between the initial post-test results and the arnount of
practice received but no statistically determinant changes with respect to the imposed rest
periods.
Arnmons et al., (1 958) investigated participants in a more complex 'Air plane
control test' whose aim was to adjust the orientation of a focal object (a mode1 plane) to a
straight 1 ine position, against various perturbing movements of the latter, using both hand
(control stick) and foot (pedals) controls. Individuals trained for 1, 8 or 40 hours in the
task and were re-tested at 1 day, lmonth, 6 months, 1 year and 2 years later. Larger
decrements in performance were revealed with increasing no-practice intervals, while
those learners who had practised less, experienced a proportionately greater decrease in
task proficiency. Re-learning, which was also studied, required more practice time the
longer the pause without training and the greater the arnount of practice before the
imposed 'rest' interval in question.
Fleishman and Parker (1962) presented differing results on a similar 'aircrafi
control' type tracking task, involving a control stick and rudder pedals to mediate the

fluctations of a target dot and centralize its position on a cathode ray oscillograph. By
increasing the initial training penod to 17 sessions of 21 one-minute trials per session,
distributed over six weeks of pract ice, the experimenters observed nominal performance
decrements in no-practice intervals of up to 24 months. Any srnall losses which did occur
were quickly recuperated within the first few minutes of retraining. Thus the level of
proficiency in the motor skill was observed to be highly dependent on the amount of an
individuals initial training in the task.
Hammerton (1963) debated the difficulty level of the velocity and positional
controls in the compensatory tracking tasks used by previous researchers to describe their
findings, as well as the degree of leaming afforded the participants, which he quatified as
overlearning. To this end, he used a more challenging second-order (acceleration) control
task and two groups differentiated by a level of overleaming in one of them. The
esperirnental task entailed moving a target dot on a cathode ray tube to a target Iine 22,5
mm away and keeping it within a 1,s-mm zone on either side of the line for 2 seconds.
Acceleration of the dot was proportional to the deflection of a thumb joystick. Volunteers
in the regular practice group trained from 40 to 110 trials in the task. while the
overleaming group practised from 90 to 170 trials. Each of these groups was re-tested at
26 weeks post-training. The results indicate decrements in performance as a result of the
no-practice interval, irrespective of the l e d n g schedule but that overlearning did
improve recall of the task.
To clarify the issue of overlearning and its effects on motor ski11 retention,
Melnick (1971) studied groups of individuals who practised a balancing ski11 on a
stabilometer and then received 0, 50, 100 or 200% of extra practice following the
Iearning criterion. At both 1 week and 1 month post-learning, immediate absolute recall
of the gross motor skill was significantly better in al1 of the overlearning conditions as
compared to the performances of participants who did not receive estra practice.
However, when the latter group were given tirne to relearn the task, their scores were
found to differ only with the 200% overlearning group and only at the one-month
retention interval.
In erercise physioIogy research, the terni 'detraining' is used to refer to the
transient and reversible nature of training-induced adaptations (McArdle. 1995).

Attempts to analyze changes. using EMG, in neuromuscular performance within strength
training/detraining paradigms have focussed on both unilateral and bilateral leg
movements and the abstention of the knee extensor/flexor muscles from a strength
training stimulus (Hakkinen & Komi, 1983; Hakkinen, Alen & Komi, 1985; Narici et al..
1989; Hortobagyi et al., 1993). Depending on an individual's distribution of motor units
within a muscle (or muscles) and his or her capability of controlling the gradation of
muscular force through such mechanisms as recruitment ancUor rate coding, for instance,
i t is possible to think of maximum strength as a motor ski11 (Vandervoort, 1992).
Preliminary research by Hakkinen and Komi (1983) included participants
accustomed to weighi training who followed a progressive resistance strength-training
program of 16-weeks duration. It combined both concentric (75% of contractions studied.
at 80-100°/b of one maximum concentric repetition. 1-6 reps per set) and eccentric (25%
of contractions, at 100-120% of one maximum concentric repetition, 1-2 reps per set)
contractions using a dynamic squat lifi exercise. An eight week detraining penod ensued
wherein al1 strength training was teminated. A significant increase in the maximal
bilateral isornetric force of the quadriceps was found post-training, concomitant with a
similar augmentation in the averaged maximum integrated EMG (IEMG) activity of the
leg extensor muscles (Le. rectus femoris, vastus lateralis and medialis), up to 12 weeks of
the training regimen. Both the isometric force and IEMG measures were observed to
decrease after detraining but not to pre-training levels. The authors attributed the decline
in neuromuscular performance to an absence of the strength training stimulus and
categorized the detraining effect as 'comparable to the opposite effects of training'.
In a secondary investigation, Hakkinen and Komi (1985) extended both the
training and detraining penods to 24 and 12 weeks respectively, along with increasing the
nurnber of repetitions for each of the concentnc (1-10) and eccentnc (3-5) contraction
exercises, described above for the sarne squatting exercise. Similar decrements in the
averaged maximum IEMG values of the quadriceps muscle group, frorn post-training
leveIs, were observed after detraining which correlated significantly with the decrease in
maximal isometric force for the sarne no-training penod. However, an improvement in
the maximal rate of isometric force production, particularly at the higher force levels
analyzed (e-g. 3000 N), was found to persist or even be further enhanced dunng the

