alternate recruitment of srp and trigger factor to the ... filerecognition particle (srp)1, composed...
TRANSCRIPT

ALTERNATE RECRUITMENT OF SIGNAL RECOGNITION PARTICLE AND TRIGGER FACTOR TO THE SIGNAL SEQUENCE
OF A GROWING NASCENT POLYPEPTIDE* Gottfried Eisner‡, Michael Moser, Ute Schäfer, Konstanze Beck§, and Matthias Müller
From the Institut für Biochemie und Molekularbiologie, Universität Freiburg, D-79104 Freiburg, Germany
Running Title: Alternate Liganding of a Nascent Signal Sequence Address correspondence to: Matthias Müller, Institut für Biochemie und Molekularbiologie, Hermann-Herder-Strasse 7, D-79104 Freiburg, Germany, Tel. 49-761-203-5265; Fax. 49-761-203-5274; E-Mail: [email protected]
Different from cytoplasmic membrane proteins, presecretory proteins of bacteria usually do not require the signal recognition particle (SRP) for targeting to the Sec translocon. Nevertheless signal sequences of presecretory proteins have been found in close proximity to signal recognition particle immediately after they have emerged from the ribosome. We show here that at the ribosome, the molecular environment of a signal sequence depends on the nature of downstream sequence elements that can cause an alternate recruitment of signal recognition particle and the ribosome-associated chaperone Trigger factor to a growing nascent chain. While signal recognition particle and Trigger factor might remain bound to the same ribosome, both ligands are clearly able to displace each other from a nascent chain. The data also imply that a signal sequence owes its molecular environment to the fact that it remains closely apposed to the ribosomal exit site during growth of a nascent secretory protein.
In Escherichia coli, most exported proteins leave the cytoplasm via the SecYEG-translocon of the inner membrane (reviewed in: refs. 1,2). Exported proteins, that remain resident in the cell, encompass periplasmic and outer membrane proteins, collectively termed secretory proteins, as well as inner membrane proteins. Targeting of both groups of proteins to the Sec-translocon occurs by two independent mechanisms. Inner membrane proteins are co-translationally targeted by the bacterial signal recognition particle (SRP)1, composed of the Ffh protein and the 4.5S RNA, in concert with the SRP receptor, FtsY (reviewed in: refs. 3,4). SRP specifically recognizes the signal anchor sequences of inner membrane proteins when they emerge from the ribosome. Experimentally, this has been demonstrated by site-directed cross-linking of Ffh to the signal anchor
sequences of nascent inner membrane proteins (5-7). In contrast, secretory proteins do not seem to undergo a specific recognition event at the ribosome but rather are post-translationally targeted to the Sec-translocon by SecA and SecB. Accordingly, nascent secretory proteins are not found cross-linked to Ffh but, like most nascent polypeptides, to the ribosome-associated chaperone Trigger factor (TF) (5-8).
From the results of a number of experiments it has been concluded that it is the low overall hydrophobicity characteristic of cleavable signal sequences of secretory proteins, that excludes an interaction with SRP (5,9-12). Surprisingly, however, nascent secretory proteins exposing only the signal sequence from the ribosome have been found cross-linked to Ffh (13,14). Only upon further growth of the nascent chain did the signal sequence move from a close contact with Ffh to one with TF. For the outer membrane protein A (OmpA), this transition was determined to occur, after 37 to 53 amino acids downstream of the signal sequence have exited the ribosome (13). A length-dependent change in the contact of a nascent signal sequence from Ffh to TF relates to the controversial issue if Ffh and TF can bind to the same ribosome at the same time. This question was raised by the disclosure of partially overlapping binding sites for both proteins at the ribosomal proteins L23 and L29 (15-21) and by studies directly addressing a competitive binding behaviour (22-24).
The finding that the vicinity of a signal sequence changes from Ffh to TF upon growth of the nascent chain might be explained by a change in the over-all affinity of the nascent chain for the two ligands. In the case of a secretory protein like pOmpA, high affinity binding sites for TF following downstream of the signal sequence would recruit TF, which thereby sequesters the adjacent signal sequence from contacts with Ffh. We therefore asked if by the same token, a downstream recognition site
1
http://www.jbc.org/cgi/doi/10.1074/jbc.M511388200The latest version is at JBC Papers in Press. Published on January 18, 2006 as Manuscript M511388200
Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

for Ffh being exposed upon continued growth of the nascent chain would have an influence on the liganding behaviour of a signal sequence. Our results show that distal sequence elements of the nascent chain in fact determine the molecular vicinity of a signal sequence and that Ffh and TF compete for binding to nascent secretory and membrane proteins.
Experimental Procedures
Strains and plasmids - The following E.
coli strains were used, MRE 600 (25), SL119 (26), C600 (Δtig) (27), and LGS322 (ΔmtlA) (28). Plasmids used were vector pKSM717 (29), p717MtlA-B expressing mannitol permease (MtlA) (6), and p717OmpA-V11 encoding pOmpA with a TAG stop codon at residue Val-11 (13).
The gene encoding the hybrid protein Omtl was constructed from plasmids p717OmpA (6,13) and p717MtlA-B. Plasmid p717OmpA was linearized at an EcoRI site located at the 3'-end of ompA and subsequently cut with SpeI to remove the ompA sequence except for the nucleotides encoding the first 26 residues including the signal sequence. This fragment was ligated onto a SpeI – EcoRI fragment of p717MtlA-Spe2 (12) encoding MtlA except for the first 46 residues. The resulting plasmid is p717Omtl. Likewise, p717Omtl-V11 harbouring a stop codon in the signal sequence was constructed using p717OmpA-V11 and p717MtlA-Spe2. To construct plasmid p717Omtl-I36 carrying a stop codon in TM2 of pOmtl at residue Ile-36, plasmid p717Omtl was cut with SpeI and HincII and ligated onto a SpeI - HincII fragment which was obtained using p717MtlA-I56 (30) as template and the primers 5’- CCGAACGACACTAGTGCAGAAGCTGG -3’ (the SpeI-l cleavage site is underlined) and 5’-ACCGTGGTTAATGGCGTTGTT-3’. All constructs made were verified by DNA sequence analysis.
In vitro synthesis - The composition of the reconstituted E. coli transcription/translation system and the purification of its components has been described previously (13). Oligodeoxynucleotides used for the synthesis of elongation-arrested nascent chains of pOmpA-50 and pOmpA-66 have been published (13). Nascent chains of pOmtl-82, pOmtl-110, pOmtl-169 were obtained using the same primers as those previously described for the synthesis of MtlA-102, MtlA-130, and MtlA-189, respectively (6,30). Elongation-arrested chains
of pOmtl-65 and pOmtl-140 were generated by 5’-TGCGCCGACGATAACGCCCAT-3’ (4 μg/25 μl reaction) and by 5’-GAAGTTAACGCCCGCAGCCAG -3’ (3 μg/25 μl reaction), respectively. If not stated otherwise, synthesis reactions were terminated after 30 min at 37°C.
