arginine vasopressin (avp) depletion in neurons of the suprachiasmatic nuclei affects the avp...
TRANSCRIPT

Arginine Vasopressin (AVP) Depletion inNeurons of the Suprachiasmatic Nuclei Affectsthe AVP Content of the ParaventricularNeurons and Stimulates AdrenocorticotrophicHormone ReleaseFelipe Gomez, Michele Chapleur, Brigitte Fernette, Claude Burlet,Jean-Pierre Nicolas, and Arlette Burlet*Laboratory of Cell Biology, INSERM U308, Nancy, France
Arginine vasopressin (AVP) produced in the hypotha-lamic suprachiasmatic nuclei (SCN) plays a role inestablishing neuroendocrine rhythms and, in particu-lar, in regulating the corticotrope axis rhythm. It hasrecently been shown that AVP from SCN inhibitscorticosteroid release. In order to investigate theinfluence of suprachiasmatic AVP on the differentpeptidergic systems through the hypothalamus, SCNneurons containing AVP were functionally lesioned byusing toxins associated with a cytotoxic monoclonalantibody (MAb) raised against AVP. Six days later, theAVP contents and AVP mRNA were measured indifferent hypothalamic and extrahypothalamic sites.Adrenocorticotrophic hormone (ACTH) concentra-tion was also measured in plasma. Microinjection ofthe AVP-MAb/toxin mixture into SCN brought abouta significant decrease in the AVP expression in SCN.This is demonstrated by the decrease in the AVPimmunoreactive content (24%, PF 0.01) and thedecrease of AVP hybridized mRNA (33%,PF 0.01).This points to the efficiency of the microinjection indecreasing the production of AVP in the injection area.Modifications of the AVP contents in the two subdivi-sions of the hypothalamic paraventricular nucleus(PVN) were also observed. AVP contents decreased inthe parvocellular subdivision (pPVN); this is coherentwith the AVP depletion in SCN since pPVN is themajor site of the SCN hypothalamic efferences. AVPcontent and AVP mRNA increased in the magnocellu-lar subdivision (mPVN); this also confirms the differ-ence in AVP synthesis regulation according to the PVNsubdivisions. The microinjection did not modify AVPexpression in supraoptic nuclei or oxytocin (OT)immunoreactive content in the main hypothalamicOT containing sites. Plasma ACTH values were double(PF 0.02) the values measured under non-specificIgG treatment 10 hr after lights on. This probably
resulted from the stimulation of the hypothalamo-pituitary-adrenal system since corticotrophin-releas-ing hormone (CRH) mRNA increased simultaneouslyby 24% (PF 0.05) in the PVN and the immunoreac-tive CRH content of the median eminence significantlydecreased (26%,PF 0.05). Overall, our data confirmthat AVP produced in the SCN inhibits the CRH-adrenocorticotrope axis in normal conditions, probablybecause of SCN projections of AVP neurons on the PVN.J. Neurosci. Res. 50:565–574, 1997.r 1997 Wiley-Liss, Inc.
Key words: vasopressin; corticotrophin-releasing fac-tor; radioimmunoassay; quantitative in situ hybridiza-tion; neuron network
INTRODUCTIONThe suprachiasmatic nucleus (SCN) of the hypo-
thalamus is a structure which has an essential role inestablishing the circadian rhythm in different physiologi-cal functions. Destruction of SCN will lead to total andirreversible loss of certain rhythms (Moore and Eichler,1972; Stephan and Nunez, 1977). More specifically, thediurnal rhythm of corticosteroid secretion (CS), whichdepends on the hypothalamic-pituitary-adrenal axis, stemsfrom the SCN. After destruction of these nuclei thecorticosterone levels lose the circadian rhythm and re-main in the morning range concentrations (Moore andEichler, 1972; Coen and MacKinnon, 1976; Abe et al.,1979) or show abnormal values (Raisman and Brown-Grant, 1977; Szafarczyk et al., 1979; Kalsbeek et al.,
*Correspondence to: Dr. Arlette Burlet, Laboratory of Cell Biology,INSERM U308, 38, rue Lionnois, 54000 Nancy, France. E-mail:[email protected]
Received 22 October 1996; Revised 10 June 1997; Accepted 10 June1997
Journal of Neuroscience Research 50:565–574 (1997)
r 1997 Wiley-Liss, Inc.

1992). This axis can be modulated by regulating either thecorticotrophin-releasing hormone (CRH) produced by theparaventricular nucleus (PVN) of the hypothalamus or theadrenocorticotrophic hormone (ACTH) in the pituitary gland.Liberation of ACTH into general circulation, in response tothe CRH, results in the release of corticosteroids from theadrenal glands (Keller-Wood and Dallmann, 1984).
Kalsbeek et al. (1992) have shown that argininevasopressin (AVP) from the dorsomedial SCN inhibitsCS through the hypothalamic paraventricular/dorsome-dian relay. There is direct and indirect evidence to supportthe hypothesis that this AVP plays a role in establishingneuroendocrine rhythms. There has been evidence ofendogenous rhythmic release of AVP from the SCN inSCN explants (Earnest and Sladek, 1986; Gillette andReppert, 1987) and rhythmic AVP expression has alsobeen reported (Noto et al., 1983; Yamase et al., 1991;Tominaga et al., 1992).
