biological studies of tabanus abactor philip texas …
TRANSCRIPT

BIOLOGICAL STUDIES OF TABANUS ABACTOR PHILIP
AND OTHER TABANIDAE OF THE
TEXAS ROLLING PLAINS
by
SCOTT RUSH KINGSTON, B.A.
A THESIS
IN
ENTOMOLOGY
Submitted to the Graduate Faculty of Texas Tech University in
Partial Fulfillment of the Requirements for
the Degree of
MASTER OF SCIENCE
Approved
Accepted
August, 1983

l^^-''' ACKNOWLEDGEMENTS
I would like to thank Dr. James Wangberg, chairman of my
committee, for his direction and guidance of this research
project and critical thesis review. To Dr. Oscar Francke
and Dr. J. M. Hellman I extend my deepest appreciation for
serving on my committee, their helpful advice, and for
reviewing this thesis. To Dr. Darryl P. Sanders (former
committee chairman) whose enthusiasm and interest made this
project possible I am indebted.
I am grateful to Dr. James Goodwin for his advice on
tabanid larvae, to Dr. L. L. Pechuman for identification of
adult tabanids, to Laurie Robbins for her plant identifica
tions, to Dr. Herb Teskey for identification of tabanid lar
vae, and to Dr. Russell Wright for identification of emerged
tabanid adults.
I am also grateful to Mr. Bill Masterson of the Masterson
Ranch, Mr. Wendel Kent of the Wight Ranch, Mr. Gad Garland
of the Garland farm, and Mr. Kermit Waters of the Waters
Ranch, whose personal cooperation, understanding, and facil
ities, have made this research possible.
I owe special thanks to my summer assistant Mrs. Michele
Thomas for her invaluable aid both in the field and in the
lab.
11

I wish to extend my gratitude to Mr. Luke Celantano and
Mr. Dennis Edwards for their computer advice, to Mr. James
Cokendolpher for his technical advice and assistance, and to
Mrs. Mary Peek for her secretarial assistance.
Special thanks are due to Mrs. Ann Busenitz, Miss Halah
Nagati, and Mr. William Rogers for their assistance in the
field. I am also grateful to Miss Tarn Garland for her help
with the survey of tabanids. Thanks are also due to Miss
Lori Barnes, Mr. Steve Denning, Mr. Jay Lindley, Mr. Emmett
Schuster, and Miss Paula Verden for their day in the field.
To my parents, brother, and sisters for their love,
encouragement, understanding, and support, I wish to extend
my deepest appreciation.
This research was conducted through facilities and finan
cial resources provided by Texas Tech University. To this
institution I am indebted.
Ill

TABLE OF CONTENTS
ACKNOWLEDGEMENTS ii
ABSTRACT v
LIST OF TABLES vi
LIST OF FIGURES vii
I. INTRODUCTION AND LITERATURE REVIEW 1
Distribution 2
Biology 4
Economic Importance 9
Control 15
Objectives 19
II. METHODS AND MATERIALS 20
Study Areas 20
General Survey 25
Biological Studies of Tabanus abactor . . . 36
III. RESULTS AND DISCUSSION U6
General Survey 46
Nighttime Habitats 71
Larval Collections 81
IV. CONCLUSIONS 87
LITERATURE CITED 89
IV

ABSTRACT
The cedar fly, Tabanus abactor Philip, is a common cattle
pest in the Rolling Plains region of Texas. Research was
conducted to learn more about the biology and distributon of
J. abactor and other Texas Tabanidae. Fifteen different
species of tabanids were collected from Childress, Wichita,
Nolan, and King counties. The Cedar fly, I. abactor, was
the predominant fly species throughout this region. The
species number and diversity were found to vary with the
topography of the site. Nighttime habits and habitats of
the Cedar fly were investigated. Adult females were found
to prefer low lying areas and large vegetation types for
nighttime resting sites. Tabanid larvae (possibly T_, abac
tor) were found in stream bottoms and other areas frequented
by the adults at night. Descriptions of the larvae and pos
sible developmental sites of T. abactor are included.
v

LIST OF TABLES
TABLE
1. Number of observations, Mean, and Standard Deviation of Tabanidae collected at five sites in the Rolling Plains of Texas during the summer of 1982 48
2. Tabanidae collected in the Texas Rolling Plains from May 14 to November 21, 1982, including the frequency, number, and period of occurrence of each species collected 55
3. Average yearly temperature and cumulative rainfall of the four trapping locations in the Texas Rolling Plains 66
4. Tabanid larvae collected on the Masterson Ranch in King Co., Texas, Summer 1982 and Spring 1983, including number, size range, and site description for larvae on each collection date 82
VI

LIST OF FIGURES
1. The Cedar Fly: Tabanus abactor Philip (female) 5
2. Map of Texas illustrating the Rolling Plains Region 21
3. Research locations in the Rolling Plains of Texas 23
4. Modified Manitoba trap used in surveying tabanids of the Texas Rolling Plains during the summer of 1982 26
5. Modified Malaise trap with cyanide jar at apex as the killing mechanism used for surveying tabanids not attracted to the Manitoba traps in the Texas Rolling Plains during the summer of 1982 27
6. Type of water source and topography nearest the Manitoba trap on the Masterson Ranch 29
7. Type of water source and topography nearest the Malaise trap on the Masterson Ranch 30
8. Type of water source and topography nearest the Manitoba trap at the Burkburnett site 31
9. Type of water source and topography nearest the Manitoba trap at the Sweetwater site 32
10. Type of water source and topography nearest the Manitoba trap at the Childress site 33
11. Materials used to study the nighttime habits and habitats of Tabanus abactor at the Masterson Ranch in King Co., Tx. during the summer of 1982 38
12. Nighttime habitat study release site A (Valley) 41
13. Nighttime habitat study release site B (Hilltop) 42
vii

14.
15.
16.
Frequency and percent of Tabanidae collected at five trapping locations in the Rolling Plains region of Texas from May 14 to November 21, 1982. . .
Seasonal occurrence of tabanids collected in the Manitoba trap at Burkburnett, Texas from May 17 to November 21, 1982
Seasonal occurrence of tabanids collected in the Manitoba trap at Childress, Texas from May 27 to November 6, 1982
47
49
50
17. Seasonal occurrence of tabanids collected in the Manitoba trap on the Masterson Ranch in King Co., Texas from May 14 to November 13, 1982 51
18. Seasonal occurrence of tabanids collected in the Malaise trap on the Masterson Ranch in King Co., Texas from May 14 to November 13, 1982 52
19. Seasonal occurrence of tabanids collected in the Manitoba trap at Sweetwater, Texas from May 28 to November 21, 1982 53
20. Seasonal occurence of Chrysops pikei in the Texas Rolling Plains from May 14 to November 21, 1982 57
21 . Seasonal occurrence of Tabanus abactor in the Texas Rolling Plains from May 14 to November 21, 1982 58
22. Seasonal occurrence of Tabanus subsimilis subsimilis in the Texas Rolling Plains from May 14 to November 21, 1982 59
23. Frequency and percent of tabanid species collected at the Masterson Ranch, King Co., Texas, Summer 1982, by Manitoba trap without CO2 61
Vlll

24. Frequency and percent of tabanid species collected at Sweetwater, Texas, Summer 1982 by Manitoba trap with and without CO2 62
25. Frequency and percent of tabanid species collected at Childress, Texas, by Manitoba trap and at the Masterson Ranch by Malaise trap, Summer 1982, with and without CO2 63
26. Frequency and percent of tabanid species collected at Burkburnett, Texas, Summer 1982 by Manitoba trap with and without CO2 64
27. Frequency and percent of tabanid species collected at Burkburnett, Childress, and Sweetwater, Texas, Summer 1982, using Manitoba traps with dry ice as an added attractant 69
28. Frequency and percent of tabanid species collected at Burkburnett, Childress, and Sweetwater, Texas, Summer 1982, using Manitoba traps without dry ice 70
29. Painted T. abactor flies under U.V. light. Fluorescent orange was the preferred color 72
30. Direction of flight of marked Tabanus abactor females from the point of release at both sites used in nighttime habitat study; Site A = Valley site, Site B = hilltop 74
31. Frequency and percent of Tabanus abactor females located at night on different vegetation and substrates at both site A and B 76
ix

32. Spatial distribution (height from the ground) of Tabanus abactor females located on vegetation at night at site A and B 78
33. Geographical preference of Tabanus abactor females at night within the Texas Rolling Plains 80
34. Larvae of unidentified Tabanus spp. (possibly T. abactor) collected from both typical and atypical tabanid habitats at the Masterson Ranch, King Co., Texas, Summer 1982 and Spring 1983 83
35. Tabanus larva (probably T. s. subsimilis) collected from manure pa^ aT the Masterson Ranch, King Co., Texas, April 30, 1983. 85

CHAPTER I
INTRODUCTION AND LITERATURE REVIEW
Tabanidae: deer flies, horse flies, and green heads are
a widely diverse and important, group of insects (Curran,
1966). Nearly three hundred species of tabanids occur in
North America and the family as a whole consists of more
blood feeding species than any other dipteran family (Borror
et al., 1976). Tabanids are common pests of livestock
throughout the United States. Adult females are obligatory
blood feeding ectoparasites and are often serious pests of
livestock and humans. Tabanids are associated with large
economic losses in livestock. Damage is incurred through
loss of blood, loss of feeding time, and transmission of
diseases in these animals. The damage these Diptera cause
is enhanced even further by the additive effects of these
factors. Loss of blood, both during and after feeding, can
be severe due to the large wounds which are produced.
Tabanus abactor Philip is the most abundant horse fly
pest in the Rolling Plains region of Texas (Davis and San
ders, 1981) and also the most important tabanid infesting
Oklahoma (Schomberg and Howell, 1955). It is commonly
referred to as the cedar fly by ranchers in the Rolling

Plains due to its assumed association with cedars, properly
redberry junipers, Juniperus pinchottii Sudw. It is a
severe pest in this region, with large populations present
during most summers. Weight losses in range cattle due to
attacks from J. abactor have been estimated to be as high as
90 kg (200 lbs) per head per season during heavy fly attacks
(Davis, 1979).
In spite of its economic importance very little is known
about the biology of T_. abactor. The ovipositional and
developmental sites have not been determined. Some authors
(Davis, 1979; Schomberg and Howell, 1955), consider that
larval development is atypical for a tabanid, with develop
ment occurring in xeric habitats. Control methods have
proven ineffective against T. abactor. Information on the
biology, distribution, and habitats are needed in order to
develop new and effective control methods against this spec
ies .
Distribution
Tabanids are distributed throughout the United States,
being more prevalent along coastal areas (salt marshes) and
wet semi-tropical regions than elswhere. Distributional
studies of Tabanidae in Texas have been mostly limited to
the eastern portion of the state. Thompson (1973a, 1973b,

1974a, 1974b, and 1976) and Thompson et al. (1977) deter
mined the distribution of several tabanid species of eastern
Texas. Twenty species were reported from the Coastal
Marshes region, 29 from the Pine Belt regions, and 25 from
the Post Oak and Coastal Prairies regions of Texas. The
Rolling Plains region was not included in this study. Sev
eral variations among the species found at these geographi
cally different sites were mentioned and described. Tabanus
abactor was not present at any of these eastern Texas loca
tions. McGregor and Schomberg (1952) provided a partial
annotated list of tabanids which were collected in areas of
severe outbreaks in Texas. All species were collected in
the eastern portions of the state except T_. abactor and T.
dorsifer Walk., which were collected only in western Texas.
Fasten et al. (1968) surveyed tabanids near three towns in
western Texas (Brackettville, Marfa, and Presidio). Large
numbers of T. abactor were collected from the Brackettville
site, but none were found at either of the other two sites.
Davis and Sanders (1981) collected six species of tabanids
from 62 counties in the Rolling Plains region of Texas.
Tabanus abactor was by far the most numerous and was present
in 46 of the 62 counties surveyed. Other species, Silvius
quadrivittatus (Say) and Tabanus subsimilis (Bellardi), were
also found in large numbers throughout this region. There

are probably more species present in the Rolling Plains than
the six reported by Davis and Sanders (1981) because col
lecting was terminated once the cedar fly was captured.
Biology
Development in most tabanid species is univoltine and
aquatic. Several authors have described the morphology
and biology of tabanids (Hansens, 1979; Pechuman, 1981;
Schwardt, 1936; Stone, 1930; Tashiro and Schwardt, 1949,
1953). Teskey (1969) provides a good coverage of the bio
nomics of the larvae of North American tabanids. Very lit
tle is known about the biology of T. abactor, especially
concerning development.
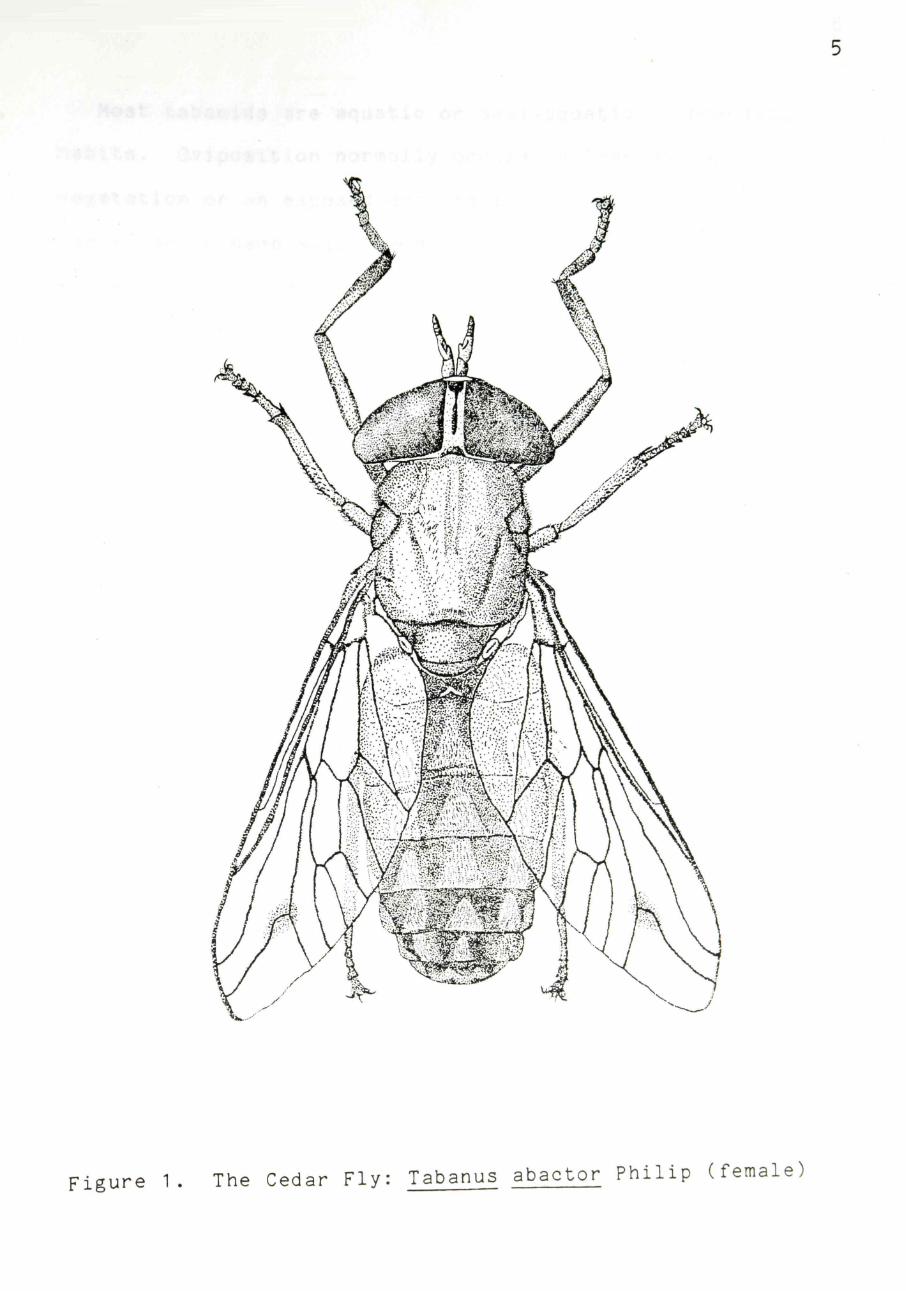
Tabanus abactor was first described from adults collected
in Eastland, Texas (Philip, 1936). Tabanus abactor is a
fairly large, robust tabanid, measuring ca. 14 mm (1/2 in)
in length. The eyes of the females are separated, as in
most tabanids, and have two green bands traversing them hor
izontally. Eyes of males are contiguous and contain only
one horizontal band. This species is easily distinguished
from other tabanids due to the three parallel rows of trian
gles along the length of the dorsum of the abdomen (one tri
plet per abdominal segment) and a faint spot at the furca
tion (branching) of wing vein R4+5 (Figure 1).
y^-.-i;-'
Figure 1. The Cedar Fly: Tabanus abactor Philip (female)

