and maize grain protein fractions
TRANSCRIPT

Clin. exp. Immunol. (1990) 79, 135-140
Antibody responses reveal differences in oral tolerance to wheatand maize grain protein fractions
R. B. JOHNSON, J. T. LABROOY & J. H. SKERRITT* Department of Medicine, (Royal Adelaide Hospital),University of Adelaide, Adelaide; and *CSIRO Wheat Research Unit, Division of Plant Industry, North Ryde, Australia
(Acceptedfor publication 28 July 1989)
SUMMARY
The influence of diet on humoral immune responses to gluten- and maize-derived proteins wasexamined using ELISA and protein blotting techniques. Mice raised on the maize-based (gluten-free)diet responded well to parenteral immunization with each of six gluten-derived protein preparations(whole gliadin, two omega-gliadin fractions, wheat salt-soluble proteins, a peptic-tryptic digest and asubtilisin digest of gluten), as serum antibody levels increased at least 300-fold in each case. Incontrast, mice raised on the wheat-based diet responded poorly to immunization with either wholegliadin or omega-gliadin and were virtually non-responsive to enzymic digest ofgluten. Diet had littleeffect on the magnitude of the antibody response to wheat salt-soluble proteins, with both groupsshowing a 300-fold increase in titre. Similarly, tolerance to alpha-zeins, the alcohol-soluble proteinsof maize, did not occur on either diet. However, some oral tolerance was observed to maize glutelin.The specificity of the various antibody responses was then analysed by immunoblotting. Followingimmunization with gluten proteins or digests, antibodies from the maize-fed mice bound more or lessequally to each of the main gliadin bands and to the glutenins while the mice on the wheat-based diethad antibody specific for omega-gliadin proteins. Serum antibodies from the maize-fed mice,immunized with either alpha-zein or maize glutelin, showed even labelling of the major maizeendosperm proteins while antibodies from mice on the wheat diet showed strong labelling of the Mr27000 and 58000 bands. These results show that diet influenced the specificity, as well as themagnitude of serum antibody responses to cereal proteins. In addition, oral tolerance appeared toaffect the humoral response to some cereal proteins more than others. Both of these findings haveimportant implications for our understanding of coeliac disease.
Keywords gliadin zein antibodies oral tolerance gluten-free diet
INTRODUCTION
Dietary antigens represent a major antigenic challenge to thehost, considering both the quantity of food passing the gastro-intestinal tract and the fact that significant levels of protein areabsorbed intact, or only partially digested (Warshaw et al.,1971). However, healthy humans (Blazer et al., 1984; LaBrooyet al., 1986) and rodents (Coombs et al., 1983; Johnson et al.,1985) respond with only low levels of circulating antibody todietary antigens. The normal immune system has the capacity todiscriminate between harmless antigens in dietary constituentsand potentially harmful substances such as bacterial toxins.Antigen feeding has been shown to produce antigen- specifichyporesponsiveness (tolerance) in both humoral and cellularimmune responses (Thomas & Parrot, 1974; Hanson et al.,1977). Such oral tolerance has been demonstrated in rodentswith dietary protein from egg (Strobel & Ferguson, 1985), soya
Correspondence: Dr R. B. Johnson, Regional Veterinary Labora-tory, P.O. Box 388, Benalla, Victoria 3672, Australia.
and milk (Pathirana et al., 1981) and wheat (Johnson et al.,1985). Evidence of oral tolerance to bovine serum albumin(BSA) in humans has also been obtained (Korenblatt et al.,1968).
One group of nutritionally and clinically important dietaryantigens are the gliadins, a complex of at least 40 aqueousalcohol-soluble proteins which serve as storage proteins in thewheat grain. The gliadins are the component of wheat which istoxic for patients with coeliac disease (Van De Kamer et al.,1953). In susceptible individuals, the ingestion of gliadin isassociated with damage to the small intestine (villous atrophy),malabsorption of nutrients and elevated serum and intestinalantibodies against these cereal antigens (reviews by Davidson &Bridges, 1987; Cole & Kagnoff, 1988; Skerritt, 1988). Thismarked immune response to gluten-derived antigens is notfound in either control subjects or patients with other types ofintestinal disease (Blazer et al., 1984; LaBrooy et al., 1986).Coeliac patients have also shown evidence of cellular hyper-sensitivity to gluten-derived antigens (O'Farrelly et al., 1984).
135

R. B. Johnson, J. T. LaBrooy & J. H. Skerritt
These observations suggest that coeliac patients may have adefect in the normal immunoregulatory mechanisms which limitsuch responses in healthy subjects.
