adult frogs derived from the nuclei of single somatic cells · adult frogs derived from the nuclei...
TRANSCRIPT

I)EVELOPhlENI‘AL BIOLOGY, 4,256-273 ( 1962)
Adult Frogs Derived from the Nuclei of Single Somatic Cells
J. B. GURDON~
Beit Memorial Research Fellow, E,mbryology Laboratory, Department of Zoology, Oxford, England
Accepted October 9, 1961
INTRODUCTION
The technique of nuclear transplantation has been used to investi- gate the question whether somatic cell nuclei undergo changes in the course of embryonic differentiation (King and Briggs, 1955; Briggs and King, 1960; Fischberg et al., 1958; Gurdon, 196Oc). Such changes may affect those constituents of a nucleus which are required for embryonic development, and changes of this kind can be discovered by observing the development of nuclear transplant embryos. It is possible that other changes take place in those parts of a nucleus which concern adult characters and reproduction, and it is to expose these (if they exist) that the following survey of adult frogs is pre- sented, each frog having been derived from the development of a single somatic nucIeus and enucleated egg. A description has already been published (Gurdon, 196Oc) of the changes in developmental capacity undergone by nuclei of endoderm cells in the frog Xenopus Zaevis, using donors ranging from blastulae to swimming tadpoles. The frogs described below have been derived from a similar range of donor embryos, and therefore provide complementary results to the former, making a more complete survey of nuclear changes in dif- ferentiating cells.
Particular interest attaches to the reproductive capacity of nuclear transplant frogs, since this provides the most critical test of whether the nuclei from which they are derived are equivalent to germ-cell nuclei in the range of differentiation that they can promote. In Xenopus, and probably in other animals, a special germinal cytoplasm
’ Present address: Division of Biology, California Institute of Technology, Pasadena, California.
256

ADULT E’HOGS FROM SINGLE SOMATIC CELL NUCLEI 257
surrounds the nuclei of all postblastula cells destined to give rise directly to germ cells, but is lacking from all somatic cells (Blackler, 1958). Some of the frogs described below are derived from the nuclei of cells which have been completely free from germ plasm for many nuclear divisions. Thus the lack or possession of fertile gametes by these frogs indicates whether the function of the germ plasm, at present unknown, is to prevent germ-cell nuclei from undergoing changes during development as suggested by Fischberg and Blacklel (1961).
MATERIAL AND METHODS
Donor embryos and recipient eggs were of the subspecies Xenopus laevi.s laeuis. The developmental stage numbers are taken from Nieuwkoop and Faber (1956). Ovulation and mating were induced by the injection of gonadotropic hormone.
The donor cells used for transplantation were taken from the vege- tal hemisphere of blastulae, and from the floor of the gut in later stages. The germ cells are present in the endoderm of early stages, but migrate dorsally and in hatched and swimming tadpoles are situated in the region of the endoderm which is lateral and dorsal to the gut lumen. Since donor cells were never taken from this region, the frogs derived from hatched and swimming tadpoles could not have been derived from germ-cell nuclei.
The nuclear transplantation technique which was followed has been described elsewhere (Elsdale et al., 1960). The great majority of transplant frogs described below carried a marker (Elsdale et al., 1960) in their nuclei, proving that they were derived from the trans- planted nucleus. The presence of the marker was determined from tail tips of growing tadpoles, and it could always be confirmed later in preparations of adult skin or internal organs.
Tadpoles and frogs were reared at approximately 21°C; tadpoles were fed on an infusion of powdered nettles, and frogs were fed when young on Tubifex worms and later on ox liver as well. Under these conditions sexually mature frogs are obtained 9-12 months after fertilization or transplantation.
Controls
A small proportion of the eggs laid, and of the frogs reared, in the laboratory develop abnormally as a result of the unnatural conditions.

