abstract comparative digestive techniques for the massive
TRANSCRIPT

Vanessa Goodman, Allison FisherZOOL 430Dr. Butler10/4/21
Comparative Digestive Techniques for the Massive Herbivorous Dinosaur: Borealopeltamarkmitchelli
Abstract
This study examines comparative digestive techniques as potential models for a large
herbivorous dinosaur living in Western Canada during the late Cretaceous period. Through the
examination of fossil records of Borealopelta markmitchelli and data concerning paleolithic flora
in the region of its discovery, its feeding tendencies and behaviors are brought to a new light.
The most outstanding concern for the nodosaur was its ability to acquire sufficient energy from
surrounding vegetation in order to sustain its metabolic demands that accompanied its large size.
By calculating digestive tract parameters on the basis of daily metabolic rate and nutritional
contents of a tree fern as a model food item, we were able to assess the ease with which B.
markmitchelli was able to survive. We hypothesized that hindgut fermentation would greatly
reduce the difficulty of meeting metabolic demands and thus allow for behaviors such as
selective feeding which are supported by morphological evidence. Our findings show that the
possession of a hindgut fermentation mechanism provides a strong explanation for how the
animal would have thrived in its summer climate and may inform further exploration about
winter survival.
Introduction
Description of B. markmitchelli
Borealopelta markmitchelli, a recently discovered species of nodosaur (placed in family
Nodosauridae, suborder Ankylosauria, and order Ornithischia), roamed Western Canada during

the Albian stage of the Early Cretaceous period, over 110 million years ago (Brown et al. 2017,
Greshko 2021). B. markmitchelli was a quadrupedal dinosaur with short legs, a wide torso, and a
tapered tail. As a member of Ankylosauria, it had a heavy coat of armor with bony osteoderm
plates and impressive keratinous spikes along its entire dorsal region. B. markmitchelli’s physical
characteristics generally resemble its more famous club-tailed relative, the Ankylosaurus,
however, the nodosaur was clubless. The species is estimated to have weighed 1300 kilograms
with a length of 5.5 meters, width of 1 meter, and height of 2.5 meters (Brown et al. 2017). Its
daily metabolic rate was previously found to be 7005 kJ/d to support average thermoregulation
and activity levels at 20oC (Fisher, Goodman 2021).
Figure 1. Sketch of Borealopelta markmitchelli with length and height dimensions (Brown et al. 2017,Fisher 2021).
Fossil Morphology
A singular fossil found in Alberta, Canada in 2011 represents the only known specimen
of the species (Henderson 2013). It is one of the best preserved specimens of an armored
dinosaur ever found, providing a holistic understanding of morphological traits which can be
used to deduce much about this animal’s life. Highlights of its preservation include completely

intact spikes, plates, and scale formations along with skin pigmentation and skull morphologies
(Brown et al. 2017). Analysis of these characteristics provide insight into the tendencies of the
nodosaur’s feeding behaviors and level of activity which can help us understand how it fulfilled
the demands of its diet. The greatest clues about the diet and digestion of B. markmitchelli were
found in the stomach contents, which were spectacularly preserved. This will be discussed after
the framework of its broader physical features and habitat have been examined.
The nodosaur family’s numerous amounts of small and dull teeth provide strong evidence
that they were herbivores who especially consumed soft types of vegetation (Colbert 1961). The
relatively narrow muzzle of B. markmitchelli suggests it practiced a more careful selection of
foodstuffs, choosing each bite with intention (Henderson, 2013). The body cavity was wide and
capacious which is common for bulky-plant eaters (Colbert 1961). Another important feature of
the dinosaur's skeleton include its greatly arched ribs which create this larger abdomen to
accommodate the organs that occupy it (Colbert 1961). It is important to recall that their diet
would also be limited by their height. B. markmitchelli’s neck suggests it was not adapted for
reaching taller foliage as its armor likely inhibited excessive upward mobility. Thus, their diet
was likely made up of plants at or below snout level, approximately 2 meters high (Brown et al.
2017).
Cladistic analyses suggest that the Stegosaurus and B. markmitchelli shared a common
bipedal ancestor. In their divergence, the nodosaur lineage evolved longer forelimbs, giving them
the edge on quadrupedal locomotion which allowed them to outcompete Stegosaurus (Colbert
1961). Their enhanced ability to walk may have supported their selective feeding strategy by
allowing them to browse through a much larger area in order to locate the most ideal food
sources.

