applying 13c stable isotope probing and the pufm … · gene to investigate anoxygenic phototrophs...
TRANSCRIPT

Applying13CStableIsotopeProbingandthepufMGenetoInvestigateAnoxygenicPhototrophsinthePinkLayerofaSaltMarshMicrobialMator,pufMadventures:SIPpingfromthegardenhoseandotherdelightsTheaWhitmanCornellUniversityMicrobialDiversity,MBL,2012Abstract:AnoxygenicphototrophsarekeyC‐fixingmembersoftheGreatSippewissettsaltmarshmicrobialmats.Thediversityandpresenceofthesemicrobesinmats,Cfixation,andthediversityofthepufMgeneinpurplesulfurandnon‐sulfurbacteriawereinvestigatedusing454sequencing,13Cstableisotopeprobing(SIP),andpufMclonelibraries.Nearbymatsarerelativelysimilarattheclasslevel,andanoxygenicphototrophcommunitiesdifferbetweenyoungandoldmats,althoughanoxygenicphototrophsinthepinklayerofthematmayonlyrepresentafewpercentofthepopulation.AnoxygenicphototrophsarelikelyfixingCinthepinklayerofthemat,althoughSIPresultsaresomewhatequivocalandpufMsequencesresemblesequencesfromdiverseknowntaxa.FutureinvestigationscouldincludedevelopmentofthepinklayercommunitiesovertimeanddeeperinvestigationofCcyclingdynamicsintheselayersandtheroleofanoxygenicphototrophstherein.(Anumberofnotesspecificallyregardingwhatwassuccessfulandwhatstepsmightbechangedareincludedinthisreportforfuturestudentsconsideringasimilarproject.)

2
1.Introduction:Anoxygenicphototrophsareafascinatinganddiversegroupoforganisms,includingpurple(Alpha‐Beta‐andGamma‐proteobacteria)andgreensulfur(Chlorobi)andnon‐sulfur(Chlrorflexi)bacteria.Usingbacteriochlorophyllsa,b,c,d,e,oreveng(inheliobacteria),theyfixcarbon(C)withouttheproductionofO2,instead,oxidisingelectrondonorsotherthanwater,suchasH2S(Achenbachetal.,2001).Withdiversepigments,theyarecapableofcapturingenergyfromwavelengthsoflightthatareinaccessibletootherphotosyntheticorganisms(Hubasetal.,2011).Bacterialphotosyntheticreactioncentres(RCs)areclassifiedbroadlyintotwogroups‐RCI‐typeandRCII‐type.RCIsproduceweakoxidantsandstrongreductants,usingaterminal,electron‐acceptingFe‐Scluster,whileRCIIsproducestrongoxidantsandweakreductants(FryantandFrigaard,2006).RCIIincludestrans‐membraneproteinsLandM,whichareconsidereddiagnosticofthoseanoxygenicphotosyntheticmicrobeswiththisreactioncentretype,includingthepurplebacteria.TheMsubunitiscodedbythepufMgene,whichhasbeensequencedinarangeofbacteria.Becausemostanoxygenicphototrophsrequiretheproductsofoxygenicphototrophy(e.g.,S2‐)toproceed,theirCfixationmaynotbeconsidered“true”primaryproduction,sinceitisindirectlyfuelledbyotherorganisms(exceptinthecasessuchaswherethereisgeologically‐derivedS2‐available,suchashotsprings).AlthoughanoxygenicphototrophsarethusperhapslessimportanttoglobalCfixationthancyanobacteriaandothertrueprimaryproducers,theycertainlyplayanimportantroleintheC,N,andSbiogeochemistryofsomeecosystems.AprimeexampleofanecosystemwhereanoxygenicphotorophsplayakeyroleisthemicrobialmatsofLittleandGreatSippewissettsaltmarshesinWoodsHole,MA.Thesematsareafascinatingconsortiumoflaminatedmicrobialcommunities,wherecyanobacteriaareoftenthepioneers,co‐dominatingthetoplayerwithdiatoms,followedbypurpleandgreenbacteria,alongwithsulfate‐reducingbacteria(Nicholsonetal.,1987).TheycanalsoincludewhitelayersorpatchesofS2‐‐oxidisingbacteria,suchasBeggiatoa,aswellassulfate‐reducingbacteria,whichcausetheprecipitationofdarkironsulfidesinthelowerlayers.Becauseoftheirimportancetothebiogeochemistryofthisecosystem,thefascinatinglydiverserangeofmetabolismsystemstheyarecapableof(particularlythepurplenon‐sulfurbacteria),andtheirmicrocharisma,IchosetoinvestigatethediversityofandevidenceofC‐fixingactivityinpurpleanoxygenicphototrophsinthepinklayerofamicrobialmatfromGreatSippewissettsaltmarshusingpufMasamarkergene.Keyquestionsinclude:(1)Howsimilararethecommunitiesoftwoadjacentmatsandayoungmat?(2)Whatistherelativeimportanceofanoxygenicphototrophsinthepinklayerofthemat?(3)AreanoxygenicphototrophsfixingCinthepinklayerofthemat?and(4)HowarethesequencesofpufMinthepinklayerrelatedtoknownpufMsequences?

3
2.Methods:2.1MatSelection,Collection,DNAExtraction,andPreliminaryAnalysesMatswerecollectedfromGreatSippewissettSaltMarshatlowtideonJuly6,2012.Theywereselectedbasedontheirrelativelywell‐developedpinklayer.Theywerecollectedbypressingasterile8.5cmdiameterx1.3cmPetriplateintothesand,removingtheshortcore,andtruncatingitsbottomwithaknife,closingthePetridishandsealingwithParafilmforthereturntriptothelab.Twopairsofmatswereselected(AandB),alongwithayounger,less‐developed,nearbymat(<2maway)forcomparison.Twoadjacentwatersampleswerecollectedinsterile15mLFalcontubesforchemicalanalyses.Inthelab,eachmatwascutinhalfwithasterilerazorblade,anda~3mmstripwascutalongthecutedge.Fromthisstrip,thepinklayerwasremoved.DNAwasextractedfromthepinklayerusingtheMoBioPowerBiofilmextractionkit.(Note:Theoptionalinitialde‐wateringstepintheprotocolwasnotnecessaryforthesemats.Inaddition,agreatermasswasusedfortheextraction(0.44g)thantheprotocolrecommended,duetothefactthatsandmadeupmuchofthe“biofilm’s”mass.)ThisDNAwasusedfortheinitial(t0)clonelibraryandthe454sequencing.ThewatersampleswereanalyzedforCO3‐andSO4‐concentrationsontheionchromatograph.ItwaschallengingtoobtainaclearpeakfortheCO3‐concentrations,duetointerferencefromtheSO4‐peak,but[CO3]wasestimatedtobewithintherangeof2mM,whichisstandardforseawater.TheSO4‐wasmeasuredtobe27mM,whichisalsowithinnormalseawaterrange.ThepHwasmeasuredas8.06,usingapHmeter.2.2454sequencingandanalysisTheDNAextractionyieldedbetween68‐128ng/µLDNA(asmeasuredontheNanoDropmachine)foreachmatsample.ThisDNAwasamplifiedfor454sequencingusing4µLoftemplateDNAina25µLreactionwithPhusionpolymerasemastermixand907Randbarcoded515Fprimers.Ultimately,1/10or1/100xdilutionswerenecessarytogetamplificationfromtheoldermatsamples.ExtractedDNAwasquantifiedusingimagingsoftwareandthentheamplifiedPCRproductwassubmittedforprocessingandanalysis.454datawerede‐multiplexedbymakingamastermapperfile,whichwasusedwiththesplit_libraries.pycommandinQIIMEtopulloutbar‐codedsequencesbetween400and450bp,withmaxbarcodeerrorsof0allowed.ThesesequenceswerethengroupedintoOTUsusingthepick_otus.pycommand(at97%ID,usinguclust),andarepresentativefromeachgroupofOTUswaschosenusingpick_rep_set.py,whichwasassignedtaxonomyusingtheassign_taxonomy.pycommandandalarge,diversetrainingsetofreferencesequencesfromtheSilvareferencedatabase,whichhadbeentrimmedtotheregionbetweenthe454primersused(Werneretal.,2012).TheOTUtablewasbuiltusingthemake_otu_table.pycommand,andrarefactioncurveswerebuiltusingmultiple_rarefactions.pywithasamplesizeof500,3resamplingtimes,andanintervalsizeof1.

