behavioral/systems/cognitive ... · behavioral/systems/cognitive...
TRANSCRIPT

Behavioral/Systems/Cognitive
Adenylyl Cyclase Type 5 Contributes to CorticostriatalPlasticity and Striatum-Dependent Learning
Mazen A. Kheirbek,1* Jon P. Britt,1* Jeff A. Beeler,2 Yoshihiro Ishikawa,4 Daniel S. McGehee,1,3 and Xiaoxi Zhuang1,2
1Committee on Neurobiology and Departments of 2Neurobiology and 3Anesthesia and Critical Care, The University of Chicago, Chicago, Illinois 60637,and 4Department of Cell Biology and Molecular Medicine, New Jersey Medical School, Newark, New Jersey 07103
Dopamine (DA)-dependent corticostriatal plasticity is thought to underlie incremental procedural learning. A primary effector of striatalDA signaling is cAMP, yet its role in corticostriatal plasticity and striatum-dependent learning remains unclear. Here, we show thatgenetic deletion of a striatum-enriched isoform of adenylyl cyclase, AC5 knock-out (AC5KO), impairs two forms of striatum-dependentlearning and corticostriatal synaptic plasticity. AC5KO mice were severely impaired in acquisition of a response strategy in the crossmaze, a striatum-dependent task requiring a correct body turn to find a goal arm. In addition, AC5KO mice were impaired in acquisitionof a motor skill, as assessed by the accelerated rotarod. Slice electrophysiology revealed a deficit in corticostriatal long-term depression(LTD) after high-frequency stimulation of tissue from AC5KO mice. LTD was rescued by activation of either presynaptic cannabinoid type1 (CB1 ) receptors or postsynaptic metabotropic glutamate receptors (mGluRs), suggesting a postsynaptic role of AC5– cAMP, upstreamof endocannabinoid release. In striatopallidal-projecting medium spiny neurons, DA D2 receptors are negatively coupled to cAMPproduction, and activation of these receptors is required for endocannabinoid release and corticostriatal LTD. Recordings from striato-pallidal neurons indicated that this is mediated by AC5, because coactivation of D2 and mGluRs could induce LTD in wild-type but not inAC5KO neurons. To further examine the role of cAMP in corticostriatal plasticity, we elevated cAMP in striatal neurons of wild-type micevia the recording electrode. Under these conditions, corticostriatal LTD was eliminated. Together, these data suggest an AC5– cAMP–endocannabinoid–CB1 signaling pathway in corticostriatal plasticity and striatum-dependent learning.
IntroductionIn humans, the dorsal striatum plays a critical role in procedurallearning, the memory for motor skills and stimulus-responsehabits, that is dissociable from hippocampus-dependent declar-ative learning, the memory for facts and events (Packard et al.,1989; Packard and White, 1991; Packard and McGaugh, 1996;Packard and Knowlton, 2002; Schroeder et al., 2002; Squire,2004; Yin et al., 2004, 2005). In rodents, lesion studies have indi-cated that striatum-dependent learning of stimulus-responsehabits is dissociable from hippocampus-dependent learning ofspatial information (Packard and McGaugh, 1996; Packard andKnowlton, 2002; Yin and Knowlton, 2006). This dissociation hasbeen documented through use of a cross maze, in which inacti-vation of the dorsal striatum impairs the animal’s ability to learnto make the correct body turn at a choice point to find the goalarm (Packard and McGaugh, 1996). The dorsal striatum receivesdense glutamatergic innervation from cortical areas, as well adopaminergic input from midbrain nuclei. The interplay be-
tween dopamine (DA) and glutamate input to medium spinyneurons (MSNs) of the dorsal striatum has provided the basis formodels of reinforcement learning that suggest DA-dependentplasticity at corticostriatal synapses is necessary for striatum-dependent learning (Calabresi et al., 1992; Schultz et al., 1997;Reynolds et al., 2001; Reynolds and Wickens, 2002). However,few studies have addressed the role of DA-dependent intracellu-lar signaling pathways that underlie this form of striatum-dependent learning and how they contribute to DA-dependentcorticostriatal plasticity.
In brain slice recordings, the predominant form of synapticplasticity seen in the dorsolateral striatum is long-term depres-sion (LTD), which requires DA D2 receptor activation, increasesin intracellular calcium, and retrograde release of endocannabi-noids that inhibit glutamate release via presynaptic cannabinoidtype 1 (CB1) receptors (Calabresi et al., 1994, 2007; Gerdeman etal., 2002; Kreitzer and Malenka, 2007). Although modulation ofintracellular cAMP content is a direct downstream effect of DAreceptor activation (Neve et al., 2004), the role of cAMP in cor-ticostriatal plasticity has remained essentially uninvestigated.Two downstream targets of cAMP-dependent protein kinase(PKA), cAMP response element binding protein (CREB), and thedopamine and cAMP-regulated phosphoprotein 32 kDa (DARPP-32)have been implicated in corticostriatal LTD (Calabresi et al.,2000; Pittenger et al., 2006), but these proteins are also regulatedby non-cAMP-dependent signaling pathways (Kang et al., 2001;Svenningsson et al., 2004), leaving no direct evidence for a con-tribution of cAMP to corticostriatal LTD.
Received July 13, 2009; revised Aug. 5, 2009; accepted Aug. 6, 2009.This work was supported in part by National Institutes of Health Grants F31MH076422 (M.A.K.), F31DA023340
(J.P.B.), F32DA020427 and RO1DA25875-2 (J.A.B.), HL059139 and GM067773 (Y.I.), DA015918 (D.S.M.), andDA022269 and MH66216 (X.Z.) and the Edward Mallinckrodt Jr. Foundation (X.Z.). We thank Wei-Jen Tang forhelpful discussions in preparing this manuscript.
*M.A.K. and J.P.B. contributed equally to this work.Correspondence should be addressed to Daniel S. McGehee, Department of Anesthesia and Critical Care, The
University of Chicago, 5841 S. Maryland Avenue, MC4028, Chicago, IL 60637. E-mail: [email protected]:10.1523/JNEUROSCI.3343-09.2009
Copyright © 2009 Society for Neuroscience 0270-6474/09/2912115-10$15.00/0
The Journal of Neuroscience, September 30, 2009 • 29(39):12115–12124 • 12115

The highly conserved cAMP second-messenger system hasbeen implicated in synaptic plasticity and learning in many dif-ferent organisms (Kandel, 2001). The enzyme that catalyzescAMP production, adenylyl cyclase (AC), has at least ninemembrane-bound isoforms that have different expression pat-terns and regulatory properties (Cooper et al., 1998; Hanouneand Defer, 2001; Wang and Storm, 2003). Calcium/calmodulin-dependent AC isoforms have been extensively studied and areessential for induction of long-term potentiation (LTP) in thehippocampus, because they couple glutamate-mediated in-creases in intracellular calcium to cAMP accumulation (Xia et al.,1993; Wu et al., 1995; Matsuoka et al., 1997; Wong et al., 1999;Wang and Storm, 2003; Nicol et al., 2005). The dorsal striatumexpresses high levels of AC5 (Matsuoka et al., 1997), a primarydownstream effector of DA receptor signaling in the striatum.Mice that lack AC5 expression [AC5 knock-outs (AC5KO)] showseverely impaired striatal D1 and D2 receptor modulation ofcAMP signaling (Iwamoto et al., 2003). In addition, recent stud-ies have indicated that AC5KO mice are unable to predict rewarddelivery in a DA-dependent Pavlovian conditioning task (Kheirbeket al., 2008). Thus, the AC5KO mouse provides an opportunity totest the contribution of cAMP to corticostriatal plasticity and dorsalstriatum-dependent learning. Here, we demonstrate a critical rolefor AC5 in a striatum-dependent learning task, response learning inthe cross maze. This deficit in striatum-dependent learning wasconfirmed in the accelerated rotarod, a motor skill learning taskcorrelated previously with plasticity in the dorsal striatum. Inaddition, we reveal a role for AC5 and cAMP signaling in corti-costriatal LTD by regulating D2-mediated retrograde release ofendocannabinoids (Costa et al., 2004; Dang et al., 2006; Yin et al.,2009).
Materials and MethodsMiceAC5KO mice were generated as described previously (Iwamoto et al.,2003) and backcrossed to C57BL/6 for nine generations. All mice testedwere 8 –12 weeks of age, and behavioral experiments were conductedduring the light phase. Wild-type (WT) littermates were used as controls,and independent groups were used for each experiment. Drd2– en-hanced green fluorescent protein (EGFP) bacterial artificial chromo-some transgenic mice were obtained from the Mutant Mouse RegionalResource Center. Drd2–EGFP mice were crossed to AC5KO backgroundto generate AC5KO/Drd2–EGFP and WT/Drd2–EGFP littermate con-trols for electrophysiological experiments. All experiments were ap-proved by the Institutional Animal Care and Use Committee of theUniversity of Chicago.
In situ hybridizationMice were decapitated, and brains were fresh frozen on dry ice. Coronalsections (20 �m) were cut and thaw mounted on slides and stored at�80° until hybridization. Sections were fixed in 4% paraformaldehydefor 10 min and digested with proteinase K, followed by acetylation for 10min and prehybridization for 2 h at 65°C in a buffer containing 50%formamide, 20� SSC, 5� Denhardt’s solution, 250 �g/ml tRNA, and500 �g/ml fish sperm DNA. For radiolabeled probe, 10 6 cpm [ 33P]rUTPprobe per 50 �l of hybridization buffer was used for overnight incubationat 65°C. Signal was visualized after 48 h development on Kodak BioMaxMR film. For digoxigenin (DIG)-labeled probes, sections were incubatedwith 400 ng/ml probes overnight at 65°C. Slides were washed and incubatedin 1:5000 dilution of anti-DIG–alkaline phosphatase (AP) antibody (Roche)for 2 h and then visualized using nitroblue-tetrazolium-chloride (NBT)/5-bromo-4-chlor-indolyl-phosphate (BCIP) staining for alkaline phos-phatase. For double labeling in situ, sections were hybridized with DIGand biotin-labeled mRNA probes simultaneously. After washing, the sec-tions were incubated in 1:5000 dilution anti-biotin–AP antibody, fol-lowed by visualization with Fast Red (Sigma). Slides were photographed,
then washed in 0.1 M glycine, pH 2.2, and incubated in anti-DIG–AP,followed by development in NBT/BCIP solution.
