a serine proteinase in drosophila embryos: yolk localization and developmental activation
TRANSCRIPT

Insect Biochem. Vol. 19, No. 7, pp. 687~91, 1989 0020-1790/89 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1989 Pergamon Press plc
A SERINE PROTEINASE IN DROSOPHILA EMBRYOS: YOLK LOCALIZATION AND DEVELOPMENTAL
ACTIVATION
MIGUEL MEDINA and CARMEN G. VALLEJO*
Instituto de Investigaciones Biom6dicas, CSIC, Facultad de Medicina, UAM, Arzobispo Morcillo, 4, 28029-Madrid, Spain
(Received 21 February 1989; revised and accepted 9 June 1989)
Abstract--A serine proteinase (molecular mass, about 25,000 Da) has been found in Drosophila mature oocytes. (1) The detected activity increases exponentially during embryogenesis. (2) The subcellular localization changes from the yolk granules, in oocytes, to the soluble fraction, in late embryos. (3) The proteinase cleaves by basic residues, but arginine is preferred (about 7-fold) over lysine.
Key Word Index: Drosophila serine proteinase, developmentally regulated proteinase, yolk granules associated proteinase, developmental changes in subeellular localization, preferred cleavage by arginine vs lysine
INTRODUCTION
We have been involved in the study of proteinases that are developmentally regulated (Garesse et al., 1980; Perona and Vallejo, 1982, 1985; Perona et al., 1987, 1988; Ezquieta and Vallejo, 1985, 1986a, b, c, 1987). Our data in the brine shrimp Artemia have indicated that there are proteinases stored in the cryptobiotic embryo and that their activities are regulated during subsequent development. In order to study whether the phenomenon of proteinase storage is more general and not necessarily related to cryptobiosis, we are now carrying out this study in Drosophila. We have already described the presence of a cathepsin B-like proteinase in oocytes and em- bryos of Drosophila and studied its developmental regulation and implication in yolk degradation (Medina et al., 1988; Medina and Vallejo, 1989). We present here evidence of a serine proteinase stored in the mature oocyte.
MATERIALS AND METHODS
Drosophila melanogaster (Oregon-R strain) was cultured at 25°C as usual (Elgin and Miller, 1977). Mature oocytes were obtained (Petriet al., 1977) taking advantage of the fact that mature oocytes adhere to glass while the immature, do not. Embryos were collected routinely for 2 h, being that these embryos were considered on average to be 1 h-old, the collected embryos were allowed to develop the additional time required (for example, 5 h to obtain 6 h-old embryos).
Homogenization and fractionation
Embryos and oocytes were dechorionated, unless other- wise indicated, prior to hand homogenization in 2-4 vol of a medium (Vallejo et al., 1981) composed of 0.3 M sucrose, 15% Ficol1400, 25 mM HEPES buffer, 60 mM KCI, 15 mM NaCl, 5 mM MgCl 2, 0.5 mM CaC12, 1 mM sodium borate,
*To whom all correspondence should be addressed.
adjusted to pH 7.5. This medium has been proven to preserve the embryonic subcellular structures (Vallejo et al., 1981; Roggen and Slegers, 1985). Subcellular fractionation was accomplished by centrifuging the homogenates at 2500rpm for 20min (500g fraction). The supernatant was further centrifuged at 15,000 rpm for 30min (27,000g fraction) and the resulting supernatant was spinned at 38,000 rpm for 60min giving a pellet (100,000g fraction) and a supernatant (soluble fraction). When only the soluble and the particulate fractions were obtained, the homogenate was centrifuged at 27,000g for 30 miu.
Proteinase assay
The proteinase activity in the resuspended particulate fractions was assayed after disruption by vortexing followed by 4-5 cycles of freezing and thawing. The assay was carried out routinely at 37°C, with 2 mg protamine (Calbiochem) and 50 mM potassium phosphate buffer, pH 7.3 in a final vol of 1 ml, according to the fluorescamine-based method of Garesse et al. (1979). One unit is, as defined previously (Garesse et al., 1979), the amount of proteolysis equivalent to the increase in fluorescence observed after labelling with fluorescamine the new or-amino-groups produced during the hydrolysis of protamine by 7 ng of trypsin (IX Sigma) at 37°C for 10min. The activity on polylysine (15-30kDa), casein and albumin (Sigma) was also determined with fluorescamine. The effect of inhibitors (Sigma, except Ep 475 which was a gift from Dr K. Hanada, Taisho Pharmaceuti- cals Co., Saitama, Japan) was determined after 20min preincubation with the enzyme at 4°C, using protamine as substrate.
