author’s choice © 2013 by the american society for ... · glycoproteomic analysis of the...
TRANSCRIPT

Glycoproteomic Analysis of the Secretome ofHuman Endothelial Cells*□S
Xiaoke Yin‡, Marshall Bern§, Qiuru Xing‡, Jenny Ho¶, Rosa Viner�, and Manuel Mayr‡**
Previous proteomics studies have partially unraveled thecomplexity of endothelial protein secretion but have notinvestigated glycosylation, a key modification of secretedand membrane proteins for cell communication. In thisstudy, human umbilical vein endothelial cells were kept inserum-free medium before activation by phorbol-12-my-ristate-13 acetate, a commonly used secretagogue thatinduces exocytosis of endothelial vesicles. In addition to123 secreted proteins, the secretome was particularly richin membrane proteins. Glycopeptides were enriched byzwitterionic hydrophilic interaction liquid chromatographyresins and were either treated with PNGase F and H2
18Oor directly analyzed using a recently developed workflowcombining higher-energy C-trap dissociation (HCD) withelectron-transfer dissociation (ETD) for a hybrid linear iontrap–orbitrap mass spectrometer. After deglycosylationwith PNGase F in the presence of H2
18O, 123 unique pep-tides displayed 18O-deamidation of asparagine, corre-sponding to 86 proteins with a total of 121 glycosylationsites. Direct glycopeptide analysis via HCD-ETD identified131 glycopeptides from 59 proteins and 118 glycosylationsites, of which 41 were known, 51 were predicted, and 26were novel. Two methods were compared: alternatingHCD-ETD and HCD-product-dependent ETD. The formerdetected predominantly high-intensity, multiply chargedglycopeptides, whereas the latter preferentially selectedprecursors with complex/hybrid glycans for fragmenta-tion. Validation was performed by means of glycoproteinenrichment and analysis of the input, the flow-through,and the bound fraction. This study represents the mostcomprehensive characterization of endothelial protein se-cretion to date and demonstrates the potential of newHCD-ETD workflows for determining the glycosylationstatus of complex biological samples. Molecular & Cel-lular Proteomics 12: 10.1074/mcp.M112.024018, 1–23,2013.
Cardiovascular disease manifests predominantly as myo-cardial ischemia, heart failure, stroke, aortic aneurysm, and
peripheral vascular disease and leads to the majority ofdeaths and disabilities worldwide. Endothelial cells (ECs) con-stitute the inner lining of all blood vessels and form the inter-face between the circulation and the vascular wall (1). Theendothelial monolayer is pivotal for maintaining vascular ho-meostasis through a balance of endothelium-derived factors(2, 3). ECs are preferred targets of cardiovascular risk factorssuch as hypercholesterolemia, diabetes, hypertension, andsmoking (1, 4). Repetitive injury is associated with a varyingdegree of endothelial dysfunction. Alterations in its anticoag-ulant and anti-inflammatory properties leave the vasculaturesusceptible to disease (5) and play a key role in the initiationand progression of cardiovascular disease (6).
Previous proteomics studies (7–13), including one by ourgroup (8), have investigated the secretome of unstimulatedhuman umbilical vein ECs (HUVECs), the most widely usedECs in cardiovascular research. Only two studies have ex-plored the secretome of HUVECs upon activation by shearstress (10) or with statin treatment (13) thus far. One studyused human microvascular ECs (9), which represent a distinctpopulation of ECs from small vessels. Yet many factors se-creted by ECs were not identified, probably because of theirlow abundance. In this study, we used a secretagogue, phor-bol ester phorbol-12-myristate-13-acetate (PMA) (14, 15), toinduce maximal protein release from serum-starved HUVECsover 45 min. In addition, we applied three different proteomicstrategies for the analysis of glycoproteins/glycopeptides tofurther enrich secreted proteins and characterize their glyco-sylation sites.
EXPERIMENTAL PROCEDURES
EC Culture—HUVECs (Lonza Group Ltd., Basel, Switzerland) werecultured on 0.1% gelatin-coated flasks in M199 medium supple-mented with 1 ng/ml endothelial cell growth factor (Sigma), 3 �g/mlendothelial growth supplement from bovine neural tissue (Sigma), 10U/ml heparin, 1.25 �g/ml thymidine, 10% fetal bovine serum (A15–108, PAA Laboratories, Velizy-Villacoublay, France), and 100 �g/mlpenicillin and streptomycin in a humidified incubator supplementedwith 5% CO2 at 37 °C. The cells were subcultured every 2 to 3 daysat a ratio of 1:4 (16).
Conditioned Medium Collection—HUVECs were cultured in com-plete medium until confluent. Then, they were washed and incubatedin M199 medium for 30 min twice before stimulation with 50 nM PMA(Sigma) in M199 medium for 45 min. The control group was incubatedwith M199 medium in the absence of PMA for 45 min. Conditionedmedia were collected and stored at �80 °C for further analysis.
Immunofluorescence Staining—HUVECs were cultured in Nuncchamber slides (Sigma-Aldrich) for 3 days. HUVECs were stimulated
From ‡The King’s British Heart Foundation Centre, King’s CollegeLondon, London SE5 9NU, UK; §Protein Metrics, San Carlos, CA94070; ¶Thermo Fisher Scientific, Hemel Hempstead, HP2 7GE, UK;�Thermo Fisher Scientific, San Jose, CA 95134
Author’s Choice—Final version full access.Received September 12, 2012, and in revised form, January 18,
2013Published, MCP Papers in Press, January 23, 2013, DOI
10.1074/mcp.M112.024018
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018
•
Research
Author’s Choice © 2013 by The American Society for Biochemistry and Molecular Biology, Inc.This paper is available on line at http://www.mcponline.org
Molecular & Cellular Proteomics 12.4 1
AQ: P
AQ: A
AQ: B

with 50 nM PMA in M199 medium for 45 min or incubated with M199medium for 45 min. The cells were fixed with 4% formaldehyde inPBS for 10 min, permeabilized with 0.1% Triton X-100 in PBS for 5min, and blocked in 5% fetal bovine serum in PBS for 30 min at 37 °C.Following 1 h of incubation with the primary antibodies, VE-cadherin(ab33168, Abcam, Cambridge, UK), and von Willebrand factor (vWF)(sc-8068, Santa Cruz Biotechnology, Santa Cruz, CA) at 37 °C, anAlexa Fluor® 594 conjugated donkey anti-rabbit IgG and an AlexaFluor® 488 conjugated donkey anti-goat IgG, respectively, wereadded, and the cells were incubated at 37 °C for 30 min. Nuclei werecounterstained with 4�,6-diamidino-2-phenylindole (D9542, Sigma)for 5 min. The slide was mounted in fluorescence mounting medium(DAKO, Denmark A/S, Glostrup, Denmark) and examined with anAxioPlan 2 fluorescence microscope (Carl Zeiss, Thornwood, NY)(17).
Proteomics Profiling of the Secretome—Conditioned media wereconcentrated with an Amicon spin column (3kD MWCO, EDO Milli-pore Corp., Billerica, MA) and separated via 4%–12% Bis-Tris SDS-PAGE (Invitrogen). Proteins were visualized via silver staining(PlusOne silver staining kit for proteins, GE Healthcare). Gel bandswere digested with modified trypsin (Promega Corp., Madison, WI)overnight on a ProGest digestion robot (Digilab Inc., Marlborough,MA) and analyzed via reverse-phase nano-flow HPLC (PepMap C18,3 �m, 100 Å, 25 cm � 75 �m inner diameter column, ThermoScientific) interfaced to an LTQ Orbitrap XL MS (Thermo Scientific)(18).
Deglycosylation—Concentrated media were mixed with deglyco-sylation buffer (150 mM NaCl, 50 mM sodium acetate, 10 mM EDTA,proteinase inhibitors, pH 6.8) supplemented with 0.05U PNGase F(Sigma), chondroitinase ABC (C3667, Sigma), and keratanase(G6920, Sigma) and incubated at 37 °C overnight (19).
Immunoblotting—Concentrated or deglycosylated media wereseparated via 4%–12% Bis-Tris gel (Invitrogen). Proteins were trans-ferred on a nitrocellulose membrane and blocked with 5% bovineserum albumin in PBS. Membranes were incubated with primaryantibody overnight at 4 °C. Secondary antibodies were incubated for1 h at room temperature. After the addition of ECL (GE Healthcare),the film was developed using a Compact X4 Automatic Processor(Xograph Healthcare Ltd., Stonehouse, UK). The following primaryantibodies were used: agrin (sc-25528, Santa Cruz Biotechnology),biglycan (ab54855, Abcam), connective tissue growth factor (sc-25440, Santa Cruz Biotechnology), fibronectin (sc-56391, Santa CruzBiotechnology), and lymphatic vessel endothelial hyaluronic acid re-ceptor 1 (AF2089, R&D Systems).
Difference Gel Electrophoresis—Conditioned media from HUVECstreated with or without PMA were concentrated using an Amicon spincolumn (3kD MWCO, Millipore) and the ReadyPrep 2D clean-up kit(Bio-Rad). The pellet was resuspended in difference gel electropho-resis lysis buffer (30 mM Tris, 8 M urea, 4% w/v CHAPS, proteaseinhibitors, pH 8.5). For each secretome sample, 15 �g of proteinswere labeled with Cy3 or Cy5. A dye swap was performed to excludepreferential labeling. Cellular extracts of HUVECs were labeled withCy2. Cy2-, Cy3-, and Cy5-labeled samples were separated via iso-electric focusing on immobilized pH gradient dry strips (18 cm, pH3–10 NL, GE Healthcare) with 30 KVH. The strips were equilibratedwith 10 mg/ml DTT in equilibration buffer (6 M urea, 2% w/v SDS, 30%v/v glycerol, 50 mM Tris, pH 8.8) for 15 min followed by 48 mg/mliodoacetamide in equilibration buffer for 15 min before separation viaSDS-PAGE at 100 W for 4 h using an Ettan DALTsix vertical electro-phoresis system (GE Healthcare) (20–22). Gels were scanned on anEttan difference gel electrophoresis imager (GE Healthcare). Imageswere overlaid with ImageQuant TL software (GE Healthcare). Com-mon spots present in both the cellular proteome and the secretomewere excised, digested with trypsin, and identified using nano-flow
HPLC-MS/MS. Detailed protocols are available on our researchgroup’s website.
Glycopeptide Enrichment—Conditioned media were desalted viathe use of Zeba spin columns (Thermo Scientific). Proteins were thenreduced by 5 mM DTT and alkylated with 25 mM iodoacetamide. Afteracetone precipitation overnight, the pellet was resuspended in 100mM triethylammonium bicarbonate (pH 8.5, Sigma) and digested withmodified trypsin (Promega) at 37 °C overnight. Peptides were labeledat a ratio of 100 �g peptides/0.8 mg Tandem Mass Tag Zero (TMT0)(Thermo Scientific) according to the manufacturer’s instruction. La-beled peptides were further enriched for glycopeptides using zwitte-rionic hydrophilic interaction liquid chromatography resin (Merck) (23).
LC/MS of Intact Glycopeptides—The glycopeptide enriched frac-tion was separated using the EASY-nLCTM nano-HPLC system(Thermo Scientific) with a Magic C18 spray tip 15 cm � 75 �m innerdiameter column (Bruker-Michrom, Auburn, CA). Gradient elution wasperformed with 4% to 30% acetonitrile in 0.1% formic acid over 60min at a flow rate of 300 nl/min. The samples were analyzed with anOrbitrap Elite hybrid MS with electron-transfer dissociation (ETD)(Thermo Scientific). The following MS and MS/MS settings were used:Fourier transform: MSn automatic gain control target � 5E4; MS/MS � 1 �scans, max ion time � 200 ms; MS � 300–1800 m/z,resolution � 60,000 at m/z 400, MS target � 1E6; dynamic exclu-sion � repeat count 1, duration 30 s, exclusion duration 90 s; higher-energy C-trap dissociation (HCD): collision energy � 35%, resolu-tion � 15,000; MSn target ion trap � 1E4, 2 �scans, max ion time �150 ms; ETD anion automatic gain control target � 2E5, charge-de-pendent ETD reaction time enabled. For alternating HCD-ETD MS/MS, the top 10 ions were analyzed. For HCD-product-dependentETD, the top 10 ions were analyzed via HCD, and product-dependentETD acquisition was triggered by product (oxonium) ions (m/z163.0812 for Hex; m/z 204.0864 for HexNAc; m/z 138.0554 for Hex-NAc fragment ion) (24).
Deglycosylation with PNGase F and H218O—Zwitterionic hydro-
philic interaction liquid chromatography resin enriched glycopeptideswere resuspended in 50 mM ammonium bicarbonate in H2
18O (97atom % 18O, Sigma) and deglycosylated with PNGase F (Sigma) for4 h at 37 °C. The samples were separated via reverse-phase nano-flow HPLC (PepMap C18, 3 �m, 100 Å, 25 cm � 75 �m innerdiameter column, Thermo Scientific) before analysis on an LTQ Or-bitrap XL MS (Thermo Scientific).
Glycoprotein Enrichment and LC/MS—ConA1 lectin resins (ThermoScientific) were used to enrich glycoproteins from concentrated con-ditioned media according to the manufacturer’s protocol. The input,glycoprotein-enriched fraction, and flow-through samples were sub-jected to trypsin digestion. The in-solution digests were separated ona Thermo Scientific Dionex UltiMate 3000 Rapid Separation LC(RSLC) system using a PepMap C18 column (3 �m, 100 Å, 50 cm �75 �m inner diameter column, Thermo Scientific). The rapid separa-tion LC system was interfaced to a Q Exactive MS (Thermo Scientific),and samples were analyzed using a top-10 HCD method.
Database Search and Data analysis—The following parameterswere used for different experiments.
(i) Gel-LC-MS/MS: Peak lists were generated by Mascot daemon(version 2.3.0, Matrix Science Ltd., London, UK) using extract_msn_com.exe and searched against the UniProt/Swiss-Prot mamma-
1 The abbreviations used are: ConA, concanavalin A; EC, endothe-lial cell; ETD, electron-transfer dissociation; GlcNAc, N-acetylgluco-samine; HCD, higher-energy C-trap dissociation; Hex, hexose;HexNAc, N-acetylhexosamine; HUVEC, human umbilical vein endo-thelial cell; PMA, phorbol-12-myristate-13-acetate; PNGase F, pep-tide: N-glycosidase F; TMT0, Tandem Mass Tag Zero; vWF, vonWillebrand factor.
Glycoproteomics of the Endothelial Secretome
2 Molecular & Cellular Proteomics 12.4
AQ: C
AQ: D
AQ: EAQ: F
AQ: G
AQ: H
AQ:H,I
AQ: J
AQ: K
AQ: L
AQ: M
Fn1
AQ: N
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

lian database (version 2012.03, 65,780 entries) using Mascot (version2.3.01, Matrix Science) with peptide tolerance � 10 ppm, MS/MStolerance � 0.8 Da, carbamidomethylation of cysteine as a fixedmodification, oxidation of methionine as a variable modification, anda maximum of two missed cleavage sites. The search results wereloaded into Scaffold software (version 3.6.2, Proteome Software Soft-ware, Inc., Portland, OR). A protein probability greater than 99%, apeptide probability greater than 95%, and a minimum number of twopeptides per protein were applied as filters to generate the protein list.Bovine contaminant proteins are listed separately.
(ii) PNGase F � H218O experiment: Thermo Scientific Proteome
Discoverer software version 1.3 was used to search against theUniProt/Swiss-Prot mammalian database (version 2012.03) usingMascot (version 2.3.01, Matrix Science) with a peptide tolerance of 10ppm; an MS/MS tolerance of 0.8 Da; carbamidomethylation of cys-teine as a fixed modification; oxidation of methionine, TMT0 label onlysine and peptide N-terminus, and deamidation (spontaneousdeamidation in ordinary water) and O18-deamidation (deglycosylationby PNGase F in H2
18O) of asparagine as variable modifications; and amaximum of two missed cleavage sites. Proteome Discoverer pro-duced a custom database containing 136 target proteins based onthis search.
(iii) Orbitrap Elite MS: Raw files were searched against the 136-protein database (along with reversed proteins as decoys) usingByonicTM (25) with a peptide tolerance of 10 ppm; an MS/MS toler-ance of 20 ppm for HCD and 0.6 Da for ETD; the carbamidomethy-lated cysteine, TMT0 label on lysine and peptide N-terminus as fixedmodifications; and oxidation of methionine, deamidation of aspara-gine and glutamine, and phosphorylation of serine and threonine asvariable modifications. ByonicTM allowed one N-glycan modificationon the N-X(not P)-S/T consensus motif per peptide, with mass andcomposition chosen from its “common human” glycan databasecontaining 350 glycan masses up to 6000 Da. Glycan modificationswere verified by the presence of corresponding glycan fragment ions,such as the HexNAc oxonium ion at 204.087 Da in HCD spectra.Peptide sequences were identified by ByonicTM from the ETD spectraand verified manually.
(iv) Q Exactive MS: Raw files were searched against the UniProt/Swiss-Prot human database (version 57.13, 20,266 entries) usingProteome Discoverer (version 1.3, Thermo Scientific) with Mascot(version 2.3.0, Matrix Science) and a peptide tolerance of 10 ppm, anMS/MS tolerance of 10 mmu, carbamidomethylation of cysteine as afixed modification, oxidation of methionine as a variable modification,and a maximum of two missed cleavage sites.
RESULTS
The Secretome of Activated ECs—HUVECs were stimu-lated with PMA, a commonly used secretagogue that inducesexocytosis of endothelial vesicles. As previously reported (26),the morphology of ECs changes from spindle-shaped toround upon PMA activation, and the rod-shaped Weibel-Pal-ade bodies, unique storage vesicles within ECs containingvWF and many other secreted proteins, fuse with the cellmembrane (Fig. 1A). In total, the secretomes of 17 primaryECs were analyzed via gel-LC-MS/MS, with or without degly-cosylation. Apart from 123 secreted proteins, the conditionedmedium of PMA-stimulated ECs was particularly rich in sur-face antigens and receptors, including many established en-dothelial markers (Table I). All identified proteins and peptidesare listed in supplemental Tables S1 and S2, respectively. Thedistribution of the frequencies and the cumulated distribution
of the number of samples in which proteins were identified areshown in supplemental Fig. S1. MS datasets of three biolog-ical replicates have been deposited in PRIDE (accession num-bers 26908–27003).