AC5 and preproenkephalin (ENK) probes were isolated from a mousebrain cDNA library using the following primer pairs: AC5, 5�CTGGAG-GAGCGTCGAGGAAA3� and 5�GGTCCTGGGAGTTGGTGTGC3�; ENK,5�GGATGCAGATGAGGGAGACA3�and 5�AAACCCGTGACCCCAAC-TGC3�. The cDNA fragment was cloned into PCRII-TOPO vector(Invitrogen) and linearized, and antisense probe was transcribed us-ing the appropriate RNA polymerase.
Behavioral experimentsWater cross maze. Experiments were conducted in a water tank (whitewalls, 100 cm in diameter), with white Plexiglas walls inserted in the mazeto make alleyways. Water was at room temperature, and white nontoxicpaint (FunStuff) was added to obscure the hidden platform. A blackcurtain surrounded the maze to reduce spatial cues. The walls of the mazewere rotated every 10 trials to further reduce the contribution ofspatial cues. A trial was started by placing the mouse in the start armfacing the wall of the tank. Once the mouse made a right or left choice,a divider closed off the arm, and the mouse was allowed to either standon the platform for 15 s (correct choice) or explore the arm of themaze without the platform for 15 s (incorrect choice). Mice were thenremoved from the maze and placed under a heat lamp for 30 s beforebeginning the next trial. Trials were started randomly from either thenorth or the south end of the maze with the goal arm always to theright of the start arm during initial acquisition and to the left ofthe start arm during reversal learning. Correct or incorrect choiceswere recorded.
Morris water maze. Experiments were conducted as described previ-ously (Vorhees and Williams, 2006). Experiments were conducted in awater tank (white walls, 100 cm in diameter), with white Plexiglas wallsinserted in the maze to make alleyways. Water was at room temperature,and white non-toxic paint (FunStuff) was added to obscure the hiddenplatform. Around the tank, white paper cutouts of stars and arrows wereplaced on the black surrounding curtain to serve as spatial cues. Micewere first trained for 2 d, three trials a day with a visible platform (8 cmdiameter) for habituation purposes and to determine any swimmingdeficits. After habituation, training trials began. A 1 cm submergedplatform (8 cm diameter) was placed in one quadrant. During train-ing, mice were started from random start positions, and latency toreach the hidden platform was measured. Mice were given three train-ing trials a day for 7 consecutive days. All measurements were madeusing a live camera and EthoVision (Noldus) software. Twenty-fourhours after the last days of training, mice were given a 1 min probe test(Vorhees and Williams, 2006) in which the platform was removed,and time spent in the target quadrant was measured by the EthoVisionsoftware.
Accelerating rotarod. Mice were placed on a rotarod apparatus(Columbus Instruments) accelerating from 4 to 40 rpm in 5 min. Trialsbegan by placing the mouse on the rod and beginning rotation. Each trialended when the mouse fell off the rod, and latency was recorded. Micewere tested for four trials a day (1 min intertrial interval) for 3 consecu-tive days.
Locomotor activity. Each mouse was placed in an acrylic open-fieldchamber 40 cm long � 40 cm wide � 37 cm high (Med Associates). Thefloor of the chamber is white, and the sides were transparent. Each cham-ber was surrounded by a black drop cloth obscuring views beyond thechamber. Illumination of open field was set to 16 lux. No backgroundnoise was provided. Infrared beams recorded the animal’s location andpath (locomotor activity) and recorded distance traveled. Data were col-lected in 5 min bins during 60 min trials.
Electrophysiological and electrochemical recordingsSlice preparation. Coronal brain slices (250 �m thick) were obtainedfrom 6- to 10-week-old mice. The mice were anesthetized with isoflurane(Abbott Laboratories) and rapidly decapitated. Their brains were trans-ferred to cold, sucrose–artificial CSF (ACSF) containing the following (inmM): 200 sucrose, 25 NaHCO3, 20 glucose, 10 ascorbic acid, 2.5 KCl, 2.5CaCl2, 1 MgCl2, and 1 NaH2PO4, pH 7.4 (saturated with 95% O2 and 5%
12116 • J. Neurosci., September 30, 2009 • 29(39):12115–12124 Kheirbek et al. • AC5 in Striatal Plasticity and Learning
CO2) (Fagen et al., 2007). Striatal brain slices were made on a vibratome(VT100S; Leica). Slices were incubated for at least 1 h in a bath circulatedat 20 ml/min with normal, 32°C ACSF containing the following (in mM):125 NaCl, 25 NaHCO3, 20 glucose, 2.5 KCl, 2.5 CaCl2, 1 MgCl2, 1NaHCO3, and 1 ascorbic acid, pH 7.4 (saturated with 95% O2 and 5%CO2) (Fagen et al., 2007). During recording, slices were superfused (2ml/min) with this same ACSF at �34°C but without the ascorbic acid andwith 10 �M (�)bicuculline methiodide to block GABAA receptor-mediated synaptic responses. DNQX at 10 �M was added to the ACSF forexperiments testing intrinsic membrane excitability. Tetrodotoxin at 1�M was included in the ACSF for the recording of miniature EPSCs(mEPSCs).
Electrophysiology. Whole-cell voltage-clamp recordings from MSNslocated in the dorsal lateral striatum were obtained under visual controlon a differential interference contrast, upright microscope with infraredillumination (Axioskop; Carl Zeiss). Recordings were obtained using2.5– 6 M� resistance pipettes made from borosilicate glass capillary tub-ing (G150-4; Warner Instruments) that was pulled on a Flaming/Brownmicropipette puller (model P-97; Sutter Instruments). To test membraneexcitability and mEPSCs, recording pipettes were filled with an internalsolution containing the following (in mM): 154 K-gluconate, 1 KCl, 1EGTA, 10 HEPES, 10 glucose, 5 ATP, and 0.1 GTP, brought to pH 7.4with KOH (Mansvelder and McGehee, 2000). To test stimulated synapticcurrents, recording pipettes were filled with an internal solution contain-ing the following (in mM): 120 CsMeSO3, 10 tetraethylammonium-Cl, 10HEPES, 10 sucrose, 5 NaCl, 5 QX-314 [2(triethylamino)-N-(2,6-dimethylphenyl) acetamine], 4 ATP-Mg2, 1.1 EGTA, and 0.3 GTP-Na2,brought to pH 7.25 with CsOH (Wang et al., 2006). Currents were mea-sured using a Multiclamp 700A amplifier (2 kHz low-pass Bessel filter)with a DigiData 1322A interface (10 kHz digitization) and pClamp 9.2software (Molecular Devices). MSNs in the dorsal lateral striatum, iden-tified by their morphology and hyperpolarized resting membrane poten-tial, were voltage clamped at �80 mV, unless noted otherwise. A bipolartungsten electrode with a 500 �m tip separation (FHC) was placed in thewhite matter 200 –300 �m dorsal to the recording electrode. Test stimuli(two pulses 25 ms apart) were delivered through a Master-8 stimulator(A.M.P.I.) every 20 s. Stimulus intensity ranged from 0.1 to 1 mA, andpulse duration ranged from 50 to 400 �s. Series resistance in the recordedcells was monitored online from a 5 mV, 20 ms depolarizing pulse deliv-ered 60 ms after the second of the paired stimuli. The series resistance was60% compensated, and cells were discarded if this varied �20% or in-creased to �30 M� during the course of the experiment. After evokedEPSC amplitudes remained relatively stable for 10 min, high-frequencystimulation (HFS) was delivered in some experiments, which consistedof four 1 s, 100 Hz trains delivered every 10 s, during which the postsyn-aptic cell was depolarized to 0 mV. EPSC amplitudes are expressed as apercentage of the baseline values, mean � SEM. For prevalence data,plasticity was determined by paired t tests comparing the 10 min baselineperiod with the last 10 min of recording.
Electrochemistry. Carbon fiber microelectrodes were fabricated bythreading a 7-�m-diameter carbon fiber (Fortafil Fibers) through boro-silicate glass capillary tubing (G150-4; Warner Instruments) that wasthen pulled on a Flaming/Brown micropipette puller (model P-97; SutterInstruments). The carbon fiber was subsequently cut to restrict thelength of the exposed fiber in the slice to �50 �m. Recording electrodeswere placed in the dorsal striatum �250 �m from a bipolar tungstenelectrode with a 500 �m tip separation (FHC). Dopamine release wasevoked by electrical stimulation (400 �A, 1 ms) delivered every 2 minwith an Iso-Flex stimulus isolator triggered by a Master-8 pulse generator(A.M.P.I.). Currents were measured using a Multiclamp 700A amplifier(10 kHz low-pass Bessel filter) with a DigiData 1322A interface (50 kHzdigitization) and pClamp 9.2 software (Molecular Devices). For fast-scancyclic voltammetry (FSCV), the electrode voltage was ramped from thebaseline voltage of �400 to �1000 mV and then back again at 300 V/sversus Ag/AgCl at 100 ms intervals. Background-subtracted FSCV wasused to visualize oxidation and reduction currents, allowing for identifi-cation of the oxidized substance and for calibration with 5 �M dopamineat the end of each experiment.