Chemical cross-linking using DSS, introduction of TAG stop codons by site-directed mutagenesis, incorporation of the site-specific cross-linker Tmd-Phe, and immunoprecipitation on 4-fold scaled-up reactions were performed as described (6,13). Translation products were resolved by SDS-PAGE using either linear 10% (Fig. 1D) and 15% (Fig. 1 B,C) acrylamide gels or, in the case of cross-linking reactions, 9-20% (Fig. 3) and 7-20% (Figs. 4,5) gradient gels, and radiolabeled proteins were visualized by phosphorimaging using a Molecular Dynamics PhosphorImager and quantified using Imagequant software (Molecular Dynamics). The preparation of INV as well as the protease protection assay have been described previously (31,32).
Miscellaneous - To express MtlA and pOmtl, the E. coli strain LGS322 (ΔmtlA) was transformed with plasmids p717MtlA-B or p717Omtl, respectively. Control cells were transformed with the vector plasmid pKSM717. Cells were grown to mid logarithmic phase in LB medium containing 50 µg/ml ampicillin and expression from plasmids was induced by adding isopropyl thio-β-D-galactoside at a final concentration of 1 mM. After an additional hour of growth, cells were harvested and a crude protein extract was obtained by sonication and low speed centrifugation (15 min, 5 000 x g) to remove debris. Extracts were further separated into soluble supernatant and membrane pellet by high speed centrifugation (1 hour, 70 000 rpm, Beckman rotor TLA100.2).
Ffh (13) and TF (14) were purified and polyclonal antibodies were raised against the purified proteins (14,32) as described. Western-blots were developed using HPR-conjugated goat anti-rabbit antibody, followed by enhanced chemiluminescence (Amersham).
RESULTS
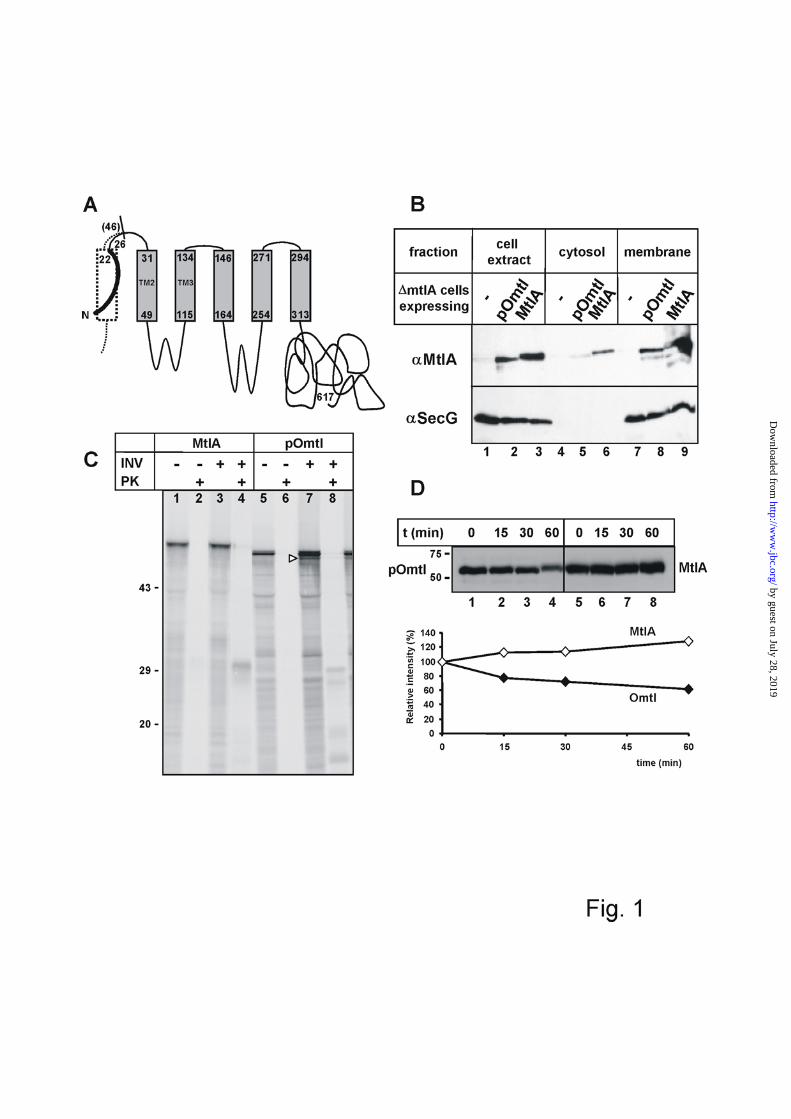
Correct localization of a polytopic membrane protein harbouring a cleavable signal sequence at its N-terminus - In order to construct a polypeptide in which a signal sequence is followed by an Ffh recognition site, i.e. a signal anchor sequence, we replaced the first 46 amino
2
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

acid residues of the inner membrane protein mannitol permease (MtlA) by the signal sequence plus the four subsequent residues of pOmpA creating the fusion protein pOmtl (Fig. 1A). MtlA has six to eight predicted transmembrane spans (TMs) with the N and the C-termini located in the cytoplasm (33,34). Despite the substitution of the first TM of MtlA for the signal sequence of pOmpA, the fusion protein pOmtl was recovered from the membrane fraction of pOmtl over-expressing E. coli cells to about the same extent as MtlA (Fig. 1B; even in wild type E. coli strains up to 20% of MtlA is found soluble following ultracentrifugation of cell extracts (35). Furthermore, when expressed in vitro by a cell-free transcription/translation system, pOmtl integrated into inside-out inner membrane vesicles (INV) as indicated by the accumulation of proteinase K-resistant fragments when INV were present (Fig. 1C, compare lanes 6 and 8). A 30 kDa protease-resistant fragment (lane 4) is diagnostic for membrane integration of MtlA (36). In the case of pOmtl, the appearance of more proteinase K fragments smaller than 30 kDa (compare lanes 4 and 8) are suggestive of a less stable membrane association when compared to MtlA which was also seen in an in vivo time course experiment (Fig. 1D). Consistent with membrane integration, the addition of INV to the in vitro synthesis reaction also generated a smaller species of Omtl most likely due to signal sequence cleavage (Fig. 1C, lane 7, arrow head). Although as expected pOmtl lacking the first TM of MtlA did not assemble into a functional permease (data not shown) it obviously was recognized and at least transiently localized to the membrane by the cell’s targeting machinery.
