biochemical studies with the vinca alkaloids ii - cancer research
TRANSCRIPT

[CANCER RESEARCH 26, 882-887,May 1966]
Biochemical Studies with the Vinca Alkaloids
II. Effect of Vinblastine on the Biosynthesis of Nucleic Acidsand Their Precursors in Rat Thymus Cells1
R. G. W. JONES,- J. F. RICHARDS, AND C. T. BEER3
Cancer Research Centre, University of British Columbia, Vancouver, B.C., Canada
Summary
Vinblastine (VLB) markedly inhibited the incorporation offormate-14C and glycine-2-14C into the DNA of rat thymus cellsuspensions. In contrast, VLB either did not affect or only slightlystimulated the labeling of the hypoxanthine and thymine in the"hydrolyzed acid-soluble" fraction. The labeling of the adenine
and guanine both in this fraction and in the RNA was slightlydecreased by the alkaloid.
Incorporation studies using formate-, glycine-, or hypoxan-thine-14C indicated that VLB interfered with reactions moreparticularly related to DNA synthesis. Time studies showedthat the labeling of the DNA purines was affected before andto a greater extent than the DNA thyminp.
The acid-soluble fraction of thymus cells incubated withformate-14C in the presence of VLB contained increased amountsof radioactive serine and glycine. This increase over the controlvalues did not appear until the incubation had been in progressfor about 2 hr, whereas the effects of the alkaloid on DNA synthesis were detectable within 0.5 hr.
Introduction
It has previously been shown (9) that certain of the Vincaalkaloids, e.g., vinblastine, vincristine, and vinleurosine, inhibit the utilization of formate-14C in the biosynthesis of nucleicacids by rat thymus cell suspensions. The DNA was affectedmuch more than the RNA. Although the correlation was notexact, in general it was found that those alkaloids which arrested the growth of experimental tumors also inhibited thesynthesis of DNA in thymus cell suspensions. In addition, thymus cell suspensions prepared from the glands of rats whichhad been given small doses of vinblastine incorporated muchless formate into DNA than cells from the glands of untreatedanimals (8). These observations suggested that the inhibitionby vinblastine of the biosynthesis of the nucleic acids in isolatedthymus cell suspensions was probably quite closely related to thebiologic activity of the compound in the intact animal.
1This work was supported by grants from the Medical ResearchCouncil of Canada and the National Cancer Institute of Canada.
2 Present address: Microbiology Unit, Department of Biochemistry, University of Oxford, England.
3Medical Research Associate, Medical Research Council ofCanada.
Received for publication December 1C, 1964; revised November29, 1965.
In the biosynthetic pathways leading from the simple precursors such as formate and glycine to the polymeric DNA thereare many intermediate reactions which might be inhibited bythe alkaloid. This communication describes the effect of vinblastine (VLB) on the synthesis de novo by thymus cell suspensions of the purine and pyrimidine bases present in the nucleotidepool. This has been studied by measuring the incorporation ofradioactive formate and glycine into the bases, since thesesimple precursors are utilized as well-established points in thebiosynthetic pathways leading to the nucleic acids. The radioactive bases are present—mainly as nucleotides—in the acid-
soluble fraction of the cell suspensions. For analytical purposesthe bases have first been liberated from the nucleotide pool byhydrolysis with perchloric acid and then separated and purifiedby conventional Chromatographie procedures.
Methods
Preparation of Thymus Cell Suspensions
Suspensions of thymus cells were prepared from glands freshlyexcised from female Wistar rats (weight, 120 ±10 gm) by themethod already described (9). The suspensions usually contained 150 ±20 mg of washed cells/ml of Robinson's medium
(pH 7.4) plus 0.1 Mglucose.
Incubaticn Conditions
Incubations were made in 50-ml Erlenmeyer flasks. The controlvessels contained 1 ml of thymus cell suspension, and formate-14Cor glycine-2-14C as in the tables, in a final volume of 2.5 ml ofRobinson's medium, pH 7.4. Other flasks contained in addition
50 Mg/ml of VLB. The flasks were closed with cotton plugs andshaken at 120 strokes/min in a water bath at 37°Cfor 3 hr. Any
departures from these conditions are described in the text.
Preparation and Hydrolysis of the Acid-soluble Fraction
At the end of the incubation, the flasks were chilled in ice andthe contents, together with 1 washing of Robinson's medium (1
ml), were transferred to centrifuge tubes. The contents of thetubes were diluted with 0.5 volume of 2.1 N HC1Ü4,cooled to0°Cfor 10 min, and centrifuged. The precipitate was resus-
pended in 1.5 ml of 0.7 N HC1O4and, after 10 min, recentrifuged.The 2 supernatants were combined to give the acid-solublefraction and the washed precipitate retained for RNA and DNAanalysis.