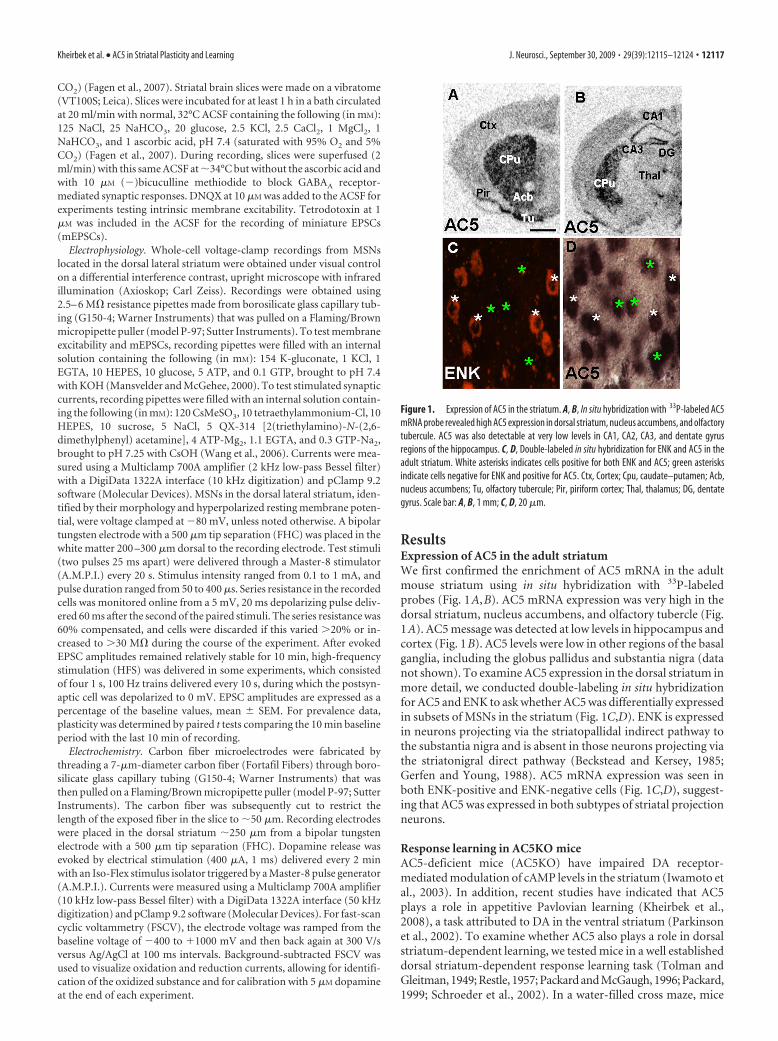
ResultsExpression of AC5 in the adult striatumWe first confirmed the enrichment of AC5 mRNA in the adultmouse striatum using in situ hybridization with 33P-labeledprobes (Fig. 1A,B). AC5 mRNA expression was very high in thedorsal striatum, nucleus accumbens, and olfactory tubercle (Fig.1A). AC5 message was detected at low levels in hippocampus andcortex (Fig. 1B). AC5 levels were low in other regions of the basalganglia, including the globus pallidus and substantia nigra (datanot shown). To examine AC5 expression in the dorsal striatum inmore detail, we conducted double-labeling in situ hybridizationfor AC5 and ENK to ask whether AC5 was differentially expressedin subsets of MSNs in the striatum (Fig. 1C,D). ENK is expressedin neurons projecting via the striatopallidal indirect pathway tothe substantia nigra and is absent in those neurons projecting viathe striatonigral direct pathway (Beckstead and Kersey, 1985;Gerfen and Young, 1988). AC5 mRNA expression was seen inboth ENK-positive and ENK-negative cells (Fig. 1C,D), suggest-ing that AC5 was expressed in both subtypes of striatal projectionneurons.
Response learning in AC5KO miceAC5-deficient mice (AC5KO) have impaired DA receptor-mediated modulation of cAMP levels in the striatum (Iwamoto etal., 2003). In addition, recent studies have indicated that AC5plays a role in appetitive Pavlovian learning (Kheirbek et al.,2008), a task attributed to DA in the ventral striatum (Parkinsonet al., 2002). To examine whether AC5 also plays a role in dorsalstriatum-dependent learning, we tested mice in a well establisheddorsal striatum-dependent response learning task (Tolman andGleitman, 1949; Restle, 1957; Packard and McGaugh, 1996; Packard,1999; Schroeder et al., 2002). In a water-filled cross maze, mice
Figure 1. Expression of AC5 in the striatum. A, B, In situ hybridization with 33P-labeled AC5mRNA probe revealed high AC5 expression in dorsal striatum, nucleus accumbens, and olfactorytubercule. AC5 was also detectable at very low levels in CA1, CA2, CA3, and dentate gyrusregions of the hippocampus. C, D, Double-labeled in situ hybridization for ENK and AC5 in theadult striatum. White asterisks indicates cells positive for both ENK and AC5; green asterisksindicate cells negative for ENK and positive for AC5. Ctx, Cortex; Cpu, caudate–putamen; Acb,nucleus accumbens; Tu, olfactory tubercule; Pir, piriform cortex; Thal, thalamus; DG, dentategyrus. Scale bar: A, B, 1 mm; C, D, 20 �m.
Kheirbek et al. • AC5 in Striatal Plasticity and Learning J. Neurosci., September 30, 2009 • 29(39):12115–12124 • 12117
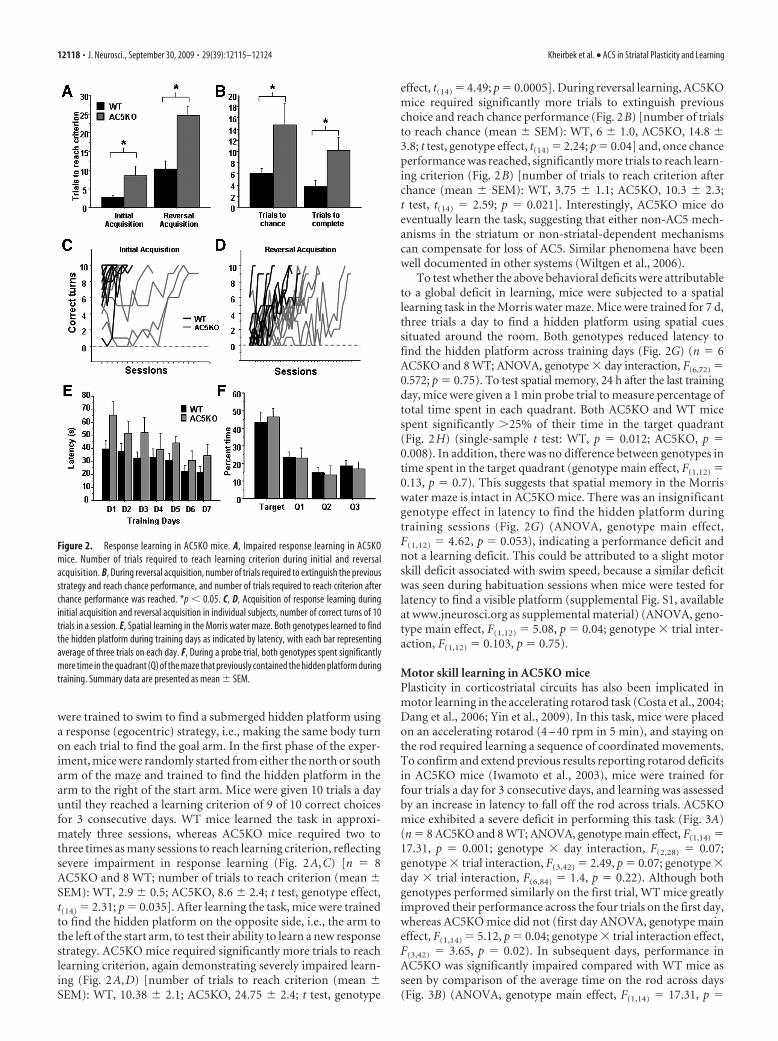
were trained to swim to find a submerged hidden platform usinga response (egocentric) strategy, i.e., making the same body turnon each trial to find the goal arm. In the first phase of the exper-iment, mice were randomly started from either the north or southarm of the maze and trained to find the hidden platform in thearm to the right of the start arm. Mice were given 10 trials a dayuntil they reached a learning criterion of 9 of 10 correct choicesfor 3 consecutive days. WT mice learned the task in approxi-mately three sessions, whereas AC5KO mice required two tothree times as many sessions to reach learning criterion, reflectingsevere impairment in response learning (Fig. 2A,C) [n 8AC5KO and 8 WT; number of trials to reach criterion (mean �SEM): WT, 2.9 � 0.5; AC5KO, 8.6 � 2.4; t test, genotype effect,t(14) 2.31; p 0.035]. After learning the task, mice were trainedto find the hidden platform on the opposite side, i.e., the arm tothe left of the start arm, to test their ability to learn a new responsestrategy. AC5KO mice required significantly more trials to reachlearning criterion, again demonstrating severely impaired learn-ing (Fig. 2A,D) [number of trials to reach criterion (mean �SEM): WT, 10.38 � 2.1; AC5KO, 24.75 � 2.4; t test, genotype
effect, t(14) 4.49; p 0.0005]. During reversal learning, AC5KOmice required significantly more trials to extinguish previouschoice and reach chance performance (Fig. 2B) [number of trialsto reach chance (mean � SEM): WT, 6 � 1.0, AC5KO, 14.8 �3.8; t test, genotype effect, t(14) 2.24; p 0.04] and, once chanceperformance was reached, significantly more trials to reach learn-ing criterion (Fig. 2B) [number of trials to reach criterion afterchance (mean � SEM): WT, 3.75 � 1.1; AC5KO, 10.3 � 2.3;t test, t(14) 2.59; p 0.021]. Interestingly, AC5KO mice doeventually learn the task, suggesting that either non-AC5 mech-anisms in the striatum or non-striatal-dependent mechanismscan compensate for loss of AC5. Similar phenomena have beenwell documented in other systems (Wiltgen et al., 2006).
To test whether the above behavioral deficits were attributableto a global deficit in learning, mice were subjected to a spatiallearning task in the Morris water maze. Mice were trained for 7 d,three trials a day to find a hidden platform using spatial cuessituated around the room. Both genotypes reduced latency tofind the hidden platform across training days (Fig. 2G) (n 6AC5KO and 8 WT; ANOVA, genotype � day interaction, F(6,72) 0.572; p 0.75). To test spatial memory, 24 h after the last trainingday, mice were given a 1 min probe trial to measure percentage oftotal time spent in each quadrant. Both AC5KO and WT micespent significantly �25% of their time in the target quadrant(Fig. 2H) (single-sample t test: WT, p 0.012; AC5KO, p 0.008). In addition, there was no difference between genotypes intime spent in the target quadrant (genotype main effect, F(1,12) 0.13, p 0.7). This suggests that spatial memory in the Morriswater maze is intact in AC5KO mice. There was an insignificantgenotype effect in latency to find the hidden platform duringtraining sessions (Fig. 2G) (ANOVA, genotype main effect,F(1,12) 4.62, p 0.053), indicating a performance deficit andnot a learning deficit. This could be attributed to a slight motorskill deficit associated with swim speed, because a similar deficitwas seen during habituation sessions when mice were tested forlatency to find a visible platform (supplemental Fig. S1, availableat www.jneurosci.org as supplemental material) (ANOVA, geno-type main effect, F(1,12) 5.08, p 0.04; genotype � trial inter-action, F(1,12) 0.103, p 0.75).