4
PleaseseeAppendixTableA.1for454metadata.2.3PrimerselectionforpufMPrimersforthepufMgenewereorderedinlab‐readyformbasedonthosedescribedinAchenbachetal(2001),knownaspufM.557FandpufM.750R.Theseprimersaredesignedtoamplifya229bpsegmentofthepufMgeneinanoxygenicphototrophs,particularlytargetingpurplesulfurandnon‐sulfurbacteria,aswellasgreennon‐sulfurbacteria.Inaddition,degeneratepufMprimersdescribedbyYutinetal(2005),knownaspufM_uniFandpufM_uniR,wereordered,butalthoughsomeproductcouldbeamplified,aPCRprogramwasnotsuccessfullyoptimizedfortheiruse,andtheywerenotusedfurtherinthisstudy.APCRprogramwithPromegaMasterMixwasdevelopedforoptimalamplificationusingtheAchenbachetal.(2001)primers,whichwas95°Cfor2min.,then30x[95°Cfor30s,63°Cfor30s,72°Cfor45s],followedby72°Cfor5min.Primerswereusedataconcentrationof0.25µMinthePCR.Sharp,cleanproductswereobtainedusingthisreactionwithDNAextractedfromthepinklayersofthemat.2.3StableIsotopeProbingofMatMicrocosmsAsinglemicrocosmwasconstructedforeachtreatment,withhalfofeachmatAandBaddedtoeach,asanattempttonormalizeformatheterogeneity.Themathalvesweregentlyplacedinthebottomof~500mLBormioliRoccoFidoItalianglasspreservejars,withrubbergasketsandmetalclampstosealthem.Thematswerejustbarelycoveredwith60mLsterileseawatersaltsolution,with27mMSO42‐andadjustedtopH8(thewaterlevelwasloweredslightlyafteronedayofincubationtoleaveabout1/3ofthematabovewaterlevel).NaHCO3wasaddedasaconcentratedsolutiontothewatertobringitto2mMHCO3‐,andanequivalentamountofNaHCO3wasinjectedintothematsat40evenly‐spacedsitespermicrocosmwithasyringe.TheonlydifferencebetweenthetwomicrocosmswasthattheNaHCO3addedtothelabelledtreatmentwaslabelledwith>99%13C,whiletheNaHCO3fortheunlabelledtreatmentwasstandardlab‐grade.(Anoteforfuturestudents‐thelabelledNaHCO3seemedmuch“fluffier”thantheunlabelled,perhapsduetoadifferentproductionprocess.)Themicrocosmswereincubatedat30°Cina24hlightincubatorfor3.5days.ThejarswereopenedbrieflyonceadaytorepeattheNaHCO3additions,foratotalof3additionsoverthecourseoftheexperiment.Attheendoftheincubation,themicrocosmswereopened,andthematsweretakenaparttocollectthepinklayerforDNAextraction.Foreachtreatment,twotubesof0.44gpinklayerhadDNAextractedforeachmatusingthePowerBiofilmDNAextractionkit,followingthepreviousprocedure.TheextractedDNAwasthencombinedintoonetube,concentratedonthevacuumcentrifuge,thengel‐purified.OnlythefragmentsofDNAestimatedtobegreaterthan4kbinlengthwereselected,inordertodecreasethetimeneededforallDNAfragmentstosettleattheirultimatedensityintheCsClgradienttube(Buckleyetal.,2007).ThegelfragmentwaspurifiedusingtheE.Z.N.A.Omegagelextractionkit,witha30µLfinalelution.Thefinalelutionstepwasrepeatedwiththesameflow‐throughsolution,toattemptto

5
getthemostDNAoffthefilter,inaconcentratedform.(Thismightnotberecommended,becausesubsequentapplicationsofthisprocedureforotherexperimentsdecreasedthequalityofthesample,perhapsbyelutingmoresalts.[Forthisapplication,saltsshouldnothavebeenanissue.])TheresultingDNAwasquantifiedontheNanoDropmachine.Gradienttubesweresetupusingagradientbuffersolutionof15mMTris‐HCL,pH8.0,15mMEDTA,and15mMKCl.ItwasmixedwithCsClsolutioninordertoobtainadensityofexactly1.762gmL‐1,usingadigitalrefractometertomeasureitsdensity.BeckmanbelltoptubeswerefilledalmosttothetopwiththeCsClbuffersolution,andthenpurifiedDNAwasaddedaftercalculatingtheamountneededtoensuretheadditionofequalamountsofDNAandTEbuffertobalancefromeachtreatment(3.9µgeach).TubeswerebalancedtopreciselythesamemassbyaddingCsCl‐buffersolution,andthencappedandinvertedseveraltimestomixthoroughly.Tubeswereplacedintherotor,withstemadaptorsaddedtopreventneckcollapse,andwerespunonanOptimaMAX‐XPBeckmanCoulterultracentrifugeat55,000rpmonaTLA110rotor(164,000xg)at20°Cfor65hoursundervacuum.Aftercentrifugation,tubeswerecarefullyremoved,twoatatime,leavingtheremainingtubesre‐balancedandspinningduringfractionation.Fractionationwasachievedbypiercingthebottomofeachtubewitha20Gneedlewiththeneckcutofftoallowdropstofallmorefreely.ThetopofthetubewasthenpiercedwithaneedleconnectedbystifftubingtoaHarvardApparatussyringepumpprogrammedtodeliver0.100mLwatermin‐1,thuspushingthedensesolutionoutoftheneedleinsertedatthebottomofthetube,into1.5mLmicrocentrifugetubes.Refractiveindexwasmeasuredandrecordedimmediatelyaftereachfractionwascollected,afterwhichtubeswerecapped.Thepumpcontinuedtoreleaseadroportwoofsolutionafterstoppingthepump,whichwascollected,butthetimeallowedforalldropstofallwassomewhatsubjective,likelyintroducingsomeerrorintotheprocedure.(Notetofutureusers‐measuringthevolumeofeachfractionatthispointishighlyrecommended,despitethefactthatitmayseemtobeasomewhatunappealingactivityat3AM.)36fractionswerecollectedforeachtube,ranginginrefractiveindexfrom1.4129to1.3921.Afterfractionation,alltubeswerede‐saltedandcleanedusingthefollowingprocedure:300µLDNAse‐freewaterwith240µLisopropanol[30minat15,000gonmicrocentrifuge;discardsupernatant];1mL70%EtOHinDNAse‐freewater[15minonmicrocentrifugeat15,000g;discardsupernatant;repeatedonce];letdryontissuesforseveralhours(ittookalmost8hoursforsamplestofinallydry‐possibleduetoresidualwater);finalelutioninsterile,DNAse‐freeTEbuffer.FortheEtOHprecipitationsteps,microcentrifugeswerekeptinthe4°CroominordertopromoteEtOHprecipitationofDNA.(ThiswouldberecommendedforallDNAcleanupstepsinfutureexperiments.)Afterfractionation,CsClremoval,andDNAcleanup,toquicklyevaluatethedistributionofpufMacrossfractions,asub‐samplefromevery3fractionswaspooledandpufMwasamplifiedusingthePCRprogramdescribedabove(givingatotalof12pooledsamplespertreatment).Uponrunningtheseproductsonagel,the