Interestingly, there is little evidence that salt-soluble proteinsfrom wheat have coeliac toxicity (Van De Kamer et al., 1953).Furthermore, coeliac patients may ingest maize with little illeffect (Anand, Piris & Truelove, 1978) and aqueous alcohol-soluble protein fractions from this cereal did not demonstrate aspecific humoral immune response in coeliacs (Skerritt andRobson, unpublished). These findings raise the possibility thatwheat gliadins and alcohol-soluble maize proteins, as well assalt-soluble wheat proteins differ in their abilities to elicithumoral and/or cellular immune responses in coeliac patients.Any differences in the intrinsic antigenicity of the cereal proteinsmay be relevant to the pathogenesis of coeliac disease, as it ispostulated that the mucosal damage in this disease is immuno-logically mediated (Davidson & Bridges, 1987).
In order to investigate whether there are differences inimmunogenicity of coeliac-toxic and non-toxic cereal proteinfractions, groups of mice were immunized parenterally withthese fractions and the resulting antibody responses werecompared by ELISA and immunoblotting methods. The in-fluence of diet on these responses was also assessed using miceraised on either wheat or maize-based diets.
MATERIALS AND METHODS
ImmunogensWheat flour (Timgalen cultivar) was extracted (2 x 1 h) at 20°Cin 10% (w/v) sodium chloride solution (20 ml/g) with centri-fugation (10 000 g, 30 min) after each step. The supernatantswere combined, dialysed and then lyophilized to yield wheatsalt-soluble proteins (WSP). The pellet from centrifugation wasextracted twice as above with 70% ethanol and the combinedsupernatants from centrifugation were dialysed and lyophilizedto yield crude gliadin. Proteolytic digests were prepared fromcommercial wheat gluten using pepsin and trypsin sequentiallyto yield 'Frazer's fraction 11 1' (PT-gluten; Frazer et al., 1959) or
subtilisin (Alcalase 0-6 1; Novo, Bazwaerd, Denmark) (Batey,1985). In the latter case, only the digested gluten that was solublein water at 20°C at 10% (w/v) was used (S-gluten). Feeding trialswith either product have indicated toxicity and mucosal damageto coeliac patients on otherwise gluten-free diets (Frazer et al.,1959; LaBrooy, Hetzel, Johnson & Skerritt, unpublished).A fraction analogous to gliadin was prepared from de-
germed maize by 70% ethanol extraction of the pellet fromsodium chloride treatment; this fraction is termed alpha-zein(Esen, 1987). The residue was extracted (2 h at 20°C) with10 ml/g 70% ethanol-I % (v/v) 2-mercaptoethanol; after centri-fugation the residue from this step was extracted (16 h at 20°C)with 10 ml/g of 50% (v/v) propan-1-ol-2% (v/v) 2-mercapto-ethanol-I% (v/v) acetic acid in water. The supernatant fromcentrifugation was neutralized with NaOH, then NaCl was
added to I M final concentration. After 20 h at 4°C, theprecipitated protein was isolated by centrifugation, dialysed andthen freeze-dried to yield maize glutelin.
Each of the two major omega-gliadin bands was isolatedfrom Timgalen gliadin by preparative electrophoresis (Caldwell,1984).
MiceBALB/c mice were bred and maintained on either a standardwheat-based diet (51% wheat grain and 18% bran and pollard),or a gluten-free diet based on 70% crushed maize (MillingIndustries, Adelaide, Australia). Groups of three to ten micewere injected intraperitoneally with 0 25 mg of the appropriateantigen emulsified in 0 2 ml Freund's complete adjuvant. Asecond injection of antigen in Freund's incomplete adjuvantfollowed 14 days later. Mice were bled prior to, and then 7 daysafter the second injection.
ELISAAntibody levels were measured using an ELISA (Johnson et al.,1985). Briefly, the wells of U-bottomed microtitre plates(Costar, Cambridge, MA) were coated with antigen at 5 ug/ml.Alkaline phosphatase-labelled anti-mouse immunoglobulin(Dako-immunoglobulins, Copenhagen, Denmark) was used asthe conjugate, p-nitrophenyl phosphate (Sigma Chemical Co.,St Louis, MO) was the substrate and the ELISA titre wasassigned as the reciprocal of the highest dilution of serum whichgave an absorbance at 405 nm of 015. This end-point wasselected because it was greater than the background binding ofthe conjugate to the coated plates (less than 0 05), and it alsomaintained consistency with previous results (Johnson et al.,1985; LaBrooy et al., 1986).
Electrophoresis and protein blottingThe compositions of the protein fractions used for immuni-zations were analysed by a sodium dodecyl sulphate poly-acrylamide gel electrophoresis (SDS-PAGE) method similar tothat used previously (Skerritt & Underwood, 1986), except thatsamples were extracted with 2% (w/v) SDS-2% 2-mercapto-ethanol in 62 mm Tris-HCl, pH 7.5; and 1 5 mm thick gels wereused; and gels were run for only 1300 Vh.