detraining period. These results led the researchers to suggest an ongoing adaptation of
the motor unit recruitment pattern subserving fast force production in experimental
participants, despite a termination of the training program.
Experiments by Narici et al., (1989) incorporated isokinetic knee extension
training (at an angular velocity of 2.09 radlsec) of the dominant leg in participants. who
were exercised for six series of ten maximal contractions, four times a week for fifieen
weeks; they subsequently detrained for the following ten weeks. The results at the end of
training revealed an increase in the cross sectional area of the quadriceps that was only
approximately iess than half of the rise in the maximal isometric force production (MVC)
of the trained leg. The changes in force output in hm, were only about half of the
increase observed in the IEMG activity of the vastus lateralis indicating that the
enhancement in MVC with training may have been influenced by a greater neural
activation as opposed to an increase in muscle hypertrophy. Moreover, the authors
suggested that increases in both IEMG and MVC measures for the untrained kg,
although not statistically significant, alluded to a supraspinal modulation of the neural
drive to the muscles. The kinetics of the changes seen in each of the above-mentioned
variables with leaming, were observed to have an equal time-course during detraining.
Ishida, Moritani and Ito, (1990) studied the relationship between modifications in
maximal voluntary strength and selected twitch contraction parameters evoked by
electrical stimulation, in an attempt to elucidate the central andlor peripheral factors
mediating the adaptations during both training and detraining. Volunteers were trained in
a heeI raising exercise whose program consisted of three sets of 15 repetitions per set
(using 70% of the maximal load on day 1) three times a week for eight weeks. They
demonstrated a significant rise in the MVC of the gastrocnemius muscle with training, in
comparison to no particular changes in the maximal twitch torque or maximal rate of
torque development in the triceps surae muscle. In contrast, eight weeks of detraining.
which resulted in no apparent decrease in MVC revealed a significant increase in the
maximum value conceming the rate of torque production. The investigators speculated
that central factors contributing to the augmentations in the maximal strength of trained
muscles might facilitate a potential for complex post-detraining adaptations in the same
muscles afler training has ceased.

Staron et al., (1991) showed similar results of a propensity for individuais to
retain strength-induced adaptations during a detraining period but what made their
findings unique was the fact that the no-training interval was extended to 32 weeks.
Specifically, women who had subscribed to a 20-week progressive resistance strength
training program, including dynamic squat, leg press and leg extension exercises.
continued to demonstrate maximal performances for the squat at eight months post-
detraining that had not significantly diminished fiom post-training measures. The values
for the other two exercises exhibited statistically defined decreases at the end of
detraining but not to pre-training levels. To assist in the evaluation of these data the
researchers noted a sustained hypertrophy of the myofibers within muscle biopsies of the
women's vastus lateralis and also suggested a retention of neural adaptations as a resuIt
of training. According to the authors, it was the acquired reserve of each of the
aforernentioned factors that decidedly aided the previously trained women in additional
re-training experiments (Le. 6 weeks), versus a second group of untrained women.
achieve maximal strength increases in each of the exercises within a comparatively
shorter time.
Hortobagyi et al., (1993) further underscored the importance of an individual's
training status as a determinant factor in the assessrnent of detraining effects on acquired
muscu1a.r strength levels, particularly among power athletes who are characterized by a
long history of strength training. To this end, the investigators studied a group of strength
ath letes, composed of power li fiers and football players, homogeneous in anthropometric
characteristics and who possessed similar weight lifting performances and workout
schedules. They were evaluated on a series of maximal strength measures including both
isometric and isokinetic knee extension/flexion forces, as well as one repetition
maximum testing of free-weight bench press and squat exercises both before and after a
14 day detraining period. An analysis of pre- and post-detraining measures revealed
statistically significant decreases in only maximal eccentric knee extension strength
performed at angular velocities of 0.87, 2.62 and 4.37 radsec. An EMG analysis of the
vastus lateralis during each of the aforementioned testing conditions could not explain the
decrements in eccentric strength since there were statistically insigni ficant reductions in
the muscle's activity level, suggesting maintenance of the neural drive to the latter.