Most tabanids are aquatic or semi-aquatic in breeding
habits. Oviposition normally occurs in layered masses on
vegetation or on exposed objects in or around streams and
ponds, or in damp soil (Pechuman, 1981; Stone, 1930). Egg
masses vary in size from 100 to 800 eggs per mass. They may
be composed of one layer, as in Chrysops species or of sev
eral layers, as in most Tabanus species (Stone, 1930). The
masses are whitish in coloration when freshly deposited,
rapidly darkening to various shades of brown or black (Kahn,
1952; Pechuman, 1981). Incubation periods vary greatly in
different climates, usually requiring from five to seven
days.
The only egg masses reported of T. abactor were those
produced by caged adults (Schomberg and Howell, 1955).
These eggs, rather than being deposited on leaves or twigs
in cages, were found on the cage floor. They were greyish
in color and in loose double tiered masses. Egg masses con
tained from 40 to 175 eggs, with a mean of 93 eggs per mass.
Incubation of these eggs lasted six days.
The recognition of tabanid larvae is often difficult due
to the lack of useful keys to the immature stages. Teskey
(1969) has provided the most comprehensive and useful cover
age of tabanid larvae, covering not only species descrip-

tions but also life histories for 81 species. Schwardt
(1931) described several tabanid larvae of Arkansas. Rob
erts and Dicke (1964) described the immatures of several
species and constructed a key to the immatures of Nearctic
tabanids.
Upon eclosion larvae migrate to the substrate or water to
complete development (Pechuman, 1981). The development of
most species occurs under moist conditions. Common larval
habitats include salt marshes, drainage ditches, and along
the banks of streams or ponds. Some species develop in
moist leaf litter or rotting logs (Pechuman, 1981; Stone,
1930). Others, such as Tabanus sulcifrons (Macquart), have
been reported to develop in areas totally devoid of moisture
(Schomberg, 1952). Wilson (1969) also noted xeric habitats
for T. sulcifrons, Tabanus fusciostatus Hine, and T. lineola
Fab. Each was collected from the dry substrate of a forest
floor. According to Schomberg and Howell (1955), Tabanus
equalis Hine and T. abactor larvae can develop in soil and
under leaves or short grasses in dry shady areas where water
does not accumulate.
Most larvae are carnivorous, preying upon any number of
worms, crustaceans, or insect larvae found in the substrate.
Some species have been found to be saprophagous, feeding
upon decaying organic matter in the soil.

8
Larval development may last from several months to sev
eral years (Stone, 1930). Tabanids are known to overwinter
only in the larval stage (Pechuman, 1981). The mature lar
vae, when preparing to pupate, move to drier soil usually
just under the soil surface (James and Harwood, 1969; Stone,
1930).
Larval development of !_. abactor is poorly understood.
Schomberg and Howell (1955) reported on 12 larvae suspected
to be T. abactor in Oaklahoma and Texas. Those found in
Oklahoma were collected from dry, grass-covered soil under a
chittimwood tree, Bumelia longinosa (Michx) Pets, where
standing water would never be found. Four larvae from Texas
were found in very dry leaf-covered soil beneath a blackjack
oak, (Quercus marilandica Muench). This was also a xeric
habitat. The larvae were described only as being white with
no typical tabanid markings. They were also described as
"lazy and sluggish" in habits compared to other Tabanus
species. Larval length was 6 mm in October, and 22 mm by
pupation the following July.
The pupal stage in tabanids lasts from one to three weeks
depending on the species and temperature. According to
Schomberg and Howell (1955) T. abactor pupation occurs
beneath the substrate and lasts 10 days. Adults emerge

through a slit on the dorsum of the pupal case. The wings
are then unfolded to dry; and after a short resting period,
flight and feeding begins. Most species are univoltine, but
some such as T. subsimilis Bellardi are multivoltine, and
others such as T. calens L. require two years to develop
(Schwardt, 1936). Schomberg and Howell (1955) suggest that
egg and larval evidence indicates T. abactor to be univol
tine .
Economic Importance
Female tabanids are notorious telmophagous (pooled blood
feeding) pests of livestock and other mammals including
humans. The females have blade-like mouthparts in which the
mandibles and maxillae act as scissors, slicing open the
skin. This action usually results in the rupturing of many
small blood vessels and provides a pool of blood on which to
feed. The blood is then sponged up with the modified
labella. Males lack mandibles and do not feed on blood.
They apparently rely upon nectar and other vegetational
juices for nutrition (Pechuman, 1981). Most females are
anautogenous, requiring a blood meal for oogenesis. Some
Tabanus and Chrysops species are autogenous, the females
being able to oviposit without a blood meal. Sugars from
nectar sources and extrafloral nectaries have been shown to

10
be important constituents in the diets of males and females
for normal behavior (Bosler and Hansens, 1974; Hocking,
1953; Kniepert, 1980; Magnarelli and Anderson, 1981; Magnar-
elli et al., 1979). Some sugars may even be converted to
and deposited as yolk during early oocyte development (Knie
pert, 1980). Supplemental carbohydrate sources probably
play only a small role in oogenesis and vitellogenesis in
anautogenous species with the ingested blood providing most
of these nutrients (Magnarelli and Anderson, 1981).
Blood feeding occurs mostly during the day. Most species
feed from late afternoon to dark (Hollander and Wright,
1980a). According to Hollander and Wright (1980a) T. abac
tor usually feeds between 3 and 9 pm CDT, with feeding rap
idly decreasing one hour after sunset. Feeding by T. abac-
tor was reported by Hollander and Wright (1980b) to occur
54% more often around the legs than elsewhere on tethered
cattle. The underline and udder also were preferred feeding
sites. Tabanids requiring a blood meal for oogenesis have
been reported to seek a host every four or five days (Hol
lander and Wright, 1980b). Estimates of blood loss from
horse flies vary considerably among species and observers.
Blood meal sizes range from .08cc to .647cc in different
species (Hollander and Wright, 1980b; Tashiro and Schwardt,
1953). Total daily blood losses range from 59 to 300 cc/an-

11
imal/day (Hollander and Wright, 1980b; Philip, 1931; Tashiro
and Schwardt, 1953; Web and Wells, 1924). Tabanus abactor
was shown by Hollander and Wright (1980b) to imbibe as much
as three times its own weight in blood in a single feeding.
Hollander and Wright (1980b) determined this intake to be
.140 cc/fly/blood meal. The blood that is ingested by the
fly only accounts for part of the total blood loss of the
animal. Exsanguination may account for more blood loss than
that which is actually imbibed (Sanders, pers. comm.). Sec
ondary bacterial infections and parasitism (screwworms) may
also occur in these wounds.
Tabanus abactor also causes much irritation during feed
ing. According to Hollander and Wright (1980b), only ^5% of
T. abactor females feed to completion due to the severe pain
that they elicit during this activity. Irritation due to
the biting and buzzing of these flies also contributes to
economic losses in livestock. Annoyance from these flies
may even cause heavier losses in weight gains than those due
to blood loss. Livestock will often stop grazing and con
gregate during peak tabanid feeding periods in an effort to
avoid the feeding of these flies. Peak tabanid feeding
periods correspond with peak grazing periods causing loss of
valuable feeding time (Webb and Wells, 1924). The animals
often inflict injury upon each other by kicking or horning

12
one another in attempting to repel these flies. Secondary
infections may also result in these wounds.
Livestock production losses due to tabanid feeding and
irritation have often been reported. Steelman (1976) stated
that in 1965 over $40,000,000 was lost due to tabanid
attacks on cattle. Decreased weight gains and decreased
milk production accounted for this loss ($30,000,000 and
$10,000,000 respectively). Steelman (1976) and Roberts and
Pund (1974) reported that weight gains steadily increased as
horse fly numbers decreased. According to Davis (1979), as
much as 90 kg (200 lbs)/animal/season can be lost in range
animal weight gains because of T. abactor.
Milk production is also hindered by high infestations of
horse flies. Decreased feeding time, loss of energy due to
irritation, and blood loss contribute to substantial losses
to the dairy industry. Milk production losses have been
reported as high as 100% during three weeks of constant
exposure to feeding horse flies (Steelman, 1976). Granett
and Hansens (1956, 1957) reported that milk production would
increase after horse fly and stable fly control. Milk qual
ity is also affected by tabanids; butterfat production can
be reduced as much as 14.8% (Bruce and Decker, 1951).
Decreases in milk quantity and quality, especially in summer

13
during high tabanid activity, may also affect the newborn
calves of dairy and beef cattle. Their growth may be
stunted and their resistance to disease and stress
decreased.
Tabanids may also be important bacterial, viral, and pro
tozoan vectors (Krinskey, 1976). The likelihood of disease
transmission is increased due to the repeated biting by
tabanids.
Horse flies are- responsible for the transmission of two
major viral diseases of livestock, hog cholera virus and
equine infectious anemia virus. Hog cholera virus is the
causitive agent of swine fever in hogs: a very acute, con
tagious disease that often results in death. According to
Tidwell et al. (1972), hog cholera virus is transmitted by
Tabanus quinquevittatus (Wiedeman), T. lineola (Fabricius),
and possibly three other species. Equine infectious anemia
virus is transmitted physically by the bites of horse flies
and other blood sucking Diptera (Hawkins et al., 1976; Issel
and Coggins, 1979; and Kemen, 1977). The disease is often
referred to as swamp fever due to its high prevalence around
swampy areas where tabanids may develop. It is a serious
chronic disease of horses and other Equidae, causing debili
tation and death in severe cases. Horse flies are particu-

14
larly a problem in eradication efforts because carrier
horses, those which elicit no visible symptoms but remain
viremic, spread the virus to healthy animals.
Bacterial diseases transmitted by tabanids are also a
major cause of loss to the livestock industry. Bacillus
anthracic (Cahn), the causitive agent of anthrax, is a
highly fatal disease of cattle and is transmitted by several
species of tabanids (Krinskey, 1976). Anthrax can be dis
seminated in a number of different ways other than by biting
flies, and the role that horse flies play in transmission is
not fully understood. Anaplasmosis, a disease caused by a
protozoan-like organism, Anaplasma marginale (Theiler), has
also been shown to be transmitted by tabanids. Anaplasmosis
is a disease which causes a severe anemic condition in cat
tle. Mortality rates have been recorded to be as high as
50% in severe outbreaks. Those animals that survive become
permanent carriers of the organism, thereby perpetuating the
disease. According to Steelman (1976) economic losses
between the years 1951 and I960 totaled $9,004,000 due to
mortality and $26,997,000 due to morbidity. Transmission is
almost totally arthropod dependent. Tabanus abactor, along
with several other tabanids, has been shown to transmit ana
plasmosis (Dikmans, 1950; Sanborn et al., 1932; Wilson et
al., 1963; Wilson and Myer, 1966).

15
Disease agents transmitted by Tabanidae are most often
carried physically from one host to another on the fly's
mouthparts. Some biological transmission occurs in certain
Chrysops species. In addition some deer flies also act as
intermediate hosts of helminths that can later cause disease
in livestock (Krinskey, 1976). Chrysops species provide the
only known means of transmission of the eye worm, Loa lea
(Guyot) in western Africa. The arterial worm Eloephora
schneideri Wehr and Dikmans is an important helminth attack
ing sheep and mule deer in southwestern portions of the
United States and has also been shown to be transmitted
biologically by tabanids. Tabanus abactor has not been
shown to be a biological carrier of any disease agent; how
ever, inadequate research has been conducted on its role as
a disease vector.
Control
Control of tabanids is very difficult due to the hardi
ness of the adults and the general lack of biological infor
mation on them. The adult female is somewhat resistant to »
insecticides and spends little time actually on the host,
thus decreasing the effective time period for chemical expo
sure (Brown and Lancaster, 1973). Varied larval habitats
and developmental time periods also complicate control
efforts.

16
Area broadcast of insecticides for adult and larval con
trol has met with little success. Historically, attempts at
using aerial applications with Lindane and DDT provided lim
ited control (Lindquist and Knipling, 1957). Control was
achieved only in open areas and little residual control was
obtained with DDT. Control of larvae in saltmarshes using
5% Dieldrin applied aerially was reported by Hansens (1956).
Larvae were eliminated from the marshes for two years post-
treatment; however, serious injury to other animal life in
the marsh accompanied this control. Synthetic pyrethroids
with short residual properties seem to be the most promising
types of insecticides for area wide control. According to
Hansens (1981) Resmethrin and Permethrin applied in ULV for
mulations are effective for controlling adult populations of
the deer fly Chrysops atlanticus Pechuman. However, control
is often only short term, sometimes just one or two days.
Practical and effective repellents for protection of
range animals against biting horse flies have not yet been
developed. Unfortunately, Stirofos-impregnated ear tags,
which are so useful in the control of horn flies and other
muscids, have no effect against •tabanids (Wilson et al.,
1978). Some protection can be obtained for dairy and feed-
lot animals by the use of mosquito repellents (Blume et al.,
1971). Repellents are impractical for use on range animals

17
because of their expense and the necessity of repeated
treatments.
Insecticides applied directly to livestock are not
totally effective either. Many chemicals such as Malathion
and Diazinon are effective for short durations against these
flies but provide little or no long term relief. Synthetic
pyrethroids, such as Permethrin, provide a longer residual
affect and can reduce fly populations for as long as two and
a half weeks (Bay et al., 1976).
Some non-chemical tactics have been attempted for tabanid
control. The Manitoba trap has reduced deer fly populations
in certain areas (Pechuman, 1981). According to Hansens
(1979) the use of such traps may decrease populations to a
tolerable level but elimination of the problem is not
achieved. Temporary impoundment of waterways, such as salt
marshes and streams, have been shown by Anderson and Kneen
(1969) to be an effective measure for eliminating deer fly
immature stages. This method is useful in certain habitats
against some species but it is too expensive and time con
suming when used on a large scale.
There are several natural enemies of tabanids some of
which may be useful in biological control (Pechuman, 1981).
Adults are preyed upon by dragonflies (Odonata), robber

18
flies (Asilidae), and wasps (Vespidae) of various genera
(Pechuman, 1981). Some sarcophagid larvae, Machronychia
spp. (near aurata, Coquillett) have been found to parasitize
tabanid adults (Thompson and Love, 1979). Thompson (1978)
and Thompson and Love (1979) suggest that the prospects of
rearing this species as a biocontrol agent of horse flies
are encouraging. Larvae and pupae are parasitized by both
dipterous and hymenopterous parasites, but these parasites
have little overall impact on.adult populations (Thompson et
al., 1980; Pechuman, 1981). Egg parasitism is common in
tabanids and often the percentage of parasitized eggs is
very high. Egg parasitism in Texas Tabanidae may be as high
as 97% (Parman, 1928). Trichogrammatidae and Scelionidae
(Hymenoptera) are found commonly parasitising tabanid eggs
(Pechuman, 1981). The potential for control of tabanids
using these insects exists; however, little research has been
conducted to determine the overall impact on populations and
feasibility of such a project in the field. More informa
tion on the oviposition sites, larval developmental sites,
and behavior of T. abactor is needed. Such knowledge may
aid in the development of a practical control strategy.