Preserved evidence of pigmentation patterns across B. markmitchelli’s armor reveal a
countershading form of crypsis (Brown et al. 2017). In combination with its durable osteoderm
exterior and large protective spikes, this dinosaur had highly evolved protection against
predation. Acrocanthosaurus, the top predator of the time, was much taller than B. markmitchelli
(Harris 1998). Brown countershading patternation would prevent detection from above or from
the side against a backdrop of soil or woody tree trunks (Brown et al 2017). This suggests
feeding times could be leisurely as they were not in constant danger of being attacked if their
attention was focused on browsing.
Paleoclimate of Western Canada
The paleolatitude of the Western Canadian area would have been about 50-60 degrees
North during the Cretaceuos period (Wan 1996). Analysis of floral fossils suggested that
summers would have been warm and humid with abundant rainfall (Wan 1996). A sequence of
Cretaceous sedimentary materials about 200-400km west of where B. markmitchelli was found,
estimates that winter temperatures could have reached as low as -15oC (Brown et al. 2020; Wan
1996). However, the mean temperature of the cold months was likely warmer. Oxygen isotope
data argues against prolonged frosting periods (Wan 1996).
Lower Cretaceous Flora
Some common flora present during the time of B. markmitchelli’s reign were
leptosporangiate ferns, conifers and other gymnosperms, ginkgophytes, and cycads (Brown et al.
2020). Most of these plants were high in fiber and low in compounds that provide quickly
available energy (Brown et al. 2020). Accordingly, the dinosaur likely had to selectively graze to
find the most attractive foods to sustain its size and lifestyle. The animal likely grazed on leafy
materials and avoided twigs, showing discretion down to each bite (Brown et al. 2020). Hummel

et al. found the metabolic energy released by in vitro fermentation of leptosporangiate ferns and
other food sources similar to what would have been available to B. markmitchelli (2008). They
showed that ferns were generally more energy dense and better sources of protein compared to
cycads and conifers (Hummel, 2008). This suggests a motive for selective feeding of ferns when
in need of quickly available energy and protein. However, due to the deciduous nature of the
Gates flora, winter foliages were more abundant in evergreens and could have made up the
majority of the animals diet during these months (Hummel, 2008).
Stomach Contents
The stomach contents of B. markmitchelli were analyzed and shown to have 88% leaf
matter, of which 85% was leptosporangiate fern remnants (Brown et al. 2020). Such a high
proportion of leaf matter to non-digestible materials suggests browsing was indeed common
practice. Given that ferns were the most abundant food item found in the fossil and that it was
the most nutritionally dense flora available to the nodosaur, digestive modeling in this study will
use the nutritional values of fern. There is no literature on the exact nutritional breakdown of the
ancient fern species. For the purpose of this study we will use nutrient ratios obtained from the
tree fern, a vegetable fern consumed by humans, to construct a diet model for B. markmitchelli.
Human-selected food items tend to be very energy dense so it is plausible to speculate that food
items available to the animal may not have been as nutritious. However, it was discovered that B.
markmitchelli had foraged for new growths after landscapes were scorched by forest fires,
evidenced by charcoal remnants in the stomach (Brown et al. 2020). This behavior may have
been opportunistic, or perhaps this is evidence that the nodosaur consistently sought out
premature plant items due to their nutritional superiority.
Digestion Patterns

