bioinformatics alignment of pairs of sequence local and global alignment methods of alignment...
Post on 18-Dec-2015
228 views
TRANSCRIPT

Bioinformatics
• Alignment of pairs of sequence
• Local and global alignment
• Methods of alignment
• Dynamic programming approach
• Use of scoring matrices and gap penalties
• PAM and BLOSUM
• Formal dynamic programming algorithm
Lecture 8

• Sequence alignment is the procedure of comparing two (pair-wise alignment) or more multiple sequences by searching for a series of individual characters or patterns that are in the same order in the sequences.
• There are two types of alignment: local and global. In global alignment, an attempt is made to align the entire sequence. If two sequences have approximately the same length and are quite similar, they are suitable for the global alignment.
• Local alignment concentrates on finding stretches of sequences with high level of matches.
Definition of sequence alignment
L G P S S K Q T G K G S - S R I W D N
Global alignment
L N - I T K S A G K G A I M R L G D A
- - - - - - - T G K G - - - - - - - -
Local alignment
- - - - - - - A G K G - - - - - - - -

• Sequence alignment is useful for discovering structural, functional and evolutionary information.
• Sequences that are very much alike may have similar secondary and 3D structure, similar function and likely a common ancestral sequence. It is extremely unlikely that such sequences obtained similarity by chance. For DNA molecules with n nucleotides such probability is very low P = 4-n. For proteins the probability even much lower P = 20 –n, where n is a number of amino acid residues
• Large scale genome studies revealed existence of horizontal transfer of genes and other sequences between species, which may cause similarity between some sequences in very distant species.
Interpretation of sequence alignment

• Dot matrix analysis
• The dynamic programming (DP) algorithm
• Word or k-tuple methods
Methods of sequence alignment

• A dot matrix analysis is a method for comparing two sequences to look for possible alignment (Gibbs and McIntyre 1970)
• One sequence (A) is listed across the top of the matrix and the other (B) is listed down the left side
• Starting from the first character in B, one moves across the page keeping in the first row and placing a dot in many column where the character in A is the same
• The process is continued until all possible comparisons between A and B are made
• Any region of similarity is revealed by a diagonal row of dots
• Isolated dots not on diagonal represent random matches
Dot matrix analysis

• Detection of matching regions can be improved by filtering out random matches and this can be achieved by using a sliding window
• It means that instead of comparing a single sequence position more positions is compared at the same time and dot is printed only if a certain minimal number of matches occur
• Dot matrix analysis can also be used to find direct and inverted repeats within the sequences
Dot matrix analysis

• Nucleic Acids Dot Plots - http://arbl.cvmbs.colostate.edu/molkit/dnadot/index.html
Dot matrix analysis: two identical sequences

• Nucleic Acids Dot Plots of genes Adh1 and G6pd in the mouse
•http://arbl.cvmbs.colostate.edu/molkit/dnadot/index.html
Dot matrix analysis: two very different sequences sequences

• Nucleic Acids Dot Plots of genes Adh1 from the mouse and rat (25 MY)
•http://arbl.cvmbs.colostate.edu/molkit/dnadot/index.html
Dot matrix analysis: two similar sequences sequences

• Nucleic Acids Dot Plots of genes Adh1 from the mouse and rat (25 MY)
•http://arbl.cvmbs.colostate.edu/molkit/dnadot/index.html
Dot matrix analysis: two similar sequences sequences; size of the sliding window increased

• The method compares every pair of characters in the two sequences and generates an alignment, which is the best or optimal.
• This is a highly computationally demanding method. However the latest algorithmic improvements and ever increasing computer capacity make possible to align a query sequence against a large DB in a few minutes.
• Each alignments has its own score and it is essential to recognise that several different alignments may have nearly identical scores, which is an indication that the dynamic programming methods may produce more than one optimal alignment. However intelligent manipulation of some parameters is important and may discriminate the alignments with similar scores.
• Global alignment program is based on Needleman-Wunsch algorithm and local alignment on Smith-Waterman. Both algorithms are derivates from the basic dynamic programming algorithm.
Dynamic programming algorithm for sequence alignment