258 (XJRDON
It is therefore essential to compare transplant frogs with control frogs from fertilized eggs, which have been reared under the same labora- tory conditions. Only those abnormalities observed in transplant frogs, but not shared by their controls, can be regarded as representing changes undergone by the donor nucleus during the development of the embryo from which it was taken.
RESULTS
Normal Frogs
Many transplant frogs are normal in size, morphology, and repro- ductive capacity. This is true of 76% of those derived from early donor nuclei (stages S-30), but of only 37% of those from advanced donor nuclei (stages 3341) . Thus transplant frogs derived from nuclei of differentiating cells are more often abnormal than those from em- bryonic cell nuclei. Since it is desirable to know whether there is any correlation between the abnormalities of transplant frogs and the origin of the nuclei from which they are derived, the abnormalities of 152 frogs obtained in the course of nuclear transplantation experi- ments are recorded in Table 1 and are discussed separately below. The figures in parentheses in the table show the total number of frogs obtained from the nuclei of any one donor embryo. Thus 2 (3) means that three frogs were derived from the same donor and that two of these showed the abnormality concerned. When the same abnormality is shown by all frogs from one donor, the abnormality is referred to as donor specific.
Adult Size and Skeletal Abnormalities
A few transplant frogs and controls grow exceptionally slowly (Table 1, column 5), but eventually reach the usual adult size and are normal in fertility. A very small proportion of frogs (Table 1, column 4) virtually cease to grow a few months after metamorphosis; these “dwarfs” remain in a juvenile condition, with undifferentiated gonads and color pattern.
Two kinds of gross physical abnormalities have been observed. One type (Table 1, column 6) consists of a deformed backbone, giving the frog a hunchback appearance and causing a corresponding displace- ment of internal organs. However, frogs of this kind are normal in other respects including that of reproductive capacity. The other type

TABL
E 1
Sunl
hlAR
Y O
F AB
XOR
NAL
ITIE
S .~
RIO
NG
I\I)U
I,T
&ocs
D
ERIV
EI)
FRO
.\I
bmoI
)ER
hl
OR
G
UT
CEL
L N
ITC
LEI"
1.
Orig
iu o
f don
or n
ucle
i (s
tage
s fro
m N
icuw
koop
an
d Fa
ber,
1056
)
N”ii
bW
?.
4.
j. 6.
7.
s.
9.
10
. K
umb~
r Tu
mor
s O
ne-y
ear-o
ld f
rogs
R
epro
durti
ve
abno
rmal
ities
of
of
do
nors
fro
gs
r)war
f R
etar
ded
Sk~l
PtZl
l R
ome
Ster
ile d
ut:
in g
row
th
nhno
rmal
ities
ga
met
es
to a
rrest
fd
!y
nf g
wm
-cel
l R
ZXk
Jnw
dc
velo
ued
deve
lorm
x~lt
Vege
tal
hem
isph
ere
and
endo
derm
ce
ll nu
clei
(S
t. 8-
30;
blas
tula
e to
he
artb
eat
tadp
oles
) 21
45
O
S
10)
27;
3(3)
6’i;r
i
2 (3
)
4%
l(l)
a(6)
2(
3)
Nor
mal
-look
ing
3 (3
) eg
gs,
bllt
all
- 12
”’ /( in
ferti
le
l(3)
l(1)
l(4)
4(5)
16
) 10
3)
18
80
2(2)
8.3
z(6)
2(
G)
17;
20; /C
lo
?:,
5 $;
Endo
derm
an
d gu
t ce
ll 3(
3)
2(3)
10
) nu
clei
(S
t. 33
-41;
Ab
norm
ally
fe
w
l(3)
hatc
hed
and
swim
min
g 12
21
eg
gs;
som
e ta
dpol
es)
OS
norm
al
1 (I)
14:1
l(1)
l(l)
l(1)
5 6
2(2)
C
TS
157;
1(4?
1
l(l)
1
2tiC
c
ll5%
__
- - I
l(l) l(2)
1
D Th
e fig
ures
in
co
lum
ns
4-10
in
dica
te
the
num
ber
of f
rogs
sh
owin
g th
e ab
norm
ality
co
ncer
ned.
Fi
gure
s in
pa
rent
hesc
bs
show
th
e to
tal
num
ber
of
frogs
ob
tain
ed
from
nu
clei
of
on
e do
nor
embr
yo;
thus
fro
gs
from
di
ffere
nt
dono
rs
arc
rccs
orde
d se
para
tely
w
ithin
ea
ch
grou
p.
The
125
frogs
fro
m
stag
e 8-
30
dono
rs
wer
e de
rived
fro
m
633
tota
l tra
nspl
anta
tions
(o
f w
hich
th
ey
repr
esen
t 20
(/h),
and
from
22
1 re
gula
r bl
astu
lae
(of
whi
ch
they
re
pres
ent
5, -‘j;).
Th
e 2T
fro
gs
from
st
age
33-4
1 nu
rlri
wer
e de
rived
fro
m
668
tota
l tra
ns-
plan
tatio
ns
and
from
13
6 re
gula
r bl
astu
lar,
of
whi
ch
they
co
nstit
ute
4yG
and
2O
’j;,
resp
ectiv
rly.