Immunoblots confirmed that proteins such as fibronectinand biglycan were constitutively secreted (Fig. 1B). Otherssuch as agrin and lymphatic vessel endothelial hyaluronic acidreceptor 1 were released upon PMA stimulation, providing an
FIG. 1. PMA treatment to stimulate EC secretion. Treatment ofHUVECs with PMA, a commonly used secretagogue, resulted in acharacteristic morphological change indicative of activation. A, im-munofluorescence staining of vWF (green) and VE-cadherin (red)shows the exocytotic effect of PMA. B, PMA increased protein se-cretion in the conditioned media as confirmed via immunoblotting. C,relative to previous studies, more than twice as many secreted andplasma membrane proteins were identified. D, overlay of intracellularand secreted proteins by means of difference gel electrophoresis. Inthe left-hand panel, proteins in conditioned media of HUVECs arestained in green (�PMA) and red (�PMA), and cellular proteins arestained in blue. Results were reproduced with different biologicalreplicates using reverse-labeling (right-hand panel: red, �PMA;green, �PMA). The protein corresponding to von Willebrand antigen2 is highlighted with a box. Common proteins in the secretome andthe cellular proteome are numbered in supplemental Fig. S2 andlisted in supplemental Table S3.
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 3
AQ: O
F1
T1
ZSI
ZSI
ZSIZSI,AQ:R
COLOR
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEI
Ext
race
llula
ran
dp
lasm
am
emb
rane
pro
tein
sid
entif
ied
inth
eH
UV
EC
-con
diti
oned
med
iaaf
ter
PM
Ast
imul
atio
n
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Cal
cium
ion-
bin
din
gp
rote
ins
Ann
exin
A1
AN
XA
1_H
UM
AN
P04
083
AN
XA
1P
lasm
am
emb
rane
Ann
exin
A2
aA
NX
A2_
HU
MA
NP
0735
5A
NX
A2
Ext
race
llula
rP
lasm
am
emb
rane
Ann
exin
A3
AN
XA
3_H
UM
AN
P12
429
AN
XA
3P
lasm
am
emb
rane
Cal
retic
ulin
CA
LR_H
UM
AN
P27
797
CA
LRE
xtra
cellu
lar
Gly
cop
rote
inC
alum
enin
CA
LU_H
UM
AN
O43
852
CA
LUE
xtra
cellu
lar
Gly
cop
rote
inC
alp
ain-
1ca
taly
ticsu
bun
itC
AN
1_H
UM
AN
P07
384
CA
PN
1P
lasm
am
emb
rane
Cal
pai
n-2
cata
lytic
sub
unit
CA
N2_
HU
MA
NP
1765
5C
AP
N2
Pla
sma
mem
bra
neC
alp
ain
smal
lsub
unit
1C
PN
S1_
HU
MA
NP
0463
2C
AP
NS
1P
lasm
am
emb
rane
Cal
synt
enin
-1C
STN
1_H
UM
AN
O94
985
CLS
TN1
Pla
sma
mem
bra
neG
lyco
pro
tein
Cal
synt
enin
-3C
STN
3_H
UM
AN
Q9B
QT9
CLS
TN3
Pla
sma
mem
bra
neG
lyco
pro
tein
Des
mog
lein
-1D
SG
1_H
UM
AN
Q02
413
DS
G1
Pla
sma
mem
bra
neG
lyco
pro
tein
Nuc
leob
ind
in-2
NU
CB
2_H
UM
AN
P80
303
NU
CB
2E
xtra
cellu
lar
Car
boh
ydra
tean
dgl
ycan
met
abol
ism
Alp
ha-a
myl
ase
1A
MY
1_H
UM
AN
P04
745
AM
Y1C
Ext
race
llula
rG
lyco
pro
tein
Exo
stos
in-l
ike
2E
XTL
2_H
UM
AN
Q9U
BQ
6E
XTL
2E
xtra
cellu
lar
Gly
cop
rote
inP
olyp
eptid
eN
-ace
tylg
alac
tosa
min
yltr
ansf
eras
e1
GA
LT1_
HU
MA
NQ
1047
2G
ALN
T1E
xtra
cellu
lar
Gly
cop
rote
inS
iala
teO
-ace
tyle
ster
ase
SIA
E_H
UM
AN
Q9H
AT2
SIA
EE
xtra
cellu
lar
Gly
cop
rote
inU
DP
-N-a
cety
lhex
osam
ine
pyr
opho
spho
ryla
seU
AP
1_H
UM
AN
Q16
222
UA
P1
Pla
sma
mem
bra
neC
oagu
latio
nan
dre
late
dp
rote
ins
Am
yloi
d-l
ike
pro
tein
2A
PLP
2_H
UM
AN
Q06
481
AP
LP2
Pla
sma
mem
bra
neG
lyco
pro
tein
Mul
timer
in-1
MM
RN
1_H
UM
AN
Q13
201
MM
RN
1E
xtra
cellu
lar
Gly
cop
rote
inP
lasm
inog
enac
tivat
orin
hib
itor
1P
AI1
_HU
MA
NP
0512
1S
ER
PIN
E1
Ext
race
llula
rG
lyco
pro
tein
Pla
smin
ogen
activ
ator
inhi
bito
r2
PA
I2_H
UM
AN
P05
120
SE
RP
INB
2E
xtra
cellu
lar
Gly
cop
rote
inTi
ssue
fact
orp
athw
ayin
hib
itor
TFP
I1_H
UM
AN
P10
646
TFP
IE
xtra
cellu
lar
Gly
cop
rote
inTi
ssue
fact
orp
athw
ayin
hib
itor
2TF
PI2
_HU
MA
NP
4830
7TF
PI2
Ext
race
llula
rG
lyco
pro
tein
Tiss
ue-t
ype
pla
smin
ogen
activ
ator
TPA
_HU
MA
NP
0075
0P
LAT
Ext
race
llula
rG
lyco
pro
tein
von
Will
ebra
ndfa
ctor
VW
F_H
UM
AN
P04
275
VW
FE
xtra
cellu
lar
Gly
cop
rote
inE
Cm
arke
rE
xtra
cellu
lar
mat
rixco
mp
onen
tsan
das
soci
ated
pro
tein
sA
grin
AG
RIN
_HU
MA
NO
0046
8A
GR
NE
xtra
cellu
lar
Gly
cop
rote
inC
olla
gen
alp
ha-2
(IV)
chai
nC
O4A
2_H
UM
AN
P08
572
CO
L4A
2E
xtra
cellu
lar
Gly
cop
rote
inC
olla
gen
alp
ha-1
(VI)
chai
nC
O6A
1_H
UM
AN
P12
109
CO
L6A
1E
xtra
cellu
lar
Gly
cop
rote
inC
olla
gen
alp
ha-1
(XII)
chai
nC
OC
A1_
HU
MA
NQ
9971
5C
OL1
2A1
Ext
race
llula
rG
lyco
pro
tein
Col
lage
nal
pha
-1(X
VIII
)ch
ain
CO
IA1_
HU
MA
NP
3906
0C
OL1
8A1
Ext
race
llula
rG
lyco
pro
tein
EG
F-co
ntai
ning
fibul
in-l
ike
extr
acel
lula
rm
atrix
pro
tein
1FB
LN3_
HU
MA
NQ
1280
5E
FEM
P1
Ext
race
llula
rG
lyco
pro
tein
Fib
rillin
-1FB
N1_
HU
MA
NP
3555
5FB
N1
Ext
race
llula
rG
lyco
pro
tein
Fib
rillin
-2FB
N2_
HU
MA
NP
3555
6FB
N2
Ext
race
llula
rG
lyco
pro
tein
Fib
rone
ctin
FIN
C_H
UM
AN
P02
751
FN1
Ext
race
llula
rG
lyco
pro
tein
Hya
luro
nan
and
pro
teog
lyca
nlin
kp
rote
in3
HP
LN3_
HU
MA
NQ
96S
86H
AP
LN3
Ext
race
llula
rLa
min
insu
bun
ital
pha
-4LA
MA
4_H
UM
AN
Q16
363
LAM
A4
Ext
race
llula
rG
lyco
pro
tein
Lam
inin
sub
unit
bet
a-1
LAM
B1_
HU
MA
NP
0794
2LA
MB
1E
xtra
cellu
lar
Gly
cop
rote
inLa
min
insu
bun
itga
mm
a-1
LAM
C1_
HU
MA
NP
1104
7LA
MC
1E
xtra
cellu
lar
Gly
cop
rote
inLy
sylo
xid
ase
hom
olog
2LO
XL2
_HU
MA
NQ
9Y4K
0LO
XL2
Ext
race
llula
rG
lyco
pro
tein
Mul
timer
in-2
MM
RN
2_H
UM
AN
Q9H
8L6
MM
RN
2E
xtra
cellu
lar
Gly
cop
rote
in
Glycoproteomics of the Endothelial Secretome
4 Molecular & Cellular Proteomics 12.4
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEI—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Nid
ogen
-1N
ID1_
HU
MA
NP
1454
3N
ID1
Ext
race
llula
rG
lyco
pro
tein
Nid
ogen
-2N
ID2_
HU
MA
NQ
1411
2N
ID2
Ext
race
llula
rG
lyco
pro
tein
Pro
lyl3
-hyd
roxy
lase
1P
3H1_
HU
MA
NQ
32P
28LE
PR
E1
Ext
race
llula
rG
lyco
pro
tein
Bas
emen
tm
emb
rane
-sp
ecifi
che
par
ansu
lfate
pro
teog
lyca
nco
rep
rote
inP
GB
M_H
UM
AN
P98
160
HS
PG
2E
xtra
cellu
lar
Gly
cop
rote
in
Big
lyca
nP
GS
1_H
UM
AN
P21
810
BG
NE
xtra
cellu
lar
Gly
cop
rote
inP
erox
idas
inho
mol
ogP
XD
N_H
UM
AN
Q92
626
PX
DN
Ext
race
llula
rG
lyco
pro
tein
SP
AR
CS
PR
C_H
UM
AN
P09
486
SP
AR
CE
xtra
cellu
lar
Gly
cop
rote
inTa
rget
ofN
esh-
SH
3TA
RS
H_H
UM
AN
Q7Z
7G0
AB
I3B
PE
xtra
cellu
lar
Gly
cop
rote
inTe
stic
an-1
TIC
N1_
HU
MA
NQ
0862
9S
PO
CK
1E
xtra
cellu
lar
Gly
cop
rote
inTh
rom
bos
pon
din
-1TS
P1_
HU
MA
NP
0799
6TH
BS
1E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Gro
wth
fact
ors
and
rela
ted
pro
tein
sC
-typ
ele
ctin
dom
ain
fam
ily11
mem
ber
AC
LC11
_HU
MA
NQ
9Y24
0C
LEC
11A
Ext
race
llula
rG
lyco
pro
tein
Cys
tein
e-ric
hm
otor
neur
on1
pro
tein
CR
IM1_
HU
MA
NQ
9NZ
V1
CR
IM1
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inC
onne
ctiv
etis
sue
grow
thfa
ctor
CTG
F_H
UM
AN
P29
279
CTG
FE
xtra
cellu
lar
Gly
cop
rote
inP
rote
inC
YR
61,
insu
lin-l
ike
grow
thfa
ctor
-bin
din
gp
rote
in10
CY
R61
_HU
MA
NO
0062
2C
YR
61E
xtra
cellu
lar
Dic
kkop
f-re
late
dp
rote
in3
DK
K3_
HU
MA
NQ
9UB
P4
DK
K3
Ext
race
llula
rG
lyco
pro
tein
Folli
stat
in-r
elat
edp
rote
in1
FSTL
1_H
UM
AN
Q12
841
FSTL
1E
xtra
cellu
lar
Gly
cop
rote
inH
epat
oma-
der
ived
grow
thfa
ctor
HD
GF_
HU
MA
NP
5185
8H
DG
FE
xtra
cellu
lar
Insu
lin-l
ike
grow
thfa
ctor
-bin
din
gp
rote
in2
IBP
2_H
UM
AN
P18
065
IGFB
P2
Ext
race
llula
rG
lyco
pro
tein
Insu
lin-l
ike
grow
thfa
ctor
-bin
din
gp
rote
in7
IBP
7_H
UM
AN
Q16
270
IGFB
P7
Ext
race
llula
rG
lyco
pro
tein
Late
nt-t
rans
form
ing
grow
thfa
ctor
bet
a-b
ind
ing
pro
tein
1LT
BP
1_H
UM
AN
Q14
766
LTB
P1
Ext
race
llula
rG
lyco
pro
tein
Late
nt-t
rans
form
ing
grow
thfa
ctor
bet
a-b
ind
ing
pro
tein
2LT
BP
2_H
UM
AN
Q14
767
LTB
P2
Ext
race
llula
rG
lyco
pro
tein
Neu
rona
lgro
wth
regu
lato
r1
NE
GR
1_H
UM
AN
Q7Z
3B1
NE
GR
1P
lasm
am
emb
rane
Gly
cop
rote
inIm
mun
ity-
and
infla
mm
atio
n-re
late
dp
rote
ins
Am
yloi
db
eta
A4
pro
tein
A4_
HU
MA
NP
0506
7A
PP
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inB
eta-
2-m
icro
glob
ulin
B2M
G_H
UM
AN
P61
769
B2M
Ext
race
llula
rG
lyco
pro
tein
Com
ple
men
tC
1qtu
mor
necr
osis
fact
or-r
elat
edp
rote
in5
C1Q
T5_H
UM
AN
Q9B
XJ0
C1Q
TNF5
Ext
race
llula
rC
omp
lem
ent
fact
orH
CFA
H_H
UM
AN
P08
603
CFH
Ext
race
llula
rG
lyco
pro
tein
Inte
rleuk
in-2
5,U
PF0
556
pro
tein
C19
orf1
0C
S01
0_H
UM
AN
Q96
9H8
C19
orf1
0E
xtra
cellu
lar
Gra
nulin
sG
RN
_HU
MA
NP
2879
9G
RN
Ext
race
llula
rG
lyco
pro
tein
Inte
rfer
on-i
nduc
edtr
ansm
emb
rane
pro
tein
1IF
M1_
HU
MA
NP
1316
4IF
ITM
1P
lasm
am
emb
rane
Gal
ectin
-1a
LEG
1_H
UM
AN
P09
382
LGA
LS1
Ext
race
llula
rG
alec
tin-3
LEG
3_H
UM
AN
P17
931
LGA
LS3
Ext
race
llula
rM
acro
pha
gem
igra
tion
inhi
bito
ryfa
ctor
aM
IF_H
UM
AN
P14
174
MIF
Ext
race
llula
rN
KG
2Dlig
and
2N
2DL2
_HU
MA
NQ
9BZ
M5
ULB
P2
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inP
entr
axin
-rel
ated
pro
tein
PTX
3P
TX3_
HU
MA
NP
2602
2P
TX3
Ext
race
llula
rG
lyco
pro
tein
Pro
tein
S10
0-A
7S
10A
7_H
UM
AN
P31
151
S10
0A7
Ext
race
llula
rP
rote
inS
100-
A8
S10
A8_
HU
MA
NP
0510
9S
100A
8E
xtra
cellu
lar
Pla
sma
mem
bra
neTu
bul
oint
erst
itial
nep
hriti
san
tigen
-lik
eTI
NA
L_H
UM
AN
Q9G
ZM
7TI
NA
GL1
Ext
race
llula
rG
lyco
pro
tein
Nuc
leas
e-se
nsiti
veel
emen
t-b
ind
ing
pro
tein
1Y
BO
X1_
HU
MA
NP
6780
9Y
BX
1E
xtra
cellu
lar
Zin
c-al
pha
-2-g
lyco
pro
tein
ZA
2G_H
UM
AN
P25
311
AZ
GP
1E
xtra
cellu
lar
Gly
cop
rote
in
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 5
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEI—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Mem
bra
nean
tigen
san
dre
cep
tors
HLA
clas
sI
hist
ocom
pat
ibili
tyan
tigen
,A
-24
alp
hach
ain
1A24
_HU
MA
NP
0553
4H
LA-A
Pla
sma
mem
bra
neG
lyco
pro
tein
HLA
clas
sI
hist
ocom
pat
ibili
tyan
tigen
,A
-30
alp
hach
ain
1A30
_HU
MA
NP
1618
8H
LA-A
Pla
sma
mem
bra
neG
lyco
pro
tein
HLA
clas
sI
hist
ocom
pat
ibili
tyan
tigen
,C
w-1
2al
pha
chai
n1C
12_H
UM
AN
P30
508
HLA
-CP
lasm
am
emb
rane
Gly
cop
rote
in
Alp
ha-2
-mac
rogl
obul
inre
cep
tor-
asso
ciat
edp
rote
inA
MR
P_H
UM
AN
P30
533
LRP
AP
1E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Bas
alce
llad
hesi
onm
olec
ule
BC
AM
_HU
MA
NP
5089
5B
CA
MP
lasm
am
emb
rane
Gly
cop
rote
inC
omp
lem
ent
com
pon
ent
C1q
rece
pto
rC
1QR
1_H
UM
AN
Q9N
PY
3C
D93
Pla
sma
mem
bra
neG
lyco
pro
tein
EC
mar
ker
Cad
herin
-13
CA
D13
_HU
MA
NP
5529
0C
DH
13P
lasm
am
emb
rane
Gly
cop
rote
inC
adhe
rin-2
CA
DH
2_H
UM
AN
P19
022
CD
H2
Pla
sma
mem
bra
neG
lyco
pro
tein
Cad
herin
-5C
AD
H5_
HU
MA
NP
3315
1C
DH
5P
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rC
D10
9an
tigen
CD
109_
HU
MA
NQ
6YH
K3
CD
109
Pla
sma
mem
bra
neG
lyco
pro
tein
CD
166
antig
enC
D16
6_H
UM
AN
Q13
740
ALC
AM
Pla
sma
mem
bra
neG
lyco
pro
tein
CD
44an
tigen
CD
44_H
UM
AN
P16
070
CD
44P
lasm
am
emb
rane
Gly
cop
rote
inC
D59
glyc
opro
tein
CD
59_H
UM
AN
P13
987
CD
59E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
CD
9an
tigen
CD
9_H
UM
AN
P21
926
CD
9P
lasm
am
emb
rane
Gly
cop
rote
inC
-typ
ele
ctin
dom
ain
fam
ily14
mem
ber
AC
LC14
_HU
MA
NQ
86T1
3C
LEC
14A
Pla
sma
mem
bra
neG
lyco
pro
tein
Dys
trog
lyca
nD
AG
1_H
UM
AN
Q14
118
DA
G1
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inE
ndog
linE
GLN
_HU
MA
NP
1781
3E
NG
Pla
sma
mem
bra
neG
lyco
pro
tein
EC
mar
ker
End
othe
lialp
rote
inC
rece
pto
rE
PC
R_H
UM
AN
Q9U
NN
8P
RO
CR
Pla
sma
mem
bra
neG
lyco
pro
tein
EC
mar
ker
Ep
hrin
typ
e-B
rece
pto
r4
EP
HB
4_H
UM
AN
P54
760
EP
HB
4P
lasm
am
emb
rane
Gly
cop
rote
inE
ndot
helia
lcel
l-se
lect
ive
adhe
sion
mol
ecul
eE
SA
M_H
UM
AN
Q96
AP
7E
SA
MP
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rLe
ucin
e-ric
hre
pea
ttr
ansm
emb
rane
pro
tein
FLR
T2FL