Antibody specificities were determined by SDS-PAGE oftotal grain protein extracts and electroblotting for 100 Vh.Antibody binding to wheat gliadinswas also assessed afterPAGEunder acidic conditions in 4-12% polyacrylamide gradient gels,and blotting to cathodally placed nitrocellulose membranes(Skerritt & Underwood, 1986). Sera were diluted in 3% BSA inphosphate-buffered saline (PBS) (50 mm sodium phosphate-0 9% sodium chloride, pH 7-2) and incubated for 90 min at20°C. The membranes were washed four times (5 min) withPBS-0 05% Tween 20. Alkaline phosphatase-labelled anti-mouse immunoglobulins (Promega, Madison, WI) diluted with2% BSA in Tris-buffered saline-Tween (TBST, 10 mM Tris-HC1, pH 8 0, containing 150 mm NaCl, I mr MgCl2, 0.1 mMZnCl2 and 0-05% Tween 20) were then added and incubated for60 min at 20°C. After four 5-min washes in TBST, substrate(Nitroblue tetrazolium, 330 ,ug/ml and 5-bromo-4-chloro-3-indololphosphate, 165 pg/ml in 100 mM Tris-HCl-100 mMNaCI- 5 mm MgCI2, pH 9 5) was added and specific antibodybinding revealed by the appearance of blue purple zones within60 min of substrate addition.
RESULTS
Composition of immunogen fractions
SDS-PAGE was performed on each immunogen preparation(Fig. 1). The total flour protein extract contained polypeptide
136

Oral tolerance to wheat and maize proteins
f g h i
kD
94 -.-a
67 - -
43--
30-^
20-1
14-4
Fig. 1. Analysis of wheat and maize immunogens and antigens bysodium dodecyl sulphate-polyacrylamide gel electrophoresis. Mole-cular weight markers are shown. a, total wheat proteins; b, Timgalenwheat gliadin; c, commercial gliadin (used in earlier work, Johnson et
al., 1985); d, peptic-tryptic digest of wheat gluten; e, subtilisin digest ofgluten; f, wheat salt-soluble proteins; g, total wheat proteins (doubleloading); h, total maize proteins; i, maize alpha-zein; and j, maizeglutelin.
species with (reduced monomer) molecular masses of up to Mr120 000. As expected, the gliadin proteins covered a mol. wt ofMr 30 000-80 000, while the peptic-tryptic digest (Frazer'sfraction 111) and the subtilisin digests of gluten were mainlycomposed of peptides up to Mr 15000 and below Mr 10000(confirmed by size exclusion HPLC), respectively. The WSPfraction consists ofseveral components ofMr 10 000-80 000; themajor components had different mobilities to those in gliadin.The electrophoretic profile of maize proteins was similar tothose published elsewhere (Esen, 1987). Two major proteinclusters (Mr 21 000-25 000) were noted in alpha-zein, the 70%ethanol extract of maize (Esen, 1987). The maize glutelinfraction contained alpha-zein and polypeptides of Mr 27000-28 000 and 58 000, along with some high mol. wt components.