Vertebrate digestion is conserved in its structure and typically follows the basic model:
mouth, esophagus, stomach, small intestine, large intestine, rectum, and anus. Across species,
there are many variations in size, shape, and other adaptations within the system in response to
diet and lifestyle (Withers 1992). An adaptation found in the fossil evidence of the nodosaur is
the presence of gastroliths which are stones held in the stomach that assist in the mechanical
breakdown of food (Brown et al. 2020). Gastroliths are common in animals that lack grinding
molars, like in the case of B. markmitchelli. Digestion tract models provide insight into how
organisms process energy and can therefore give clues into various other traits like feeding
behaviors and adaptations.
The aim of this study is to create two digestion models: one of basic nutrient acquisition
via mechanical and chemical breakdown followed by small intestine absorption, and one with
use of fermenting micro symbionts for additional energy acquisition from otherwise
non-digestible plant fiber, like cellulose. The possibility of foregut fermentation is unlikely
considering there is no record of foregut fermentation being used in reptile species (Farlow
1987). It’s unlikely that an animal of such massive size would be able to efficiently obtain
enough energy for growth and locomotion from directly digesting plants with low concentrations
of available carbohydrates and proteins. Previous studies find that it is more likely that giant
herbivorous dinosaurs relied on fermentation as a means to utilize the high volumes of cellulose
present in the available flora (Farlow 1987).
Hypothesis
We suspect that fermentation was an essential mechanism for optimal digestion of
low-energy plant materials in B. markmitchelli. By modeling both direct digestion and digestion
supplemented with hindgut fermentation, we will quantify the expected increase in usable energy

obtained by fermentation. The confirmation of this adaptation would yield strong supporting
evidence for the allowance of other behaviors, such as selective feeding, which have been
speculated by many researchers on the basis of morphology.
Methods
Direct Digestion:
Digestion tract dimensions and structure were modeled using the nutritional content of a
tree fern, Diplazium esculentum.
Energy Contribution of the Tree Fern
Using data from the USDA FoodData Central on percentage composition (%
composition) of each nutrient present in tree fern coupled with energy density (ED) of each
nutrient (carbohydrate, protein, fat), we were able to calculate the net amount of energy per gram
of food that the dinosaur would obtain with consumption (2021).
𝐸𝑛𝑒𝑟𝑔𝑦 𝑐𝑜𝑛𝑡𝑟𝑖𝑏𝑢𝑡𝑖𝑜𝑛 (𝑘𝐽/𝑔) = % 𝑐𝑜𝑚𝑝𝑜𝑠𝑖𝑡𝑖𝑜𝑛 ✕ 𝐸𝐷 (𝑘𝐽/𝑔)
Table 1. Percents of nutrient out of the total mass of a tree fern and the energy densities of each nutrient.
Nutrient Percent of total mass (USDA) Energy density (kJ/g)(Withers 1992)
Carbohydrate 11% (3.7% fiber)= 7.3% net 15.9
Protein 0.29% 18.15
Lipid 0.07% 39.2
Water 88.6% 0
The energy contributions were summed to find the total energy density (EDtot) of a tree
fern.
Mass of Food Assimilated

The mass of tree fern required to assimilate per day (ma) was found using daily metabolic
rate (DMR=7005kJ/day) (Fisher, Goodman 2021).
𝑚𝑎 (𝑔/𝑑𝑎𝑦) = 𝐷𝑀𝑅 (𝑘𝐽/𝑑𝑎𝑦) ÷ 𝐸𝐷
𝑡𝑜𝑡 (𝑘𝐽/𝑔)
Additionally, amounts of each nutrient assimilated per day were found and converted to
moles per day (M) using the amount assimilated (ma) in g/day, the percent composition, and the
specific molecular weight (Mw) in g/mol.
M=𝑚
𝑎× % 𝑐𝑜𝑚𝑝
𝑀𝑤
Dietary fiber was originally treated as an indigestible food source when building this
model before considering fermenting adaptations.
Mass of Food Ingested
The amount of food that would need to be ingested (mi) in order to meet the energetic
demands of daily metabolic rate was calculated using an assimilation efficiency value of 50%.
This number is comparable to previous estimates of other megaherbivore dinosaurs based on the
high fiber content of available forage (Weaver 1983).
𝑚𝑖 (𝑔/𝑑𝑎𝑦) = 𝑚
𝑎÷ 0. 5
Number of Food Items
The mass of a single fern frond was calculated using dimensions of the plant to find its
volume. Volume of a rectangle divided by 2 may accurately describe the shape of the plant,
(Figure 2).
v (m3)=𝑙×𝑤×ℎ
2