The acid-soluble fraction was adjusted to pH 4.5-5.0 with
882 CANCER RESEARCH VOL. 26
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from
Biochemical Studies with Vinca Alkaloids. II
5 N KOH and, after standing 6 hr at 4°C,centrifuged. The super
natant was removed and the precipitate (mainly potassiumperchlorate) was washed in 5 ml of ice-cold water. The combinedsupernatant and washing was evaporated to dryness (air streamat 70°C),and the residue was hydrolyzed by heating for 60 minat 100°Cwith 2 ml of 72% HC1O4. The latter was removed as
the insoluble potassium salt, and the supernatant was adjustedto pH 4-4.5 to give the hydrolyzed acid-soluble fraction.
Separation and Purification of Furine and Pyrimidine Bases
ION-EXCHANGECHROMATOGRAPHY.The purine and pyrimidincbases in the hydrolyzed acid-soluble fraction were separatedon a column of Dowex 50-H+ ion-exchange resin (200-400 mesh)by Cohn's method (3) as modified by Wells and Winzler (12).
After the sample had been applied, the column was developedwith a linearly increasing concentration of hydrochloric acid(4 N HC1, 500 ml, slowly feeding into a mixing bottle initiallycontaining water, 500 ml). The column effluent was continuouslymonitored for compounds absorbing in the range 265-280 mju
Gilson Medical Electronics model 2651 F ultraviolet absorption
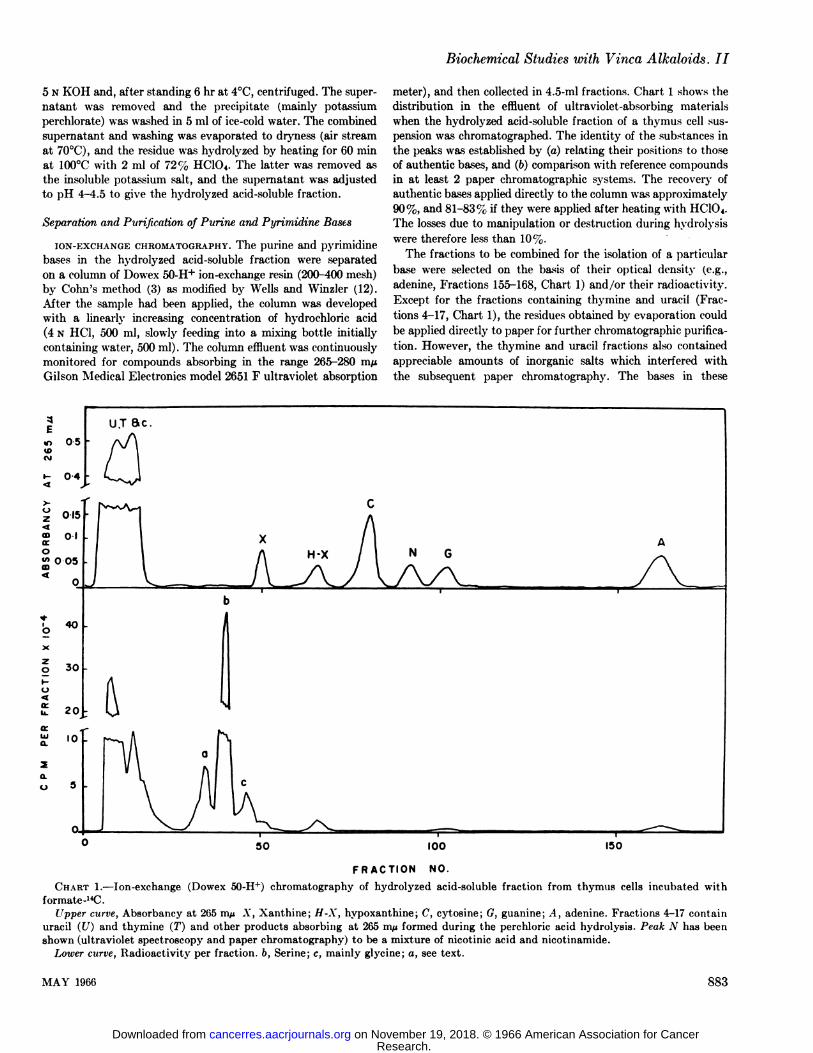
meter), and then collected in 4.5-ml fractions. Chart 1 shows thedistribution in the effluent of ultraviolet-absorbing materialswhen the hydrolyzed acid-soluble fraction of a thymus cell suspension was chromatographed. The identity of the substances inthe peaks was established by (a) relating their positions to thoseof authentic bases, and (b) comparison with reference compoundsin at least 2 paper Chromatographie systems. The recovery ofauthentic bases applied directly to the column was approximately90%, and 81-83% if they were applied after heating with HC1O<.