RT2
_HU
MA
NO
4315
5FL
RT2
Pla
sma
mem
bra
neG
lyco
pro
tein
Gua
nine
nucl
eotid
e-b
ind
ing
pro
tein
sub
unit
bet
a-2-
like
1aG
BLP
_HU
MA
NP
6324
4G
NB
2L1
Pla
sma
mem
bra
neH
LAcl
ass
Ihi
stoc
omp
atib
ility
antig
en,
alp
hach
ain
EH
LAE
_HU
MA
NP
1374
7H
LA-E
Pla
sma
mem
bra
neG
lyco
pro
tein
Inte
rcel
lula
rad
hesi
onm
olec
ule
1IC
AM
1_H
UM
AN
P05
362
ICA
M1
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rIn
terc
ellu
lar
adhe
sion
mol
ecul
e2
ICA
M2_
HU
MA
NP
1359
8IC
AM
2P
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rIn
tegr
inal
pha
-2IT
A2_
HU
MA
NP
1730
1IT
GA
2P
lasm
am
emb
rane
Gly
cop
rote
inIn
tegr
inal
pha
-5IT
A5_
HU
MA
NP
0864
8IT
GA
5P
lasm
am
emb
rane
Gly
cop
rote
inIn
tegr
inal
pha
-6IT
A6_
HU
MA
NP
2322
9IT
GA
6P
lasm
am
emb
rane
Gly
cop
rote
inIn
tegr
inb
eta-
1IT
B1_
HU
MA
NP
0555
6IT
GB
1P
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rP
rote
inja
gged
-1JA
G1_
HU
MA
NP
7850
4JA
G1
Pla
sma
mem
bra
neG
lyco
pro
tein
Pro
tein
jagg
ed-2
JAG
2_H
UM
AN
Q9Y
219
JAG
2P
lasm
am
emb
rane
Gly
cop
rote
inJu
nctio
nala
dhe
sion
mol
ecul
eA
JAM
1_H
UM
AN
Q9Y
624
F11R
Pla
sma
mem
bra
neG
lyco
pro
tein
BTB
/PO
Zd
omai
n-co
ntai
ning
pro
tein
KC
TD12
KC
D12
_HU
MA
NQ
96C
X2
KC
TD12
Pla
sma
mem
bra
neK
inec
tinK
TN1_
HU
MA
NQ
86U
P2
KTN
1P
lasm
am
emb
rane
Gly
cop
rote
inLy
soso
me-
asso
ciat
edm
emb
rane
glyc
opro
tein
1LA
MP
1_H
UM
AN
P11
279
LAM
P1
Pla
sma
mem
bra
neG
lyco
pro
tein
Low
-den
sity
lipop
rote
inre
cep
tor
LDLR
_HU
MA
NP
0113
0LD
LRP
lasm
am
emb
rane
Gly
cop
rote
inLo
w-d
ensi
tylip
opro
tein
rece
pto
r-re
late
dp
rote
in5
LRP
5_H
UM
AN
O75
197
LRP
5P
lasm
am
emb
rane
Gly
cop
rote
inLy
mp
hatic
vess
elen
dot
helia
lhya
luro
nic
acid
rece
pto
r1
LYV
E1_
HU
MA
NQ
9Y5Y
7LY
VE
1P
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rH
epat
ocyt
egr
owth
fact
orre
cep
tor
ME
T_H
UM
AN
P08
581
ME
TE
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Cat
ion-
ind
epen
den
tm
anno
se-6
-pho
spha
tere
cep
tor
MP
RI_
HU
MA
NP
1171
7IG
F2R
Pla
sma
mem
bra
neG
lyco
pro
tein
C-t
ype
man
nose
rece
pto
r2
MR
C2_
HU
MA
NQ
9UB
G0
MR
C2
Pla
sma
mem
bra
neG
lyco
pro
tein
Glycoproteomics of the Endothelial Secretome
6 Molecular & Cellular Proteomics 12.4
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEI—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Cel
lsur
face
glyc
opro
tein
MU
C18
MU
C18
_HU
MA
NP
4312
1M
CA
MP
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rN
euro
ligin
-1N
LGN
1_H
UM
AN
Q8N
2Q7
NLG
N1
Pla
sma
mem
bra
neG
lyco
pro
tein
Neu
rona
lcel
lad
hesi
onm
olec
ule
NR
CA
M_H
UM
AN
Q92
823
NR
CA
MP
lasm
am
emb
rane
Gly
cop
rote
inN
euro
pili
n-1
NR
P1_
HU
MA
NO
1478
6N
RP
1E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Neu
rop
ilin-
2N
RP
2_H
UM
AN
O60
462
NR
P2
Pla
sma
mem
bra
neG
lyco
pro
tein
Neu
rotr
imin
NTR
I_H
UM
AN
Q9P
121
NTM
Pla
sma
mem
bra
neG
lyco
pro
tein
Pro
toca
dhe
rin-1
0P
CD
10_H
UM
AN
Q9P
2E7
PC
DH
10P
lasm
am
emb
rane
Gly
cop
rote
inP
roto
cad
herin
-12
PC
D12
_HU
MA
NQ
9NP
G4
PC
DH
12P
lasm
am
emb
rane
Gly
cop
rote
inP
roto
cad
herin
gam
ma-
A11
PC
DG
B_H
UM
AN
Q9Y
5H2
PC
DH
GA
11P
lasm
am
emb
rane
Gly
cop
rote
inP
roto
cad
herin
gam
ma-
A12
PC
DG
C_H
UM
AN
O60
330
PC
DH
GA
12P
lasm
am
emb
rane
Gly
cop
rote
inP
roto
cad
herin
gam
ma-
B7
PC
DG
J_H
UM
AN
Q9Y
5F8
PC
DH
GB
7P
lasm
am
emb
rane
Gly
cop
rote
inP
roto
cad
herin
-1P
CD
H1_
HU
MA
NQ
0817
4P
CD
H1
Pla
sma
mem
bra
neG
lyco
pro
tein
Pro
toca
dhe
rin-9
PC
DH
9_H
UM
AN
Q9H
C56
PC
DH
9P
lasm
am
emb
rane
Gly
cop
rote
inP
rogr
amm
edce
lld
eath
1lig
and
2P
D1L
2_H
UM
AN
Q9B
Q51
PD
CD
1LG
2E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Pla
tele
ten
dot
helia
lcel
lad
hesi
onm
olec
ule
PE
CA
1_H
UM
AN
P16
284
PE
CA
M1
Pla
sma
mem
bra
neG
lyco
pro
tein
EC
mar
ker
Ple
xin-
D1
PLX
D1_
HU
MA
NQ
9Y4D
7P
LXN
D1
Pla
sma
mem
bra
neG
lyco
pro
tein
Inac
tive
tyro
sine
-pro
tein
kina
se7
PTK
7_H
UM
AN
Q13
308
PTK
7P
lasm
am
emb
rane
Gly
cop
rote
inR
ecep
tor-
typ
ety
rosi
ne-p
rote
inp
hosp
hata
sed
elta
PTP
RD
_HU
MA
NP
2346
8P
TPR
DP
lasm
am
emb
rane
Gly
cop
rote
inR
ecep
tor-
typ
ety
rosi
ne-p
rote
inp
hosp
hata
seF
PTP
RF_
HU
MA
NP
1058
6P
TPR
FP
lasm
am
emb
rane
Gly
cop
rote
inR
ecep
tor-
typ
ety
rosi
ne-p
rote
inp
hosp
hata
seka
pp
aP
TPR
K_H
UM
AN
Q15
262
PTP
RK
Pla
sma
mem
bra
neG
lyco
pro
tein
Pol
iovi
rus
rece
pto
rP
VR
_HU
MA
NP
1515
1P
VR
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inP
olio
viru
sre
cep
tor-
rela
ted
pro
tein
2P
VR
L2_H
UM
AN
Q92
692
PV
RL2
Pla
sma
mem
bra
neG
lyco
pro
tein
EC
mar
ker
Rou
ndab
out
hom
olog
1R
OB
O1_
HU
MA
NQ
9Y6N
7R
OB
O1
Pla
sma
mem
bra
neG
lyco
pro
tein
Rou
ndab
out
hom
olog
4R
OB
O4_
HU
MA
NQ
8WZ
75R
OB
O4
Pla
sma
mem
bra
neG
lyco
pro
tein
Syn
dec
an-4
SD
C4_
HU
MA
NP
3143
1S
DC
4E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Sem
apho
rin-4
DS
EM
4D_H
UM
AN
Q92
854
SE
MA
4DP
lasm
am
emb
rane
Gly
cop
rote
inS
emap
horin
-6B
SE
M6B
_HU
MA
NQ
9H3T
3S
EM
A6B
Pla
sma
mem
bra
neG
lyco
pro
tein
Tyro
sine
-pro
tein
pho
spha
tase
non-
rece
pto
rty
pe
sub
stra
te1
SH
PS
1_H
UM
AN
P78
324
SIR
PA
Pla
sma
mem
bra
neG
lyco
pro
tein
Sta
bili
n-1
STA
B1_
HU
MA
NQ
9NY
15S
TAB
1P
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rTr
ansf
errin
rece
pto
rp
rote
in1
TFR
1_H
UM
AN
P02
786
TFR
CE
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Tyro
sine
-pro
tein
kina
sere
cep
tor
Tie-
1TI
E1_
HU
MA
NP
3559
0TI
E1
Pla
sma
mem
bra
neG
lyco
pro
tein
Tyro
sine
-pro
tein
kina
sere
cep
tor
UFO
UFO
_HU
MA
NP
3053
0A
XL
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inV
ascu
lar
end
othe
lialg
row
thfa
ctor
rece
pto
r2
VG
FR2_
HU
MA
NP
3596
8K
DR
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rV
ascu
lar
end
othe
lialg
row
thfa
ctor
rece
pto
r3
VG
FR3_
HU
MA
NP
3591
6FL
T4E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
EC
mar
ker
Ver
ylo
w-d
ensi
tylip
opro
tein
rece
pto
rV
LDLR
_HU
MA
NP
9815
5V
LDLR
Pla
sma
mem
bra
neG
lyco
pro
tein
Mis
cella
neou
sm
emb
rane
pro
tein
sB
rain
acid
solu
ble
pro
tein
1B
AS
P1_
HU
MA
NP
8072
3B
AS
P1
Pla
sma
mem
bra
neD
naJ
hom
olog
sub
fam
ilyB
mem
ber
4D
NJB
4_H
UM
AN
Q9U
DY
4D
NA
JB4
Pla
sma
mem
bra
neR
NA
-bin
din
gp
rote
inE
WS
EW
S_H
UM
AN
Q01
844
EW
SR
1P
lasm
am
emb
rane
Nck
-ass
ocia
ted
pro
tein
1N
CK
P1_
HU
MA
NQ
9Y2A
7N
CK
AP
1P
lasm
am
emb
rane
Na(
�)/
H(�
)ex
chan
gere
gula
tory
cofa
ctor
NH
E-R
F2N
HR
F2_H
UM
AN
Q15
599
SLC
9A3R
2P
lasm
am
emb
rane
Pol
ymer
ase
Ian
dtr
ansc
ript
rele
ase
fact
orP
TRF_
HU
MA
NQ
6NZ
I2P
TRF
Pla
sma
mem
bra
neS
erum
dep
rivat
ion-
resp
onse
pro
tein
SD
PR
_HU
MA
NO
9581
0S
DP
RP
lasm
am
emb
rane
Sus
hire
pea
t-co
ntai
ning
pro
tein
SR
PX
2S
RP
X2_
HU
MA
NO
6068
7S
RP
X2
Ext
race
llula
rE
ryth
rocy
teb
and
7in
tegr
alm
emb
rane
pro
tein
STO
M_H
UM
AN
P27
105
STO
MP
lasm
am
emb
rane
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 7
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEI—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Mis
cella
neou
sse
cret
edp
rote
ins
Pep
tidyl
-gly
cine
alp
ha-a
mid
atin
gm
onoo
xyge
nase
AM
D_H
UM
AN
P19
021
PA
ME
xtra
cellu
lar
Gly
cop
rote
inA
ngio
poi
etin
-2A
NG
P2_
HU
MA
NO
1512
3A
NG
PT2
Ext
race
llula
rG
lyco
pro
tein
End
othe
lin-1
ED
N1_
HU
MA
NP
0530
5E
DN
1E
xtra
cellu
lar
End
othe
lialc
ell-
spec
ific
mol
ecul
e1
ES
M1_
HU
MA
NQ
9NQ
30E
SM
1E
xtra
cellu
lar
Gly
cop
rote
inP
rote
inFA
M3C
FAM
3C_H
UM
AN
Q92
520
WN
T16
Ext
race
llula
rE
pid
idym
alse
cret
ory
pro
tein
E1
NP
C2_
HU
MA
NP
6191
6N
PC
2E
xtra
cellu
lar
Gly
cop
rote
inP
rogr
amm
edce
lld
eath
pro
tein
10P
DC
10_H
UM
AN
Q9B
UL8
PD
CD
10P
lasm
am
emb
rane
Pro
lact
in-i
nduc
ible
pro
tein
PIP
_HU
MA
NP
1227
3P
IPE
xtra
cellu
lar
Gly
cop
rote
inS
ulfh
ydry
loxi
das
e1
QS
OX
1_H
UM
AN
O00
391
QS
OX
1E
xtra
cellu
lar
Gly
cop
rote
inS
ecre
togl
obin
fam
ily1D
mem
ber
2S
G1D
2_H
UM
AN
O95
969
SC
GB
1D2
Ext
race
llula
rTh
iore
dox
ina
THIO
_HU
MA
NP
1059
9TX
NE
xtra
cellu
lar
Thym
osin
bet
a-4
TYB
4_H
UM
AN
P62
328
TMS
B4X
Ext
race
llula
rP
rote
ase
inhi
bito
rsC
ysta
tin-C
CY
TC_H
UM
AN
P01
034
CS
T3E
xtra
cellu
lar
Gly
cop
rote
inLe
ukoc
yte
elas
tase
inhi
bito
rIL
EU
_HU
MA
NP
3074
0S
ER
PIN
B1
Ext
race
llula
rIn
ter-
alp
ha-t
ryp
sin
inhi
bito
rhe
avy
chai
nH
2IT
IH2_
HU
MA
NP
1982
3IT
IH2
Ext
race
llula
rG
lyco
pro
tein
Ser
pin
B9
SP
B9_
HU
MA
NP
5045
3S
ER
PIN
B9
Ext
race
llula
rM
etal
lop
rote
inas
ein
hib
itor
1TI
MP
1_H
UM
AN
P01
033
TIM
P1
Ext
race
llula
rG
lyco
pro
tein
Met
allo
pro
tein
ase
inhi
bito
r2
TIM
P2_
HU
MA
NP
1603
5TI
MP
2E
xtra
cellu
lar
Pro
teas
esA
ngio
tens
in-c
onve
rtin
gen
zym
eA
CE
_HU
MA
NP
1282
1A
CE
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inE
Cm
arke
rD
isin
tegr
inan
dm
etal
lop
rote
inas
ed
omai
n-co
ntai
ning
pro
tein
10A
DA
10_H
UM
AN
O14
672
AD
AM
10P
lasm
am
emb
rane
Gly
cop
rote
in
Am
inop
eptid
ase
BA
MP
B_H
UM
AN
Q9H
4A4
RN
PE
PE
xtra
cellu
lar
Am
inop
eptid
ase
NA
MP
N_H
UM
AN
P15
144
AN
PE
PP
lasm
am
emb
rane
Gly
cop
rote
inB
one
mor
pho
gene
ticp
rote
in1
BM
P1_
HU
MA
NP
1349
7B
MP
1E
xtra
cellu
lar
Gly
cop
rote
inC
athe
psi
nB
CA
TB_H
UM
AN
P07
858
CTS
BE
xtra
cellu
lar
Gly
cop
rote
inC
athe
psi
nD
CA
TD_H
UM
AN
P07
339
CTS
DE
xtra
cellu
lar
Gly
cop
rote
inC
athe
psi
nZ
CA
TZ_H
UM
AN
Q9U
BR
2C
TSZ
Ext
race
llula
rG
lyco
pro
tein
Car
box
ypep
tidas
eQ
CB
PQ
_HU
MA
NQ
9Y64
6C
PQ
Ext
race
llula
rG
lyco
pro
tein
Dip
eptid
ylp
eptid
ase
2D
PP
2_H
UM
AN
Q9U
HL4
DP
P7
Ext
race
llula
rG
lyco
pro
tein
Dip
eptid
ylp
eptid
ase
3D
PP
3_H
UM
AN
Q9N
Y33
DP
P3
Pla
sma
mem
bra
neE
ndop
lasm
icre
ticul
umam
inop
eptid
ase
1E
RA
P1_
HU
MA
NQ
9NZ
08E
RA
P1
Ext
race
llula
rG
lyco
pro
tein
Furin
FUR
IN_H
UM
AN
P09
958
FUR
INP
lasm
am
emb
rane
Gly
cop
rote
inG
amm
a-gl
utam
ylhy
dro
lase
GG
H_H
UM
AN
Q92
820
GG
HE
xtra
cellu
lar
Gly
cop
rote
inS
erin
ep
rote
ase
HTR
A1
HTR
A1_
HU
MA
NQ
9274
3H
TRA
1E
xtra
cellu
lar
Insu
lin-d
egra
din
gen
zym
eID
E_H
UM
AN
P14
735
IDE
Ext
race
llula
rP
lasm
am
emb
rane
Inte
rstit
ialc
olla
gena
seM
MP
1_H
UM
AN
P03
956
MM
P1
Ext
race
llula
rG
lyco
pro
tein
Str
omel
ysin
-2M
MP
10_H
UM
AN
P09
238
MM
P10
Ext
race
llula
rM
atrix
met
allo
pro
tein
ase-
14M
MP
14_H
UM
AN
P50
281
MM
P14
Pla
sma
mem
bra
ne72
kDa
typ
eIV
colla
gena
seM
MP
2_H
UM
AN
P08
253
MM
P2
Ext
race
llula
rG
lyco
pro
tein
Lyso
som
alP
ro-X
carb
oxyp
eptid
ase
PC
P_H
UM
AN
P42
785
PR
CP
Pla
sma
mem
bra
neG
lyco
pro
tein
Ser
ine
pro
teas
e23
PR
S23
_HU
MA
NO
9508
4P
RS
S23
Ext
race
llula
rG
lyco
pro
tein
Ub
iqui
tinca
rbox
yl-t
erm
inal
hyd
rola
se14
UB
P14
_HU
MA
NP
5457
8U
SP
14P
lasm
am
emb
rane
Glycoproteomics of the Endothelial Secretome
8 Molecular & Cellular Proteomics 12.4
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEI—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Sig
nalt
rans
duc
tion
pro
tein
sA
den
ylyl
cycl
ase-
asso
ciat
edp
rote
in1
CA
P1_
HU
MA
NQ
0151
8C
AP
1P
lasm
am
emb
rane
Cel
ldiv
isio
nco
ntro
lpro
tein
42ho
mol
ogC
DC
42_H
UM
AN
P60
953
CD
C42
Pla
sma
mem
bra
neC
onta
ctin
-ass
ocia
ted
pro
tein
-lik
e3
CN
TP3_
HU
MA
NQ
9BZ
76C
NTN
AP
3E
xtra
cellu
lar
Pla
sma
mem
bra
neG
lyco
pro
tein
Ad
apte
rm
olec
ule
crk
CR
K_H
UM
AN
P46
108
CR
KP
lasm
am
emb
rane
Ras
GTP
ase-
activ
atin
gp
rote
in-b
ind
ing
pro
tein
1G
3BP
1_H
UM
AN
Q13
283
G3B
P1
Pla
sma
mem
bra
neG
row
thar
rest
-sp
ecifi
cp
rote
in6
GA
S6_
HU
MA
NQ
1439
3G
AS
6E
xtra
cellu
lar
Gly
cop
rote
inIn
terf
eron
-ind
uced
guan
ylat
e-b
ind
ing
pro
tein
1G
BP
1_H
UM
AN
P32
455
GB
P1
Ext
race
llula
rG
uani
nenu
cleo
tide-
bin
din
gp
rote
inG
(i)su
bun
ital
pha
-2G
NA
I2_H
UM
AN
P04