Instead, the investigators indicated a probable decrease in the hypertrophy of fast twitch
muscle fibers associated with eccentric strength gains. Thus when the initial training level
of an athlete is very hi& short-term detraining periods do not necessarily have any
deleterious effects on the specific adaptations in maximal concentric forces developed
with long-term strength training.
The consequences of reduced training periods on neuromuscular performance
have also been evaluated for both previously trained and untrained individuals. Research
conducted by Graves et al., (1988) involved two groups of participants (previously
untrained for at teast a year) who had been undergoing variabte resistance, bilateral knee
extension strength training programs for penods of ten and eighteen weeks. Individuals
who were trained two days a week and three days a week in both of the training regimes
experienced significant increases in maximal bilateral isometric strength as well as in the
amount of resistance used during training. All of these people were subsequently engaged
in twelve weeks of reduced training fiequency. Those who had trained at 2 days per week
were randomly assigned to both a 1 day a week group and a detraining group while
rnembers who exercised for 3 days a week were relegated to a 2 days per week group and
a 1 day a week group. The findings demonstrated no statistically significant reductions in
either of the above mentioned strength measures for those groups in which training had
been reduced to 2 days a week and 1 day per week. The detraining group, on the other
hand. exhibited a decrease of more than half of their original post-training isometric
strength.
Hakkinen et al., (1991) investigated reduced training in ten strength athletes
composed of power lifters and body builders, each of whom possessed a systematic
strength training background of 5 to 10 years. Al1 of the athletes participated in a three-
week experimental period incorporating a two-week 'regular* heavy resistance strength
training program for the leg extensor muscles followed by a one week period in which
the overalI volume of training was reduced by 50%. The regular program consisted of a
squat-lifi exercise of 18-22 contractions per session at 70%-100% of one maximum
repetition (1RM) and a leg press or knee extension exercise of 20 to 40 contractions per
session at 60%-80% of IRM, every second day. The results showed no changes arnong
the athIetes in bilateral MVC dunng leg extension, maximal force per cross sectional area

(force/CSA) of the quadriceps (each of which were found to be comelated), and maximal
averaged IEMG of the vastus lateralis, medialis and rectus femoris, before and afier the
three-week period. The researchers found, however, that they could divide the group into
five 'best' athletes and fwe 'other' athletes based on a person's value of maximal
force/CSA of the quadriceps. This distinction between the two groups revealed that the
'best' athletes exhibited significant increases in both bilateral MVC and averaged IEMG
during the reduced training period as opposed to the 'others' who did not. The
investigators concluded that in the case of highly trained strength athletes, who may have
at~ained their maximum in neuromuscular performance. fùrther improvements in
maximal strength ancUor neural activation may be accrued not fiom 'normal' training
periods but rather fiom a short penod of 'reduced' training.
As was presented within section 1.2 of the introduction, a separate experimental
mode1 based on the spinal stretch r e m (SSR) has aided in defining the underlying
processes andlor substrates accounting for the acquisition and sustenance of a simple
motor ski Il (Wolpaw, 1994). The learning paradigm involved training monkeys to
maintain a constant elbow angle (90°) against an extension torque for a pre-specified time
interval. If the averaged EMG activity of the focal biceps was found to be in a certain
range. a small additional extension torque would be provided to extend the eIbow and
elicit the biceps SSR. Liquid reward was given to the monkeys 200 msec aller the extra
torque. The animals were trained under one of three different modes, a control mode, in
which the reward always followed the additional torque, established the base-line
amplitude of the response. Converseiy, under each of an SSR-up or SSR-down mode.
reward was only given if EMG activity was greater (SSR-up) or less (SSR-down) than
the control value.
Leaming in either of the two experimental modes, characterized by 3,000 to
6,000 trials per day over a period of 2 to 17 months, elicited a different learning cuwe
than that seen in traditional motor learning paradigms. An immediate change of
approximately 8% in the specific direction of a mode was observed on the first day of
training, which preceded that modes gradua1 development over weeks, in which 80%-
90% of the change actually occurred (Wolpaw, Braitman & Segal, 1983; Wolpaw &
O'Keefe, 1984). lnvestigators suggested that the initial phase development process

entailed an operant conditioning of one or more descending spinal cord pathways acting
on the reflex arc mediating the SSR and that the chronic daily presence of this activity
produced a persistent structural and/or biochemical alteration in the spina1 cord. The fact
that these changes were observed to persist over pauses in training of 10-38 days
supported the notion of an adaptive plasticity in the spinal cord (Wolpaw et al.. 1986).
Further studies on the reversal and redevelopment of SSR change, demonstrated
similar incremental evolutions in the face of opposite mode imposition followed by
reimposition of the original mode (each of which also survived non-performance breaks)
that reinforced the hypothesis of a persistent segmental alteration (Wolpaw, 1983).
Moreover, the predominance of the SSR phenornenon in the focal agonist (biceps) as
compared to other synergistic (brachialis and brachioradialis) and antagonist (triceps)
muscles underscored the feasibility of the SSR system as a substrate for the study of
memory in primates.
Recent motor control research, focussing on the acquisition and retention of
motor skills has investigated complex tasks as well as visual imaging of the brain itself.
in order to elucidate the resulting CNS adaptations to each of the preceding phenornena.
Kami et al., (1995) noted both substantial improvements and significant retention arnong
participants in the learning of a motor task that required the rapid tapping of accurate
finger sequences in opposition to the thumb of the non-dominant hand. A training
schedule incorporating 10 to 20 minutes of practice per day for five weeks resulted in a
performance index that was two times the speed of accurate sequencing exhibited at the
beginning of practice. The performance enhancement displayed a fast early rising phase
within the first w-eek that was followed by a more gradua1 evolution of acquisition to
asymptote by week four. The task showed linle transfer to the other hand and did not
generalize to the execution of a different control sequence of equivalent component
movements. The researchers did find however, that the MW of local blood oxygenation
level-dependent (BOLD) signals elicited in the primary motor cortex (M 1 ) demonstrated
a consistently larger activation area for the trained sequence in cornparison to the control
sequence. The tirne course in the development of the size differences between the trained
and the control sequences of the Ml response corresponded with the maximal asymptote
performance of the trained sequence, suggesting that leaming is charactenzed by a new