The losses due to manipulation or destruction during hydrolysiswere therefore less than 10%.
The fractions to be combined for the isolation of a particularbase were selected on the basis of their optical density (e.g.,adenine, Fractions 155-168, Chart 1) and/or their radioactivity.
Except for the fractions containing thymine and uracil (Fractions 4-17, Chart 1), the residues obtained by evaporation could
be applied directly to paper for further Chromatographie purification. However, the thymine and uracil fractions also containedappreciable amounts of inorganic salts which interfered withthe subsequent paper chromatography. The bases in these
100
FRACTION NO.
150
CHART 1.—Ion-exchange (Dowex 50-H+) chromatography of hydrolyzed acid-soluble fraction from thymus cells incubated withformate-14C.
Upper curve, Absorbancy at 205 m/i A", Xanthine; H-X, hypoxanthine; C, cytosine; G, guanine; .1, adenine. Fractions 4-17 contain
uracil (17) and thymine (T) and other products absorbing at 265 m/* formed during the perchloric acid hydrolysis. Peak N has beenshown (ultraviolet spectroscopy and paper chromatography) to be a mixture of nicotinic acid and nicotinamide.
Lower curve, Radioactivity per fraction, b, Serine; c, mainly glycine; a, see text.
MAY 1966 883
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from

R. G. W. Jones, J. F. Richards, and C. T. Beer
fractions were therefore absorbed on a column of charcoal(1.0 x 0.7 cm in diameter; Darco G 60) which was then washedfree from salts with 15 ml 0.1 N HC1. The absorbed bases wereeluted with 20 ml of a mixture of pyridine-water-ethanol1,10:45:45) which was then evaporated to dryness (air streamat 70°C).Final traces of pyridine were removed by adding 2 ml
of 1% ammonia and reevaporating. The residue was applieddirectly to paper for chromatography as described below.
PAPERCHKOMATOGRAPHY.The bases obtained by evaporatingthe fractions from the column were applied to Whatman No. 1paper in 0.1-0.3 ml of 0.1 N HC1. Each base was run consecutively in at least 2 systems (9). The bases, visualized by inspection in ultraviolet light, were eluted from excised portions of thechromatogram with 1 ml 0.1 N HC1, and their spectral puritywas verified in a recording spectrophotometer (Can-, Model 11).
The amount of a base recovered was calculated from the opticaldensity at the appropriate wave length, and the associatedradioactivity was then determined as described below. From thedata the specific radioactivity of the base was calculated.
Measurement of Radioactivity
Most samples, including fractions from ion-exchange columns,were assayed in a liquid scintillation counter (Packard Tri-Carb).The sample vials usually contained 0.2 ml of the aqueous radioactive solution, 1.0 ml of l M Hyamine in methanol, and 9.0ml of scintillation fluid of composition: toluene, 600 ml; ethanol,400 ml; 9:4-oz's-2-(phenyloxazolyl) benzene, 100 mg; diphenyl-oxazole, 4 gm. Appropriate corrections were made for "quenching." In some cases samples which were free from salts were
also assayed by drying aliquots on platinum planchets andcounting in a windowless gas-flow detector (Nuclear-Chicago,Model D47). All counts presented under "Results" are expressed
in relationship to standard samples counted by the gas flowmethod.
The total radioactivity due to thymine, hypoxanthine, andadenine in some of the hydrolyzed acid-soluble fractions wasdetermined by isotopie dilution. The radioactivity of the adenineand hypoxanthine could also be estimated by summing thecounts in the appropriate fractions from the ion-exchange column, from which these bases were recovered radiochemicallypure and in high yield.
TABLE 1INCORPORATIONOF FORMATE-UCINTOBASESOF
THYMUSCELL SUSPENSIONS
Results
Incorporation of Formate-14C into Acid-soluble Furine
Pijrimidine Compounds by Thymus Cell Suspensionsand
The effect of VLB (50 pg/ml) on the incorporation of for-mate-14C into the bases of the hydrolyzed acid-soluble fraction
and of the DNA and RNA is shown in Table 1. Although thealkaloid markedly reduced the specific activity of the bases inthe DNA (55-65%), it appeared to stimulate slightly the incorporation of the isotope into the thymine and hypoxanthineof the acid-soluble fraction (+23 and +16%, respectively).The alkaloid had very little effect on the labeling of the adenine,guanine, and xanthine in the acid-soluble fraction and slightlyinhibited the incorporation of formate into the adenine andguanine of the RNA, but the effect was much less than on thebases of the DNA. The total radioactivity of some of the bases
BASEISOLATEDThymineAdenineGuanineHypoxanthineXanthineADDITIONVLB"VLBVLBVLBVLBSOURCE
AND SPECIFIC ACTIVITY OPBASE(cpm//jmoleX10-")Hydrolyzed
acid-solubiefraction276339
(123)<>120116
(96.5)41.343.8
(106)194224.8
(116)139.3141.8
(102)RNA20.716.9
(81)9.78.7
(90)DNA19.28.7
(45)3.01.1
(35)2.30.95
(40.3)
" Vinblastine, 50 Mg/ml, present from start of incubation period
(3 hr).6 Figures in parentheses = (specific activity of base from VLB
incubation/specific activity of base from control incubation) X100.