Figure 2. Approximating the volume of a ground fern (Goodman, 2021).
The density of water (d) in kg/m3 was chosen as an accurate representation of the density
of the plant since the USDA reports tree fern having 88% water content (2021). To find the mass
of each frond (mfrond), this density was multiplied by the calculated volume (m3) of one frond.
mfrond (kg)= d v×
Using this calculated mass we were able to deduce the number of food items that the
dinosaur would have to consume per day for both digestive tract models.
Number of items per day= 𝑚𝑖
÷ 𝑚𝑓𝑟𝑜𝑛𝑑
Rate of Nutrition Uptake in the Small Intestine
Since carbohydrates are the most abundant nutrient of the food source, it was considered
as the limiting factor when investigating the digestive tract surface area and dimensions. The rate
of glucose uptake in the small intestines of reptiles (R1) was corrected to the body temperature of
our animal during its daytime digestive phase (31.56oC) using a Q10 conversion (Fisher,
Goodman 2021; Withers 1992).
Rcorrect=R1 (2.5)(Tb-37)/10×
Surface Area of the Gut

The required surface area (SA) of the gut was computed using the daily intake in number
of moles of carbohydrates (Mcarb) and the corrected uptake rate (mol/day/cm2).
SA (cm2)=Mcarb Rcorrect÷
Gut Length
Gut diameter was estimated based on the dimensions of a cololite fossil, which is
believed to represent the approximate size of the posterior stomach (Brown et al, 2020). Looking
at the attachment sites of stomach and small intestine in other animals and knowing intestine
diameter is highly conserved, 5 cm seems to be an appropriate diameter (d) of the small intestine
(Withers 1992).
The surface area of the gut is represented by that of a cylinder. Accordingly, we were able
to speculate gut length.
Length (cm)=𝑆𝐴 ÷ π ÷ 𝑑
Fermentation Model:
As an alternative model for digestion, we speculated that B. Markmitchelli utilized
hindgut fermentation via symbiotic microorganisms to increase the nutritional yield of its foods.
Hummel et al. performed an in vitro study to mimic hindgut fermentation in herbivorous
dinosaurs using live microbes fed a variety of dried foliage types. The amount of gas produced
by the microbes was measured and converted to energy released using a standard regression
established for similar plant materials (2008). This was corrected to metabolizable energy (ME)
using previously established coefficients which reflect assimilation efficiency of the energy
produced by fermentation (Weaver 1983). The tree ferns analyzed in this study were found to
have produced 6,900 kJ of metabolizable energy for every kilogram of dry matter allowed to
completely ferment (Hummel et al. 2008).

Energy Produced by Fermentation of the Tree Fern
The ME of dry mass for tree ferns was converted to ME of fresh mass to reflect the actual
state of the food as it was eaten by B. markmitchelli. Knowing a tree fern is 88.6% water, it can
be assumed that dry mass occupies 11.4% of fresh mass (USDA 2021). The energy supplied by
fermentation of fresh ferns was found using:
MEfresh (kJ/kg) =MEdry 0.114×
The ME (kJ) acquired by fermenting the daily amount of fresh ferns ingested in kg (mi)
was found using:
MEdaily = mi MEfresh×
Comparing the Energy Contribution of Fermentation
The amount of fresh ferns ingested each day (mi) had been calculated to supply exactly
enough energy to support the daily metabolic rate via direct digestion. To compare the amount of
energy acquired from the same amount of material through direct digestion and fermentation, the
values were subtracted.
Difference in energy uptake (kJ) = DMR - MEdaily
To find the amount of food required if fermentation was the only mechanism used in food
breakdown (mfermenting), the daily metabolic rate (in kJ) was divided by the ME of fresh ferns (in
kJ/kg).
mfermenting = DMR MEfresh÷
The number of food items required per day for fermentation-only breakdown was found
using the mass of fresh ferns required (mfermenting) divided by the mass of each frond (mfrond):
Number of items per day= 𝑚𝑓𝑒𝑟𝑚𝑒𝑛𝑡𝑖𝑛𝑔
÷ 𝑚𝑓𝑟𝑜𝑛𝑑
Results

Direct Digestion
Table 1. Energy values and contributions of macronutrients in the tree fern (USDA 2021,Withers 1992).
Nutrient Percent of totalmass (USDA)
Energy density(kJ/g) (Withers1992)
Energycontribution(kJ/g)
Moles of foodassimilated(mol/day)
Carbohydrate 11% (-3.7%fiber)=7.3% net
15.9 1.1607 2.3
Protein 0.29% 18.15 0.05264 1.2
Lipid 0.07% 39.2 0.02744 0.015
Water 88.6% 0 0 -
Total - - 1.24 -
The tree fern was found to provide 1.24kJ of energy per gram of weight (1.1607kJ/g of
carbohydrate, 0.05264kJ/g of protein, and 0.02744kJ/g of lipid) (Table 1). To support the daily
metabolic rate of B. markmitchelli (7005 kJ/d), it must have assimilated 5,649 g of tree-fern
equivalent food per day. To assimilate this quantity at 50% efficiency, it must have consumed
11.3 kg or 243.2 full fern fronds per day. Given the need to uptake 2.3 moles of carbohydrate
daily, the surface area of the small intestine would have been 7419cm2 with a length of 4.73
meters.