In extracts of embryos of Drosophila only two proteolytic activities have been detected on protamine. One is active at acid pH and is completely inhibited by specific inhibitors of cysteine proteinases (Medina et al., 1988) and the other is active in the neutral to alkaline pH range and is the proteinase studied here.
Percoll density gradient
A Percoll gradient in 0.25 M sucrose, l0 mM HCl-Tris pH 7.4 was performed by centrifuging a 30% solution at 50,000 g for 45 min. The activity was assayed after 4-5 cycles of freezing and thawing of the fractions.
687

688 MIGUEL MEDINA and CAgMEN G. VALLEJO
ioo
>-
>
t~ > ~c
...i
/ /
e'.o ;o 81o ~o pH
Fig. 1. pH-dependent activity of the neutral proteinase of Drosophila. The proteinase was assayed in 50 mM buffer (© O, phosphate; A A, borate) at the pH indi- cated. The soluble fraction of 21 h-old embryos was used as the source of enzyme. The activity was also assayed with the purified enzyme and with that of early embryos (6 h)
presenting a similar profile (not shown).
Biogel P-60 chromatography A Biogel P-60 column (73 x 1.2 cm) was equilibrated in
50mM potassium phosphate buffer pH 7.5, 0.1 M potas- sium chloride, 20% glycerol. The column was calibrated with the protein markers ovalbumin (1; 43 kDa), chymo- trypsinogen A (2; 25 kDa) and ribonuclease (3; 13.7 kDa).
RESULTS
We have found a proteinase activity in Drosophila embryos with optimum activity in the neutral to alkaline pH range (Fig. 1). We present here different characteristics of the proteinase.
Developmental regulation
The assay of the proteinase in mature oocytes and after fertilization indicated that the activity was de- velopmentally regulated. The activity was detected in the mature oocytes and increased exponentially dur-
Table 1. Subcellular localization of the neutral proteinase of Drosophila in early embryos
Fraction 500 g 27,000 g 1000,000 g Soluble 3.5 h-old embryos 68 7 2 23 The homogenate was obtained and centrifuged as indicated in
Materials and Methods. The data stand for the percentual distribution of the proteinase activity. The activity of the hom- ogenate is as in Fig. 2. The data represent the average of 2 experiments,
ing the embryonic development, especially from 8 h onwards (Fig. 2).
Developmental changes in subcellular distribution
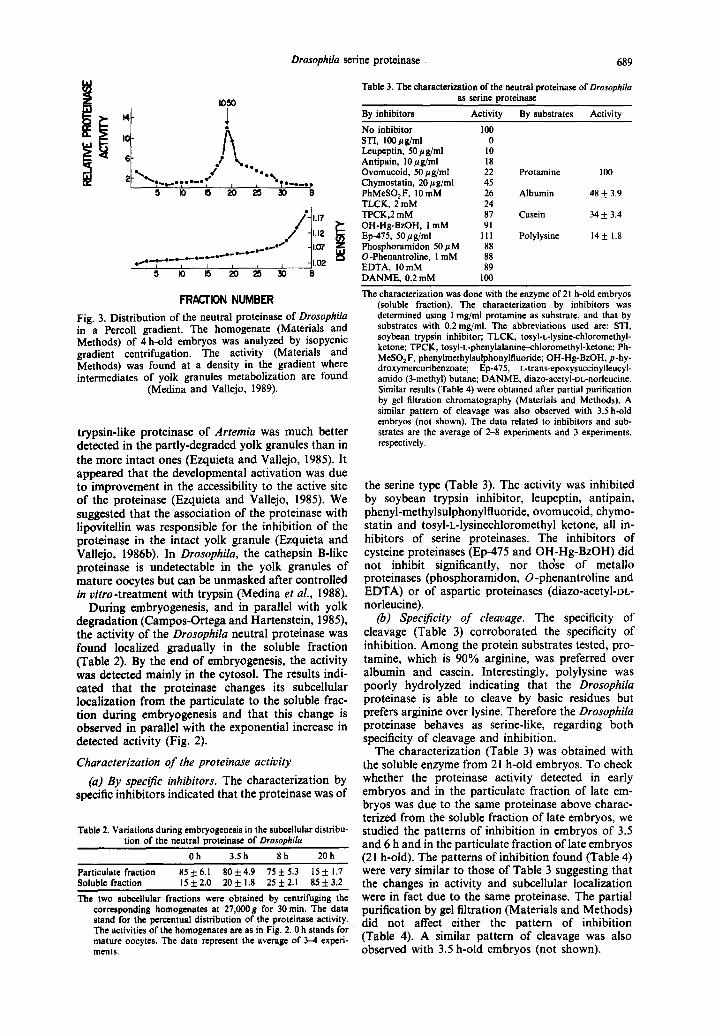
The study of the subceUular distribution of the proteinase activity in late embryos indicated that the enzyme was localized in the soluble fraction. The same study in oocytes or early embryos indicated that the activity was mainly associated with the 500g fraction (Table 1). This fraction contains, in early embryos, yolk granules and nuclei by microscopic and biochemical observations (Medina et al., 1988; Medina and VaUejo, 1989). The