19
Objectives
The four objectives of this study were: (1) to survey
the tabanids of the Rolling Plains region of Texas, (2) to
determine developmental site(s) for T. abactor and other
tabanid species, (3) to investigate the nocturnal habits and
habitats of T_, abactor, and (4) to determine the location
and duration of larval development in T. abactor.
Davis and Sanders (1981) conducted a partial survey in
the Rolling Plains but a comprehensive survey has not been
completed. My survey was initiated in an effort to deter
mine what other species are present in addition to T. abac
tor within this area and to note the economic impact these
flies may have on the area cattle industry. Information on
interspecific competition between tabanids present was also
sought during this study. The nighttime habits and habitats
of T. abactor were studied to determine oviposition and
other behavior patterns that may have significance for con
trol. Descriptions of larvae and life cycle of T. abactor
given by Schomberg and Howell (1955) are inconclusive. Ade
quate descriptions of the larvae and life cycle of T. abac
tor are necessary if control methods aimed at the larval
stage are to be initiated.

CHAPTER II
METHODS AND MATERIALS
Study Areas
The Rolling Plains of Texas, as described by Correl and
Johnston (1979), is an area of approximately 9,600,000 hec
tares (24,000,000 acres) of gently rolling to moderately
rough plains topography, (Figure 2). It is dissected by
numerous intermittent stream valleys flowing easterly to
southeasterly. Approximately 66% of this area is primarily
used for the grazing of cattle. The elevation ranges from
240 to 900 meters, with the soils varying from coarse sands
along streams to tight clays or red-bed clays and shales.
The annual rainfall ranges from about 56 cm. in the west to
almost 76 cm. in the east. May and September are normally
the high rainfall months, with a summer dry period and high
evaporation rates typical during most seasons. The vegeta
tion is dominated by range grasses, such as tabosa (Hilaria
mutica, Buckl.) and others, with honey mesquite, (Prosopis
glandulosa Torrey), junipers (Juniperus spp.) and other
tree species. Weed species such as broomweed (Gutierrezia
spp.) and sandburs (Cenchrus spp.) are also common invaders
due to overgrazing.
20
21
^^•^^^^^r rr;:^^-rP5isr:
Figure 2. Map of Texas illustrating the Rolling Plains Region (lighter portion).

22
Research began in the summer of 1982 and was conducted on
four ranches in four Rolling Plains counties: King, Chil
dress, Wichita, and Nolan, Texas (Figure 3). All four coun
ties and ranches were included in the survey. Biological
investigations of T. abactor were limited to the Masterson
Ranch in King Co., Tx.
Trapping Locations
The central trapping site was located on the Masterson
Ranch ca. 24 km (15 mi) northeast of Guthrie, Tx. It occu
pies an area of over 12,000 hectares (30,000 acres) with the
dominant vegetation including range grasses, redberry juni
pers (Juniperus pinchottii Sudw.), and some honey mesquite
( . glandulosa) . The overall topography is very rugged to
moderately hilly, with numerous intermittent stream valleys.
The south fork of the Wichita River flows through the south
ern part of the ranch, and it along with several man-made
watering tanks and troughs provide water for the cattle
grazing on the ranch.
The second trapping location was the Kermit Waters Ranch
ca. 96 km (60 mi) north of the Masterson site. This ranch
encompassed ca. 3,200 hectares (8,000 acres) ca. 8.8 km (5.5
mi) northeast of Childress, in Childress Co., Texas. It is
much less rugged with gently rolling hills and few intermit-

23
Figure 3- Research locations in the Rolling Plains of Texas.
^ = Masterson Ranch, King Co., Tx. • = Waters Ranch, Childress Co., Tx.
= Garland Farm, Wichita Co., Tx. = Wight Ranch, Nolan Co., Tx.

24
tent streams. Some water tanks occur on this ranch also,
most of which are fed by wells rather than by streams. The
vegetation consists of various grasses and honey mesquite
(_P. glandulosa) . Junipers do not occur at this site.
The third trapping location was on the Gad Garland farm,
ca. 8 km. (5 mi) east of Burkburnett, in Wichita Co., Texas,
This site is on the far eastern edge of the Rolling Plains
and is relatively flat with only a few gently rolling hills.
It was the most open of all the sites with grassy meadows,
bordered by wheat and cotton fields. A few large trees
occur along the edges of a nearby stream.
The fourth study area was ca. 11.2 km. (7 mi) southeast
of Sweetwater on the Wight Ranch in Nolan Co., Texas. The
Wight Ranch consists of ca. 960 ha. (2,400 acres) of gently
rolling to rough terrain. This area most closely resembles
that of the Masterson Ranch, but is less rugged. The pre
dominant vegetation is also similar to the Masterson Ranch.
Running streams and man-made stocktanks are present as water
sources.

25
General Survey
Flies were collected at each locality by the use of modi
fied Manitoba traps designed by Thorsteinson et al. (1964),
modified by Davis and Sanders (1981) (Figure 4). A large
35.56 cm (14 in) copper float was painted glossy black and
hung in the center of the trap as an attractant due to the
positive tropism of Tabanids toward dark moving objects
(Bracken et al. , 1962; Ezell, 1973; Granger, 1970; Hansens,
1947; and Thompson, 1969). Cyanide kill jars were con
structed and placed at the end of the wire cone as the kill
ing mechanism. In addition, one 17 ft. Malaise trap (Bio-
quip model 2875A), was also used at the Masterson Ranch, to
capture those species not attracted to the Manitoba trap
(Roberts, 1972; and Steyskal, 1981). The Malaise trap was
modified from its original design by the addition of an
inverted cyanide kill jar at the trap apex (Figure 5).
Henceforth the Manitoba trap and the Malaise trap on the
Masterson Ranch are termed the 'Masterson site' and the 'Ma
laise site' respectively in all figures and discussion.
Manitoba traps were placed on the Masterson Ranch, Burk
burnett site, Childress site, and on the Sweetwater site on
May 14, May 17, May 27, and May 28, 1982, respectively. The
Malaise trap was placed on the Masterson Ranch on May 21 ,
1982.

26
Figure 4. Modified Manitoba trap used in surveying tabanids of the Texas Rolling Plains during the summer of 1982.

27
0 '
-'».>ijr
Figure 5. Modified Malaise trap with cyanide jar at apex as the killing mechanism used for surveying tabanids not attracted to the Manitoba traps in the Texas Rolling Plains during the summer of 1982.

28
Each trap was placed in close proximity to a water
source. The Manitoba trap at the Masterson Ranch was con
structed on the site which corresponded to Davis' (1979)
northeast location. An open valley was to the east of the
trap with the breastwork of a large watertank to the west
(Figure 6). The Malaise trap was placed on the southeast
portion of the ranch near the south fork of the Wichita
River. It was constructed between dense vegetation on one
side and a large cliff on the other which could constitute a
flyway for various Diptera (Figure 7). The Manitoba trap at
Burkburnett was located adjacent to a stream (Gilbert creek)
in an open field (Figure 8). The trap at Sweetwater was
situated ca. 20 meters south of a stream that fed a large
water tank ca. 100 meters from the trap (Figure 9). The
Childress trap was ca. 300 meters west of a large spring fed
watering tank (Figure 10).
Exclosures were constructed at each site except for the
Burkburnett site where an existing corral kept the horses
out. A three strand barbed wire fence (ca. 5 sq.m.) was
constructed around each of the other traps, including the
Malaise trap, to protect livestock from the cyanide.
According to Joyce and Hansens (1968), Tabanidae are
reported to fly 0.3 to 0.9 m (1-3 ft.) above the surface of
the ground when searching for a blood meal. Therefore, the

29
Figure 6. Type of water source and topography nearest the Manitoba trap on the Masterson Ranch.

30
t i| fciEi 1 Ig 1 1 II . . ^ ^
^::T*^im-!^ '.•9.:,^* ' ^ •~%L ty^'^ i. ^ •'"• • ' x , .
BlmW/ii1^lm/.l
' i %
•^^»t^ " \
r.»lB Vl^-'. «'-,.«:-. J,
^ ^ ^ ^ ^ ^ ^ • » ^ t ^
' ••" •' l l \ ~??^>s
^••''jir'k
•_rjjj^ ,
P m 9 '^BH
'>•. •.
• ^ - - - •
\
. ,
Figure 7. Type of water source and topography nearest the Malaise trap on the Masterson Ranch.

31
^
Figure 8. Type of water source and topography nearest the Manitoba trap at the Burkburnett site.

32
Figure 9. Type of water source and topography nearest the Manitoba trap at the Sweetwater site.

33
'.% -
Figure 10 Type of water source Manitoba trap at the
and topography Childress site
nearest the

34
base of the plexiglass portion of the trap and the equator
of the large black ball, used as an attractant, were kept
one meter from the substrate. The plexiglass and black ball
were kept clean throughout the summer to ensure continual
attraction to flies. Vegetation within the exclosures was
kept short to allow easy access and high visibility to the
flies.
Approximately 22.5 kg. of dry ice (50 lbs.) was added
every other week to the Manitoba traps at Childress and
Sweetwater in an effort to obtain a greater diversity and
density of flies (Blume et al., 1972; Hansens et al., 1971;
and Roberts, 1970). The efficiency of dry ice baited traps
versus non baited traps was also tested. Dry ice was used
at the Burkburnett site on June 17 and July 28 1982. The
use of dry ice was later discontinued at this site for prac
tical reasons. Dry ice was placed in 3.5 cm thick styro-
foam coolers with perforated lids to allow carbon dioxide to
escape. Each container was placed under the black ball for
maximum effect.
Catches were removed weekly and cyanide jars replaced
when necessary. Flies at the Burkburnett site were not col
lected as regularly due to the distance and time involved.
They were instead removed about once every two weeks. All

35
collected specimens were placed in a freezer for future pin
ning and labelling.
Fly traps were removed from each site after the first
hard freeze (0 C or less for 2 or more days) and after two
consecutive weeks of empty traps. Traps were removed on 6
November 1982 at Childress, 13 November at the Masterson
Ranch (both Malaise and Manitoba traps), and 21 November at
Sweetwater and Burkburnett.
Representatives of each species captured were sent to Dr,
L. L. Pechuman at Cornell University, Ithaca, New York for
species determination. Species separation of collected
tabanids was sometimes difficult due to damage incurred to
them by other insects (mostly grasshopers) collected in the
traps. Unidentifiable specimens were removed from the sam
ple and discarded.
The effectiveness of Malaise and Manitoba traps in col
lecting tabanids was compared. The abundance, seasonal
occurrence, and date of peak populations of tabanids were
determined for each trapping location. The abundance and
period of occurrence of each species collected in the Roll
ing Plains were also determined. The date for peak field
populations was determined for those collected most often
(n>5). A comparison among species of tabanids to the cli-

36
mate (U. S. Environmental Data Survice, 1981a, 1981b), geog
raphy, and vegetation of each site was also conducted in an
effort to determine factors which may influence the species
composition at each site.
Biological Studies of
Tabanus abactor
Nighttime Habits and Habitats
The nighttime habits and habitats of T. abactor were
determined through a capture recapture technique. Flies
were collected, marked with fluorescent paint, and located
in the field after dark using a hand-held black light to
determine their location and behavior at night.
Collecting of adults was facilitated by the natural
attraction of T. abactor females to the sides and tires of a
light blue pickup truck while driving through the ranch at
16 to 32 kmh. (10 - 20 mph.). This attraction was due to
the truck's temperature, appearance, and movement. They were
then collected from the truck with an aerial net at various
locations on the ranch. During late summer, when fly num
bers diminished, dry ice was added to the back of the pickup
under a camper shell as an additional attractant. The cam
per shell was left open when driving and usually facing down

37
wind when the vehicle was not moving to attract more flies.
Flies were then collected from within the camper shell as
well as from other areas of the vehicle.
Flies were placed in 850 ml. ice cream containers with
four Kim Wipes © to lessen injury. The lids of the contain
ers were punctured before being placed into an ice cooler to
slow the flies' movements and facilitate marking (Figure 11)
Containers were removed from the cooler within 0.5 hour and
the flies were then removed individually for painting. This
removal was simplified by the attachment of a funnel-like
device to the ice cream container lid after the center was
removed. The funnel was comprised of two screens; one of
rigid 2 mm. metal mesh, with an inner liner of softer plas
tic mesh. These were constricted at the tip to allow only
one fly to exit at a time. The uppermost 2 cm. of a 50 ml.
(2.5 cm. diameter) plastic container with lid was attached
over this opening to further enable the release of only one
fly (Figure 11).
Many types of marking systems have been used for disper
sion studies of tabanids, including radioactive isotopes,
colored chalks, reflective paints, and fluorescent powders
(Beesly and Crewe, 1963; Bennett and Smith, 1968; Eddy et
al., 1962; Fales et al., 1964; Sheppard et al., 1973, 1980;

38
Figure 11 Materials used to study the nighttime habits and habitats of Tabanus abactor at the Masterson Ranch in King Co. , TT'. during the summer of 1982.

39
Thornhill and Hays, 1972; and Turner and Gerhardt, 1965).
Fluorescent paints and powders were experimented with in
this study. A fluorescent powder (zinc oxide) from Ferro
Corporation was used after Sheppard et al. (1973, 1980) who
used a fluorescent powder. Day Glo (B), in determining flight
routes in Louisiana. Marking and relocation using zinc
oxide powder proved very efficient; however, flies marJced
with this material, often flew immediately to vegetation and
began removing it. The material may have been harmful to
the flies, altering their behavior. Different colors were
also not available, so for these reasons fluorescent powder
was not used in this study.
An effective marker was a water based fluorescent Crayola
R Poster Paint (NO. 116 Non-Toxic Crayola (B) ) . Although
Beesly and Crewe (1963) showed no detrimental reactions to
oil base paints on Chrysops silacea, water-based paints show
less potential for harm. They also wear off quicker allow
ing the use of only a few colors and only one pattern as
compared to the complex patterns and color combinations used
by Beesly and Crewe in their studies. Six different colors
(fluorescent orange, green, pink, yellow, red, and blue)
were used to differentiate the various daily release sites
and dates (Figure 11).

40
Flies were hand painted with a drop of paint on the dor
sum of the thorax using a small paint brush. Most flies
were held by the legs during painting, with the wings being
held for those flies attempting flight. Originally flies
were kept until evening to be released. This resulted in
high mortality rates. The release of flies after dusk
resulted in some healthy flies, but these flies would not
fly. Consequently flies were released immediately after
marking, taking note of the general direction of flight
(with or against the wind). Any individuals unable to fly
normally immediately after marking were removed from the
population.
Flies were collected and released between 3 and 8 pm CST
which is the peak of T_, abactor activity (Hollander and
Wright, 1980a). The release time was shifted slightly as
the season progressed to allow at least two hours of flight
time before sunset.
Two release sites were selected. The first site (site A)
was the low valley where the Manitoba trap was located (Fig
ure 12). The second site (site B) was located on top of a
large mesa with a steep slope directly to the south and a
lesser sloped, but just as deep, valley to the north (Figure
13). There was heavy vegetation at the base of the southern
ravine where moisture was always present seeping from the

41
Figure 12. Nighttime habitat study release site A (Valley)

42
•^iif- %•
Figure 13. Nighttime habitat study release site B (Hilltop).