• The alignment procedure depends upon scoring system, which can be based on probability that 1) a particular amino acid pair is found in alignments of related proteins (pxy); 2) the same amino acid pair is aligned by chance (pxpy); 3) introduction of a gap would be a better choice as it increases the score.
• The ratio of the first two probabilities is usually provided in an amino acid substitution matrix. There are many such matrices, two of them PAM and BLOSUM are considered later.
• The score for the gap introduction and its extension is also calculated from the matrices and represent a prior knowledge and some assumptions. One of them is quite simple, if negative cost of a gap is too high a reasonable alignment between slightly different sequences will be never achieved but if it is too low an optimal alignment is hardly possible. Other assumptions are based on sophisticated statistical procedures.
Description of the dynamic programming algorithm

Scoring a sequence alignment with a gap penalty
Sequence 1 V D S - C Y
Sequence 2 V E S L C Y
Score 4 2 4 -11 9 7 Score = sum of amino acid pair scores (26)
minus single gap penalty (11) = 15
As two sequences may differ, it is likely to have non-identical amino acids placed in the corresponding positions. In order to optimise the alignment gap(s) may be introduced, which may reflect losses or insertions, which occurred in the past in the sequences. Introduction of gaps causes penalties. Scores gained by each match are not always the same, for instance two rare amino acids will score more than two common.

Derivation of the dynamic programming algorithm
1. Score of new = Score of previous + Score of new
alignment alignment (A) aligned pair
V D S - C Y V D S - C Y
V E S L C Y V E S L C Y
15 = 8 + 7
2. Score of = Score of previous + Score of new
alignment (A) alignment (B) aligned pair
V D S - C V D S - C
V E S L C V E S L C
8 = -1 + 9
3. Repeat removing aligned pairs until end of alignments is reached

• Consider building this alignment in steps, starting from the initial match (V/V) and then sequentially adding a new pair until the alignment is complete, at each stage choosing a pair from all the possible matches that provides the highest score for the alignment up to that point.
• If the full alignment has the highest possible (or optimal) score, then the old alignment from which it was derived (A) by addition of the aligned Y/Y pair must also have been optimal up to that point in the alignment.
• In this manner, the alignment can be traced back to the first aligned pair that was also an optimal alignment.
• The example, which we have considered, illustrates 3 choices: 1. Match the next character(s) in the following position(s); 2. Match the next character(s) to a gap in the upper sequence; 3. Add a gap in the lower sequence.
Description of the dynamic programming algorithm

• It is critical to have reasonable scoring schemes accepted by the scientific community for DNA and proteins and for different types of alignments
• The wealth of information accumulated in the gene/protein banks was utilised with dynamic programming procedure to create such matrices for scoring matches and separately penalties for gaps introduction and extensions
• Matrices for DNA are rather similar as there are only two options purine & pyrimidine and match & mismatch
• Proteins are much more complex and the number of option is significant
• PAM and other matrices are represented in log odds scores, which is the ratio of chance of amino acid substitution due to essential biological reason to the chance of random substitution
• There are many different PAMs, which are representing different evolutionary scenarios. PAM 250 represents a level of 250% of changes expected in 2500 MY
• PAM is more suitable for studying quite distant proteins, BLOSUM is for more conserved proteins of domains
Scoring matrices

Scoring matrices: PAM (Percent Accepted Mutation)
Amino acids are grouped according to to the chemistry of the side group: (C) sulfhydryl, (STPAG)-small hydrophilic, (NDEQ) acid, acid amide and hydrophilic, (HRK) basic, (MILV) small hydrophobic, and (FYW) aromatic. Log odds values: +10 means that ancestor probability is greater, 0 means that the probability are equal, -4 means that the change is random. Thus the probability of alignment YY/YY is 10+10=20, whereas YY/TP is –3-5=-8, a rare and unexpected between homologous sequences.