fact that the oocyte has only one nucleus but is loaded with yolk granules (Campos-Ortega and Hartenstein, 1985) suggested that the proteinase was associated with these embry- onic structures. We have further purified the proteinase activity by isopycnic density gradient cen- trifugation. The experiment was carried out with a homogenate of embryos of 4 h development. At this time, over 50% of yolk granules have already started degradation (Medina and Vallejo, 1989). The proteinase activity (Fig. 3) in the Percoll gradient was detected mainly at a density of 1.05, where we have observed intermediates of degradation of yolk gran- ules to band (Medina and VaUejo, 1989). Intact yolk granules are detected at about density 1.07 where a small activity was observed (Fig. 3). This low activity probably indicates that the proteinase is more difficult to detect (Materials and Methods) in the intact yolk granule than in the granule which has already undergone metabolization. The activity of the
i 60
3O
.~ I0
U)
y 5 I0 15 ~0
O~ytw Embr~o~lle~ ( h ) Hatching
Fig. 2, Developmental activation of the neutral proteinase of Drosophila. Mature oocytes and embryos were obtained as indicated in Materials and Methods. The embryos were then homogenized (Materials and Methods) without previous dechorionization, to allow disruption during homogenization of subcellular organeltes in order to avoid latency. The activity was determined in the whole homogenates
and is expressed in units per mg protein. The data represent the average of 3-8 experiments.
Drosophila serine proteinase
5 I0 15 20 25 30 B
*t1.17 " / / 1''.2 !
, , , ~ " -41.07 e ,= .o_o_e - -e .e - - e=" e ~' I i I I ~ J 11.02
5 IO 15 20 25 :30 B
FRACTION N U M B E R
Fig. 3. Distribution of the neutral proteinase of Drosophila in a Percoll gradient. The homogenate (Materials and Methods) of 4 h-old embryos was analyzed by isopycnic gradient centrifugation. The activity (Materials and Methods) was found at a density in the gradient where intermediates of yolk granules metabolization are found
(Medina and Vallejo, 1989).
trypsin-like proteinase of Artemia was much better detected in the partly-degraded yolk granules than in the more intact ones (Ezquieta and Vallejo, 1985). It appeared that the developmental activation was due to improvement in the accessibility to the active site of the proteinase (Ezquieta and Vallejo, 1985). We suggested that the association of the proteinase with lipovitellin was responsible for the inhibit ion of the proteinase in the intact yolk granule (Ezquieta and Vallejo, 1986b). In Drosophila, the cathepsin B-like proteinase is undetectable in the yolk granules of mature oocytes but can be unmasked after controlled in vitro-treatment with trypsin (Medina et al., 1988).