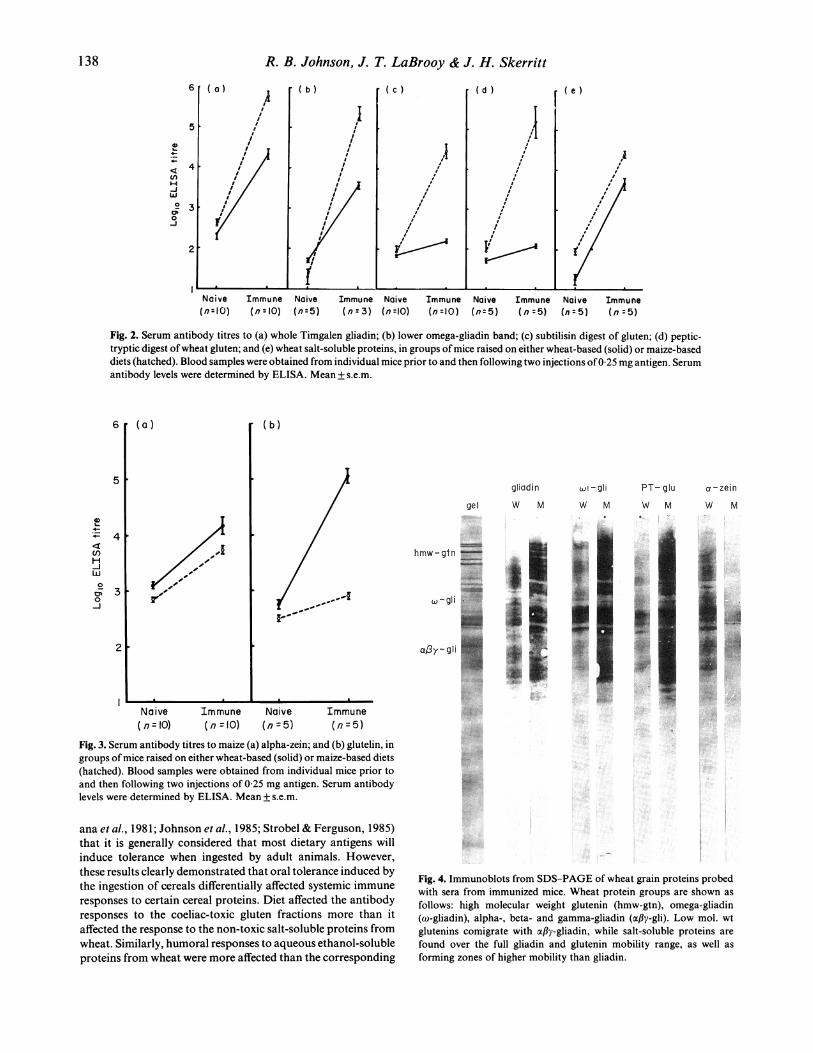
Effect of diet on antibody titresAntibody titres were determined prior to, and then followingimmunization. Naive mice from both dietary groups hadmeasurable antibody titres (200-400) to whole gliadin (Fig. 2).However, while the immunization of mice on a wheat-contain-ing diet with gliadin resulted in a 90-fold increase in antibodytitres, a considerably greater (1800-fold) increase was noted withthe animals on a gluten-free diet. Similar trends were noted withthe omega-gliadins, although levels of antibody to these pro-
teins in immunized mice were lower. Selective elevations ofantibodies specific for the protein digests were seen in im-munized mice on the gluten-free diet (Fig. 2). These mice showeda 300-fold (S-gluten) and a 1800-fold (PT-gluten) elevation,
while mice on a wheat-containing diet (similarly immunized),showed a less than three-fold increase.
In contrast, diet had little influence on antibody responses inmice immunized with WSP, with both groups of mice showing300-fold increases in antibody titres (Fig. 2). Furthermore, therewas no evidence of tolerance to maize alcohol-soluble protein inmice on either diet. Titres to alpha-zein were quite high beforeimmunization in mice on both diets (Fig. 3). In contrast, mice ona maize-containing diet showed little change in antibodyresponse to maize glutelin after immunization, although markedincreases in titre were seen in mice on a wheat-containing diet.
Antibody specificityPossible effects of diet on antibody specificity were studied byprobing immunoblots of SDS-PAGE fractionations of wheatand maize total protein extracts with pooled sera from miceimmunized with selected wheat and maize protein fractionsrespectively. In order to analyse more closely the gliadinspecificity of the sera, acidic-buffer gel electrophoresis andimmunoblotting were also performed.
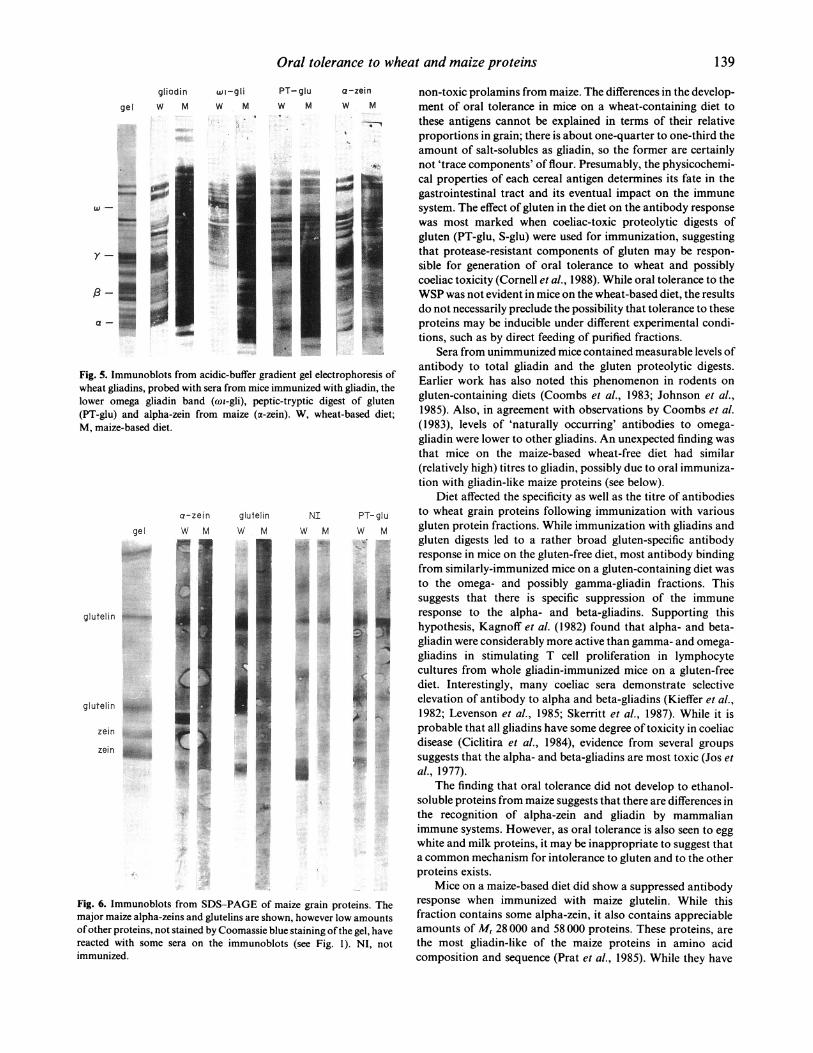
To minimize the possible effects of antibody concentrationon apparent specificity, sera from mice immunized with wheatgluten fractions were assayed at a number of dilutions (resultsfor 1:500 dilution for mice on wheat-containing diets and1:2500 dilution for maize-containing diets are shown). A broadantibody response to the full range of gliadins and glutenins wasnoted for mice on a