cancer research de novo lipogenesis protects cancer cells ... · densely, modulation of lipogenesis...
TRANSCRIPT

Tum
DeRadMem
EvelieSebasJelle MGuido
Abst
Intro
Devassociin tumwith afor ma
AuthorEndocrNetworGeneticK.U. LeCenter,Oncolo
Note: SOnline
E. Rysmcontrib
CorresmentalHerestr32-16-3
doi: 10
©2010
www.a
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
Canceresearch
or and Stem Cell Biology
novo Lipogenesis Protects Cancer Cells from Freeicals and Chemotherapeutics by Promoting
R
brane Lipid Saturation
n Rysman1, Koen Brusselmans1, Katryn Scheys1, Leen Timmermans1, Rita Derua2,tian Munck3, Paul P. Van Veldhoven4, David Waltregny5, Veerle W. Daniëls1,
achiels1, Frank Vanderhoydonc1, Karine Smans6, Etienne Waelkens2, Verhoeven1, and Johannes V. Swinnen1ractAct
cancerreportphospshowgenicenzymlipid smoreand redensediffusiThesechang
poorerny tumo
s' Affiliatiinology, 2Divk (LiMoNe)s, and 4Labuven, LeuveUniversitygy Research
upplementa(http://cance
an, K. Bruuted equally
ponding AMedicine anaat 49 bus 945934; E-m
.1158/0008-
American A
acrjourna
Downlo
ivation of de novo lipogenesis in cancer cells is increasingly recognized as a hallmark of aggressives and has been implicated in the production of membranes for rapid cell proliferation. In the current, we provide evidence that this activation has a more profound role. Using a mass spectrometry–basedholipid analysis approach, we show that clinical tumor tissues that display the lipogenic phenotypean increase in the degree of lipid saturation compared with nonlipogenic tumors. Reversal of the lipo-switch in cancer cells by treatment with the lipogenesis inhibitor soraphen A or by targeting lipogenices with small interfering RNA leads to a marked decrease in saturated and mono-unsaturated phospho-pecies and increases the relative degree of polyunsaturation. Because polyunsaturated acyl chains aresusceptible to peroxidation, inhibition of lipogenesis increases the levels of peroxidation end productsnders cells more susceptible to oxidative stress–induced cell death. As saturated lipids pack morely, modulation of lipogenesis also alters lateral and transversal membrane dynamics as revealed byon of membrane-targeted green fluorescent protein and by the uptake and response to doxorubicin.data show that shifting lipid acquisition from lipid uptake toward de novo lipogenesis dramaticallyes membrane properties and protects cells from both endogenous and exogenous insults. These findingse important new insights into the role of de novo lipogenesis in cancer cells, and they provide a rationale
providfor the use of lipogenesis inhibitors as antineoplastic agents and as chemotherapeutic sensitizers. Cancer Res;70(20); 8117–26. ©2010 AACR.
as a rethe acare afftransltion. I
duction
elopment and progression of cancer are frequentlyated with increased de novo production of fatty acidsor cells. Activation of de novo lipogenesis correlates
prognosis and shorter disease-free survivalr types (1–4). This metabolic change occurs
creaseand istransfnomy(5). Inchemilencincell prIn sevis obsdent oThis smay ethe nefactorfew ofIn t
lipoge
ons: 1Laboratory for Experimental Medicine andision of Biochemistry, 3Light Microscopy and ImagingVIB, Department of Molecular and Developmentaloratory of Lipid Biochemistry and Protein Interactions,n, Belgium; 5Department of Urology and GIGA CancerHospital of Liège, Liege, Belgium; and 6Ortho Biotechand Development, Beerse, Belgium
ry data for this article are available at Cancer Researchrres.aacrjournals.org/).
sselmans, K. Scheys, L. Timmermans, and R. Deruato this work.
uthor: Johannes V. Swinnen, Laboratory for Experi-d Endocrinology, K.U. Leuven, Gasthuisberg, O&N 1,02, 3000 Leuven, Belgium. Phone: 32-16-330533; Fax:ail: [email protected].
5472.CAN-09-3871
ssociation for Cancer Research.
ls.org
Research. on April 16, 202cancerres.aacrjournals.org aded from
sult of common oncogenic insults and is mediated bytivation of multiple lipogenic enzymes. These enzymesected at all levels of regulation including transcription,ation, protein stabilization, and protein phosphoryla-ncreased lipid production has been linked to an in-d need for membranes during rapid cell proliferation,considered to be part of a more general metabolic
ormation that provides cancer cells with more auto-in terms of their supply of building blocks for growthsupport of this hypothesis, blockade of lipogenesis bycal inhibitors or RNA interference (RNAi)–mediated si-g of lipogenic enzymes or their regulators attenuatesoliferation and ultimately leads to cell death (6–16).eral cancer types, overexpression of lipogenic enzymeserved very early in cancer development and is indepen-f the proliferative status of the individual cells (17, 18).uggests that the role of tumor-associated lipogenesisxtend beyond bulk membrane biosynthesis to meeteds of rapid cell proliferation. Activation of growthsignaling and protection from cell death are just athe emerging novel roles of this pathway (19–22).
his study, we investigated the most direct effects ofnesis, specifically the changes in the lipid composition8117
0. © 2010 American Association for Cancer

of canalterattumorcarcining mrationtherap
Mate
Cell cLNC
were o(betwmorphcultursuppleand linbovineA was(HelmGermatainedfatty a(ACC)testedmicroquentContrwereLipofe
ClinicFres
normagone ataticperformor tijacentblottinby theStudie
[2-14C2-14
shamLipidsmetho
QuantriglyPho
tated
Gas cAliq
ted w
acidicmassa SupeTotalto theacid u
AnalyionizaTiss
v/v). Cmethyaddedtutedlipidsspectrion trBiosys(Adviolows:−55 eVmonitspeciewas utope eweresoftwa
LipidEqu
sonicacommResea
ImmuEqu
(NuPAbraneACC (TechnImagedensit
DeterfluorefluoreCell
were tcencesylateddiffusFV100Bio-RaA 40550 mWwere r
Rysman et al.