43
rocky ground. Large trees and an intermittent stream, where
standing water was sometimes present, were in the valley to
the north of this release site.
Marked J. abactor females were sought one hour after sun
set using a hand held blacklight. A Safari ® light (Burgess
Batt Division UL © Model 165/3 Gould Inc. Pat. No. 204903)
was used with a GE F8T5 BLB black light bulb (Figure 11).
The vegetation and substrate of the immediate and surround
ing areas of the release site were scanned as thoroghly as
possible for 2 - 3 hours each night, weather permitting.
Upon locating a fly, its position was marked with a red flag
for determining the flight distance the following day. The
vegetation on which it rested, the geographical location
(hill, sloped areas, or bottomland), height on the plant,
and habitat description (presence or absence of cover) were
noted. Congregation among flies was also recorded.
Larval Collections
The search for T. abactor larvae followed larval collect
ing techniques similar to those of Philip (1928) and Freeman
and Hansens (1972). Larvae were collected from streams and
ponds with soil sieves of different sizes. U.S. Standard
Sieve Series No. 6 with an opening of 3.36 mm. was used most
often, providing an opening small enough to halt larval

44
escape and large enough to allow the removal of loose soil
that was agitated with water. In addition, some large lar
vae were floated out during the sifting of soil with water.
These methods, although slow, were chosen over others due to
the increased possibility of success and the small amount of
damage the larvae would sustain during capture. Larval
searches were made in some atypical habitats for tabanids as
well, such as dry stream bottoms, around the base of vegeta
tion, and in manure pads and soil. Larvae were searched for
in these habits by manually digging and sifting through the
substrate in the field.
Most of the collected larvae were brought back to the
laboratory for rearing. Four types of rearing containers
were used. The majority of the larvae were placed in 400
ml. beakers with 100 to 150 ml of the substrate (sand & mud)
in which they were collected. The substrate was kept moist
at all times. The second group of larvae were placed in
34x17x9 cm. plastic shoe boxes half filled with their sub
strate at a slanting gradient from over half filled at the
upper end down to nothing at the lower'end. Water was
placed in these containers to maintain a constant moisture
gradient. Both types of containers were covered with perfo
rated plastic lids. All larvae were maintained at room
temperature and lighting conditions. The food for the lar-

45
vae was initially blood worms (chironomid larvae) and other
organic material found in the substrate but on March 16,
1982, following the advise of Dr. James Goodwin, all remain
ing larvae and any to be found later were fed beef liver
once a week. Two larvae were also placed into large glass
petri dishes of sand to facilitate observations at this time
as well. Some larvae were collected from atypical tabanid
habitats. These larvae were placed, with their natural sub
strate, into 275 ml. (9 mm. dia. x 5 mm. deep) plastic con
tainers with perforated lids. An attempt was made to main
tain the moisture of the substrate at the same level as it
was in nature.
Some larvae were preserved immediately in 75/& ethyl alco
hol after collecting for species identification. Represen
tatives of these larvae were sent to Dr. H. J. Teskey at the
Biosystematics Research Institute in Canada for determina
tion .

CHAPTER III
RESULTS AND DISCUSSION
General Survey
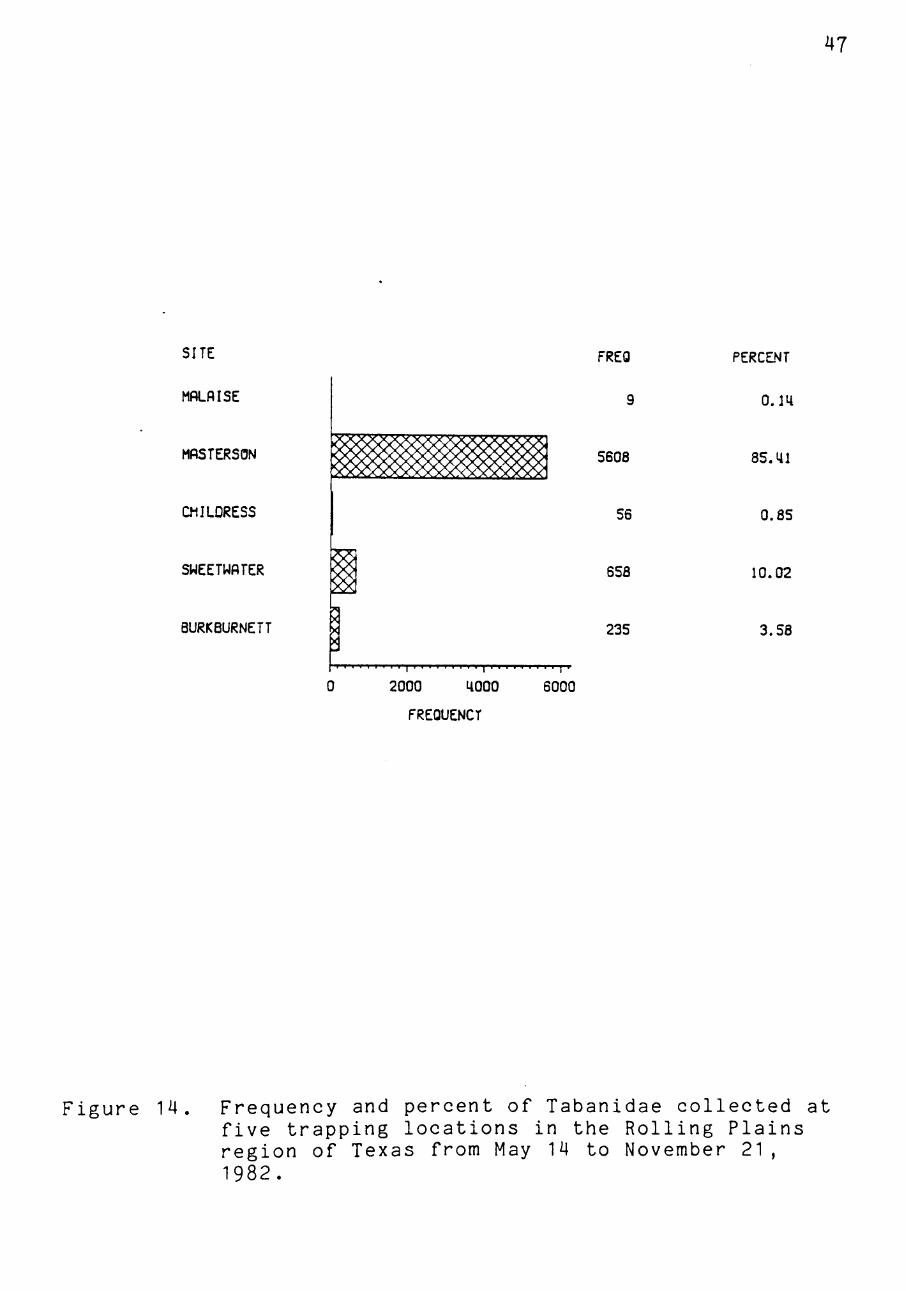
A total of 6,584 tabanids were collected from four Mani
toba traps and one Malaise trap in the study area. More
flies were collected by the Manitoba trap (x=79.5 +_ 256.7
S.D., n = 82) than were collected by the Malaise trap (x = 0.5 +_
1.61, n=19) which collected only nine flies, less than ^% of
the total (t=2.80, d.f.=8l, p<0.01). The most productive
trapping location was at the Masterson Ranch using the Mani
toba trap. Although no dry ice was used as an attractant
source for the Masterson site, over 85% of the flies col
lected during the survey were collected there (Figure 14).
Tabanids were collected in ever decreasing numbers at the
Sweetwater, Burkburnett, Childress, and Malaise sites
respectively (Figure 14). A significantly greater number of
flies were collected at the Masterson site than at any other
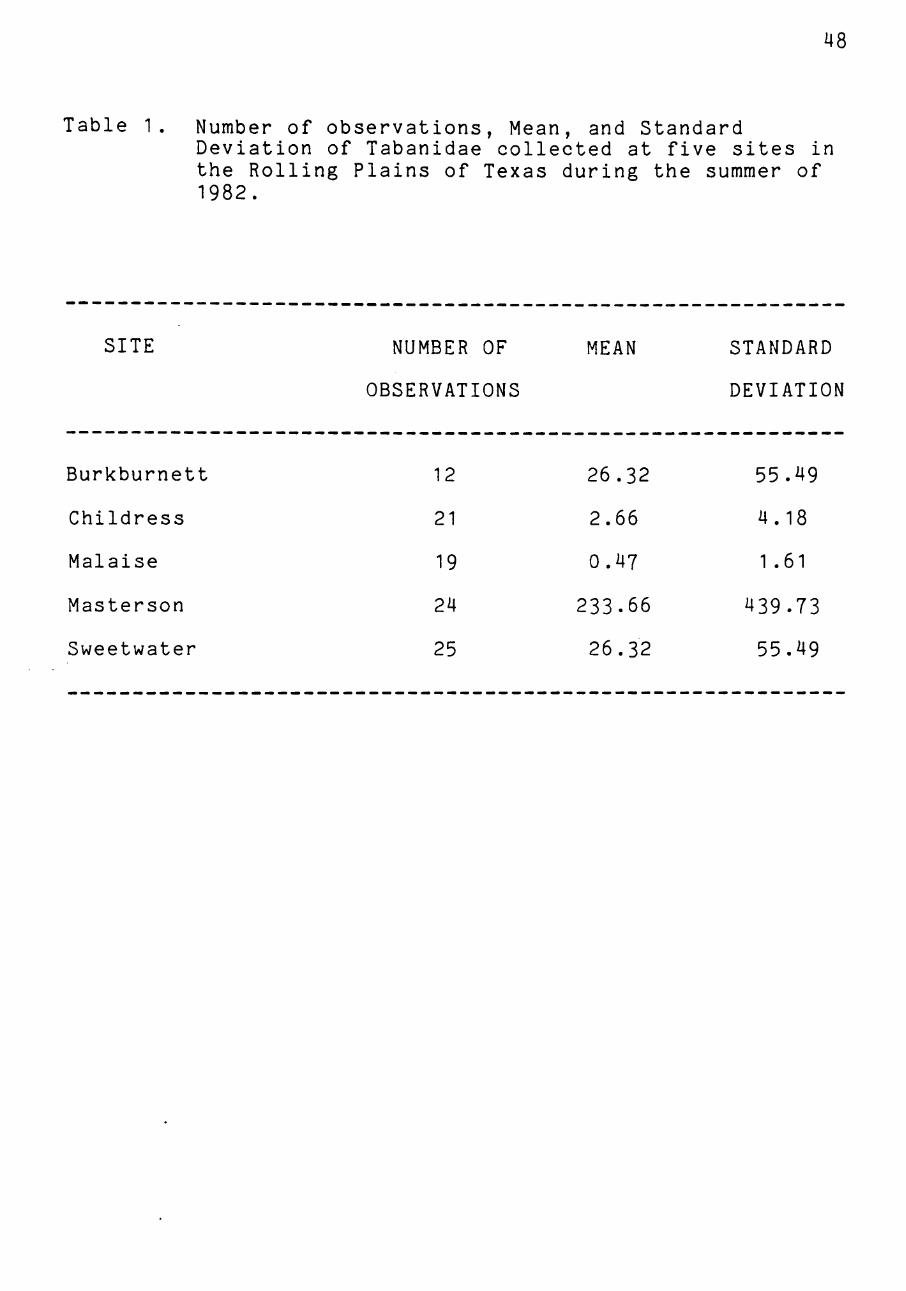
trapping location, using T tests at the 95% level of signif
icance (Table 1).
Peak populations were noted on July 30, July 29, July 16,
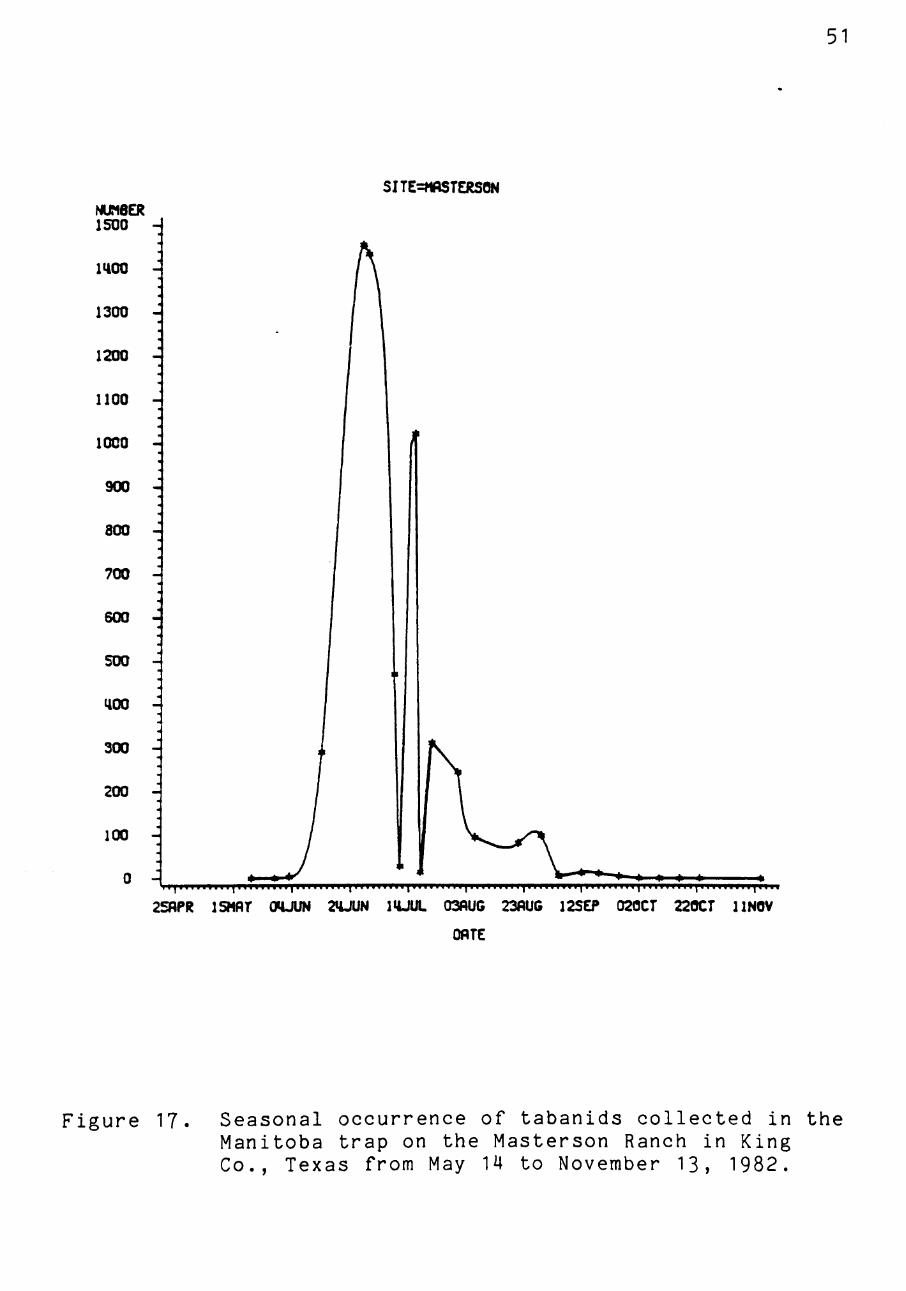
June 28, and August 2, for the Burkburnett, Childress,
Malaise, Masterson, and Sweetwater sites, respectively (Fig
ures 15-19). Those flies collected from Burkburnett, Chil
dress, and Sweetwater reflected a tendency for population
46
47
SITE
MflLfllSE
MRSTERSON
CMILORESS
SWEETWATER
BURKBURNETT
FREO
5608
56
658
235
PERCENT
o.m
85. m
0.85
10.02
3.58
1 1 ' 1 1 1 1 ' ' ' ' ' ' ' ' I ' ' ' ' ' ' ' ' ' 1 ' T - i - f - i - T 0 2000 4000 6000
FREQUENCr
Figure 14. Frequency and percent of Tabanidae collected at five trapping locations in the Rolling Plains region of Texas from May 14 to November 21, 1982.
48
Table 1. Number of observations. Mean, and Standard Deviation of Tabanidae collected at five sites in the Rolling Plains of Texas during the summer of 1982.
SITE NUMBER OF
OBSERVATIONS
MEAN STANDARD
DEVIATION
Burkburnett
Childress
Malaise
Masterson
Sweetwater
12
21
19
24
25
26.32
2.66
0.47
233.66
26.32
55.49
4.18
1 .61
439.73
55.49

49
SITE=8URK8URNETT fAJHSER 90
80
70
60
SO
HO
30
20
JO
» » i i i i i i i i » i » »
ISHflT T
OUJUN 2HJUN liUUL
QflTE
• • 1 ' '
OSflUG 23RUG 12SEP
Figure 15. Seasonal occurrence of tabanids collected in the Manitoba trap at Burkburnett, Texas from May 17 to November 21, 1982.

50
SITE=aiIU5RESS
NUnSCR
m
13
12
11
10
8
04JUN 24JUN 220CT 11NOV
Figure 16. Seasonal occurrence of tabanids co l l ec t ed in the Manitoba t r ap at Ch i ld r e s s , . Texas from May 27 to November 6, 1982.
51
SITE=mSTERSON NUMBER 1500 -}
moo -
1300 -^
1200 -j
1100 -
1000 -j
900 -
800 -I
700 -J
600 -j
500 -j
1400 ^
300 -
200 -J
100 -
0 - * » » * •i H 111 n 11111111 n 11111111 n 1111 11 11111 1111111111)1 ) 111 n 11111111 n I n I
2SnPR ISHflr aHJW ZUUH mJUL 03AUG 23AUC 12SEP 020CT 220CT IINOV
DATE
Figure 17. Seasonal occurrence of tabanids collected in the Manitoba trap on the Masterson Ranch in King Co., Texas from May 14 to November 13, 1982.