899
GN
AI2
Pla
sma
mem
bra
neG
lyp
ican
-1G
PC
1_H
UM
AN
P35
052
GP
C1
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inH
edge
hog-
inte
ract
ing
pro
tein
HH
IP_H
UM
AN
Q96
QV
1H
HIP
Ext
race
llula
rP
lasm
am
emb
rane
Gly
cop
rote
inH
istid
ine
tria
dnu
cleo
tide-
bin
din
gp
rote
in1a
HIN
T1_H
UM
AN
P49
773
HIN
T1P
lasm
am
emb
rane
Inte
grin
-lin
ked
pro
tein
kina
seIL
K_H
UM
AN
Q13
418
ILK
Pla
sma
mem
bra
neR
asG
TPas
e-ac
tivat
ing-
like
pro
tein
IQG
AP
1IQ
GA
1_H
UM
AN
P46
940
IQG
AP
1P
lasm
am
emb
rane
cAM
P-d
epen
den
tp
rote
inki
nase
typ
eII-
alp
hare
gula
tory
sub
unit
KA
P2_
HU
MA
NP
1386
1P
RK
AR
2AP
lasm
am
emb
rane
Ras
-rel
ated
pro
tein
Rab
-18
RA
B18
_HU
MA
NQ
9NP
72R
AB
18P
lasm
am
emb
rane
Ras
-rel
ated
pro
tein
Rab
-5C
RA
B5C
_HU
MA
NP
5114
8R
AB
5CP
lasm
am
emb
rane
Ras
-rel
ated
C3
bot
ulin
umto
xin
sub
stra
te1
RA
C1_
HU
MA
NP
6300
0R
AC
1P
lasm
am
emb
rane
Ras
-rel
ated
pro
tein
Ral
-AR
ALA
_HU
MA
NP
1123
3R
ALA
Pla
sma
mem
bra
neR
as-r
elat
edp
rote
inR
ap-1
bR
AP
1B_H
UM
AN
P61
224
RA
P1B
Pla
sma
mem
bra
neG
TPas
eN
Ras
RA
SN
_HU
MA
NP
0111
1N
RA
SP
lasm
am
emb
rane
Ras
-rel
ated
pro
tein
Rab
-11A
RB
11A
_HU
MA
NP
6249
1R
AB
11A
Pla
sma
mem
bra
neR
ho-r
elat
edG
TP-b
ind
ing
pro
tein
Rho
CR
HO
C_H
UM
AN
P08
134
RH
OC
Pla
sma
mem
bra
neR
ho-a
ssoc
iate
dp
rote
inki
nase
2R
OC
K2_
HU
MA
NO
7511
6R
OC
K2
Pla
sma
mem
bra
neR
as-r
elat
edp
rote
inR
-Ras
2R
RA
S2_
HU
MA
NP
6207
0R
RA
S2
Pla
sma
mem
bra
neP
rote
inS
100-
A6a
S10
A6_
HU
MA
NP
0670
3S
100A
6P
lasm
am
emb
rane
Pro
tein
S10
0-A
10a
S10
AA
_HU
MA
NP
6090
3S
100A
10P
lasm
am
emb
rane
Sw
itch-
asso
ciat
edp
rote
in70
SW
P70
_HU
MA
NQ
9UH
65S
WA
P70
Pla
sma
mem
bra
neN
ED
D8-
activ
atin
gen
zym
eE
1re
gula
tory
sub
unit
ULA
1_H
UM
AN
Q13
564
NA
E1
Pla
sma
mem
bra
neTr
ansp
ort-
rela
ted
pro
tein
sA
P-2
com
ple
xsu
bun
ital
pha
-1A
P2A
1_H
UM
AN
O95
782
AP
2A1
Pla
sma
mem
bra
neA
P-2
com
ple
xsu
bun
ital
pha
-2A
P2A
2_H
UM
AN
O94
973
AP
2A2
Pla
sma
mem
bra
neA
DP
-rib
osyl
atio
nfa
ctor
1A
RF1
_HU
MA
NP
8407
7A
RF1
Pla
sma
mem
bra
neA
DP
-rib
osyl
atio
nfa
ctor
6A
RF6
_HU
MA
NP
6233
0A
RF6
Pla
sma
mem
bra
neA
DP
-rib
osyl
atio
nfa
ctor
-lik
ep
rote
in3
AR
L3_H
UM
AN
P36
405
AR
L3P
lasm
am
emb
rane
Bet
a-ar
rest
in-1
AR
RB
1_H
UM
AN
P49
407
AR
RB
1P
lasm
am
emb
rane
Chl
orid
ein
trac
ellu
lar
chan
nelp
rote
in1
CLI
C1_
HU
MA
NO
0029
9C
LIC
1P
lasm
am
emb
rane
Chl
orid
ein
trac
ellu
lar
chan
nelp
rote
in4
CLI
C4_
HU
MA
NQ
9Y69
6C
LIC
4P
lasm
am
emb
rane
Clu
ster
inC
LUS
_HU
MA
NP
1090
9C
LUE
xtra
cellu
lar
Gly
cop
rote
inC
oato
mer
sub
unit
bet
aC
OP
B_H
UM
AN
P53
618
CO
PB
1P
lasm
am
emb
rane
EH
dom
ain-
cont
aini
ngp
rote
in1
EH
D1_
HU
MA
NQ
9H4M
9E
HD
1P
lasm
am
emb
rane
EH
dom
ain-
cont
aini
ngp
rote
in2
EH
D2_
HU
MA
NQ
9NZ
N4
EH
D2
Pla
sma
mem
bra
neP
alm
itoyl
-pro
tein
thio
este
rase
1P
PT1
_HU
MA
NP
5089
7P
PT1
Ext
race
llula
rG
lyco
pro
tein
Pro
tein
S10
0-A
13S
10A
D_H
UM
AN
Q99
584
S10
0A13
Ext
race
llula
r
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 9
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

explanation for why previously unidentified proteins (8, 10)were found in the present analysis (Fig. 1C). An overlay be-tween secreted (Cy3 and Cy 5; green and red color) andcellular (Cy 2; blue color) proteins is shown in Fig. 1D. Com-mon spots were numbered (supplemental Fig. S2) and iden-tified via LC-MS/MS (supplemental Table S3). Certain pro-teins, such as von Willebrand antigen 2 (a propeptide of vWF,AA 23–763), were clearly more abundant in the secretome ofPMA-treated HUVECs.
The Endothelial Glycoproteome—Among the 1252 identi-fied proteins were 253 extracellular or plasma membraneproteins (approximately 20%) related to cell adhesion, bloodcoagulation, hemostasis, signaling transduction, and proteintransportation, of which 166 were known glycoproteins (TableI). To further characterize this subproteome, we employed aglycoproteomics approach. Secreted proteins were precipi-tated and digested with trypsin, and tryptic peptides werelabeled with TMT0 to increase their charge state prior toenrichment by means of zwitterionic hydrophilic interactionliquid chromatography purification (24). For glycosite identifi-cation, an indirect and a direct strategy were pursued (Fig.2A): (i) digestion with PNGase F in the presence of 18O waterto label the conversion of asparagine to aspartic acid upon theremoval of N-glycans, and (ii) alternating HCD and ETD (HCD-
TAB
LEI—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDU
niP
rot
acce
ssio
nnu
mb
erG
ene
nam
eC
ellu
lar
com
pon
ent
Gly
cop
rote
inE
Cm
arke
r
Sol
ute
carr
ier
fam
ily12
mem
ber
2S
12A
2_H
UM
AN
P55
011
SLC
12A
2P
lasm
am
emb
rane
Pro
activ
ator
pol
ypep
tide
SA
P_H
UM
AN
P07
602
PS
AP
Ext
race
llula
rG
lyco
pro
tein
Syn
taxi
n-b
ind
ing
pro
tein
1S
TXB
1_H
UM
AN
P61
764
STX
BP
1P
lasm
am
emb
rane
Syn
taxi
n-b
ind
ing
pro
tein
3S
TXB
3_H
UM
AN
O00
186
STX
BP
3P
lasm
am
emb
rane
Tran
smem
bra
neem
p24
dom
ain-
cont
aini
ngp
rote
in10
TME
DA
_HU
MA
NP
4975
5TM
ED
10P
lasm
am
emb
rane
Gly
cop
rote
inV
esic
le-a
ssoc
iate
dm
emb
rane
pro
tein
-ass
ocia
ted
pro
tein
AV
AP
A_H
UM
AN
Q9P
0L0
VA
PA
Pla
sma
mem
bra
ne
aTh
ese
pro
tein
sw
ere
also
iden
tifie
din
the
cellu
lar
pro
teom
eac
cord
ing
toth
ed
iffer
ence
gele
lect
rop
hore
sis
anal
ysis
pre
sent
edin
the
sup
ple
men
tald
ata.
FIG. 2. Glycoproteomics. A, glycopeptide identification workflow.Comparison of direct and indirect glycopeptide detection using HCD-ETD and 18O-deamidation after PNGase F � H2
18O treatment, re-spectively: identified unique glycopeptides (B), unique glycosylationsites (C), and unique glycoproteins (D).
Glycoproteomics of the Endothelial Secretome
10 Molecular & Cellular Proteomics 12.4
ZSI
ZSI
F2
COLOR
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

alt-ETD) or HCD-product-dependent ETD (HCD-pd-ETD)fragmentation on an Orbitrap Elite MS (24).
There was little overlap in the numbers of glycopeptides(Fig. 2B) and glycosylation sites (Fig. 2C) identified via thedirect (HCD-ETD) and the indirect (PNGase F � H2
18O) meth-ods. Better agreement was observed at the protein level (Fig.2D). With the indirect (PNGase F � H2
18O) method, 27 pep-tides were identified with N[�2.99] modification at non-con-sensus sequence, out of 1139 total identified peptides withN[�2.99]. This anomaly rate of 2.4% (27/1139) combines therate of false identifications and the rate of chance deamida-tions in 18O water that were not in the consensus sequence ofglycosylation (i.e. N-X(not P)-S/T). All glycopeptides identifiedare listed in Table II and supplemental Table S4. Three spectra(full MS, HCD, and ETD) from a neuronal cell adhesion mole-cule (UniProt accession number Q92823) (AA - 222FNHTQ-TIQQK231) are presented in Fig. 3.
For the same samples, HCD-pd-ETD revealed 28 known,25 potential, and 16 novel glycosylation sites based on 209identified spectra; HCD-alt-ETD revealed 20 known, 32 po-tential, and 14 novel glycosylation sites from 110 identifiedspectra. The HCD-alt-ETD method selected mostly precur-sors with higher intensities, higher charge, and smaller m/z(Fig. 4A). Several large glycopeptides were detected via onlyHCD-alt-ETD, and more low-abundant glycopeptides weredetected via HCD-pd-ETD. There was limited overlap in theidentified glycopeptides but better agreement in the proteinlevel (Fig. 4B). Among the 319 total glycopeptides identified inthe conditioned media, 31 were attached with a trimannosylcore (-HexNAc2Hex3) or truncated core (-HexNAc2Hex), 50with high mannose (-HexNAc2Hex4–9), and 238 with complex/hybrid glycans. Notably, HCD-pd-ETD detected almost twiceas many complex/hybrid glycoforms as HCD-alt-ETD(Fig.4C).
Validation of Glycoproteins—To validate the glycosylationstatus, we performed additional analysis before and afterglycoprotein enrichment with affinity resins of ConA lectin(n � 4) using a Q Exactive MS (Thermo Scientific). We thencompared the number of identified spectra in the glycopro-tein-enriched fraction, the flow-through, and the input (sup-plemental Table S5). For most glycoproteins, a higher spectralcount was observed in the glycoprotein-enriched fractionthan in the original input and/or the flow-through. Represent-ative examples (fibronectin, neuronal cell adhesion molecule,tyrosine-protein-kinase-like 7, and vWF) are shown in Fig. 5A.Non-glycosylated proteins, such as annexin A2 and alpha-enolase, were more abundant in the flow-through. Glycopro-teins identified in all three methods are highlighted in Fig. 5B.
Confirmation of Predicted Glycosylation Sites—The hemo-static protein vWF is the main protein stored within Weibel-Palade bodies (27). After secretagogue stimulation, Weibel-Palade bodies undergo exocytosis, releasing vWF filaments.vWF is one of the few known proteins containing the ABOblood group signature, which is formed by different glycans.
Although the released glycan composition of this protein hasbeen investigated extensively (28, 29), experimental evidencefor many putative glycosylation sites is still missing. The cov-erage obtained for vWF in our proteomics analysis is shown inFig. 6A. The precursor protein consists of homologous unitssuch as the VWF type A, C, and D domains and a C-terminalcystine know (CTCK). The vWF propeptide (D1-D2, AA 23–763) is separated from the remaining domains of mature vWF(AA 764–2813) via furin-mediated proteolytic cleavage. Weconfirmed 6 N-glycosylation sites. Notably, three N-glycosy-lation sites were located within the propeptide (AA 23–763).Examples of ETD spectra are shown in Fig. 6B.
DISCUSSION
This study represents a significant advance over the exist-ing proteomics literature on ECs. Unlike other cell types, ECsdo not tolerate prolonged serum starvation, and their suscep-tibility to cell death upon serum withdrawal poses a majorchallenge for proteomic workflows targeting their secretome.We performed secretome analysis after 45 min of PMA stim-ulation combined with enrichment strategies for glycopro-teins and glycopeptides. Glycopeptides were analyzed viathree complementary MS techniques: the detection of 18Oasparagine deamidation after digestion with PNGase F inH2
18O, HCD-alt-ETD, and HCD-pd-ETD using an OrbitrapElite MS.
The Endothelial Secretome—The secretagogue PMA mini-mized EC death by allowing a shorter incubation period underserum-free conditions while increasing coverage in the pro-teomic analysis by inducing the exocytosis of intracellularstorage vesicles (14) such as Weibel-Palade bodies. Theseunique storage vesicles in ECs play a major role in hemostasisand cell-to-cell communication. Using this approach, manymore proteins were identified than in any previous proteomicsstudy on ECs, including known endothelial surface markerssuch as endoglin (CD105), integrin beta-1 (CD29), tyrosine-protein kinase receptor Tie-1, and junctional adhesion mole-cule A; secreted growth factors (i.e. C-type lectin domainfamily 11 member A); co-receptors (i.e. neuropilin-1 (co-re-ceptor for VEGF-A)); proteases(i.e. furin); and inflammatorymediators (i.e. macrophage migration inhibitory factor), toname just a few. Short-term PMA treatment does not releasemicroparticles (30), as shedding events make it difficult todiscern intracellular from secreted/membrane proteins. In adirect comparison of the cellular proteome and the secretomeutilizing difference gel electrophoresis, 70 out of 96 proteinsanalyzed were present in both samples, representing �10%of the visible protein spots in the secretome.