Cance8118
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
cer cells, and assessed the consequences of theseions on cancer cell biology. Our findings suggest that-associated lipogenesis protects cancer cells fromogenic- and therapeutic-associated insults by promot-embrane lipid saturation. They also provide aale for the use of lipogenesis inhibitors as chemo-eutic sensitizers.
rials and Methods
ulture and treatmentsaP, 22Rv1, PC-3, Du145, BT474, and HCT116 cellsbtained from the American Type Culture Collectioneen 2008 and 2010) and authenticated by checkingology and by karyotyping in December 2009. Cells wereed in RPMI 1640, DMEM/F-12, or McCoy's 5A mediummented with 10% FCS (Invitrogen). Palmitic, linoleic,olenic acid (Sigma) were complexed to fatty acid–freeserum albumin (Invitrogen) as described (8). Soraphenkindly provided by Drs. Klaus Gerth and Rolf Jansenholtz-Zentrum für Infektionsforschung, Braunschweig,ny; refs. 23, 24). Doxorubicin and verapamil were ob-from Sigma. Small interfering RNAs (siRNA) targetingcid synthase (FASN) and acetyl-CoA carboxylase-αwere obtained from Ambion. Multiple siRNA sets were. Those with the least off-target effects as judged byarray analysis (data not shown) were selected for subse-experiments (s883 and s5030). Silencer Select Negativeol #1 siRNA from Ambion was used as control. Cellsreverse transfected with siRNAs (50 nmol/L) usingctamine RNAiMAX (Invitrogen).
al tissue specimensh, snap-frozen prostate cancer tissues and matchingl samples were obtained from patients who had under-radical retropubic prostatectomy for localized pros-
carcinoma. Harvesting of the tissue samples wasmed as previously described (25). The normal and tu-ssues were identified by histologic analysis of areas ad-to the tissue that was used for lipid and Westerng analysis. The use of clinical samples was approvedLocal Commission for Medical Ethics and Clinical
s at the University of Liège.
]acetate incorporation assay and TLC analysisC–labeled acetate (57 mCi/mmol; 2 μCi/dish; Amer-International) was added to the culture medium.were extracted according to a modified Bligh-Dyerd and analyzed by TLC analysis as described (9).
tification of total cellular phospholipids,cerides, and cholesterolspholipids, triglycerides, and cholesterol were quanti-as described (26–28).
hromatographic analysis of fatty acyl chains
uots of Bligh-Dyer lipid extracts (9) were supplemen-ith tricosanoic acid as internal standard, subjected toband-calcul
r Res; 70(20) October 15, 2010
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
methanolysis, and analyzed by gas chromatography–spectrometry (Trace GC-MS, Thermo Finnigan) usingrcap 5HT column (15 m × 0.25 mm; 0.10 μm; Alltech).ion current signals, obtained in EI+ mode, were relatedinternal standard signal and converted to nmol fattysing experimentally obtained relative response factors.
sis of intact phospholipid species by electrospraytion tandem mass spectrometryue or cells were homogenized in 1 N HCl/CH3OH (1:8,HCl3, 200 μg/mL of the antioxidant 2,6-di-tert-butyl-4-lphenol (Sigma; ref. 29), and lipid standards were. The organic fractions were evaporated and reconsti-in CH3OH/CHCl3/NH4OH (90:10:1.25, v/v/v). Phospho-were analyzed by electrospray ionization tandem massometry (ESI-MS/MS) on a hybrid quadrupole linearap mass spectrometer (4000 QTRAP system, Appliedtems) equipped with a robotic nanoflow/ion sourcen Biosciences). The collision energy was varied as fol-prec 184, 50 eV; nl 141, 35 eV; nl 87, −40 eV; prec 241,. The system was operated in the multiple reactionoring (MRM) mode for quantification of individuals. Typically, a 3-minute period of signal averagingsed for each spectrum. Data were corrected for 13C iso-ffects if the contribution was >10%. Corrected datapresented as heat maps using the HeatMap Builderre (Clifton Watt, Stanford University).
peroxidation product assayal amounts of cells were scraped in ice-cold PBS. Aftertion, 3,000 × g supernatants were analyzed using aercial lipid peroxidation assay kit (Oxford Biomedicalrch).
noblotting analysisal amounts of protein were loaded onto precast gelsGE, Invitrogen), transferred to nitrocellulose mem-s, and incubated with antibodies against FASN (30),Cell Signaling Technology), α-tubulin (Cell Signalingology), and β-actin (Sigma) as described (8, 9). TheMaster 1D software (GE Healthcare) was used forometric quantification.
mination of lateral membrane dynamics byscence recovery after photobleaching andscence diffusion after photoactivations, grown on Lab-Tek II chambered cover glass (Nunc),ransfected with pAcGFP1-F (Clontech) for FRAP (fluores-recovery after photobleaching) or with PA-GFP-F (farne-photoactivatable form of GFP; ref. 31) for fluorescence
ion on photoactivation using an Olympus FluoView0 confocal microscope (Olympus America, Inc.) and ad Radiance 2100 confocal system, respectively (31–33).-nm laser diode was used for bleaching (10 seconds at) and activation. Full-frame images (512 × 512 pixels)
ecorded using a 488-nm argon laser and a 500- to 600-nm
pass emission filter. Changes in fluorescence wereated using the FluoView FV1000 software (Olympus).Cancer Research
0. © 2010 American Association for Cancer

DeteraccumTo
treatephatiddoxor450 nmthe NBrubicias thebicinwerelong-prubiciemissi
Cell dAt t
ent anble anexcluscombi
StatisThe
way AvaluesAll dafigure
Resu
Revercells oand cTo s
lular lcells, w(furtheinhibitthe lipprevioincorppholip(Fig. 1phospCholesBec
size podesatuseries,canceracyl cwouldacyl sextracmethyphen
(16:0),The lesors o(contaThe ncontenfor quacyl chcellula38% t9% toBas
chains(Fig. 1chaintions,mentFig. 1phatidsaturaspecieturatiodepensaturafor phplemenouncThe
not unall lipotherbreastline (HLNCaPmembknockinsteasomewreflec3-foldlabelean 8-fdata sphenomonowith mthe rely tha
ActivaincreahumaTo
is assoin vivotrol ti
Lipogenesis Promotes Membrane Lipid Saturation
www.a
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
mination of flip-flop rate and doxorubicinulationmeasure the flip-flop rate of doxorubicin, cells wered with 10 μmol/L 7-nitrobenzo-2-oxa-1,3-diazole phos-ylethanolamine (NBD-PE; Invitrogen), exposed toubicin, and monitored in a fluorimeter (excitation,; emission, 510 nm) at 37°C. The difference betweenD fluorescence 2 seconds after the addition of doxo-
n and the NBD fluorescence 4 minutes later was usedmeasurement for the relative flip-flop rate of doxoru-(34). To measure doxorubicin accumulation, the cellsanalyzed using a fluorescence microscope (515-nmass emission filter) 30 minutes after addition of doxo-n (10 μmol/L) or fluorimetrically (excitation, 490 nm;on, 580 nm).