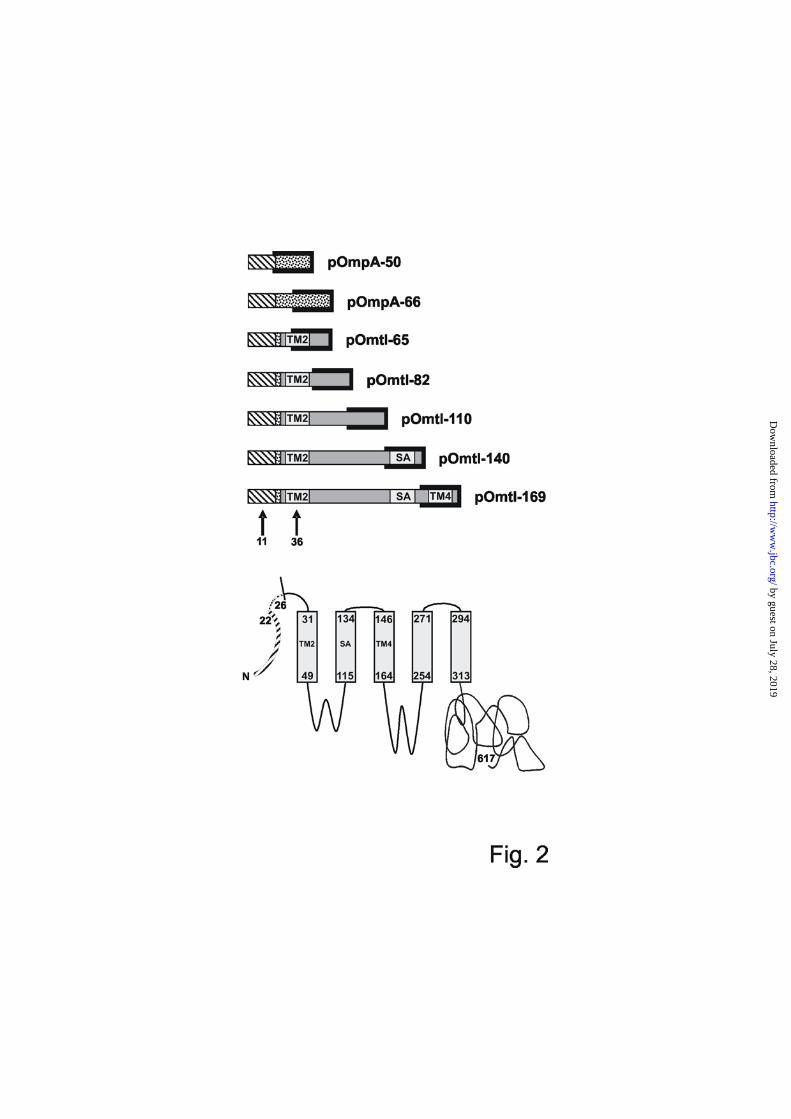
A downstream signal anchor sequence mediates contact of the N-terminal signal sequence with Ffh - Next a series of elongation-arrested chains of pOmtl (Fig. 2) were synthesized in vitro. These nascent pOmtl chains carried a photoactivatable derivative of phenylalanine (Tmd-Phe; L-4’-(3-[trifluoromethyl]-3H-diazirin-3-yl) phenylalanine) either within the hydrophobic core of the pOmpA signal sequence (position 11) or within TM2 of MtlA (position 36; arrows). Fig. 2 also illustrates the various sequence portions of pOmtl being exposed from the ribosomal exit tunnel. Following synthesis in the presence of Ffh and TF, the various radioactively labelled pOmtl nascent chains were irradiated with UV light and the obtained cross-
links were visualized by SDS-PAGE and phosphorimaging (Fig. 3). For comparison two short nascent chains of pOmpA carrying Tmd-Phe at position 11 in the signal sequence were also included (panel A). In the latter cases a prominent photo-adduct of the pOmpA signal sequence was obtained (X) that was immunoprecipitated by anti-Ffh antibodies (Fig. 3A, lanes 3 and 6). When the shortest nascent chain of pOmtl (pOmtl-65) was UV-irradiated (Fig. 3A, lanes 7 – 10) a more discrete Ffh-adduct (X), which was masked by an non further characterized translation product (●), became visible after immunoprecipitation with anti-Ffh antibodies (lane 9). Thus the signal sequence of pOmpA contacts Ffh also when present in short nascent chains of pOmtl. As expected, pOmtl-65 nascent chains carrying the cross-linker at position 36 within TM2 of MtlA, did not show this adduct (Fig. 3B, lanes 1 – 3) since this cross-linking site is predicted to be buried within the ribosomal exit tunnel (cf. Fig. 2). In pOmtl-82 exposing the entire TM2 of MtlA (cf. Fig. 2) the Ffh-adduct of the signal sequence became more prominent (Fig. 3A, lanes 11 – 13, upper panel) whereas immunoprecipitation revealed also some faint contact with TF (asterisk, lane 14). The same ligands were picked up when the cross-linker was placed into TM2 of MtlA, now with a relative increase in number of TF-adducts (Fig. 3B, lanes 4 – 7). Irrespective of the position of Tmd-Phe, nascent chains of pOmtl-110 exposing about 30 amino acids downstream of TM2 of MtlA (cf. Fig. 2) overwhelmingly cross-linked to TF (Fig. 3A, lanes 15 – 18, upper panel and B, lanes 8 – 11). Hence in pOmtl, the signal sequence of pOmpA and the TM2 of MtlA share the same molecular environment as previously observed for the signal sequence of pOmpA (13): they loose contact to Ffh in favour of one with TF when the nascent chain increases in length.