Motor skill learning in AC5KO micePlasticity in corticostriatal circuits has also been implicated inmotor learning in the accelerating rotarod task (Costa et al., 2004;Dang et al., 2006; Yin et al., 2009). In this task, mice were placedon an accelerating rotarod (4 – 40 rpm in 5 min), and staying onthe rod required learning a sequence of coordinated movements.To confirm and extend previous results reporting rotarod deficitsin AC5KO mice (Iwamoto et al., 2003), mice were trained forfour trials a day for 3 consecutive days, and learning was assessedby an increase in latency to fall off the rod across trials. AC5KOmice exhibited a severe deficit in performing this task (Fig. 3A)(n 8 AC5KO and 8 WT; ANOVA, genotype main effect, F(1,14) 17.31, p 0.001; genotype � day interaction, F(2,28) 0.07;genotype � trial interaction, F(3,42) 2.49, p 0.07; genotype �day � trial interaction, F(6,84) 1.4, p 0.22). Although bothgenotypes performed similarly on the first trial, WT mice greatlyimproved their performance across the four trials on the first day,whereas AC5KO mice did not (first day ANOVA, genotype maineffect, F(1,14) 5.12, p 0.04; genotype � trial interaction effect,F(3,42) 3.65, p 0.02). In subsequent days, performance inAC5KO was significantly impaired compared with WT mice asseen by comparison of the average time on the rod across days(Fig. 3B) (ANOVA, genotype main effect, F(1,14) 17.31, p
Figure 2. Response learning in AC5KO mice. A, Impaired response learning in AC5KOmice. Number of trials required to reach learning criterion during initial and reversalacquisition. B, During reversal acquisition, number of trials required to extinguish the previousstrategy and reach chance performance, and number of trials required to reach criterion afterchance performance was reached. *p 0.05. C, D, Acquisition of response learning duringinitial acquisition and reversal acquisition in individual subjects, number of correct turns of 10trials in a session. E, Spatial learning in the Morris water maze. Both genotypes learned to findthe hidden platform during training days as indicated by latency, with each bar representingaverage of three trials on each day. F, During a probe trial, both genotypes spent significantlymore time in the quadrant (Q) of the maze that previously contained the hidden platform duringtraining. Summary data are presented as mean � SEM.
12118 • J. Neurosci., September 30, 2009 • 29(39):12115–12124 Kheirbek et al. • AC5 in Striatal Plasticity and Learning
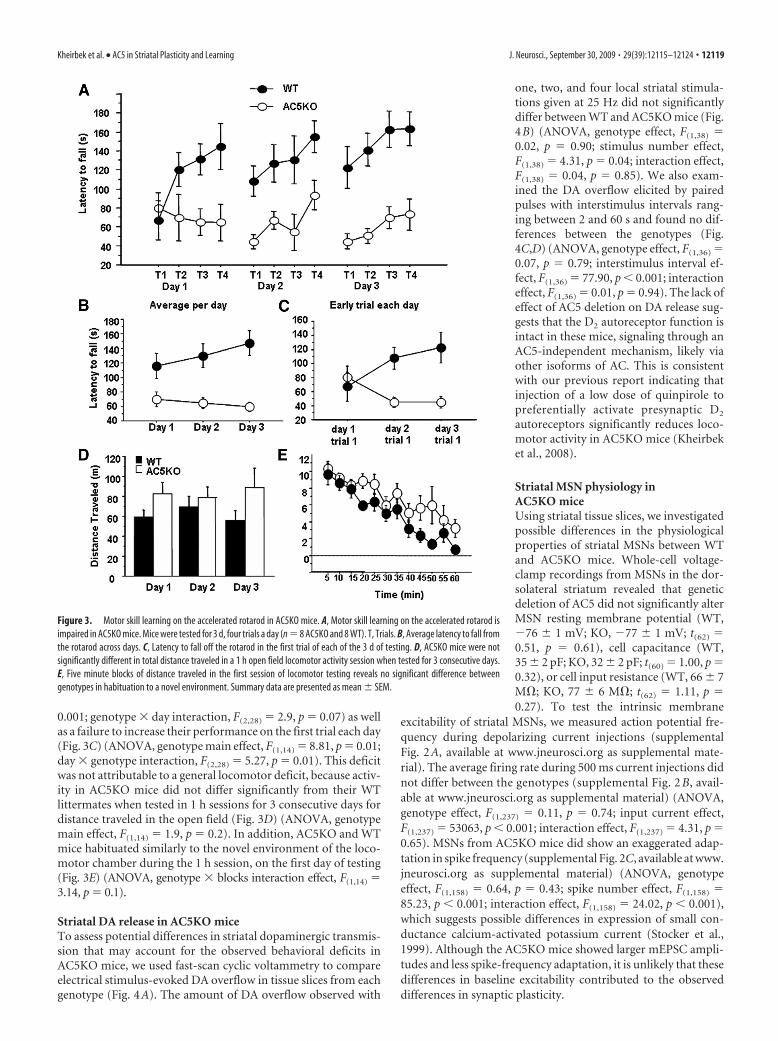
0.001; genotype � day interaction, F(2,28) 2.9, p 0.07) as wellas a failure to increase their performance on the first trial each day(Fig. 3C) (ANOVA, genotype main effect, F(1,14) 8.81, p 0.01;day � genotype interaction, F(2,28) 5.27, p 0.01). This deficitwas not attributable to a general locomotor deficit, because activ-ity in AC5KO mice did not differ significantly from their WTlittermates when tested in 1 h sessions for 3 consecutive days fordistance traveled in the open field (Fig. 3D) (ANOVA, genotypemain effect, F(1,14) 1.9, p 0.2). In addition, AC5KO and WTmice habituated similarly to the novel environment of the loco-motor chamber during the 1 h session, on the first day of testing(Fig. 3E) (ANOVA, genotype � blocks interaction effect, F(1,14) 3.14, p 0.1).
Striatal DA release in AC5KO miceTo assess potential differences in striatal dopaminergic transmis-sion that may account for the observed behavioral deficits inAC5KO mice, we used fast-scan cyclic voltammetry to compareelectrical stimulus-evoked DA overflow in tissue slices from eachgenotype (Fig. 4A). The amount of DA overflow observed with
one, two, and four local striatal stimula-tions given at 25 Hz did not significantlydiffer between WT and AC5KO mice (Fig.4B) (ANOVA, genotype effect, F(1,38) 0.02, p 0.90; stimulus number effect,F(1,38) 4.31, p 0.04; interaction effect,F(1,38) 0.04, p 0.85). We also exam-ined the DA overflow elicited by pairedpulses with interstimulus intervals rang-ing between 2 and 60 s and found no dif-ferences between the genotypes (Fig.4C,D) (ANOVA, genotype effect, F(1,36) 0.07, p 0.79; interstimulus interval ef-fect, F(1,36) 77.90, p 0.001; interactioneffect, F(1,36) 0.01, p 0.94). The lack ofeffect of AC5 deletion on DA release sug-gests that the D2 autoreceptor function isintact in these mice, signaling through anAC5-independent mechanism, likely viaother isoforms of AC. This is consistentwith our previous report indicating thatinjection of a low dose of quinpirole topreferentially activate presynaptic D2
autoreceptors significantly reduces loco-motor activity in AC5KO mice (Kheirbeket al., 2008).
Striatal MSN physiology inAC5KO miceUsing striatal tissue slices, we investigatedpossible differences in the physiologicalproperties of striatal MSNs between WTand AC5KO mice. Whole-cell voltage-clamp recordings from MSNs in the dor-solateral striatum revealed that geneticdeletion of AC5 did not significantly alterMSN resting membrane potential (WT,�76 � 1 mV; KO, �77 � 1 mV; t(62) 0.51, p 0.61), cell capacitance (WT,35 � 2 pF; KO, 32 � 2 pF; t(60) 1.00, p 0.32), or cell input resistance (WT, 66 � 7M�; KO, 77 � 6 M�; t(62) 1.11, p 0.27). To test the intrinsic membrane
excitability of striatal MSNs, we measured action potential fre-quency during depolarizing current injections (supplementalFig. 2A, available at www.jneurosci.org as supplemental mate-rial). The average firing rate during 500 ms current injections didnot differ between the genotypes (supplemental Fig. 2 B, avail-able at www.jneurosci.org as supplemental material) (ANOVA,genotype effect, F(1,237) 0.11, p 0.74; input current effect,F(1,237) 53063, p 0.001; interaction effect, F(1,237) 4.31, p 0.65). MSNs from AC5KO mice did show an exaggerated adap-tation in spike frequency (supplemental Fig. 2C, available at www.jneurosci.org as supplemental material) (ANOVA, genotypeeffect, F(1,158) 0.64, p 0.43; spike number effect, F(1,158) 85.23, p 0.001; interaction effect, F(1,158) 24.02, p 0.001),which suggests possible differences in expression of small con-ductance calcium-activated potassium current (Stocker et al.,1999). Although the AC5KO mice showed larger mEPSC ampli-tudes and less spike-frequency adaptation, it is unlikely that thesedifferences in baseline excitability contributed to the observeddifferences in synaptic plasticity.
Figure 3. Motor skill learning on the accelerated rotarod in AC5KO mice. A, Motor skill learning on the accelerated rotarod isimpaired in AC5KO mice. Mice were tested for 3 d, four trials a day (n 8 AC5KO and 8 WT). T, Trials. B, Average latency to fall fromthe rotarod across days. C, Latency to fall off the rotarod in the first trial of each of the 3 d of testing. D, AC5KO mice were notsignificantly different in total distance traveled in a 1 h open field locomotor activity session when tested for 3 consecutive days.E, Five minute blocks of distance traveled in the first session of locomotor testing reveals no significant difference betweengenotypes in habituation to a novel environment. Summary data are presented as mean � SEM.