6
fractionswherethetwotreatmentsdivergedwereidentified,andthenamplifiedindividuallyforgreaterresolution.2.4CloneLibrariesandAnalysisThreeclonelibrarieswereconstructedfrompufMampliconsfromDNAextractedfrom(1)thecombinedintialmatsamplesAandB,(2)thefinallabelledmicrocosm,and(3)thefinalunlabelledmicrocosm.Forthemicrocosmsamples,DNAwasusedfromthepufMproductinthefourheaviestlaneswithvisibleproductsfromthehigh‐resolutionPCRreaction(4visiblelanesforthelabelled,1visiblelanefortheunlabelled).Fortheinitialmatsample,allpufMDNAwasused.Foralltreatments,theproductswerecutoutofthegelandpurifiedusingtheMilliporeDNAGelExtractionkit.CloningwasperformedfollowingtheTopoTAIsomeraseInvitrogenkitwithcompetentE.colicells.Theligationstepwasincreasedto2hoursinordertomaximizeligationofthepufMamplicon.Theplasmidwaselectroporatedintothecells,whichwerethenshakenfor1hourandplatedonampicillin‐treatedplatesat100and10µL.Clonesappearedinalltreatmentsafter12hours,andwerepickedtofill96‐wellplatesofampicillin‐treatedgrowthmedia.Ultimately,onlyhalfoftheinitialsample,20clonesfromtheunlabelled,and72clonesfromthelabelledtreatmentsweresequenced.Forclonelibrarydataanalysis,plasmidsequenceswereremovedfromthesequenceddatausingthecross_matchcommandinPhrap.SequencesweretranslatedtoproteinsusingExPASyTranslator,andthenBLASTedagainsttheNCBIproteindatabasetoensurethecorrectorientationandreadingframewasselected.InitialandfinalclonelibrarysequenceswerepooledwiththeGenBankreferenceproteinsequencesfromthepufMtreedevelopedbyAsaoetal.(2011).(AfewadditionalsequenceswereaddedaftertreeconstructiontoattempttofillinbrancheswithlowcoveragebyknownsequencesbyBLASTingsequencesfromthosebranchesagainsttheNCBIdatabase.)OrganismsandtheiraccessionnumbersarenotedintheAppendixTableA3.SequenceswerealignedusingMuscle,andthenonlytheportionsofthegenethatallclonesandreferencesequencescontainedwerewereselectedusingJalview.AlignedandtrimmedsequenceswereimportedintoArb,andaneighbour‐joiningtreewasconstructed,usingthePAMcorrectionfactorforproteins.Sequencesfromanybranchesthatwere“backwards”weredeleted,andthetreere‐drawn.ThetreewasdesignedinFigTree.Anadditionaltreewasconstructedusingonlytheclonelibrarysequences,usingthesamemethods.GCcontentwascalculatedforeachsequenceusingExcel,andthenmappedontotheclonelibrarytreeusingiTOL.Inaddition,16SandpufMsequenceswereobtainedfromDNAextractedfromtwoisolatesfrompurplenon‐sulfurbacterialenrichmentscreatedduringthefirsthalfofthecourse.These16SsequenceswereBLASTedagainsttheNCBIdatabaseandthepufMsequencespositionedinthepufMcloneandreferencesequencetree.PleaseseeAppendixTableA.2forclonelibrarymetadata.

7
2.5QuantitativePCR(qPCR)andDataProcessingThe16SandpufMcopynumbersfromtheSIPfractionswerequantifiedusingqPCR.AQuiagenQuantiFastSYBRGreenPCRkitwasusedforDNAamplificationof16S,using907Rand515FprimersandpufM,usingthesameprimersusedelsewhereinthisreport.1µLoftemplateDNAwasaddedforeachreaction.Aplasmid‐based16Sampliconfromourinitialgroupprojectclonelibraryplatewasusedasthetemplateforthe16Sstandardcurve,whileaplasmidfromtheinitialmatpinklayerpufMclonelibrarywasusedforthepufMstandardcurve.TheseplasmidswereselectedbasedontheirstrongamplificationinatestPCRrunandhighDNAconcentrationintheplate.Thereactionswere25µL,andwereamplifiedonaStepOnePlusreal‐timePCRsystemfromAppliedBiosystems.AnysamplesthatwereamplifiedwithCTvalues(wherethefluorescencecrossesthediagnosticthreshold)aboveorbelowthestandardcurvewereexcludedfromthedataanalysis.3.ResultsandDiscussion:3.1454SequencingofMats’PinkLayersThe454sequencesonthemats’pinklayersyieldedthousandsofhitsFigures1‐3).ClassificationusingtherepresentativesetdatabasedesignedbyKellyGravuerandChuckPepe‐RamseysuggestedthatCyanobacteria,Gamma‐,Delta‐,andAlphaproteobacteriadominatethemat.Thisisnotsurprising,asCyanobacteriaarevisiblypresent.
Figure1.Dominanttaxa(classlevel)ofoldermatA’spinklayer,asdeterminedby454sequencing(n=14570).
CyanobacteriaSubsectioniii
Gammaproteobacteria
Deltaproteobacteria
Notclassiuied
Alphaproteobacteria
Chloroplasts
Flavobacteria
Spirochaetes
Holophagae
CyanobacteriSubsectioni

8
Figure2.Dominanttaxa(classlevel)ofoldermatB’spinklayer,asdeterminedby454sequencing(n=12106).
Figure3.Dominanttaxa(classlevel)ofyoungermat’spinklayer,asdeterminedby454sequencing(n=9493).
CyanobacteriaSubsectioniii
Gammaproteobacteria
Deltaproteobacteria
Notclassiuied
Alphaproteobacteria
Chloroplasts
Flavobacteria
Spirochaetes
Holophagae
CyanobacteriSubsectioni
CyanobacteriaSubsectioniii
Gammaproteobacteria
Deltaproteobacteria
Notclassiuied
Alphaproteobacteria
Chloroplasts
Flavobacteria
Spirochaetes
Holophagae

9
DifferencesbetweenthetwoadjacentmatsAandB(Figures1and2)attheclasslevelwerenotstriking,andeventheyoungermatbroadlyresembledtheoldermats,withtheexceptionofagreaterdominanceofgammaproteobacteriaovercyanobacteriaintheyoungermat.Thisindicatesthat,althoughcertainlyheterogeneousatbothmacro‐andsub‐micronscales,directlyadjacentmatsamplescanberelativelysimilar,atleastataphylogeneticlevel‐functionalheterogeneitywasnotinvestigatedhere.ThesimilaritybetweentheadjacentoldermatsAandBpersistedwhenconsideringonlythetaxathatwereexpectedtobecapableofanoxygenicphototrophy,althoughdifferencesbetweenthedominanceofThiorhodococcusandThirhodovibrio(bothpurplesulfurbacteria)werenotable(Figures4and5).Theyoungermatwaslesssimilartotheoldermatswithregardstotheanoxygenicphototrophs,withThiorhodovibriobeingdominant,alongwithRoseospira(purplenon‐sulfurbacteria)andThiorhodococcus.Thetaxaidentifiedasanoxygenicphototrophsmadeup3%ofthetotal16S454hitsintheoldermats,butrepresented10%ofthehitsintheyoungermat,possiblyindicatingagreaterrelativeabundanceofthesetaxa.
Figure4.Dominantanoxygenicphototrophictaxa(genuslevel,exceptforCongregibacterandErythrobacter‐phylumlevel)ofoldermatA’spinklayer,asdeterminedby454sequencing(n=462).
ThiorhodococcusThiorhodovibrioCongregibacterRoseospiraMarinicellaChlorobiThiocapsaErythrobacterDeuluviicoccusRhodospirillaceaRhodocyclaceae