and more extensive representation in the motor cortex. Additional retention experiments
reinforced a long lasting effect of the graduai learning process, as a persistence of
superior performance in the task and in the evoked M l response was observed after 10
and 2 1 weeks of no extra training.
In their study, Brashers-Krug et al., (1996) investigated the underiying
consolidation process of an experimental task in motor memory. They trained a total of
seventy individuals in the use of robotic manipulandum to make 10 cm rectilinear
reaching rnovements fiom a central position to eight different target locations, at a
moderate Pace (i-e. 500 msec). An initial target set consisting o f a senes of 192
movements was first used to establish baseline motor response patterns before the
introduction of a second set in which the manipulandum provided perturbing forces in a
clockwise direction. The participants were required to learn to compensate for these
forces in an effort to continue to guide their rnovements to the targets (task 1). The
performance index of learning was the degree to which the participants could adjust to
the extra forces in reproducing their baseline trajectories
The investigators found that a control group tested 24 hours after the learning of
task 1, exhibited a significant irnprovement in their performance with respect to post-
training values. An experimental group, which received training in a different pattern of
forces (conter clockwise) immediately after the learning of task 1, demonstrated a
reduced learning capacity or negative transfer in the second task. When tested a day iater
for retention of the first task, the performance level of the experimental group was
comparable to the index of learning observed after training but it did not show any
additional improvement, as had been noted for the control group. Therefore the extra
training in task 2 apparently compromised the retention of the first task (retrograde
interference). Intervening with the second task at 5 minutes, 1 hour and four hours post-
training, in three other separate groups revealed that only the 4-hour group could display
a statistically significant increase in the retention of task 1. However, both negative
transfer and retrograde interference were seen to decrease monotonically with increasing
no-training intervals. According to the researchers, there would thus exist a small time
time-window in hurnan motor memory wherein a more fragile representation of memory
is stored before it is transformed or consolidated into a more solid state.

In a follow-up study, Shadmer and Brashers-hg (1996) trained another group of
volunteers in a longer practice paradigrn of three target sets for each of the two different
tasks described above, that were separated by a 24-hour interval between training
sessions. Significant improvements in perfomance were observed at two and three weeks
beyond the completion of training for tasks one and two respectively while a control
group, which was only trained in first task, continued to show a high level of leaming at 5
months afler the initial practice session. Two separate intemal models of motor action
had therefore been consolidated in long-term motor memory when the temporal distance
between the learning of two independent skills was 24 hrs. The investigators also re-
evaluated the time course in the consolidation of the motor skills based on the newly
protracted practice period (to ensure a learning plateau) and a recall session that was
extended to 1 week post-training instead of 24 hours (employed to nulliS, any effects the
leaming of task 2 might have had on the subsequent recall of task 1 - anterograde
interference). They found a significant recall of the first task at 5,5 hours following the
completion of training in task 2 but that the retention level approached the performance
index of the control group only at 24 hn post-learning.
In yet another study, Shadmer and Holcomb (1997) provided evidence of a
reorganization of the neural representation of human motor memory to support the
îïndings of Brashers-Krug et al., (1996) and Shadrner and Brasher-Kmg, (1996)
regarding the stability of the intemal mode1 of a motor task, aHer only a few hours
following acquisition. The authors procured positron emission tomography (PET) scans
of the brain as participants leamed the task of a novel mechanical system (identical to
that used in the previous studies). Measures of regional central blood flow (rCBF) were
correlated with total motor output over areas of the prefrontal cortex dunng training of
the task. Participants were then monitored a second time at 5,s hours post-learning. a
period that signified consolidation of the motor ski11 in the above research. The
investigators observed new regions where activations correlated significantly with motor
output, the premotor, posterior parietal and cerebeller cortex, concomitant with a
reduction in the preceding correlation for the prefiontal areas.

Appeadix 2
QUESTIONNAIRE
Age :
Height: m Weight: kgs
Arm preference: please pnnt a response of either "Lefi" or "Right" for each of the
questions below, in the spaces provided.
1) Which hand do you employ when writing?
7) M i c h hand do you ernploy when grasping the handle of a racquet (i.e.
badminton, squash, tennis) or tooi such as a harnmer?
3) Which side do you use when holding and stnking with a baseball bat?
4) Which hand do you use when throwing a baseball or softball?
5) Which hand is used when pitching underarm in softball?
6) Which hand is employed when striking a punching bag for power?