in the acid-soluble fraction was determined by isotopie dilutionanalysis. The total counts of the adenine, hypoxanthine, andthymine from the VLB incubations were 98, 122, and 127%,respectively, of the control incubation values. The figuresparallel the effect of the alkaloid on the specific activities (Table1). There was, therefore, no significant accumulation of labelednucleotide precursors in the acid-soluble fraction even thoughthe alkaloid markedly inhibited the final labeling of the DXA.
Comparison of Effects of VLB on Formate, Glycine, andHypoxamhine Utilization by Thymus Cell Suspensions
Since glycine is also a precursor in the synthesis de novo ofinosinic acid and subsequently other purine nucleotides, theeffect of VLB on the incorporation of formate-14C and of glycine-2-14Cinto the purine bases was compared. The results are shown
in Table 2, in which the specific activities of the purine basesisolated from the VLB-treated suspension are expressed aspercentages of the control values. The specific activity of thehypoxanthine isolated from the acid-soluble fraction of thecells incubated with formate-14C in the presence of VLB was
substantially higher (+29%) than the control. However, VLBhad little effect on the specific activity of the hypoxanthine(+7%) when glycine-2-14C was the substrate. Table 2 shows
that this differential in the relative incorporation of the 2 precursors applied not only to hypoxanthine but to all the purinebases isolated from the acid-soluble fraction. With glycine assubstrate the specific activities of both the adenine and guaninein the VLB incubations were significantly below the controlvalues. Although this is in apparent contrast to the results whenformate was the substrate (Tables 1 and 2), relative to hypoxanthine the specific activities of the adenine, guanine, and