Figure 3. Gastrointestinal model for B. Markmitchelli (Goodman, 2021).
Fermentation Model
Knowing the metabolizable energy by fermentation (ME) per kg of dry mass of tree ferns
is 6,900 kJ and fresh ferns are 88.6% water, the ME of fresh ferns was found to be 786.6 kJ/kg.
This would mean if the animal ingested the same amount of fresh mass as it needed to support
the DMR using direct digestion, complete fermentation would yield 8,889 kJ. In comparison to
the amount acquired by direct digestion (7005 kJ), fermentation would supply an additional
1,884 kJ of usable energy from the same mass of food. If DMR was supported by fermentation
alone, B. markmitchelli would have to consume 8.9kg of fresh ferns per day, a reduction of 2.4kg
per day compared to direct digestion. This translates to 191.6 individual fronds consumed per
day.
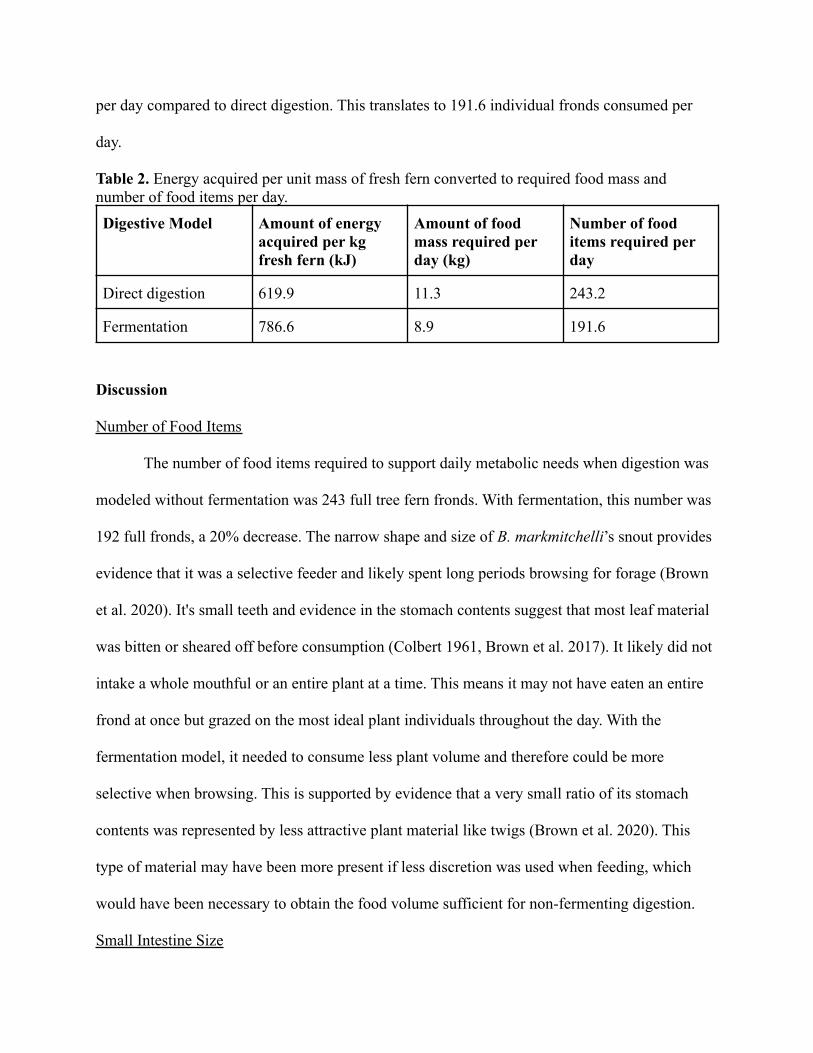
Table 2. Energy acquired per unit mass of fresh fern converted to required food mass andnumber of food items per day.
Digestive Model Amount of energyacquired per kgfresh fern (kJ)
Amount of foodmass required perday (kg)
Number of fooditems required perday
Direct digestion 619.9 11.3 243.2
Fermentation 786.6 8.9 191.6
Discussion
Number of Food Items
The number of food items required to support daily metabolic needs when digestion was
modeled without fermentation was 243 full tree fern fronds. With fermentation, this number was
192 full fronds, a 20% decrease. The narrow shape and size of B. markmitchelli’s snout provides
evidence that it was a selective feeder and likely spent long periods browsing for forage (Brown
et al. 2020). It's small teeth and evidence in the stomach contents suggest that most leaf material
was bitten or sheared off before consumption (Colbert 1961, Brown et al. 2017). It likely did not
intake a whole mouthful or an entire plant at a time. This means it may not have eaten an entire
frond at once but grazed on the most ideal plant individuals throughout the day. With the
fermentation model, it needed to consume less plant volume and therefore could be more
selective when browsing. This is supported by evidence that a very small ratio of its stomach
contents was represented by less attractive plant material like twigs (Brown et al. 2020). This
type of material may have been more present if less discretion was used when feeding, which
would have been necessary to obtain the food volume sufficient for non-fermenting digestion.
Small Intestine Size