10
Figure5.Dominantanoxygenicphototrophictaxa(genuslevel,exceptforCongregibacterandErythrobacter‐phylumlevel)ofoldermatB’spinklayerasdeterminedby454sequencing(n=334).
Figure6.Dominantanoxygenicphototrophictaxa(genuslevel)ofyoungermat’spinklayer,asdeterminedby454sequencing(n=1018).
ThiorhodococcusThiorhodovibrioCongregibacterRoseospiraMarinicellaChlorobiThiocapsaErythrobacterDeuluviicoccusRhodospirillaceaRhodocyclaceaeChloroulexi
ThiorhodococcusThiorhodovibrioCongregibacterRoseospiraMarinicellaChlorobi
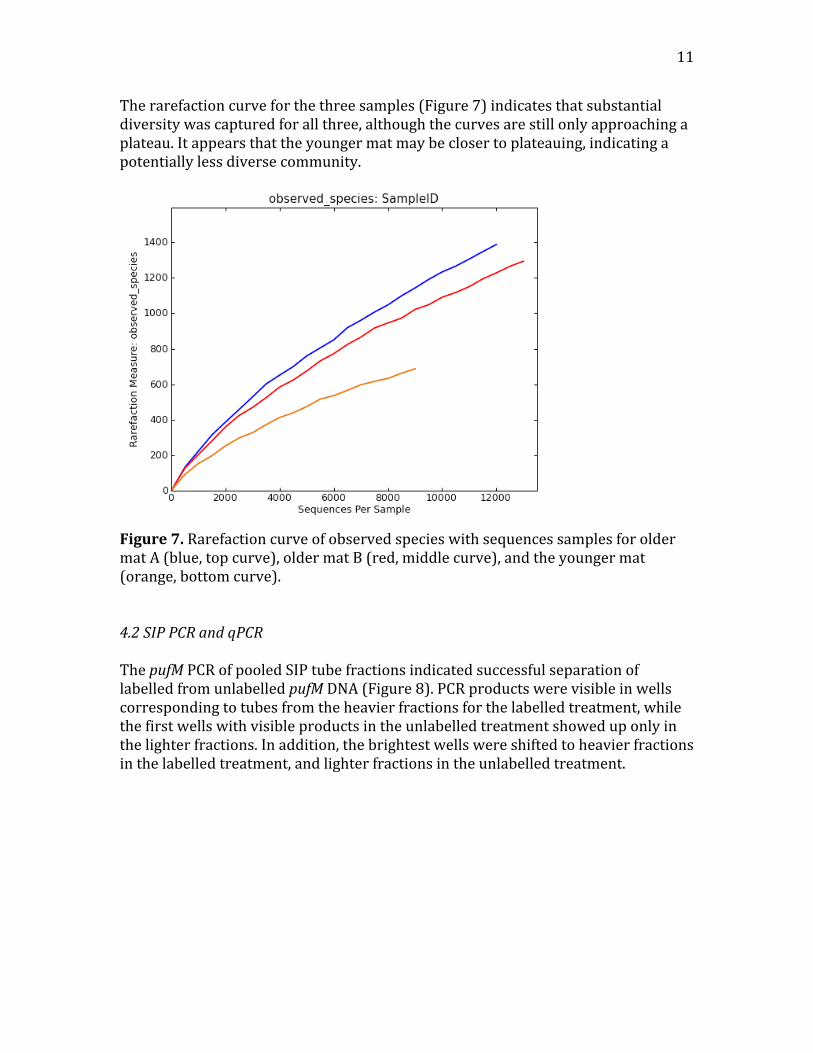
11
Therarefactioncurveforthethreesamples(Figure7)indicatesthatsubstantialdiversitywascapturedforallthree,althoughthecurvesarestillonlyapproachingaplateau.Itappearsthattheyoungermatmaybeclosertoplateauing,indicatingapotentiallylessdiversecommunity.
Figure7.RarefactioncurveofobservedspecieswithsequencessamplesforoldermatA(blue,topcurve),oldermatB(red,middlecurve),andtheyoungermat(orange,bottomcurve).4.2SIPPCRandqPCRThepufMPCRofpooledSIPtubefractionsindicatedsuccessfulseparationoflabelledfromunlabelledpufMDNA(Figure8).PCRproductswerevisibleinwellscorrespondingtotubesfromtheheavierfractionsforthelabelledtreatment,whilethefirstwellswithvisibleproductsintheunlabelledtreatmentshoweduponlyinthelighterfractions.Inaddition,thebrightestwellswereshiftedtoheavierfractionsinthelabelledtreatment,andlighterfractionsintheunlabelledtreatment.
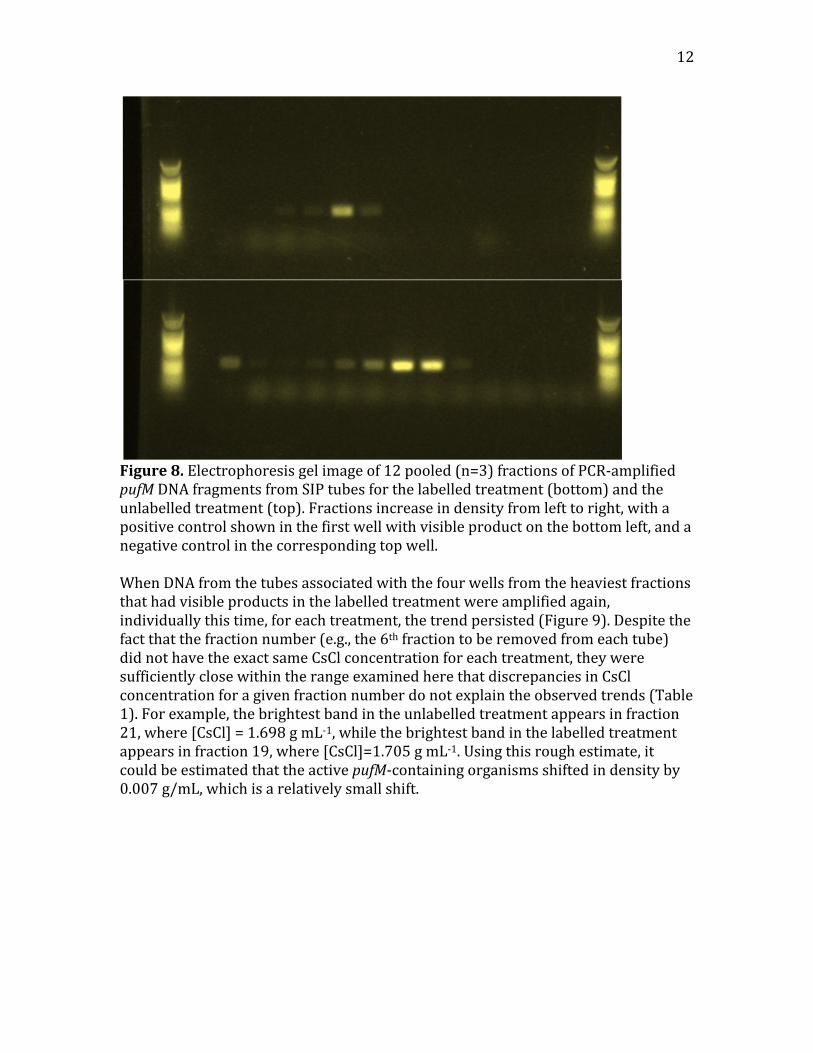
12
Figure8.Electrophoresisgelimageof12pooled(n=3)fractionsofPCR‐amplifiedpufMDNAfragmentsfromSIPtubesforthelabelledtreatment(bottom)andtheunlabelledtreatment(top).Fractionsincreaseindensityfromlefttoright,withapositivecontrolshowninthefirstwellwithvisibleproductonthebottomleft,andanegativecontrolinthecorrespondingtopwell.WhenDNAfromthetubesassociatedwiththefourwellsfromtheheaviestfractionsthathadvisibleproductsinthelabelledtreatmentwereamplifiedagain,individuallythistime,foreachtreatment,thetrendpersisted(Figure9).Despitethefactthatthefractionnumber(e.g.,the6thfractiontoberemovedfromeachtube)didnothavetheexactsameCsClconcentrationforeachtreatment,theyweresufficientlyclosewithintherangeexaminedherethatdiscrepanciesinCsClconcentrationforagivenfractionnumberdonotexplaintheobservedtrends(Table1).Forexample,thebrightestbandintheunlabelledtreatmentappearsinfraction21,where[CsCl]=1.698gmL‐1,whilethebrightestbandinthelabelledtreatmentappearsinfraction19,where[CsCl]=1.705gmL‐1.Usingthisroughestimate,itcouldbeestimatedthattheactivepufM‐containingorganismsshiftedindensityby0.007g/mL,whichisarelativelysmallshift.
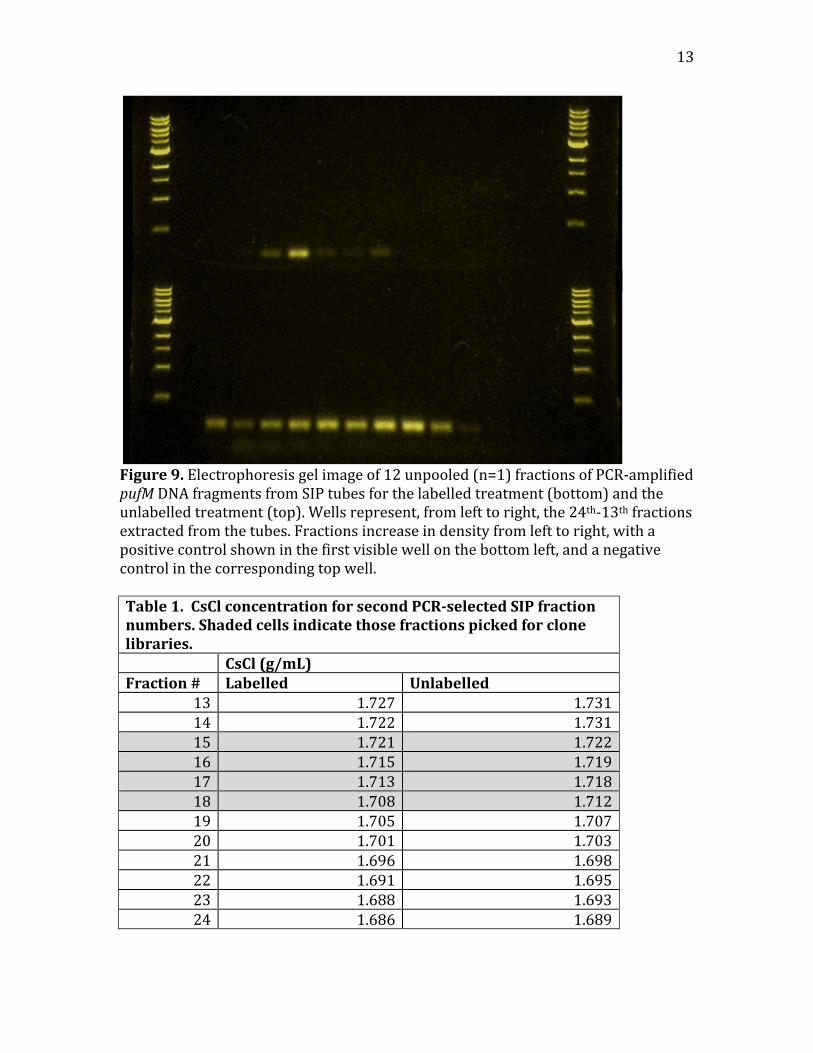
13