wheat-free diets after immunization with thevarious gluten fractions and digests (Fig. 4), but mice on wheat-containing diets had most reactivity to proteins of Mr 60000-80000. Immunoblots from acidic-buffer gels confirmed thatthese were omega-gliadins (Fig. 5). Broader gliadin (but notglutenin) cross-reactivity was noted with sera from alpha-zein-immunized mice at dilutions up to 1: 2500. Sera from un-immunized mice did not label gliadins on immunoblots atdilutions greater than 1:200.
Although titres to alpha-zein were similar in zein-immunizedmice on either diet, antibody specificities differed (Fig. 6).Antibody from maize-fed mice bound to each of the main maizeendosperm proteins, while selectivity for Mr 27000 and Mr58 000 protein was noted for the mice on a wheat diet.Immunoblots of sera from glutelin-immunized mice on themaize diet also showed even labelling of major maize proteins,while sera from those on the wheat diet showed strongerlabelling of the Mr 28 000 and 58 000 bands. The converse wastrue for serum from mice immunized with the PT-digest ofgluten; antibodies from mice on the maize-diet bound selectivelyto the Mr 58 000 glutelin while equal labelling of bands was seenwith antibodies from mice on the wheat-diet.
DISCUSSION
Each of the eight cereal antigens tested was capable ofgenerating a serum antibody response when injected parenter-ally into mice. The results confirmed previous findings thatmarked oral tolerance to gliadin does occur (Johnson et al.,1985), and emphasize that much of the poor antibody responseto prolamins noted by other investigators (Baldo, Sutton &Wrigley, 1982; Blazer et al., 1984; Skerritt, 1988) was not due tointrinsically lower immunogenicity.
Oral tolerance has been observed to such a wide range ofantigens (Thomas & Parrot, 1974; Hanson et al., 1977; Pathir-
a b c d e
137
':
A... ..M
R. B. Johnson, J. T. LaBrooy & J. H. Skerritt
U)
I-I
1-4
w
0
Naive Immune Naive Immune Naive Immune Naive Immune Naive Immune
(nIO0) (n=10) (n=5) (n:3) (n:=0) (n=10) (n:5) (n=5) (n=5) (n=5)
Fig. 2. Serum antibody titres to (a) whole Timgalen gliadin; (b) lower omega-gliadin band; (c) subtilisin digest of gluten; (d) peptic-
tryptic digest ofwheat gluten; and (e) wheat salt-soluble proteins, in groups ofmice raised on either wheat-based (solid) or maize-based
diets (hatched). Blood samples were obtained from individual mice prior to and then following two injections of0 25 mg antigen. Serum
antibody levels were determined by ELISA. Mean + s.e.m.