The resultant small intestine surface area and length from the direct digestion model is
seemingly small. For an herbivore of such size, a small intestine of less than 5 meters is likely an
underestimation (Brown et al. 2017). This could have been the result of the food source
referenced. Because no available sources give the macronutrient breakdown of a more accurate
fern type, the USDA databank supplied data on a human consumable tree fern (USDA 2021).
Use of this human food source likely overestimates the amount of available energy and
carbohydrates and underestimates non-digestible material as humans don’t possess the cellulose
digesting capabilities that most herbivores utilize (Withers 1992). In reality, the ferns consumed
by B. markmitchelli likely contained a much higher concentration of cellulose which wouldn’t
have been broken down into digestible material by mechanical and chemical processes alone.
The amount of usable carbohydrates would likely represent a lower fraction in the overall mass
of food consumed. Therefore, the energy density of the food source would be lower, the amount
consumed would be higher, and the small intestine would be longer to increase absorption
opportunity.
Though it is clear that some confounding factors may have contributed to a smaller small
intestine than expected based on body size, there is some biological evidence that may
rationalize the function of this decreased size. Primarily, the presence of gastroliths found in the
cololite fossil of the posterior stomach (Brown et al. 2020). B. markmitchelli was not known to
have an intensive food milling apparatus in the mouth so it likely relied on gastric grinding to
increase nutrient availability of food consumed. This type of processing is advantageous because
it decreases the time needed to attain adequate digestibility in the gastrointestinal tract (Farlow
1987). For animals without this type of breakdown, low digestibility is compensated with a
longer small intestine and a longer duration of digestion. The presence of gastroliths accounted

for 50% of the stomach composition in the cololite fossil (Brown et al. 2020). In order to
accommodate this spatial demand, the stomach was probably enlarged and other organs, like the
small intestine, were consequently decreased.
Importance of Fermentation in Winter Months
The fermentation model demonstrated that substantially more energy could have been
attained in its presence. In the event that the diet of B. markmitchelli was less nutritionally dense
than we had estimated, hindgut fermentation would ensure that energy needs were met with ease.
In summer months, this mechanism would enable selective feeding when food resources were
abundant. However, in winter months, food resources were diminished and less selectivity would
have been exercised. Most of the plants in the region’s flora were deciduous, with the exception
of a handful of evergreens. The foliage of the evergreens would have been significantly less
nutritious than the plants that make up the summer diet (Hummel et al. 2008). Fermentation
processes would have been crucial to meet the dinosaur’s metabolic needs throughout the cold
season. Surely, the amount of food mass would increase greatly, but the capacious body would
have enabled the intake. Increased amount of food intake in winter months could also justify the
seemingly small gut length that was calculated in dependence on nutritionally dense food items
present in the summer. Though fern leaf matter made up the majority of the animals stomach
contents at the time of its death, digestive tract surface area and length may be better assumed
using the nutritionally poor food items of the winter months as limiting factors.
Conclusion
Based on the comparative models for digestion with and without fermentation, we can
speculate that fermentation by symbiotic microorganisms in the gut played an important role in
digestion. Fermentation requires a decreased load of food mass consumed daily which