Figure9.Electrophoresisgelimageof12unpooled(n=1)fractionsofPCR‐amplifiedpufMDNAfragmentsfromSIPtubesforthelabelledtreatment(bottom)andtheunlabelledtreatment(top).Wellsrepresent,fromlefttoright,the24th‐13thfractionsextractedfromthetubes.Fractionsincreaseindensityfromlefttoright,withapositivecontrolshowninthefirstvisiblewellonthebottomleft,andanegativecontrolinthecorrespondingtopwell.Table1.CsClconcentrationforsecondPCRselectedSIPfractionnumbers.Shadedcellsindicatethosefractionspickedforclonelibraries. CsCl(g/mL)Fraction# Labelled Unlabelled
13 1.727 1.73114 1.722 1.73115 1.721 1.72216 1.715 1.71917 1.713 1.71818 1.708 1.71219 1.705 1.70720 1.701 1.70321 1.696 1.69822 1.691 1.69523 1.688 1.69324 1.686 1.689
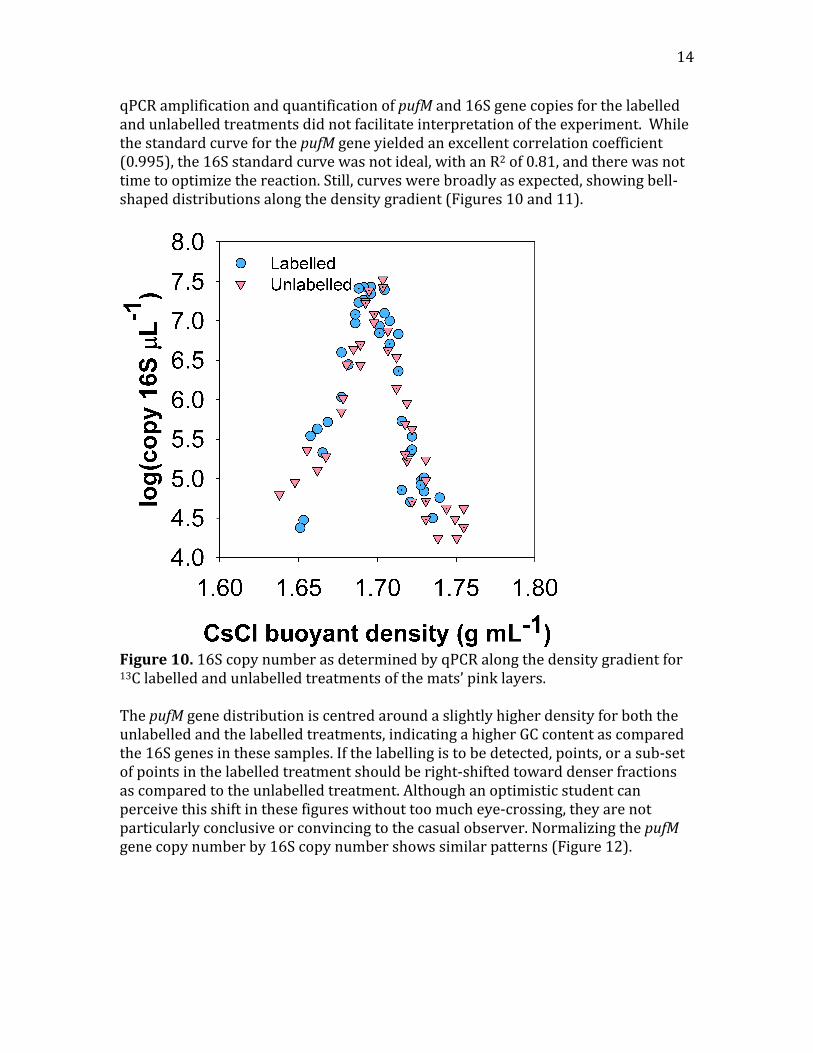
14
qPCRamplificationandquantificationofpufMand16Sgenecopiesforthelabelledandunlabelledtreatmentsdidnotfacilitateinterpretationoftheexperiment.WhilethestandardcurveforthepufMgeneyieldedanexcellentcorrelationcoefficient(0.995),the16Sstandardcurvewasnotideal,withanR2of0.81,andtherewasnottimetooptimizethereaction.Still,curveswerebroadlyasexpected,showingbell‐shapeddistributionsalongthedensitygradient(Figures10and11).
Figure10.16ScopynumberasdeterminedbyqPCRalongthedensitygradientfor13Clabelledandunlabelledtreatmentsofthemats’pinklayers.ThepufMgenedistributioniscentredaroundaslightlyhigherdensityforboththeunlabelledandthelabelledtreatments,indicatingahigherGCcontentascomparedthe16Sgenesinthesesamples.Ifthelabellingistobedetected,points,orasub‐setofpointsinthelabelledtreatmentshouldberight‐shiftedtowarddenserfractionsascomparedtotheunlabelledtreatment.Althoughanoptimisticstudentcanperceivethisshiftinthesefigureswithouttoomucheye‐crossing,theyarenotparticularlyconclusiveorconvincingtothecasualobserver.NormalizingthepufMgenecopynumberby16Scopynumbershowssimilarpatterns(Figure12).
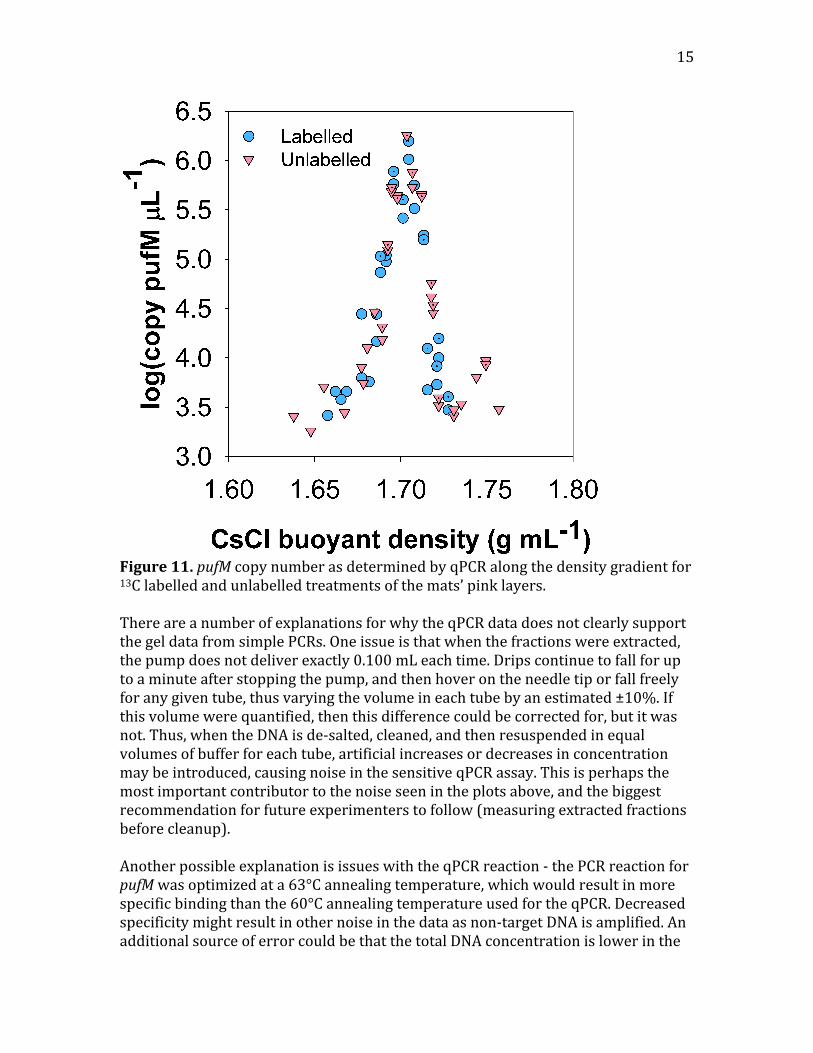
15
Figure11.pufMcopynumberasdeterminedbyqPCRalongthedensitygradientfor13Clabelledandunlabelledtreatmentsofthemats’pinklayers.ThereareanumberofexplanationsforwhytheqPCRdatadoesnotclearlysupportthegeldatafromsimplePCRs.Oneissueisthatwhenthefractionswereextracted,thepumpdoesnotdeliverexactly0.100mLeachtime.Dripscontinuetofallforuptoaminuteafterstoppingthepump,andthenhoverontheneedletiporfallfreelyforanygiventube,thusvaryingthevolumeineachtubebyanestimated±10%.Ifthisvolumewerequantified,thenthisdifferencecouldbecorrectedfor,butitwasnot.Thus,whentheDNAisde‐salted,cleaned,andthenresuspendedinequalvolumesofbufferforeachtube,artificialincreasesordecreasesinconcentrationmaybeintroduced,causingnoiseinthesensitiveqPCRassay.Thisisperhapsthemostimportantcontributortothenoiseseenintheplotsabove,andthebiggestrecommendationforfutureexperimenterstofollow(measuringextractedfractionsbeforecleanup).AnotherpossibleexplanationisissueswiththeqPCRreaction‐thePCRreactionforpufMwasoptimizedata63°Cannealingtemperature,whichwouldresultinmorespecificbindingthanthe60°CannealingtemperatureusedfortheqPCR.Decreasedspecificitymightresultinothernoiseinthedataasnon‐targetDNAisamplified.AnadditionalsourceoferrorcouldbethatthetotalDNAconcentrationislowerinthe