George Sarantinos .M.A. Candidate McGill University Department of Physical Education 475 Pine Ave. West Montreal, Quebec, H2 W l S4
Appendix 3
Dear:
Your interest regarding participation in this study is very welcomed and much appreciated. The purpose of this experiment is to inquire into the electromyographic changes in and between selected muscles of the lefl upper limb and chest following both training and afier periods of no practice.
There will be a total of nine testing sessions to occur in the exercise physiology laboratory of McGi11 University's Seagram's Sport Science Centre. The first of these sessions will entail the measurement of your height and weight, and include the completion of a questionnaire to determine right arm/hand preference. For your own comfort and to facilitate the testing protocol, you will be asked to dress in shortdlight pants and a t-shirt.
Prior to actual data collection, you will be placed in the chair of an experimental table using velcro straps to lirnit movement of the waist and upper torso. A plastic apparatus will be secured to the top of your forearm using velcro straps once again, as well as athletic tape. Six adhesive surface EMG electrodes will be placed on six sites of your left upper limb and chest. These are the back of each of the shoulder and upper a m , the left side of the chest below the collar bone, the front of the upper arm and above the forearm. The EMG electrodes will provide information concerning the attendant eIectricaI activity of each of the muscles under study during both training and no-practice. In order to optimize conductivity at the electrodelskin interface sites, shaving of local surface hair will be required. Furthemore, each of the six sites will be outlined using a non-toxic permanent ink marker to ensure the sarne electrode placement for each testing session.
Subsequently, you will be asked to perfonn (3) three sets of twenty-five (25) trials of an unrestrained movement using the left ami in the horizontal plane. The task will need to be performed as quickly as possible, beginning from a 'home' and terminating at an end 'target' position. You will repeat the protocol of the motor task for four (4) consecutive days (including the first) and then return to the lab for one set of ten trials of the task one day afier the training period and at 1, 2, 4, 6, and 8 weeks post-training. During this time you will be requested to refrain tiom any type of strength training involving the upper body as this may influence your testing performance.
By the end of the training period you may experience some soreness of the biceps muscle due to the nature of the task. The use of EMG electrodes to record the electrical activity of the six muscles assumes no nsk.
118

You can be assured that a11 persona1 and data coIIected will be held in the strictest of confidence by the experimenter. Your files will be numerically encoded so as not to reveal your tme identity at any time; neither during the testing sessions nor in the final paper.
Via this experiment significant knowledge will be gained conceming neuromuscular hc t ioning when sufficiently trained and pnmarily how abstaining fiom practice may affect either an athlete's or even a non-athIete's neuromuscular performance. If you decide to proceed as a participant, you will have the unwaivering option of withdrawing fiom this investigation at any time and request to have al1 persona1 information and data destroyed. No reasons need be given, no questions will be asked.
1 thank you and welcome any inquiries you rnay have.
Sincerely,
George Sarantinos, M A . Candidate

Appendix 4
Informed Consent Form
1 , g a n t permission to George Sarantinos
to proceed with:
1. The measurement o f Anthropometric components (Height and Weight).
7 -. The placement o f surface adhesive EMG electrodes to record the myoelectric activity of six (6) muscles of the left upper limb and chest.
3 . The collection o f 450 trials of data of a 'fast' two-joint motor task, during both training and retention conditions for the purpose of investigating the "Learning and Retention Adaptations of Myoelectric Activity During a Novel Multi-Joint Task".
1 have read and clearly understood an explanation o f the nature, procedures, purposes, risks and benefits of the proposed research in which 1 will be participating. 1 am fully aware that this research project is investigational, that 1 am at liberty to withdraw from the study at my own discretion, at any time, by simply asking to do so and request to have al1 persona1 information and data files destroyed. With this in mind and having clarïfied any further persona1 inquiries with the investigator, I voluntarily consent to the methods/procedures enumerated above.
Signature:
Witness :

CERTlFlCATE OF ETHICAL ACCEP INVOtVlNG HUMAN
tABtLlW FOR RESEARCH SUBJECTS
A review cornmittee consistirtg of thma cf the follcwing membem:
1. P d . E. t u a thaue . 1. Prof. M. Maguire
2 . Prof. R. Ghorh 2. Prof. N. Jeckson
3. Frof. M. Downey 3. Prof. H. Pemault
has examined the application for certification of the etkical acceptabilay of the projec! titled:
The Ef fecrs of Detraining au Neuromscular I h i l t i - joint Coordiaaciou. -
The review cornmittee considers the researcn procedures. as explained by the appllcant in this application, to be acceptable an ethical grounds.
L- (Sig ned)

Appendix 6
ANOVA table for Performance Time data - Experimental group
Source Type of test d. f. F P
Learning Within-Subjects Contrasts (1,9) 29.3 1 0.0001
Retention Within-Subjects Conri-asts
Day 5 vs. Ret 1 (1 ,9) 0.22 0.639 Ret 1 vs. Ret 2 (1 ,9) 10.35 0.01 1 Ret 2 vs. Ret 3 ( L 9 ) 0.53 0.387 Ret 3 vs. Ret 4 ( 1-9) 0.68 0.432 Ret 4 vs. Ret 5 ( 1-9) 0.67 0.433