During embryogenesis, and in parallel with yolk degradation (Campos-Ortega and Hartenstein, 1985), the activity of the Drosophila neutral proteinase was found localized gradually in the soluble fraction (Table 2). By the end of embryogenesis, the activity was detected mainly in the cytosol. The results indi- cated that the proteinase changes its subcellular localization from the particulate to the soluble frac- tion during embryogenesis and that this change is observed in parallel with the exponential increase in detected activity (Fig. 2).
Characterization of the proteinase activity
(a) By specific inhibitors. The characterization by specific inhibitors indicated that the proteinase was of
Table 2. Variations during embryogenesis in the subcellular distribu- tion of the neutral proteinase of Drosophila
Oh 3.5h 8h 20h Particulate fraction 85 + 6.1 80_+ 4.9 75 + 5.3 15 ± 1.7 Soluble fraction 15-t-2.0 20±1.8 25+2.1 85±3.2 The two subccllular fractions were obtained by centrifuging the
corresponding homogenates at 27,000g for 30 rain. The data stand for the pereentual distribution of the proteinase activity~ The activities of the homogenates are as in Fig. 2. 0 h stands for mature ooeytes. The data represent the average of 3-4 experi- ments.
689
Table 3. The characterization of the neutral proteinase of Drosophila as serine proteinase
By inhibitors Activity By substrates Activity No inhibitor 100 STI, 100/Jg/ml 0 l.~upeptin, 50/~g/ml 10 Antipain, 10/~g/ml 18 Ovomucoid, 50/~g/ml 22 Chymostatin, 20 #g/ml 45 PhMeSO2F, 10 mM 26 TLCK, 2 mM 24 TPCK,2 mM 87 OH-Hg-BzOH, 1 mM 91 Ep-475, 50/~g/ml 111 Phosphoramidon 50/~M 88 O-Phenantroline, 1 mM 88 EDTA, 10 mM 89 DANME, 0.2 mM 100
Protamine 100
Albumin 48 ± 3.9
Casein 34 + 3.4
Polylysine 14 + 1.8
The characterization was done with the enzyme of 21 h-old embryos (soluble fraction). The characterization by inhibitors was determined using I mg/ml protamine as substrate, and that by substrates with 0.2 mg/ml. The abbreviations used are: ST1, soybean trypsin inhibitor; TLCK, tosyI-L-lysine-ehloromethyl- ketone; TPCK, tosyI-L-pbenylalanine--chloromethyl-ketone; Ph- MeSO2F, phenylmethylsulphonylfluoride; OH-Hg-BzOH, p-hy- droxymercuribenzoate; Ep-475, L-trans-epoxysuccinylleucyl- amido (3-methyl) butane; DANME, diazo-acetyl-DL-norleucine. Similar results (Table 4) were obtained after partial purification by gel filtration chromatography (Materials and Methods). A similar pattern of cleavage was also observed with 3.5 h-old embryos (not shown). The data related to inhibitors and sub- strates are the average of 2-8 experiments and 3 experiments, respectively.
the serine type (Table 3). The activity was inhibited by soybean trypsin inhibitor, leupeptin, antipain, phenyl-methylsulphonyifluoride, ovomucoid, chymo- statin and tosyI-L-lysinechloromethyl ketone, all in- hibitors of serine proteinases. The inhibitors of cysteine proteinases (Ep-475 and OH-Hg-BzOH) did not inhibit significantly, nor thdse of metallo proteinases (phosphoramidon, O-phenantroline and EDTA) or of aspartic proteinases (diazo-acetyl-DL- norleucine).
(b) Specificity of cleavage. The specificity of cleavage (Table 3) corroborated the specificity of inhibition. Among the protein substrates tested, pro- tamine, which is 90% arginine, was preferred over albumin and casein. Interestingly, polylysine was poorly hydrolyzed indicating that the Drosophila proteinase is able to cleave by basic residues but prefers arginine over lysine. Therefore the Drosophila proteinase behaves as serine-like, regarding both specificity of cleavage and inhibition.