Extension of the pOmtl nascent chain by another 30 amino acids (pOmtl-140) did not change the predominant liganding of TM2 of MtlA to TF (Fig. 3B, cf. lanes 8 – 15), whereas the signal sequence of pOmtl-140 showed about as many contacts with Ffh as with TF (Fig. 3A, lanes 19 – 22). Remarkably, however, as soon as TM3 of MtlA, which by its topogenic information is a signal anchor sequence, became fully exposed in pOmtl-169 nascent chains (cf. Fig. 2), the preceding signal sequence almost exclusively cross-linked to Ffh (Fig. 3A, lanes 23 – 26, upper panel) and also TM2 of MtlA showed relatively many more Ffh-adducts than
3
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

in the shorter pOmtl-140 and pOmtl-110. Obviously the signal sequence regained contact to Ffh upon exposure of a downstream signal anchor sequence. Incidentally, incorporation of Tmd-Phe into TM3 of MtlA had revealed multiple Ffh binding sites (30; and data not shown). The cross-linking data illustrated in Fig. 3 therefore suggest that it is this late signal anchor TM3 that recruits SRP to the nascent chain of pOmtl-169 and that the signal sequence and TM2 must be so close that both resume contact to Ffh. The formation of such a helix bundle-type structure of nascent pOmtl-169 at the ribosomal exit site is supported by data shown in Fig. 3A (lanes 23 - 26): in addition to Ffh, the cross-linker placed in the signal sequence yields a strong adduct of pOmtl-169 with a 8 kDa protein (black star) which is immunoprecipitated with antibodies against ribosomal protein L29 (lower panel). Cross-linking to L29 was not observed for TM2 (Fig. 3B, lanes 16 – 17). On the other hand, also the shorter chains pOmtl-82 and pOmtl-110 were found to cross-link to ribosomal proteins L29 and L23 (Fig. 3A, lanes 11 – 18, black and white stars, respectively). These findings indicate that even after exposure of more than 130 amino acids the signal sequence of nascent pOmtl is not freely mobile but remains closely apposed to the ribosomal exit site.
Ffh and TF compete for the same nascent chains - Amongst the various pOmtl nascent chains analyzed thus far, pOmtl-110 and pOmtl-169 display the most distinct liganding behaviour. When the two species are synthesized in the presence of both Ffh and TF, the signal sequence present in pOmtl-110 picks TF as interaction partner (“TF binder”) and that of pOmtl-169 Ffh (“SRP binder”). Thus the molecular vicinity of the signal sequence in the folding pOmtl nascent chain appears to be determined by the nature of downstream sequence sections. These findings do, however, not allow for predicting the fate of the disfavoured ligand; while it is virtually not found in contact with the nascent pOmtl chain it might still remain bound to the synthesizing ribosome via L23/L29.
In the experiments summarized in Fig. 3, cross-links of a given nascent chain obtained with either of the two positions of Tmd-Phe were not always entirely identical. Thus by comparison with the signal sequence, TM2 showed relatively more contacts with TF than with SRP (cf. pOmtl-82 and pOmtl-140 in panels A and B). While this finding could reflect
a closer location of TM2 to the ribosome-bound TF, it raised the general question as to the influence of the sidedness of the cross-linker on the liganding pattern. In theory, the changing cross-linking partners of a growing pOmtl nascent chain could also be the result of a change in conformation of a folded nascent chain at the ribosomal exit site. In order to render the results less dependent on the orientation of a single cross-linker we employed the NH2-group-specific cross-linker disuccinimidyl suberate (DSS), as the N-terminal part of pOmtl including TM2 contains five lysyl residues. Likewise it seemed to be important to address the influence of various concentrations of Ffh and TF on the cross-linking behaviour of pOmtl nascent chains. In the particular cell-free transcription/translation system used here, the endogenous concentration of Ffh is so low that Ffh-cross-links to MtlA and its SRP-dependent integration into membrane vesicles are only observed if purified Ffh is added (6,32). On the contrary, sufficient depletion of the cell-free system of TF requires preparing its components from a TF knock-out mutant strain.
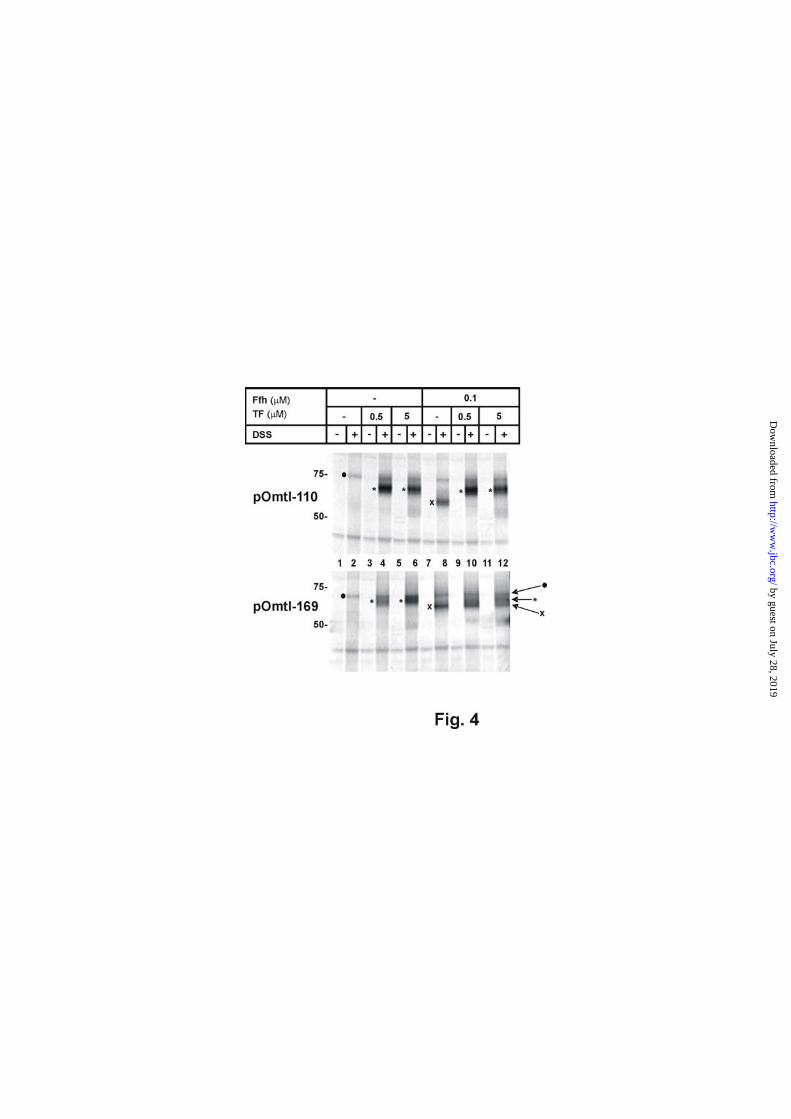
Fig. 4 illustrates the cross-linking results obtained for pOmtl-110 and pOmtl-169 synthesized under these adapted conditions. In the absence of Ffh and TF, DSS did not give rise to any specific adduct (lanes 1 and 2; the ones marked with ● were present under all conditions tested). When synthesized in the presence of 0.5 μM TF, a pronounced cross-linking product of pOmtl-110 with a protein of the size of TF appeared (upper panel, lane 4, asterisk). This concentration of TF had previously been determined to be present in similar cell-free systems (37). Obviously 0.5 μM TF were saturating for the interaction with pOmtl-110, because raising the level of TF to 5 μM did not improve the yield of the cross-linking product (lane 6), although even this concentration of TF amounts to only 10% of its estimated intracellular concentration (38). Different from previous experiments (Fig. 3), in which the concomitant presence of Ffh and TF predominantly led to TF-adducts of pOmtl-110, a clear interaction with Ffh was observed when pOmtl-110 nascent chains were synthesized in the complete absence of TF (lane 8, X). The amount of Ffh sufficient to reveal this interaction (0.1 μM) is the same as that required to see Ffh activity in vitro and furthermore compares fairly well with the estimated intracellular concentration of Ffh. Cross-linking of pOmtl-
4
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

110 to Ffh was, however, completely over-ridden by 0.5 μM TF (lane 10).