eath assayhe indicated time after compound exposure, the adher-d floating cells were collected and combined. The via-d dead cells were counted using the trypan blue dyeion assay (9). Synergism was determined using thenation index (CI) method (35).
tical analysisresults were analyzed by a Student's t test or by one-NOVA using a Tukey multiple comparison test. Pof <0.05 were considered to be statistically significant.ta presented represent means ± SE, as indicated in thelegends.
lts and Discussion
sal of the lipogenic phenotype depletes cancerf saturated and mono-unsaturated acyl chainsauses a shift toward polyunsaturationtudy the effect of tumor-associated lipogenesis on cel-ipid composition, we treated LNCaP prostate cancerhich have a high lipogenic activity, with soraphen Ar referred to as soraphen). Soraphen is a highly potentor of ACC (23, 24, 36) and is an ideal tool to reverseogenic phenotype in cancer cells (12). Consistent withus studies (12), soraphen treatment decreased theoration of 14C- from 2-14C–labeled acetate into phos-ids and triglycerides by 7- and 13-fold, respectivelyA) and led to a 1.7- and 4.2-fold reduction of the totalholipid and triglyceride levels, respectively (Fig. 1B).terol levels remained unaffected.ause mammalian cells have a limited ability to synthe-lyunsaturated fatty acids de novo, as they lack the Δ12rase required to produce fatty acids of the ω3 and ω6we hypothesized that de novo lipogenesis would enrichcells mainly with saturated and/or mono-unsaturatedhains and that reversal of the lipogenic phenotypecause a selective reduction in the levels of these latterpecies. To test this hypothesis, the total cellular lipidts were methanolysed, and the generated fatty acyl
l esters were analyzed by gas chromatography. Sora-treatment decreased the total levels of palmitoylWestecorde
acrjournals.org
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
stearoyl (18:0), and oleoyl (18:1) species by ∼2-fold.vels of polyunsaturated acyl chain species, the precur-f which were obligatorily derived from the mediumining 10% serum), substantially increased (Fig. 1C).et result was (a) an overall decrease in cellular acylt, confirming the importance of de novo lipogenesisantitative lipid supply, and (b) a major shift in cellularain composition toward polyunsaturation; the relativer contribution of saturated acyl chains dropped fromo 29% and polyunsaturated fatty acids increased from20%.ed on our observation that most of the labeled acylwere incorporated into phospholipids in cancer cellsA) and given the importance of phospholipid acylcomposition in numerous membrane-related func-we further investigated the effects of soraphen treat-on intact phospholipids by ESI-MS/MS. As shown inD, soraphen treatment substantially decreased phos-ylcholine (PC) species with zero or one degree of un-tion (both acyl chains combined) up to 4-fold, whereass with polyunsaturated chains (more than three unsa-ns in both chains combined) increased up to 8-fold,ding on the species. A similar trend toward decreasedtion and increased polyunsaturation was also observedosphatidylethanolamine and phosphatidylserine (Sup-ntary Fig. S1); however, these effects were less pro-ed and less general compared with those of PC.observed changes in membrane lipid saturation wereique to LNCaP cells. Similar effects were observed inogenic cancer cell lines tested. These included threeprostate cancer cell lines (22Rv1, PC-3, and Du145), acancer cell line (BT474), and a colorectal cancer cellCT116; Fig. 1D). Changes were most outspoken inand in HCT116 cells. Importantly, a similar shift in
rane lipid saturation was seen after siRNA-mediateddown of the lipogenic enzymes FASN or ACC (siRNA)d of treatment with soraphen (Fig. 1E). Effects werehat less pronounced than after soraphen treatment,
ting the less potent inhibition of lipogenesis [2- toreduction in the incorporation of 14C- from 2-14C–d acetate into lipids (Supplementary Fig. S2) versusold reduction after soraphen treatment]. Overall, thesetrongly support the hypothesis that the lipogenictype in cancer cells provides cells with saturated and-unsaturated acyl chains, which replenishes the cellsembrane components and simultaneously increases
lative degree of saturation of phospholipids, particular-t of PC.
tion of the lipogenic pathway is associated withsed saturation of phospholipids in vivo inn tumor specimensassess whether the lipogenic phenotype of cancer cellsciated with an increased saturation of phospholipids, prostate tumor specimens and normal matching con-ssues were analyzed for overexpression of FASN by
rn blot analysis, and the phospholipid profiles were re-d. Three of the five matched samples (tumor versusCancer Res; 70(20) October 15, 2010 8119
0. © 2010 American Association for Cancer

contromor tiby ESIgenictumor
increaa decrpared
FigureLNCaPphosphfrom coC, effecchromasoraphMS (pre(expreswith 50 nmol/L siRNA targeting FASN or ACC, respectively. Lipid profiling was performed after 72 to 96 h in two pairs of samples (1–2). The heat mapsshow th
Rysman et al.
