by tessa gordon* and r. b. stein from the department of
TRANSCRIPT

J. Physiol. (1982), 323, pp. 307-323 307With 8 text-fturesPrinted in Great Britain
TIME COURSE AND EXTENT OF RECOVERY IN REINNERVATEDMOTOR UNITS OF CAT TRICEPS SURAE MUSCLES
BY TESSA GORDON* AND R. B. STEINFrom the Department of Physiology, University of Alberta, Edmonton,
Canada T6G 2H7
(Received 16 April 1981)
SUMMARY
1. Nerve and muscle properties were studied in single motor units of triceps suraemuscles in the cat using chronic recording techniques and intramuscular micro-stimulation. Recordings were made before and at intervals up to 18 months after anerve was sectioned and sutured either to its distal stump (nerve-nerve suture) or toa muscle directly (nerve-muscle suture). Thus, each nerve and muscle served as itsown control for recovery after reinnervation.
2. Following a delay all muscles recovered their preoperative tension afternerve-nerve suture with a single exponential having a time constant of 1-2 months.Only half the muscles recovered their preoperative tensions after nerve-musclesutures. Muscles which did not recover fully also had a slower time course of recovery.
3. The estimated number of motor units did not increase significantly later than2 months after nerve section and suture. Further recovery of muscle tension is dueto increased unit tension, rather than increasing numbers ofreinnervated motor units.Unit tension recovered completely in all muscles, but did not become enlarged, evenwhen muscles apparently remained partially denervated.
4. The latency ofcompound nerve potentials often recovered completely, althoughthe amplitude of the potential remained depressed. Single motor axonal potentialsrecovered to control levels after reinnervation of muscle with a time constant similarto that for the recovery of motor unit tension. Therefore, two distinct populationsofmotor axons contribute to the compound potential: reinnervating axons whose sizerecovers fully, and disconnected axons which remain atrophic. Incomplete recoveryof the compound potential amplitude mainly results from a failure of all axons toremake peripheral connexions.
5. Thus, formation of functional nerve-muscle connexions completely reverses theeffects of axotomy on nerve and muscle. Reinnervated motor units recovered theirpreoperative size, whether or not much of the muscle remained denervated.
* Present address and address for correspondence: Department of Pharmacology, University ofAlberta, Edmonton, Alberta, Canada T6G 2H7.

T. GORDON AND R. B. STEIN
INTRODUCTION
Axons in the central stump of severed peripheral nerves atrophy (Gutmann& Sanders, 1943) with consequent reduction in conduction velocity (Cragg &Thomas, 1961). The extent of atrophy is not related to streaming of axoplasm intoregenerating axons since axons atrophy similarly when ligated to prevent regenerationor surgically repaired to encourage regeneration (Davis, Gordon, Hoffer, Jhamandas& Stein, 1978). Recovery after nerve section and repair is never as complete as aftera simple crush injury, since only a fraction of the fibres regenerate into the distalstump across the scar tissue of the suture line (Gutmann & Sanders, 1943). Thelatency of the compound action potential recorded proximal to suture line usuallyrecovers to preoperative values, but the amplitude never reaches control levels (Daviset al. 1978).Formation of peripheral connexions is the most important factor for maturation
of regenerating fibres (Weiss & Taylor, 1944; Sanders & Young, 1946) independent ofthe target of regeneration (Aitken, Sharman & Young, 1947). Thus, incompleterecovery of the compound action potential amplitude is consistent with regenerationof insufficient numbers of large fibres to restore the original bimodal fibre distributionin hind limb nerves (Gutmann & Sanders, 1943). Since recovery of muscle tensionis often complete when the compound potential remains depressed, either a similarfraction of sensory and motor fibres successfully reinnervate muscle (and thesuccessful motor axons enlarge their field to innervate all muscle fibres) or most motoraxons are successful in reinnervation even when sensory reinnervation is far fromcomplete. Sensory fibres would then be differentially affected in regeneration as theyare during atrophy (Hoffer, Stein & Gordon, 1979; Gordon, Hoffer, Jhamandas &Stein, 1980).To decide between these alternatives and to correlate recovery of nerve and muscle
more directly, wesampled the population ofmotoraxonsthat successfully reinnervatedmuscle fibres using a modified microstimulation method (Stephens & Taylor, 1975;Taylor & Stevens, 1976; Garnett, O'Donovan, Stephens & Taylor, 1979). For eachregenerating nerve fibre that was stimulated intramuscularly, we recorded (1) evokedunit potentials with implanted chronic recording electrodes on the central stump ofthe resutured nerves, (2) associated unit electromyographic activity and (3) twitchtension. Our results indicate that nerve and muscle fibres recover completely iffunctional connexions are remade and that there are two separate populations ofnerve and muscle fibres after regeneration. Reinnervated fibres recover preoperativelevels, while fibres that do not form nerve-muscle connexions remain atrophic.Greatly enlarged motor units, as suggested by electromyographic studies in man(Yahr, Herz, Moldaver & Grundfest, 1950; Rosenfalck & Buchthal, 1970), were notobserved in the present experiments when muscles recovered their preoperativetension and the compound nerve potentials remained below control values. Incom-plete recovery of compound potential amplitude in these nerves must be largely dueto poorer recovery of sensory fibres than motor fibres. Motor units did not becomeenlarged even when relatively few motor nerves were -successful in growing to themuscle. In that case, many muscles remained denervated. A brief account of someof the results has been presented elsewhere (Gordon & Stein, 1980).
308