Anatomical studies have also demonstrated theexistence of AVP projections from the SCN into therostral PVN and subparaventricular region of the hypo-thalamus (Hoormeman and Buijs, 1982; Watts et al.,1987). There is no evidence of a direct connectionbetween AVP projections from the SCN and parvocellularCRH-producing neurons in the PVN. Buijs et al. (1993)concluded that the SCN transmits information related toCS via interneurons in and around the PVN. Thisconclusion is supported by the fact that the PVN hasextensive intranuclear connections (Van der Pol andPowley, 1978) which may enable the information to bepassed on to the CRH-containing neurons or to anotherrelay in the axis. In this paper, we describe anothermethod of investigation of the AVP relationship betweenSCN and PVN. A mixture containing cellular toxins and aspecific antibody to AVP [AVP-monoclonal antibody(MAb)/toxins] was microinjected directly into the SCN.We have previously reported specific inhibition of AVPrelease (Burlet et al., 1992a) and synthesis (Walker et al.,1997) when AVP-MAb/toxins were microinjected intothe supraoptic nucleus (SON) or PVN, respectively. Wealso showed that the incorporation of a specific radioac-tive tracer was totally inhibited in AVP-producing neu-rons (Burlet et al., 1991) and our hypothesis is that themicroinjection decreased AVP synthesis at the injectionsite. In our present experiment, we checked the efficiencyof the microinjection into SCN by measuring the AVPexpression in and outside the injection site. The plasmalevel of ACTH was also measured, bearing in mindBuijs’s hypothesis.
MATERIALS AND METHODSSixty male adult Long Evans rats (250–300 g) were
kept in individual cages under controlled temperature
(23 6 1°C) and lighting (12/12 hr dark/light cycle).Drinking water and food (powdered lab chow) wereprovided ad libitum. After 15 days habituation, stainlesssteel guide cannulae (11 mm G-27) were implantedbilaterally 2.5 mm above the SCN using a stereotaxicapparatus (A/P:20.3 from b; L: 60.3; D: 5.4 mm;Paxinos and Watson, 1982). The rats were allowed torecover for 4 days and then a Silastic catheter was placedin the right jugular artery of each rat. The catheter wasconnected to a stainless steel cannula (G-20). Thiscannula and the two guide cannulae were fixed to theskull by dental cement. The blood sampling cannularemained closed by a plastic cap. Injections of antibody/toxins mixture were given 5 days after the secondoperation.
Antibody/Toxins MixtureThe mouse monoclonal antibody (AVP-MAb) was
prepared from mouse hybridoma generously provided byP.Y. Couraud (Institut Pasteur, Paris, France). It belongsto the IgG 2a isotype. AVP-MAb cross-reactivity wasstudied by radioimmunoassay against different AVP struc-tural analogues (Burlet et al., 1992a). It increases diuresisand decrease AVP excretion when it is injected insupraoptic or paraventricular nuclei (Burlet et al., 1992a).These biological effects do not depend on the plasmacomplement. In this experiment, the mixture was injectedas an ascites fluid (5 mg/ml IgG) mixed (v/v) with cellulartoxins: the ricin A chain (100 nM) and monensin (50 nM).The ricin A chain does not penetrate living cells by itself,but once inside, it denaturates the 60S ribosomal subunitand thus inhibits protein synthesis (Carrasco et al., 1975;Endo and Tsurugi, 1987). Monensin is a carboxylicionophore which increases the intralysosomal pH anddecreases the intracellular degradation of the ricin Achain (Grimde, 1983). A group of 30 rats (treated group)was injected with the AVP-MAb/toxins mixture.
Non-specific IgG (nsIgG) was prepared by repeatedammonium sulfate (50%) precipitation of rat serum,followed by dialysis against 0.9% saline. Immunocyto-chemistry was carried out with the IgG preparation to ruleout the possibility of any specific cerebral epitope recog-nition. A mixture of toxins and nsIgG (instead of theAVP-MAb/toxins mixture) was injected into the 30 ratcontrol group.
Cerebral MicroinjectionThe rats were kept in their home cages and were
allowed to move freely. The mixture containing toxinsand either AVP-MAb or nsIgG was injected (0.25 µl/nucleus) using steel needles (Hamilton, 33-gauge, 12 mmlength) under continuous slow pressure over a period of 5min (Harvard Apparatus syringe infusion pump 22,
566 Gomez et al.

Dover, MA). Each rat received one daily injection on 3successive days 2 hr after the beginning of the light cycle.
Blood SamplingA plastic catheter full of a heparin solution (1
mg/ml) was connected to the blood sampling cannula.Two hundred microliters of blood was collected beforeand after the central microinjection, and in both cases 6 hrand 10 hr after the beginning of the light cycle. Sampleswere transferred to a 1 mlplastic tube to be centrifuged(10 mm at 2,500 T/min). One hundred microliters ofsupernatant plasma was recovered for further analysis.
Radioimmunological assay (RIA) of AVP, Oxytocin(OT), CRH, and ACTH
Six days after the final injection, the rats weredecapitated, and the brains were collected and stored at280°C. Serial sections of 300 µm were cut and discretehypothalamic sites were micropunched, namely, medial(MPO) and lateral preoptic nucleus (LPO), SCN, magno-cellular division (mNPV) and parvocellular division(pNPV) of paraventricular nucleus, supraoptic nucleus(SON), arcuate nucleus (ARC), lateral hypothalamus(LH), bed nucleus of the stria terminalis (BST), retrochi-asmatic subdivision of supraoptic nucleus (SON-R),ventromedian nucleus (VMN), and dorsomedian nucleus(DMN). Other sites were microdissected: the centralnucleus of the amygdala (ACe), the median eminence(ME), and the neurohypophysis (NH).