Similarly, the SRP binder pOmtl-169 was also able to cross-link to TF, if this was the only ligand exogenously added during synthesis (Fig. 4, lower panel, compare lanes 2 and 4, asterisk). In this case, however, the number of TF-adducts increased if the concentration of TF was raised ten fold (lanes 4 and 6). Again in accordance with the results shown in Fig. 3, the Ffh cross-links of pOmtl-169 (Fig. 4, lane 8, X) largely persisted in the presence of 0.5 μM TF (lane 10). Interestingly at the higher concentration of TF, contacts of pOmtl-169 to both ligands were obtained (lane 12, asterisk and X). Hence the results presented in Fig. 4 clearly indicate that both nascent chains, pOmtl-110 and pOmtl-169, in principle can interact with each ligand, but that there is an obvious, concentration-dependent competition between TF and SRP for their substrates.
TF and Ffh can displace each other from a nascent chain - The experiments presented thus far are consistent with the notion that a pOmtl nascent chain synthesized in the presence of the two ligands TF and SRP recruits the one for which it has exposed high affinity binding sites. These are for SRP the signal anchor-type TM3 - nota bene not the stop-transfer-type TM2 - and for TF obviously areas around TM2 and the subsequent loop of MtlA (cf. Fig. 1A). If in fact downstream domains of the nascent chain ultimately recruit the ligands of a signal sequence, it should be possible to displace a weak ligand by a post-translational addition of a strong one. This has been addressed by the experiments shown in Fig. 5. First we confirmed that pOmtl-110 and pOmtl-169 chains still cross-linked to their favoured ligands even if those were absent during synthesis and only post-translationally added (lanes 1 – 4 and 15 – 18). As already shown in Fig. 4, synthesis of pOmtl-110 in the mere presence of Ffh led to the smaller Ffh-adduct (Fig. 5, lane 6, X). Remarkably, post-translational incubation of these Ffh-pOmtl-110 complexes with 0.5 μM TF completely displaced the co-translationally bound Ffh resulting in a sole TF-adduct (lane 8, asterisk). As pointed out above, displacement is again meant to indicate removal from nascent pOmtl but not necessarily from the ribosome. If this displacement of Ffh by TF from pOmtl-110 was the result of a higher overall affinity of this nascent chain for TF, allowing for more binding of Ffh during synthesis by raising its concentration 10-fold (compare lanes 6 and 10)
should partially resist the displacement by TF. In fact, in these conditions pOmtl-110 was found cross-linked to both Ffh and TF (lanes 12 – 14). In much the same manner, TF the disfavoured signal sequence ligand of pOmtl-169 when bound co-translationally (lane 20) was virtually completely displaced by a post-translational addition of Ffh (lanes 22 -24). If these findings obtained in vitro using the model protein pOmtl can be extrapolated to the in vivo situation, our results collectively indicate that a bacterial nascent chain possessing an N-terminal signal sequence alternates between contacts to SRP and TF depending on the downstream exposure of high affinity binding sites for either ligand.
DISCUSSION
We have performed a comprehensive analysis of the molecular environment of a bacterial signal sequence when it emerges on the ribosome during ongoing synthesis. The signal sequence of pOmpA whose overall hydrophobicity falls short of those of classical SRP substrates (8), nevertheless interacts with Ffh when nascent chains are so short that they expose little more than the signal sequence from the ribosome (Figs. 2 and 3 and ref. 13). Upon emergence of TF binding sites in longer nascent chains, TF becomes the predominant cross-linking partner of the pOmpA signal sequence. This is exemplified by pOmtl-110 in Fig. 6A and C. Only when pOmtl-110 is synthesized in the complete absence of TF, does Ffh remain a cross-linking partner. If Ffh and TF are concomitantly present during synthesis, potential cross-links of pOmtl-110 with Ffh are overcome by those with TF. The same result is obtained when TF is added to pre-formed Ffh-pOmtl-110 complexes, indicating that TF can in fact displace Ffh from the signal sequence upon growth of the nascent chain. Displacement could partially be prevented by raising the concentration of Ffh ten fold during the initial binding period. These results are consistent with a competition of TF and SRP for their binding sites on an emerging nascent chain. When the nascent chain of pOmtl has grown to a size of 169 amino acids (Fig. 6 B and D) it recruits again Ffh with a clear competitive advantage over TF. This is inferred from the predominant cross-linking of pOmtl-169 to Ffh when TF and SRP were simultaneously present during synthesis. The prevalence of Ffh-cross-links could only be reduced by performing synthesis with ten times more TF. The
5
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

preferential binding of pOmtl-169 to Ffh is most clearly demonstrated by the fact that TF is displaced from pOmtl-169 by a post-translational incubation with Ffh. A positive correlation between the extent of cross-linking and the binding affinity of both ligands for a given nascent chain is indicated by the mutual displacement of the ligands and the influence that the concentration of the ligands exerted on the cross-linking intensities. The validity of this cross-linking approach is underscored by the fact that, where comparable, the employment of the site-specific photoprobe and of the more global chemical cross-linker gave identical results (compare Fig. 3 with Figs. 4 and 5). Moreover, the carbene generated by UV-irradiation from the diazirine group of Tmd-Phe, covalently links only to ligands that come closer than 2 Å, which is unlikely to frequently happen by casual collisions with non-functional binding partners. In order to evaluate the dependence of the cross-linking intensity on the ligand concentration we have varied the ratio of the effective concentrations of both ligands rather than attempting to establish the intracellular concentrations of TF and SRP in our in vitro conditions. The reason is that for both purified ligands it is difficult to estimate the fraction of active material which seems particularly relevant for the oxidation-sensitive Ffh (39). Furthermore, the functionally meaningful concentration of TF is probably that of the ribosome-bound chaperone which again is difficult to determine accurately. Following estimations of intracellular concentration, probably every ribosome has at least one molecule of TF bound, whereas the level of Ffh would only be sufficient to saturate 1 – 10% of the ribosomes. If this assumption is correct, a simultaneous binding of Ffh and TF (23,24) would be limited to only a fraction of ribosomes. In any event, our results addressing only the interactions of Ffh and TF with the nascent polypeptide do not exclude such a situation (cf. Fig. 6 C and D). The model drawn in Fig. 6C suggests that TF when recruited to the exposed nascent pOmtl-110 chain, sequesters the signal sequence from a direct association with Ffh. The possibility of an auxiliary function of TF in influencing the accessibility of the signal sequence of a secretory protein to SRP has previously been discussed (6,13). While this has remained a point at issue (11), the finding of a loss of Ffh interaction upon growth of a nascent
secretory protein strongly suggests that Ffh cross-links when documented for short presecretory proteins do not necessarily prove that the protein is a functional SRP substrate (13,14,40). The cross-linking results presented here are consistent with a model in which high affinity binding sites for TF and Ffh when emerging on the ribosome, determine the liganding of the N-terminal signal sequence. In contrast to the signal anchor-type TM3 of MtlA, the stop-transfer TM2 did not prove to be such a high affinity binding site for Ffh. One major difference between both topogenic sequences is the restriction of positively charged amino acid residues to either flanking end (41). Thus while positive charges usually precede a signal anchor sequence they are located at the distal end of a stop-transfer sequence. It is therefore conceivable that SRP in addition to a hydrophobic TM recognizes positive charges upstream of a signal anchor sequence as recently addressed (42,43). Whether or not TM3 of pOmtl-169 in order to recruit Ffh to the ribosome-nascent chain-complexes has to be fully exposed on the ribosomes has not been studied in any detail here. However, the increase in Ffh-cross-links relative to those to TF when comparing pOmtl-140 and pOmtl-110 (cf. Fig. 3) might suggest that recruitment of Ffh to the nascent chain was already initiated when TM3 was still hidden in the ribosomal exit tunnel (cf. Fig. 2). There are precedents for such a signalling between the ribosomal exit tunnel and downstream processes (44,45).