Biological Importance of Glycosylation—Glycosylation iskey for the stability and solubility of secreted and membraneproteins. It is the most complex post-translational modifica-tion (31) and mediates extracellular matrix network assembly,cell–cell interactions, and cell–matrix interactions. Unlikepolynucleotides and polypeptides, which have a linear struc-
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 11
T2,ZSI
F4
ZSI
F5
F6
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEII
Gly
cop
eptid
esid
entif
ied
via
the
HC
D-E
TDm
etho
d
Pro
tein
nam
eU
niP
rot
IDP
eptid
eG
lyco
site
Typ
eG
lyca
nsO
bse
rved
m/z
Z�
Mas
s(p
pm
)
Afa
min
AFA
M_H
UM
AN
$DIE
NFN
(�17
02.5
82)ˆS
TQk
N33
Kno
wn
Hex
8Hex
NA
c284
3.35
94
�5.
8A
min
opep
tidas
eN
AM
PN
_HU
MA
N$k
LN(�
892.
318)
YTL
SQ
GH
RV
VLR
N12
8K
now
nH
ex3H
exN
Ac2
781.
916
4�
2.8
$NA
N(�
2042
.720
)ˆSS
PV
AsT
TPS
AS
ATT
NP
AS
ATT
LDQ
Sk
N42
Nov
elH
ex4H
exN
Ac4
Neu
Ac2
1095
.276
5�
0.3
Alp
ha-N
-ace
tylg
luco
sam
inid
ase
AN
AG
_HU
MA
N$S
VY
N(�
1257
.450
)cS
GE
AcR
GhN
RS
PLV
RN
503
Pot
entia
lH
ex4H
exN
Ac3
954.
927
41.
1A
ngio
poi
etin
-2A
NG
P2_
HU
MA
N$k
IVTA
TVN
(�56
8.21
2)N
SV
LQk
N24
0P
oten
tial
Hex
1Hex
NA
c268
9.64
54
�0.
3$S
GhT
TNG
IYTL
TFP
N(�
1038
.375
)STE
EIk
N30
4P
oten
tial
Hex
3Hex
NA
c2d
Hex
176
3.56
55
�1.
4A
ttra
ctin
ATR
N_H
UM
AN
$DLD
MFI
N(�
1241
.455
)AS
kN
1198
Kno
wn
Hex
3Hex
NA
c3d
Hex
194
8.45
03
6.7
$GcS
cFS
DW
QG
PG
cSV
PV
PA
N(�
1095
.397
)QS
FWTR
N32
5P
oten
tial
Hex
3Hex
NA
c310
77.7
084
�2.
1$G
cScF
SD
WQ
GP
GcS
VP
VP
AN
(�89
2.31
7)Q
SFW
TRE
EY
SnL
kN
325
Pot
entia
lH
ex3H
exN
Ac2
1040
.061
5�
2.4
Cad
herin
-2C
AD
H2_
HU
MA
N$E
QIA
RFH
LRA
HA
VD
InG
NQ
VE
NP
IDIV
INV
IDM
ND
NR
PE
FLH
QV
WN
(�17
51.6
24)G
TVP
EG
Sk
N27
3K
now
nH
ex4H
exN
Ac4
Neu
Ac1
930.
011
92.
8
Cad
herin
-5C
AD
H5_
HU
MA
N$E
N(�
1054
.370
)ˆIS
EY
HLT
AV
IVD
kN
112
Kno
wn
Hex
4Hex
NA
c281
5.14
64
0.7
$ELD
RE
VY
PW
YN
(�12
41.4
55)L
TVE
Ak
N44
2K
now
nH
ex3H
exN
Ac3
dH
ex1
954.
972
410
.0C
D10
9an
tigen
CD
109_
HU
MA
N$L
NLY
LDS
VN
(�10
38.3
75)ˆE
TQFc
VN
IPA
VR
N13
55N
ovel
Hex
3Hex
NA
c2d
Hex
193
8.44
74
1.5
$kkN
(�15
40.5
29)IT
kN
279
Pot
entia
lH
ex7H
exN
Ac2
793.
410
4�
0.9
$QN
(�18
48.6
40)ˆS
TMFS
LTP
EN
SW
TPk
N51
3N
ovel
Hex
8Hex
NA
c2d
Hex
110
72.7
194
1.0
CD
59gl
ycop
rote
inC
D59
_HU
MA
N$T
AV
N(�
1954
.704
)ˆcsS
DFD
AcL
ITk
N43
Kno
wn
Hex
4Hex
NA
c5N
euA
c110
77.7
084
9.1
Com
ple
men
tfa
ctor
IC
FAI_
HU
MA
N$F
LNN
(�10
54.3
70)G
TcTA
EG
kN
103
Kno
wn
Hex
4Hex
NA
c293
9.09
43
�4.
0$L
ISN
(�35
34.2
44)ˆc
Sk
N49
4P
oten
tial
Hex
8Hex
NA
c6d
Hex
1Neu
Ac3
1207
.501
45.
7C
AP
-Gly
dom
ain-
cont
aini
nglin
ker
pro
tein
1C
LIP
1_H
UM
AN
$GE
N(�
1257
.450
)ˆAS
Ak
N12
63N
ovel
Hex
4Hex
NA
c380
2.35
93
�1.
4
$GE
N(�
1784
.635
)AS
Ak
N12
63N
ovel
Hex
6Hex
NA
c497
0.43
73
10.7
$EP
SA
TPP
ISN
(�21
88.7
41)L
TkN
187
Nov
elH
ex11
Hex
NA
c299
9.19
24
�6.
9$A
NE
N(�
1200
.428
)ˆAS
FLQ
kSIE
DM
TVk
N97
1N
ovel
Hex
4Hex
NA
c2d
Hex
198
0.72
44
2.6
$AN
EN
(�12
16.4
23)ˆA
SFL
QkS
IED
MTV
kN
971
Nov
elH
ex5H
exN
Ac2
985.
232
411
.0$A
NE
N(�
1257
.449
)AS
FLQ
kSIE
DM
TVk
N97
1N
ovel
Hex
4Hex
NA
c398
9.48
94
8.2
$AN
EN
(�14
44.5
34)A
SFL
QkS
IED
MTV
kN
971
Nov
elH
ex3H
exN
Ac4
dH
ex1
1036
.764
49.
3E
phr
inty
pe-
Are
cep
tor
2E
PH
A2_
HU
MA
N$T
AS
VS
IN(�
892.
317)
QTE
PP
kVR
LEG
RN
435
Kno
wn
Hex
3Hex
NA
c285
6.69
44
�0.
4Fi
bro
ussh
eath
-int
erac
ting
pro
tein
2FS
IP2_
HU
MA
N$I
GW
EY
ES
TN(�
1751
.624
)ISR
N14
23N
ovel
Hex
4Hex
NA
c4N
euA
c185
8.37
34
0.8
$TIT
FSA
N(�
1362
.481
)VS
SH
EhT
Yk
N16
75N
ovel
Hex
5Hex
NA
c2d
Hex
191
2.92
34
3.0
$GG
IN(�
892.
318)
ISG
QG
SIIS
AQ
VS
PTR
N21
5N
ovel
Hex
3Hex
NA
c210
20.5
093
1.5
$EN
SN
(�12
00.4
28)F
SQ
LALS
NE
ILLG
HkE
kN
2216
Nov
elH
ex4H
exN
Ac2
dH
ex1
708.
363
66.
3$m
PIE
N(�
1444
.534
)LS
SIQ
Qk
N28
24N
ovel
Hex
3Hex
NA
c4d
Hex
182
5.39
84
0.8
$YN
(�22
04.7
72)k
N42
7N
ovel
Hex
5Hex
NA
c4N
euA
c210
26.7
793
7.7
N-a
cety
lglu
cosa
min
e-6-
sulfa
tase
GN
S_H
UM
AN
$LV
kRLE
FTG
ELN
(�20
18.7
08)ˆN
TYIF
YTS
DnG
YH
TGQ
FSLP
IDkR
N31
7K
now
nH
ex6H
exN
Ac3
dH
ex1N
euA
c169
6.02
710
�9.
6$G
PG
IkP
N(�
1540
.529
)QTS
kN
362
Pot
entia
lH
ex7H
exN
Ac2
835.
657
4�
0.8
Gol
gin
sub
fam
ilyA
mem
ber
4G
OG
A4_
HU
MA
N$E
LEhV
N(�
1735
.630
)ˆLS
Vk
N16
12N
ovel
Hex
3Hex
NA
c4d
Hex
1Neu
Ac1
848.
139
4�
3.9
$kE
LEH
VN
(�10
38.3
75)L
SV
kN
1612
Nov
elH
ex3H
exN
Ac2
dH
ex1
752.
648
40.
8$k
ELE
HV
N(�
1524
.534
)LS
Vk
N16
12N
ovel
Hex
6Hex
NA
c2d
Hex
187
4.18
64
�1.
6$S
LQE
NkN
(�12
57.4
50)Q
Sk
N58
5N
ovel
Hex
4Hex
NA
c377
7.39
14
10.2
$TR
ILE
LES
SLE
kSLQ
EN
kN(�
1216
.423
)QS
kN
585
Nov
elH
ex5H
exN
Ac2
938.
686
5�
2.3
Inte
rcel
lula
rad
hesi
onm
olec
ule
2IC
AM
2_H
UM
AN
$GS
LEV
N(�
2028
.741
)ˆcS
TTcN
QP
EV
GG
LETS
LDk
N47
Kno
wn
Hex
5Hex
NA
c610
39.8
615
5.1
$HY
LVS
N(�
568.
212)
ISH
DTV
LQcH
FTcS
Gk
N82
Kno
wn
Hex
1Hex
NA
c290
5.68
74
�0.
6IC
OS
ligan
dIC
OS
L_H
UM
AN
$IA
RTP
SV
NIG
ccIE
NV
LLQ
QN
(�24
57.8
77)L
TVG
SQ
TGN
DIG
ER
N22
5P
oten
tial
Hex
8Hex
NA
c5d
Hex
172
4.21
99
�5.
4$I
AR
TPS
VN
IGcc
IEN
VLL
N(�
2594
.937
)ˆLTV
GS
QTG
ND
IGE
RN
225
Pot
entia
lH
ex4H
exN
Ac6
dH
ex1N
euA
c266
7.90
610
0.1
Inte
rleuk
in-6
rece
pto
rsu
bun
itb
eta
IL6R
B_H
UM
AN
$SH
LQN
(�56
8.21
2)Y
TVN
ATk
LTV
NLT
ND
RY
LATL
TVR
NLV
Gk
N37
9K
now
nH
ex1H
exN
Ac2
725.
691
76.
5
Glycoproteomics of the Endothelial Secretome
12 Molecular & Cellular Proteomics 12.4
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEII—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDP
eptid
eG
lyco
site
Typ
eG
lyca
nsO
bse
rved
m/z
Z�
Mas
s(p
pm
)
$SH
LQN
(�56
8.21
2)Y
TVN
ATk
LTV
NLT
ND
RY
LATL
TVR
NLV
Gk
N37
9K
now
nH
ex1H
exN
Ac2
1015
.568
59.
4$L
TVN
(�14
60.5
29)L
TND
RY
LATL
TVR
NLV
Gk
N39
0K
now
nH
ex4H
exN
Ac4
877.
848
5�
8.4
$QN
(�56
8.21
2)cS
QH
ES
SP
DIs
HFE
RN
818
Nov
elH
ex1H
exN
Ac2
733.
814
48.
8In
tegr
inal
pha
-3IT
A3_
HU
MA
N$D
VR
kLLL
SIN
(�89
2.31
7)V
TNTR
N65
6P
oten
tial
Hex
3Hex
NA
c210
28.5
543
�2.
2$A
HcV
WLE
cPIP
DA
PV
VTN
(�13
62.4
81)V
TVk
N92
6P
oten
tial
Hex
5Hex
NA
c2d
Hex
186
4.22
35
8.3
Inte
grin
bet
a-1
ITB
1_H
UM
AN
$kN
kNV
TN(�
1444
.534
)ˆRS
kN
97P
oten
tial
Hex
3Hex
NA
c4d
Hex
188
8.71
04
2.8
Junc
tiona
lad
hesi
onm
olec
ule
CJA
M3_
HU
MA
N$I
WN
(�15
81.5
55)V
TRR
DS
ALY
RcE
VV
AR
ND
RN
104
Pot
entia
lH
ex6H
exN
Ac3
1139
.528
4�
4.8
$IW
N(�
892.
318)
ˆVTR
N10
4P
oten
tial
Hex
3Hex
NA
c264
3.30
33
�0.
7Ly
soso
me-
asso
ciat
edm
emb
rane
glyc
opro
tein
1LA
MP
1_H
UM
AN
$GH
TLTL
N(�
1378
.476
)FTR
N10
3K
now
nH
ex6H
exN
Ac2
921.
421
3�
0.4
$GH
TLTL
N(�
1540
.529
)FTR
N10
3K
now
nH
ex7H
exN
Ac2
975.
439
3�
0.4
$YN
VS
GTN
GTc
LLA
SM
GLQ
LN(�
2042
.720
)ˆLTY
ER
kDN
TTV
TRLL
nIN
PN
kN
241
Kno
wn
Hex
4Hex
NA
c4N
euA
c273
4.66
110
6.4
$kD
N(�
2059
.735
)TTV
TRN
249
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
861.
641
41.
0$k
DN
(�22
04.7
72)T
TVTR
N24
9K
now
nH
ex5H
exN
Ac4
Neu
Ac2
898.
156
45.
8$k
DN
(�23
50.8
30)T
TVTR
N24
9K
now
nH
ex5H
exN
Ac4
dH
ex1N
euA
c293
4.41
44
�0.
2$k
DN
(�25
69.9
05)T
TVTR
N24
9K
now
nH
ex6H
exN
Ac5
Neu
Ac2
989.
440
46.
2$k
DN
(�25
70.9
25)T
TVTR
N24
9K
now
nH
ex6H
exN
Ac5
dH
ex2N
euA
c198
9.69
04
1.0
$kD
N(�
2571
.946
)TTV
TRN
249
Kno
wn
Hex
6Hex
NA
c5d
Hex
498
9.69
04
�2.
4$k
DN
(�27
15.9
63)ˆT
TVTR
N24
9K
now
nH
ex6H
exN
Ac5
dH
ex1N
euA
c210
31.1
904
�2.
0$k
DN
(�27
15.9
63)T
TVTR
N24
9K
now
nH
ex6H
exN
Ac5
dH
ex1N
euA
c210
25.6
964
�0.
9$k
DN
(�27
17.9
78)T
TVTR
N24
9K
now
nH
ex8H
exN
Ac7
1025
.949
4�
1.2
$kD
N(�
2861
.000
)TTV
TRN
249
Kno
wn
Hex
6Hex
NA
c5N
euA
c310
62.2
114
3.1
$kD
N(�
2864
.036
)TTV
TRN
249
Kno
wn
Hex
8Hex
NA
c7d
Hex
110
62.4
664
1.2
$kD
N(�
3007
.058
)TTV
TRN
249
Kno
wn
Hex
6Hex
NA
c5d
Hex
1Neu
Ac3
1098
.476
44.
4$A
AN
(�17
02.5
81)G
SLR
N32
2K
now
nH
ex8H
exN
Ac2
872.
711
32.
6$A
AN
(�18
64.6
34)G
SLR
N32
2K
now
nH
ex9H
exN
Ac2
926.
395
33.
5Ly
soso
me-
asso
ciat
edm
emb
rane
glyc
opro
tein
2LA
MP
2_H
UM
AN
$EkP
EA
GTY
SV
NN
GN
(�10
54.3
70)D
TcLL
ATm
GLQ
LNIT
QD
kN
229
Kno
wn
Hex
4Hex
NA
c210
48.5
065
0.8
Lyso
som
alal
pha
-glu
cosi
das
eLY
AG
_HU
MA
N$Q
VV
EN
(�10
95.3
97)ˆM
TRA
HFP
LDV
QW
ND
LDY
MD
SR
N39
0K
now
nH
ex3H
exN
Ac3
905.
408
50.
9$G
AY
TQV
IFLA
RN
(�10
54.3
70)N
TIV
NE
LVR
N88
2K
now
nH
ex4H
exN
Ac2
918.
718
41.
4P
rote
in-l
ysin
e6-
oxid
ase
LYO
X_H
UM
AN
$AE
N(�
1054
.370
)QTA
PG
EV
PA
LSN
LRN
144
Pot
entia
lH
ex4H
exN
Ac2
762.
369
48.
1$D
PG
AA
VP
GA
AN
AS
AQ
QP
RTP
ILLI
RD
N(�
2432
.884
)RN
97P
oten
tial
Hex
3Hex
NA
c6d
Hex
1Neu
Ac2
1106
.316
5�
5.0
Lym
pha
ticve
ssel
end
othe
lial
hyal
uron
icac
idre
cep
tor
1LY
VE
1_H
UM
AN
$AN
(�13
62.4
81)D
SN
PN
EE
Skk
TDk
N28
9N
ovel
Hex
5Hex
NA
c2d
Hex
198
4.72
74
5.8
Lyso
som
alal
pha
-man
nosi
das
eM
A2B
1_H
UM
AN
$GFk
DH
FTFc
LN(�
892.
317)
ISIc
PLS
QTA
AR
FQV
IVY
nPLG
Rk
N49
7P
oten
tial
Hex
3Hex
NA
c298
7.50
46
�1.
2H
epat
ocyt
egr
owth
fact
orre
cep
tor
ME
T_H
UM
AN
$TLL
RN
(�17
51.6
24)ˆS
sGcE
AR
RD
EY
RN
405
Pot
entia
lH
ex4H
exN
Ac4
Neu
Ac1
1040
.946
46.