RECOVERY OF REINNER VATED MOTOR UNITS
METHODS
The methods for constructing and implanting chronic recording electrodes enclosed within silasticcuffs have been described previously (Stein, Nichols, Jhamandas, Davis & Charles, 1977). Silasticcuffs containing platinum-iridium electrodes were implanted in ten cats under aseptic conditionsaround the sciatic nerve and one or more of its branches, to be used for stimulation and recording.Measurements were made of the maximum compound evoked potential under Fluothane anaesthe-sia, as shown diagramatically in Fig. 1. Either the lateral gastrocnemius soleus (LGS) or the medial
Triceps suraeSciatic CPS
LGS Tens~~i~onE.m.g.
Fig. 1. Schematic diagram of chronic stimulation and recording of compound and unitaction potentials on the sciatic nerve and the nerve branches supplying the lateralgastrocnemius soleus (LGS) and the medial gastrocnemius (MG) muscles which form partof the triceps sure group of ankle extensor muscles. Tension and e.m.g. were recordedsimultaneously in response to either (1) stimulation of the whole muscle nerve via theelectrodes contained in the silastic cuffs around theMG andLGS nerves or (2) intramuscularmicrostimulation of single motor units with a needle electrode (see text for further details).MG and LGS nerves were cut distal to the eilastic cuffs.
gastrocnemius (MG) nerve was stimulated maximally and a compound action potential wasrecorded on the sciatic nerve simultaneously with the electromyographic activity (e.m.g.) and thetwitch contraction of the corresponding muscle. The methods for recording e.m.g. and isometriccontraction of the muscle have been described in detail previously (Davis et al. 1978). Briefly, thee.m.g. was recorded from the surface ofthe muscle by implanted electrodes attached to a flat Silasticsheet. The torque of ankle extension (also referred to as plantar flexion) was recorded by placingthe cat's foot into a specially designed boot. The boot was free to rotate about a line coaxial withthe ankle joint and was connected to a tension transducer (either Grass FT03 or FT1O) located4 cm from the axis of rotation. The force directly measured (N) is given here, and should bemultiplied by 004 to yield torque in Nm. The leg was stabilized at two other points by placingit between two rods and fixing a short metal plate down over the lateral or medial surface of thelower leg. Since the animals were studied chronically, care was taken not to occlude the blood supplyor damage the skin. Finally, the compound evoked potential was recorded on the muscle nerve whenthe whole sciatic nerve was stimulated.To measure the action potentials of single axons and their associated unit twitch tension and
e.m.g. response, we modified the intramuscular microstimulation introduced by Stephens & Taylor(1975). A bipolar stimulating electrode was contstructed to confine the spread of current to veryshort distances. A fine Ag wire, which was coated with insulating Teflon except for the tip, wasinserted into the barrel of a 26-gauge needle and fixed with epoxy resin. The electrode was insertedpercutaneously into the muscle belly near the location of the motor point. The current intensitywas increased to levels high enough to evoke single neural spikes, associated with all-or-none e.m.g.
309