Kheirbek et al. • AC5 in Striatal Plasticity and Learning J. Neurosci., September 30, 2009 • 29(39):12115–12124 • 12119

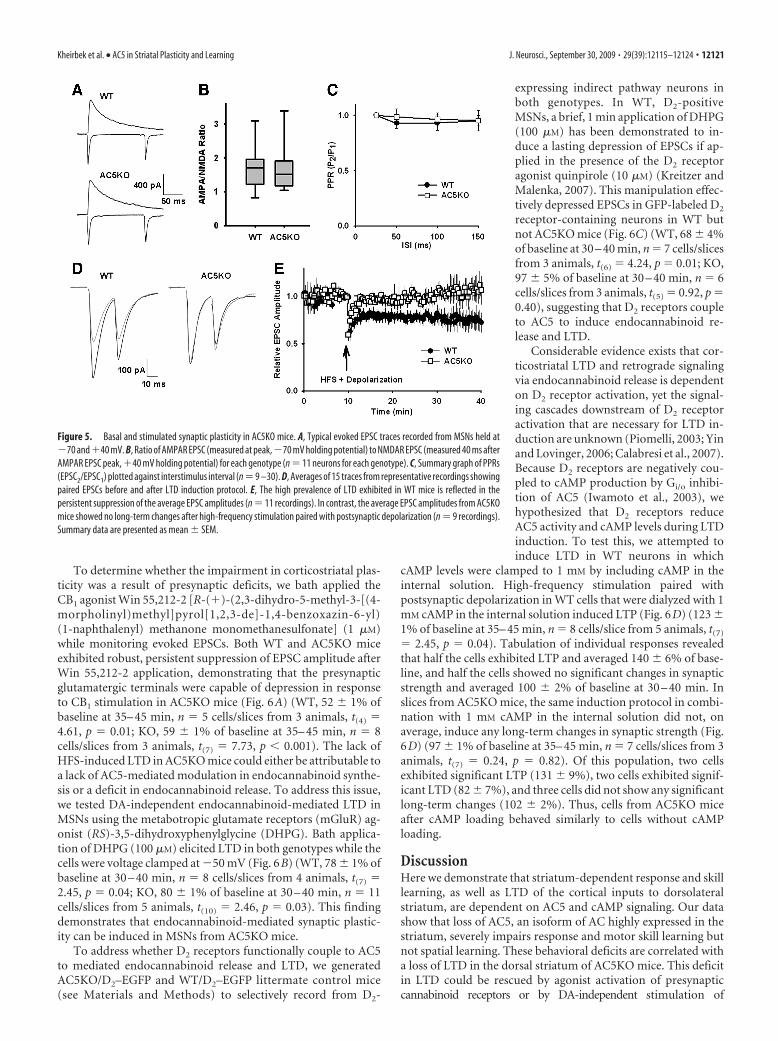
Corticostriatal plasticity inAC5KO miceTo examine the synaptic properties of theglutamatergic afferents to MSNs, we re-corded mEPSCs in MSNs from both WTand AC5KO mice (supplemental Fig. 3A,available at www.jneurosci.org as supple-mental material). There was no significantdifference in mEPSC frequency betweenthe genotypes (supplemental Fig. 3B,available at www.jneurosci.org as supple-mental material) (t(30) 1.08, p 0.29),but AC5KO mice did have larger-amplitude mEPSCs (Fig. 5C) (t(30) 2.96,p 0.006). This suggests that the AC5KOmice may have more synaptic AMPA re-ceptors (AMPARs) on MSNs than the WTmice. To assess possible differences in thestatus of synaptic plasticity and presynap-tic function, we measured the AMPAR/NMDAR ratio and paired-pulse ratio(PPR) between the two genotypes. Forthese studies, we electrically stimulatedthe white matter adjacent to the dorsolat-eral striatum to preferentially activate cor-tical afferents to MSNs (Fig. 5A). TheAMPAR/NMDAR ratio was not differentbetween the WT and AC5KO mice (Fig.5B) (t(19) 0.44, p 0.66). The fact thatAC5KO mice exhibited larger mEPSCamplitudes without any difference inAMPAR/NMDAR ratio suggests that theMSNs in the AC5KO mice had an in-creased density of both AMPA andNMDA receptors. This may be a conse-quence of homeostatic plasticity attribut-able to differences in network activity in theabsence of AC5 (Turrigiano, 2007). Consistent with the similarity inmEPSC frequency, the paired-pulse ratio did not differ betweenthe genotypes (Fig. 5C) (ANOVA, genotype effect, F(1,110) 0.45, p 0.50; interstimulus interval effect, F(3,110) 0.92, p 0.43;interaction effect, F(3,110) 0.19, p 0.91), suggesting that presyn-aptic release probability is similar between the genotypes.
The best characterized form of plasticity in the dorsal striatumis LTD of corticostriatal synapses (Calabresi et al., 2007). Thisform of plasticity is induced by pairing high-frequency stimula-tion of glutamatergic afferent fibers with postsynaptic depolar-ization (Choi and Lovinger, 1997). To assess possible differencesin corticostriatal plasticity in AC5KO mice, we compared LTDinduction in brain slices from AC5KO and WT mice. Corticalglutamatergic inputs to MSNs in the dorsolateral striatum werestimulated by an extracellular electrode placed in the adjacentwhite matter (Fig. 5D). High-frequency stimulation (four 1 s, 100Hz trains delivered every 10 s) paired with postsynaptic depolar-ization (to 0 mV) produced LTD on average in recordings fromWT mice (Fig. 5E) (73 � 8% of baseline at 30 – 40 min; n 11cells/slices from 8 animals; t(10) 2.83, p 0.02). This LTDaccompanied an increase in the paired-pulse ratio (109 � 2% ofbaseline at 30 – 40 min; t(10) 2.53, p 0.03), which is consistentwith previous observations that corticostriatal LTD is medi-ated presynaptically by a decreased release probability (Choiand Lovinger, 1997). In tissue slices from AC5KO mice, the
same HFS protocol did not induce long-term plasticity onaverage (Fig. 5E) (105 � 8% of baseline at 30 – 40 min; n 9cells/slices from 7 animals; t(8) 1.79, p 0.86). Individualrecordings showed some heterogeneity in the type and magni-tude of plasticity observed after the LTD induction protocol.In WT recordings, 7 of 11 cells showed significant LTD (t test,p 0.05) that averaged 59 � 7% of baseline at 30 – 40 min, and4 of 11 cells showed no significant change in synaptic strength(t test, p � 0.05). Such heterogeneity is in agreement with pre-vious studies (Kreitzer and Malenka, 2007). In AC5KO slices,only two cells exhibited a low-magnitude LTD (85% for both),four cells showed no significant long-term changes, and threecells exhibited low-magnitude LTP (131 � 14%). Thus, the mag-nitude and prevalence of striatal LTD were both significantlylower in AC5KO mice relative to WT. Although the AC5KO miceshowed larger mEPSC amplitudes and less spike-frequency adap-tation (supplemental Figs. 2, 3, available at www.jneurosci.org assupplemental material), it is unlikely that these differences inbaseline excitability contributed to the observed differences insynaptic plasticity. The firing properties to prolonged depo-larization would not affect the presynaptic stimulation, be-cause each stimulus was �0.4 ms, even during HFS. Whereasstronger quantal responses could affect plasticity, strongerpostsynaptic excitation and calcium entry should have en-hanced plasticity, but we saw the opposite effect in the slicesfrom AC5KO mice.
Figure 4. Dorsal striatal DA overflow in AC5KO mice. A, Sample traces of dopamine oxidation currents elicited with one, two,and four local striatal stimulations given at 25 Hz in WT and AC5KO mice (top). Subtracted voltammograms obtained from thesweeps that elicited the peak dopamine oxidation currents (bottom). B, The amount of dopamine overflow elicited with one, two,or four local electrical stimulations does not significantly differ between genotypes (n 8 WT and 6 AC5KO). C, Sample traces ofdopamine oxidation currents elicited with paired pulses using interstimulus intervals ranging between 2 and 60 s. Paired pulseswere delivered in 2 min intervals but concatenated here for display purposes. D, The relative amount of dopamine overflowobtained from a second stimulus given at varying interstimulus intervals does not differ between the genotypes (n 4 WTand 4 AC5KO).