Cance8120
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
l) showed a substantial overexpression of FASN in tu-ssue (Fig. 2). Analysis of the phospholipid composition-MS/MS showed a different profile for PC in the lipo-
e siRNA/siCtrl ratio (expressed as log2) for different PC species.
tumors compared with the nonlipogenic tumors. Thes with increased FASN expression showed a consistent
port owith t
r Res; 70(20) October 15, 2010
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
se in saturated and mono-unsaturated acyl chains andease in polyunsaturated species in tumor tissue com-with matching normal tissue (Fig. 2). These data sup-
1. Effect of inhibition of de novo lipogenesis on the cellular lipid composition of cancer cells in vitro. A, effect of soraphen on lipid synthesis.cells were treated with soraphen (100 nmol/L) or vehicle (control) for 24 h. During the last 4 h, 2-[14C]acetate was added. 14C incorporation inolipids (PL), triglycerides (TG), and cholesterol (C) was analyzed by TLC. Columns, mean (n = 4); bars, SE. *, P < 0.05, significantly differentntrol. B, effect of soraphen on cellular lipid content of LNCaP cells 72 h after addition of soraphen (100 nmol/L) or vehicle (control; n = 3).t of soraphen treatment on cellular lipid acyl chain composition of LNCaP cells. Lipid extracts as in B were hydrolyzed and analyzed by gastography (n = 3). The scheme represents the contribution of the fatty acyl chains listed relative to their sum (in nmol/μg DNA) in both control anden-treated cells. D, effect of soraphen treatment on intact PC species in different cancer cell lines. Lipid extracts as in B were analyzed by ESI-MS/cursor 184) in the MRM mode. Lipid profiling was performed in three pairs of samples (1–3). The heat maps show the soraphen/control ratiosed as log2) for different PC species. E, effect of RNAi targeting lipogenic genes on intact PC species. LNCaP and HCT116 cells were transfected
ur findings with the lipogenesis inhibitor soraphen andhe siRNA-mediated knockdown of lipogenic enzymes
Cancer Research
0. © 2010 American Association for Cancer

in cantumorphosp
Moducells alipidSatu
chainsphysictheir scals ewhich
lipid stimatemaloncell dhydrogroupare mour obsis insaturapids o
Figureof FASnormalized to β-actin expression. Values are expressed as a ratio (log ) of tumor over matching normal tissue. The relative abundances of PC species wererecorde al rati
FiguresuscepperoxidpalmiticLNCaP(100 nmpalmiticlast 2 hH2O2. Elipid peperoxidbars, ScontrolsignificH2O2 aacid onanalyzeThe hea(expres
Lipogenesis Promotes Membrane Lipid Saturation
www.a
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
cer cell lines, and they provide in vivo evidence that-associated lipogenesis increases the saturation ofholipids in human tumors.
lation of de novo fatty acid synthesis in cancerffects the susceptibility of cellular membranes toperoxidationrated, mono-unsaturated, and polyunsaturated acyldramatically differ in terms of their structural andochemical properties. One of the key differences isusceptibility to peroxidation. In this process, free radi-
2
d by ESI-MS/MS in the MRM mode. The heat map shows the tumor/norm
xtract electrons from lipids in cellular membranes,leads to the formation of oxidized lipid spec
increa
sed as log2) for different PC species.
acrjournals.org
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
pecies have important biological functions and may ul-ly degrade into smaller reactive products includingdialdehydes and 4-hydroxyalkenals, which can causeamage when expressed at high levels (37–39). As thegens in between double bonds in methylene (CH2)s are particularly reactive, polyunsaturated acyl chainsuch more susceptible to peroxidation. Consistent withservation that modulation of de novo fatty acid synthe-cancer cells affects the balance between (mono-un)ted and polyunsaturated acyl chains in the phospholi-f cellular membranes, soraphen treatment significantly
o (expressed as log2) for different PC species.
2. Changes in phospholipid composition in lipogenic and nonlipogenic prostate tumor tissue specimens versus matched normal tissue. ExpressionN in matched pairs of malignant versus normal prostate tissue specimens was measured by Western blotting analysis, and the expression was
sed the levels of lipid peroxidation products (Fig. 3A).
ies. These Treatment with exogenous H2O2, which produces higher levels3. Modulation of lipogenesis affects thetibility of cellular membranes to lipidation. A, effects of soraphen, H2O2, andacid on lipid peroxidation products.cells were treated with soraphenol/L) or vehicle (control) and withacid (75 μmol/L) for 72 h. During the, the cells were exposed to 200 μmol/Lqual amounts of cells were analyzed forroxidation products using a lipidation assay kit. Columns, mean (n = 4);E. *, P < 0.05, significantly different fromwithout H2O2 exposure; #, P < 0.05,antly different from both treatment withnd soraphen alone. B, effect of palmiticthe PC profile. PC species wered by ESI-MS/MS in the MRM mode.t map shows the soraphen/control ratio
Cancer Res; 70(20) October 15, 2010 8121
0. © 2010 American Association for Cancer

of freeproduacyl cnous psuppocer cethe de
De nocells tThe
their dand mvestigaed thecell deH2O2-markeby trypacted
diumpolyunpronoity tonot durathersultsBT474siRNAcreaseas illuthesetectsby cha
Tumodynamcomm
Figuresoraph(PUFA;siRNA tfor 72 hcell viabH2O2 a
Rysman et al.