6 ( a )
51'
0
V)H
w
0
CA
0
4
2
Naive Immune Naive Immune(n= 10) (n =10) (n =5) (n =5)
Fig. 3. Serum antibody titres to maize (a) alpha-zein; and (b) glutelin, ingroups ofmice raised on either wheat-based (solid) or maize-based diets(hatched). Blood samples were obtained from individual mice prior toand then following two injections of 0-25 mg antigen. Serum antibodylevels were determined by ELISA. Mean + s.e.m.
ana et al., 1981; Johnson et al., 1985; Strobel & Ferguson, 1985)that it is generally considered that most dietary antigens willinduce tolerance when ingested by adult animals. However,these results clearly demonstrated that oral tolerance induced bythe ingestion of cereals differentially affected systemic immuneresponses to certain cereal proteins. Diet affected the antibodyresponses to the coeliac-toxic gluten fractions more than itaffected the response to the non-toxic salt-soluble proteins fromwheat. Similarly, humoral responses to aqueous ethanol-solubleproteins from wheat were more affected than the corresponding
( b )
glicdin wi-gli PT- glu a-zein
gel W M W M W M W M
~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~hmw-gin I iI I I I I i
at4y-gli
Fig. 4. Immunoblots from SDS-PAGE of wheat grain proteins probed
with sera from immunized mice. Wheat protein groups are shown as
follows: high molecular weight glutenin (hmw-gtn), omega-gliadin
(co-gliadin), alpha-, beta- and gamma gliadin (afiy gli). Low mol. wt
glutenins comigrate with afly gliadin, while salt-soluble proteins are
found over the full gliadin and glutenin mobility range, as well as
forming zones of higher mobility than gliadin.
138
A I
Oral tolerance to wheat and maize proteins
gel
y -
a-
a-
gliodinW M
i.. ............
re
i:.V -L.
wi-gilW m
N:
PT-glu a-zein
W M W M
t ~ ~ ~
Fig. 5. Immunoblots from acidic-buffer gradient gel electrophoresis ofwheat gliadins, probed with sera from mice immunized with gliadin, thelower omega gliadin band (con-gli), peptic-tryptic digest of gluten(PT-glu) and alpha-zein from maize (a-zein). W, wheat-based diet;M, maize-based diet.
a-zein glutelin NI PT-glu
gel W M W. M W M W Mgelg
glutelin
in.t.alin
zein
zein
Fig. 6. Immunoblots from SDS-PAGE of maize grain proteins. Themajor maize alpha-zeins and glutelins are shown, however low amountsofother proteins, not stained by Coomassie blue staining ofthe gel, havereacted with some sera on the immunoblots (see Fig. 1). NI, notimmunized.
non-toxic prolamins from maize. The differences in the develop-ment of oral tolerance in mice on a wheat-containing diet tothese antigens cannot be explained in terms of their relativeproportions in grain; there is about one-quarter to one-third theamount of salt-solubles as gliadin, so the former are certainlynot 'trace components' of flour. Presumably, the physicochemi-cal properties of each cereal antigen determines its fate in thegastrointestinal tract and its eventual impact on the immunesystem. The effect of gluten in the diet on the antibody responsewas most marked when coeliac-toxic proteolytic digests ofgluten (PT-glu, S-glu) were used for immunization, suggestingthat protease-resistant components of gluten may be respon-sible for generation of oral tolerance to wheat and possiblycoeliac toxicity (Cornell et al., 1988). While oral tolerance to theWSP was not evident in mice on the wheat-based diet, the resultsdo not necessarily preclude the possibility that tolerance to theseproteins may be inducible under different experimental condi-tions, such as by direct feeding of purified fractions.
Sera from unimmunized mice contained measurable levels ofantibody to total gliadin and the gluten proteolytic digests.Earlier work has also noted this phenomenon in rodents ongluten-containing diets (Coombs et al., 1983; Johnson et al.,1985). Also, in agreement with observations by Coombs et al.(1983), levels of 'naturally occurring' antibodies to omega-gliadin were lower to other gliadins. An unexpected finding wasthat mice on the maize-based wheat-free diet had similar(relatively high) titres to gliadin, possibly due to oral immuniza-tion with gliadin-like maize proteins (see below).
Diet affected the specificity as well as the titre of antibodiesto wheat grain proteins following immunization with variousgluten protein fractions. While immunization with gliadins andgluten digests led to a rather broad gluten-specific antibodyresponse in mice on the gluten-free diet, most antibody bindingfrom similarly-immunized mice on a gluten-containing diet wasto the omega- and possibly gamma-gliadin fractions. Thissuggests that there is specific suppression of the immuneresponse to the alpha- and beta-gliadins. Supporting thishypothesis, Kagnoff et al. (1982) found that alpha- and beta-gliadin were considerably more active than gamma- and omega-gliadins in stimulating T cell proliferation in lymphocytecultures from whole gliadin-immunized mice on a gluten-freediet. Interestingly, many coeliac sera demonstrate selectiveelevation of antibody to alpha and beta-gliadins (Kieffer et al.,1982; Levenson et al., 1985; Skerritt et al., 1987). While it isprobable that all gliadins have some degree of toxicity in coeliacdisease (Ciclitira et al., 1984), evidence from several groupssuggests that the alpha- and beta-gliadins are most toxic (Jos etal., 1977).