Models reflecting the influence of downstream sequence sections on the liganding pattern of a signal sequence are depicted in Fig. 6 C and D. They are based on the findings that the pOmpA signal sequence remains attached to ribosomal proteins L23 and L29 necessarily leading to a certain degree of folding of the nascent chains near the ribosomal exit site. As the cross-linking behaviour of pOmtl-110 and pOmtl-169 did not change when the ligands were added post-translationally (Fig. 5), TF and Ffh are probably not involved in such an initial folding. The acquisition of a more compact structure of the ribosome-associated nascent chain by anchoring its N-terminus near the ribosomal exit site is fully compatible with the observed displacement of ligands.
Using vacant ribosomes, Buskiewicz et al. (23) reported that binding of the SRP receptor, FtsY, to ribosome-bound Ffh excludes TF from the ribosome. To examine the effect of
6
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

FtsY under our experimental conditions, pOmtl-110 nascent chains were co-translationally charged with 0.1 μM Ffh and post-translationally incubated with 0.5 μM TF. As in Fig. 5, TF was able to displace Ffh from these nascent chains, even in the presence of 0.5 μM FtsY and 5 mM GMP-PNP added to allow a stable Ffh-FtsY interaction. This finding indicates that FtsY did not interfere with TF-binding at least to these ribosome-associated nascent chains of pOmtl. Similarly we also investigated interaction of SecA with nascent pOmtl-chains, as SecA was found to associate with the signal sequence of
short pOmpA nascent chains (13,31,46). SecA cross-links were in fact obtained with pOmtl nascent chains ranging in size from 65 up to 169 residues. As the SecA cross-links did not appear to compete with those to TF and Ffh, they could either result from nascent chain-independent binding of SecA to ribosomes (46-48) or also reflect interactions with released nascent chains (6). In any event, these results do not help settle the issue (31,46) if SecA indeed is able to mediate membrane-targeting of nascent chains in E. coli.
REFERENCES
1. de Keyzer, J., van der Does, C., and Driessen, A. J. (2003) Cell. Mol. Life. Sci. 60,
2034-2052 2. Müller, M., Koch, H. G., Beck, K., and Schäfer, U. (2001) Prog. Nucleic Acid Res.
Mol. Biol. 66, 107-157 3. Koch, H. G., Moser, M., and Müller, M. (2003) Rev. Physiol. Biochem. Pharmacol.
146, 55-94 4. Pool, M. R. (2005) Mol. Membr. Biol. 22, 3-15 5. Valent, Q. A., de Gier, J.-W. L., von Heijne, G., Kendall, D. A., ten Hagen-Jongman,
C. M., Oudega, B., and Luirink, J. (1997) Mol. Microbiol. 25, 53-64 6. Beck, K., Wu, L. F., Brunner, J., and Müller, M. (2000) EMBO J. 19, 134-143 7. Valent, Q. A., Kendall, D. A., High, S., Kusters, R., Oudega, B., and Luirink, J. (1995)
EMBO J. 14, 5494-5505 8. Beha, D., Deitermann, S., Müller, M., and Koch, H. G. (2003) J. Biol. Chem. 278,
22161-22167 9. de Gier, J. W., Scotti, P. A., Saaf, A., Valent, Q. A., Kuhn, A., Luirink, J., and von
Heijne, G. (1998) Proc. Natl. Acad. Sci. U. S. A. 95, 14646-14651 10. Kim, J., Rusch, S., Luirink, J., and Kendall, D. A. (2001) FEBS Lett. 505, 245-248 11. Lee, H. C., and Bernstein, H. D. (2001) Proc. Natl. Acad. Sci. U. S. A. 98, 3471-3476 12. Neumann-Haefelin, C., Schäfer, U., Müller, M., and Koch, H. G. (2000) EMBO J. 19,
6419-6426 13. Eisner, G., Koch, H. G., Beck, K., Brunner, J., and Müller, M. (2003) J. Cell Biol.
163, 35-44 14. Chevalier, N., Moser, M., Koch, H. G., Schimz, K. L., Willery, E., Locht, C., Jacob-
Dubuisson, F., and Müller, M. (2004) J. Mol. Microbiol. Biotechnol. 8, 7-18 15. Gu, S. Q., Peske, F., Wieden, H. J., Rodnina, M. V., and Wintermeyer, W. (2003)
RNA 9, 566-573 16. Kramer, G., Rauch, T., Rist, W., Vorderwülbecke, S., Patzelt, H., Schulze-Specking,
A., Ban, N., Deuerling, E., and Bukau, B. (2002) Nature 419, 171-174 17. Pool, M. R., Stumm, J., Fulga, T. A., Sinning, I., and Dobberstein, B. (2002) Science
297, 1345-1348 18. Halic, M., Becker, T., Pool, M. R., Spahn, C. M., Grassucci, R. A., Frank, J., and
Beckmann, R. (2004) Nature 427, 808-814 19. Ferbitz, L., Maier, T., Patzelt, H., Bukau, B., Deuerling, E., and Ban, N. (2004) Nature
431, 590-596 20. Baram, D., Pyetan, E., Sittner, A., Auerbach-Nevo, T., Bashan, A., and Yonath, A.