Bilateral tissue samples were placed in 500 µl 0.2 NHCl in microfuge tubes and stored at280°C untilassayed.
AVP and OT contents of the micropunched areaswere measured using RIA developed in our laboratoryand previously described (Burlet et al., 1992b). AVP andOT were extracted by ultrasonic disruption and proteinconcentrations were determined from an aliquot (100 µl)of homogenate by the Lowry et al. (1951) method. Therest of the homogenate was centrifuged at 4000 rpm for20 min at 4°C. The supernatants were then divided inorder to make a separate estimation of AVP and OTcontents. AVP antibody (P1) was raised in rabbit. Itcross-reacts 100% with AVP and Lys-vasopressin, 1.25%with vasotocin, 0.07% with pressinoic acid, and 0.002%with OT. Assay sensitivity is 1 pg/tube; intra- andinterassay variations are 1.4% and 12%, respectively. OTantibody (L15) was also raised in rabbit. It cross-reacts100% with OT and mesotocin and does not cross-reactwith the precursor molecule of OT. Its cross-reactivitywith isotocin, tocinoic acid, and AVP is less than 0.001%.Assay sensitivity is 1 pg/tube; intra- and interassayvariations are 3% and 8%, respectively.
CRH content was extracted by ultrasonic disruptionof the areas micropunched from two other groups of rats.Supernatants were lyophilized and reconstituted in RIAbuffer [0.05 M Tris-HCl, 1% bovine serum albumin(BSA), 0.1% Triton-X-100, pH 7.5] and then incubatedwith CRH antibody (d5 1:10,000; Neosystem,Strasbourg, France) at 4°C for 48 hr. The tracer (125I-Tyrrat/human CRH, 2,200 Ci/mM; NEN, Boston, MA) wasadded. Horse serum (100 µl) was added before sharingbound peptide with charcoal-dextran mixture. After cen-trifugation (4,000 rpm for 30 min at 4°C), the sampleradioactivity was measured in the supernatant using agamma counter coupled to a microcomputer (MDA 312system, Kontron, Ve´lizy, France). CRH antibody reactedwith human or rat (Tyr)-CRH but did not cross-react(,0.001%) with AVP, OT, TRH, LHRH, or Substance P.The sensitivity of CRH RIA was 10 pg/tube. The intra-and interassay coefficients of variation were 6.5% and9%, respectively.
Plasma ACTH concentration (250 µl) and standardwere measured with the ACTH Allegro kit (MallinkrodtMedical, France). This used a rabbit anti-ACTH serumand the sensitivity of the assay was 1 pg/tube of plasma.The intra- and interassay coefficients of variation were4.2% and 6.4%, respectively.
In Situ Hybridization of AVP and CRH mRNATheAVP probe was a 27-base oligomer complemen-
tary to bases 964–990 of the AVP gene (Ivell and Richter,1984; Sherman et al., 1986). The CRH probe was a46-base oligomer directed against bases 523–567 of theCRH gene (Jingami et al., 1985). The probes were 38-endlabeled with35S and terminal deoxynucleotyl-transferasebefore being purified in Sephadex G25 columns (Tebu,LePerray-en-Yvelines, France).
Four rats per group were sacrificed under theconditions described above (RIA). The brains wereremoved and stored at280°C until sectioned in 12 µmfrontal sections. The brain sections of AVP-MAb- ornsIgG-treated rats were collected onto the same gelatin-coated slides, were treated with paraformaldehyde solu-tion (4%) for 10 min, and then were dehydrated in gradedethanol and finally stored at280°C until hybridization.All manipulations were performed under RNAse-freeconditions.
We performed in situ hybridization according to theLaurent-Huck and Felix (1991) protocol, carefully respect-ing the conditions required for quantification (homog-enous thickness of sections, simultaneous treatment ofcontrol and experimental brain sections, large quantity oflabeled probes in order to treat numerous slides simulta-neously, etc.). After a prehybridization treatment, thesections were recovered with 30 µl of hybridization buffercontaining 3–3.63 105 cpm 35S-labeled probe and
AVP-MAb/Toxins Into SCN and Plasma ACTH 567

coverslipped before being incubated at 37°C in a moistchamber. The imperfect hybrids were then disrupted bysuccessive washings. The sections were next dehydratedin graded ethanol, air-dried prior to being exposed tob-Max Hyperfilm (Amersham International, Buckingham-shire, England) for 3 days for AVP and 6 days for CRH.One slide of radioactive standards prepared from brainpaste was simultaneously exposed.
The sections were then dipped into a liquid emul-sion (NTB2, Kodak), developed in Kodak D19 after 10(AVP) and 30 (CRH) days of exposure, and fixed inthiosulfite. After being washed in water and dehydrated ingraded ethanol, the sections were mounted in Canadabalsam.
The specificity of the radioactive signal was checkedby the usual methods. The signal did not appear when thelabeled probe was omitted; it strongly decreased when a10-fold amount of unlabeled probe was added to labeledprobe, and disappeared when a 100-fold amount ofunlabeled probe was used.
The hybridized signals were measured on the filmautoradiograms from four adjacent sections of the medialregion of SCN, SON, or PVN. The optical density of thehybridized areas was quantified using a Biocom (Biocom,Les Ulis, France) image analysis system. Optical densi-ties were converted to dpm/mg of tissue, using theradioactive paste standards (Laurent-Huck and Felix,1991).