16
unlabelledthaninthelabelledsamples,resultinginnodetectionatlowerhigh‐enddensitiesthaninthelabelledsamples,resultinginanapparent,butincorrect,shifttowardheavyfractioninthelabelledfractions.Thisissomewhatlesslikelyofanexplanation,though,because(1)the16SqPCRdataarerelativelysimilarbetweensamples,withtheunlabelledsampleshavingsimilarlyhighmaximumcopynumbers,and(2)ifthiswerethecase,wewouldexpectthebrightestbandswithineachtreatmenttostilloccuratthesamedensity,whichtheydonot,asdiscussedabove.Themostappealingexplanationfortheapparentdiscrepancyisthattheshiftwasverysubtle,andwasdetectableusingsimplePCR,butwasobscuredintheerrorsintheqPCRdataintroducedbythefactorsdiscussedabove.Indeed,thedifferencein[CsCl]betweenthebrightestpeaksoftheunlabelledandlabelledfractionsisonly0.007g/mL,whichiswithinthenoiseoftheqPCRdata.
Figure12.pufM/16ScopynumberratioasdeterminedbyqPCRalongthedensitygradientfor13Clabelledandunlabelledtreatmentsofthemats’pinklayers.

17
ThereasonthisshiftwassubtleislikelyduetotherelativestocksandfluxesofHCO3‐inthemats.Whileseawatermaycontainaround2mMHCO3‐atanygiventime,itonlymakesupafractionofthetotalCinthemat.Forexample,ifthemat’sisonaverage~3%Cbymass(Rothermichetal.,2000),at130g/mat,adding2.88mgHCO3‐‐Conthreeoccasions,resultsinanincreaseintotalmatCofonly0.23%,eventhoughtheHCO3‐concentrationwouldbedoubledtoquadrupled.IfmatCturnoverishigh,throughheterotrophicaswellasphotosyntheticactivity,thentheaddedlabelwouldbequicklydiluted.ThishighlightsaclassicSIPquandary:howtoaddthelabelledsubstrateinachemicallyandbiologicallycomplexsystemwithoutriskingmassiveandenvironmentallyirrelevantoverloadononehandorlabeldilutionontheother.Infutureprojectswithinthescopeofthiscourse,thebestrecommendationmightbetobeheavy‐handedwithsubstrateadditionsinordertoseeclearresults,butifecologicalrelevanceisimportant,otheroptions,ordifferentconceptualframeworksmaybenecessary.4.3SIPCloneLibrariesandPhylogeneticAnalysesThecloningreactionsweresuccessfulfortheinitialandtwofinalmatDNAextractions,yieldingpickablecoloniesonallplates.(Thelonger,2+hourligationstepwaslikelyanimportantfactorinensuringefficientligation,andwouldberecommendedforfuture“functionalgene”clonelibraries.Theplateswithplasmid‐bearingcoloniesfromtheunlabelledtreatmentyieldedonly35coloniesintotal,whichwastobeexpected,sincelessDNAwasadded,inchoosingPCRproductsfromtheheavierfractionsonly.Theclonelibrariesallreturnedbetween85‐87%useablesequences,aftersequencetranslationandalignment.Theneighbour‐joiningtreeconstructedfromthetranslatedandalignedpufMsequencesindicatesdiversegroupsofpufMsequences(Figure13).Becausethelabelledfractionsaredistributedacrossthetreeandarenotclusterednearanysetofinitialsequences,itappearsthatanylabelincorporationintothepufMgenewasnotrestrictedtoasmallgroupofanoxygenicphototrophs.However,inaddition,thefewunlabelledclonesthatweresequencedappearthroughoutthetree.Lookingclosely,itappearsthatthepinkclonesthatdoshowuptendtobethosethatareheavierinGCcontent,whilethelabelledcloneshavebothhighandlowGCcontent.Thiscouldindicatethatthelow‐GCclonesfromthelabelledtreatmentwerepresentintheheavierfractionsonlybecauseoftheirincorporationofthe13Clabel.However,thisargumentissomewhattenuous,because,althoughthesamplestakenforcloningwerefromthesamefractionnumbers,thesefractionscorrespondedtoslightlydifferentCsClconcentrations(Table1).
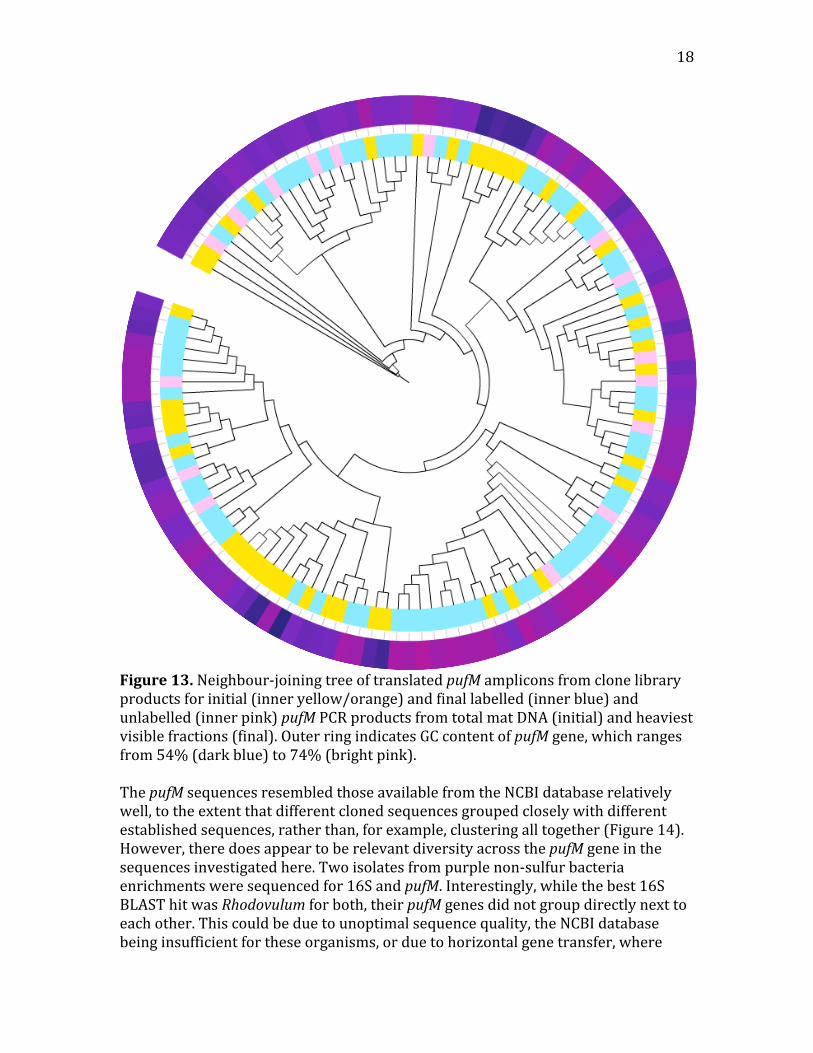
18
Figure13.Neighbour‐joiningtreeoftranslatedpufMampliconsfromclonelibraryproductsforinitial(inneryellow/orange)andfinallabelled(innerblue)andunlabelled(innerpink)pufMPCRproductsfromtotalmatDNA(initial)andheaviestvisiblefractions(final).OuterringindicatesGCcontentofpufMgene,whichrangesfrom54%(darkblue)to74%(brightpink).ThepufMsequencesresembledthoseavailablefromtheNCBIdatabaserelativelywell,totheextentthatdifferentclonedsequencesgroupedcloselywithdifferentestablishedsequences,ratherthan,forexample,clusteringalltogether(Figure14).However,theredoesappeartoberelevantdiversityacrossthepufMgeneinthesequencesinvestigatedhere.Twoisolatesfrompurplenon‐sulfurbacteriaenrichmentsweresequencedfor16SandpufM.Interestingly,whilethebest16SBLASThitwasRhodovulumforboth,theirpufMgenesdidnotgroupdirectlynexttoeachother.Thiscouldbeduetounoptimalsequencequality,theNCBIdatabasebeinginsufficientfortheseorganisms,orduetohorizontalgenetransfer,where