12120 • J. Neurosci., September 30, 2009 • 29(39):12115–12124 Kheirbek et al. • AC5 in Striatal Plasticity and Learning
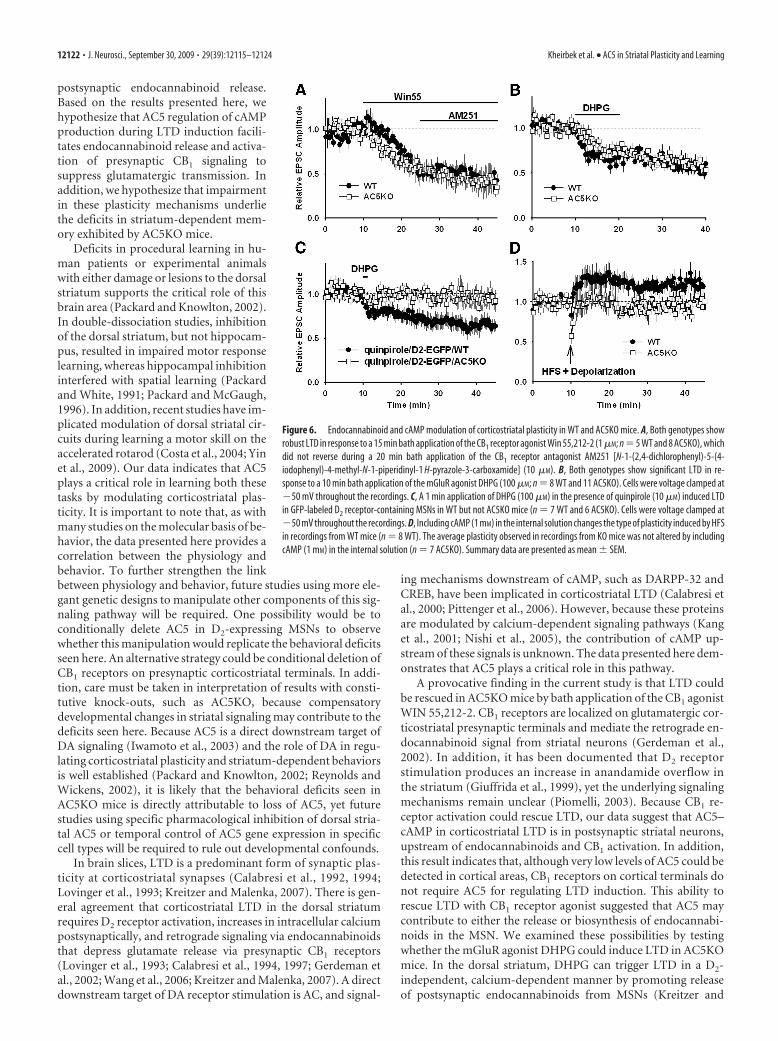
To determine whether the impairment in corticostriatal plas-ticity was a result of presynaptic deficits, we bath applied theCB1 agonist Win 55,212-2 [R-(�)-(2,3-dihydro-5-methyl-3-[(4-morpholinyl)methyl]pyrol[1,2,3-de]-1,4-benzoxazin-6-yl)(1-naphthalenyl) methanone monomethanesulfonate] (1 �M)while monitoring evoked EPSCs. Both WT and AC5KO miceexhibited robust, persistent suppression of EPSC amplitude afterWin 55,212-2 application, demonstrating that the presynapticglutamatergic terminals were capable of depression in responseto CB1 stimulation in AC5KO mice (Fig. 6A) (WT, 52 � 1% ofbaseline at 35– 45 min, n 5 cells/slices from 3 animals, t(4) 4.61, p 0.01; KO, 59 � 1% of baseline at 35– 45 min, n 8cells/slices from 3 animals, t(7) 7.73, p 0.001). The lack ofHFS-induced LTD in AC5KO mice could either be attributable toa lack of AC5-mediated modulation in endocannabinoid synthe-sis or a deficit in endocannabinoid release. To address this issue,we tested DA-independent endocannabinoid-mediated LTD inMSNs using the metabotropic glutamate receptors (mGluR) ag-onist (RS)-3,5-dihydroxyphenylglycine (DHPG). Bath applica-tion of DHPG (100 �M) elicited LTD in both genotypes while thecells were voltage clamped at �50 mV (Fig. 6B) (WT, 78 � 1% ofbaseline at 30 – 40 min, n 8 cells/slices from 4 animals, t(7) 2.45, p 0.04; KO, 80 � 1% of baseline at 30 – 40 min, n 11cells/slices from 5 animals, t(10) 2.46, p 0.03). This findingdemonstrates that endocannabinoid-mediated synaptic plastic-ity can be induced in MSNs from AC5KO mice.
To address whether D2 receptors functionally couple to AC5to mediated endocannabinoid release and LTD, we generatedAC5KO/D2–EGFP and WT/D2–EGFP littermate control mice(see Materials and Methods) to selectively record from D2-
expressing indirect pathway neurons inboth genotypes. In WT, D2-positiveMSNs, a brief, 1 min application of DHPG(100 �M) has been demonstrated to in-duce a lasting depression of EPSCs if ap-plied in the presence of the D2 receptoragonist quinpirole (10 �M) (Kreitzer andMalenka, 2007). This manipulation effec-tively depressed EPSCs in GFP-labeled D2
receptor-containing neurons in WT butnot AC5KO mice (Fig. 6C) (WT, 68 � 4%of baseline at 30 – 40 min, n 7 cells/slicesfrom 3 animals, t(6) 4.24, p 0.01; KO,97 � 5% of baseline at 30 – 40 min, n 6cells/slices from 3 animals, t(5) 0.92, p 0.40), suggesting that D2 receptors coupleto AC5 to induce endocannabinoid re-lease and LTD.
Considerable evidence exists that cor-ticostriatal LTD and retrograde signalingvia endocannabinoid release is dependenton D2 receptor activation, yet the signal-ing cascades downstream of D2 receptoractivation that are necessary for LTD in-duction are unknown (Piomelli, 2003; Yinand Lovinger, 2006; Calabresi et al., 2007).Because D2 receptors are negatively cou-pled to cAMP production by Gi/o inhibi-tion of AC5 (Iwamoto et al., 2003), wehypothesized that D2 receptors reduceAC5 activity and cAMP levels during LTDinduction. To test this, we attempted toinduce LTD in WT neurons in which
cAMP levels were clamped to 1 mM by including cAMP in theinternal solution. High-frequency stimulation paired withpostsynaptic depolarization in WT cells that were dialyzed with 1mM cAMP in the internal solution induced LTP (Fig. 6D) (123 �1% of baseline at 35– 45 min, n 8 cells/slice from 5 animals, t(7)
2.45, p 0.04). Tabulation of individual responses revealedthat half the cells exhibited LTP and averaged 140 � 6% of base-line, and half the cells showed no significant changes in synapticstrength and averaged 100 � 2% of baseline at 30 – 40 min. Inslices from AC5KO mice, the same induction protocol in combi-nation with 1 mM cAMP in the internal solution did not, onaverage, induce any long-term changes in synaptic strength (Fig.6D) (97 � 1% of baseline at 35– 45 min, n 7 cells/slices from 3animals, t(7) 0.24, p 0.82). Of this population, two cellsexhibited significant LTP (131 � 9%), two cells exhibited signif-icant LTD (82 � 7%), and three cells did not show any significantlong-term changes (102 � 2%). Thus, cells from AC5KO miceafter cAMP loading behaved similarly to cells without cAMPloading.
DiscussionHere we demonstrate that striatum-dependent response and skilllearning, as well as LTD of the cortical inputs to dorsolateralstriatum, are dependent on AC5 and cAMP signaling. Our datashow that loss of AC5, an isoform of AC highly expressed in thestriatum, severely impairs response and motor skill learning butnot spatial learning. These behavioral deficits are correlated witha loss of LTD in the dorsal striatum of AC5KO mice. This deficitin LTD could be rescued by agonist activation of presynapticcannabinoid receptors or by DA-independent stimulation of
Figure 5. Basal and stimulated synaptic plasticity in AC5KO mice. A, Typical evoked EPSC traces recorded from MSNs held at�70 and�40 mV. B, Ratio of AMPAR EPSC (measured at peak,�70 mV holding potential) to NMDAR EPSC (measured 40 ms afterAMPAR EPSC peak, �40 mV holding potential) for each genotype (n 11 neurons for each genotype). C, Summary graph of PPRs(EPSC2/EPSC1) plotted against interstimulus interval (n 9 –30). D, Averages of 15 traces from representative recordings showingpaired EPSCs before and after LTD induction protocol. E, The high prevalence of LTD exhibited in WT mice is reflected in thepersistent suppression of the average EPSC amplitudes (n 11 recordings). In contrast, the average EPSC amplitudes from AC5KOmice showed no long-term changes after high-frequency stimulation paired with postsynaptic depolarization (n 9 recordings).Summary data are presented as mean � SEM.
Kheirbek et al. • AC5 in Striatal Plasticity and Learning J. Neurosci., September 30, 2009 • 29(39):12115–12124 • 12121
postsynaptic endocannabinoid release.Based on the results presented here, wehypothesize that AC5 regulation of cAMPproduction during LTD induction facili-tates endocannabinoid release and activa-tion of presynaptic CB1 signaling tosuppress glutamatergic transmission. Inaddition, we hypothesize that impairmentin these plasticity mechanisms underliethe deficits in striatum-dependent mem-ory exhibited by AC5KO mice.
Deficits in procedural learning in hu-man patients or experimental animalswith either damage or lesions to the dorsalstriatum supports the critical role of thisbrain area (Packard and Knowlton, 2002).In double-dissociation studies, inhibitionof the dorsal striatum, but not hippocam-pus, resulted in impaired motor responselearning, whereas hippocampal inhibitioninterfered with spatial learning (Packardand White, 1991; Packard and McGaugh,1996). In addition, recent studies have im-plicated modulation of dorsal striatal cir-cuits during learning a motor skill on theaccelerated rotarod (Costa et al., 2004; Yinet al., 2009). Our data indicates that AC5plays a critical role in learning both thesetasks by modulating corticostriatal plas-ticity. It is important to note that, as withmany studies on the molecular basis of be-havior, the data presented here provides acorrelation between the physiology andbehavior. To further strengthen the linkbetween physiology and behavior, future studies using more ele-gant genetic designs to manipulate other components of this sig-naling pathway will be required. One possibility would be toconditionally delete AC5 in D2-expressing MSNs to observewhether this manipulation would replicate the behavioral deficitsseen here. An alternative strategy could be conditional deletion ofCB1 receptors on presynaptic corticostriatal terminals. In addi-tion, care must be taken in interpretation of results with consti-tutive knock-outs, such as AC5KO, because compensatorydevelopmental changes in striatal signaling may contribute to thedeficits seen here. Because AC5 is a direct downstream target ofDA signaling (Iwamoto et al., 2003) and the role of DA in regu-lating corticostriatal plasticity and striatum-dependent behaviorsis well established (Packard and Knowlton, 2002; Reynolds andWickens, 2002), it is likely that the behavioral deficits seen inAC5KO mice is directly attributable to loss of AC5, yet futurestudies using specific pharmacological inhibition of dorsal stria-tal AC5 or temporal control of AC5 gene expression in specificcell types will be required to rule out developmental confounds.