Statistical AnalysisData of two rat groups were recorded in several Stat
View program tables: one-way analysis of variance(ANOVA) and post-hoc tests were performed to analyzethe radioimmunoassay data and Mann-Whitney U-test forin situ hybridization data. A probability of 95% wasconsidered significant.
RESULTSDuring the microdissection and in situ hybridiza-
tion analysis, it was observed that the guide cannulaewere correctly located above the SCN and the tissue wasin good condition. The histological study revealed nomechanical damage and the cannula marks were observed2.5 mm above the injection site.
AVP Immunoreactive ContentsNeither the nsIgG nor AVP-MAb injections dis-
rupted the physiological relationships in the AVP immu-noreactive contents of the micropunched areas (Fig. 1).The highest AVP content was observed in the SON, wherein both groups of rats, the AVP value was 5-fold the valuein the SCN and double the value in the PVN. Preoptic
nuclei showed traces of AVP (,1 ng/mg of proteins) andin other AVP non-synthesizing nuclei such as the LH,DMN, and VMN the AVP content remained from 10 to 30times lower than in the SON.
The AVP-MAb injection specifically decreased theAVP content of SCN. Six days after the injection, itdecreased by 26% (F 1,285 8.35; t5 2.89; P , 0.01)the AVP immunoreactive content of SCN. It also induceda significant, albeit different, effect on the two PVNsubdivisions. The parvocellular division of the PVN(pPVN) revealed an AVP content which was lower (42%;F 1,285 4.26; t5 2.58;P , 0.02) in treated rats than incontrol rats. On the contrary, the AVP content in themagnocellular PVN division (mPVN) was higher intreated animals (51%; F 1,285 5.33; t5 2.51;P , 0.02)than in nsIgG-treated rats.
The AVP-MAb injection did not alter the AVPcontent of other brain sites such as the LPO, MPO, LH,DMN, VMN, and ARC. There was no significant differ-ence between treated and control animals. The AVPcontent was not significantly modified in the ME or in theNH (data not shown).
OT Immunoreactive ContentsThe microinjection of AVP-MAb/toxins mixture
into the SCN did not affect the OT content of micro-punched areas (Fig. 2). OT distribution was identical innsIgG and AVP-MAb injected rats. OT content in the
Fig. 1. Vasopressin contents of different hypothalamic sites ofLong Evans rats sacrificed 6 days after simultaneous injectionsof AVP-MAb and toxins (stripes, n5 16) or non-specific ratIgG and toxins (points, n5 16) near the SCN. Values representthe mean6 SEM. *P , 0.05 vs. control. **P , 0.01 vs.control. ARC, arcuate nuclei (n.); LH, lateral hypothalamus;mPVN, magnocellular subdivision of paraventricular n.; pPVN,parvocellular subdivision of paraventricular n.; SON-R, retro-chiasmatic subdivision of supraoptic n.; SON, antechiasmaticsubdivision of supraoptic n.
568 Gomez et al.

SON was double that in both PVN divisions and was 10times higher than the peptide content in the DMN andVMN.
CRH Immunoreactive ContentsThe microinjection of AVP-MAb/toxins mixture
into the SCN modified the CRH immunoreactive contentof some hypothalamic areas (Fig. 3; F 5,645 40.3;P , 0.001). It did not change the PVN content, butsignificantly decreased by 25% the content of ARC(P , 0.05) and by 26% the content of ME (P , 0.05). Itdid not affect the CRH content of the other sites (data notshown).
Quantitative In Situ HybridizationThe AVP-MAb injection specifically decreased the
AVP mRNA hybridized in the SCN (Figs. 4, 5): a 33%decrease (U5 5, P , 0.01) in the SCN when measured 6days later (Fig. 5A), but no change in the SON (Fig. 5B).The two PVN subdivisions were affected in differentways: a 23% increase in the mPVN (U5 10, P , 0.05;Figs. 4, 5B) and no change in the pPVN (Fig. 5B).
The AVP-MAb/toxins injection into the SCN in-creased by 24% the CRH mRNA hybridized in the PVN(Figs. 6, 7; U5 11,P , 0.05).
ACTH Level in Jugular PlasmaThe jugular plasma ACTH was significantly higher
in treated rats than in nsIgG injected or intact rats (Fig. 8;F 5,565 18.52;P , 0.001). It was equal in all groups ofrats when it was measured in the middle of the lightperiod. It significantly increased in all rats at the end ofthe dark period (P , 0.01), but in AVP-MAb-treated ratsit was double (P , 0.02) the value in nsIgG-treated ratsand increased by 48% (P , 0.05) the value in controlrats.
DISCUSSIONThis experiment shows that three microinjections of
an AVP-MAb/ricin A/monensin mixture into the SCN ofthe rat brain significantly decreased the AVP expressionin the SCN 6 days after the final injection. In addition, theAVP-MAb/toxin injection affected only the AVP expres-sion in the two hypothalamic nuclei, the SCN and thePVN. It increased CRH expression in the PVN, as well asplasma level of ACTH, and significantly decreased theimmunoreactive content of ME. This proves that theAVP-MAb/toxin stimulated the hypothalamo-pituitary-adrenal axis. We conclude that our results support thehypothesis of the inhibitory suprachiasmatic AVP’s roleon the hypothalamo-adrenocorticotrope axis of the rat.This inhibition partially disappeared after the immunologi-cal lesion of the suprachiasmatic AVP neurons.