T. GORDON AND R. B. STEINand twitch responses. A large number of motor units could be sampled by adjusting the positionof the needle and the intensity of the stimulus voltage. The duration of the stimulation was alwaysless than 20 ssec to avoid direct stimulation of muscle. The stimulus intensity was adjusted andthe all-or-none firing of single motor units carefully checked by monitoring the neural as well asthe unitary e.m.g. responses.
Unit twitch tensions were reliably recorded, but movement of the needle as a result of maximaltetanic forces often precluded the recording of unit tetanic tension. However, since the twitchtension of single units was linearly related to the tetanic tension (T. Gordon & R. B. Stein,unpublished observations) the measurement of twitch tension provided an adequate measure ofthe motor unit size. Contraction speed was assessed by the contraction time of the units recordedas the time from stimulation to peak tension. Since unit responses were small relative to evokedresponses of whole nerve and muscle, between twenty-five and forty responses were averaged usinga Digital Equipment Co. 11/34 laboratory computer.
Control data were obtained after implantation of recording devices. The LGS or MG nerves werecut distal to the recording electrodes and sutured either to the distal stump of the nerve (nerve-nervesuture) or to the muscle directly (nerve-muscle suture). Single units were studied in six cats forperiods of 100-570 days after nerve repair. Voltage was recorded differentially on the nerve acrossthe impedance of the electrodes in the silastic cuff. The triphasic electrode configuration (Stein et al.1977) was coupled by a step-up transformer with an approximate gain of 15 (Hammond 585D)to a low noise preamplifier (Grass P-15). The transformer provided the valuable improvement ofsignal-to-noise ratio essential for recording single unit neural potentials. Voltage was converted firstinto current by dividing by the impedance of the electrodes and then normalized by the averageimpedance of all the recording cuffs used (3 kil) to give calibrated neural signals in units ofmicrovolts.
RESULTS
Recovery of muscle tensionGutmann & Sanders (1943) noted that the number of axons which regenerated
across the suture line of cut and sutured nerves was extremely variable, and axonnumbers in the distal nerve stump was always less than that in the central stump,even a year after suture. Yet we found that recovery of reinnervated muscle tensionwas surprisingly good in response to stimulation of the nerve proximal to the suturewhether the cut nerve was sutured to its distal stump (nerve-nerve suture) or directlyto a muscle (nerve-muscle suture). All muscles recovered preoperative tension afternerve-nerve sutures (mean = 105%, range 93-113 %). Three out of six musclestotally recovered after nerve-muscle suture (mean = 110 %), while the other threerecovered only 15-50% of control tension.The time course of recovery was related to the success of reinnervation, since
muscles that recovered more completely also recovered more quickly. Recovery oftwitch and maximal tetanic tension followed a simple exponential, after a delay asshown for two medial gastrocnemius muscles in Fig. 2. In Fig. 2A, a MG musclerecovered 96% of control tension after nerve-nerve suture as compared to 50% foranother MG muscle after nerve-muscle suture (Fig. 2 B). The recovery curves appearsimilar, but the time scales differ by a factor of two in the two parts of the Figure.Thus, the time course of recovery of the MG muscle after nerve-muscle suture wasmore than twice as long as that after nerve-nerve suture. In seven determinationsof recovery curves for muscles that regained more than 90% of preoperative tensionlevels, the time constant of recovery was 35 + 4 days (mean+s.E.). This compareswith 117+17 days for three muscles which recovered less than 50% of their
310
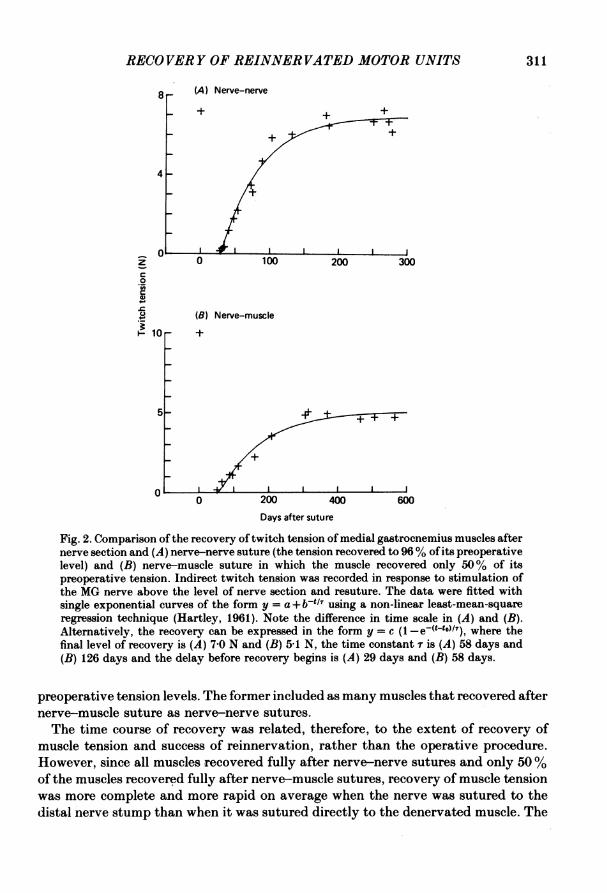
RECOVERY OF REINNERVATED MOTOR UNITS 311
8 (A) Nerve-nerve
8-4-
01o 100 200 300
C0
(B) Nerve-muscle
1-10 -1+
5- +~~~ +
o 200 400 600
Days after suture
Fig. 2. Comparison of the recovery oftwitch tension of medial gastrocnemius muscles afternerve section and (A) nerve-nerve suture (the tension recovered to 96% of its preoperativelevel) and (B) nerve-muscle suture in which the muscle recovered only 50% of itspreoperative tension. Indirect twitch tension was recorded in response to stimulation ofthe MG nerve above the level of nerve section and resuture. The data were fitted withsingle exponential curves of the form y = a+ b-ti? using a non-linear least-mean-squareregression technique (Hartley, 1961). Note the difference in time scale in (A) and (B).Alternatively, the recovery can be expressed in the form y = c (1 -e(t-to)/7), where thefinal level of recovery is (A) 7 0 N and (B) 5-1 N, the time constant r is (A) 58 days and(B) 126 days and the delay before recovery begins is (A) 29 days and (B) 58 days.
preoperative tension levels. The former included as many muscles that recovered afternerve-muscle suture as nerve-nerve sutures.The time course of recovery was related, therefore, to the extent of recovery of
muscle tension and success of reinnervation, rather than the operative procedure.However, since all muscles recovered fully after nerve-nerve sutures and only 50%of the muscles recovered fully after nerve-muscle sutures, recovery of muscle tensionwas more complete and more rapid on average when the nerve was sutured to thedistal nerve stump than when it was sutured directly to the denervated muscle. The