Scoring matrices: BLOSUM62 (BLOcks amino acid SUbstitution Matrices)
Ideology of BLOSUM is similar but it is calculated from a very different and much larger set of proteins, which are much more similar and create blocks of proteins with a similar pattern

• This diagram indicates the moves that are possible to reach a certain position (i,j) starting from the previous row and column at position (i -1, j-1) or from any position in the same row or column
• Diagonal move with no gap penalties or move from any other position from column j or row i, with a gap penalty that depends on the size of the gap
Formal description of dynamic programming algorithm
i -x
i -1
j -1i -y j
i
Si - x,j - wx Si –1, j- 1 + s(ai , bj)
Si, j - y - wy Si, j

Formal description of dynamic programming algorithm
For two sequences a = a1, a2,..ai and b = b1, b2, ..bj, where Sij = S ( a1,…ai, b1,…bj) then
where Sij is the score at position at i in sequence a and j in sequence b, s(aibj) is score for aligning the character at positions i and j, wx is the penalty for a gap of length x in sequence a, and wx is the penalty for a gap of length y in sequence b.
Note that Sij is a type of running best score as the algorithm moves through every position in the matrix
Sij = max { Si – 1, j – 1 + s(aibj),
max (Si – x, j - wx),
x 1
max (Si j- y - wx),
y 1
}
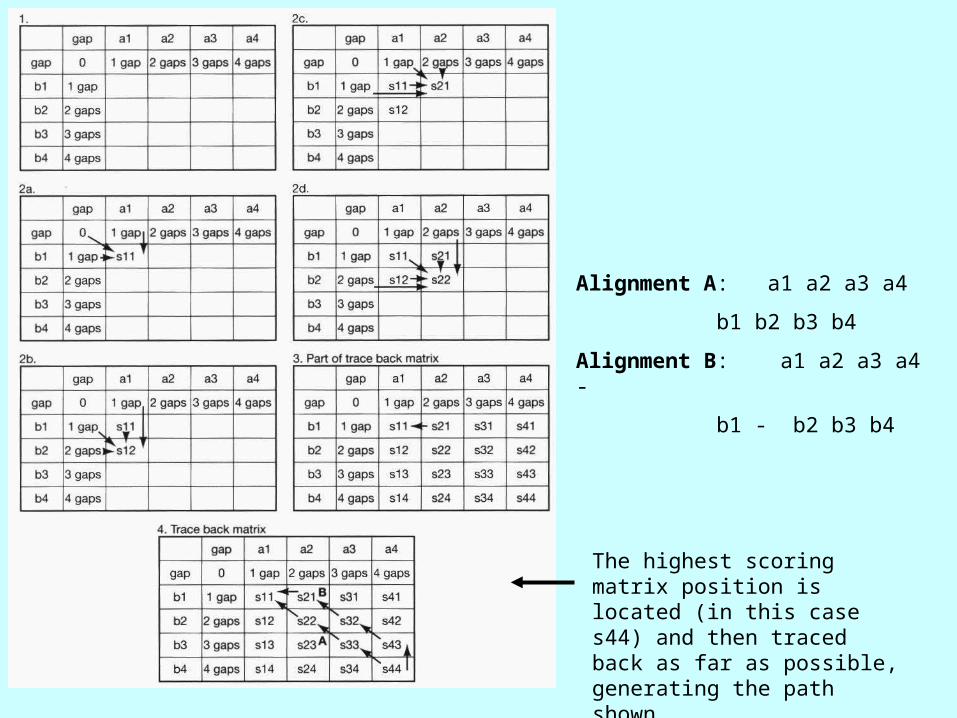
Alignment A: a1 a2 a3 a4
b1 b2 b3 b4
Alignment B: a1 a2 a3 a4 -
b1 - b2 b3 b4
The highest scoring matrix position is located (in this case s44) and then traced back as far as possible, generating the path shown