The finding that oral tolerance did not develop to ethanol-soluble proteins from maize suggests that there are differences inthe recognition of alpha-zein and gliadin by mammalianimmune systems. However, as oral tolerance is also seen to eggwhite and milk proteins, it may be inappropriate to suggest thata common mechanism for intolerance to gluten and to the otherproteins exists.
Mice on a maize-based diet did show a suppressed antibodyresponse when immunized with maize glutelin. While thisfraction contains some alpha-zein, it also contains appreciableamounts of Mr 28 000 and 58 000 proteins. These proteins, arethe most gliadin-like of the maize proteins in amino acidcomposition and sequence (Prat et al., 1985). While they have
139

140 R. B. Johnson, J. T. LaBrooy & J. H. Skerritt
some immunological relationship to gliadin (Skerritt, 1988) theyare very different from alpha-zein (Esen, 1985). The antibodyblotting patterns ofmice on a wheat-based diet, immunized withalpha-zein, differed from those of similar mice immunized withmaize glutelin.
These studies highlight the complexity involved in analysingimmune responses to dietary antigens. The results demonstratedthat diet had a marked influence on both the magnitude andspecificity of the humoral immune response to cereal proteins.Serum antibody responses to the coeliac-toxic gluten-derivedwheat proteins and digests were suppressed as a result of gluteningestion. A connection between protease-resistance and im-munogenicity is suggested, but requires further study. This iseven truer for the relationship between immunogenicity andcoeliac-toxicity. Continuation of this work may provide furtherinsights into both the regulation of humoral and cellularimmune responses to dietary antigens and the role of suchresponse in coeliac disease.
ACKNOWLEDGMENTSWe wish to thank Miss Susan Clare for technical assistance. This workwas supported by grants from the NH & MRC, the University ofAdelaide, the Royal Adelaide Hospital and the CSIRO-UniversitiesCollaborative Research Fund.
REFERENCES
ANAND, B.S., PIRIS, J. & TRUELOVE, S.C. (1978) The role of variouscereals in coeliac disease. Q. J. Med. 185, 101.
BALDO, B.A., SUTTON, R. & WRIGLEY, C.W. (1982) Grass allergens, withparticular reference to cereals. Prog. Allergy, 30, 1.
BATEY, I.L. (1985) Enzymic solubilization of wheat gluten. J. appl.Biochem. 7, 423.
BLAZER, S., NAVEH, Y., BERANT, M., MERZBACH, D. & SPERBER, S.(1984) Serum IgG antibodies to gliadin in children with celiac diseaseas measured by an immunofluorescence method. J. pediatr. Gastro-enterol. Nutr. 3, 205.
CALDWELL, K.A. (1984) Purification of cereal prolamins by means ofpreparative PAGE at acid pH. Biochem. Physiol. Pflanzen. 179, 31.
CICLITIRA, P.J., EVANS, D.J., FAGG, N.L.K., LENNOX, E.S. & DOWLING,R.H. (1984) Clinical testing of gliadin fractions in coeliac patients.Clin. Sci. 66, 357.
COLE, S.G. & KAGNOFF, M.F. (1985) Celiac disease. Annu. Rev. Nutr. 5,241.
COOMBS, R.R.A., KIEFFER, M., FRASER, D.R. & FRAZIER, P.J. (1983)Naturally developing antibodies to wheat gliadin fractions and toother cereal antigens in rabbits, rats and guinea pigs on normallaboratory diets. Int. Arch. Allergy appl. Immunol. 70, 200.
CORNELL, H.J., AURICCHIO, R.S., DE RITIS, G., DE VINCENZI, M.,MAIURI, L., RAIA, V. & SILVANO, V. (1988) Intestinal mucosa ofceliacs in remission is unable to abolish toxicity of gliadin peptides onin vitro developing fetal rat intestine and cultured atrophic celiacmucosa. Pediatr. Res. 24, 233.
DAVIDSON, A.G.F. & BRIDGES, M.A. (1987) Coeliac disease: a critical
review of aetiology and pathogenesis. Clin. chim. Acta, 163, 1.ESEN, A. (1987) A proposed nomenclature for the alcohol-soluble
proteins (zeins) of maize (Zea mays L.). J. Cereal Sci. 5, 117.FRAZER, A.C., FLETCHER, R.F., Ross, C.A.C., SHAW, B., SAMMONS,
H.G. & SCHNEIDER, R. (1959) Gluten-induced enteropathy. The effectof partially digested gluten. Lancet, ii, 252.