T. GORDON AND R. B. STEINfastest time course of recovery of tension after nerve suture was similar to after acrush injury (Stein, Gordon, Hoffer, Davis & Charles, 1980) and recovery of muscletension was considerably more rapid than recovery of compound neural potentials(102 + 11 days for ten nerves; Davis et al. 1978) as will be considered further below.
Motor unit tensionThe recovery of tension from the whole muscle and from single motor units was
compared in six of the ten cats and the results are shown in Table 1. When musclesfirst became reinnervated, motor units were significantly smaller in terms of their
TABLE 1. Recovery of whole muscle tension (six cats) compared to recovery of unit twitch tensionsin the same animals. Early reinnervation represents the period 2-3 months and late reinnervationrepresents the period 5-18 months after the nerve was severed. Means+S.E. of means are shown.
Reinnervated Musclerecovery
Preoperative Early Late (%)Twitch (N) 7-4+1-7 1-7+0-8 55+30 79Tetanus (N) 33-9+6-2 7-8+2-5 24-4+ 12-3 72Unit tension (mN) 36-3 + 30 126 + 09 433 + 2-6 119Total no. of units studied 322 215 598Unit tension (% ofpreoperative twitch tension) 0-49 0-17 0-585 119Unit tension (% ofreinnervated muscle tension) 0 49 0 74 0-79 161
No. of units per muscle 204 135 127 62
twitch tension than control preoperative units. Motor unit tension recovered inparallel with the tension developed by the whole muscle in response to nervestimulation. The time course of the recovery of muscle tension and unit tension wasstrikingly similar, as illustrated in Fig. 3.
In all six muscles studied, the mean twitch tension of motor units recovered to orslightly above control preoperative levels, even when the whole muscle did not fullyrecover its preoperative value. In three cats, where muscle tension recovered less than50% of preoperative tension levels, mean (± s.E. of mean) tension of reinnervatedunits was 48-8+ 3-8 compared to 39-8 + 4-7 mN for units prior to nerve section in thesame muscles. These values are not statistically significatnt different from oneanother or from the values for all six cats given in Table 1. Unit tension expressedas a percentage of reinnervated muscle tension is enlarged (Table 1), as reportedpreviously (Bagust & Lewis, 1974), but the absolute unit tension after reinnervationor the unit twitch tension as a percentage of the preoperative twitch tension was notsignificantly different from preoperative values.The number of units was estimated during early stages of reinnervation when the
motor units had not fully recovered and at later times when reinnervated motor unittensions had fully recovered ('late reinnervation' in the Table and following Figures).The number of reinnervated motor units estimated for each cat from the ratio ofmuscle tension to motor unit tension during early stages of reinnervation, was notdifferent from later stages when muscles had recovered maximally (Table 1),
312

RECOVERY OF REINNER VATED MOTOR UNITS
.2c
H
iC
4.0
._2
c
wH
c0_T
8
4.
313
(A) Muscle twitch
_ + .9- +
(B) Muscle tetanus40 - x X_
20 -x
60 (C) Motor unit twitch
30-
0 100 200 300
Days after suture
Fig. 3. Recovery of (A) twitch tension, (B) tetanic tension and (C) unit twitch tension aftersection of the LGS nerve and suture of the central to the distal stump of the nerve. Eachpoint in (C) represents the mean + s.E. of twenty-one to forty-two single motor units. Asimple exponential was fitted to the raw data as in Fig. 2. Note the similar time coursefor recovery of the muscle and motor unit tension (r = (A) 37, (B) 36 and (C) 27 days).The delay before recovery began was approximately 35 days in all parts of the Figure.
consistent with parallel recovery ofmotor unit tension and muscle tension. Therefore,the extent of recovery of the whole muscle mainly depends on the number of nervefibres that grow to the muscle and successfully make nerve-muscle connexions. Sincemuscles tend to recover more fully after nerve-nerve suture than suture of nervedirectly to muscles, the chances of regenerating nerve fibres finding their way backto muscle and forming functional connexions are higher after the former than thelatter surgical procedure.
0

T. GORDON AND R. B. STEIN
I(A) Control
201
10
T (B) Early reinnervationI
T T
I
T
I
T ,T.1.I I i I II II I I I L -1 1 I I I I
10 100
Twitch tension (mN)
Fig. 4. Logarithmic distribution of unit twitch tension of single motor units of six cats.Thirty to one hundred and seventy units were sampled from each of six muscles (A)preoperatively (control) and after reinnervation, (B) 2-3 months after nerve and repair(early reinnervation) and (C) 5-18 months after surgical repair (late reinnervation).Standard error bars are shown.
The distribution of unit tensions was highly skewed, as noted previously byWuerker, McPhedran & Henneman (1965) and by Milner-Brown, Stein & Yemm(1973). Itbecomes more symmetric when the tension axis is divided into approximatelyequally spaced bins on logarithmic co-ordinates, as shown in Fig. 4. The mean of thisdistribution (equivalent to the geometric mean of the original distribution) is thenmore equal to the mode. After reinnervation, changes occur both in the mean andthe variance of this distribution. Initially, the mean tension was smaller, as was therange of tensions (Fig. 4B). Consequently, the histogram of unit tension, plotted as
40
301CaR
0-
.0
Ez
201-
101
v . .HU
314
n]