260 GURDON
of abnormality consists of a projecting lower jaw, usually accom- panied by a blister on its apex. This abnormality (Table 1, column 7) may interfere with feeding but is not regularly associated with any other physical or reproductive incapacity.
Apart from dwarfism, the abnormalities described above are to some extent donor specific, They are found with a high frequency among frogs of certain genetic constitutions. When recipient eggs from one frog are used for many nuclear transfers, these abnormalities may occur among the frogs from one donor embryo but be entirely absent from those of other donors; they may be found also among the majority of frogs from the fertilized eggs of one parentage, but among none of those from others. On the other hand, by no means all frogs of a similar or identical genetic constitution show the abnor- mality, a fact suggesting an environmental influence on the expres- sion of this abnormality, Whatever the explanation of these abnor- malities, they cannot be consistently associated with frogs derived from transplanted nuclei, since they are found as frequently among frogs from fertilized eggs.
Color Pattern
There is a very considerable variation in color pattern among in- dividuals of the subspecies Xenopus Eaevis laeuis. Pattern is appar- ently genetically controlled, since all transplant frogs derived from the nuclei of the same donor embryo are almost identical in pattern, but are consistently different in this respect from frogs derived from another donor embryo. When two frogs having different patterns are mated together there is a great variation in the patterns of their progeny. These observations indicate that many genes are concerned in the expression of pattern. If this is so, the complete lack of varia- tion in pattern among transplant frogs derived from the nuclei of one donor embryo shows that, in respect of these genes at least, somatic nuclei do not differ qualitatively from each other.
Sex Ratio
The sex ratio of frogs reared from fertilized eggs under laboratory conditions does not differ from equality (111 females and 117 males among I.4 matings). It has been found, without exception, that trans- plant frogs derived from embyonic cell nuclei of any one donor are of the same sex. This indicates that the sex of a donor embryo may

ADULT FROGS FROM SINGLE SOMATIC CELL NUCLEI 261
be deduced from that of the transplant frogs derived from it. The number of female-producing donors should therefore equal the num- ber of male-producing ones. Table 1, column 2, shows that the pro- portion of female- and male-producing donors between stages 8 and 30 is effectively equal, as would be expected. However, among frogs derived from nuclei of stages 3341 there is an excess of female donors (Table l), and most of the male-producing donors gave rise only to sterile frogs. This departure from equality in the number of female- and of male-producing donors suggests the possibility that endoderm nuclei from aclvuncect &nor stages are unable to give rise to normal male differentiation, so that sex reversal occurs. However, too few adult frogs have so far been obtained from such tadpole nuclei to make it certain that they differentiate as females significantly more commonly than as males.
Reproductive Capacity
The fertility of frogs reared in the laboratory is very variable. Al- though in many matings, over 90% of eggs are fertile, and of these only 2-3% develop abnormally, it sometimes happens that healthy and normal frogs lay eggs of which most are infertile, and of those that are fertilized, up to 30% may develop abnormally. It is also found that fertility, and the normality of developing tadpoles, may vary greatly from one mating to the next of the same frogs. A comparison of frogs reared under natural as opposed to laboratory conditions supports the belief that low fertility and abnormal development of offspring are largely attributable to unknown deficiencies of the laboratory conditions and to the artificial induction of ovulation by hormone injection.
Each transplant frog has been mated to different frogs not derived from transplanted nuclei. By an appropriate series of matings, it would be possible to make a genetic analysis of the nuclei of a transplant frog, and so discover any recessive gene changes in the transplanted nucleus. However the results of such matings will not be described in this article, which is concerned with the capacity of a transplant frog to give normal offspring when mated to a wild-type frog, and hence with the ability of the transplanted nucleus to give rise to normal adult cell types including germ cells.
Male and female transplant frogs which have given over 30% fertile eggs of which over 70% develop normally are regarded as normal in