Cance8122
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
radicals, induced a further increase in peroxidationcts. Interestingly, partial replenishment of saturatedhains by supplementation of the medium with exoge-almitic acid (Fig. 3B) largely reversed these changes,rting the idea that enhanced lipogenesis renders can-lls less susceptible to lipid peroxidation by limitinggree of phospholipid polyunsaturation.
vo lipogenesis determines the sensitivity of cancero oxidative stress–induced cell deathre is growing evidence that oxidized phospholipids andegradation products play a key role in the inductionediation of cellular apoptosis (40–43). Therefore, we in-ted whether modulation of de novo lipogenesis affect-sensitivity of cancer cells to oxidative stress–inducedath. The native LNCaP cells were fairly resistant toinduced cell death, but pretreatment with soraphendly increased their death in response to H2O2, as shown
an blue staining (Fig. 4A). These effects were counter-by exogenous palmitic acid. Interestingly, when the me-In aofmem
ility. Columns, mean (n = 3); bars, SE. *, P < 0.05, significantly different from contrond soraphen/siRNA alone.
r Res; 70(20) October 15, 2010
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
was supplemented with a mixture of saturated andsaturated fatty acids, the rescue effect was much lessunced. These data indicate that the increased sensitiv-oxidative stress observed after soraphen treatment ise to the general decrease in the amount of lipids butto the change in lipid (un)saturation. Concordant re-were obtained in 22Rv1 cells (Fig. 4A) and in PC-3,, and HCT116 cells (Supplementary Fig. S3). Similarly,-mediated knockdown of the lipogenic pathway in-d the sensitivity to oxidative stress–induced cell death,strated for LNCaP and HCT116 cells (Fig. 4B). Overall,data support the idea that increased lipogenesis pro-cancer cells from oxidative stress–induced cell deathnging the extent of saturation of cellular membranes.
r-associated fatty acid synthesis alters membraneics and affects the uptake and response toon chemotherapeutics
ddition to its effect on lipid peroxidation, modulationbrane lipid composition is known to have amajor effect4. De novo lipogenesis in cancer cells determines their sensitivity to oxidative stress–induced cell death. A, LNCaP and 22Rv1 cells were treated withen (100 nmol/L) or vehicle (control) and with palmitic acid (75 μmol/L) or with a mixture of 10% palmitic, 45% linoleic, and 45% linolenic acid75 μmol/L total) for 72 h. During the last 24 h, cells were exposed to 300 μmol/L H2O2. B, LNCaP and HCT116 cells were transfected with 50 nmol/Largeting FASN or ACC (siRNA), respectively, or with control siRNA (siCtrl). Cells were treated with palmitic acid (75 μmol/L) or PUFA (75 μmol/L total). During the last 24 h, cells were exposed to 300 or 400 μmol/L H2O2, respectively. Cells were collected and stained with trypan blue to assess
#
l/siCtrl; , P < 0.05, significantly different from both treatment withCancer Research
0. © 2010 American Association for Cancer

on theine whcancerof sormembfers toplanereferslateramembques:To pewith tsylatioand mis medbleachhad acontro(SuppLNCa
farnesfixed tof thesuredtivatiowas leer diffresultcreaseTra
ferredby spepoundas doxtry intnousuptakof fattdoxor
Figuregreen flAfter 72Points,B, analysoraphfarnesytime = 4 s); bars, SE. Data are expressed as the % fluorescence intensity compared with the fluorescence intensity in the region of photoactivation at time 0.*, P < 0
Lipogenesis Promotes Membrane Lipid Saturation
www.a
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
mobility of membrane components (38, 44). To exam-ether modulation of de novo fatty acid synthesis incells exerts similar effects, we investigated the effect
aphen on both the lateral and transversal mobility ofrane components in LNCaP cells. Lateral mobility re-the movement of membrane components within theof one membrane leaflet, whereas transversal mobilityto movement from one leaflet to the other. To analyzel membrane mobility, we followed the movement ofrane-targeted GFP using two complementary techni-FRAP and fluorescence diffusion after photoactivation.rform FRAP, we transiently transfected LNCaP cellshe plasmid pAcGFP1-F that encodes GFP with a farne-n tag. We bleached the fluorescence in a small areaeasured the recovery of fluorescence in this area, whichiated by neighboring GFP molecules moving into theed area. As shown in Fig. 5A, soraphen-treated cellsfaster fluorescence recovery rate compared withl cells. Similar results were obtained with BT474 cells
.05, significantly different from control.
lementary Fig. S4). In the complementary experiment,P cells were transfected with a plasmid encoding a
by mofluore
acrjournals.org
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