8S4 CANCER RESEARCH VOL. 26
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from
Biochemical Studies with Vinca Alkaloids. II
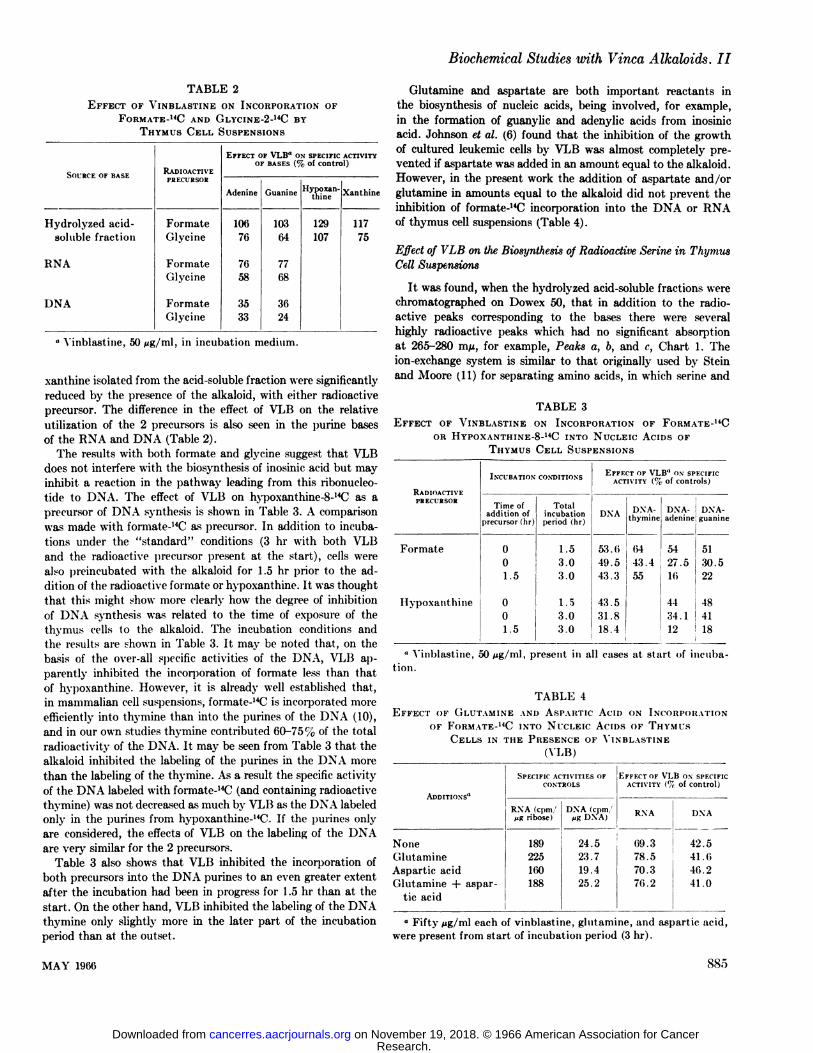
TABLE 2EFFECTOF VINBLASTINEON INCORPORATIONOF
FORMATE-14CANDGLYCINE-2-14CBYTHYMUSCELL SUSPENSIONS
SOURCEorBASEHydrolyzed
acid-solublefractionRNADNARADIOACTIVE
PRECURSORFormate
GlycineFormateGlycineFormateGlycineEFFECT
OF VLB" ox SPECIFICACTIVITYOF BASES(% ofcontrol)Adenine1067676583533Guanine103
6477683624Hypoxan-
thine129107Xanthinc117
75
0 Vinblastine, 50 pg/ml, in incubation medium.
xanthine isolated from the acid-soluble fraction were significantlyreduced by the presence of the alkaloid, with either radioactiveprecursor. The difference in the effect of VLB on the relativeutilization of the 2 precursors is also seen in the purine basesof the RNA and DNA (Table 2).
The results with both formate and glycine suggest that VLBdoes not interfere with the biosynthesis of inosinic acid but mayinhibit a reaction in the pathway leading from this ribonucleo-tide to DNA. The effect of VLB on hypoxanthine-8-14C as a
precursor of DNA synthesis is shown in Table 3. A comparisonwas made with formate-14C as precursor. In addition to incubations under the "standard" conditions (3 hr with both VLB
and the radioactive precursor present at the start), cells werealso preincubated with the alkaloid for 1.5 hr prior to the addition of the radioactive formate or hypoxanthine. It was thoughtthat this might show more clearly how the degree of inhibitionof DNA synthesis was related to the time of exposure of thethymus cells to the alkaloid. The incubation conditions andthe results are shown in Table 3. It may be noted that, on thebasis of the over-all specific activities of the DNA, VLB apparently inhibited the incorporation of formate less than thatof hypoxanthine. However, it is already well established that,in mammalian cell suspensions, formate-14C is incorporated more
efficiently into thymine than into the purines of the DNA (10),and in our own studies thymine contributed 60-75% of the total
radioactivity of the DNA. It may be seen from Table 3 that thealkaloid inhibited the labeling of the purines in the DNA morethan the labeling of the thymine. As a result the specific activityof the DNA labeled with formate-"C (and containing radioactive
thymine) was not decreased as much by VLB as the DNA labeledonly in the purines from hypoxanthine-14C. If the purines only
are considered, the effects of VLB on the labeling of the DNAare very similar for the 2 precursors.
Table 3 also shows that VLB inhibited the incorporation ofboth precursors into the DNA purines to an even greater extentafter the incubation had been in progress for 1.5 hr than at thestart. On the other hand, VLB inhibited the labeling of the DNAthymine only slightly more in the later part of the incubationperiod than at the outset.
Glutamine and aspartate are both important reactants inthe biosynthesis of nucleic acids, being involved, for example,in the formation of guanylic and adenylic acids from inosinicacid. Johnson et al. (6) found that the inhibition of the growthof cultured leukemic cells by VLB was almost completely prevented if aspartate was added in an amount equal to the alkaloid.However, in the present work the addition of aspartate and/orglutamine in amounts equal to the alkaloid did not prevent theinhibition of formate-I4C incorporation into the DNA or RNAof thymus cell suspensions (Table 4).
Effect of VLB on the Biosynthesis of Radioactive Serine in ThymusCell Suspensions
It was found, when the hydrolyzed acid-soluble fractions werechromatographed on Dowex 50, that in addition to the radioactive peaks corresponding to the bases there were severalhighly radioactive peaks which had no significant absoqitionat 265-280 m/i, for example, Peaks a, b, and c, Chart 1. Theion-exchange system is similar to that originally used by Steinand Moore (11) for separating amino acids, in which serine and
TABLE 3EFFECT OF VINBLASTINEON INCORPORATIONOF FORMATE-'*C
ORHyPOXANTHINE-8-I4CINTONUCLEICACIDSOFTHYMUSCELL SUSPENSIONS
RADIOACTIVEPRECURSORFormateHypoxanthineINCUBATION
CONDITIONSTime
ofaddition of
precursorfhr)001.5001.5Totalincubationperiod(hr)1.53.03.01.53.03.0EFFECT
OF VLB0 ON SPECIFICACTIVITY(% ofcontrols)DNA53.649.543.343.531.818.4DNA-thymine6443.455DNA-adenine5427.5164434.112DN'A-guanine5130.522484118
1Yinblastine, 50 Mg/ml> present in all cases at start of incuba
tion.