The modifications in the AVP expression did notresult from either mechanical destruction of AVP neurons
Fig. 2. OT contents of different hypothalamic sites of LongEvans rats sacrificed 6 days after simultaneous injections ofAVP-MAb and toxins (stripes, n5 16) or non-specific rat IgGand toxins (points, n5 16) near the SCN. Values represent themean 6 SEM. There is no significant difference betweenAVP-MAb/toxin injected rats and control rats. ARC, arcuatenuclei (n.); DMN, dorsomedian n.; mPVN, magnocellularsubdivision of paraventricular n.; pPVN, parvocellular subdivi-sion of paraventricular n.; SON-R, retrochiasmatic subdivisionof supraoptic n.; SON, antechiasmatic subdivision of supraopticn.; VMN, ventromedian n.
Fig. 3. CRH contents of different hypothalamic sites of LongEvans rats sacrificed 6 days after simultaneous injections ofAVP-MAb and toxins (stripes, n5 16) or non-specific rat IgGand toxins (points, n5 16) near the SCN. Values represent themean6 SEM. *P , 0.05 vs. control. ARC, arcuate nuclei (n.);ME, median eminence; PVN, paraventricular n.
AVP-MAb/Toxins Into SCN and Plasma ACTH 569

in the SCN or from a non-specific reaction to theAVP-MAb injection: in all studied animals, the mark ofthe injecting needle was located just above the SCNwhich was intact, and the rats injected with the nsIgG
showed normal AVP balance as described previously(Burlet et al., 1992a).
The possibility of interferences between the in-jected monoclonal AVP antibody and the polyclonal
Fig. 4. Darkfield photomicrographs of brain coronal sections showing AVP hybridized mRNA in theSCN (A) and PVN (B) of rats sacrificed 6 days after the injection of non-specific IgG and toxins (A,B)orAVP-MAb and toxins (A8,B8) near the SCN. The immunological treatment decreased the hybridizedsignals in the SCN but increased the signals in the mPVN. CO, chiasma opticum. Bar5 0.1 mm.
570 Gomez et al.

antibody used in the specific RIA was ruled out for anumber of reasons. First, the injected antibody was notdetectable after the 5th hr. It was found stained inefferents far from the injection site 24 hr later (Burlet etal., 1987, 1991). Modifications in AVP expression wereobserved 6 days after the final AVP-MAb injection.Second, the AVP-MAb did not bind iodinated AVP andtherefore could not interfere with the polyclonal antibodyused in the specific RIA. Moreover, this polyclonalantibody was 1,000 times more efficient in binding AVPthan AVP-MAb. Consequently, any antigen-antibody as-sociation formed with the injected AVP-MAb would havebeen disrupted in the presence of our polyclonal AVPantibody.
The long-lasting decrease of AVP expression in theSCN proves the effectiveness of immunolesions in impair-ing AVP production in certain neurons. We have previ-ously reported that the use of toxins, associated toantibodies on the CRH parvocellular neurons in the PVN,inhibits the endocrine functions of the CRH neurons(Menzaghi et al., 1991, 1992a,b), decreases the CRHcontents (Menzaghi et al., 1993) and CRH mRNA(Walker et al., 1997) in the hypothalamus, and causesmorphological damages of the CRH neurons (Menzaghiet al., 1992a,b). We consider that the decreased AVPexpression in the SCN following the AVP-MAb/toxininjections proves the efficiency of this method. Percent-age of peptide level modifications may be considered low,but the immunotoxin activity was always variable in vivo,depending on numerous factors (internalization rate,intracellular distribution, target cell type; Iwahashi et al.,1993; Dosio et al., 1996).
Significant modifications of the AVP level andexpression resulting from the immunological lesion of theSCN/AVP neurons were restricted to brain areas which
are functionally linked with the SCN. The SCN and thePVN were the only sites in the whole peptide systemwhich were affected by the treatment. This observationagrees with the thesis that AVP from the SCN does notbelong to the AVP hypothalamus-pituitary system. AVPefferents from the SCN innervate periventricular sites aswell as the PVN, but do not show a specific relationshipwith the SON (Van der Pol and Powley, 1978). The AVPlevel decreased in pPVN where the major SCN efferentsproject, but increased in mPVN and did not change inSON, which is not known to connect SCN (Van der Poland Powley, 1978). Moreover, the OT system, which iscomposed of magnocellular neurons in the SON andPVN, was not perturbed, as long as the OT contentsremained unchanged. Functional lesion did not lead to awidespread modification of AVP expression in all of theAVP synthesis sites. This fact supports our previousstatement that the direct effect of the AVP-MAb/toxinsmixture was limited to the injection site and causes anindirect imbalance in the interconnected nuclei.
The existence of opposite effects in the two PVNsubdivisions supports the hypothesis that they are regu-lated by two different mechanisms. A similar phenom-enon has already been observed when AVP-MAb wasinjected into the PVN (Rafai et al., 1995). This decreasedthe AVP content of both pPVN and SCN, but increasedthat of mPVN. The increase of the AVP content of mPVNmay be caused by a decrease in inhibitory action of theAVP neurons in the SCN on those in the mPVN. Howcould these opposite effects in mPVN and pPVN occursimultaneously? Anatomical cartography of efferent AVPprojections from the SCN shows that these fibers inner-vate the two PVN divisions in different ways. Results ofPha-L tract-tracing combined with immunocytochemicalstudies have shown the existence of AVP projections fromthe SCN along the rostral and periventricular parts of thePVN (Buijs et al., 1993). However, neither these findingsnor axon degeneration studies have succeeded in demon-strating that there is a major input of AVP fibers from theSCN in the mPVN and in the medial pPVN (Hoormemanand Buijs, 1982; Buijs et al., 1993). This leads us to thinkthat the SCN could have a direct effect on the AVPneurons in the pPVN but not in the mPVN. Evidence ofthe existence of an indirect neuronal relationship supportsthe hypothesis that there is a relay via interneuronsbetween the SCN efferents and post-synaptic neurons inthe PVN (Buijs et al., 1993). Given the lack of detectableeffects in other nuclei, we would therefore suggest thatthe main peptidergic interaction of the AVP-containingneurons is established in the PVN.