In brain slices, LTD is a predominant form of synaptic plas-ticity at corticostriatal synapses (Calabresi et al., 1992, 1994;Lovinger et al., 1993; Kreitzer and Malenka, 2007). There is gen-eral agreement that corticostriatal LTD in the dorsal striatumrequires D2 receptor activation, increases in intracellular calciumpostsynaptically, and retrograde signaling via endocannabinoidsthat depress glutamate release via presynaptic CB1 receptors(Lovinger et al., 1993; Calabresi et al., 1994, 1997; Gerdeman etal., 2002; Wang et al., 2006; Kreitzer and Malenka, 2007). A directdownstream target of DA receptor stimulation is AC, and signal-
ing mechanisms downstream of cAMP, such as DARPP-32 andCREB, have been implicated in corticostriatal LTD (Calabresi etal., 2000; Pittenger et al., 2006). However, because these proteinsare modulated by calcium-dependent signaling pathways (Kanget al., 2001; Nishi et al., 2005), the contribution of cAMP up-stream of these signals is unknown. The data presented here dem-onstrates that AC5 plays a critical role in this pathway.
A provocative finding in the current study is that LTD couldbe rescued in AC5KO mice by bath application of the CB1 agonistWIN 55,212-2. CB1 receptors are localized on glutamatergic cor-ticostriatal presynaptic terminals and mediate the retrograde en-docannabinoid signal from striatal neurons (Gerdeman et al.,2002). In addition, it has been documented that D2 receptorstimulation produces an increase in anandamide overflow inthe striatum (Giuffrida et al., 1999), yet the underlying signalingmechanisms remain unclear (Piomelli, 2003). Because CB1 re-ceptor activation could rescue LTD, our data suggest that AC5–cAMP in corticostriatal LTD is in postsynaptic striatal neurons,upstream of endocannabinoids and CB1 activation. In addition,this result indicates that, although very low levels of AC5 could bedetected in cortical areas, CB1 receptors on cortical terminals donot require AC5 for regulating LTD induction. This ability torescue LTD with CB1 receptor agonist suggested that AC5 maycontribute to either the release or biosynthesis of endocannabi-noids in the MSN. We examined these possibilities by testingwhether the mGluR agonist DHPG could induce LTD in AC5KOmice. In the dorsal striatum, DHPG can trigger LTD in a D2-independent, calcium-dependent manner by promoting releaseof postsynaptic endocannabinoids from MSNs (Kreitzer and
Figure 6. Endocannabinoid and cAMP modulation of corticostriatal plasticity in WT and AC5KO mice. A, Both genotypes showrobust LTD in response to a 15 min bath application of the CB1 receptor agonist Win 55,212-2 (1 �M; n 5 WT and 8 AC5KO), whichdid not reverse during a 20 min bath application of the CB1 receptor antagonist AM251 [N-1-(2,4-dichlorophenyl)-5-(4-iodophenyl)-4-methyl-N-1-piperidinyl-1 H-pyrazole-3-carboxamide] (10 �M). B, Both genotypes show significant LTD in re-sponse to a 10 min bath application of the mGluR agonist DHPG (100 �M; n 8 WT and 11 AC5KO). Cells were voltage clamped at�50 mV throughout the recordings. C, A 1 min application of DHPG (100 �M) in the presence of quinpirole (10 �M) induced LTDin GFP-labeled D2 receptor-containing MSNs in WT but not AC5KO mice (n 7 WT and 6 AC5KO). Cells were voltage clamped at�50 mV throughout the recordings. D, Including cAMP (1 mM) in the internal solution changes the type of plasticity induced by HFSin recordings from WT mice (n 8 WT). The average plasticity observed in recordings from KO mice was not altered by includingcAMP (1 mM) in the internal solution (n 7 AC5KO). Summary data are presented as mean � SEM.
12122 • J. Neurosci., September 30, 2009 • 29(39):12115–12124 Kheirbek et al. • AC5 in Striatal Plasticity and Learning

Malenka, 2005). AC5KO mice exhibited a DHPG–LTD compa-rable with WT mice. This suggests that the impaired HFS-induced LTD in AC5KO mice is not attributable to a deficit inendocannabinoid signaling but rather to impaired DA modula-tion of endocannabinoid release during the induction protocol.Finally, because D2 receptor stimulation promotes endocannabi-noid release in vivo (Giuffrida et al., 1999) and D2 receptor acti-vation is required for LTD induction (Calabresi et al., 1994,1997), we directly tested whether D2 receptor coupling to AC5 isrequired for endocannabinoid production and LTD. Using micewith genetically labeled D2-expressing striatopallidal neurons, weshowed that, although a short application of DHPG in the pres-ence of quinpirole could induce LTD in WT cells, it could not inAC5KO cells. This suggests that the D2 receptor directly couplesto AC5 to regulate endocannabinoid-mediated LTD.
Our data further suggest that regulation of cAMP levels byAC5 may be required for corticostriatal LTD, because corticos-triatal LTD can be effectively eliminated by elevated postsynapticcAMP levels in WT striatal neurons. Such a manipulation wouldblock D2-mediated inhibition of cAMP levels, which is similar tothat seen after genetic deletion of D2 receptors or chronic haloperi-dol treatment that also eliminate corticostriatal LTD (Centonze etal., 2004). The data suggest that D2 activation inhibits tonic AC5activity during LTD induction. The mechanism of tonic AC5activation is unknown, but one possibility is the adenosine A2a
receptors that colocalize with D2 receptors in the striatum andcouple to AC5 to increase cAMP production (Fink et al., 1992;Lee et al., 2002). Future studies examining the consequence ofbrief application of A2a antagonist on this form of LTD mayprovide insight into the mechanism of tonic AC5 activation inD2-expressing neurons.
In addition, elevated postsynaptic cAMP levels in WT cellsresulted in a net LTP of corticostriatal synapses, which was alsoseen with D2 receptor knock-out or chronic haloperidol treat-ment (Centonze et al., 2004). In our experiments, the elevation ofcAMP in the MSN may also contribute to LTP induction, becausetwo receptors that are positively coupled to cAMP signaling, A2a
and D1 receptors, contribute to LTP induction at corticostriatalsynapses (Reynolds et al., 2001; Reynolds and Wickens, 2002;Shen et al., 2008). Elevation of cAMP levels in AC5KO mice didnot, on average, alter synaptic strength, suggesting impairment inthe ability to strengthen synapses in AC5KO. This is not surpris-ing, because cAMP signaling contributes to LTP induction atthese inputs, as mentioned.
Independent of the LTP effects, our data shows that D2-mediatedchanges in cAMP are necessary for LTD at the glutamatergic inputsto striatal MSNs. A major goal for future investigations will be theidentification of downstream cAMP-responsive proteins thatcouple DA receptors and AC5 activity to the induction of plastic-ity at corticostriatal synapses. The control of endocannabinoidrelease is poorly understood, which is a major challenge in thefield, as recent studies illustrate (Adermark and Lovinger, 2007).Focusing on cAMP-dependent pathways may help identify novelmolecular mechanisms that contribute to this important media-tor of striatal plasticity.
In conclusion, the current study elucidates the critical role ofAC5, an isoform of AC highly expressed in the striatum, instriatum-dependent response learning. The data provide insightinto the molecular mechanisms underlying LTD induction in thedorsal striatum and how DA regulates this form of plasticity viaan AC5– cAMP– endocannabinoid–CB1 signaling pathway. It es-tablishes a framework for analysis of the contribution of cortico-striatal plasticity to dorsal striatum-dependent behaviors.
ReferencesAdermark L, Lovinger DM (2007) Retrograde endocannabinoid signaling at
striatal synapses requires a regulated postsynaptic release step. Proc NatlAcad Sci U S A 104:20564 –20569.
Beckstead RM, Kersey KS (1985) Immunohistochemical demonstration of dif-ferential substance P-, met-enkephalin-, and glutamic-acid-decarboxylase-containing cell body and axon distributions in the corpus striatum of the cat.J Comp Neurol 232:481–498.
Calabresi P, Maj R, Pisani A, Mercuri NB, Bernardi G (1992) Long-termsynaptic depression in the striatum: physiological and pharmacologicalcharacterization. J Neurosci 12:4224 – 4233.
Calabresi P, Pisani A, Mercuri NB, Bernardi G (1994) Post-receptor mech-anisms underlying striatal long-term depression. J Neurosci 14:4871-4881.
Calabresi P, Saiardi A, Pisani A, Baik JH, Centonze D, Mercuri NB, BernardiG, Borrelli E (1997) Abnormal synaptic plasticity in the striatum of micelacking dopamine D2 receptors. J Neurosci 17:4536 – 4544.
Calabresi P, Gubellini P, Centonze D, Picconi B, Bernardi G, Chergui K,Svenningsson P, Fienberg AA, Greengard P (2000) Dopamine andcAMP-regulated phosphoprotein 32 kDa controls both striatal long-termdepression and long-term potentiation, opposing forms of synaptic plas-ticity. J Neurosci 20:8443– 8451.
Calabresi P, Picconi B, Tozzi A, Di Filippo M (2007) Dopamine-mediatedregulation of corticostriatal synaptic plasticity. Trends Neurosci30:211–219.
Centonze D, Usiello A, Costa C, Picconi B, Erbs E, Bernardi G, Borrelli E,Calabresi P (2004) Chronic haloperidol promotes corticostriatal long-term potentiation by targeting dopamine D2L receptors. J Neurosci24:8214 – 8222.
Choi S, Lovinger DM (1997) Decreased probability of neurotransmitter re-lease underlies striatal long-term depression and postnatal developmentof corticostriatal synapses. Proc Natl Acad Sci U S A 94:2665–2670.
Cooper DM, Karpen JW, Fagan KA, Mons NE (1998) Ca 2�-sensitive ad-enylyl cyclases. Adv Second Messenger Phosphoprotein Res 32:23–51.
Costa RM, Cohen D, Nicolelis MA (2004) Differential corticostriatal plas-ticity during fast and slow motor skill learning in mice. Curr Biol14:1124 –1134.
Dang MT, Yokoi F, Yin HH, Lovinger DM, Wang Y, Li Y (2006) Disruptedmotor learning and long-term synaptic plasticity in mice lackingNMDAR1 in the striatum. Proc Natl Acad Sci U S A 103:15254 –15259.
Fagen ZM, Mitchum R, Vezina P, McGehee DS (2007) Enhanced nicotinicreceptor function and drug abuse vulnerability. J Neurosci 27:8771– 8778.