TABLE 4EFFECTOF GLUTAMINEANDASPARTICACIDON INCORPORATION
OF FORMATE-14CINTONUCLEICACIDSOF THYMUSCELLSIN THE PRESENCEOF VINBLASTINE
(VLB)
ADDITIONS"NoneGlutamineAspartic
acidGlutamine + aspar-
tic acidSPECIFIC
ACTIVITIESOFCONTKOLSRXA
(cpm/Hg ribose)189225160188DNA
(cpm/«gDNA)24.523.719.4
25.2EFFECT
OFVLB ox SPECIFICACTIVITY(% ofcontrol)RNA69.378.570.3
76.2DNA42.541.646.241.0
" Fifty /ig/ml each of vinblastine, glutamine, and aspartic acid,
were present from start of incubation period (3 hr).
MAY 1966SX",
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from
R. G. W. Jones, J. F. Richards, and C. T. Beer
glycine were eluted in positions corresponding quite closely toPeaks b and c in Chart 1. It was established by isotopie dilutionanalysis and Chromatographie comparison with authenticamino acids that over 90% of the radioactivity in Peak b wasdue to serine. It was also shown by similar chromâtographicprocedures that the radioactive serine was present in the original(i.e., unhydrolyzed) acid-soluble fraction and that only smalllosses (approximately 8%) occurred during treatment withperchloric acid. The recovery of serine after acid hydrolysis andion-exchange separation was 85%, i.e., high enough to givereliable quantitative information.
It was invariably found that there was more radioactive serinein the acid-soluble fraction of thymus cells which had beenincubated with formate-14C in the presence of VLB than in the
corresponding fraction from cells incubated without the alkaloid.The actual increase varied considerably from one experimentto another (Table 5), and in 1 case was 256% of the control.Serine may be formed by the enzymatic condensation of an"active 1-carbon" unit with glycine and it was found that whenglycine-2-14C was used as substrate VLB caused a rise in serine
radioactivity which was almost identical with that obtainedwhen formate-14C was the substrate (Table 5).
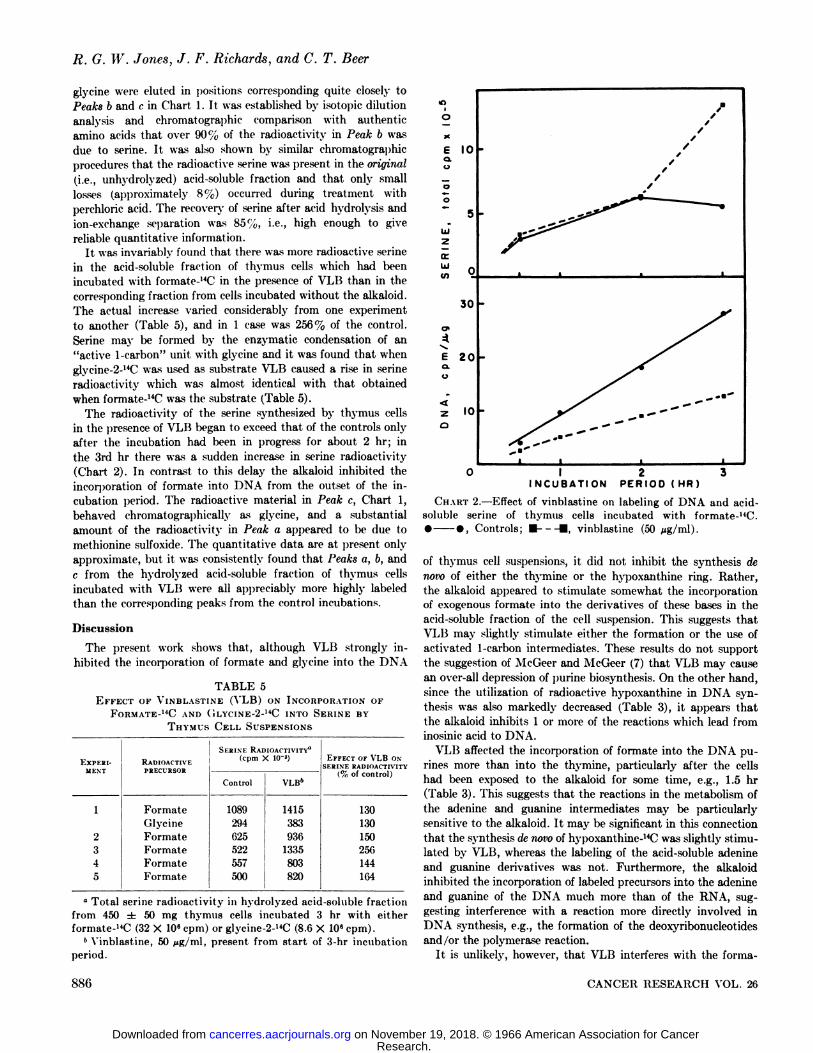
The radioactivity of the serine synthesized by thymus cellsin the presence of VLB began to exceed that of the controls onlyafter the incubation had been in progress for about 2 hr; inthe 3rd hr there was a sudden increase in serine radioactivity(Chart 2). In contrast to this delay the alkaloid inhibited theincorporation of formate into DNA from the outset of the incubation period. The radioactive material in Peak c, Chart 1,behaved chromatographically as glycine, and a substantialamount of the radioactivity in Peak a appeared to be due tomethionine sulfoxide. The quantitative data are at present onlyapproximate, but it was consistently found that Peaks a, b, andc from the hydrolyzed acid-soluble fraction of thymus cellsincubated with VLB were all appreciably more highly labeledthan the corresponding peaks from the control incubations.