HANSON, D.G., VAZ, N.M., MAIA, L.C.S., HORNBROOK, M.M., LYNCH,J.M. & Roy, C.A. (1977) Inhibition of specific immune responses byfeeding protein antigens. Int. Arch. Allergy appl. Immunol. 55, 526.
JOHNSON, R.B., LABROOY, J.T., SHEARMAN, D.J.C. & DAVIDSON, G.P.(1985) The effect of diet on systemic immune responses to wheatgliadin. Aust. J. exp. Biol. med. Sci. 63, 299.
Jos, J., CHARBONNIER, L., MOUGENOT, J.F., MOSSE, J. & REY, J. (1977)Isolation and characterization of the toxic fraction of wheat gliadinin coeliac disease. In Perspectives in Coeliac Disease (ed. by B.McNicholl, C. F. McCarthy & P. F. Fottrell) p. 75. MTP Press,Lancaster.
KAGNOFF, M.F., AUSTIN, R.K., JOHNSON, H.C.L., BERNARDIN, J.E.,DIETLER, M.D. & KASARDA, D.D. (1982) Celiac sprue: correlationwith murine T-cell responses to wheat gliadin components. J.Immunol. 129, 2693.
KIEFFER, M., FRAZIER, P.J., DANIELS, N.W.R. & COOMBS, R.R.A. (1982)Wheat gliadin fractions and other cereal antigens reactive withantibodies in the sera of coeliac patients. Clin. exp. Immunol. 50, 65 1.
KORENBLATT, P.E., ROTHBERG, R.M., MINDEN, P. & FARR, R.S. (1968).Immune responses in humans after oral and parenteral exposure to
bovine serum albumin. J. Allergy, 41, 226.LABROOY, J.T., HOHMANN, A.W., DAVIDSON, G.P., HETZEL, P.A.S.,JOHNSON, R.B. & SHEARMAN, D.J.C. (1986) Intestinal and serum
antibody in coeliac disease: a comparison using ELISA. Clin. exp.
Immunol. 66, 661.LEVENSON, S.D., AUSTIN, R.K., DIETLER, M.D., KASARDA, D.D. &KAGNOFF, M.F. (1985) Specificity of antigliadin antibody in coeliacdisease. Gastroenterol. 89, 1.
O'FARRELLY, C., WHELAN, C.A., FEIGHERY, C.F. & WEIR, D.G. (1984)Suppressor cell activity in coeliac disease induced by alpha-gliadin, a
dietary antigen. Lancet, ii, 1305.PATHIRANA, C., GOULDING, N.J., GIBNEY, M.J., PITTS, J.M., GAL-
LAGHER, P.J. & TAYLOR, T.G. (1981) Immune tolerance produced bypre- and postnatal exposure to dietary antigens. Int. Arch. Allergyappl. Immunol. 66, 114.
PRAT, S., CORTADAS, J., PUIGDOMENECH, P. & PALAU, J. (1985) Nucleicacid (cDNA) and amino acid sequences of the maize endospermprotein glutelin-2. Nucl. Acids. Res. 5, 1493.
SKERRITT, J.H. (1988) Immunochemistry of cereal grain storage pro-
teins. In Advances in Cereal Science and Technology Vol IX (ed. byY. Pomeranz) p. 263, AACC Press, St Paul, MN.
SKERRITT, J.H., JOHNSON, R.B., HETZEL, P.A.S., LABROOY, J.T.,SHEARMAN, D.J.C. & DAVIDSON G.P. (1987) Variation of serum andintestinal gluten antibody specificities in coeliac disease. Clin. exp.
Immunol. 68, 189.SKERRITT, J.H. & UNDERWOOD, P.A. (1986) Specificity characteristics ofmonoclonal antibodies to wheat grain storage proteins. Biochim.Biophys. Acta, 874, 245.
STROBEL, S. & FERGUSON, A. (1985) Oral tolerance-induction andmodulation. Klin. Paediat. 197, 297.
THOMAS, H.C. & PARROTT, D.M.V. (1974) The induction of tolerance to
a soluble protein antigen by oral administration. Immunology, 27,631.
VAN DE KAMER, J.H., WEIJERS, H.A. & DICKE, W.K. (1953) Coeliacdisease. IV. An investigation into the injurious constituents of wheatin connection with their action on patients with coeliac disease. Actapaediatr. 42, 223.
WARSHAW, A.L., WALKER, W.A., CORNELL, R. & ISSELBACHER, K.J.(1971) Small intestinal permeability to macromolecules. Trans-mission of horseradish peroxidase into mesenteric lymph and portalblood. Lab. Invest. 25, 675.