ylated photoactivatable form of GFP (PA-GFP-F). At aime point after photoactivation of GFP in a small spotplasma membrane, the fluorescence intensity was mea-as a function of the distance from the spot of photoac-n. The slope of the curve for the soraphen-treated cellsss steep than that for the control cells, reflecting a high-usion rate (Fig. 5B). Together with the FRAP data, theses indicate that inhibition of fatty acid synthesis in-s the lateral mobility of membrane components.nsversal mobility of membrane components, also re-to as flip-flop, occurs at a low rate unless it is facilitatedcific transporters. However, for certain exogenous com-s including commonly used chemotherapeutics, suchorubicin, passive flip-flop is a major mechanism of en-o the cells (34). Because treatment of cells with exoge-polyunsaturated fatty acids is known to promote thee of doxorubicin (45), we determined whether inhibitiony acid synthesis would promote membrane flip-flop andubicin uptake. The flip-flop of doxorubicin was assessed
5. Modulation of lipogenesis alters membrane dynamics. A, FRAP analysis of the effect of soraphen on the lateral dynamics of farnesylateduorescent protein (GFP). LNCaP cells were transfected with a farnesylated GFP construct and treated with soraphen (100 nmol/L) or vehicle (control).h, the fluorescence was bleached in a specific region (white squares). The fluorescence recovery was analyzed at the indicated time points.mean (n = 41–44); bars, SE. Values from soraphen-treated cells were significantly different from control (P < 0.05) from 1 to 43 s after bleaching.sis of lateral dynamics of photoactivatable GFP. LNCaP cells were transfected with a photoactivatable farnesylated GFP construct and treated withen (100 nmol/L) or vehicle (control). After 72 h, the diffusion of fluorescence along the plasma membrane was analyzed after photoactivation oflated GFP in a specific region (white squares). The fluorescence was measured as a function of the distance from this region. Points, mean (n = 4; at
nitoring doxorubicin-mediated quenching of NBD-PE, ascently labeled phospholipid that incorporates into
Cancer Res; 70(20) October 15, 2010 8123
0. © 2010 American Association for Cancer
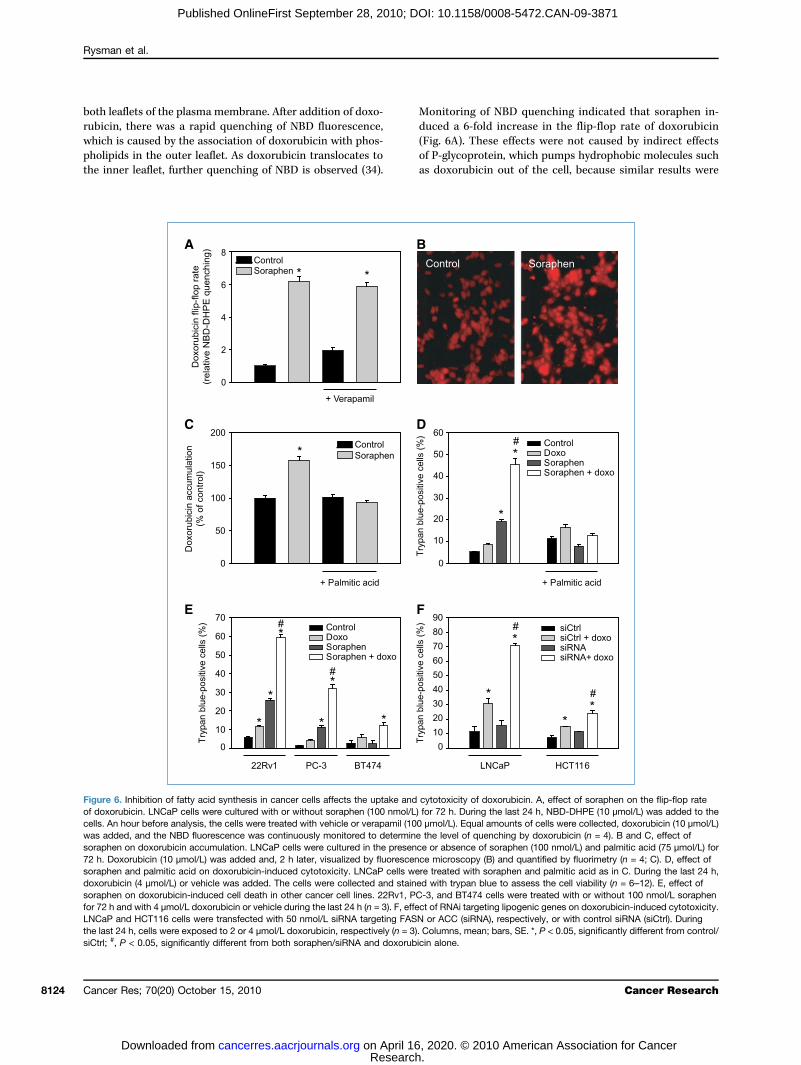
both lerubiciwhichpholipthe in
Monitduced(Fig. 6
Figureof doxocells. Awas adsoraph72 h. Dsoraphdoxorusoraphfor 72 hLNCaPthe lastsiCtrl; #
Rysman et al.
Cance8124
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
aflets of the plasma membrane. After addition of doxo-n, there was a rapid quenching of NBD fluorescence,is caused by the association of doxorubicin with phos-
ids in the outer leaflet. As doxorubicin translocates toner leaflet, further quenching of NBD is observed (34).of P-gas dox
24 h, cells were exposed to 2 or 4 μmol/L doxorubicin, respectively (n = 3). Colum, P < 0.05, significantly different from both soraphen/siRNA and doxorubicin alon
r Res; 70(20) October 15, 2010
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
oring of NBD quenching indicated that soraphen in-a 6-fold increase in the flip-flop rate of doxorubicinA). These effects were not caused by indirect effects
lycoprotein, which pumps hydrophobic molecules suchorubicin out of the cell, because similar results were6. Inhibition of fatty acid synthesis in cancer cells affects the uptake and cytotoxicity of doxorubicin. A, effect of soraphen on the flip-flop raterubicin. LNCaP cells were cultured with or without soraphen (100 nmol/L) for 72 h. During the last 24 h, NBD-DHPE (10 μmol/L) was added to then hour before analysis, the cells were treated with vehicle or verapamil (100 μmol/L). Equal amounts of cells were collected, doxorubicin (10 μmol/L)ded, and the NBD fluorescence was continuously monitored to determine the level of quenching by doxorubicin (n = 4). B and C, effect ofen on doxorubicin accumulation. LNCaP cells were cultured in the presence or absence of soraphen (100 nmol/L) and palmitic acid (75 μmol/L) foroxorubicin (10 μmol/L) was added and, 2 h later, visualized by fluorescence microscopy (B) and quantified by fluorimetry (n = 4; C). D, effect ofen and palmitic acid on doxorubicin-induced cytotoxicity. LNCaP cells were treated with soraphen and palmitic acid as in C. During the last 24 h,bicin (4 μmol/L) or vehicle was added. The cells were collected and stained with trypan blue to assess the cell viability (n = 6–12). E, effect ofen on doxorubicin-induced cell death in other cancer cell lines. 22Rv1, PC-3, and BT474 cells were treated with or without 100 nmol/L soraphenand with 4 μmol/L doxorubicin or vehicle during the last 24 h (n = 3). F, effect of RNAi targeting lipogenic genes on doxorubicin-induced cytotoxicity.and HCT116 cells were transfected with 50 nmol/L siRNA targeting FASN or ACC (siRNA), respectively, or with control siRNA (siCtrl). During
ns, mean; bars, SE. *, P < 0.05, significantly different from control/e.