Fink JS, Weaver DR, Rivkees SA, Peterfreund RA, Pollack AE, Adler EM,Reppert SM (1992) Molecular cloning of the rat A2 adenosine receptor:selective co-expression with D2 dopamine receptors in rat striatum. BrainRes Mol Brain Res 14:186 –195.
Gerdeman GL, Ronesi J, Lovinger DM (2002) Postsynaptic endocannabi-noid release is critical to long-term depression in the striatum. Nat Neu-rosci 5:446 – 451.
Gerfen CR, Young WS 3rd (1988) Distribution of striatonigral and striato-pallidal peptidergic neurons in both patch and matrix compartments: anin situ hybridization histochemistry and fluorescent retrograde tracingstudy. Brain Res 460:161–167.
Giuffrida A, Parsons LH, Kerr TM, Rodríguez de Fonseca F, Navarro M,Piomelli D (1999) Dopamine activation of endogenous cannabinoidsignaling in dorsal striatum. Nat Neurosci 2:358 –363.
Hanoune J, Defer N (2001) Regulation and role of adenylyl cyclase iso-forms. Annu Rev Pharmacol Toxicol 41:145–174.
Iwamoto T, Okumura S, Iwatsubo K, Kawabe J, Ohtsu K, Sakai I, HashimotoY, Izumitani A, Sango K, Ajiki K, Toya Y, Umemura S, Goshima Y, Arai N,Vatner SF, Ishikawa Y (2003) Motor dysfunction in type 5 adenylylcyclase-null mice. J Biol Chem 278:16936 –16940.
Kandel ER (2001) The molecular biology of memory storage: a dialoguebetween genes and synapses. Science 294:1030 –1038.
Kang H, Sun LD, Atkins CM, Soderling TR, Wilson MA, Tonegawa S (2001)An important role of neural activity-dependent CaMKIV signaling in theconsolidation of long-term memory. Cell 106:771–783.
Kheirbek MA, Beeler JA, Ishikawa Y, Zhuang X (2008) A cAMP pathwayunderlying reward prediction in associative learning. J Neurosci 28:11401–11408.
Kheirbek et al. • AC5 in Striatal Plasticity and Learning J. Neurosci., September 30, 2009 • 29(39):12115–12124 • 12123

Kreitzer AC, Malenka RC (2005) Dopamine modulation of state-dependentendocannabinoid release and long-term depression in the striatum.J Neurosci 25:10537–10545.
Kreitzer AC, Malenka RC (2007) Endocannabinoid-mediated rescue of stri-atal LTD and motor deficits in Parkinson’s disease models. Nature445:643– 647.
Lee KW, Hong JH, Choi IY, Che Y, Lee JK, Yang SD, Song CW, Kang HS, LeeJH, Noh JS, Shin HS, Han PL (2002) Impaired D2 dopamine receptorfunction in mice lacking type 5 adenylyl cyclase. J Neurosci 22:7931–7940.
Lovinger DM, Tyler EC, Merritt A (1993) Short- and long-term synapticdepression in rat neostriatum. J Neurophysiol 70:1937–1949.
Mansvelder HD, McGehee DS (2000) Long-term potentiation of excitatoryinputs to brain reward areas by nicotine. Neuron 27:349 –357.
Matsuoka I, Suzuki Y, Defer N, Nakanishi H, Hanoune J (1997) Differentialexpression of type I, II, and V adenylyl cyclase gene in the postnataldeveloping rat brain. J Neurochem 68:498 –506.
Neve KA, Seamans JK, Trantham-Davidson H (2004) Dopamine receptorsignaling. J Recept Signal Transduct Res 24:165–205.
Nicol X, Muzerelle A, Bachy I, Ravary A, Gaspar P (2005) Spatiotemporallocalization of the calcium-stimulated adenylate cyclases, AC1 and AC8,during mouse brain development. J Comp Neurol 486:281–294.
Nishi A, Watanabe Y, Higashi H, Tanaka M, Nairn AC, Greengard P (2005)Glutamate regulation of DARPP-32 phosphorylation in neostriatal neu-rons involves activation of multiple signaling cascades. Proc Natl Acad SciU S A 102:1199 –1204.
Packard MG (1999) Glutamate infused posttraining into the hippocampusor caudate-putamen differentially strengthens place and response learn-ing. Proc Natl Acad Sci U S A 96:12881–12886.
Packard MG, Knowlton BJ (2002) Learning and memory functions of thebasal ganglia. Annu Rev Neurosci 25:563–593.
Packard MG, McGaugh JL (1996) Inactivation of hippocampus or caudatenucleus with lidocaine differentially affects expression of place and re-sponse learning. Neurobiol Learn Mem 65:65–72.
Packard MG, White NM (1991) Dissociation of hippocampus and caudatenucleus memory systems by posttraining intracerebral injection of dopa-mine agonists. Behav Neurosci 105:295–306.
Packard MG, Hirsh R, White NM (1989) Differential effects of fornix andcaudate nucleus lesions on two radial maze tasks: evidence for multiplememory systems. J Neurosci 9:1465–1472.
Parkinson JA, Dalley JW, Cardinal RN, Bamford A, Fehnert B, Lachenal G,Rudarakanchana N, Halkerston KM, Robbins TW, Everitt BJ (2002)Nucleus accumbens dopamine depletion impairs both acquisition andperformance of appetitive Pavlovian approach behaviour: implicationsfor mesoaccumbens dopamine function. Behav Brain Res 137:149 –163.
Piomelli D (2003) The molecular logic of endocannabinoid signalling. NatRev Neurosci 4:873– 884.
Pittenger C, Fasano S, Mazzocchi-Jones D, Dunnett SB, Kandel ER, BrambillaR (2006) Impaired bidirectional synaptic plasticity and proceduralmemory formation in striatum-specific cAMP response element-bindingprotein-deficient mice. J Neurosci 26:2808 –2813.
Restle F (1957) Discrimination of cues in mazes: a resolution of the place-vs.-response question. Psychol Rev 64:217–228.
Reynolds JN, Wickens JR (2002) Dopamine-dependent plasticity of corti-costriatal synapses. Neural Netw 15:507–521.
Reynolds JN, Hyland BI, Wickens JR (2001) A cellular mechanism ofreward-related learning. Nature 413:67–70.
Schroeder JP, Wingard JC, Packard MG (2002) Post-training reversible in-activation of hippocampus reveals interference between memory systems.Hippocampus 12:280 –284.
Schultz W, Dayan P, Montague PR (1997) A neural substrate of predictionand reward. Science 275:1593–1599.
Shen W, Flajolet M, Greengard P, Surmeier DJ (2008) Dichotomous dopa-minergic control of striatal synaptic plasticity. Science 321:848 – 851.
Squire LR (2004) Memory systems of the brain: a brief history and currentperspective. Neurobiol Learn Mem 82:171–177.
Stocker M, Krause M, Pedarzani P (1999) An apamin-sensitive Ca 2�-activated K � current in hippocampal pyramidal neurons. Proc Natl AcadSci U S A 96:4662– 4667.
Svenningsson P, Nishi A, Fisone G, Girault JA, Nairn AC, Greengard P(2004) DARPP-32: an integrator of neurotransmission. Annu Rev Phar-macol Toxicol 44:269 –296.
Tolman EC, Gleitman H (1949) Studies in spatial learning; place and re-sponse learning under different degrees of motivation. J Exp Psychol39:653– 659.
Turrigiano G (2007) Homeostatic signaling: the positive side of negativefeedback. Curr Opin Neurobiol 17:318 –324.
Vorhees CV, Williams MT (2006) Morris water maze: procedures for as-sessing spatial and related forms of learning and memory. Nat Protoc1:848 – 858.
Wang H, Storm DR (2003) Calmodulin-regulated adenylyl cyclases: cross-talk and plasticity in the central nervous system. Mol Pharmacol63:463– 468.
Wang Z, Kai L, Day M, Ronesi J, Yin HH, Ding J, Tkatch T, Lovinger DM,Surmeier DJ (2006) Dopaminergic control of corticostriatal long-termsynaptic depression in medium spiny neurons is mediated by cholinergicinterneurons. Neuron 50:443– 452.
Wiltgen BJ, Sanders MJ, Anagnostaras SG, Sage JR, Fanselow MS (2006)Context fear learning in the absence of the hippocampus. J Neurosci26:5484 –5491.
Wong ST, Athos J, Figueroa XA, Pineda VV, Schaefer ML, Chavkin CC,Muglia LJ, Storm DR (1999) Calcium-stimulated adenylyl cyclase activ-ity is critical for hippocampus-dependent long-term memory and latephase LTP. Neuron 23:787–798.
Wu ZL, Thomas SA, Villacres EC, Xia Z, Simmons ML, Chavkin C, PalmiterRD, Storm DR (1995) Altered behavior and long-term potentiation intype I adenylyl cyclase mutant mice. Proc Natl Acad Sci U S A92:220 –224.
Xia Z, Choi EJ, Wang F, Blazynski C, Storm DR (1993) Type I calmodulin-sensitive adenylyl cyclase is neural specific. J Neurochem 60:305–311.
Yin HH, Knowlton BJ (2006) The role of the basal ganglia in habit forma-tion. Nat Rev Neurosci 7:464 – 476.
Yin HH, Lovinger DM (2006) Frequency-specific and D2 receptor-mediated inhibition of glutamate release by retrograde endocannabinoidsignaling. Proc Natl Acad Sci U S A 103:8251– 8256.
Yin HH, Knowlton BJ, Balleine BW (2004) Lesions of dorsolateral striatumpreserve outcome expectancy but disrupt habit formation in instrumentallearning. Eur J Neurosci 19:181–189.
Yin HH, Ostlund SB, Knowlton BJ, Balleine BW (2005) The role of thedorsomedial striatum in instrumental conditioning. Eur J Neurosci22:513–523.
Yin HH, Mulcare SP, Hilario MR, Clouse E, Holloway T, Davis MI, HanssonAC, Lovinger DM, Costa RM (2009) Dynamic reorganization of striatalcircuits during the acquisition and consolidation of a skill. Nat Neurosci12:333–341.
12124 • J. Neurosci., September 30, 2009 • 29(39):12115–12124 Kheirbek et al. • AC5 in Striatal Plasticity and Learning