262 GURDON
reproductive capacity, since frogs reared in the laboratory from fer- tilized eggs may not give better results. Transplant frogs which give poor results on one occasion may prove normal in fertility in subse- quent matings. Only those frogs which fail to prove fertile and to give normal offspring to the extent described above, on at least three occasions, are recorded as abnormal in Table 1 (columns 8 and 9) and are discussed below. The remaining frogs which are normal in reproductive capacity show that the somatic nuclei from which they are derived have not undergone any changes in respect of genes required for normal gametogenesis.
Description of reproductive abnormalities. The only abnormality found among 125 frogs derived from nuclei of embryos up to the heartbeat stage concerns 6 females from a single blastula donor. Two of these females gave normal eggs, but the other 4 gave normal- looking eggs which were nevertheless entirely infertile; each of the 4 females was mated to a normal male several times, and also nuclei were transferred to their eggs, but in every case not one egg began to cleave. This type of abnormality differs from those described be- low, since it did not interfere with the growth and differentiation of the gametes in any apparent way. The germ cells were not deficient in numbers nor inviable during oogenesis.
Among the frogs derived from advanced donor stages there were five females and four males with reproductive abnormalities. Three females were derived from the same stage 33 donor. Two of these regularly laid exceptionally few eggs (approximately 500), which developed normally; they were presumably deficient in the number of their germ cells, since no dying oocytes were found in their ovaries. The third female from this donor was entirely lacking any gonads, though oviducts were present. Its rudimentary genital ridges had
FIGS. 1A and 1B. Normal testis of a frog derived from the transplantation of a late gastrula endoderm nucleus. The dense patches are clumps of mature sperm.
FIGS. 2A and 2B. Sterile testis of a transplant frog from a gut-cell nucleus of a swimming tadpole (stage 40). Nests of primary spermatocytes are normal (as in Fig. lB), but later stages in spermatogenesis are seen as pycnotic nuclei (small spheres) or abnormal sperm.
FIGS. 3A and 3B. Sterile testis of a transplant frog from a gut-cell nucleus of a swimming tadpole (stage 41). Nests of primary spermatocytes are normal, but nuclei become pycnotic and disappear during or after the second meiotic di- vision.

ADULT FROGS FROM SINGLE SOMATIC CELL NUCLEI 263

264 GURDON
differentiated as fat body, and its germ cells must have never existed or have died at a very early stage.
A female derived from a stage 40 donor, had entirely sterile ovaries; in this case (Figs. 5A and 5B), oocytes became arrested and died just before oocyte growth began. A very few oocytes (approximately 0.1%) began growth, but none reached the stage of yolk formation.
Another female, from stage 33, was killed when 1 year old and was found to have very immature ovaries (Figs. 6A and 6B) ; these con- tained many oocytes, all of which had abnormal germinal vesicles with large “nucleoli” often attached in threads (Fig. 6B). None of the large oocytes had begun yolk formation, but since many had reached this stage, it seems that they may all have suffered from an abnormality preventing yolk formation and resulting in arrest. Some oocytes were observed to be dying at earlier stages, This frog con- tained groups of abnormal loose cells in the ovary (Fig. 6B), and also contained many abnormal cells in its blood suggesting leukemia; it is conceivable that the arrest and lethality of the oocytes could be a result of the abnormal cells in the blood and ovaries of this individual.
All four male frogs derived from the stage 37 or older donors were sterile. Figures 2A and 2B show sections of the testis of a frog from a stage 40 donor. Another frog from the same donor had testes of similar histological appearance. Although a few apparently normal sperm can be seen, both these frogs were entirely sterile when mated several times to normal females. Figures 2A and 2B show that their testes are normal up to the first meiotic division. During or soon after this division the majority of spermatocyte nuclei become pycnotic and
FIGS. 4A and 4B. Sterile testis of a transplant frog from a gut-cell nucleus of a hatched tadpole ( stage 39 ) Germ cells are entirely lacking from this gonad, which mainly consists of interstitial cells (Fig. 4B), for the most part arranged in tubules (Fig. 4A).
FIGS. 5A and 5B. Sterile ovary of a transplant frog from a gut-cell nucleus of a swimming tadpole (stage 40). Normal nests of premeiotic nuclei can be seen ( Fig, 5A), but at this stage nuclei become pycnotic ( Fig. 5B). A very few oocytes (not shown) began, but did not complete, the growth phase.
FIGS. 6A and 6B. Sterile ovary of a transplant-frog from an endoderm nucleus of a hatching tadpole (stage 33). Oocyte growth is apparently arrested before yolk formation (as in Fig. 6A), but oocytes may die at any time up to this stage (dark blobs in Fig. 6A). The “nucleoli” of oocyte germinal vesicles are abnormally large and are often retained on the chromosome threads (Fig. 6B), though normally free at this stage. The groups of abnormal, possibly cancerous, cell nuclei (Fig. 6B) are probably derived from ovarian follicle cells.
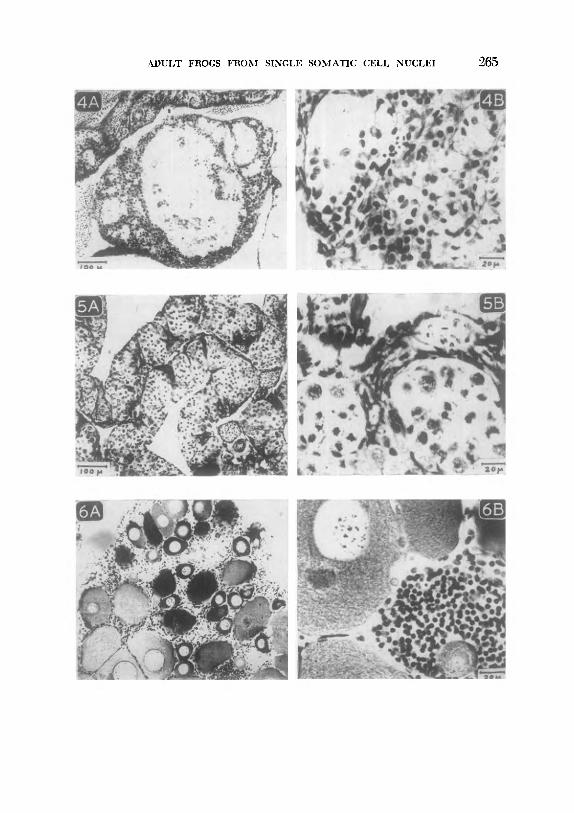
ADULT FROGS FROM SINGLE SOMATIC CELL NUCLEI 265