The characterization (Table 3) was obtained with the soluble enzyme from 21 h-old embryos. To check whether the proteinase activity detected in early embryos and in the particulate fraction of late em- bryos was due to the same proteinase above charac- terized from the soluble fraction of late embryos, we studied the patterns of inhibition in embryos of 3.5 and 6 h and in the particulate fraction of late embryos (21 h-old). The patterns of inhibit ion found (Table 4) were very similar to those of Table 3 suggesting that the changes in activity and subcellular localization were in fact due to the same proteinase. The partial purification by gel filtration (Materials and Methods) did not affect either the pattern of inhibition (Table 4). A similar pattern of cleavage was also observed with 3.5 h-old embryos (not shown).

690 MIGUEL MEDINA and CAR~N G. VALLEJO
Table 4. The characterization of the neutral proteinase of Drosophila embryogenesis
3.5h 6h 21h
H H H S P Ep
No inhibitor 100 100 100 100 100 100 STI, 100/~g/ml 0 0 0 0 0 0 Leupeptin, 50 #g/ml 13 9 16 10 18 13 Antipain, 10 gg/ml 22 25 20 18 33 - - Ovomucoid, 50 #g/ml - - - - 26 22 - - 18 Chymostatin, 20 ttg/ml - - - - 48 45 - - 39 PhMeSO 2 F, 10 mM 44 54 28 26 - - - - TLCK, 2 mM - - - - 11 24 - - 22 TPCK,2 mM - - - - 103 87 - - - - OH-Hg-BzOH, 1 mM - - - - 82 91 - - 86 Ep-475, 50/~g/ml 98 104 106 111 - - - - Phosphoramidon 50 #M - - - - 82 88 - - 96 O-Phenantroline, 1 mM - - - - -- 88 75 - - EDTA, 10mM 87 93 - - 89 78 - - DANME, 0.2 mM - - - - 105 100 - - - -
during
The abreviations and the conditions of assay are as under Table 3. The data of 3.5, 6 and 21 h were obtained with the complete homogenates (H) of embryos of those times of development. The characterization of the particulate (P) and soluble (S) fraction of 21 h-embryos is also shown as well as that of the soluble activity after partial purification (Ep) by gel filtration (Materials and Methods)• The data obtained with the soluble fraction of late embryos (21 h-old) are also in Table 3 and are shown here for comparison•
(c) Michaelis-Menten kinetics. W e h a v e deter - m i n e d the s a t u r a t i o n cu rve o f t he p r o t e i n a s e vs the r o u t i n e s u b s t r a t e p r o t a m i n e . T h e resu l t s i nd i ca t ed h y p e r b o l i c k ine t ics . T h e Km va lue ca l cu l a t ed f r o m a L i n e w e a v e r - B u r k p lo t (Fig. 4) was 36 / ~M . T h e fac t t h a t on ly a s lope was o b s e r v e d in the rec ip roca l p lo t o f t he k ine t ic d a t a s u g g e s t e d t h a t we were dea l i ng w i th on ly o n e p ro t e inase .
(d) Molecular weight determination. T h e m o l e c u l a r we igh t o f t he ser ine p r o t e i n a s e was d e t e r m i n e d af te r f i l ter ing t he so lub le f r a c t i on o f late e m b r y o s (21 h- o ld) t h r o u g h a Biogel P-60 c o l u m n (Fig. 5) c a l i b r a t ed w i th p r o t e i n m a r k e r s . T h e va l ue d e t e r m i n e d w a s 25,000. T h e fac t t h a t o n l y a p e a k o f ac t iv i ty was o b s e r v e d a lso s u g g e s t s t h a t t he ac t iv i ty de t ec t ed is d u e to on ly o n e p ro t e inase .