Appendix 7
ANOVA table for the PerCormance Time differences between the Experimental and Control groups
Source Type of test d. f. F P
Learn-Ret Within-Subjects Effects (2,26) 13.416 0.0001
Learn-Ret * Group Within-Subjects Effects ( 2 3 ) 7.252 0.003
Learn-Ret 1 Within-Subjects (Day 1 - Day 5) Contrasts (1,13) 18.195 0.001
Learn-Ret 2 Within-Subjects (Day 5 - Ret 5 ) Contrasts (1,131 8.625 0.01 2
Learn-Ret l*
Group Within-Subjects (Day 1 - Day 5) Contrats W 3 ) 5.749 0.032
Learn-Ret 2* Group Within-Subjects (Day 5 - Ret 5) Contrasts ( 1 , 13) 8.758 0.01 1
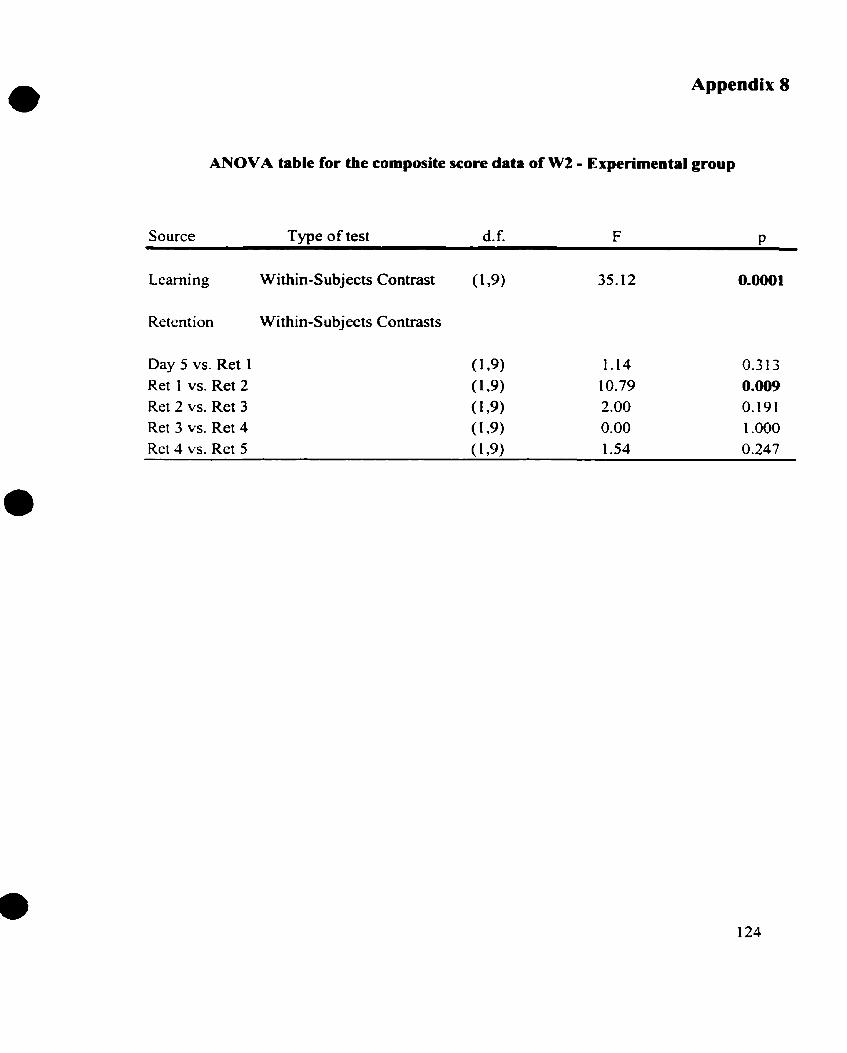
Appendix 8
ANOVA table for the composite score data of W2 - Experimental group
Source Type of test d. f. F P
Leaming Within-Subjects Contrast (1,9) 35-12 0.0001
Retention Within-Subjects Contrasts
Day 5 vs. Ret 1 Ret 1 vs. Ret 2 Ret 2 vs. Ret 3 Ret 3 vs. Ret 4 Ret 3 vs. Ret 5

Appendix 9
ANOVA table for the Composite Score differences of W2 between the Experimental and Control groups
Source Type of test d. f. F P
Learn-Ret Within-Subjects Effects (2-26) 9.39 0.00 1
Leam-Ret * Group Within-Subjects Effects (2-26) 4.289 0.025
Lem-Ret 1 Within-Subjects (Day 1 - Day 5) Contrasts (1,13) 12.165 0.004
Learn-Ret 2 Within-Subjects (Day 5 - Ret 5) Contrasts (1,13) 5.476 0.036
Lem-Ret 1 * Croup Within-Subjects (Day 1 - Day 5) Contrasts (1,13) 0.238 0.634
Learn-Ret 2* Group Within-Subjects (Day 5 - Ret 5) Contrasts (1,13) 10.004 0.007

APPENDICES 10 - 14
The following appendices contain the data of the third waveform (W3) of the
SVD analysis for both the experimental and control group members in each of their
leaming and retention conditions. An attempt was made to organize this data in a rnanner
that would highlight the agonist and antagonist muscles central to the execution of the
motor task as revealed by the eigenvector loadings for each of the muscles studied. To
this end, the names of the relevant agonist andior antagonist muscles for a particular
experimental condition are first provided within an appendix table (A) in their
abbreviated form. The purpose is to provide a visual presentation of the interrelationships
of the muscles in each of the extensor and flexor groups involved in the production of the
motor task and how these relationships change across experirnental conditions.
According to the magnitude and polarity of their respective eigenvector loadings
the muscles are listed in order of decreasing magnitude and as either firing sequentially
(e.g. PD vs. LO) or synchronously (e.g. LO + LA). A second appendix table (B) displays
the actual eigenvector loadings or muscle coefficients which categorize the representat ion
of agonist and antagonist muscles seen in the preceding appendix.