266 (:URLloN
accumulate in the lumen of the tubules. A frog derived from a stage 13 donor and from the same recipient eggs as the above frogs was normally fertile; sections of its testes are shown in Figs. 1A and 1B for comparison. It is usual to see occasional pycnotic nuclei among large numbers of normal spermatozoa.
A frog derived from a stage 41 donor had a macroscopically normal testis on one side, but had an oviduct and no gonad at all on the other side. From sections of the testis (Figs. 3A and 3B), it can be seen that no normal sperm has been formed, and that the majority of spermatocytes became abnormal and died during or after their first meiotic division.
A frog derived from stage 39 had no oviducts or macroscopically visible gonads. When sectioned, the genital ridges (Figs. 4A and 4B ) were found to consist of interstitial cells partially organized in the form of tubules, but entirely lacking any germ cells. It is probable that the germ cells died during larval growth, since “testes” of this kind are found in animals in which 0-nucleolated (lethal) germ cells have been transplanted (Blackler and Fischberg, personal com- munication) .
Tumors
Four cases of tumors affecting various internal organs were ob- served among transplant frogs from advanced donor stages, but none among those from early donors. Two out of 15 tetraploid transplant frogs, not shown in the table, were also found to suffer from cancer. The incidence of cancers among experimental and control frogs will be considered elsewhere (Balls, unpublished).
Serial Transplant Frogs
When an originally transplanted nucleus gives rise to a blastula whose nuclei are used for second transplantations, the frogs resulting from these second (or subsequent) transfers are called serial trans- plant frogs. Serial transplant frogs derived from a blastula transplant embryo should be genetically identical to each other, and therefore any differences between them must be due to an initially cytoplasmic or environmental cause. Since the abnormalities recorded in columns 4-7 of Table 1 were found in only a few individuals among several genetically identical frogs from the same donor, this provides strong support for the conclusion arrived at above, that these abnormalities