4$V
IVQ
PD
QN
(�20
42.7
20)ˆˆ
FTG
LIA
GV
VS
ISTA
LLLL
LGFF
LWLk
kRk
N93
0P
oten
tial
Hex
4Hex
NA
c4N
euA
c288
6.22
58
3.5
$VIV
QP
DQ
N(�
2174
.799
)ˆFTG
LIA
GV
VS
ISTA
LLLL
LGFF
LWLk
kRN
930
Pot
entia
lH
ex5H
exN
Ac6
dH
ex1
1140
.772
64.
0In
ters
titia
lcol
lage
nase
MM
P1_
HU
MA
N$A
FQLW
SN
(�20
34.7
03)ˆV
TPLT
FTk
N14
3N
ovel
Hex
7Hex
NA
c3N
euA
c110
65.9
924
3.1
$AFQ
LWS
N(�
2059
.735
)VTP
LTFT
kN
143
Nov
elH
ex5H
exN
Ac4
dH
ex1N
euA
c110
65.9
994
0.3
$AFQ
LWS
N(�
2075
.730
)ˆVTP
LTFT
kN
143
Nov
elH
ex6H
exN
Ac4
Neu
Ac1
1076
.245
40.
2$A
FQLW
SN
(�20
75.7
30)V
TPLT
FTk
N14
3N
ovel
Hex
5Hex
NA
c4d
Hex
2Neu
Gc1
1070
.250
41.
4$A
FQLW
SN
(�20
76.7
50)V
TPLT
FTk
N14
3N
ovel
Hex
6Hex
NA
c4d
Hex
210
70.2
514
�1.
0$A
FQLW
SN
(�20
92.7
45)V
TPLT
FTk
N14
3N
ovel
Hex
7Hex
NA
c4d
Hex
210
74.2
514
0.2
$AFQ
LWS
N(�
2350
.830
)ˆVTP
LTFT
kN
143
Nov
elH
ex5H
exN
Ac4
dH
ex1N
euA
c211
44.7
694
�0.
3$A
FQLW
SN
(�23
50.8
30)V
TPLT
FTk
N14
3N
ovel
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
1139
.021
4�
1.9
$AFQ
LWS
N(�
2351
.851
)VTP
LTFT
kN
143
Nov
elH
ex5H
exN
Ac4
dH
ex3N
euA
c111
39.2
774
�0.
9
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 13
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEII—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDP
eptid
eG
lyco
site
Typ
eG
lyca
nsO
bse
rved
m/z
Z�
Mas
s(p
pm
)
Mul
timer
in-1
MM
RN
1_H
UM
AN
$IN
ALk
kPTV
N(�
1200
.428
)LTT
VLI
GR
N10
20K
now
nH
ex4H
exN
Ac2
dH
ex1
957.
031
41.
5$G
AR
LFV
LLS
SLW
SG
GIG
LN(�
1241
.455
)ˆNS
kN
21P
oten
tial
Hex
3Hex
NA
c3d
Hex
110
01.0
064
�3.
0$L
FVLL
SS
LWS
GG
IGLN
(�56
8.21
2)N
Sk
N21
Pot
entia
lH
ex1H
exN
Ac2
756.
162
4�
0.8
$ID
N(�
1095
.397
)ˆIS
LTV
ND
VR
NTY
SS
LEG
kN
344
Kno
wn
Hex
3Hex
NA
c397
6.97
34
�1.
1$I
nN(�
568.
212)
LTV
SLE
ME
kN
576
Pot
entia
lH
ex1H
exN
Ac2
803.
746
3�
1.8
$kIE
N(�
892.
317)
LTS
AV
NS
LNFI
IkN
680
Pot
entia
lH
ex3H
exN
Ac2
868.
469
4�
3.4
$LN
QS
NFQ
kmY
QM
FN(�
2262
.815
)ˆETT
SQ
VR
N82
8P
oten
tial
Hex
5Hex
NA
c5d
Hex
1Neu
Ac1
1069
.470
52.
0$A
LEA
kSIH
LSIn
FFS
LN(�
568.
212)
kN
921
Pot
entia
lH
ex1H
exN
Ac2
819.
195
4�
5.2
$SIH
LSIN
FFS
LN(�
892.
318)
ˆkN
921
Pot
entia
lH
ex3H
exN
Ac2
721.
609
4�
6.7
C-t
ype
man
nose
rece
pto
r2
MR
C2_
HU
MA
N$V
TPA
cN(�
2059
.735
)TS
LPA
QR
N69
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
925.
654
4�
2.1
$SN
(�20
59.7
35)V
TkN
954
Pot
entia
lH
ex5H
exN
Ac4
dH
ex1N
euA
c110
20.1
223
0.2
Nck
-ass
ocia
ted
pro
tein
5N
CK
P5_
HU
MA
N$E
RG
PQ
GQ
GH
GR
MA
LNLQ
LSD
TDD
N(�
2018
.708
)ˆE
TFD
ELH
IES
SD
Ek
N58
5N
ovel
Hex
6Hex
NA
c3d
Hex
1Neu
Ac1
1127
.492
6�
3.7
Lyso
som
alp
rote
inN
CU
-G1
NC
UG
1_H
UM
AN
$LLH
TAD
TcQ
LEV
ALI
GA
sPR
GN
(�12
41.4
54)R
N23
0P
oten
tial
Hex
3Hex
NA
c3d
Hex
110
10.2
234
�1.
4N
atur
alcy
toto
xici
tytr
igge
ring
rece
pto
r3
ligan
d1
NR
3L1_
HU
MA
N$L
N(�
2172
.745
)SS
QE
DP
GTV
YQ
cVV
RH
AS
LHTP
LRN
216
Pot
entia
lH
ex10
Hex
NA
c2d
Hex
110
73.4
775
�4.
3
Neu
rona
lcel
lad
hesi
onm
olec
ule
NR
CA
M_H
UM
AN
$IP
AN
(�27
15.9
63)k
N10
09P
oten
tial
Hex
6Hex
NA
c5d
Hex
1Neu
Ac2
927.
404
4�
1.4
$IP
AN
(�29
60.0
68)ˆˆ
kN
1009
Pot
entia
lH
ex5H
exN
Ac7
dH
ex1N
euA
c210
00.1
814
6.5
$FN
(�13
78.4
76)H
TQTI
kN
223
Pot
entia
lH
ex6H
exN
Ac2
768.
611
40.
9$F
N(�
1378
.476
)HTQ
TIQ
Qk
N22
3P
oten
tial
Hex
6Hex
NA
c210
24.4
803
2.2
$SS
RE
RP
PTF
LTP
EG
N(�
1710
.598
)AS
Nk
N27
6K
now
nH
ex5H
exN
Ac3
Neu
Ac1
1062
.994
4�
0.4
Nuc
lear
rece
pto
r-in
tera
ctin
gp
rote
in3
NR
IP3_
HU
MA
N$L
ME
TN(�
568.
212)
LSk
N72
Nov
elH
ex1H
exN
Ac2
651.
345
38.
7
Pla
smin
ogen
activ
ator
inhi
bito
r1
PA
I1_H
UM
AN
$GN
(�16
94.6
03)ˆM
TRLP
RLL
VLP
kFS
LETE
VD
LRk
N28
8K
now
nH
ex4H
exN
Ac3
dH
ex1N
euA
c110
63.9
535
3.4
$GN
(�20
59.7
35)M
TRN
288
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
955.
394
30.
5$G
N(�
2350
.830
)ˆmTR
N28
8K
now
nH
ex5H
exN
Ac4
dH
ex1N
euA
c279
9.31
34
�3.
7$G
N(�
2351
.851
)ˆmTR
N28
8K
now
nH
ex5H
exN
Ac4
dH
ex3N
euA
c179
8.81
44
�5.
6P
late
let-
der
ived
grow
thfa
ctor
sub
unit
BP
DG
FB_H
UM
AN
$LLH
GD
PG
EE
DG
AE
LDLN
(�18
64.6
34)m
TRS
HS
GG
ELE
SLA
RG
RR
N63
Nov
elH
ex9H
exN
Ac2
1176
.921
5�
5.5
Sec
reto
ryp
hosp
holip
ase
A2
rece
pto
rP
LA2R
_HU
MA
N$M
QD
TSG
HG
VN
TsD
MY
PM
PN
TLE
YG
N(�
2204
.772
)ˆˆR
TYk
N11
23N
ovel
Hex
5Hex
NA
c4N
euA
c210
14.9
136
�1.
9
Pro
colla
gen-
lysi
ne,2
-ox
oglu
tara
te5-
dio
xyge
nase
2
PLO
D2_
HU
MA
N$Y
FN(�
1856
.656
)YTV
kVLG
QG
EE
WR
N63
Pot
entia
lH
ex5H
exN
Ac3
dH
ex1N
euA
c110
74.7
444
0.2
Pho
spho
lipid
tran
sfer
pro
tein
PLT
P_H
UM
AN
$qLL
YW
FFY
DG
GY
IN(�
2157
.783
)ˆAS
AE
GV
SIR
TGLE
LSR
N11
7P
oten
tial
Hex
4Hex
NA
c6N
euA
c111
18.8
985
0.2
$IY
SN
(�14
03.5
07)H
SA
LES
LALI
PLQ
AP
LkN
398
Kno
wn
Hex
4Hex
NA
c3d
Hex
110
33.7
824
1.5
Ple
xin-
C1
PLX
C1_
HU
MA
N$I
AN
(�15
24.5
34)ˆF
TSD
VE
YS
DD
Hch
LILP
DS
EA
FQD
VQ
GkR
HR
N13
08N
ovel
Hex
6Hex
NA
c2d
Hex
112
03.3
425
�1.
8$I
AN
(�22
04.7
73)F
TSD
VE
YS
DD
hcH
LILP
DS
EA
FQD
VQ
GkR
N13
08N
ovel
Hex
5Hex
NA
c4N
euA
c263
8.68
910
5.9
$VIL
GE
N(�
1298
.476
)LTS
NcP
EV
IYE
IkN
407
Kno
wn
Hex
3Hex
NA
c498
5.48
34
�1.
8$E
LcQ
N(�
1378
.476
)kN
548
Pot
entia
lH
ex6H
exN
Ac2
873.
730
38.
0$D
VcI
QFD
GG
NcS
SV
GS
LSY
IALP
HcS
LIFP
ATT
WIS
GG
QN
(�14
03.5
08)ˆI
TMM
GR
N77
1P
oten
tial
Hex
4Hex
NA
c3d
Hex
160
7.27
911
0.5
$EN
DN
FN(�
1054
.370
)ISk
N87
1N
ovel
Hex
4Hex
NA
c286
1.72
53
�4.
1$E
ND
NFN
(�25
22.9
16)ˆI
Sk
N87
1N
ovel
Hex
5Hex
NA
c7N
euA
c110
19.4
384
7.5
Tyro
sine
-pro
tein
kina
se-l
ike
7P
TK7_
HU
MA
N$S
AN
(�30
07.0
58)A
SFN
IkN
116
Kno
wn
Hex
6Hex
NA
c5d
Hex
1Neu
Ac3
1102
.722
42.
1$S
AN
(�30
26.0
90)A
SFN
IkN
116
Kno
wn
Hex
9Hex
NA
c7d
Hex
111
07.4
794
1.3
$ATV
FAN
(�17
10.5
98)G
SLL
LTQ
VR
PR
N28
3K
now
nH
ex5H
exN
Ac3
Neu
Ac1
945.
458
40.
0$R
QD
VN
(�16
86.5
86)IT
VA
TVP
SW
LkN
405
Pot
entia
lH
ex7H
exN
Ac2
dH
ex1
991.
485
43.
2P
entr
axin
-rel
ated
pro
tein
PTX
3P
TX3_
HU
MA
N$A
TDV
LN(�
1419
.502
)kTI
LFsY
GTk
N22
0P
oten
tial
Hex
5Hex
NA
c398
6.47
44
�4.
0
Glycoproteomics of the Endothelial Secretome
14 Molecular & Cellular Proteomics 12.4
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018
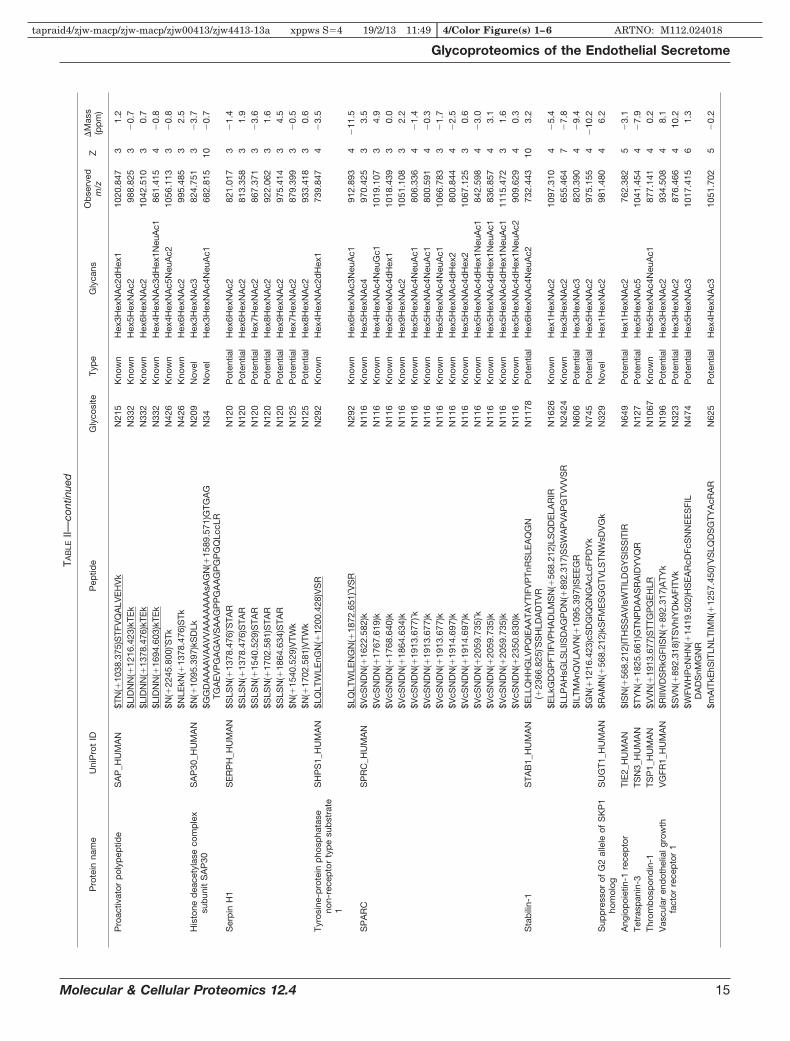
TAB
LEII—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDP
eptid
eG
lyco
site
Typ
eG
lyca
nsO
bse
rved
m/z
Z�
Mas
s(p
pm
)
Pro
activ
ator
pol
ypep
tide
SA
P_H
UM
AN
$TN
(�10
38.3
75)S
TFV
QA
LVE
HV
kN
215
Kno
wn
Hex
3Hex
NA
c2d
Hex
110
20.8
473
1.2
$LID
NN
(�12
16.4
23)k
TEk
N33
2K
now
nH
ex5H
exN
Ac2
988.
825
3�
0.7
$LID
NN
(�13
78.4
76)k
TEk
N33
2K
now
nH
ex6H
exN
Ac2
1042
.510
30.
7$L
IDN
N(�
1694
.603
)kTE
kN
332
Kno
wn
Hex
4Hex
NA
c3d
Hex
1Neu
Ac1
861.
415
4�
0.8
$N(�
2245
.800
)ˆSTk
N42
6K
now
nH
ex4H
exN
Ac5
Neu
Ac2
1056
.113
3�
0.8
$NLE
kN(�
1378
.476
)STk
N42
6K
now
nH
ex6H
exN
Ac2
995.
485
32.
5H
isto
ned
eace
tyla
seco
mp
lex
sub
unit
SA
P30
SA
P30
_HU
MA
N$N
(�10
95.3
97)k
SD
LkN
209
Nov
elH
ex3H
exN
Ac3
824.
751
3�
3.7
$GG
DA
AA
AV
AA
VV
AA
AA
AA
AsA
GN
(�15
89.5
71)G
TGA
GTG
AE
VP
GA
GA
VS
AA
GP
PG
AA
GP
GP
GQ
LccL
RN
34N
ovel
Hex
3Hex
NA
c4N
euA
c168
2.81
510
�0.
7
Ser
pin
H1
SE
RP
H_H
UM
AN
$SLS
N(�
1378
.476
)ˆSTA
RN
120
Pot
entia
lH
ex6H
exN
Ac2
821.
017
3�
1.4
$SLS
N(�
1378
.476
)STA
RN
120
Pot
entia
lH
ex6H
exN
Ac2
813.
358
31.
9$S
LSN
(�15
40.5
29)S
TAR
N12
0P
oten
tial
Hex
7Hex
NA
c286
7.37
13
�3.
6$S
LSN
(�17
02.5
81)S
TAR
N12
0P
oten
tial
Hex
8Hex
NA
c292
2.06
23
1.6
$SLS
N(�
1864
.634
)STA
RN
120
Pot
entia
lH
ex9H
exN
Ac2
975.
414
34.
5$N
(�15
40.5
29)V
TWk
N12
5P
oten
tial
Hex
7Hex
NA
c287
9.39
93
�0.
5$N
(�17
02.5
81)V
TWk
N12
5P
oten
tial
Hex
8Hex
NA
c293
3.41
83
0.6
Tyro
sine
-pro
tein
pho
spha
tase
non-
rece
pto
rty
pe
sub
stra
te1
SH
PS
1_H
UM
AN
$LQ
LTW
LEnG
N(�
1200
.428
)VS
RN
292
Kno
wn
Hex
4Hex
NA
c2d
Hex
173
9.84
74
�3.
5
$LQ
LTW
LEN
GN
(�18
72.6
51)ˆV
SR
N29
2K
now
nH
ex6H
exN
Ac3
Neu
Ac1
912.