RECOVERY OF REINNER VATED MOTOR UNITS 315in Fig. 4 or as a cumulative histogram in Fig. 5, is shifted left along the tension axisto smaller values. The cumulative distribution gives the fraction of units whichdevelops tension less than or equal to the value indicated. For example, 50% of theearly reinnervated units develop 8 mN or less, compared to about 16 mN forpreoperative units. A cumulative distribution with a logarithmic scale is useful sincethe distribution will be shifted in parallel to the left if all units were reduced to a
100
80 -
20 -CL0
40
E 40
+ Early reinnervation
20gF -E Late reinnervation
C1 10 100 1000
Twitch tension (mN)
Fig. 5. Change in the cumulative probability distribution of single unit tension duringreinnervation. Single motor unit tension is plotted as a cumulative probability distributionto compare distributions of preoperative and reinnervated motor units. Note that thedistribution is shifted to the left along the X-axis during early reinnervation to smallertension values but the motor units recover their preoperative tension and the distributionof the fully recovered reinnervated motor units overlaps the distribution for thepopulation of units studied preoperatively.
constant fraction of their preoperative value. The cumulative tension distribution forearly reinnervated units in Fig. 5 is clearly not shifted to the left in parallel sincethe lower limit of tension was not changed.
After reinnervation, the tensions recovered and the cumulative distributionsshifted back to overlap the control curve. The differences between the distributionsin Fig. 5 were tested for significance by applying the Kolmogorov-Smirnov test(Fisz, 1963). The two distributions for control and late reinnervation were notsignificantly different, but were significantly different from that forearly reinnervation(P < 00001). Although the cumulative distribution of recovered units was notsignificantly different from the preoperative distribution, the variance was somewhathigher (compare Fig. 4 parts A and C). The reasons for the change in shape of thetension distribution will be considered in the Discussion.

316 T. GORDON AND R. B. STEIN
0-7 (A) Compound AP latency
0-6 -
E
0-4
0-4 [ I I I' '
(B) Compound AP amplitude0-6 -
E0-4
0-2 _
(C) Axon AP amplitude2-4
X x
2-0
1-6
0 100 200 300 400
Days after suture
Fig. 6. Change in action potential (AP) latency (A) and amplitude for whole nerves (B)and for single axons (C). A and B, sciatic nerve was stimulated and compound potentialsrecorded on the MG and LGS nerves proximal to the site of section and suture. In C, singleaxons were stimulated intra-muscularly and unitary potentials were recorded on the MGand LGS nerves proximal to level of section.
Recovery of resutured nervesFig. 6 shows changes in the latency and amplitude ofcompound potentials recorded
on the central stump of the cut and sutured nerves. The points are the geometricmeans of four nerves that were studied for a period of at least 400 days after nervesuture. Initially, the latency increased and the amplitude declined after nerve section,as occurs when nerves are cut and ligated to prevent regeneration (Davis et al. 1978).The latency began to decrease 40-60 days after the nerves had been cut and shortlyafter the time when tension could first be measured in reinnervated muscle. As shown

RECOVERY OF REINNER VATED MOTOR UNITS 317previously, the decline in amplitude of the potential also halted some time afterreinnervation, but the amplitude of the compound potential did not recover much,even 400 days after nerve repair.That the time to the first peak of the compound potential recovered, indicates that
some axons do fully recover after nerve-muscle connexions are made. The compoundamplitude may not recover to the same extent because of the wide dispersion oflatencies of axons that have and have not been successful in remaking peripheralconnexions. The recovery of potentials in single motor axons that did reinnervatemuscles was much better than the recovery of the compound potentials, as shownin Fig. 6C. When nerves first reinnervate muscle fibres, the mean axon potentialsare significantly smaller than preoperative values, but within 1-2 months thepotentials recover toward preoperative levels. The axon potentials have recoveredto control levels by 4 months after nerve repair with a time constant of 37 days whenthe compound potential is just beginning to increase in size, and the latency of thepotential has not yet recovered preoperative levels. The latency of the compoundpotentials did return toward control values after more than a year, but the amplitudeof the compound potential never fully recovered.The complete recovery of axons that successfully remade connexions shows that
the discrepancy between recovery of the amplitude and the latency of the compoundpotentials may be accounted for by the recovery ofaxons that have remade peripheralconnexions, with continuing atrophy in axons that do not remake peripheralconnexions to a lower stable level (Davis et al. 1978; Gordon et al. 1980). Differencesbetween sensory and motor fibres may also be responsible, since sensory fibres aremore severely affected by axotomy and recovery of activity in peripheral nervesduring locomotion was always less complete in sensory fibres (Gordon et al. 1980).We estimated the degree of atrophy of axotomized sensory and motor fibres bymeasuring the total charge delivered to the dorsal and ventral roots respectively whenstimulating the central stump of the cut peripheral nerve (See Hoffer et al. 1979 formethod) in final acute experiments. The ratio of the charge delivered to the dorsal(DRC) and ventral (VRC) roots by stimulating the LGS nerve is 0-38 for controlnerves. We previously found that this ratio fell to 0 100 or less 300 days after nervesection and ligation showing that sensory axons were more severely affected byaxotomy than motor fibres (Hoffer et al. 1979). For LGS muscles which recoveredless than 50% of control tension, the root ratio of DRC to VRC was between 0-042and 0 055, 500 days or more after nerve section and nerve-muscle suture, consistentwith greater atrophy of sensory fibres isolated from the periphery than motor fibreswithout nerve-muscle connexions. Where more than 90 % of control preoperativetension levels returned, the root ratio returned toward control levels 300 days afternerve-nerve suture, indicating that sensory axons that reinnervate peripheralend-organs recover as well as motor axons which reinnervate muscle.The mean of the amplitude of axon potentials recorded chronically in reinnervated
motor units did not change significantly more than 100 days following nerve suture.That data have been grouped together and compared with neural potentials recordedpreoperatively (Fig. 7A) and during early reinnervation (Fig. 7B). Note that theamplitudes of axon potentials are more symmetrically distributed than the unittensions. The distribution is unimodal since the units were sampled with intramuscular