52
NUHSER 7
6
SITE=t1flU)ISE
H
T T 4 1 - * - * - • - • » » * » * I i i i i i i m m ^trrrr^FTwrjvrrrr^r^rjr.^'mrrrrTjr.^www^ry^r.^n^^^'trrjttf i n i rrTTTTTTyrT^
2SBPR ISMflT OqjUN 2«UUN lUJUL OSflUC 23flUG 12SEP 020Cr 220CT UNOV
OPTE
Figure 18. Seasonal occurrence of tabanids collected in the Malaise trap on the Masterson Ranch in King Co., Texas from May 14 to November 13, 1982.

53
SITE=SHEETHflTER
NUNSER 220
200
180
180 -
140
120 -
100 -
80 -
60 -
40
20 -1
0 -• p^yT<^^^^H^^^^^ifWi»W>TTTT<^'>TTTT^^*^^^^^^T**^^*'''" ' < |IIH.^^^TT^^ ;
25APR ISMPT 04JUN 2(UUN 14JUL 03RUG 23PUG 12SEP 020CT 220CT 11NOV
ORTE
Figure 19. Seasonal occurrence of tabanids collected in the Manitoba trap at Sweetwater, Texas from May 28 to November 21, 1982.

54
peaks to occur later in the season, near the end of July, a
difference of an entire month from the Masterson site, where
fly populations peaked near the end of June. This differ
ence may be important in the timing of broadcast insecti
cides for their control.
A total of 15 species were collected within the Rolling
Plains region (Table 2). As would be expected, all but four
of these were females. Males are not normally collected at
traps since they are not attracted to them for feeding pur
poses. However some males were collected. Two T. abactor
males were collected, one at the Masterson site and one at
Sweetwater. One Tabanus atratus Fabr. male and one Tabanus
subsimilis subsimilis Bellardi male were collected at the
Burkburnett site. In addition one dead male T_. abactor was
found lying in a dry stream bottom on the Masterson Ranch.
Traps were empty 45 of 106 times inspected. The low cap
ture rate was largely due to the poor efficiency of the
Malaise trap at the Masterson Ranch site and the Manitoba
trap at the Childress site, collecting flies on only 3 of 19
and 11 of 21 dates, respectively. The continuation of trap
ping for two weeks beyond the first hard freeze also con
tributed to the high frequency of empty traps.
55
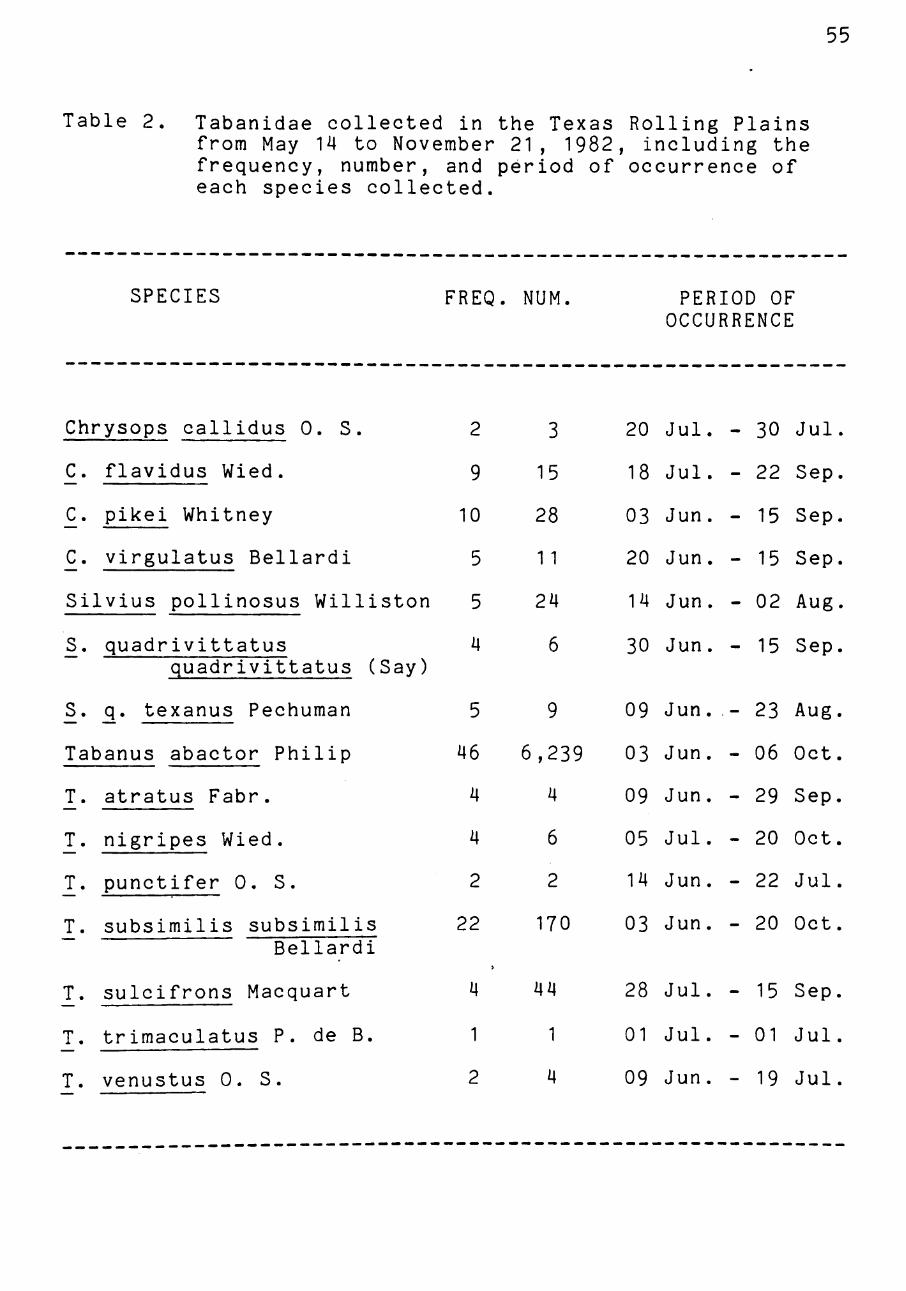
Table 2. Tabanidae collected in the Texas Rolling Plains from May 14 to November 21, 1982, including the frequency, number, and period of occurrence of each species collected.
SPECIES FREQ. NUM. PERIOD OF OCCURRENCE
Chrysops callidus 0. S
C. flavidus Wied.
—' Pi^^i Whitney
C_. virgulatus Bellardi
Silvius pollinosus Williston
^. quadrivittatus
quadrivittatus (Say)
^. q, texanus Pechuman
Tabanus abactor Philip T. atratus Fabr.
T. nigripes Wied.
T. punctifer 0. S.
T . subsimilis subsimilis Bellardi
2
9
10
5
5
4
5
46
4
4
2
22
3
15
28
1 1
24
6
9
6,239
4
6
2
170
20
18
03
20
14
30
09
03
09
05
14
03
Jul.
Jul.
Jun .
Jun.
Jun .
Jun.
Jun.
Jun.
Jun.
Jul.
Jun.
Jun.
- 30
- 22
- 15
- 15
- 02
- 15
- 23
- 06
- 29
- 20
- 22
- 20
Jul.
Sep.
Sep.
Sep.
Aug.
Sep.
Aug.
Oct.
Sep.
Oct.
Jul.
Oct.
T. sulcifrons Macquart
T. trimaculatus P. de B
T. venustus 0. S.
4 44
1 1
2 4
28 Jul. - 15 Sep
01 Jul. - 01 Jul
09 Jun. - 19 Jul.

56
The probability of collecting T. abactor in the Rolling
Plains was found to be significantly greater than the prob
ability of collecting any other tabanid species (X =
5323.14, d.f.=l4, p<0.05). Tabanus abactor was collected
most often, present in 46 of the 106 collection dates (Table
2). Tabanus subsimilis subsimilis Bellardi and the deer fly
Chrysops pikei Whitney were also collected quite often
(Table 2). Tabanus abactor was also present within the
Rolling Plains for the longest period of any tabanid during
the summer (Table 2). Field observations were consistent
with trapping data. Tabanus abactor females were seen to
attack cattle during the same period and at the same inten
sity as they were collected in traps.
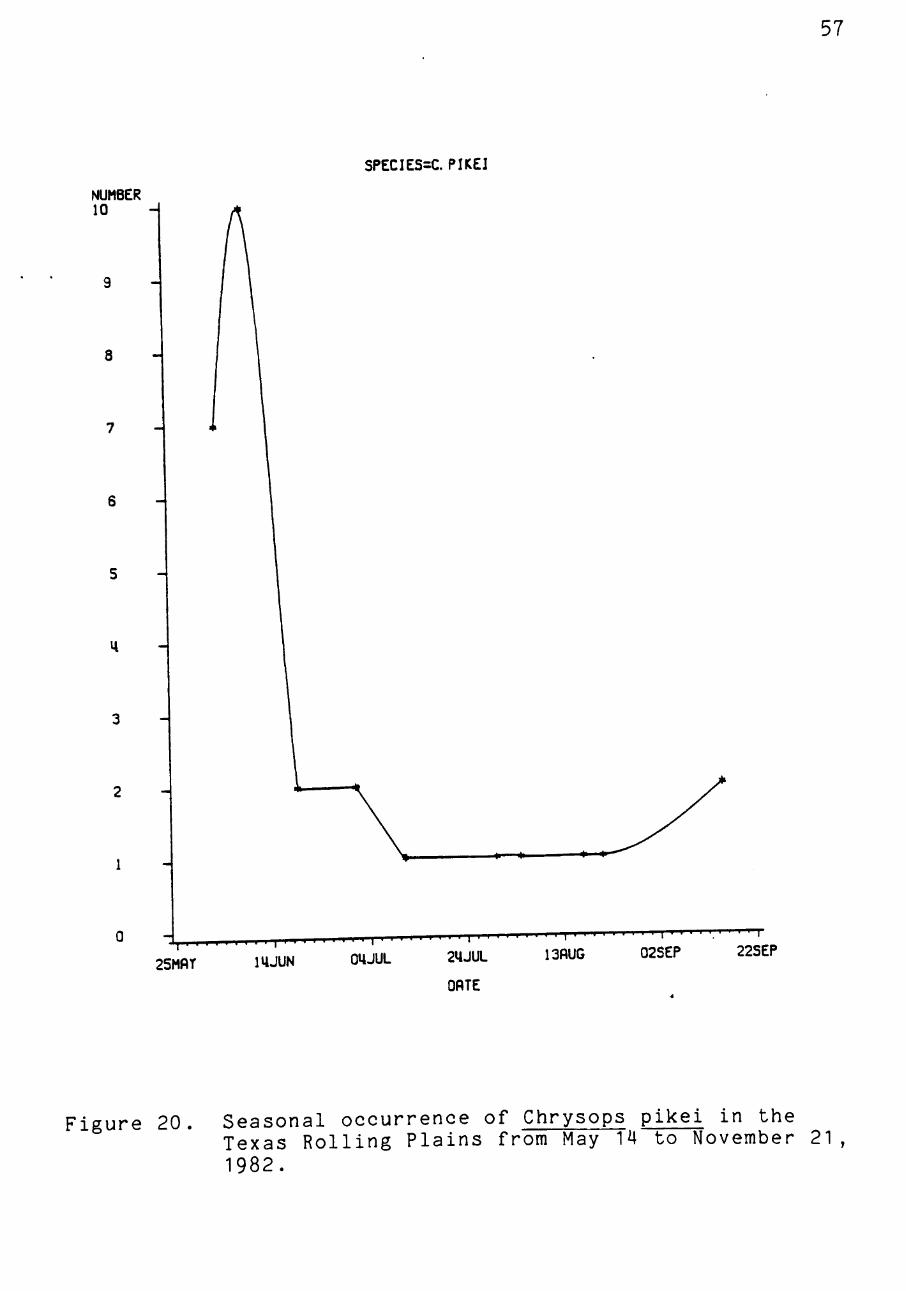
Chrysops pikei, T. abactor, and T^. _s. subsimilis were
collected on more than five occasions within the study area.
Chrysops pikei, collected at both the Childress and Master-
son sites, showed peak populations in late May and June,
although perhaps the population could have peaked before
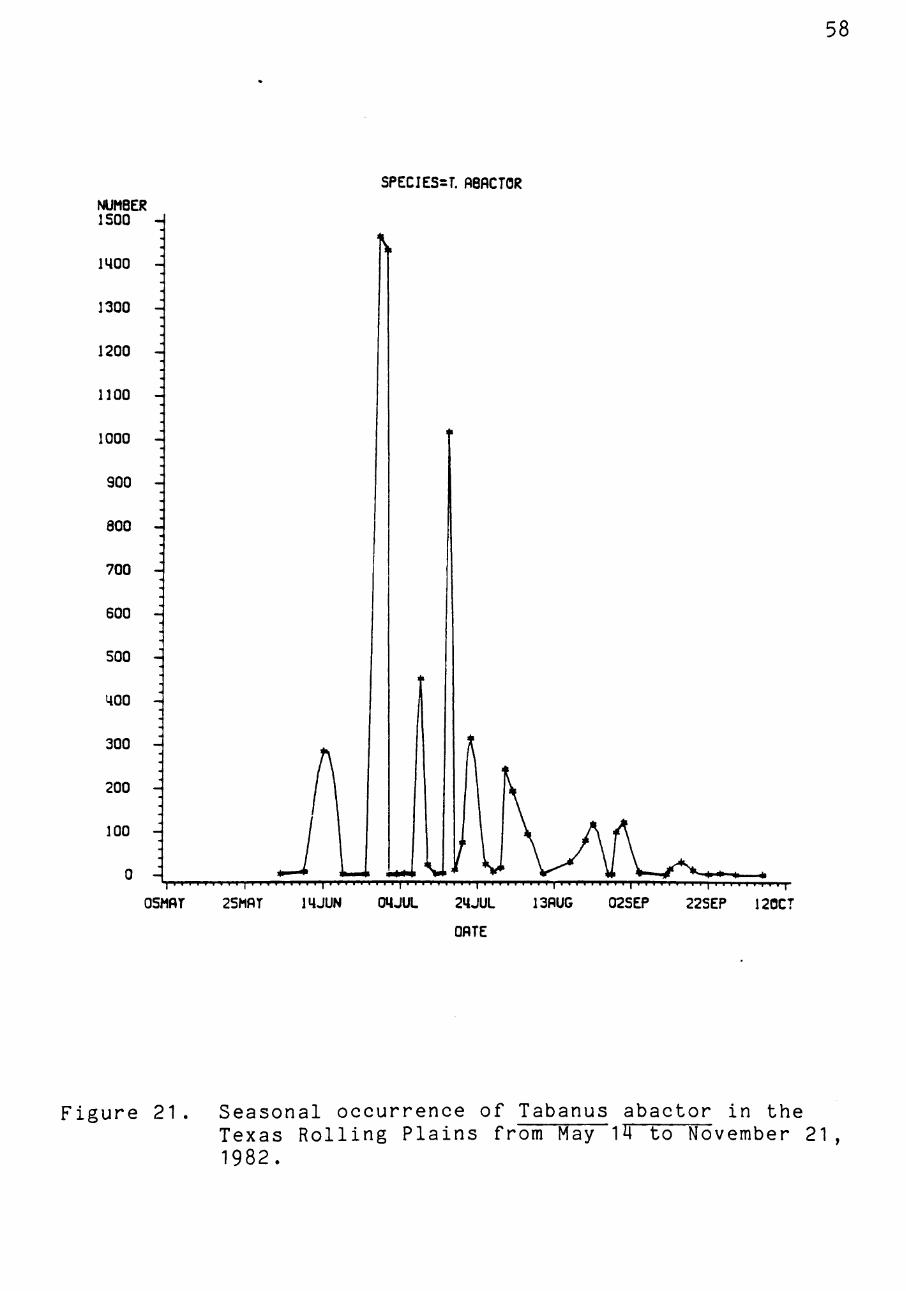
traps were constructed (Figure 20). Populations of T. abac-
tor peaked in late June to early July, with flies being
present throughout the summer from early June till frost in
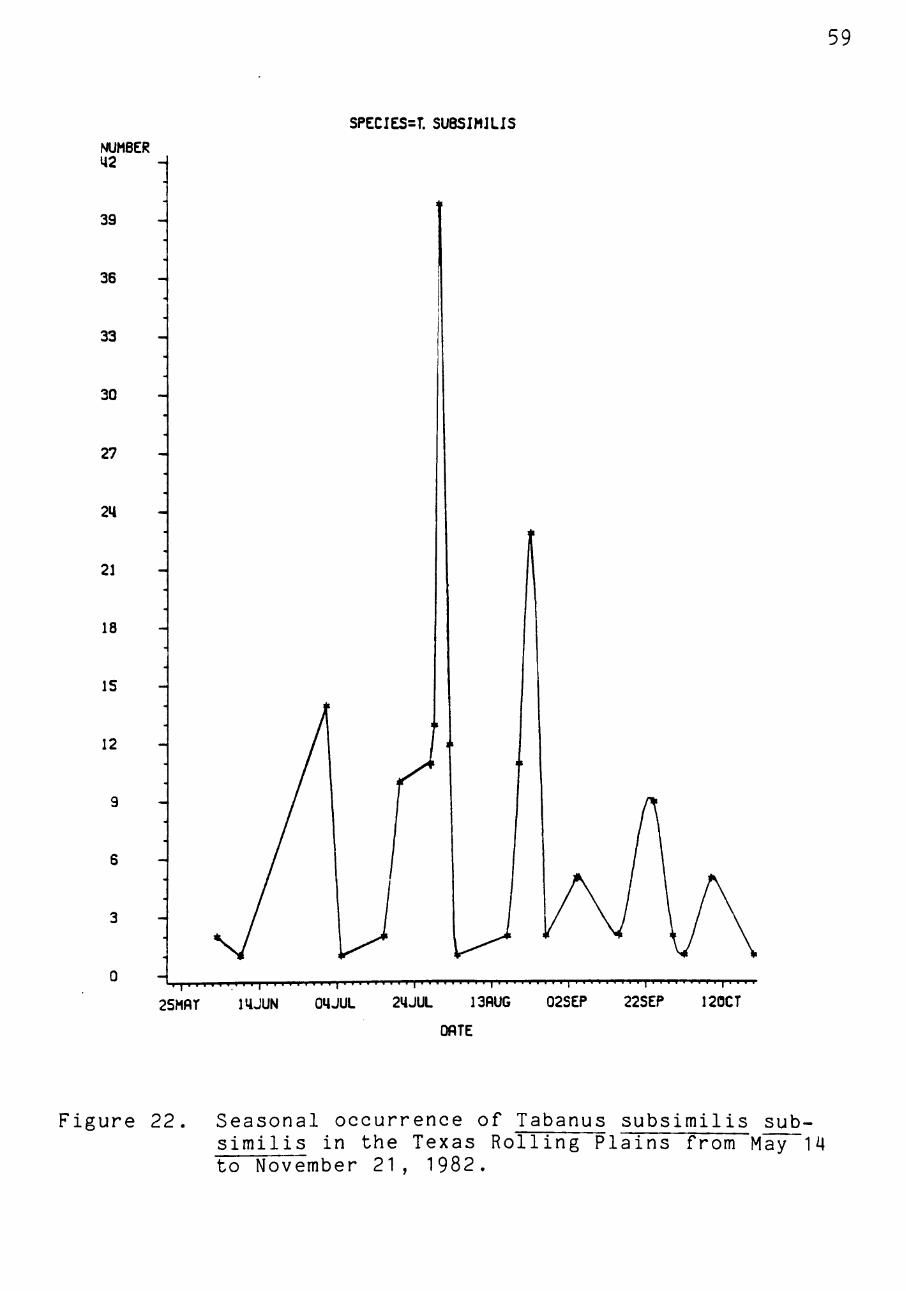
October (Figure 21). Tabanus £. subsimilis populations
peaked later in the season with the highest populations
present during late July (Figure 22).
57
SPECIES=C. PIKEI
NUMBER 10
-¥• *-
• ' ' ' I I I I I I
25MflT 14 JUN OUJUL 24JUL
DATE
13flUG 02SEP 22SEP
Figure 20. Seasonal occurrence of Chrysops pikei in the Texas Rolling Plains from May 14 to November 21 , 1982.
58
SPECIES=T. flBflCTOR
NUMBER 1500 -I
moo -
1300 ^
1200 ^
1100 -J
1000 -;
900 -J
800 -
700 ^
600 -_
500 -|
400 ^
300 -|
200 -J
100 -
0 - u OSMflT 25MflT 14JUN
* * • >• *
I I I I I I I I 1 1 1 1 1 1
04JUL 24JUL
ORTE
• I I I I I I I I I I I I I I I . . . . . T r I J
13flUG 02SEP 22SEP 120CT
Figure 21 . Seasonal occurrence of Tabanus abactor in the Texas Rolling Plains from May 14 to November 21, 1982.
59
SPECIES=r. SUBSIMILIS
NUMBER 42
39
36
33
30
27
24
21
18
IS
12
1 1 1 1 I ' ' ' I I 1 I I
25MflT 14JUN 04JUL 24JUL 13flUG 02SEP 22SEP 120CT
DATE
Figure 22. Seasonal occurrence of Tabanus subsimilis sub-similis in the Texas Rolling Plains from May 14 to November 21, 1982.