accommodates more selective feeding behaviors. Additionally, food particle reduction by gastric
grinding made digestion more efficient and nutrient uptake more available even with a reduced
small intestine length. The combination of these digestive feeding mechanisms meant that B.
markmitchelli was able to grow and maintain its massive body size even when available food
sources had generally low energy density.
To support and improve these findings, a more accurate reference for nutrient
composition of the fern species actually consumed by B. markmitchelli would yield more
realistic dimensions of the digestive tract. Furthermore, exploring the implications of the
vegetation available and consumed in the winter months may provide further insight to the
significance of hindgut fermentation in B. markmitchelli.
ReferencesBrown, C. M., Henderson, D. M., Vinther, J., Fletcher, I., Sistiaga, A., Herrera, J., & Summons,
R. E. (2017). An Exceptionally Preserved Three-Dimensional Armored Dinosaur RevealsInsights into Coloration and Cretaceous Predator-Prey Dynamics. Current biology : CB,27(16), 2514–2521.e3. https://doi.org/10.1016/j.cub.2017.06.071
Brown, C. M., Greenwood, D. R., Kalyniuk, J. E., Braman, D. R., Henderson, D. M.,Greenwood, C. L., & Basinger, J. F. (2020). Dietary palaeoecology of an EarlyCretaceous armoured dinosaur (Ornithischia; Nodosauridae) based on floral analysis ofstomach contents. Royal Society open science, 7(6), 200305. Retrieved from:Dietary palaeoecology of an Early Cretaceous armoured dinosaur (Ornithischia;Nodosauridae) based on floral analysis of stomach contents
Colbert, E. H. (1961). Dinosaurs: Their discovery and their world. Dutton.Farlow, J. O. (1987). Speculations About the Diet and Digestive Physiology of Herbivorous
Dinosaurs. Paleobiology, 13(1), 60–72. http://www.jstor.org/stable/2400838Fisher, A., Goodman, V. (2021). Metabolism and Thermoregulation of a Massive Herbivorous
Ectotherm in Cretaceous Western Canada.Fooddata Central Search Results. FoodData Central. (n.d.). Retrieved October 1, 2021, from
https://fdc.nal.usda.gov/fdc-app.html#/food-details/170464/nutrients.Greshko, M. (2021, May 3). This is the best dinosaur fossil of its kind ever found. Magazine.
Retrieved September 11, 2021, fromhttps://www.nationalgeographic.com/magazine/article/dinosaur-nodosaur-fossil-discover

YHarris, J. D. (1998). A Reanalysis of Acrocanthosaurus atokensis, its Phylogenetic Status, and
Paleobiogeographic Implications, Based on a New Specimen from Texas: Bulletin 13(Vol. 13). New Mexico Museum of Natural History and Science.
Henderson, D. D. (2013, May 13). A one-in-a-billion dinosaur find. The Guardian. RetrievedSeptember 11, 2021, fromhttps://www.theguardian.com/science/lost-worlds/2013/may/13/dinosaurs-fossils
Hummel, J., Gee, C. T., Südekum, K. H., Sander, P. M., Nogge, G., & Clauss, M. (2008). In vitrodigestibility of fern and gymnosperm foliage: implications for sauropod feeding ecologyand diet selection. Proceedings. Biological sciences, 275(1638), 1015–1021.https://doi.org/10.1098/rspb.2007.1728
Stewart, R. (n.d.). U.S. Forest Service. Forest Service Shield. Retrieved October 3, 2021, fromhttps://www.fs.fed.us/wildflowers/plant-of-the-week/athyrium_filix-femina.shtml.
Weaver J.C. The improbable endotherm: the energetics of the sauropod dinosaur Brachiosaurus.Paleobiology. 1983;9:173–182.
Wan Z. 1996. The Lower Cretaceous flora of the Gates Formation from western Canada. PhDthesis, University of Saskatchewan, Saskatoon, Canada. Retrieved fromhttps://harvest.usask.ca/handle/10388/etd-10202004-235747
Withers, P. C. 1992. Comparative Animal Physiology. Saunders College Pub. 949:93-104
AppendixCalculationsWithout fermentationDMR = 7005 kJ/day @ 20oCCarbohydrates
● 11% - 3.7% of which is fiber● Net 7.3% Carbohydrates● .073 * 15.9 kJ/g = 1.1607 kJ/g
Lipids● .0007 * 39.2 kJ/g = .02744 kJ/g
Protein● .0029 * 18.15 kJ/g = .05264 kJ/g
Non-Digestible material● 3.7% fiber
Water● 88.6%
Net energy from ferns● 1.1607 kJ/g + .02744 kJ/g + .05264 kJ/g = 1.24 kJ/g
Food mass assimilated to support DMR of 5766 kJ/day