318 T. GORDON AND R. B. STEINmicro-stimulation and included only axons with nerve-muscle connexions. Theamplitudes ofpotentials from newly reinnervated axons are similarly distributed, butthe distribution is shifted toward smaller values. When the data are plotted as acumulative distribution on logarithmic coordinates, a parallel shift toward smaller
30 (
(A) Control
201-
10
40F
40
c
0.
301-
20
10
ol40fE- (C) Late reinnervation
30 _
201-
10_
O03
Axon AP amplitude (pV)
Fig. 7. Distribution of axon potential amplitude of single motor units of six cats of (A)preoperative control units, (B) units during early reinnervation (2-3 months after nervesection and repair) and (C) units during late reinnervation (5-18 months post-suture).Thirty to 170 units were sampled from the same muscles before, early and late afterreinnervation. Standard error bars are shown.
potentials is seen (Fig. 8). All potentials are reduced 30-40% and the differencebetween the two distributions is statistically different at the 0-01 % level.Axon potentials of motor axons which did not grow across the suture line and
remake nerve-muscle connexions, were recorded proximal to the suture in final acuteexperiments on the same cats that were studied chronically, in response to stimulationof teased ventral roots. Stimulation of these units elicited no muscle tension. Their

RECOVERY OF REINNER VATED MOTOR UNITS 319cumulative frequency distribution is plotted and compared with reinnervating unitsin Fig. 8. The units with no peripheral connexions, shown as A in the Figure, weresignificantly smaller than control and fully recovered reinnervated units. Althoughthe curve for these units is slightly left of the early reinnervated motor units on thex-axis, the two distributions were not significantly different. This shows that duringthe first 2-3 months when reinnervating axons are remaking nerve-muscle connexions,
100
80
0
60
E 40 / No reinnervationX Control+ Early reinnervation
20 _1 Late reinnervation
0 1.0 2-0 5-0 1-0Axon AP amplitude (gV)
Fig. 8. Change in the cumulative probability distribution of single axonal potentialamplitudes during the course ofreinnervation are compared with distribution ofpotentialsrecorded from axons that did not remake nerve-muscle connexions in the same animals(all potentials were recorded proximal to the site ofnerve section and suture). During earlyreinnervation (2-3 months after axotomy), the distribution of axon potentials (n = 58)is shifted to smaller values which were not significantly different from values for axonswithout reinnervation 300-570 days after axotomy (n = 157). The distribution of re-innervated units moves back along the X-axis with time to resemble the distribution forthe same units sampled preoperatively.
their axons had atrophied almost as much as axons which remained withoutconnexions for 300-570 days. This is consistent with rapid axonal atrophy immediatelyafter axotomy. The decay of axonal amplitude follows exponential time course witha time constant of 1-2 months (Davis et al. 1978). Thus, when all motor axons aresampled in resutured nerves proximal to the suture line, the distribution of axonalpotential amplitude is bimodal with clearly separated peaks for axons that have andhave not remade peripheral connexions.With time, the axons recover in diameter and the size and distribution of their
potentials are shifted again to larger values. The cumulative histogram of recoveredmotor units overlaps andwas not statistically different from the curve for preoperativeaxons, even at the points of maximum diversion. These findings suggests that alla-motoneurones (1) atrophy to the same extent after axotomy which is consistent

320 T. GORDON AND R. B. STEINwith the recent findings of Milner & Stein (1981), (2) recover in parallel when theymake functional nerve-muscle connexions and that (3) formation of nerve-muscleconnexions is able to completely reverse the attrophic changes in the central stumpof the nerve resulting from axotomy.
DISCUSSION
Our results show conclusively for the first time that atrophic changes occurring in-the proximal stump of cut and sutured motor nerves and in denervated muscle fibresare totally reversed when the nerve axons successfully reinnervate skeletal muscle.Unsuccessful axons continue to atrophy and reach the same final level as axons inligated nerves which were prevented from regenerating. Previous methods utilizingcompound potentials or histological cross-sections could not distinguish axons thatmade peripheral connexions from those that had not.
Since thenumberofmotorunitsremained constantduringthe period ofreinnervationfollowing 1-3 months after suture, recovery-of motor axons from this point is directlyrelated to increase in diameter of axons with nerve-muscle connexions. Gutmann &Sanders (1943) found more large axons histologically 1 year after nerve suture thanat 200 days. This continuing reconstitution of the fibre diameter spectrum may bedue to later arrivals of sensory axons. Brown & Butler (1976) observed that axonsof muscle spindle afferents had reduced conduction velocities 6-8 months afterreinnervation, whereas y-motor axons had normal conduction velocities. In fact, priormotor innervation may be important for the complete recovery of spindles (Ip,Vrbova' & Westbury, 1977).
Shortly following reinnervation, motor unit tensions were significantly less thancontrol values, and the range was markedly reduced. The smaller tension ispresumably a consequence of denervation atrophy of muscle fibres (Brown-Sequard,1853; Sunderland, 1978). The reduced range probably results from two causes. Thelarge differences in contractile speed and size between fast and slow muscles arereduced after denervation (Lewis, 1972; Syrovy, Gutmann & Melichnat, 1972). Inaddition, motor units in early reinnervated muscle are probably heterogeneous intheir histochemical properties and not homogeneous as in normal units (Nemeth,Pette & Vrbova', 1980). Such a mixing would result in units of intermediate propertiesand a reduced range of tensions.
Alternatively, we might have missed a number of very small units with our chronictechniques, but there are two arguments against this explanation for the reducedrange of tensions. First, the distribution of motor axons sizes was uniformly shiftedtoward smaller diameters, so that there did not seem to be any bias against samplingsmall axons. Secondly, in final acute experiments (T. Gordon & R. B. Stein, unpub-lished observations) on some of the same animals studied chronically, ventral rootfilaments were dissected and single unit properties were studied in a more conventionalfashion. Comparison of results from chronic and acute studies indicates that ifanything, large units were missed out in the chronic recordings, because the largetwitch tensions generated tended to dislodge the electrode used for microstimulationand prevent reliable measurements.With time after reinnervation the level and distribution of tensions produced by