60
Although more flies were collected from the Masterson
Ranch location, more species were collected at the Burkbur
nett site (Figures 23-26). This may be due in part to the
use of CO2 at this site as an attractant. The Masterson and
Sweetwater locations yielded the same number of species
although the species were quite different (Figures 23, 24).
Both sites had a high percentage of T. abactor (99/& and 87%
respectively). The presence of T. abactor in similar num
bers at both sites indicates the presence of some common
factor or factors which are beneficial to its development.
Tabanus abactor was the only species collected in the
Malaise trap (Figure 25). Tabanus abactor was also the most
prevalent species at the Burkburnett site, followed by J. ^.
subsimilis, Tabanus sulcifrons Mackquart, and C_, pikei
respectively (Figure 26). As might be suspected, those
sites with greater numbers of a single species (T. abactor)
seem to have fewer numbers of species present and those
which are present are only present in small quantities.
The Childress site produced very few flies, 56 in all,
with T. s, subsimilis being collected most frequently (Fig
ure 25). It is possible that the location of the trap could
have been the reason for the low capture rate; however, the
owner also confirms the low density of horse flies on his
ranch (Mr. Waters, pers. comm.).

61
SITE=MflSTERSON
SPECIES
T. PUNCTIFER
T. ATRATUS
T. ABflCTOR
S. POLLINOSUS
S. aOUflORIVIT
S. Q. TEXRNUS
C. FLflVIDIS
C. PIKEI
T. SUBSIMILIS
^
FREO
5566
21
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 111111
I 2000 4000 6000
FREQUENCY
PERCENT
0.04
0.02
99.25
0.37
0.09
0.11
0.05
0.04
0.04
Figure 23. Frequency.and percent of tabanid species collected at the Masterson Ranch, King Co., Texas, Summer 1982, by Manitoba trap without CO2•

62
SITE=SWEETWflTER
SPECIES
T. ATRATUS
T. ABACTOR
S. POLLINOSUS
S. Q. TEXANUS
C. FLAVIDIS
T. NIGRIPES
C. VIRGULATUS
T. SUBSIMILIS
C.CALLIOUS
:^
I
FREO
574
10
11
51
1 1 1 1 1 1 1 1 1 1 1 1
PERCENT
0.15
87.23
0.46
0.15
1.52
0.76
1.67
7.75
0.30
r * • ' • I ' • • ' I • ' ' ' I ' • ' • I • • • • 1
0 100 200 300 400 500 600
FREOUENCT
LEGEND: C02 N \ \ \ 1 PRESENT fyV/yl ABSENT
Figure 24. Frequency and percent of tabanid species collected at Sweetwater, Texas, Summer 1982 by Man itoba trap with and without CO2.

63
SITE=CHILORESS
SPECIES
T. ATRATUS
T. ABACTOR
C. FLAVIDIS
T. SUBSIMILIS
^
FREO
0 10 20 30 40 50
FREOUENCT
46
PERCENT
1.79
14.29
1.79
82.14
SPECIES
T. ABACTOR ^
SITE=MALAISE
I T^TTTT H ^ I T TT^yTV t f f T l l T f r t ^
FREO
0 1 2 3 4 5 6 7 8 9
FREOUENCT
PERCENT
100.00
LEGEND: C02 N W M PRESENT ry//A ABSENT
Figure 25. Frequency and percent of tabanid species collected at Childress, Texas, by Manitoba trap and at the Masterson Ranch by Malaise trap. Summer 1982, with and without CO2.

64
SITE=8URICBURNETT
SPECIES
I ATRATUS
T. ABACTOR
S. 0. OUADRIVIT.
S. Q. TEXANUS
C. FLAVIDIS
C. PIKEI
T. VENUSTUS
T. TRIMACULATUS
T. SULCIFRONS
T. NIGRIPES
T. SUBSIMILIS
C. CALLIDUS
±:
:^ Zl
(A
FREQ
82
26
44
71
PERCENT
0.43
34.89
0.43
0.85
0.43
11.06
1.70
0.43
18.72
0.43
JO. 21
0.43
0 20 40 60 80 100
FREOUENCT
LEGEND: C02 N \ \ \ 1 PRESENT \///A ABSENT
Figure 26. Frequency and percent of tabanid species collected at Burkburnett, Texas, Summer 1982 by Manitoba trap with and without CO2.

65
A comparison of the rainfall, temperature, vegetation,
and topography among these sites suggests why T. abactor
predominates at the Masterson, Sweetwater, and Burkburnett
sites but not at the Childress location (Table 3). It
appears that the amount of rainfall present at a site corre
lates positively to the number of species present (Table 3,
Figs. 23-26). Rainfall alone probably cannot account for
the difference in species between sites. The temperature
differences between sites may also have an affect on tabanid
development (Table 3, Figure 14).
The vegetation and topography of these sites may have a
greater influence on tabanid development than the weather
since the availability of water is ultimately dependent on
the drainage of the site. The vegetation at each site is
generally indicative of the climate, soil types, topography,
and subsequent drainage and water availability of the site.
The main difference in these sites is the presence or
absence of junipers (Juniperus spp.). This is due mostly to
soil composition and terrain. Junipers are especially com
mon on poor, dry, rocky, mesa type areas and in limestone
and gypeus soils (Correll and Johnston, 1979). These areas
are also characterized by numerous valleys and intermittent
streams where tabanids may develop. The Childress site
lacks junipers, with honey mesquite (?_. glandulosa) as the

66
Table 3. Average yearly temperature and cumulative rainfall of the four trapping locations in the Texas Rolling Plains.
LOCATION AVERAGE CUMMULATIVE TEMP, rc) RAINFALL (mm)
Burkburnett'' 17.83 69.14
(Wichita Falls)
Childress'' 16.61 52.50
Masterson Ranch^ 18.81 53.57 (Guthrie)
Sweetwater'' 17.39 56.36 (Roscoe)
(1) Based on 30 year averages.
(2) Based on 1981 and 1982 averages (U.S. Environmental Data Service, 1981 ,1982) .

67
predominant vegetation. This site also lacks valleys and
streams, as previously noted, being less rugged than the
other sites. The only water present is in man-made water
tanks fed by springs. The absence of development sites
other than ponds, in conjunction with low rainfall and
cooler temperatures, probably explains why so few species
are found at this location. The fields at the Burkburnett
site have been cleared of unwanted plant species and natural
vegetation is more scarce. Numerous streams, ponds, and
several vegetation types can be found at this site. These
varied habitats, along with high temperatures and rainfall,
probably account for the large diversity of species col
lected there.
Tabanus abactor was present at all sites, developing best
in areas of rugged terrain with numerous valleys and inter
mittent streams (Masterson and Sweetwater sites). Its pres
ence at Burkburnett in smaller numbers than at the Masterson
and Sweetwater sites indicates either the lack of suitable
development sites, or possibly competetion with other spec
ies developing at the Burkburnett site but not at the other
sites. The absence of large numbers of J. abactor at the
Childress site is probably due to the combination of unsui
table habitats, low rainfall, lower temperatures, and possi
bly competition with "!_. s^. subsimilis.

68
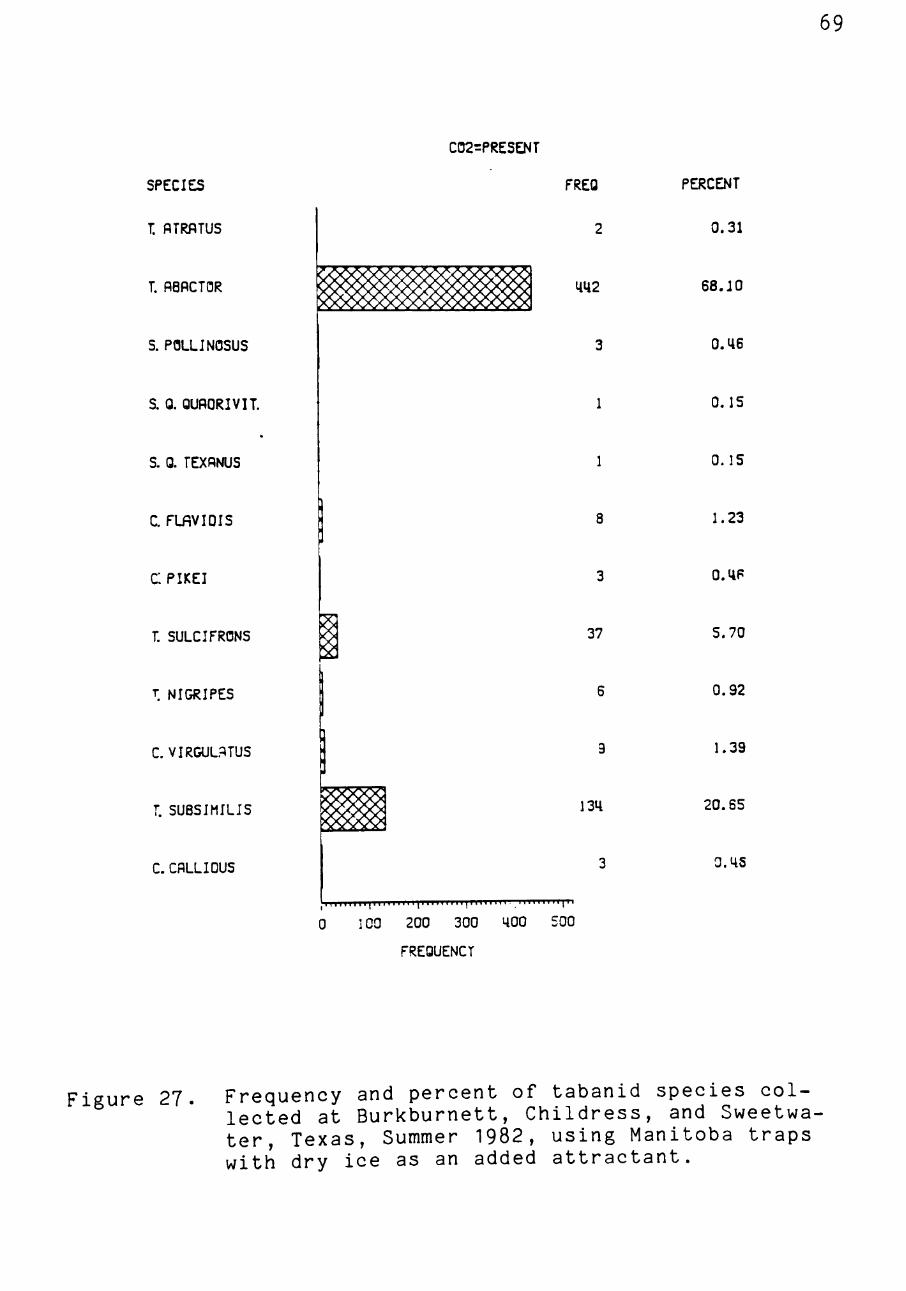
Carbon dioxide, used at the Childress, Sweetwater, and
Burkburnett sites, aided in the collection of tabanids.
Overall, a greater number of flies were collected using dry
ice (x = 31 + 57.0, n = 21) than without it (1? = 8 + 21.0, n = 37,
Figures 27, 28). This difference, however, was not shown to
be statistically significant (t=1.76, d.f.=23, p>0.05). The
lack of significant difference between the presence or
absence of CO2 could have been due to the method employed.
The holes in the lids of the styrofoam coolers containing
dry ice sometimes became frosted over, thus decreasing the
effective amount of CO2 released. The location of holes on
the sides of coolers may have prevented this.
Two more species were collected using dry ice than were
collected using the Manitoba trap alone (Figures 27, 28).
Only two species which were collected at Burkburnett, Ta
banus trimaculatus P. de B. and Tabanus venustus 0. S., were
collected only in the absence of dry ice (Figure 28). This
compares to four species being collected only in the pres
ence of dry ice (Chrysops callidus 0. S., Tabanus nigripes
Wied., Silvius pollinosus Williston, and Si 1 viu-s quadrivit-
tatus quadrivittatus Say, Figure 27). The use of CO2 seems
warranted when surveying tabanids as more individuals and
species are collected.
69
C02=PRESENT
SPECIES
T. ATRATUS
T. ABACTOR
S. POLLINOSUS
S. Q. OUAORIVIT.
S. Q. TEXANUS
C. FLflVIDIS
C: PIKEI
T. SULCIFRONS
r NIGRIPES
C.VIRGUL3TUS
T. SUBSIMILIS
C.CALLIDUS
FREQ
442
37
134
1 1 1 I I I I I I 1 1 1 1 1 1 I I 1 1 |i 1 1 " " ' 1 1 1 " " ' 11 I " " " " I "
0 ICO 200 300 400 500
FREOUENCT
PERCENT
0.31
68.10
0.46
0.15
0.15
1.23
0.4F
5.70
0.92
1.39
20.65
45
Figure 27. Frequency and percent of tabanid species collected at Burkburnett, Childress, and Sweetwater, Texas, Summer 1982, using Manitoba traps with dry ice as an added attractant.