The analysis of jugular plasma showed an increaseof the ACTH level in blood after the AVP-MAb/toxinsinjection. This is coherent with the findings of Kalsbeek
Fig. 5. A,B: Measurements of AVP hybridized mRNA indifferent hypothalamic nuclei of Long Evans rats sacrificed 6days after simultaneous injections of AVP-MAb and toxins(stripes, n5 4) or nsIgG and toxins (points, n5 4) near theSCN. Values represent the mean6 SEM. *P , 0.05 vs. control.
AVP-MAb/Toxins Into SCN and Plasma ACTH 571
et al. (1992) showing an approximately 7-fold increase ofthe basal corticosterone level after the infusion of an AVPantagonist in the PVN. These results confirm that PVNplays a role in increasing corticotrope activity resultingfrom SCN lesions. The AVP-MAb/toxin injections intothe SCN increased the CRH mRNA in PVN and bloodACTH level. It also decreased the CRH immunoreactivecontent of the ME which was usually associated withCRH hypersecretion (Hauger et al., 1994). Moreover, theAVP from the pPVN can act as a CRH agonist for ACTHrelease in the pituitary (Antoni, 1992; Franci et al., 1993).However, the injection decreased the AVP immunoreac-tive content in pPVN and did not affect the AVPhybridized mRNA. Our hypothesis is that the AVP-MAbinjection decreased AVP content of SCN efferents inpPVN, but increased CRH synthesis and release.
The present study contributes evidence that thedifferent peptidergic interactions could be an outlet forcircadian information from the SCN. AVP originating inthe SCN acts at the PVN level on AVP and CRH neurons.Plasma ACTH increased when AVP expression wasdepleted in the SCN. Our overall data strongly supportthe hypothesis that suprachiasmatic AVP has an inhibitoryeffect on the hypothalamus-pituitary adrenal system. Itremains to determine the differential effect of SCN lesionon the daily rhythm of plasma ACTH.
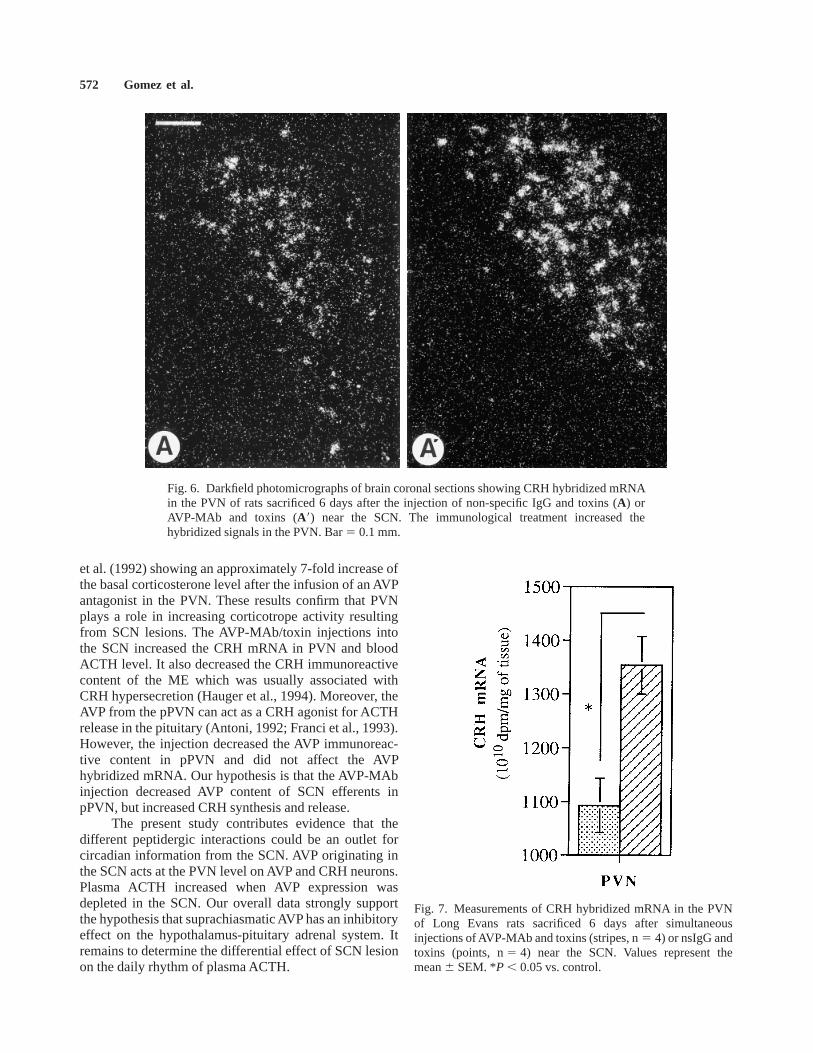
Fig. 6. Darkfield photomicrographs of brain coronal sections showing CRH hybridized mRNAin the PVN of rats sacrificed 6 days after the injection of non-specific IgG and toxins (A) orAVP-MAb and toxins (A8) near the SCN. The immunological treatment increased thehybridized signals in the PVN. Bar5 0.1 mm.