RECOVERY OF REINNER VATED MOTOR UNITSsingle motor units returned to control values, but the variance was increasedsomewhat. The increase in range and variance compared to early reinnervationpresumably results from motoneurones respecifying muscle properties, so that eachmotor unit is again composed of muscle fibres which are homogeneous in size (Gordon& Stein, 1980) and histochemistry. Instead ofthe mosaic pattern seen histochemicallyin normal muscle, there tends to be a block of fibres after reinnervation which allstain similarly (Kugelberg, Edstrom & Abbruzzese, 1970). Randomness in thereinnervation of these blocks of fibres may also contribute to the increased varianceof motor unit sizes.The twitch tensions of reinnervated motor units never became significantly larger
than control values, even when the tension in the whole muscle remained quitedepressed. Bagust & Lewis (1974) found enlarged motor units after reinnervation ofthe flexor digitorum longus muscle in the cat. However, they expressed mean motorunit tension as a fraction of whole muscle tension. If whole muscle tensions werereduced after reinnervation in their study, motor unit size could be increased relativeto the smaller over-all tension, but not in absolute units. E.m.g. studies in humansalso suggest an increased size of motor units after regeneration (Buchthal &Schmalbruch, 1980), but the increased e.m.g. could result from the block pattern ofreinnervation. In one of the few human studies where both e.m.g. and tension wereassessed after reinnervation, Milner-Brown, Stein & Lee (1974) found greatlyincreased e.m.g. signals with little or no increase in twitch tensions.We do not deny that motor units can become increased in tension in disease states
such as poliomyelitis or in partial denervation produced experimentally. However,there are significant differences in the time and distances involved in these latterconditions. If a motoneurone dies as a result of a disease process, while otherssupplying adjacent muscle fibres remain healthy, reinnervation can take placeimmediately by the formation of sprouts which would only have to grow a fractionof a millimetre. In this way, the twitch tension of motor units would be increasedabove normal levels in polio' patients or in other forms of motoneurone disease(Milner-Brown et al. 1974). Similarly, after partial denervation in experimentalanimals, sprouting motoneurones can increase their peripheral field of innervationat least 5 times (Brown & Ironton, 1978).With the physical barriers to regeneration, it is remarkable that motor axons reach
and modify enough muscle fibres that the distributions of motor unit tension becomestatistically indistinguishable from control values within a few months of nerveinjury. Rather than motor unit size increasing substantially under these conditions,the residual muscle atrophy indicates to a first approximation the fraction of axonsthat has been unsuccessful in reinnervating the muscle. With good surgical tech-niques, early repair of a sectioned muscle nerve in experimental animals does leadto complete restoration of muscle tension and even the detailed organization of motorunit types (Gordon & Stein, 1980).
This research was supported by grants from the Canadian Medical Research Council, MuscularDystrophy Association of Canada, and Alberta Heritage Foundation for Medical Research. We aregrateful to Dr K. Pearson for his comments on the manuscript and to Dr L. A. Davis for surgicalhelp.
11 PHY 323
321