‘I’AB
LE
2 AB
SOR
~IAL
ITIE
S A~
ION
G
SER
IAL
TRAN
SFER
FR
OG
S
1.
DO
WXS
” 2.
Sr
tmbe
r an
d Bt
ngf
of
dono
rs
3.
N”:fb
prr
frogs
8.
0.
10.
Rep
rodu
ctive
ab
norm
nlitie
s Tu
mor
s
Som
e St
erile
du
e ga
met
es
to
BTF&
fu
lly
of
germ
-cel
l de
velo
ped
devc
lopm
eut
Don
or
embr
yos
12 0
an
d 2
1 1
1 -
-
(con
trols
) lO
C?
9%
57;
5 C
y’
0 5’
A,
lst-t
rans
fer
frogs
4
26
1 3
4 -
-
from
ab
ove
Blas
ttlla
e 47
; 12
7;:
165;
~.
____
____
Seria
l tra
nsfe
r 5
frogs
fro
m
abov
e I%
&ula
e
0 Fo
ur
blas
tula
e fro
m
the
orig
inal
do
nor
embr
yos
gave
ris
e,
afte
r tra
nspl
anta
tion
of
thei
r nu
clei
, to
th
e fir
st-tr
ansf
er
frogs
. Fi
ve
blas
tula
e w
ere
take
n fro
m
the
first
-tran
sfw
rm
bryo
s an
d ga
ve
rise
aftrr
tra
nspl
anta
tion
to
the
seria
l tra
nsfe
r fro
gs.

268 CURDON
of growth and skeleton are not genetic, but are the result of labora- tory conditions.
Any consistent difference between first-transfer frogs and serial transfer frogs from the same original donor embryo might be re- garded as direct consequences of serial transplantation. In fact no such differences were found (Table 2), thus supporting the belief that serial transplantation does not cause cumulative damage to nuclei.
Further details and photographs of serial transplant frogs have been published elsewhere (Gurdon, 1961, 1962); the latter are com- pared with first-transfer frogs and frogs reared directly from sibs of the donor embryos themselves.
DISCUSSION AND CONCLUSIONS
The Significance of Abnormal Transplant Frogs
Many transplant frogs from nuclei of all donor stages are normal. Of the abnormalities from which the remainder suffered, retarded growth and skeletal malformations are found with a similar frequency in frogs from fertilized eggs, and so can be safely attributed to general causes, not to specific properties of transplanted nuclei. Tumors have been found more frequently among frogs derived from nuclei of ad- vanced rather than early donor stages, but at present too few cases have been found to support a causal connection between tumor susceptibility and the origin of frogs from advanced donor nuclei. The sterility from which transplant frogs suffer is of various kinds and appears to be caused by different factors. Only four out of 125 frogs from early donor nuclei were sterile. These were all derived from one blastula donor and suffered from a peculiar form of sterility (page 262), which was exactly the same in each frog. This kind of sterility is not characteristic of blastula nuclei in general and was probably due to an abnormal condition of some of the nuclei of the particular blastula donor used in this case. Altogether 9 frogs from advanced donor nuclei suffered from reproductive abnormalities. Since two of these were deficient only in the number of germ cells and contained some normal gametes, their abnormality cannot be attributed to a condition of the original transplanted nucleus, from which their normal gametes must have been derived. The remaining seven cases of sterility will be considered in more detail, since they

AIXJLT FROGS FROM SINGLE SOMATIC CELL NUCLEI 269
could be due to a natural condition of the transplanted nuclei acquired during cellular differentiation,
Of these seven frogs, two entirely lacked germ cells, which may never have existed or may have been eliminated during tadpole stages. In the other five cases the majority of germ cells became arrested at different stages during gametogenesis; however, a very few germ cells passed these lethal stages, and some formed morpho- logically, though not functionally, normal gametes. Thus there ap- peared to be no specific abnormality, in these cases, that brought about the arrest and death of germ cells.
There are many possible causes for sterility. An abnormality or deficiency of germ plasm, as often occurs in overmature eggs, is be- lieved to lead to sterility ( Witschi, 1952). However, this is unlikely to explain the present cases, since parallel transplantations of early and advanced donor nuclei into eggs from one female have shown that all the frogs from early donors were fertile, while sterile frogs occurred only among those derived from advanced donors. The in- frequent occurrence of sterile frogs among controls reared directly from fertilized eggs indicates that all cases of sterile transplant frogs cannot be attributed to sterility of the particular donor embryos from which they were derived. The question of technical damage to trans- planted nuclei has been investigated (Gurdon, 1960b); it was found that a twofold increase in the length of exposure of cells to dis- aggregating and storage saline solutions did not give any consequent increase in developmental abnormalities of transplant embryos. It seems unlikely that the nuclear constituents concerned with fertility should be damaged by the technique while those concerned with development are not. However, on these grounds alone technical damage cannot be entirely excluded as a cause for sterility.
A possible explanation of sterile transplant frogs is that the nuclei from which they were derived have undergone cytoplasmically in- duced changes during the development of the donor embryo. This hypothesis may be related to certain information about the germ plasm in Xenopus (Blackler, 1958). Primordial germ cells are situated in the endoderm from the late blastula until the swimming tadpole stage, when they migrate into the genital ridges. During this time their nuclei are surrounded by germ plasm. Germ cells do not divide while in the endoderm but commence division when they reach the genital ridges, at which time the germ plasm undergoes a change