70
C02=ABSENT
SPECIES
T. ATRATUS .
T. ABACTOR
S. Q. TEXANUS
C FLAVIDIS
C. PIKEI
T. VENUSTUS
T. TRIMACULATUS
T. SULCIFRONS
C. VIRGULATUS
T. SUBSIMILIS
X
FREO
222
23
34
l l l l l l l I I [ I I I I I I I I H I I I I I I I I I I I M l l l l M ] I I I l l l l l l | l l
0 50 100 150 200 250
FREOUENCT
PERCENT
0.33
74.00
0.67
1.33
7.67
1.33
0.33
2.33
0.67
11.33
Figure 28 . Frequency and percent of tabanid species collected at Burkburnett, Childess, and Sweetwater, Texas, Summer 1982, using Manitoba traps without dry ice.

71
Nighttime Habitats
The handling and painting of flies was not detrimental to
their normal activities. In most cases marked and released
individuals were seen to return immediately to the vehicle
or to me and attempt feeding. Tabanus abactor females
marked on previous days were also seen to return to attack.
The flies could be easily located using the hand-held black-
light and during dark nights could be seen 20 meters or more
away. This distance diminished rapidly, however, during
bright moonlit nights. The colors most observable in the
field were fluorescent orange and yellow. Several arachnids
(scorpions and spiders) fluoresced with the same frequency
as the fluorescent yellow paint which caused some difficulty
in locating marked flies. Fluorescent orange proved to be
the best color and was used as often as possible because of
this (Figure 29).
Out of a total release of 2,046 painted T. abactor
females, 104 were found. This corresponds to a 5% location
rate. In five instances flies were found on the second eve
ning after release, and one fly was even located on the
third night following release.
A total of 670 flies were released at site A (Valley
site) and 1,376 were released at site B (Hilltop site).

72
Figure 29. Painted T. abactor flies under U.V. light. Fluoresc'ent orange was the preferred color.

73
From these, 72 were located at site A and 32 at site B.
This corresponds to a location rate of 11% for site A and 2%
for site B. This difference is probably due to the geo
graphical dissimilarity between site A and B. The flies at
site B (Hilltop) also flew significantly further than those
at sight A (Valley) (x=242.0 + 221.9 m, n=32; x=26.3 + 29.8
m, n=32, respectively, t=-5.4, d.f.=32, p<0.01). This dif
ference in flight distance for each site probably also
accounts for the larger percentage of flies collected at
site A than at site B.
The flies after release usually flew with the wind rather
than against it. This is unusual since most insects orient
themselves against the wind when in flight. Generally, the
flies at site A flew in an east and west direction along the
valley (Figure 30). Those at site B flew with the wind,
sometimes for long distances, with one being located .8 km.
(.5 mi.) from the release site (Figure 30). Direction was
not taken into account initially; this explains why the
direction of flight is recorded for only 28 of the 72 flies
located at site A. In addition to flying with southerly
winds, flies also flew down into the valleys to the north
and south of the release site where larger vegetation and
water were often present.

74
FREOUENCT
30 ^
25 -
20 -
IS -
10 -
s -
N
DIRECTION
LECENOJ SITE 5 3 3 ^ 8
Figure 30. Direction of flight of marked Tabanus abactor females from the point of release at both sites used in nighttime habitat study; Site A=Valley site. Site B=Hilltop.

75
Flies remained stationary at night unless disturbed.
Tabanus abactor females were found to rest on several types
of vegetation (Figure 31). The occurrence of both the
grasses and the branch substrates signifies flies which pre
ferred to land on objects close to the ground. One fly was
found resting on the black ball attractant source (Sphere)
used on the Manitoba trap located at release site A. This
points to the possibility that some flies may remain on an
animal at night, although this was not examined in the
field.
There appears to be a vegetational preference for Junip-
erus pinchottii Sudw. (redberry juniper; Cedar, Figure 31),
but if the plant community as a whole is taken into account,
this is not so. Junipers are found to represent over 50% of
the plant community, perhaps as much as 70%. Larger trees
such as mesquite _P. glandulosa, elm Celtis occidentalis (L.)
and sumack Rhus microphylla (Engelm) are found in much lower
densities. The other plant species on which flies were
found: (Condalia hookeri M.C., Celtis reticulata Torr.,
Juglans microcarpa Berl., Rhus aromatica Ait., and Ziziphus
obtusifolia T. and G.) are also present in much lower densi
ties than junipers.

76
PLANT
Dead Branch on Ground
C. hookeri
C. occidentalis
C. reticulata
Grasses
J. microcarpa
£. pinchottii
£. glandulosa
R. aromatica
R. microphylla
Sphere on Manitoba Trap
Z, ob tus i fo l i a
FREO
13
6
11
3U
m
m
^ t ^ ^ ^ W ^ ^ ^ ^ . 1 I I I I I I I I ^p^^-rT^^^^y. . r^^"^^r^^^T
10 20
FREOUENCT
30 40
PERCENT
0.96
1.92
12.50
S.77
10.58
0.96
32.69
13.46
5.77
13.46
0.96
0.96
Figure 31. Frequency and percent of Tabanus abactor females located at night on different vegetation and substrates at both site A and B.

77
A significantly greater number of flies were found to
prefer cryptic habitats over more open types (x=4.M j- 6.0,
n=17; x=1.0 + 2.2, n=17, of flies located/date for leafy and
open habitats, respectively; t=2.7, d.f.=l6, p<0.05).
Approximately 72% of those located were found on leafy
areas, with only 2S% preferring some other type of environ
ment .
The distance flies were found from the ground corre
sponded to the type of plant on which they were found (Fig
ure 32). The average size of junipers and other smaller
vegetation accounts for the presence of 44 flies (42%) found
between 1.05 - 1.50 meters (Figure 32).
The flies' geographical location also had an impact on
their spatial distribution. A large number of flies were
found above 2.25 meters at site B (Figure 32). These were
in large trees at the edges of dry stream beds in a valley.
It is also possible that other flies at higher resting sites
were not observed due to the limited reach of the black-
light.

78
FREOUENCT
4 5 H
40 -
35
30 -
25 -
20 -
15 -
10 -
5 -
3- . 55 . 5 5 - 1 . 0 5 1 . 0 5 - 1 . 5 0
HEIGHT IN METERS
LEGENO! SITE I i fl &-666<l 8
1 , .50-2.25 >2 .25
Figure 32. Spatial distribution (height from the ground) of Tabanus abactor females located on vegetation at night at site A and B.

79
Congregation was observed once between one painted and
one unpainted fly. Flies were also distributed unevenly
throughout the ranch, with large populations being located
in certain regions.
Flies showed a marked geographical preference for shel
tered low-lying valleys and slopes over more open, flat
hilltops (Figure 33). Overall, 86% of the flies located
preferred these low-lying areas to more open hilltops.
Those flies released at site A remained in the valley (38%)
or moved toward the slopes (62%), never moving to the open
hilltops (Figure 33). The slopes at this site usually cor
responded with the breastworks of a dam with heavy vegeta
tion growing on it. Those flies released at site B (hill
top) preferred to fly to the valley, 450 meters away (47%)
or slopes (25%), rather than remaining on the hilltops
(28%).
The adults' preference for low-lying sheltered areas may
indicate that similar locations are utilized as feeding and
breeding sites. These areas could also'be the best targets
for broadcast insecticide treatments. Broadcast sprays
applied in early morning or late evening before adults
become active may reduce T. abactor populations.

80
PERCENTflGE
35 -I
30 -
25 -
20 -
;5 -
10 -
s -1
R 8
j — HILLTOP
14%
R 6
SLOPE
44%
fl 8 SITE
VALLEY ! OESC
42%
Figure 33. Geographical preference of Tabanus abactor females at night within the Texas Roiling Plains.

81
Larval Collections
A total of 57 tabanid larvae were collected from the Mas
terson Ranch (Table 4). Fifty-three larvae were collected
from typical tabanid habitats and five were collected from
dry (atypical) sites. Larvae were collected from the litto
ral zone of both watering tanks and small watering holes
found in intermittent stream beds. Larvae were usually
found under 2.5-4 cm. of mud, but some were collected away
from the shore under 2-3 cm. of water. The larvae collected
from these habitats appeared to be the same species. In
addition two other larvae of the same species were collected
from dried stream beds, an atypical habitat for tabanids.
Larvae were light colored, with dark bands at the junction
of each segment, and were usually very active (Figure 34).
Two of these larvae pupated and one male T_. atratus emerged
on April 24, 1983.
According to H. J. Teskey (Biosystematics Research Insti
tute, Canada), representatives of these larvae were all of
the same species but definitely were not J. atratus. He
also states that these larvae have not been previously iden
tified. Those Tabanus species from this region which have
not been described are T. abactor, T. cheliopterus, T.
equalis, T. nefarius, T. quaesitus, and T. texanus (H. J.

82
Table 4. Tabanid larvae collected on the Masterson Ranch in King Co., Texas, Summer 1982 and Spring 1983, . including number, size range, and site description for larvae on each collection date.
Larva Date Number Size Range Site in mm. Description
Unidentified
M It
H i t
»1 U
tl It
It It
It It
Ii 11
It I I
T. atratus 1
Unidentified
It 11
It It
Aug.
Aug.
Sep.
Sep.
Sep.
Sep.
Oct.
Oct.
Oct.
Oct.
Mar .
Mar .
Apr .
Apr .
26,
27,
5,
12,
18,
25
2
2
16
23
13
13
30
30
82
82
82
82
82
r 82
, 82
, 82
, 82
, 82
, 83
, 83
, 83
, 83
1
1
5
4
4
5
2
7
1
2
2
1
1
3
2.5X.7
2.5X.5
2.5X.3-5.5X.7
1.5X.2-3.5X.4
1.7X.2-5.7X.5
1.5X-3.5X1 .5
3.IX.6-3.5X.7
.9X.1-3.5X1.5
1 .5X.2
3.IX.6-3.5X.7
2.3X.3-4.5X.4
3.8X.5
3.2X.6
.7X.1-1.8X.3
A
A
B
B
A
B
C
B
C
C
C
D
D
E
Site Description:
A = Intermittent stream bed with small water puddle. B = Intermittent stream bed with large water puddle. C = Large water tank, sandy - rocky shore-line. D = Dry intermittent stream bed. E = Manure pad, xeric habitat. 1 = Pupated or Emerged.

83
Figure 34. Larvae of unidentified Tabanus spp. (possibly T abactor) collected from both~typical and atypi-"caT tabanid habitats at the Masterson Ranch, King Co., Texas, Summer 1982, and Spring 1983.

84
Teskey, pers. comm.). If the remaining larvae are not i;*
atratus, then the probability that they are T. abactor is
high because none of the other species mentioned have been
collected from this site. Also, adults of the other species
are too small to develop from the largest of the larvae col
lected (H. J. Teskey, pers. comm.). It appears that two
species have been collected, one being T. atratus, and the
other possibly T. abactor.
All but one of the other larvae taken into the laboratory
for rearing died. Those larvae placed in sand died within 3
days, apparently due to the abrasion of the substrate and
the higher moisture present. Most of those placed in 400
ml. beakers also died due to an uncontrolled increase in
temperature in the laboratory. The remaining larva has not
yet undergone metamorphosis.
Three larvae of a third species were collected on April
30, 1983 from two adjacent manure pads in xeric habitats.
The manure pads were dry on the surface but were slightly
moist in the center where the larvae were found. Other
insect larvae (Scarabaeidae), which could act as a food
source, were found in this habitat as well. Larvae were
yellowish in color, with no striations and were very slug
gish in their activity (Figure 35).

85
Figure 35. Tabanus larva (probably T. s. subsimilis) col-lected^from manure pad aT tHe Master son Ranch , King Co. , Texas, April 30, 1983.

86
Two of these larvae pupated on May 15, and May 13, 1983.
Pupae were obtect and dark brown to black with spines pro
jecting from the posterior portion of each abdominal segment
and stout spines projecting from the tip of the last abdomi
nal segment. Pupation lasted 11 days with one male and one
female Tabanus species emerging on May 26 and 29 respec
tively. These were both identified by R. Wright at Oklahoma
State University as probably being Tabanus subsimilis sub
similis . However, this species belongs to a species complex
with Tabanus lineola, and in some cases there is some uncer
tainty, as to the differences between these two species with
the possibility that a third, and undescribed species, is
also present.

CHAPTER IV
CONCLUSIONS
Tabanus abactor is the most abundant and important horse
fly pest infesting the Rolling Plains of Texas. It was
present at all, and predominated at most, sites within the
study area. Tabanus abactor appears to develop best in
areas of rugged terrain, with numerous valleys, intermittent
streams, and abundant junipers. Tabanids were more abundant
in regions with higher annual temperatures and rainfall.
Tabanus abactor is the dominant species throughout the Roll
ing Plains except in the northernmost region. Tabanus s.
subsimilis is the dominant species of this region, perhaps
due to the moderate terrain, cooler temperatures, and lower
rainfall. The nighttime resting locations of T_. abactor
also reveal their preference for low lying valley and stream
locations. Their attraction to these areas may prove useful
in control. Applications of insecticide to these areas in
early morning or late evening during the periods of the year
of peak tabanid abundance could significantly reduce tabanid
populations.
Larvae of T. atratus and an unknown tabanid (possibly T.
abactor) were found to develop in and around watering tanks
and streams and in some cases were able to resist desicca-
87

88
tion well enough to exist in dry stream beds. Application
of insecticides with short residual activity to the littoral
zone of water tanks may also help decrease populations of T_.
abactor and other tabanid species. The discovery of T. ^.
subsimilis larvae in the very atypical habitats of manure
pads is also significant. Other species (possibly T. abac
tor) may also develop there. If so, larvicidal feed addi
tives fed to range cattle could be used to render these
habitats inhospitable to tabanid development.
More research on the control of T. abactor is needed.
Before progress in the control of T_. abactor can be
achieved, the location and duration of larval development of
this pest must be determined. Other areas in need of
research include: 1) efficacy tests on insecticides directed
toward larvae and adults applied to the areas mentioned, 2)
determination of actual losses in weight gains due to
tabanid attacks, and 3) the possibility of using biological
control against both larvae and adults of T. abactor.