T. GORDON AND R. B. STEIN
REFERENCES
AITKEN, J. T., SHARMAN, M. & YOUNG, J. Z. (1947). Maturation of regenerating nerve fibres withvarious peripheral connections. J. Anat. 81, 1-22.
BAGUST, J. & LEWIS, D. M. (1974). Isometric contractions of motor units in self-reinnervated fastand slow twitch muscles of the cat. J. Physiol. 237, 91-102.
BROWN, M. C. & BUTLER, R. G. (1976). Regeneration of afferent and efferent fibres to musclespindles after nerve injury in adult cats. J. Physiol. 260, 253-266.
BROWN, M. C. & IRONTON, R. (1978). Sprouting and regression of neuromuscular synapses inpartially denervated mammalian muscles. J. Physiol. 278, 325-348.
BROWN-SEQUARD, H. (1853). Experimental Researches Applied to Physiology and Pathology. NewYork: H. Bailliers Press.
BUCHTHAL, F. & SCHMALBRUCH, H. (1980). Motor units of mammalian muscle. Physiol. Rev. 60,90-142.
CRAGG, G. B. & THOMAS, P. K. (1961). Changes in conduction velocity and fibre size proximal toperipheral nerve lessons. J. Physiol. 157, 315-327.
DAVIS, L. A., GORDON, T., HOFFER, J. A., JHAMANDAS, J. & STEIN, R. B. (1978). Compound actionpotentials recorded from mammalian nerves following ligation or resuturing. J. Physiol. 285,543-559.
FISz, M. (1963). Probability Theory and Mathematical Statistics. New York: Wiley.GARNETT, R. A. F., O'DONOVAN, M. J., STEPHENS, J. A. & TAYLOR, A. (1979). Motor unit
organization of human medial gastrocnemius. J. Physiol. 287, 33-43,GORDON, T., HOFFER, J. A., JHAMANDAS, J. & STEIN, R. B. (1980). Long-term effects of axotomy
on neural activity during cat locomotion. J. Physiol. 303, 243-263.GORDON, T. & STEIN, R. B. (1980). Rematching of nerve and muscle properties in reinnervated catmotor units. In Plasticity of Muscle, ed. PETTE, D., pp. 283-296. Berlin: de Gruyter.
GUTMANN, E. & SANDERS, F. K. (1943). Recovery of fibre number and diameter in the regenerationof peripheral nerves. J. Physiol. 101, 489-518.
HARTLEY, H. 0. (1961). The modified Gauss-Newton method for the fitting of non-linear regressionfunctions by least squares. Technometrics 3, 269-280.
HOFFER, J. A., STEIN, R. B. & GORDON, T. (1979). Differential atrophy of sensory and motor fibersfollowing section of cat peripheral nerves. Brain Res. 178, 347-361.
IP, M. C., VRBOVA, G. & WESTBURY, D. R. (1977). The sensory reinnervation of the cat followingdenervation and de-efferentation. Neuroscience 2, 423-434.
KUGELBERG, E., EDSTR6M, L. P. & ABBRUZZESE, M. (1970). Mapping of motor units in experi-mentally reinnervated rat muscle. J. Neurol. Neurosurg. Psychiat. 33, 319-329.
LEWIS, D. M. (1972). The effect of denervation of the mechanical and electrical responses of fastand slow mammalian twitch muscle. J. Physiol. 222, 51-75.
MILNER, T. E. & STEIN, R. B. (1981). The effects of axotomy on the conduction of action potentialsin peripheral sensory and motor nerve fibres. J. Neurol. Neurosurg. Psychiat. 44, 485-496.
MILNER-BROWN, H. S., STEIN, R. B. & YEMM, R. (1973). The contractile properties ofhuman motorunits during voluntary isometric contractions. J. Physiol. 228, 285-306.
MILNER-BROWN, H. S., STEIN, R. B. & LEE, R. G. (1974). The contractile and electrical propertiesofhuman motor units in neuropathies and motor neuron disease. J. Neurol. Neurosurg. Psychiat.37, 670-676.
NEMETH, P. M., PETTE, D. & VRBOVi, G. (1980). Malate dehydrogenase homogeneity of single fibresof the motor unit. In Plasticity of Muscle, ed. PETTE, D., pp. 45-54. Berlin: de Gruyter.
ROSENFALCK, P. & BUCHTHAL, F. (1970). On the concept of the motor subunit. Int. J. Neurosci.1, 27-37.
SANDERS, F. K. & YOUNG, J. Z. (1946). The influence of peripheral connexion on the diameter ofregenerating nerve fibres. J. exp. Biol. 22, 203-212.
STEIN, R. B., NICHOLS, T. R., JHAMANDAS, J., DAVIS, L. & CHARLES, D. (1977). Stable long-termrecordings from cat peripheral nerves. Brain Res. 128, 21-38.
STEIN, R. B., GORDON, T., HOFFER, J. A., DAVIS, L. A. & CHARLES, D. (1980). Long-term recordingsfrom cat peripheral nerves during degeneration and regeneration: Implications for human nerverepair and prosthetics. In Nerve Repairs: Its Clinical and Experimental Basis, ed. JEWETT, C. L.& MCCAROLL, H. R., pp, 166-176. St Louis: C. V. Mosby.
322

RECOVERY OF REINNER VATED MOTOR UNITSSTEPHENS, J. A. & TAYLOR, A. (1975). Human motor unit contractions studied by controlled
intramuscullar microstimulation. J. Physiol. 252, 8-9.SUNDERLAND, S. (1978). Nerves and Nerve Injuries. Edinburgh: Livingstone.SYROVY, I., GUTMANN, E. & MELICHNI, J. (1972). The effect of degeneration on contraction andmyosin properties of fast and slow rabbit and cat muscles. Phy8iol. bohemo8lov. 21, 353-359.
TAYLOR, A. & STEPHENS, J. A. (1976). Study of human motor unit contractions by controlledintramuscular stimulation. Brain Res. 117, 331-335.
WEISS, P. & TAYLOR, A. C. (1944). Further experimental evidence against 'neurotropism' in nerveregeneration. J. exp. Zool. 95, 233-257.
WUERKER, R. B., MCPHEDRAN, A. M. & HENNEMAN, E. (1965). Properties of motor units in aheterogeneous pale muscle (M. Gastrocnemius) of the cat. J. Neurophy8iol. 28, 85-99.
YAHR, M. D., HERz, E., MOLDAVER, J. & GRUNDFEST, H. (1950). Electromyographic patterns inreinnervated muscle. Arch8 Neurol. Psychiat., Chicago 63, 728-738.
YOUNG, J. Z. (1951). Growth and plasticity in the nervous system. Proc. R. Soc. B 139, 18-37.
11-2
323