19
organismsofthesame“genus”couldhavepufMsequencesthataremoresimilartoorganismsofdifferentgenera.Futureworkherecouldincludedevelopingmoreisolatesfromwhichtosequenceboth16SandpufMgenes,andexpandingthisapproach.
Figure14.Neighbour‐joiningtreeoftranslatedpufMsequencesfromclonelibrariesandselectedpufMsequencesfromtheNCBIdatabase.Groupsarecollapsedinordertofacilitatevisualization,butarecolouredtoindicatewhetherclonesfromtheunlabelledlibraryarepresent.Forthosewheretheyarenotpresent,groupsincludeclonesfromthefinallabelledtreatmentand/orfromtheinitialmatsequencesandclonelibrary.5.ConclusionsandFutureDirections:ThepinklayersofmicrobialmatsinGreatSippewissettsaltmarshhavediversecommunitiesofanoxygenicphototrophsthatlikelyshiftoverthecourseoftheirdevelopment,withThiorhodovibriopossiblybeingafirstcolonizerofyoungmats.Theseanoxygenicphototrophslikelymakeuponlyasmallsubsetofthetotalbacterialpopulation(ontheorderof3‐10%ofthe45416Samplicons).However,theyarelikelyimportantdriversofCfixationinthislayer,althoughsupportofthisfindingfromSIPresultsisnotasrobustasitmighthavebeen,anditisnotapparentwhethermostoronlysometaxaareactive.Inaddition,pufMsequencesfromthese

20
matsresemblethosefrompublishedsequences,althoughsomediversityiscertainlyapparent,withsomesequencesnotmatchingpublishedsequencesverywell.Infutureresearch,itwouldbeveryinterestingtotakeamorephysiologicalapproachtothesequestions,measuringCcyclingexplicitly,andperhapstakingabroaderapproachtothecommunity,investigatingotherlayersofthemat,andeventransferofCcompoundsbetweenorganismswithinthemat.Inaddition,followingevolutionoftheanoxygenicphototrophicmatcommunitiesovertime,asmatsdevelop,oracrossafalsetimeseriessimilartotheapproachtakenhere,couldbeaninterestinglineofinvestigation.UsingaheavierlabelforSIP,oranotherapproachtoavoiddilutionthroughinherentCcyclinginthematcommunity,couldhelpproducelessequivocalresults.ForinvestigatingpufMdiversityfurther,itwouldbeidealtouseordevelopacuratedpufMdatabasetowhichsequencescouldbecompared,toensurebroadercoverageofexistingsequences.Includingmoreknownisolateswithboth16SandpufMsequenceswouldalsobeinteresting,andusingdegeneratepufMprimers,pufMprimersdesignedtotargetotheranoxygenicphototrophscouldalsohelpexpandfutureresearchquestions.6.Acknowledgements:Iwouldliketothankthecourseinstructors,DanandSteve,forthisamazingexperiencethissummerinWoodsHole.ParticularthanksgotoDanforhishelpwithSIPandSteveforthesweetjamsessionsonthedock.Inaddition,theTAsallprovidedinvaluableresourcesandsupportduringthecourse,especiallyChuck’shelpwithbioinformatics,Mallory’shelpwithqPCRandmyprojectdesign,Verena’shelpwiththeanoxygenicphototrophs,andSuiquan’shelpwiththeionchromatograph.ThanksalsotoAshleyforhertirelesssupportofthecourse.Iwouldalsoliketothankmyclassmatesforbeingsoamazing:Iamsogladtohavegottentoknowyouall,andamalreadyexcitedforthenexttimeourpathscross.Thiswholecoursehasbeensoinspiring,andIamexcitedtoseewhereittakesmenext.IwouldalsoliketothankthePfizerInc.EndowedScholarship,theMiltonL.ShifmanEndowedScholarship,andtheDOEgranttotheMicrobialDiversitycourseforfinancialsupport,aswellastheCornellCropandSoilSciencedepartment’ssummerfundingsupport.7.References:Achenbach,L.A.,Carey,J.,Madigan,M.T.(2001)PhotosyntheticandPhylogeneticPrimersforDetectionofAnoxygenicPhototrophsinNaturalEnvironments.AppliedandEnvironmentalMicrobiology,67:2922‐2926.Asao,M.,Pinkart,H.C.,Madigan,M.T.(2011)DiversityofextremophilicpurplephototrophicbacteriainSoapLake,aCentralWashington(USA)sodalake.EnvironmentalMicrobiology,13:2146‐2157.Bryant,D.A.,Frigaard,N.‐U.(2006)Prokaryoticphotosynthesisandphototrophyilluminated.TrendsinMicrobiology,14:488‐496.