LITERATURE CITED
Anderson, J. F., and F. R. Kneen. 1969. The temporary impoundment of salt marshes for the control of coastal deer flies. Mosquito News. 29(2):239-243.
Bay, D. E., N. C. Ronald, and R. L. Harris. 1976. Evaluation of a synthetic pyrethroid for tabanid control on horses and cattle. Southwest. Entomol. 1(4):198-203.
Beesley, W. N., and W. Crewe. 1963. The bionomics of Chrysops silacea Austen. II. The biting rhythm and disper-sal in rain-forest. Ann. Trop. Med. Parasitol. 57:191-203.
Bennett, G. F., and S. M. Smith. 1968. Phosphorus for marking Tabanidae (Diptera). Mosquito News. 28(4):559- 568.
Blume, R. R., J. A. Miller, J. L. Eschle, J. J. Matter, and M. 0. Pickens. 1972. Trapping tabanids with a modified Malaise trap baited with CO,. Mosquito News. 32(1):90-95.
Blume, R. R., R. H. Roberts, J. L. Eschle, and J. J. Matter. 1971. Tests of aerosols of Deet for protection of livestock from biting flies. J. Econ. Entomol. 64 (5):1193-1196.
Borror, D. J., D. M. Delong, and C. A. Triplehorn. 1976. An Introduction to the Study of Insects. 4th ed. Holt, Rinehart and Winston, Dallas. 852 pp.
Bosler, E. M., and E. J. Hansens. 1974. Natural feeding behavior of adult saltmarsh greenheads and its relation to oogenesis. Ann. Entomol. Soc. Am. 67:321-324.
Bracken, G. K., W. Havec, and A. Thorsteinson. 1962. The orientation of horse flies and deer flies (Tabanidae Diptera). II. The role of some visual factors in the attractiveness of decoy silhouettes. Can. J. Zool. 40:685-695.
Brown, L., and J. L. Lancaster. 1973. Controlling horse flies on cattle. Ark. Farm Res. 22(5):6.
89

90
Bruce, W. N., and G. C. Decker. 1951. Tabanid control on dairy and beef cattle with synergized pyrethrins. J. Econ. Entomol. 44:154-159.
Correll, D. S., and M. C. Johnston. 1979. Manual of the vascular plants of Texas. Texas Research Foundation, Renner, Texas. 1881 pp.
Curran, C. H. 1966. Family Tabanidae - The horse flies, pp. 148-154. In The Families and Genera of North American Diptera. 2nd ed. Henry Tripp, New York.
Davis, S. G. 1979. Seasonal and geographical distribution of Tabanus abactor Philip and associated species in the Texas Rolling Plains. A Thesis in Entomology. Texas Tech University. Lubbock, Tx.
Davis S. G., and D. P. Sanders. 1981. Seasonal and geographical distribution of Tabanus abactor Philip in the Texas Rolling Plains. Southwest, tntomol. 6(2):8l-86.
Dikmans, G. 1950. The transmission of anaplasmosis. Am. J. Vet. Res. 11 :5-l6.
Fasten, E. R., M. A. Price, and 0. H. Grahm. 1968. The collection of biting flies in west Texas with Malaise and animal baited traps. Mosquito News. 28(3):465-469.
Eddy, G. W., A. R. Roth, and F. W. Plapp. 1962. Studies on the flight habits of some marked insects. J. Econ. Entomol. 55(5) : 603-607.
Ezell, W. B. Jr. 1973. Interspecific variation in the flight responses of tabanids (Insecta: Diptera) to certain visual and chemical stimuli. Bull. South Carolina Acad. Sci. 35:100-1 11 .
Fales, J. H., 0. F. Bodenstein, G. D. Mills, and L. H. Wes-sel. 1964. Preliminary studies on face fly dispersion. Ann. Entomol. Soc. Am. 57(1) : 135-137 .
Freeman, J., and E. J. Hansens. 1972. Collecting larvae of the salt marsh greenhead Tabanus nigrovittatus and related species in New Jersey: Comparison of Methods. Environ. Entomol. 1 (5 ): 653-658.
Granett, P., and E. J. Hansens. 1956. The effect of biting fly control on milk production. J. Econ. Entomol. 49(4) :465-467.

91
Granett, P., and E. J. Hansens. 1957. Futher observations on the effect of biting fly control on milk producing cattle. J. Econ. Entomol. 50:332-336.
Granger, C. A. 1970. Trap design and color as factors in trapping -the saltmarsh greenhead fly. J. Econ. Entomol. 63(5) :1670-1672.
Hansens, E. J. 1947. Greenhead flies (Tabanus nigrovittatus) like dark colours. New Jersey Agric. 29:3-4.
Hansens, E. J. 1956. Granulated insecticides against greenhead (Tabanus) larvae in the salt marsh. J. Econ. Entomol. 49:401-403.
Hansens, E. J. 1979. Review: Tabanidae of the east coast as an economic problem. J. New York Entomol. Soc. 87(4):312-318.
Hansens, E. J. 1981. Resmethrin and Permethrin sprays to reduce anoyance from a deer fly Chrysops atlanticus. J. Econ. Entomol. 74:3-4.
Hansens, E. J., E. M. Bosler, and J. W. Robinson. 1971. Use of traps for study and control of saltmarsh greenhead flies. J. Econ. Entomol. 64(6):1481-1486 .
Hawkins, J. A., W. V. Adams Jr., B. H. Wilson, C. J. Issel, and E. E. Roth. 1976. Transmission of equine infectious anemia virus by Tabanus fuscicostatus. J. Am. Vet. Med. Assoc. 68(1):63-64.
Hocking, B. 1953. The intrinsic range and speed of flight of insects. Trans. R. Entomol. Soc. London. 104:223-345.
Hollander, A. L., and R. E. Wright. 1980a. Daily activity cycles of Oklahoma Tabanidae (Diptera). Environ. Entomol. 9:600-604.
Hollander, A. L., and R. E. Wright. 1980b. Impact of tabanids on cattle: blood meal size and preferred feeding sites. J. Econ. Entomol. 73(3) : 431-433.
Issel, C. J., and L. Coggins. 1979. Equine infectious anemia: Current knowledge. J. Am. Vet. Assoc. 174(7): 727-733.

92
James, M. I., and R. F. Harwood. 1969. Horse flies, deer flies and snipe flies. pp.223-233. In Herm's Medical Entomology. 6th ed. The Macmillan Company. Collier-Macmillan Ltd. London.
Joyce, J. M., and E. J. Hansens. 1968. The influence of weather on the activity and behavior of greenhead flies, Tabanus nigriovittatus Macquart and T. lineola Fabricius. J. New York Entomol. Soc. 76:72-80.
Kahn, N. H. 1952. Oviposition and hatching in some species of Tabanidae. Ann. Entomol. Soc. Am. 43:550-553.
Kemen, M. J. 1977. Equine infectious anemia: The controversy continues. The Cornell Veterinarian. 67(2):177-189.
Kniepert, F. W. 1980. Blood-feeding and nectar-feeding in adult Tabanidae (Diptera). Oecologia. 46(1):125-129 .
Krinskey, W. L. 1976. Animal disease agents transmitted by horse flies and deer flies (Diptera: Tabanidae). J. Med. Entomol. 13(3) : 225-275 .
Lindquist, A. W., and E. F. Knipling. 1957. Recent advances in veterinary entomology. Ann. Rev. Entomol. 2:181-202.
Magnarelli, L. A., and J. F. Anderson. 1981. Sugar feeding by female tabanids (Diptera: Tabanidae) and its relation to gonotropic activity. J. Med. Entomol. 18(5): 429-433.
Magnarelli, L. A., J. F. Anderson, and J. H. Thorne. 1979. Diurnal nectar-feeding of salt marsh Tabanidae (Diptera). Environ. Entomol. 8(3) : 544-548 .
McGregor, W. S., and 0. C. Schomberg. 1952. A partial annotated list of species of the Tabanidae. J. Econ. Entomol. 45(4):746.
Parman, D. C. 1928. Experimental dissemination of the egg parasite Phanurus emersoni Girault and biological notes on the species. U.S.D.A. Circular No. 18. 7 pp.
Pechuman, L. L. 1981. The horse flies and deer flies of New York (Diptera, Tabanidae). Search. 18:1-66.

93
Philip, c. B. 1928. Methods of collecting and rearing the immature stages of Tabanidae (Diptera). J. Parasitol. 14(4):243-253.
Philip, C. B. 1931. The Tabanidae (horseflies) of Minnesota with special reference to their biologies and taxonomy. Minn. Agric. Exp. Sta. Tech. Bull. No. 80. 128 pp.
Philip, C. B. 1936. New Tabanidae (horseflies) with notes on certain species of the longus group of Tabanus. Ohio J. of Science. 36:149-156.
Roberts, R. H. 1970. Tabanidae collected in a Malaise trap baited with CO2. Mosquito News. 30(1):52-53.
Roberts, R. H. 1972. The effectiveness of several types of Malaise traps for the collection of Tabanidae and Cu-licidae. Mosquito News. 32(4 ) :542-547 .
Roberts, R. H., and R. J. Dicke. 1964. The biology and taxonomy of some immature Neartic Tabanidae (Diptera). Ann. Entomol. Soc. Am. 57:31-40.
Roberts, R. H., and W. A. Pund. 1974. Control of biting flies on beef steers: Effect on performance in pasture and feedlot. J. Econ. Entomol. 67:232-234.
Sanborn, C. E., G. W. Stiles, and L. H. Mai. 1932. Preliminary experiments in the transmission of anaplasmosis by horseflies. Oaklohoma Agric. Exp. Sta. Bull. No. 204. 15 pp.
Schomberg, 0. 1952. Larval habitat of Tabanus sulcifrons in Oklahoma. J. Econ. Entomol. 45(4) :747.
Schomberg, 0. D., and D. E. Howell. 1955. Biological notes on Tabanus abactor Philip and equalis Hine. J. Econ. Entomol. 48(5):6l8-619 .
Schwardt, H. H. 1931. The biology of Tabanus lineola Fabr. Ann. Entomol. Soc. Am. 24:409-416.
Schwardt, H. H. 1936. Horse flies of Arkansas. Ark. Agric. Exp. Sta. Bull. No. 332. 66 pp.
Sheppard, D. C., B. M. Wilson, and J. A. Hawkins. 1973. A devise for self-marking of Tabanidae. Environ. Entomol. 2:960-961.

94
Sheppard, C., B. H. Wilson, B. R. Farthing. 1980. Flight routs of Tabanidae in a Louisiana bottomland hardwood forest. Environ. Entomol. 2(5):489-491.
Steelman, C. D. 1976. Effects of external and internal arthropods on domestic livestock production. Ann. Rev. Entomol. 21 : 155-178.
Steyskal, G. C. 1981. A bibliography of the Malaise trap. Proc. Entomol. Soc. Wash. 83(2):225-229.
Stone, A. 1930. The bionomics of some Tabanidae (Diptera). Ann. Entomol. Soc. Am. 23:261-274.
Tashiro, H., and H. H. Schwardt. 1949. Biology of the major species of horse flies of central New York. J. Econ. Entomol. 42(2) :269-272.
•Tashiro, H., and H. H. Schwardt. 1953. Biological studies of horse flies in New York. J. Econ. Entomol. 46(5):8l3-822.
Teskey, H. J. 1969. Larvae and pupae of some eastern North American Tabanidae (Diptera). Mem. Entomol. Soc. Can. 63:1-147.
Thompson, P. H. 1969. Collecting methods for Tabanidae (Diptera). Ann. Entomol. Soc. Am. 62(1):50-57.
Thompson, P. H. 1973a. Tabanidae (Diptera) of Texas, I. Costal marsh species. West Galvaston Bay; Incidence, frequency, abundance and seasonal distribution. Proc. Entomol. Soc. Wash. 75(3) : 359-364.
Thompson, P. H. 1973b. Tabanidae (Diptera) of Texas, II. Pine Belt species, Huntsville State Park; Incidence, frequency, abundance and seasonal distribution. Proc. Entomol. Soc. Wash. 75(4) : 430-435.
Thompson, P. H. 1974a. Tabanidae (Diptera) of Texas, III. Post Oak Belt species, Navasota River watershed; Incidence, frequency, abundance and seasonal distribution. Proc. Entomol. Soc. Wash. 76(1):35-38.
Thompson, P. H. 1974b. Tabanidae (Diptera) of Texas, IV. Pine Belt species, the big thicket; Incidence, frequency, abundance and seasonal distribution. Proc. Entomol. Soc. Wash. 76(3) : 315-321 .

95
Thompson, P. H. 1976. Tabanidae (Diptera) of Texas, V. second year collections from the Huntsville State Park and the Navasota River Watershed. Proc. Entomol. Soc. Wash. 78(3):376-381 .
Thompson, P. H. 1978. Parasitism of adult Tabanus sub-similis subsimilis Bellardi (Diptera: TabanidaeTTy a
' multigrammine sarcophagid (Diptera: Sarcophagidae). Proc. Entomol. Soc. Wash. 80(1):67-74.
Thompson, P. H., R. R. Blume, and A. Aga. 1977. Tabanidae (Diptera) of Texas, VI. Costal Prairie species, Victoria County. Proc. Entomol. Soc. Wash. 79(2):266-269 .
Thompson, P. H., and C. V. Love. 1979. Potential of a multigrammine sarcophagid in biological control of tabanids. Southwest. Entomol. 4(4) :298-303.
Thompson, P. H., S. M. Meola, and J. M. Thompson. 1980. Dead-end parasitism of bombyliid larvae in tabanid adults. Southwest. Entomol. 5(1):12-15.
Thornhill, A. R., and K. L. Hays. 1972. Dispersal and flight activities of some species of Tabanus (Diptera: Tabanidae). Environ. Entomol. 1 :602-606.
Thorsteinson, A. J., G. K. Bracken, and W. Hanec. 1964. The Manitoba horse fly trap. Can. Entomol. 96:166.
Tidwell, M. A., W. D. Dean, G. P. Combs, D. W. Anderson, W. 0. Cowart, and R. C. Axtell. 1972. Transmission of hog cholera virus by horseflies (Tabanidae: Diptera). Am. J. Vet. Res. 33(3):6l5-622.
Turner, E. C , and R. R. Gerhardt. 1965. A material for rapid marking of face flies for dispersal studies. J. Econ. Entomol. 58:584-585.
U.S. Environmental Data Service. 1981a. Climatological Data, Annual Summary: Texas. Ashville, N.C., National Climatic Center. 86(13) 32 pp.
U.S. Environmental Data Service. 1981b. Climatological Data: Texas. Ashville, N.C., National Climatic Center 87(1-12) 781 pp.
Web J. L., and R. W. Wells. 1924. Horseflies: biologies ' and relation to western agricultures. U.S.D.A. Bull. No. 1218. 36 pp.

96
Wilson, B. H. 1969. Tabanid larval habitats and population densities in an Alluvial area in southern Louisiana. Ann. Entomol. Soc. Am. 62(5) : 1203-1204.
Wilson, B. H., E. C. Burns, W. T. Oglesby, R. B. Meyer, T. E. Rogers, and J. Wimblery. 1963. The effect of horse fly control on rate of infection of bovine anaplasmosis under field conditions in Louisiana. J. Econ. Entomol. 56:578-579.
Wilson, B. H., and R. B. Meyer. 1966. Transmission studies of bovine anaplasmosis with horseflies, Tabanus fasci-costatus and Tabanus nigrovittatus. Am. J. Vet. Res. 27(114) :367-3^T:
Wilson, N. L., J. E. Huston, and D. I. Davis. 1978. Effectiveness of stirofos impregnated ear tags for control of horn flies and horse flies on cattle in central Texas. Southwest. Vet. ^1(^ ) : 1Q7-19Q . 01 norn iiies ana norse iiies on cauLie Texas. Southwest. Vet. 31(3) : 197-199 .