(2005) Proc. Natl. Acad. Sci. U. S. A. 102, 12017-12022
7
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

21. Schlünzen, F., Wilson, D. N., Tian, P., Harms, J. M., McInnes, S. J., Hansen, H. A., Albrecht, R., Buerger, J., Wilbanks, S. M., and Fucini, P. (2005) Structure 13, 1685-1694
22. Ullers, R. S., Houben, E. N., Raine, A., ten Hagen-Jongman, C. M., Ehrenberg, M., Brunner, J., Oudega, B., Harms, N., and Luirink, J. (2003) J. Cell Biol. 161, 679-684
23. Buskiewicz, I., Deuerling, E., Gu, S. Q., Jockel, J., Rodnina, M. V., Bukau, B., and Wintermeyer, W. (2004) Proc. Natl. Acad. Sci. U. S. A. 101, 7902-7906
24. Raine, A., Ivanova, N., Wikberg, J. E., and Ehrenberg, M. (2004) Biochimie 86, 495-500
25. Cammack, K. A., and Wade, H. E. (1965) Biochem. J. 96, 671-680 26. Lesley, S. A., Brow, M. A., and Burgess, R. R. (1991) J. Biol. Chem. 266, 2632-2638 27. Deuerling, E., Schulze-Specking, A., Tomoyasu, T., Mogk, A., and Bukau, B. (1999)
Nature 400, 693-696 28. Grisafi, P. L., Scholle, A., Sugiyama, J., Briggs, C., Jacobson, G. R., and Lengeler, J.
W. (1989) J. Bacteriol. 171, 2719-2727 29. Maneewannakul, S., Maneewannakul, K., and Ippen-Ihler, K. (1994) Plasmid 31, 300-
307 30. Beck, K., Eisner, G., Trescher, D., Dalbey, R. E., Brunner, J., and Müller, M. (2001)
EMBO Rep. 2, 709-714 31. Behrmann, M., Koch, H. G., Hengelage, T., Wieseler, B., Hoffschulte, H. K., and
Müller, M. (1998) J. Biol. Chem. 273, 13898-13904 32. Koch, H. G., Hengelage, T., Neumann-Haefelin, C., MacFarlane, J., Hoffschulte, H.
K., Schimz, K. L., Mechler, B., and Müller, M. (1999) Mol. Biol. Cell 10, 2163-2173 33. Sugiyama, J. E., Mahmoodian, S., and Jacobson, G. R. (1991) Proc. Natl. Acad. Sci.
U. S. A. 88, 9603-9607 34. Vervoort, E. B., Bultema, J. B., Schuurman-Wolters, G. K., Geertsma, E. R., Broos, J.,
and Poolman, B. (2005) J. Mol. Biol. 346, 733-743 35. Aboulwafa, M., and Saier M. H., Jr. (2003) Mol. Microbiol. 48, 131-141 36. Werner, P. K., Saier, M. H., Jr., and Müller, M. (1992) J. Biol. Chem. 267, 24523-
24532 37. Agashe, V. R., Guha, S., Chang, H. C., Genevaux, P., Hayer-Hartl, M., Stemp, M.,
Georgopoulos, C., Hartl, F. U., and Barral, J. M. (2004) Cell 117, 199-209 38. Patzelt, H., Kramer, G., Rauch, T., Schonfeld, H. J., Bukau, B., and Deuerling, E.
(2002) Biol. Chem. 383, 1611-1619 39. Ezraty, B., Grimaud, R., El Hassouni, M., Moinier, D., and Barras, F. (2004) EMBO J.
23, 1868-1877 40. Sijbrandi, R., Urbanus, M. L., ten Hagen-Jongman, C. M., Bernstein, H. D., Oudega,
B., Otto, B. R., and Luirink, J. (2003) J. Biol. Chem. 278, 4654-4659 41. Goder, V., and Spiess, M. (2001) FEBS Lett. 504, 87-93 42. Peterson, J. H., Woolhead, C. A., and Bernstein, H. D. (2003) J. Biol. Chem. 278,
46155-46162 43. Batey, R. T., Rambo, R. P., Lucast, L., Rha, B., and Doudna, J. A. (2000) Science 287,
1232-1239 44. Liao, S., Lin, J., Do, H., and Johnson, A. E. (1997) Cell 90, 31-41 45. Houben, E. N., Zarivach, R., Oudega, B., and Luirink, J. (2005) J. Cell Biol. 170, 27-
35 46. Karamyshev, A. L., and Johnson, A. E. (2005) J. Biol. Chem. 280, 37930-37940 47. Hoffschulte, H. K., Drees, B., and Müller, M. (1994) J. Biol. Chem. 269, 12833-12839 48. Liebke, H. H. (1987) J. Bacteriol. 169, 1174-1181 49. Bernabeu, C., and Lake, J. A. (1982) Proc. Natl. Acad. Sci. U. S. A. 79, 3111-3115
8
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

FOOTNOTES
* We thank Dr. Hans-Georg Koch for a critical reading of the manuscript. Antibodies against SecG were a generous gift from Drs. Hajime Tokuda and Ken-ichi Nishiyama (University of Tokyo, Japan) and those against ribosomal proteins from Dr. Richard Brimacombe (Max Planck Institute of Molecular Genetics, Berlin, Germany). This work was supported by grants from the Deutsche Forschungsgemeinschaft (Sonderforschungsbereich 388 and Graduiertenkolleg 434). M.M. is also supported by grant LSHG-CT-2004-005257 of the European Union.
‡ Present address: Biozentrum, University of Basel, Klingelbergstrasse 50/70, CH-4056 Basel, Switzerland
§ Present address: Centre for Vascular Research, University of New South Wales, Gate 9, High Street, Randwick, NSW 2031, Australia
1 The abbreviations used are: DSS, disuccinimidyl suberate; INV, inside-out, inner membrane vesicles; SRP, signal recognition particle; TF, Trigger factor; TM, transmembrane span; Tmd-Phe, L-4’-(3-[trifluoromethyl]-3H-diazirin-3-yl) phenylalanine.