Discussion
The present work shows that, although VLB strongly inhibited the incorporation of formate and glycine into the DNA
TABLE 5EFFECTOF VINBLASTINE(VLB) ON INCORPORATIONOF
FOKMATE-14CANDGLYCINE-2-14CINTOSERINEBYTHYMUSCELL SUSPENSIONS
EXPEÕI-1IENT12345RADIOACTIVEPRECURSORFormateGlycine
FormateFormateFormateFormateSERINE
RADIOACTIVITY"(cpm X10-')Control1089294
625522557500VLB614153839361335803820EFFECT
OF VLBONSERINERADIOACTIVITY(%
of control)130130
150256144164
E IOa.
30
20o,
z IOo
" Total serine radioactivity in hydrolyzed acid-soluble fraction
from 450 ± 50 mg thymus cells incubated 3 hr with eitherformate-14C (32 X 10e cpm) or glycine-2-14C (8.6 X IO6cpm).
'' Vinblastino, 50 >ig/ml, present from start of 3-hr incubation
period.
INCUBATION PERIOD (HR)
CHART 2.—Effect of vinblastine on labeling of DNA and acid-soluble serine of thymus cells incubated with formate-uC.• », Controls; • • vinblastine (50 jug/ml).
of thymus cell suspensions, it did not inhibit the synthesis denovo of either the thymine or the hypoxanthine ring. Rather,the alkaloid appeared to stimulate somewhat the incorporationof exogenous formate into the derivatives of these bases in theacid-soluble fraction of the cell suspension. This suggests thatVLB may slightly stimulate either the formation or the use ofactivated 1-carbon intermediates. These results do not supportthe suggestion of McGeer and McGeer (7) that VLB may causean over-all depression of purine biosynthesis. On the other hand,since the utilization of radioactive hypoxanthine in DNA synthesis was also markedly decreased (Table 3), it appears thatthe alkaloid inhibits 1 or more of the reactions which load frominosinic acid to DNA.
VLB affected the incorporation of formate into the DNA pu-rines more than into the thymine, particularly after the cellshad been exposed to the alkaloid for some time, e.g., 1.5 hr(Table 3). This suggests that the reactions in the metabolism ofthe adenine and guanine intermediates may be particularlysensitive to the alkaloid. It may be significant in this connectionthat the synthesis de novoof hypoxanthine-14C was slightly stimulated by VLB, whereas the labeling of the acid-soluble adenineand guanine derivatives was not. Furthermore, the alkaloidinhibited the incorporation of labeled precursors into the adenineand guanine of the DNA much more than of the RNA, suggesting interference with a reaction more directly involved inDNA synthesis, e.g., the formation of the deoxyribonucleotidesand/or the polymerase reaction.
It is unlikely, however, that VLB interferes with the forma
ssi i CANCKl! liKSKAlìCH VOL. 20
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from

Biochemical Studies with Vinca Alkaloids. II
tion of deoxyribonucleotides as o class, since the synthesis ofthe acid-soluble thymine intermediates was not inhibited (Table1). This indicates that the incorporation of formate-14C into the
DNA thymine is markedly reduced because the alkaloid inhibits 1 or more of the reactions which lie between thymidylicacid and DNA. These reactions include the kinase-catalyzedsteps leading to the formation of thymidine triphosphate andthe polymerase reaction. However, the depletion of the supplyof any of the other concurrently required deoxyriboside tri-phosphates would also inhibit the incorporation of the labeledthymine precursor.
The present work shows that VLB interferes with the biosynthesis of DNA in thymus cells at some point after the formation of the simpler pyrimidine and purine nucleotides. However, as far as is known at present, the reactions involved inthe biosynthesis of DNA are common to most mammaliantissues. The differences in the sensitivity of tissues to VLB wouldseem, therefore, to be related to aspects of the metabolism ofthe cells other than those directly concerned with DNA synthesis.