21
Buckley,D.H.,Huangyutitham,V.,Hsu,Shi‐Fang,andNelson,T.A.(2007)Stableisotopeprobingwith15NachievedbydisentanglingtheeffectsofgenomeG+CcontentandisotopeenrichmentonDNAdensity.AppliedandEnvironmentalMicrobiology,73:3189‐3195.Hubas,C.,Jesus,B.,Passarelli,C.,Jenathon,C.(2011)Toolsprovidingnewinsightintocoastalanoxygenicpurplebacterialmats:reviewandperspectives.ResearchinMicrobiology,162:858‐868.Nicholson,J.A.M.,Stolz,J.F.,Pierson,B.K.(1987)StructureofamicrobialmatatGreatSippewissettMarsh,CapeCode,Massachusetts.FEMSMicrobialEcology,45:343‐364.Rothermich,M.M.,Guerrero,R.,Lenz,R.W.,Goodwin,S.(2000)Characterization,seasonaloccurrence,anddielfluctuationofPoly(hydroxyalkanoate)inphotosyntheticmicrobialmats.AppliedandEnvironmentalMicrobiology,66:4279‐4291.Werner,J.J.,Koren,O.,Hugenholtz,P.,DeSantis,T.Z.,Walters,W.A.,Caporaso,J.G.,Angenent,L.T.,Knight,R.,Ley,R.E.(2012)Impactoftrainingsetsonclassificationofhigh‐throughputbacterial16srRNAgenesurveys.TheISMEJournal,6:94:103.

22
AppendixTableA.145416SMetadataSampleID#
Description Furtherdescription
Origin Sitecharacteristics
LinkerPrimer
Thea3P Oldermat,adjacentto4P(<2cmapart)
AACCGTTAA
Thea4P Oldermat,adjacentto3P(<2cmapart)
TTACCGTTA
Thea5P
Toplayerofmatsamplecollectedin8.5cmdiameterx1.3cmdepthPetridish;centrestripcutoutwithrazor;0.44gpinklayerextracted;DNAamplifiedwithMoBioPowerBiofilmkit
Youngermat,nearby3Pand4P(<2maway)
ACCGGTTCC
TheaRB Redblobs0.1‐0.5cmwide,nestledinthemuckatthebaseofgrassbythewaterline
Uponinspection,seemedtohavetinywhiterectangles(bones?)
GreatSippewissettSaltMarsh,collectedatlowtideonJuly6,2012
AdjacentwaterpH=8.06,[SO42‐]=27mM
AACCTACTA
TableA.2CloneLibraryMetadataSampleID/File Description,origin,
sitecharacteristicsPrimersusedforamplification
PlateIDs
TLW_pufM23 FromSIPexperimentdescribedinthisreport‐labelledandunlabelledclones
CellsX1,X2,andA3,B3,C3,andD3allfromSIPexperiment’sunlabelledclones;Remainingcells(E3,F3,G3,H3,andX4‐X12)arefromSIPexperiment’s13C‐labelledclones
TLW_t0 CombinedsamplefromThea3PandThea4P(see454dataabove)
pufM557FandpufM750R
Half‐plate’sworthofinitialsample
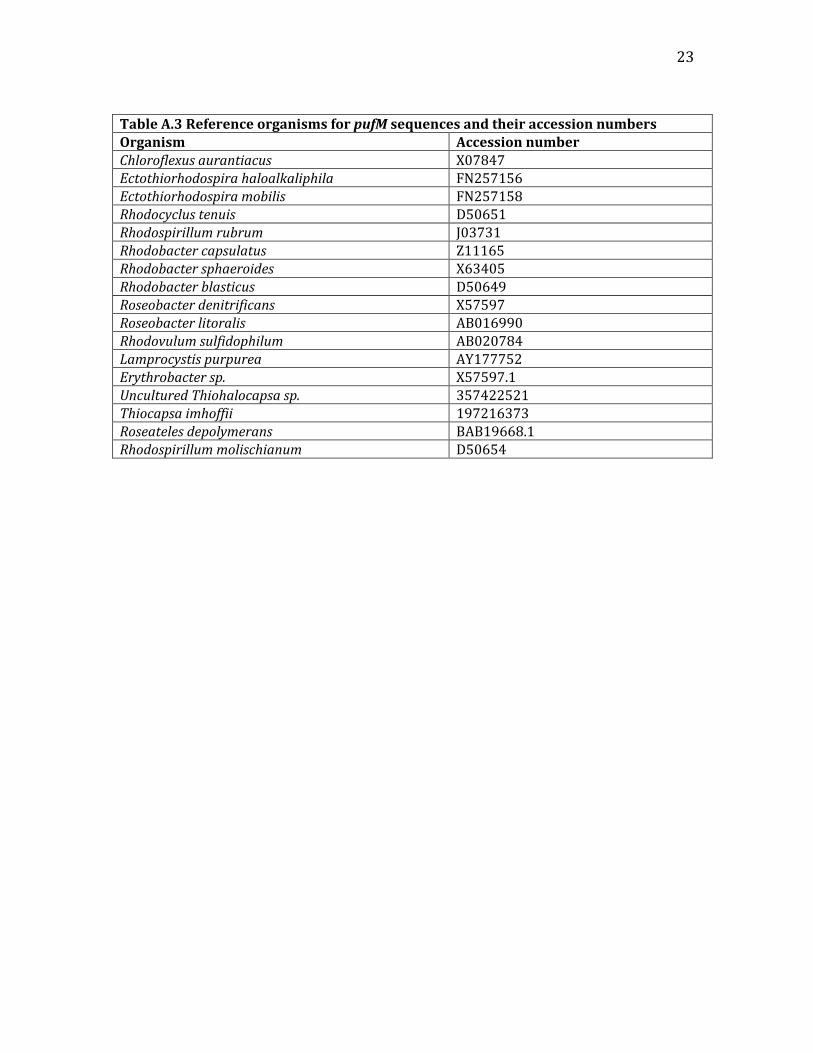
23
TableA.3ReferenceorganismsforpufMsequencesandtheiraccessionnumbersOrganism AccessionnumberChloroflexusaurantiacus X07847Ectothiorhodospirahaloalkaliphila FN257156Ectothiorhodospiramobilis FN257158Rhodocyclustenuis D50651Rhodospirillumrubrum J03731Rhodobactercapsulatus Z11165Rhodobactersphaeroides X63405Rhodobacterblasticus D50649Roseobacterdenitrificans X57597Roseobacterlitoralis AB016990Rhodovulumsulfidophilum AB020784Lamprocystispurpurea AY177752Erythrobactersp. X57597.1UnculturedThiohalocapsasp. 357422521Thiocapsaimhoffii 197216373Roseatelesdepolymerans BAB19668.1Rhodospirillummolischianum D50654