DISCUSSION
W e h a v e f o u n d a ser ine p r o t e i n a s e in the m a t u r e o o e y t e s of Drosophila, w h i c h a p p e a r s to be a s s o c i a t e d wi th yo lk g r anu l e s . T h e ac t iv i ty i nc rease s e x p o n e n - t ia l ly d u r i n g e m b r y o g e n e s i s a n d , in paral le l , c h a n g e s i ts loca l i za t ion to t he so lub le f rac t ion . W e h a v e desc r ibed p r e v i o u s l y a t ryps in - l ike p r o t e i n a s e in t he
b r ine s h r i m p Artemia w h o s e ac t iv i ty i nc reased expo- nen t i a l ly d u r i n g la rva l d e v e l o p m e n t • T h e Artemia n e u t r a l p r o t e i n a s e was a lso local ized in yo lk g r a n u l e s in t he d o r m a n t e m b r y o a n d a p p e a r e d g r a d u a l l y in the
! 4.6
4.4 !: 1.0 1.2 1.4 1.6 1.8
v/vo Fig. 5. The molecular weight of the neutral proteinase of Drosophila. The molecular weight was determined by filtra- tion of the enzyme sample through a calibrated Biogel P-60 column as indicated in Materials and Methods. A molecular mass of about 25 kDa was calculated from the elution
volume indicated by the arrow.
~ '~I/ i I , r 0,04
0.5 I 1.5 2 Km:36pM
S (rag I rnl )
fly
o~8°'12 ~ 5 I0 15 20
I / S ( m g / m l )
Fig. 4. Effect of the substrate concentration on the initial velocity of the neutral proteinase of Drosophila. Protamine was used as substrate at the concentrations indicated. As also indicated, the Km was calculated from a Lineweaver-Burk plot of the data. The soluble fraction of 21 h-old embryos was used as the source
of enzyme.

Drosophila serine proteinase 691
cytosol during the larval development (Ezquieta and Vallejo, 1985), in parallel with yolk degradation (Perona et al., 1987, 1988). The comparative data on Drosophila and Artemia may suggest that the pres- ence of a serine proteinase in the yolk granules is a common feature at the beginning of development. We do not believe that these proteinases are involved in the degradation of yolk in the corresponding system (Ezquieta and Vallejo, 1986b; Perona et al., 1987; Medina et al., 1988). The degradation of yolk in both systems has been attributed to a cysteine, cathepsin B-like proteinase (Perona and Vallejo, 1985; Medina et al., 1988; Medina and Vallejo, 1989) and we have found in both cases that lysosomotropic agents in- hibit yolk degradation (Ezquieta and Vallejo, 1986c; Perona et al., 1987; Medina et al., 1988). Despite the similarities described between the proteinases of Drosophila and Artemia, there appear to be differ- ences in the respective characterizations. The Artemia proteinase behaves as trypsin-like, being inhibited only by trypsin inhibitors (Ezquieta and Vallejo, 1985, 1987) and cleaving by arginine and lysine with similar affinity (Ezquieta and Vallejo, unpublished). The Drosophila proteinase cleaves lysine poorly and is inhibited by a broader spectrum of inhibitors. These characteristics have lead us to classify it as a serine proteinase.
With regard to the possible role of the Drosophila serine proteinase during development, we have sug- gested recently (Medina et al., 1988) that it might be involved in the limited proteolysis that produces activation of the cysteine, cathepsin B-like proteinase proposed to be involved in yolk degradation in Drosophila.
Recently (Indrasith et al., 1988), a serine proteinase has been described in Bombix mori. The molecular weight, specificity of cleavage and inhibition observed with specific inhibitors were very similar to those of the Drosophila enzyme. The activity is also develop- mentally regulated, with a pattern similar to the one found in Drosophila (this paper) and Artemia (Ezquieta and Vallejo, 1987). The only data reported in relation with the subcellular localization was that the enzyme was 90% soluble towards the end of the stage, when yolk is already digested. It would be interesting to study the subcellular localization dur- ing embryogenesis. As suggested above, the presence of a serine proteinase in yolk granules may be a common feature of different embryonic systems.
Acknowledgements--This work has been supported by Comisi6n Asesora de Investigaci6n Cientifica y T~cnica. We thank Dr B. Ezquieta for critical reading of the manuscript.
REFERENCES
Campos-Ortega J. A. and Harstentein V. (1985) The Embry- onic Development of Drosophila melanogaster. Springer, Berlin.