● 7005 kJ/day / 1.24 kJ/g = 5649 g/day● Mass of Carbs
○ 5649 g/day * .073 = 412.4 g Carbohydrates/day● Mass of Lipids
○ 5649 g/day * .0007 = 3.95 g Lipids/day● Mass of Protein
○ 5649 g/day * .029 = 163.8 g Protein/day● Convert grams to moles
○ 412.4 g / 180 g/mol = 2.3 mol Carbohydrates/day○ 3.95 g / 256 g/mol = .015 mol Lipids/day○ 163.8 g / 135 g/mol = 1.2 mol Protein/day
Food mass ingested to support DMR of 7005 kJ/day● Assume 50% digestive efficiency based on high fiber diet (Weaver 1983)● 5649 g/day / 0.5 = 11298 g/day
Number of food items per day● Average ground fern grows to be about 2ft tall and 1 ft wide with a thickness of 5 mm
(Stewart, 2021)● Volume of a rectangle with these dimensions divided by 2 may accurately describe the
shape of the plant, see figure 1.● V =(l*w*h)/2 = (.6096 m *.3048 m*.0005m)/2● V = 4.645*10^-5 m^3● Since the plant is around 90% water we assume its density to be equivalent to water
(1000 kg/m^3)● 1000kg/m3*(4.645*10^-5)=0.04645● M = 0.04645 kg or 46.45 g● 11298 g/day / 46.45 g/fern = 243.2 full fern fronds
Surface area of the intestine● Limiting nutrient: carbohydrates● Rate of glucose uptake in reptiles 357nmole/min/cm2 at 37oC (Withers 1992)● Daytime body temp: 31.56oC● Q10 correction for glucose uptake: 357(2.5)(31.56-37)/10=216.9nmole/min/cm2 at daytime
body temp● 216.9nmole=0.0000002169mole/min/cm2=0.00031mol/day/cm2
● (1cm2/0.00031mol/day)*2.3 mol Carbohydrates/day=7419cm2
Calculate intestine length and intestine volume● 5cm diameter, SA=pi*d*l● 7419cm2=pi*5*l● Length=472.6cm long=4.73meters
Fermentation model

● DMR=7005 kJ/day● Metabolizable energy by fermentation of tree ferns = 1kg of dry matter = 6,900 kJ ME
(Hummel et al. 2008)Conversion of dry mass to wet mass
● 88.6% water, 11.4% nutrient matter● dry mass=11.4% of wet mass
Energy density of fresh ferns● 0.114kg dry = 1kg wet● 6900kJ * (0.114kg dry/1kg wet)=786.6 kJ/kg fresh fern
Amount of fresh fern eaten per day=11298g/day=11.3kg/day● 11.3kg of fresh fern eaten in a day to meet DMR of 7005kJ with direct digestion● 11.3kg*786.6kJ/kg=8,889kJ● 8889-7005=1884 additional kJ of energy acquired from fermentation with the same
amount of fernsWith only fermentation
● 7005kJ/day / 786.6 kJ/kg fresh fern = only 8.9kg would have to be eaten per day● Reduction of 2.4kg food/day● Food items/day = 8900 g/ 46.45 g/fern = 191.6 fronds
Respective ContributionsAbstract was written by Vanessa. Introduction, Methods, Analysis and Discussion were
written and edited by Allison and Vanessa. Results were written by Allison. Image of thedinosaur was illustrated by Allison. Images of the digestive tract and fern were illustrated byVanessa.