Fig. 7. Measurements of CRH hybridized mRNA in the PVNof Long Evans rats sacrificed 6 days after simultaneousinjections of AVP-MAb and toxins (stripes, n5 4) or nsIgG andtoxins (points, n5 4) near the SCN. Values represent themean6 SEM. *P , 0.05 vs. control.
572 Gomez et al.

ACKNOWLEDGMENTSWe thank Ms. A. Claudon and the members of the
Laboratory of Functional Explorations in Endocrinologyand Metabolism (Nancy, CHU) for performing the ACTHradioimmunoassays.
REFERENCES
Abe K, Kronig J, Greer AA, Critchlow (1979): Effects of destruction ofthe suprachiasmatic nuclei on the circadian rhythms in plasmacorticosterone, body temperature, feeding and plasma thyrotro-pin. Neuroendocrinology 29:119–131.
Antoni FA (1992): Vasopressinergic control of pituitary adrenocortico-tropin secretion comes of age. Front Neuroendocrinol 14:76–122.
Buijs RM, Markman M, Nunes-Cardoso B, Hou Y, Shinn S (1993):Projection of the rat suprachiasmatic nucleus to stress relatedareas in the rat hypothalamus: A light and electron microscopicstudy. J Comp Neurol 335:42–54.
Burlet A, Leon-Henri B, Robert F, Arahmani A, Fernette B, Burlet C(1987): Monoclonal anti-vasopressin (VP) antibodies penetrateinto VP neurons in-vivo. Exp Brain Res 65:629–638.
Burlet A, Haumont-Pellegri B, Tankosic P, Arahmani A, Fernette B,Nicolas JP, Burlet C (1991): The monoclonal antibody toneuropeptide: A new tool to act in vivo on peptidergic neuronactivity. In Ben Greenstein D (ed): ‘‘Neuroendocrine Research
Methods.’’ London: Hatwood Academic Publishers, pp 355–382.
Burlet A, Chateau-Chapleur M, Haumont-Pellegri B, Menzaghi F,Jansen B, Fernette B, Nicolas JP, Burlet C (1992a): Long-termreduction of vasopressin excretion induced by the centralinjection of an immunoconjugate (antibody to vasopressinlinked to ricin A chain). Neuroscience 50:965–973.
Burlet A, Jhanwar-Uniyal M, Chapleur-Chateau M, Burlet CR, Leibow-itz SF (1992b): Effect of food deprivation and refeeding on theconcentration of vasopressin and oxytocin in discrete hypotha-lamic sites. Pharmacol Biochem Behav 43:897–905.
Carrasco L, Fernandez-Puentes C, Vasquez D (1975): Effects of ricinon ribosomal sites involved in the interaction of the elongationfactors. Eur J Biochem 54:499–503.
Coen CW, MacKinnon PCB (1976): Serotonin involvement in oestro-gen-induced luteinizing hormone release in ovariectomised rats.J Endocrinol 71:49P–50P.
Dosio F, Franceshi A, Ceruti M, Brusa P, Cattel L, Colombatti M(1996): Enhancement of ricin toxin A chain immunotoxintoxicity: Synthesis, ionophoretic ability, and in vitro activity ofmonensin derivatives. Biochem Pharmacol 52:157–166.
Earnest D, Sladek CD (1986): Circadian rhythms of vasopressinrelease from individual rat suprachiasmatic explants in vitro.Brain Res 382:129–133.
Endo Y, Tsurugi K (1987): RNA N-glycosidase activity of ricinA-chain. Mechanism of action of the toxic lectin ricin eukary-otic ribosomes. J Biol Chem 262:8128–8130.
Franci CR, Anselmo-Franci JA, Kozlowski GP, McCann SM (1993):Actions of endogenous vasopressin and oxytocin on anteriorpituitary hormone secretion. Neuroendocrinology 57:693–699.
Gillette MU, Reppert SM (1987): The hypothalamic suprachiasmaticnuclei: Circadian patterns of vasopressin secretion and neuronalactivity in vitro. Brain Res Bull 19:135–139.
Grimde B (1983): Effects of carboxylic ionophores on lysosomalprotein degradation in rat hepatocytes. Exp Cell Res 149:27–31.
Hauger RL, Thrivikraman KV, Plostky PM (1996): Aged-relatedalterations of hypothalamo-pitutary-adrenal axis function inmale Fisher 344 rats. Endocrinology 134:1528–1536.
Hoorneman EMD, Buijs BRM (1982): Vasopressin fiber pathway inthe rat brain following suprachiasmatic nucleus lesioning. BrainRes 243:235–241.
Ivell R, Richter D (1984): Structure and comparison of the oxytocinand vasopressin genes from rat. Proc Natl Acad Sci USA81:2006–2010.
Iwahashi T, Okochi E, Ariyoshi K, Watake H, Amann E, Mori S, TsuruoT, Ono K (1993): Specific targeting and killing activity ofanti-P-glycoprotein monoclonal antibody MRK16 directedagainst intrinsically multidrug-resistant human colorectal carci-noma cell lines in the nude mouse model. Cancer Res 53:5475–5482.
Jingami H, Mizuno N, Takahashi H, Shibahara S, Furutani Y, Imura H,Numa S (1985): Cloning and sequence analysis of cDNA for ratcorticotropin-releasing factor precursor. FEBS Lett 191:63–68.
Kalsbeek A, Buijs RM, VanHeerikhuize JJ, Arts M, VanderWoude TP(1992): Vasopressin-containing neurons of the suprachiasmaticnuclei inhibit corticosterone release. Brain Res 580:62–67.