VLB causes metaphase arrest of dividing cells (2, 5). Creaseyand Markiw (4) have recently shown that VLB may interferein vivo with the synthesis of soluble RNA in the Ehrlich ascitestumor. They suggested that this might lead to disturbancesin amino acid metabolism and interfere with the synthesis ofprotein such as that needed in the mitotic apparatus. It wasfound here that in the presence of VLB there was an increase inthe total radioactivity of certain amino acids (serine, glycineand probably methionine) in the acid-soluble fraction of thymuscells incubated with formate-14C. This increase may be due
either to an enhanced synthesis of these compounds or to theirdecreased utilization as might result from a blockage in proteinsynthesis. The effect of VLB on the radioactivity of the serineappeared only after the incubation had been in progress forabout 2 hr, whereas the radioactivity of the DNA was affectedfrom the start. This suggests that the 2 effects are only indirectlyrelated.
There is insufficient information at present to explain thebiologic activities of VLB in satisfactory biochemical terms.There is evidence both here and elsewhere (I, 4, 9) that thealkaloid interferes with nucleic acid metabolism, but it is notcertain whether this is a direct effect or the result of a biochemicaldisturbance in some other part of the molecular organization of
the cell. More detailed studies are in progress in attempts toanswer this.
Acknowledgments
The authors are indebted to Dr. R. L. Noble for providingfacilities and for helpful discussion and advice, and to Dr. GordonSvoboda of the Eli Lilly Co., Indianapolis, for a gift of vinblastine.
References
1. Beer, C. T. Biochemical Studies with Vincaleukoblastine.Can. Cancer Conf., 4: 355-01, 1901.
2. Cardinali, G., Cardinali, G., and Blair, J. The Stathmo-kinetic Effect of Vincaleukoblastine on Xormal Bone Marrowand Leukemic Cells. Cancer Res., el: 1542-44, 1901.
3. Cohn, W. E. The Separation of Purine and Pyrimidine Basesand of Nucleotides by Ion Exchange. Science, 109: 377-78,1949.
4. Creasey, W. A., and Markiw, M. E. Biochemical Effects ofthe Vinca Alkaloids. I. Effects of Vinblastine on Nucleic AcidSynthesis in Mouse Tumor Cells. Biochem. Pharmacol., 13:135-42, 1904.
5. Cults, J. H. The Effect of Vincaleukoblastine on DividingCells in Vivo. Cancer Res., SI: 108-72, 1901.
0. Johnson, I. S., Wright, H. F., Svoboda, G. H., and Vlantis, J.Antitumor Principles Derived from Vinca rosea Linn. I. Vincaleukoblastine and Leurosine. Cancer Res., 20: 1010-22, 1900.
7. McGeer, P. L., and McGeer, E. G. Effect of Amethopterinand Vincaleukoblastine on Urinary 4-Amino-5-irnidazolecar-boxamide. Biochem. Pharmacol., 12: 297-98, 1903.
8. Richards, J. F., and Beer, C. T. Some Effects of Vinca Alkaloids on Nucleic Acid Metabolism. Lloydia, 27: 340-51, 1904.
9. Richards, J. F., Jones, R. G. W., and Beer, C. T. BiochemicalStudies with Vinca Alkaloids. I. Effect on Nucleic AcidFormation by Isolated Cell Suspensions. Cancer Res., 26: 876-81, 1966.
10. Smellie, R. M. S., Thomson R. Y., and Davidson, J. X . TheNucleic Acid Metabolism of Animal Cells in vitro. I. TheIncorporation of CH-Formate. Biochim. Biophys. Acta, 29:59-74, 1958.
11. Stein, W. H., and Moore, S. Chromatographie Determinationof Amino Acids of Proteins. Cold Spring Harbor Symp.Quant. Biol., 14: 179-90, 1949.
12. Wells, W., and Winzler, J. W. Metabolism of Human Leucocytes in Vitro. III. Incorporation of Formate-C14 into CellularComponents of Leukemic Human Leucocytes. Cancer Res.,19: 1080-90, 1959.
MAY 1906 887
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from

1966;26:882-887. Cancer Res R. G. W. Jones, J. F. Richards and C. T. Beer Precursors in Rat Thymus CellsVinblastine on the Biosynthesis of Nucleic Acids and Their Biochemical Studies with the Vinca Alkaloids: II. Effect of
Updated version
http://cancerres.aacrjournals.org/content/26/5/882
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/26/5/882To request permission to re-use all or part of this article, use this link
Research. on November 19, 2018. © 1966 American Association for Cancercancerres.aacrjournals.org Downloaded from