Cancer Research
0. © 2010 American Association for Cancer

obtainamil (by a sdoxorurimetr22Rv1Additiof sortizedwhichdoxorsuscepculturinducepretreup to∼50%the CIogenorescuewere oPC-3 ailarly,sitized(Fig. 6cancertogethdeath,agentsthe uschemo
Conc
Theincrea
progreprovidrect emembtivatinare lestect caIt alsoefficacsuggescant aboth cstudyinhibitors for therapeutic intervention, particularly as thera-py sen
Disclosure of Potential Conflicts of Interest
No p
Acknowledgments
We t
Grant
K.U.Flanderof Attrafor ScieK. Schepostdoc
Theof pageaccorda
Refe1. Bru
SinSp
2. Kuold
3. Metyp
4. SwcanCa
5. Deoglife
6. PizKuce
7. Kusel199
8. Bruintgecel
Lipogenesis Promotes Membrane Lipid Saturation
www.a
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
ed in the presence of the P-glycoprotein inhibitor verap-Fig. 6A). The increased flip-flop rate was accompaniedignificant increase in the intracellular accumulation ofbicin, as assessed by fluorescence microscopy and fluo-ic analysis of cellular extracts and as shown also forand PC-3 cells (Fig. 6B and C; Supplementary Fig. S5).on of exogenous palmitic acid counteracted the effectsaphen (Fig. 6C). Soraphen treatment markedly sensi-LNCaP cells to the cytotoxic effects of doxorubicin,was consistent with the increased accumulation ofubicin in soraphen-treated cells and their increasedtibility to cell death. LNCaP cells grown under standarde conditions were fairly resistant to doxorubicin-d cell death (8% cell death at 4 μmol/L; Fig. 6D), butatment with soraphen, which alone induced death in20% of the cells, increased the rate of cell death to. This potent effect was synergistic, as assessed usingmethod (Supplementary Fig. S6; ref. 35). Addition of ex-us palmitic acid counteracted these effects and largelyd the cells from death (Fig. 6D). Concordant resultsbtained with the prostate cancer cell lines 22Rv1 andnd with the breast cancer cell line BT474 (Fig. 6E). Sim-knockdown of the lipogenic pathway with siRNA sen-the cancer cells for doxorubicin-induced cytotoxicity
F). These findings suggest that increased lipogenesis incells may limit the uptake of chemotherapeutics and,er with the mentioned changes in susceptibility to cellmay render cancer cells less susceptible to cytotoxic. Conversely, these data provide a new rationale fore of lipogenesis inhibitors to increase the efficacy oftherapeutic agents.
lusion
w that the implications of Rece
erference-mediated silencing of the acetyl-CoA-carboxylase-αne induces growth inhibition and apoptosis of prostate cancerls. Cancer Res 2005;65:6719–25.
9. DeRNattsis
10. Kriinh20
11. Krine18
12. Beaclec
13. ChcaRe
14. Haca
15. DoWB(THhu11
acrjournals.org
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
ssion, extend beyond its commonly accepted role ining lipids for rapid cell proliferation, as we provide di-vidence that cancer cells promote saturation of theirranes and modulate their biophysical properties by ac-g de novo lipogenesis. As (mono-un)saturated lipidss susceptible to lipid peroxidation, this shift may pro-ncer cells from lipid peroxidation–mediated cell death.alters membrane dynamics and affects the uptake andy of chemotherapeutics. The findings from this studyt that tumor-associated lipogenesis confers a signifi-dvantage to cancer cells, as it helps them to survivearcinogenic- and therapeutic-mediated insults. Thisalso provides a novel strategy for the use of lipogenesis
sitizers.
otential conflicts of interest were disclosed.
hank Drs. Klaus Gerth and Rolf Jansen for soraphen A.
Support
Leuven grants OT/07/068A and GOA 2008/16; Research Foundation-s grant G.0590.08; “Stichting tegen Kanker”; and Interuniversity Polesction Programme–Belgian State, Prime Minister's Office, Federal Officentific, Technical and Cultural affairs. E. Rysman, L. Timmermans,ys, and V.W. Daniëls are research assistants and K. Brusselmans is atoral fellow of the Research Foundation-Flanders.costs of publication of this article were defrayed in part by the paymentcharges. This article must therefore be hereby marked advertisement innce with 18 U.S.C. Section 1734 solely to indicate this fact.