893
4�
11.5
SP
AR
CS
PR
C_H
UM
AN
$VcS
ND
N(�
1622
.582
)kN
116
Kno
wn
Hex
5Hex
NA
c497
0.42
53
3.5
$VcS
ND
N(�
1767
.619
)kN
116
Kno
wn
Hex
4Hex
NA
c4N
euG
c110
19.1
073
4.9
$VcS
ND
N(�
1768
.640
)kN
116
Kno
wn
Hex
5Hex
NA
c4d
Hex
110
18.4
393
0.0
$VcS
ND
N(�
1864
.634
)kN
116
Kno
wn
Hex
9Hex
NA
c210
51.1
083
2.2
$VcS
ND
N(�
1913
.677
)ˆkN
116
Kno
wn
Hex
5Hex
NA
c4N
euA
c180
6.33
64
�1.
4$V
cSN
DN
(�19
13.6
77)k
N11
6K
now
nH
ex5H
exN
Ac4
Neu
Ac1
800.
591
4�
0.3
$VcS
ND
N(�
1913
.677
)kN
116
Kno
wn
Hex
5Hex
NA
c4N
euA
c110
66.7
833
�1.
7$V
cSN
DN
(�19
14.6
97)k
N11
6K
now
nH
ex5H
exN
Ac4
dH
ex2
800.
844
4�
2.5
$VcS
ND
N(�
1914
.697
)kN
116
Kno
wn
Hex
5Hex
NA
c4d
Hex
210
67.1
253
0.6
$VcS
ND
N(�
2059
.735
)ˆkN
116
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
842.
598
4�
3.0
$VcS
ND
N(�
2059
.735
)kN
116
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
836.
857
43.
1$V
cSN
DN
(�20
59.7
35)k
N11
6K
now
nH
ex5H
exN
Ac4
dH
ex1N
euA
c111
15.4
723
1.6
$VcS
ND
N(�
2350
.830
)kN
116
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
909.
629
40.
3S
tab
ilin-
1S
TAB
1_H
UM
AN
$ELL
QH
HG
LVP
QIE
AA
TAY
TIFV
PTn
RS
LEA
QG
N(�
2366
.825
)ˆSS
HLD
AD
TVR
N11
78P
oten
tial
Hex
6Hex
NA
c4N
euA
c273
2.44
310
3.2
$ELk
GD
GP
FTIF
VP
HA
DLM
SN
(�56
8.21
2)LS
QD
ELA
RIR
N16
26K
now
nH
ex1H
exN
Ac2
1097
.310
4�
5.4
$LLP
AH
sGLS
LIIS
DA
GP
DN
(�89
2.31
7)S
SW
AP
VA
PG
TVV
VS
RN
2424
Kno
wn
Hex
3Hex
NA
c265
5.46
47
�7.
8$I
LTM
AnQ
VLA
VN
(�10
95.3
97)IS
EE
GR
N60
6P
oten
tial
Hex
3Hex
NA
c382
0.39
04
�9.
4$G
N(�
1216
.423
)cS
DG
IQG
NG
AcL
cFP
DY
kN
745
Pot
entia
lH
ex5H
exN
Ac2
975.
155
4�
10.2
Sup
pre
ssor
ofG
2al
lele
ofS
KP
1ho
mol
ogS
UG
T1_H
UM
AN
$RA
MN
(�56
8.21
2)kS
FME
SG
GTV
LSTN
WsD
VG
kN
329
Nov
elH
ex1H
exN
Ac2
981.
480
46.
2
Ang
iop
oiet
in-1
rece
pto
rTI
E2_
HU
MA
N$I
SN
(�56
8.21
2)IT
HS
SA
VIs
WTI
LDG
YS
ISS
ITIR
N64
9P
oten
tial
Hex
1Hex
NA
c276
2.38
25
�3.
1Te
tras
pan
in-3
TSN
3_H
UM
AN
$TY
N(�
1825
.661
)GTN
PD
AA
SR
AID
YV
QR
N12
7P
oten
tial
Hex
5Hex
NA
c510
41.4
544
�7.
9Th
rom
bos
pon
din
-1TS
P1_
HU
MA
N$V
VN
(�19
13.6
77)S
TTG
PG
EH
LRN
1067
Kno
wn
Hex
5Hex
NA
c4N
euA
c187
7.14
14
0.2
Vas
cula
ren
dot
helia
lgro
wth
fact
orre
cep
tor
1V
GFR
1_H
UM
AN
$RIIW
DS
RkG
FIIS
N(�
892.
317)
ATY
kN
196
Pot
entia
lH
ex3H
exN
Ac2
934.
508
48.
1$S
VN
(�89
2.31
8)TS
VhI
YD
kAFI
TVk
N32
3P
oten
tial
Hex
3Hex
NA
c287
6.46
64
10.2
$WFW
HP
cNH
N(�
1419
.502
)HS
EA
RcD
FcS
NN
EE
SFI
LD
AD
SnM
GN
RN
474
Pot
entia
lH
ex5H
exN
Ac3
1017
.415
61.
3
$mA
ITkE
hSIT
LNLT
IMN
(�12
57.4
50)ˆV
SLQ
DS
GTY
AcR
AR
N62
5P
oten
tial
Hex
4Hex
NA
c310
51.7
025
�0.
2
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 15
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

TAB
LEII—
cont
inue
d
Pro
tein
nam
eU
niP
rot
IDP
eptid
eG
lyco
site
Typ
eG
lyca
nsO
bse
rved
m/z
Z�
Mas
s(p
pm
)
von
Will
ebra
ndfa
ctor
VW
F_H
UM
AN
$YFN
(�20
59.7
35)ˆk
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
776.
330
4�
4.7
$YFN
(�20
59.7
35)ˆk
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
1034
.440
3�
1.2
$YFN
(�20
59.7
35)k
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
1027
.114
3�
0.1
$YFN
(�20
59.7
36)k
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
770.
587
4�
0.6
$YFN
(�22
04.7
72)k
N15
6P
oten
tial
Hex
5Hex
NA
c4N
euA
c210
75.7
983
3.0
$YFN
(�22
21.7
89)k
N15
6P
oten
tial
Hex
6Hex
NA
c4d
Hex
1Neu
Ac1
1081
.467
30.
8$Y
FN(�
2350
.830
)ˆˆk
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
854.
851
4�
3.2
$YFN
(�23
50.8
30)ˆk
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
848.
856
4�
0.9
$YFN
(�23
50.8
30)ˆk
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
1131
.473
3�
0.2
$YFN
(�23
50.8
30)k
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
843.
362
40.
5$Y
FN(�
2350
.830
)kN
156
Pot
entia
lH
ex5H
exN
Ac4
dH
ex1N
euA
c211
24.1
453
�0.
7$Y
FN(�
2350
.831
)ˆkN
156
Pot
entia
lH
ex5H
exN
Ac4
dH
ex1N
euA
c284
8.85
74
0.0
$YFN
(�23
50.8
31)k
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
843.
362
41.
2$Y
FN(�
2351
.851
)ˆkN
156
Pot
entia
lH
ex5H
exN
Ac4
dH
ex3N
euA
c184
9.61
34
�0.
2$Y
FN(�
2366
.825
)ˆkN
156
Pot
entia
lH
ex5H
exN
Ac4
dH
ex1N
euA
c1N
euG
c185
3.09
84
�9.
4
$YFN
(�23
66.8
25)k
N15
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
Neu
Gc1
1129
.813
30.
5
$YFN
(�27
15.9
63)ˆk
N15
6P
oten
tial
Hex
6Hex
NA
c5d
Hex
1Neu
Ac2
940.
387
4�
3.7
$YFN
(�27
15.9
63)k
N15
6P
oten
tial
Hex
6Hex
NA
c5d
Hex
1Neu
Ac2
934.
643
4�
0.8
$YFN
(�27
16.9
83)k
N15
6P
oten
tial
Hex
6Hex
NA
c5d
Hex
3Neu
Ac1
934.
898
4�
1.5
$YFN
(�28
61.0
01)k
N15
6P
oten
tial
Hex
6Hex
NA
c5N
euA
c397
1.40
94
3.7
$YFN
(�30
07.0
58)k
N15
6P
oten
tial
Hex
6Hex
NA
c5d
Hex
1Neu
Ac3
1007
.667
4�
2.2
$YFN
(�30
09.0
74)k
N15
6P
oten
tial
Hex
8Hex
NA
c710
07.9
234
0.6
$YFN
(�30
26.0
89)k
N15
6P
oten
tial
Hex
9Hex
NA
c7d
Hex
110
12.6
764
�2.
0$Y
FN(�
3372
.190
)kN
156
Pot
entia
lH
ex7H
exN
Ac6
dH
ex1N
euA
c310
98.9
514
�0.
9$G
DIL
QR
VR
EIR
YQ
GG
N(�
1378
.476
)RN
1574
Kno
wn
Hex
6Hex
NA
c290
9.43
94
�0.
2$G
DIL
QR
VR
EIR
YQ
GG
N(�
568.
212)
RTN
TGLA
LRN
1574
Kno
wn
Hex
1Hex
NA
c291
2.98
44
�4.
0$Y
QG
GN
(�20
59.7
35)R
N15
74K
now
nH
ex5H
exN
Ac4
dH
ex1N
euA
c199
3.41
13
1.2
$AS
PP
SS
ScN
(�20
59.7
35)IS
SG
Em
Qk
N21
1P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
1073
.955
4�
2.4
$AS
PP
SS
ScN
(�23
50.8
30)ˆI
SS
GE
mQ
kN
211
Pot
entia
lH
ex5H
exN
Ac4
dH
ex1N
euA
c211
51.7
364
8.9
$AS
PP
SS
ScN
(�23
50.8
30)IS
SG
Em
Qk
N21
1P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
1146
.231
40.
3$N
(�20
59.7
35)V
ScP
QLE
VP
VcP
SG
FQLS
ckN
2546
Kno
wn
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
984.
037
5�
3.6
$GQ
VY
LQcG
TPcN
(�20
59.7
35)L
TcR
N66
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac1
1053
.444
40.
9$G
QV
YLQ
cGTP
cN(�
2075
.730
)LTc
RN
666
Pot
entia
lH
ex6H
exN
Ac4
Neu
Ac1
1057
.695
42.
2$G
QV
YLQ
cGTP
cN(�
2076
.750
)LTc
RN
666
Pot
entia
lH
ex6H
exN
Ac4
dH
ex2
1058
.193
4�
0.6
$GQ
VY
LQcG
TPcN
(�23
50.8
30)L
TcR
N66
6P
oten
tial
Hex
5Hex
NA
c4d
Hex
1Neu
Ac2
1126
.222
44.
5B
ovin
ep
rote
ins
Alp
ha-1
-aci
dgl
ycop
rote
inA
1AG
_BO
VIN
$QN
(�28
61.0
00)ˆG
TLS
kN
104
Pot
entia
lH
ex6H
exN
Ac5
Neu
Ac3
1020
.428
40.
4$Q
N(�
2861
.000
)GTL
Sk
N10
4P
oten
tial
Hex
6Hex
NA
c5N
euA
c310
14.9
344
1.8
$QN
(�28
80.0
31)G
TLS
kN
104
Pot
entia
lH
ex9H
exN
Ac7
1019
.690
40.
5$N
PE
YN
(�28
14.0
10)ˆˆ
kN
57P
oten
tial
Hex
5Hex
NA
c7N
euA
c210
18.9
234
6.2
Alp
ha-f
etop
rote
inFE
TA_B
OV
IN$A
EN
(�12
00.4
28)A
TEcF
ETk
N19
7P
oten
tial
Hex
4Hex
NA
c2d
Hex
198
3.42
83
�5.
8$A
EN
(�22
04.7
72)A
TEcF
ETk
N19
7P
oten
tial
Hex
5Hex
NA
c4N
euA
c298
8.91
34
0.1
$AN
(�22
04.7
72)F
TEIQ
kN
251
Pot
entia
lH
ex5H
exN
Ac4
Neu
Ac2
901.
651
42.
5A
lpha
-2-H
S-g
lyco
pro
tein
FETU
A_B
OV
IN$k
LcP
DcP
LLA
PLN
(�22
04.7
72)D
SR
N15
6K
now
nH
ex5H
exN
Ac4
Neu
Ac2
905.
409
50.
3$k
LcP
DcP
LLA
PLN
(�28
61.0
00)D
SR
N15
6K
now
nH
ex6H
exN
Ac5
Neu
Ac3
1036
.654
5�
1.0
$LcP
DcP
LLA
PLN
(�22
04.7
72)D
SR
N15
6K
now
nH
ex5H
exN
Ac4
Neu
Ac2
1043
.195
4�
1.7
Pep
tide
mod
ifica
tion
sym
bol
s:$,
N-t
erm
inal
TMT0
lab
ellin
g(�
224.
152)
;ˆ,N
aad
duc
ton
glyc
an(�
21.9
82);
ˆˆ,2
Na
add
ucto
ngl
ycan
(�43
.964
);c,
carb
amid
omet
hyla
tion
ofcy
stei
ne(�
57.0
21);
h,ox
idat
ion
ofhi
stid
ine
(�15
.995
);k,
TMT0
lab
elin
gof
lysi
ne(�
224.
152)
;m,o
xid
atio
nof
met
hion
ine
(�15
.995
);n,
dea
mid
atio
nof
asp
arag
ine
(�0.
984)
;q,G
ln-�
pyr
o-G
luan
dlo
ssof
TMT0
(�24
1.17
9);
s,p
hosp
hory
latio
nof
serin
e(�
79.9
66).
Und
erlin
edgl
ycop
eptid
esw
ere
also
det
ecte
dvi
ath
eP
NG
ase
F�
H218O
met
hod
(sup
ple
men
talT
able
S4)
.
Glycoproteomics of the Endothelial Secretome
16 Molecular & Cellular Proteomics 12.4
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

ture, sugars tend to be arranged in branched polymers, re-sulting in an exponential increase of possible polysaccharidecombinations. Theoretically, just six monosaccharides can
give rise to 1012 different glycan structures. This high diversityof protein-bound glycans requires a combination of differenttechniques. For example, new MS-based methods were de-
FIG. 3. HCD-pd-ETD fragmentation. Full MS showing the different glycoforms of the same peptide sequence (A). Characteristic oxoniumion detected by HCD at m/z � 204.09 (B). This HexNAc signature triggered an ETD scan to identify the peptide sequence and confirm theglycosylation site (C).
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 17
COLOR
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

veloped to profile the cell surface N-glycoproteome as adifferentiation marker for stem cells (32). We applied a com-bination of different glycoproteomics techniques to furtherenrich for secreted and shed membrane proteins and revealpotential glycosylation sites within the endothelial secretome.Glycoproteins play important roles in many biological pro-cesses related to ECs, such as angiogenesis, in which thestructural change of the glycans will determine the attachment
property of cells and influence cell-to-cell interactions (33).Interestingly, vWF is a glycoprotein produced uniquely by ECsand megakaryocytes. Previous publications investigatingvWF isolated from plasma failed to identify glycosylation siteswithin the propeptide (29). In plasma, the concentration of thepropeptide is about one-tenth of the concentration of maturevWF (34, 35). In the conditioned medium of ECs, however, weobserved several glycopeptides of the propeptide. Thus, the
FIG. 4. Comparison of HCD-pd-ETD and HCD-alt-ETD. The two methods, HCD-pd-ETD (blue) and HCD-alt-ETD (red), displayed distinctdistributions of the observed m/z, charge state, mass of identified peptides (M�H), and glycan mass, as well as the intensity of the precursorions and the ByonicsTM score (all y-axes). The x-axes represent index numbers after proteins were sorted by their corresponding y-axis valuefrom lower to higher (A). There was limited overlap in the identified glycopeptides (B). C, the HCD-pd-ETD method preferentially identifiedcomplex/hybrid glycans.
Glycoproteomics of the Endothelial Secretome
18 Molecular & Cellular Proteomics 12.4
COLOR
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

endothelial secretome allowed us to interrogate the glycosy-lation sites of von Willebrand antigen 2, the N-terminal cleav-age product of vWF that aids N-terminal multimerization andprotein compartmentalization of mature vWF in storagegranules.
Conventional Methods for Glycoproteomics—As reviewedelsewhere (36), conventional glycoproteomic methods involvethe enrichment of glycoproteins (typically with lectins likeConA and wheat germ agglutinin), cleavage of the glycans,and identification of the remaining peptide sequence. Themost widely used method for detecting N-glycopeptides isdigestion by PNGase F. PNGase F cleaves the GlcNAc mol-
ecule closest to the peptide (37). After PNGase F treatment,formerly N-linked glycosylated peptides are identified basedon the conversion of Asn to Asp (deamidation) in the consen-sus motif for N-linked glycosylation (sequence N-X(not P)-S/T). This method has two major caveats. The first of these is ahigh false positive rate due to spontaneous deamidation. Asn-Gly sites, in particular, are prone to spontaneous deamidation(38–40). To reduce false positives, PNGase F treatment isperformed in 18O water, adding a larger tag of 2.99 Da.Importantly, all known glycosyltransferases that mediate N-linked glycosylation are supposed to recognize a consensusmotif, and this consensus sequence for N-linked glycosylation
FIG. 5. Glycoprotein enrichment for validation. A, spectral count of input, glycoprotein-enriched fraction (GP), and flow-through fraction(FT) from representative glycoproteins and non-glycoproteins. B, complementarity of the different methods (HCD-ETD, PNGase F � H2
18Otreatment, and glycoprotein enrichment). Only 18 glycoproteins were consistently identified.
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 19
COLOR
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

FIG. 6. Sequence coverage for vWF. A, schematic illustration of vWF sequence. Coverage is highlighted in green, and potentialglycosylation sites are shown in red. A large hexagon indicates a glycosylation site with a reference in the Uniprot database. By using theHCD-ETD (H) or PNGase F (P) method, we confirmed six N-glycosylation sites on vWF. B, ETD spectra of glycopeptides identified via HCD-ETD(N156, N211, N666, N1574). The following abbreviations are used: a, y, g, k � TMT modified Ala, Tyr, Gly, and Lys, respectively; c �carboxyamidomethylation of Cys; m � oxidation of Met.
Glycoproteomics of the Endothelial Secretome
20 Molecular & Cellular Proteomics 12.4
COLOR
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

must be taken into consideration (41). 2) The second caveat isthat after PNGase F cleavage, the released sugars can beanalyzed separately, but the link to the identified peptideswith deamidated amino acids is lost (42, 43). Ideally, intactglycopeptides are analyzed directly via MS/MS even in com-plex biological samples.