Appendix 10A
Dav 1 Dav 5 Partici~ant:
Pl LO vs. PD . . .LA
Br - -. -Tu .
P2 . > .
LA vs. PD LA.+ U) H> Bic . ' 8 ,
., --. . , . . , , . ..*- .. -
P3 LO vs. PD ,?- ,-a :: , - . . . . , :-. . .
Pec P5 LA \-S. PD PD-vsLA+LO
Bic Bt P6 LA "' 'LA vs. .m.
Bic vs.Pec S. - - . - . .- . . .W. - ,
P7 LA vs. PD Y* '" .mmA L1 '
Bic
-
Pl0 PD vs. LA + LO L A vs. PD Bic Pec \-S. Br + Bic
Within muscle group representations of W3 for the leaming penod (Day 1 to Day 5) of the experimental group. P7 is presented in the bold font. The white and grey areas denote a local and global pattern of the waveform respectively. The muscles are presented in order of decreasing magnitude according to their eigenvector coefficients.

Appendix 10B
Day 1 Dav 5
P3
P4
1 -0.46/-0.31 vs. 0.33 1 0.61 . 1
P7
0.66 vs. -0.5 1
-0.4 0.61 vs. -0.51
Eigenvector coeffkients of the within muscle group representations depicted in Appendix 10A, for the learning period @ay 1 to Day 5) o f the expenmental group. The data for P7 are presented in the bold font. The white and grey areas denote a local and global shape of the waveforms for W3.
-0.55 VS. 0.49/037 I . - A S 4
I
-0.71 vs. 0.65
-0.63 vs.0.40 0.56 vs. -0.32
Pl0
+ - _ -0.57 . J
: ,-,-@!W va^ -46f
-0.70 0.57 VS. -0.39/-0-30 1
-0.53 \-S. 0.33/0.30 0.48 vs. -0.39

Appendix 1 1 A
w Ret 1 Ret 2 Participant:
,_ - - z$;+--~?, *:-':y ::.-:-A - : * ? t.JTp+-z: +:+;;-y :- Pec .- . . ,-. . .- , - . . . - ,.., , . . L - . .-.- ". =,< '.
PS .*vt:;Zb-+. u: - 2 ;- ,PD - ; - mI$&:m -+: 7.. ." , * ~0
I: . ABiC I Pec vs. Bic I Pec vs. Bic T
P6
- .- .
P8 PD VS. LA + LO . PD vs:-LO ,+ Lh PD vs. LO * . B a . . .
r . -. 1 BrR + Bic rs. Pec
BrR LA.--, PD
1 Pec vs. BrR + Bic 1 BrR vs. Bic + Pec 1 BER "
Within muscle group representations of W3 for the short-terni motor memory period (Ret 1 and Ret 2) of the experimental group. Day 5 of the learning period is included for comparative purposes. P7 is presented in the bold font. The white and grey areas denote a local and global pattern of the waveform respectively. The muscles are presented in
order of decreasing magnitude according to their eigenvector coefficients. The asterix (*),
t LO vs. TI *
cross (t) and double cross ($) superscnpts highlight a decrement, persistence or
improvement in motot coordination respective1 y.
129
Bk * LO vs. LA

Appendix 11 B
Da? 5 Ret 1
. . -a54 t 0.37 f P4 -0.71 vs. 0.65 0.51 vs. -0.40
p6 1 O=I2+~.4.33 - 1 -0.52 vs. 0.34 *
P9 1 0f4v&o-31 1 -0.55 \-S. 0.50 -0.45 vs. 0.44
Pl0 0.48 vs. -0.39
1 0.57 vs. -0.39/-0.30 ( -0.77 vs. 0.54/0.31
Ret 2
4.70 vs. 0.64
-0.55 vs . 0.31
0.56 vs. -0.37t
Eigenvector coefficients of the within muscle group representations depicted in Appendix 11A, for the short-tenn motor memory period (Ret 1 and Ret 2) of the expenmental group. Day 5 of the learning period is also included for comparative purposes. P7 is presented in the bold font. The white and grey areas denote a local and global pattern
of the waveform respectively. The asterix (*), cross (t) and double cross ($) highlight a
decrement, persistence or improvement in motor coordination respectively.