(XJRDON
in staining properties. There is therefore a remarkable correlation between the association of germ-cell nuclei with germ plasm and their lack of division during this time. Thus the main function of the germ plasm is evidently to inhibit germ-cell division. It may be im- portant for the germ-cell nuclei to be protected from the endoderm cytoplasm (Fischberg and Blackler, 1961)) since this might induce changes in them as it appears to do in endoderm nuclei (Gurdon, 196Oc). In the case of gall midges, it has been demonstrated that an association with germ plasm is essential to prevent the germ-line nuclei from undergoing the same changes (loss of chromatin) as the somatic nuclei ( Geyer-Duszynska, 1959; Bantock, 1961). In Xenopus the germ plasm may also inhibit division in order to prevent the germ cells from proliferating before they have reached the germ ridges, which are not formed until the swimming tadpole stage.
The results reported above are consistent with this general hy- pothesis. The sterile transplant frogs could thus be regarded as show- ing the kind of way in which endoderm nuclei may change, and one of the functions of the germ plasm may be to prevent germ-cell nuclei from changing in this way too. On the other hand, the fact that the majority of all tested nuclei from advanced donor stages have not undergone such changes, and that the abnormalities ob- served differ from one case to the next, indicates that these changes are unspecific and are not causally connected with the differentiation of endoderm cells. However, until it is possible to test a greater number of gut-cell nuclei of swimming tadpoles for their ability to give rise to fertile frogs, the real significance of the sterile frogs described above must remain uncertain.
The Significance of Normal Transplant Frogs
None of the 125 frogs from early donor embryos indicate that any natural changes acquired during cellular differentiation have taken place in the nuclei from which they are derived. The abnormalities of these frogs can be attributed to other factors. Of the 27 frogs from advanced donors, 9 suffered from abnormalities that might be due to natural changes in the transplanted nuclei (5 from sterility, 2 from sterility and tumors, and 2 from tumors only), while the re- maining 18 (67%) were either normal or suffered from abnormalities attributable to other causes. These results indicate that, of the nuclei

ADULT FROGS FHOM SINGLE SOMATIC CELL NUCLEI 271
trom which frogs were obtained, all those from early donors and two- thirds of those from advanced donors contain all genes required for the formation and functioning of normal adult frogs.
The possibility has been pointed out (Gurdon, 196Oc; Briggs and King, 1960) that a reversal of the properties of nuclei could take place as a result of transplantation in Xenopus eggs. A nucleus from one species of Xenopus, after synthesis from the egg cytoplasm of another species, undergoes a genetic change which limits the range of genetic information that it can impart. The serial transplantation of nuclei between two species of Xenopus has shown that the nuclei do not undergo any reversal of this change (Gurdon, in press). Thus the range of genetic information contained in transplanted nuclei has apparently not been acquired as a result of their transplantation, but was contained in these nuclei when they were part of the original donor cells. It is worth recalling here that all transplant frogs from hatched and swimming tadpole donors must have been derived from endoderm nuclei, not from germ-cell nuclei (page 257).
It must be remembered that by no means all transplanted nuclei give rise to adult frogs. The proportion of total transplantations which do so in Xenopus is about 30% for blastula nuclei, 10% for stage 30 nuclei, 6% for hatched tadpole nuclei, and 3% for stage 3941 swim- ming tadpole nuclei (Gurdon, 196Oc). Since all nuclei from early donor stages and two-thirds of those from advanced donors are ap- parently capable of giving rise to normal frogs, it is possible to cal- culate the minimum percentage of endoderm cell nuclei with a com- plete range of genetic information required for the formation of a nor- mal individual: 30% of blastula cells; 10% of stage 30 cells; 4% (ys of 6%) of hatched tadpole cells, and 2% of swimming tadpole cells. It is stressed that these results show the minimum proportion of cells whose nuclei are capable of giving rise to normal frogs. It is not cer- tain that those nuclei which fail to give rise to normal frogs are genetically changed in any way. There is in fact some evidence (Gurdon, in press) that those transplanted nuclei which do not give rise to regular blastulae are not effectively tested for their develop- mental capacity. If this is so, the nuclei which have given rise to adult frogs should be considered as a proportion of blastula transplant embryos and not of total transplantations. About 20% of the blastulae from nuclei of hatched and swimming-tadpole gut cells give rise to