Novel HCD-ETD Method—HCD fragmentation mostlybreaks glycosidic bonds, whereas ETD preserves the glycanattachment and fragments the peptide backbone, providingmore complete peptide sequence information. CurrentMS/MS acquisition strategies for glycopeptide analysis relyon the acquisition of MS/MS spectra for all precursor ions. Inthis study, HCD was employed to generate glycan oxoniumions and trigger an ETD spectrum in a data-dependent man-ner. HCD presents the sugar signatures within the low m/zrange, which are otherwise lost as a result of the one-third ruleof ion trap fragmentation (44). Glycopeptides with terminalHexNAc generate typically an m/z 204.0864 oxonium ion andits fragments at m/z 168.0653 and 138.0550. The oxonium ionand its fragments are measured with the high mass accuracyof the Orbitrap analyzer, and the unambiguous identificationof the glycan oxonium ion generated by the HCD scan servesas a diagnostic marker for glycopeptides. This approach wascompared against conventional HCD-alt-ETD scans using acomplex biological sample. The HCD-alt-ETD preferentiallydetects higher charged and higher intensity precursor ionsthan HCD-pd-ETD. This might be because (i) a higher chargeincreases ETD fragmentation efficiency, resulting in moreidentified glycopeptides; (ii) high-charged precursors did notproduce HCD spectra of sufficient quality to trigger ETDbased on the diagnostic oxonium ions; or (iii) more abundantpeptides were selected in HCD-alt-ETD because the instru-ment duty cycle is less efficient than in HCD-pd-ETD. Overall,the combination of multiple MS methods used in our studyprovides greater confidence in the identification of glycopep-tides than studies relying on a single approach and offerscomplementary advantages in the assessment of the glyco-proteome, notably, the simultaneous identification of the pep-tide sequence, the glycosylation site, and the glycancomposition.
Study Limitations—N-linked and O-linked glycosylation arethe two most common forms of glycosylation in mammals(45). Only N-linked glycosylation was analyzed in the presentstudy. Unlike N-linked glycosylation, O-linked glycosylationhas no consensus site (46). This makes the analysis of O-linked glycopeptides a more daunting task (47). Lectins arewidely used for glycoprotein enrichment. There are manytypes of lectins binding to different sugars, such as ConA(binds to �-D-mannosyl and �-D-glucosyl residues) and wheatgerm agglutinin (binds to GlcNAc�1–4GlcNAc�1–4GlcNAc-and N-acetylneuraminic acid). Here we used only ConA as aproof of principle to demonstrate the complementary resultsof multiple glycoprotein identification methods. ConA isknown to display nonspecific avidity for hydrophobic ligands
such as certain domains of tropomyosin (48). Furthermore,the standard protocol for the ConA glycoprotein enrichmentkit is not optimized for cleanliness, and several known non-glycoproteins were also detected in the eluate samples. Se-quential washes with low- and high-ionic-strength buffersbefore elution might have reduced this contamination (49).Also, mixing different lectins would increase the coverage ofthe glycoproteome in biological samples (39). Additional ef-forts are needed for a complete structural characterization ofprotein glycosylation; in particular, the quantitation of theoccupancy rates and the identification of the glycan structureas complex/hybrid glycans cannot be discerned via our cur-rent MS approach.
CONCLUSIONS
Cardiovascular diseases arise from exposure to risk factorsthat induce complex pathophysiological perturbations of en-dothelial protein secretion. The recent advent of new pro-teomic technologies has enabled us to obtain information onthe dynamic regulation of endothelial protein secretion. Wepresent results from an extensive glycoproteomic analysiswith information on glycan composition obtained via a directMS method. Future proteomics studies linking endothelialsecretory processes to cardiovascular risk factors and endo-thelial dysfunction will provide valuable insights about themechanisms contributing to cardiovascular disease.
Acknowledgment—We thank Dr. Sarah Langley for assistance withthe gene ontology annotation.
* This work was funded by the Department of Health via a NationalInstitute for Health Research (NIHR) Biomedical Research Centreaward to Guy’s and St. Thomas’ NHS Foundation Trust in partnershipwith King’s College London and King’s College Hospital NHS Foun-dation Trust. Dr. M. Mayr is supported by a Senior Research Fellow-ship of the British Heart Foundation.
□S This article contains supplemental material.** To whom correspondence should be addressed: Prof. Manuel
Mayr, Cardiovascular Division, King’s British Heart Foundation Cen-tre, King’s College London, 125 Coldharbour Lane, London SE5 9NU,UK, Tel.: �44 (0) 20 7848 5132, E-mail: [email protected].
REFERENCES
1. Mayr, M., Mayr, U., Chung, Y. L., Yin, X., Griffiths, J. R., and Xu, Q. (2004)Vascular proteomics: linking proteomic and metabolomic changes. Pro-teomics 4, 3751–3761
2. Pula, G., Perera, S., Prokopi, M., Sidibe, A., Boulanger, C. M., and Mayr, M.(2008) Proteomic analysis of secretory proteins and vesicles in vascularresearch. Proteomics Clin. Appl. 2, 882–891
3. Pober, J. S., and Sessa, W. C. (2007) Evolving functions of endothelial cellsin inflammation. Nat. Rev. Immunol. 7, 803–815
4. Cai, H., and Harrison, D. G. (2000) Endothelial dysfunction in cardiovasculardiseases: the role of oxidant stress. Circ. Res. 87, 840–844
5. Libby, P., and Simon, D. I. (2001) Inflammation and thrombosis: the clotthickens. Circulation 103, 1718–1720
6. Ross, R. (1999) Atherosclerosis—an inflammatory disease. N. Engl. J. Med.340, 115–126
7. Pellitteri-Hahn, M. C., Warren, M. C., Didier, D. N., Winkler, E. L., Mirza,S. P., Greene, A. S., and Olivier, M. (2006) Improved mass spectrometricproteomic profiling of the secretome of rat vascular endothelial cells. J.Proteome Res. 5, 2861–2864
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 21
ZSI
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

8. Tunica, D. G., Yin, X., Sidibe, A., Stegemann, C., Nissum, M., Zeng, L.,Brunet, M., and Mayr, M. (2009) Proteomic analysis of the secretome ofhuman umbilical vein endothelial cells using a combination of free-flowelectrophoresis and nanoflow LC-MS/MS. Proteomics 9, 4991–4996
9. Flora, J. W., Edmiston, J., Secrist, R., Li, G., Rana, G. S., Langston, T. B.,and McKinney, W. (2008) Identification of in vitro differential cell secre-tions due to cigarette smoke condensate exposure using nanoflow cap-illary liquid chromatography and high-resolution mass spectrometry.Anal. Bioanal. Chem. 391, 2845–2856
10. Burghoff, S., and Schrader, J. (2011) Secretome of human endothelial cellsunder shear stress. J. Proteome Res. 10, 1160–1169
11. Hocking, S. L., Wu, L. E., Guilhaus, M., Chisholm, D. J., and James, D. E.(2010) Intrinsic depot-specific differences in the secretome of adiposetissue, preadipocytes, and adipose tissue-derived microvascular endo-thelial cells. Diabetes 59, 3008–3016
12. Griffoni, C., Di Molfetta, S., Fantozzi, L., Zanetti, C., Pippia, P., Tomasi, V.,and Spisni, E. (2011) Modification of proteins secreted by endothelialcells during modeled low gravity exposure. J. Cell. Biochem. 112,265–272
13. Brioschi, M., Lento, S., Tremoli, E., and Banfi, C. (2013) Proteomic analysisof endothelial cell secretome: a means of studying the pleiotropic effectsof Hmg-CoA reductase inhibitors. J. Proteomics (In press)
14. Castagna, M., Takai, Y., Kaibuchi, K., Sano, K., Kikkawa, U., and Nishizuka,Y. (1982) Direct activation of calcium-activated, phospholipid-dependentprotein kinase by tumor-promoting phorbol esters. J. Biol. Chem. 257,7847–7851
15. Jacobs, J. M., Waters, K. M., Kathmann, L. E., Camp, D. G., 2nd, Wiley,H. S., Smith, R. D., and Thrall, B. D. (2008) The mammary epithelial cellsecretome and its regulation by signal transduction pathways. J. Pro-teome Res. 7, 558–569
16. Zampetaki, A., Kiechl, S., Drozdov, I., Willeit, P., Mayr, U., Prokopi, M.,Mayr, A., Weger, S., Oberhollenzer, F., Bonora, E., Shah, A., Willeit, J.,and Mayr, M. (2010) Plasma microRNA profiling reveals loss of endothe-lial miR-126 and other microRNAs in type 2 diabetes. Circ. Res. 107,810–817
17. Margariti, A., Zampetaki, A., Xiao, Q., Zhou, B., Karamariti, E., Martin, D.,Yin, X., Mayr, M., Li, H., Zhang, Z., De Falco, E., Hu, Y., Cockerill, G., Xu,Q., and Zeng, L. (2010) Histone deacetylase 7 controls endothelial cellgrowth through modulation of beta-catenin. Circ. Res. 106, 1202–1211
18. Yin, X., Cuello, F., Mayr, U., Hao, Z., Hornshaw, M., Ehler, E., Avkiran, M.,and Mayr, M. (2010) Proteomics analysis of the cardiac myofilamentsubproteome reveals dynamic alterations in phosphatase subunit distri-bution. Mol. Cell. Proteomics 9, 497–509
19. Didangelos, A., Yin, X., Mandal, K., Baumert, M., Jahangiri, M., and Mayr,M. (2010) Proteomics characterization of extracellular space compo-nents in the human aorta. Mol Cell. Proteomics 9, 2048–2062
20. Mayr, M., Grainger, D., Mayr, U., Leroyer, A. S., Leseche, G., Sidibe, A.,Herbin, O., Yin, X., Gomes, A., Madhu, B., Griffiths, J. R., Xu, Q., Tedgui,A., and Boulanger, C. M. (2009) Proteomics, metabolomics, and immu-nomics on microparticles derived from human atherosclerotic plaques.Circ. Cardiovasc. Genet. 2, 379–388
21. Pula, G., Mayr, U., Evans, C., Prokopi, M., Vara, D. S., Yin, X., Astroulakis,Z., Xiao, Q., Hill, J., Xu, Q., and Mayr, M. (2009) Proteomics identifiesthymidine phosphorylase as a key regulator of the angiogenic potential ofcolony-forming units and endothelial progenitor cell cultures. Circ. Res.104, 32–40
22. Urbich, C., De Souza, A. I., Rossig, L., Yin, X., Xing, Q., Prokopi, M.,Drozdov, I., Steiner, M., Breuss, J., Xu, Q., Dimmeler, S., and Mayr, M.(2011) Proteomic characterization of human early pro-angiogenic cells. J.Mol. Cell. Cardiol. 50, 333–336
23. Hagglund, P., Bunkenborg, J., Elortza, F., Jensen, O. N., and Roepstorff, P.(2004) A new strategy for identification of N-glycosylated proteins andunambiguous assignment of their glycosylation sites using HILIC enrich-ment and partial deglycosylation. J. Proteome Res. 3, 556–566
24. Saba, J., Dutta, S., Hemenway, E., and Viner, R. (2012) Increasing theproductivity of glycopeptides analysis by using higher-energy collisiondissociation-accurate mass-product-dependent electron transfer disso-ciation. Int. J. Proteomics 2012, 560391
25. Bern, M., Cai, Y., and Goldberg, D. (2007) Lookup peaks: a hybrid of denovo sequencing and database search for protein identification by tan-dem mass spectrometry. Anal Chem. 79, 1393–1400
26. Antonov, A. S., Lukashev, M. E., Romanov, Y. A., Tkachuk, V. A., Repin,V. S., and Smirnov, V. N. (1986) Morphological alterations in endothelialcells from human aorta and umbilical vein induced by forskolin andphorbol 12-myristate 13-acetate: a synergistic action of adenylate cy-clase and protein kinase C activators. Proc. Natl. Acad. Sci. U.S.A. 83,9704–9708
27. Sadler, J. E. (1998) Biochemistry and genetics of von Willebrand factor.Annu. Rev. Biochem. 67, 395–424
28. Matsui, T., Titani, K., and Mizuochi, T. (1992) Structures of the asparagine-linked oligosaccharide chains of human von Willebrand factor. Occur-rence of blood group A, B, and H(O) structures. J. Biol. Chem. 267,8723–8731
29. Canis, K., McKinnon, T. A., Nowak, A., Haslam, S. M., Panico, M., Morris,H. R., Laffan, M. A., and Dell, A. (2012) Mapping the N-glycome of humanvon Willebrand factor. Biochem. J. 447, 217–228
30. Simak, J., Holada, K., and Vostal, J. G. (2002) Release of annexin V-bindingmembrane microparticles from cultured human umbilical vein endothelialcells after treatment with camptothecin. BMC Cell Biol. 3, 11
31. Hart, G. W., and Copeland, R. J. (2010) Glycomics hits the big time. Cell143, 672–676
32. Wollscheid, B., Bausch-Fluck, D., Henderson, C., O’Brien, R., Bibel, M.,Schiess, R., Aebersold, R., and Watts, J. D. (2009) Mass-spectrometricidentification and relative quantification of N-linked cell surface glyco-proteins. Nat. Biotechnol. 27, 378–386
33. Leth-Larsen, R., Lund, R. R., and Ditzel, H. J. (2010) Plasma membraneproteomics and its application in clinical cancer biomarker discovery.Mol. Cell. Proteomics 9, 1369–1382
34. Madabhushi, S. R., Shang, C., Dayananda, K. M., Rittenhouse-Olson, K.,Murphy, M., Ryan, T. E., Montgomery, R. R., and Neelamegham, S.(2012) von Willebrand factor (VWF) propeptide binding to VWF D’D3domain attenuates platelet activation and adhesion. Blood 119,4769–4778
35. van Mourik, J. A., Boertjes, R., Huisveld, I. A., Fijnvandraat, K., Pajkrt, D.,van Genderen, P. J., and Fijnheer, R. (1999) von Willebrand factor pro-peptide in vascular disorders: a tool to distinguish between acute andchronic endothelial cell perturbation. Blood 94, 179–185
36. Pan, S., Chen, R., Aebersold, R., and Brentnall, T. A. (2011) Mass spec-trometry based glycoproteomics—from a proteomics perspective. Mol.Cell. Proteomics 10, R110.003251
37. Maley, F., Trimble, R. B., Tarentino, A. L., and Plummer, T. H., Jr. (1989)Characterization of glycoproteins and their associated oligosaccharidesthrough the use of endoglycosidases. Anal. Biochem. 180, 195–204
38. Flatmark, T. (1967) Multiple molecular forms of bovine heart cytochrome c.V. A comparative study of their physicochemical properties and theirreactions in biological systems. J. Biol. Chem. 242, 2454–2459
39. Zielinska, D. F., Gnad, F., Wisniewski, J. R., and Mann, M. (2010) Precisionmapping of an in vivo N-glycoproteome reveals rigid topological andsequence constraints. Cell 141, 897–907
40. Angel, P. M., Lim, J. M., Wells, L., Bergmann, C., and Orlando, R. (2007) Apotential pitfall in 18O-based N-linked glycosylation site mapping. RapidCommun. Mass Spectrom. 21, 674–682
41. Palmisano, G., Melo-Braga, M. N., Engholm-Keller, K., Parker, B. L., andLarsen, M. R. (2012) Chemical deamidation: a common pitfall in large-scale N-linked glycoproteomic mass spectrometry-based analyses. J.Proteome Res. 11, 1949–1957
42. Wuhrer, M., Deelder, A. M., and Hokke, C. H. (2005) Protein glycosylationanalysis by liquid chromatography-mass spectrometry. J. Chromatogr. BAnalyt. Technol. Biomed. Life Sci. 825, 124–133
43. Morelle, W., and Michalski, J. C. (2007) Analysis of protein glycosylation bymass spectrometry. Nat. Protoc. 2, 1585–1602
44. Zhang, M. Y., Pace, N., Kerns, E. H., Kleintop, T., Kagan, N., and Sakuma,T. (2005) Hybrid triple quadrupole-linear ion trap mass spectrometry infragmentation mechanism studies: application to structure elucidationof buspirone and one of its metabolites. J. Mass Spectrom. 40,1017–1029
45. Tissot, B., North, S. J., Ceroni, A., Pang, P. C., Panico, M., Rosati, F.,Capone, A., Haslam, S. M., Dell, A., and Morris, H. R. (2009) Glycopro-teomics: past, present and future. FEBS Lett. 583, 1728–1735
46. Hang, H. C., Yu, C., Kato, D. L., and Bertozzi, C. R. (2003) A metaboliclabeling approach toward proteomic analysis of mucin-type O-linkedglycosylation. Proc. Natl. Acad. Sci. U.S.A. 100, 14846–14851
Glycoproteomics of the Endothelial Secretome
22 Molecular & Cellular Proteomics 12.4
AQ: Q
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018

47. Zhao, P., Viner, R., Teo, C. F., Boons, G. J., Horn, D., and Wells, L. (2011)Combining high-energy C-trap dissociation and electron transfer disso-ciation for protein O-GlcNAc modification site assignment. J. ProteomeRes. 10, 4088–4104
48. Owen, J. B., Di Domenico, F., Sultana, R., Perluigi, M., Cini, C., Pierce,W. M., and Butterfield, D. A. (2009) Proteomics-determined differences inthe concanavalin-A-fractionated proteome of hippocampus and inferior
parietal lobule in subjects with Alzheimer’s disease and mild cognitiveimpairment: implications for progression of AD. J. Proteome Res. 8,471–482
49. Ghosh, D., Krokhin, O., Antonovici, M., Ens, W., Standing, K. G., Beavis,R. C., and Wilkins, J. A. (2004) Lectin affinity as an approach to theproteomic analysis of membrane glycoproteins. J. Proteome Res. 3,841–850
Glycoproteomics of the Endothelial Secretome
Molecular & Cellular Proteomics 12.4 23
tapraid4/zjw-macp/zjw-macp/zjw00413/zjw4413-13a xppws S�4 19/2/13 11:49 4/Color Figure(s) 1–6 ARTNO: M112.024018