Appeodix 12A
Ret 2 Ret 3
1 1 P D vs. LO 1 1. y m.-PDD BrR + Bic vs. Pec :, . -*: mw pe: :..- I
PD U v s . PD $ Pec FS. Br Pec
P6
Within muscle group representations of W3 for the first long-term motor memory condition (Ret 3) of the experimental group. Ret 2, of the short-term motor memory period is also included for comparative purposes. P7 is presented in the bold font. The white and grey areas denote a local and global pattern of the wavefoxm respectively. The muscles are presented
in order of decreasing magnitude according to their eigenvector coefficients. The asterix (*),
cross (7) and double cross ($) superscripts highlight a decrement, penistence or improvement in motor coordination respectively.
LO vs. LA
Pec vs. Bic f 1
LA vs. LO ? Pec vs. Bic

Appendix 12B
Ret 2 Ret 3
P6 1 -0.55 vs. 0.31 1 0.12 rs. -0.36 t
P7
P9 1 0.40 * 1 -0.74 rs. 0.40 $
p8
0.56 \-S. -0.37t
-0.78 vs. 0.55 *
Eigenvector coefficients of the within muscle group representations depicted in in Appendix 12A, for the first long-term motor memory condition (Ret 3) of the experimental group. Ret 2, of the short-term motor rnernory period is also included for comparative purposes. f 7 is presented in the bold font. The white and g e y areas denote a local and global
pattern of the waveform respectively. The asterix (*), cross (t) and double cross ($) highlight
a decrement, persistence or improvement in motor coordination respectively.
0.59 vs. -0.53
&6'9 m. -492 +
-0.52 vs. 0.47 * -0.37/-0.31 vs. 0.43
Pl0
a @SS:VS.' i as+- $7 - 447 4-0.38 - -
0.68 vs. -0.47
'0.65 vr. 451Q171A3 -
0.41
-036 vs. 0-37.033

Appeadix 13A
Ret 3 Ret 4 Ret 5 Panici~ant:
1 Pec vs. Bic 1 Bic + BrR vs. Pec
PD vs. LA
- - .BIRs . .-*.,.l I * . . . - - BrR vs. Bic
Pl0
Within muscle group representations of W3 for the second and third long-term motor rnemory conditions (Ret 4 and Ret 5) of the experimental group. Ret 3 is also included for comparative purposes. P7 is presented in the bold font. The white and grey areas denote a local and global pattern of the waveform respectively. The muscles are presented in order of
decreasing magnitude according to their eigenvector coefficients. The asterix (*), cross (P) and double cross ($) superscnpts highlight a decrement, persistence or improvement in motor coordination respectively.
133
Pec
P D v s . U + L O Bic + BA vs. ~ e c
LO $
Bic t PDvs.LO +LA

Appendix 13B
Ret 3 Ret 4 Ret 5 Partici~ant:
4.e~. 038 -- -0.44/-0.40 * -0.58/-031or; a37 - -+ ,
- . - . - P9 -0.74 vs . 0.10 $ -:@4-vr 431- * 0.67 +&&3/-0:~i
. . <
- . -- g r . .
. . - 0.41 rs~;@/ia~~ii;-.. . - ..
r . . , . , . . . . ... pl0 . . - .0a7~0.&! . 0.38 $ - 0;68.vs.-û.54/-0;34
0.59 vs. -0.54
0-9-W. OS, . .* -
Eigenvector coefficients of the within muscle group representations depicted in Appendix 13A, for the second and third long-term motor memory conditions (Ret 4 and Ret 5) of the experimental group. Ret 3 is also included for comparative purposes. The data for P7 is presented in the bold font. The white and grey areas denote a local and global pattern
of the waveform respectively. The asterix (*), cross (t) and double cross ($) highlight a
decrement, persistence or improvement in motor coordination respectively.
-0.48/-0.34 vs. 0.30
. 0œ-W:m. * L
Ofi2 . 3 4.@-~.'0.59

Appendix 14A
Dav 1 Dav 5 Ret 5 Partici~ant:
LA + LO vs. PD
Within muscle group adaptations of W3 for Day 1 , Day 5 and Ret 5 conditions of the control group. The white and grey areas denote a local and global pattern of the
the wavefom respectively. The muscles are presented in order of decreasing magnitude
according to their eigenvector coefficients. The asterix (*), cross (t) and double cross ($)
superscripts highlight a decrement, persistence or improvement in motor coordination respectively.

Appendix 148
Eigenvector coefficients of the within muscle group adaptations depicted in Appendix 14A, for the control group conditions @ay 1, Day 5 and Ret 5). The white and grey areas denote
a local and global pattern of the wavefom respectively. The asterix (*), cross (t) and
double cross ($) highlight a decrement, persistence or improvement in motor coordination respectively.
w w Ret 5 Partici~ant:
Cl
C2
C3
C4
C5
-0.47
0.85
0-63vs--0.37
0.12 vs. -0.37/-0.37
-0.39
0.71 YS. -0.47
0.82 vs. -0.35
-0.3 1 -0.71rs.0.49
0.42
-0.50 T
0.73
-0.64-~~.-0-53,l-0.34~-
. - . ,, . 0.4z . f * ,'.y - - - ' - - % 6. - &%/-O& ys. 0~45:
0.62 f 0.78 m. 4.48
.f - ~
~ ; ~ ~ ' ~ j ~ ~ ~ ; 3 4 ~ ' : ~ n , ~ J ~ ~ ' - .,. . . + :<- -'x-..d ,.
- . - - . .
a65 VS. -0.58/-0.30
0.37
-Q17i.%-&@
0.39 * . i 5 2 rr. 0.42 *
0.66 W. -0.33
-0.70.+ OJl "' : 0$.1 . ,
. .
c- -:,
0-42 *