272 GURDON
adult frogs (Gurdon, 196Oc), and hence, if the above supposition is correct, at least 13% (51 of 20%) of such nuclei contain all the genetic information required for the formation of a normal frog.
The existence of a normal adult individual capable of normal re- production constitutes proof that the zygote or other nucleus, from which it was derived, contains a full complement of those genes required for the formation and functioning of a normal individual. The results described here therefore show that some swimming-tad- pole gut cells contain nuclei which are, to this extent, genetically un- changed from the zygote condition, The question whether those gut- cell nuclei which did not give rise to frogs are genetically changed, cannot yet be answered.
SUMMARY
A survey has been made of over 150 adult frogs which have been obtained by the transplantation of nuclei from endoderm cells of Xenopus Levis donors ranging from late blastulae to swimming tad- poles.
Many of the transplant-frogs from donors of all ages were entirely normal, and many of the abnormalities observed were not connected with the condition of the transplanted nuclei. Frogs derived from the nuclei of differentiating cells were more often abnormal than those derived from embryonic cells. Out of 27 frogs derived from nuclei of hatched tadpole gut cells, 7 were sterile.
The results demonstrate that at least 30% of blastula nuclei and at least 4% of hatched tadpole gut-cell nuclei contain a complete range of the genetic information necessary for the formation and functioning of a normal adult individual.
The author is very grateful to Dr. M. Fischberg and Dr. A. W. Blackler for their help in the interpretation of gonad abnormalities; also to the latter and to Dr. R. Briggs for their comments on this manuscript. He also wishes to thank Mrs. A. Gibson for her care of the frogs required for this work.
REFERENCES
BANTOCK, C. ( 1961). Chromosome elimination in Cecidomyidae. Nature 190, 466-467.
BLACKLER, A. W. ( 1958). Contribution to the study of germ-cells in the Anura. J. Embryol. Exptl. Morphol. 6, 491-503.
BRIGGS, R., and KING, T. J. (1960). Nuclear transplantation studies on the early gastrula (Rana pipiens). I. Nuclei of presumptive endoderm. Develop. BZoZ. 2, 252270.

ADULT FROGS FROM SINGLE SOMATIC CELL NUCLEI 273
ELSDALE, T. R., GURDON, J. B., and FISCHBERG, M. ( 1960). A description of the technique for nuclear transplantation in Xenopus luevis. .l. Embryol. ExptZ. Morphol. 8, 437444.
FISCHBERG, M., and BLACKLER, A. W. ( 1961). How cells specialize. Sci. American 205, 124-140.
FISCHBERG, M., GURDON, J. B., and ELSDALE, T. R. ( 1958). Nuclear transfer in Amphibia and the problem of the potentialities of the nuclei of differ- entiating tissues. Exptl. Cell Research Suppl. 6, 161-178.
GEYER-DUSZY&SKA, I. ( 1959 ) . Experimental research on chromosome elimination in Cecidomyidae (Diptera). .I. Exptl. Zool. 141, 391-448.
GURDON, J. B. (1960b). Factors responsible for the abnormal development of embryos obtained by nuclear transplantation in Xenopus Zuevis. J. Embryol. Exptl. Morphol. 8, 327-340.
GURDON, J. B. (196Oc). The developmental capacity of nuclei taken from dif- ferentiating endoderm cells of Xenopus Zueois. J. E,mbryoZ. ExptZ. Morphol. 8, 505526.
GURDON, J. B. (1961). The transplantation of nuclei between two subspecies of Xerwpus Zuevis. Heredity 16, 305-315.
GURDON, J. B. ( 1962). Multiple genetically-identical frogs. J. Heredity, in press. CURDON, J. B. (in press). The transplantation of nuclei between two species of
Xenopus. KING, T. J., and BRIGGS, R. (195.5). Changes in the nuclei of differentiating
gastrula cells, as demonstrated by nuclear transplantation. hoc. N&Z. Acad. Sci. U. S. 41, 321-325.
NIEUWKOOP, P. D., and FABER, J. (1956). “Normal Table of Xenopus Laez;is ( Daudin) .” North-Holland Publishing Co., Amsterdam.
WITSCHI, E. ( 1952). Overripeness of the egg as a cause of twinning and terato- genesis: A review. Cancer Research 12, 763-786.