Keller-Wood M, Dallmann MF (1984): Corticosteroid inhibition ofACTH secretion. Endocrinology 5:1–24.
Laurent-Huck F, Felix JM (1991): Measurement of oxytocin andvasopressin gene expression by in situ hybridization. In ConnPM (ed): ‘‘Methods in Neuroscience.’’ Vol 5. London: Aca-demic Press, pp 159–182.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951): Proteinmeasurement with the Folin phenol reagent. J Biol Chem193:265–275.
Fig. 8. Plasma ACTH levels of Long Evans rats 6 and 10 hrafter the beginning of the light period: before treatment (emptycolumns, n5 20), or after AVP-MAb/toxins (stripes, n5 8) ornsIgG/toxins (points, n5 8) injection near the SCN. Valuesrepresent the mean6 SEM. All values measured 10 hr afterlights on are significantly higher (P , 0.01) than the respectivevalues measured 6 hr after lights on. ns: non-significant vs.nsIgG. *P , 0.05: treated vs. before treatment.`P , 0.02:treated vs. nsIgG.
AVP-MAb/Toxins Into SCN and Plasma ACTH 573

Menzaghi F, Burlet A, Van Oers JVAW, Tilders FH, Nicolas JP, BurletC (1991): Long term inhibition of stress induced ACTH releaseby intracerebral administration of a monoclonal antibody to ratcorticotropin releasing factor together with ricin A chain andmonensin. J Neuroendocrinol 3:469–475.
Menzaghi F, Burlet A, Chateau-Chapleur M, Nicolas JP, Burlet C(1992a): Alteration of pituitary-adrenal responses to adrenalec-tomy by the immunological targeting of CRH neurons. NeurosciLett 135:49–52.
Menzaghi F, Burlet A, Van Oers JVAW, Barbanel G, Tilders FH,Nicolas JP, Burlet C (1992b): A new perspective for the study ofcentral neuronal network implicated in stress. In Kvetnansky R,McCarty R, Axelrod J (eds): ‘‘Stress: Neuroendocrine andMolecular Approachs.’’ New York: Gordon Breach, pp 439–448.
Menzaghi F, Heinrichs S, Merlo Pich E, Tilders FH, Koob GF (1993):Functional impairment of hypothalamic corticotropin releasingfactor neurons with immunotargeted toxins enhances foodintake induced by neuropeptide Y. Brain Res 618:76–82.
Moore RY, Eichler VB (1972): Loss of circadian adrenal corticosteronerhythm following suprachiasmatic lesions in the rat. Brain Res42:201–206.
Noto T, Hashimoto H, Doi Y, Nakajima T, Kato N (1983): Biorhythmof arginine-vasopressin in the paraventricular, supraoptic andsuprachiasmatic nuclei of rats. Peptides 4:875–878.
Paxinos G, Watson C (1982): ‘‘The Rat Brain in Stereotaxic Coordi-nates.’’ New York: Academic Press.
Rafai H, Chateau-Chapleur M, Haumont-Pellegri B, Fernette B, AngelE, Burlet C, Nicolas JP, Burlet A (1995): The immunologicalimpairment of vasopressin (AVP) neurons into paraventricularnuclei modifies AVP expression in suprachiasmatic nuclei.Neurosci Lett 199:147–151.
Raisman G, Brown-Grant K (1977): The ‘‘suprachiasmatic syndrome.’’Endocrine and behaviour abnormalities following lesions of thesuprachiasmatic nuclei in female rats. Proc R Soc 198:417–438.
Sherman TG, McKelvy JF, Watson SJ (1986): Vasopressin mRNAregulation in individual hypothalamic nuclei: A Northern and insitu hybridization analysis. J Neurosci 6:1685–1694.
Stephan FK, Nunez AA (1977): Elimination of circadian rhythms indrinking activity, sleep and temperature by isolation of thesuprachiasmatic nucleus. Behav Biol 20:1–16.
Szafarczyk A, Ixart G, Malaval F, Nouguier-Soule´ J, Assenmacher I(1979): Effects of lesions of the suprachiasmatic nuclei andp-chlorophenylalanine on the circadian rhythms of adrenocorti-cotrophic hormone and corticosterone in the plasma, andlocomotor activity of rats. J Endocrinol 83:1–16.
Tominaga K, Shinihara K, Otori Y, Fukuhara C, Inouye ST (1992):Circadian rhythms of vasopressin content in the suprachias-matic nucleus of the rat. NeuroReport 3:809–812.
Van der Pol AN, Powley T (1978): A fine-granaid anatomical analysisof the role of the rat suprachiasmatic nucleus in circadianrhythms of feeding and drinking. Brain Res 160:307–326.
Walker CD, Tankosic P, Tilders FH, Burlet A (1997): Immunotargetedlesions of paraventricular CRH and AVP neurons in developingrats reveal the pattern of maturation of the systems and theirfunctional importance. J Neuroendocrinol 9:23–45.
Watts AG, Swanson LW, Sanchez-Watts G (1987): Efferent projectionsof the suprachiasmatic nucleus. Studies using anterogradetransport of Phaseolus vulgaris leucoagglutinin in rat. J CompNeurol 258:204–229.
Yamase K, Takahashi S, Nomura K, Kawashima S (1991): Circadianchanges in arginine vasopressin level in the suprachiasmaticnuclei in the rat. Neurosci Lett 130:255–258.
574 Gomez et al.