FIGURE LEGENDS FIG. 1. Replacing the first signal anchor sequence of MtlA by a cleavable signal sequence does not interfere with membrane localization. A, Topography model and construction of pOmtl. The 22 amino acid long signal sequence of pOmpA (bold black line) plus the first four residues of mature OmpA were fused to an N-terminally truncated version of MtlA lacking 46 amino acids including the first TM (dotted lines). The 5 remaining TMs of MtlA are represented by grey boxes with the numbers indicating the first and last residue of each predicted TM. A recently proposed topography model of MtlA differs from the one shown by predicting two additional TMs between TM4 and TM5 (34). The about 300 residue long soluble C-terminal part of MtlA is cytosolic. Numbering starts with the first amino acid of pOmtl. B, MtlA and pOmtl were expressed in E. coli strain LGS322 lacking the mtlA-gene (ΔmtlA) from plasmids p717MtlA-B or p717Omtl. Control cells were transformed with vector pKSM717. Soluble and membrane fractions were each subjected to SDS-PAGE in a 1 : 1 protein ratio (corresponding to 0.4 : 1 equivalent volumes). They were analyzed by Western-blotting using antibodies raised against MtlA (αMtlA) and SecG (αSecG) as a marker protein for the inner membrane. C, Full length MtlA and pOmtl were synthesized by a reconstituted E. coli transcription/translation system in the presence or absence of inside-out inner membrane vesicles (INV). Radiolabelled translation products were separated by SDS-PAGE and visualized by phosphorimaging. The molecular masses of marker proteins are indicated. Half of each reaction was digested with proteinase K (PK). Addition of INV leads to the generation of membrane protected fragments of MtlA and pOmtl that are resistant toward proteinase K (lanes 4 and 8). In the case of pOmtl, addition of INV also causes signal sequence cleavage (open arrow head). D, Plasmid-encoded pOmtl and MtlA were expressed in E. coli strain LGS322 (ΔmtlA). At zero time, protein synthesis was blocked by the addition of spectinomycin to a final concentration of 100 µg/ml. Aliquots were removed at the indicated time points, precipitated with 5% TCA, resuspended in gel loading buffer, and analyzed by SDS-PAGE and Western-blotting using antibodies raised against MtlA (upper panel). Scanned blots were quantified using Imagequant software (lower panel). The data shown are representative of two parallel experiments. FIG. 2. Structure of the ribosome-associated nascent chains of pOmpA and pOmtl used. The length of the various nascent chains is indicated by the numbers of amino acid residues. The signal sequence and downstream mature sequence sections of pOmpA are represented by hatched and stippled boxes, respectively. TMs and interspersed loops of MtlA are drawn as light and dark grey boxes, respectively. TM2 and TM4 are indicated, TM3 is denominated SA (signal anchor sequence). The black clamp symbolizes the ribosomal exit tunnel assumed to bury 30 amino acid residues of each nascent chain (49). Arrows indicate the two positions of the cross-linker Tmd-Phe at amino acid positions 11 and 36. The bottom diagram depicts full size pOmtl for comparison. FIG. 3. The molecular vicinity of a nascent signal sequence depends on emerging downstream sequence sections. Ribosome-associated pOmpA and pOmtl nascent chains of the indicated chain
9
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from

lengths were synthesized by a reconstituted transcription/translation system prepared from E. coli. Truncation of the mRNAs was achieved by the addition of complementary oligodeoxynucleotides and RNaseH cleavage (31). Stop (UAG) codons at the indicated positions were suppressed by tRNAsup charged with Tmd-Phe. In vitro reactions were supplemented with 80-100 nM Ffh. UV-irradiation (UV) resulted in adducts with Ffh (X, 50 kDa) and TF (asterisk, migrating on SDS-PAGE at 58 kDa) and with the ribosomal proteins L23 (white star, 11 kDa) and L29 (black star, 8 kDa) as identified by co-immunoprecipitation with specific antibodies (αFfh, αTF, αL23, αL29). TF-adducts (brackets) often resolve into several species (13). ● labels an UV-independent transcription/translation product of the Omtl-DNAs. A, UV-induced cross-links obtained with Tmd-Phe replacing Val at position 11 within the hydrophobic core of the signal sequence of pOmpA. B, UV-induced cross-links obtained with Tmd-Phe replacing Ile at position 36 within TM2 of MtlA. FIG. 4. TF and SRP compete for nascent pOmtl chains. Nascent chains of pOmtl-110 and pOmtl-169 were expressed in vitro using stop codon-less mRNAs. Different from the experiments summarized in Fig. 3, the subcellular components required for in vitro translation had now been prepared from a TF null mutant strain. Purified Ffh and TF were present during synthesis at the indicated concentrations. Where indicated cross-linking was induced by 2.5 mM DSS. Marked are unspecific cross-links (●) and those to the 50 kDa Ffh (X) and TF (asterisk) running on SDS-PAGE with an apparent mass of 58 kDa. FIG. 5. TF and SRP mutually displace each other from nascent pOmtl chains. Nascent chains of pOmtl-110 and pOmtl-169 were synthesized as described in the legend to Fig. 4 in the presence of the ligands indicated in row co-translational. In addition they were subsequently incubated for 10 min at 37 °C with the ligands listed in row post-translational. Where indicated cross-linking was performed with DSS and immunoprecipitation carried out with antisera directed against Ffh and TF (αFfh, αTF). FIG. 6. Competitive cross-linking of TF and SRP to pOmtl-nascent chains. A, Model of the conformation of pOmtl-110 at the ribosomal exit site represented by ribosomal protein L23 (hatched area). The signal sequence is depicted as grey cylinder, TM2 as white cylinder. Listed are the experimental conditions in which pOmtl-110 was incubated with SRP and TF (co- or post-translationally) and the major observed cross-links, with the arrow indicating a change of ligands. B, As in panel A, except that results obtained for the longer pOmtl-169 chain are summarized. The front grey cylinder represents the signal anchor-type TM3. The interaction of the signal sequence with ribosomal protein L29 is experimentally verified by direct cross-linking. C, As indicated by the cross-linking results summarized in panel A (coloured rows), TF is the preferred binding partner of pOmtl-110 interacting with its signal sequence and areas around TM2. SRP might remain bound to the synthesizing ribosome. D, As indicated by the cross-linking results summarized in panel B (coloured rows), SRP is the preferred binding partner of pOmtl-169 being recruited to the signal sequence by TM3. TF might remain bound to the synthesizing ribosome.
10
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from




Gottfried Eisner, Michael Moser, Ute Schäfer, Konstanze Beck and Matthias Müllersequence of a growing nascent polypeptide
Alternate recruitment of signal recognition particle and trigger factor to the signal
published online January 18, 2006J. Biol. Chem.
10.1074/jbc.M511388200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 28, 2019http://w
ww
.jbc.org/D
ownloaded from