Elgin S. C. R. and Miller D. W. (1977) Mass rearing of flies and mass production and harvesting of embryos. In The Genetics and Biology of Drosophila (Edited by Ashburner
M. and Wright T. R. G.), Vol. 2a, pp. 112-121. Academic Press, New York.
Ezquieta B. and Vallejo C. G. (1985) The trypsin-like proteinase of Artemia: yolk localization and develop- mental activation. Comp. Biochem. Physiol. 8211, 731-736.
Ezquieta B. and Vallejo C. G. (1986a) Diguanosine 5", 5"-pl; p4-tetraphosphate causes specific inhibition and desensitization of Artemia trypsin-like proteinase in the hydrolysis of a high-affinity, arginine-rich substrate. Biochim. biophys. Acta 883, 380-382.
Ezquieta B. and Vallejo C. G. (1986b) Lipovitellin inhibition of Artemia trypsin-like proteinase: a role for a storage protein in regulating proteinase activity during develop- ment. Arch. Biochem. Biophys. 250, 410-417.
Ezquieta B. and Vallejo C. G. (1986c) Artemia trypsin-like proteinase: developmental activation is inhibited by a lysosomotropic agent. Biol. Cell 58, 227-232.
Ezquieta B. and Vallejo C. G. (1987) Artemia trypsin-like proteinase: a developmentally regulated proteinase. In Artemia Research and its Applications. Vol. 2, Biochem- istry and Molecular Biology (Edited by Declair W., Moens L., Slegers H., Jaspers E. and Sorgeloos P.), pp. 53-65. Universa Press, Wetteren.
Garesse R., Castell J. V., Vallejo C. G. and Marco R. (1979) A fluorescamine-based sensitive method for the assay of proteinase capable of detecting the initial cleavage steps of a protein. Eur. J. Biochem. 99, 253-259.
Garesse R., Perona R., Marco R. and Vallejo C. G. (1980) The unmasking of proteolytic activity during the early development of Artemia salina. Eur. J. Biochem. 106, 225-231.
Indrasith L. S., Sasaki T. and Yamashita O. (1988) A unique protease responsible for selective degradation of a yolk protein in Bombyx mori. J. biol. Chem. 263, 1045-1051.
Medina M. and Vallejo C. G (1989) The maternal origin of acid hydrolases in Drosophila and their relation with yolk degradation. Dev. Growth. Diff. 31, 241-247.
Medina M., Le6n P. and Vallejo C. G. (1988) Drosophila cathepsin B-like proteinase. A suggested role in yolk degradation. Arch. Biochem. Biophys. 263, 355-363.
Perona R. and Vallejo C. G. (1982) The lysosomal proteinase of Artemia. Purification and characterization. Eur. J. Biochem. 124, 357-362.
Perona R. and Vallejo C. G. (1985) Acid hydrolases during Artemia development. A role in yolk degradation. Comp. Biochem. Physiol. 81B, 993-1000.
Perona R., Ezquieta B. and Vallejo C. G. (1987) The degradation of yolk in Artemia. In Artemia Research and its Applications. 1Iol. 2, Biochemistry and Molecular Biology (Edited by Declair W., Moens L., Slegers H., Jaspers E. and Sorgeloos P.), pp. 105-123. Universa Press, Wetteren.
Perona R., B6s J. C. and Vallejo C. G. (1988) The degra- dation of yolk in the brine shrimp Artemia. Involvement of the lysosomal system by biochemical and morpho- logical studies. Biol. Cell 63, 361-366.
Petri W. H., Wyman M. and Kafatos F. C. (1977) The Genetics and Biology of Drosophila (Edited by Ashburner M. and Wright T. R. G), Vol. 2a. Academic Press, New York.
Roggen E. and Slegers H. (1985) Isolation and characteriz- ation of cytoplasmic poly(A) polymerase from crypto- biotic gastrulae of Artemia salina. Fur. J. Biochem. 147, 225-232.
Vallejo C. G., Perona R., Garesse R. and Marco R. (1981) The stability of the yolk granules of Artemia. An im- proved method for their isolation and study. Cell Diff. 10, 343-356.