ived 11/16/2009; revised 06/14/2010; accepted 07/18/2010; published
results from this study sho sed lipogenesis, which frequently accompanies cancer OnlineFirst 09/28/2010.rencessselmans K, Swinnen JV. The lipogenic switch in cancer. In:gh KK, Costello LC, editors. Mitochondria and cancer. New York:ringer; 2009, p. 39–59.hajda FP. Fatty acid synthase and cancer: new application of anpathway. Cancer Res 2006;66:5977–80.nendez JA, Lupu R. Fatty acid synthase and the lipogenic pheno-e in cancer pathogenesis. Nat Rev Cancer 2007;7:763–77.innen JV, Brusselmans K, Verhoeven G. Increased lipogenesis incer cells: new players, novel targets. Curr Opin Clin Nutr Metabre 2006;9:358–65.Berardinis RJ, Lum JJ, Hatzivassiliou G, Thompson CB. The biol-y of cancer: metabolic reprogramming fuels cell growth and pro-ration. Cell Metab 2008;7:11–20.er ES, Jackisch C, Wood FD, Pasternack GR, Davidson NE,hajda FP. Inhibition of fatty acid synthesis induces programmedll death in human breast cancer cells. Cancer Res 1996;56:2745–7.hajda FP, Jenner K,Wood FD, et al. Fatty acid synthesis: a potentialective target for antineoplastic therapy. Proc Natl Acad Sci U S A4;91:6379–83.sselmans K, De Schrijver E, Verhoeven G, Swinnen JV. RNA
Schrijver E, Brusselmans K, Heyns W, Verhoeven G, Swinnen JV.A interference-mediated silencing of the fatty acid synthase geneenuates growth and induces morphological changes and apopto-of LNCaP prostate cancer cells. Cancer Res 2003;63:3799–804.del SJ, Axelrod F, Rozenkrantz N, Smith JW. Orlistat is a novelibitor of fatty acid synthase with antitumor activity. Cancer Res04;64:2070–5.del SJ, Lowther WT, Pemble CW. Fatty acid synthase inhibitors:w directions for oncology. Expert Opin Investig Drugs 2007;16:17–29.ckers A, Organe S, Timmermans L, et al. Chemical inhibition ofetyl-CoA carboxylase induces growth arrest and cytotoxicity se-tively in cancer cells. Cancer Res 2007;67:8180–7.ajes V, Cambot M, Moreau K, Lenoir GM, Joulin V. Acetyl-CoArboxylase α is essential to breast cancer cell survival. Cancers 2006;66:5287–94.tzivassiliou G, Zhao F, Bauer DE, et al. ATP citrate lyase inhibitionn suppress tumor cell growth. Cancer Cell 2005;8:311–21.nnelly C, Olsen AM, Lewis LD, Eisenberg BL, Eastman A, Kinlaw. Conjugated linoleic acid (CLA) inhibits expression of the Spot 14
RSP) and fatty acid synthase genes and impairs the growth ofman breast cancer and liposarcoma cells. Nutr Cancer 2009;61:4–22.Cancer Res; 70(20) October 15, 2010 8125
0. © 2010 American Association for Cancer

16. Kin14:tar
17. Piysynsio106
18. Milthecar
19. Littfatcel
20. Fioacipla200
21. MigenzIns
22. VaOvtyr20
23. Beandlide
24. Gephlulobio
25. Waexpcer162
26. Vacohu
27. Vabareqlay
28. Vaof
29. Milcel
30. Va
Roince
31. Pase18
32. Sadotre
33. RuveaJ E
34. Rethrme
35. ChshAd
36. ShforaseCe
37. Deme
38. RatieAc
39. ScdaCh
40. Wecarel20
41. Tain
42. Fruph
43. Dupeac
44. Stitie
45. Da
Rysman et al.
Cance8126
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871
law WB, Quinn JL, Wells WA, Roser-Jones C, Moncur JT. Spota marker of aggressive breast cancer and a potential therapeuticget. Endocrinology 2006;147:4048–55.athilake CJ, Frost AR, Manne U, et al. The expression of fatty acidthase (FASE) is an early event in the development and progres-n of squamous cell carcinoma of the lung. Hum Pathol 2000;31:8–73.graum LZ, Witters LA, Pasternack GR, Kuhajda FP. Enzymes offatty acid synthesis pathway are highly expressed in in situ breastcinoma. Clin Cancer Res 1997;3:2115–20.le JL, Wheeler FB, Fels DR, Koumenis C, Kridel SJ. Inhibition ofty acid synthase induces endoplasmic reticulum stress in tumorls. Cancer Res 2007;67:1262–9.rentino M, Zadra G, Palescandolo E, et al. Overexpression of fattyd synthase is associated with palmitoylation of Wnt1 and cyto-smic stabilization of β-catenin in prostate cancer. Lab Invest8;88:1340–8.ita T, Ruiz S, Fornari A, et al. Fatty acid synthase: a metabolicyme and candidate oncogene in prostate cancer. J Natl Cancert 2009;101:519–32.zquez-Martin A, Colomer R, Brunet J, Lupu R, Menendez JA.erexpression of fatty acid synthase gene activates HER1/HER2osine kinase receptors in human breast epithelial cells. Cell Prolif08;41:59–85.dorf N, Schomburg D, Gerth K, Reichenbach H, Hofle G. Isolationstructure elucidation of soraphen A1, a novel antifungal macro-from Sorangium cellulosum. Liebigs Ann Chem 1993:1017–21.
rth K, Bedorf N, Irschik H, Hofle G, Reichenbach H. The sora-ens: a family of novel antifungal compounds from Sorangium cel-sum (Myxobacteria). I. Soraphen A1α: fermentation, isolation,logical properties. J Antibiot (Tokyo) 1994;47:23–31.ltregny D, Alami Y, Clausse N, de Leval J, Castronovo V. Over-ression of the homeobox gene HOXC8 in human prostate can-correlates with loss of tumor differentiation. Prostate 2002;50:–9.n Veldhoven PP, Bell RM. Effect of harvesting methods, growthnditions and growth phase on diacylglycerol levels in culturedman adherent cells. Biochim Biophys Acta 1988;959:185–96.n Veldhoven PP, Swinnen JV, Esquenet M, Verhoeven G. Lipase-sed quantitation of triacylglycerols in cellular lipid extracts:uirement for presence of detergent and prior separation by thin-er chromatography. Lipids 1997;32:1297–300.n Veldhoven PP, Meyhi E, Mannaerts GP. Enzymatic quantitationcholesterol esters in lipid extracts. Anal Biochem 1998;258:152–5.
ne S, Ivanova P, Forrester J, Alex BH. Lipidomics: an analysis oflular lipids by ESI-MS. Methods 2006;39:92–103.n de Sande T, De Schrijver E, Heyns W, Verhoeven G, Swinnen JV.ondruline
r Res; 70(20) October 15, 2010
Research. on April 16, 202cancerres.aacrjournals.org Downloaded from
le of the phosphatidylinositol 3′-kinase/PTEN/Akt kinase pathwaythe overexpression of fatty acid synthase in LNCaP prostate can-r cells. Cancer Res 2002;62:642–6.tterson GH, Lippincott-Schwartz J. A photoactivatable GFP forlective photolabeling of proteins and cells. Science 2002;297:73–7.uer H, Putz V, Fischer K, Hescheler J, Wartenberg M. Increasedxorubicin uptake and toxicity in multicellular tumour spheroidsated with DC electrical fields. Br J Cancer 1999;80:1204–13.nions J, Brach T, Kuhner S, Hawes C. Photoactivation of GFP re-ls protein dynamics within the endoplasmic reticulum membrane.xp Bot 2006;57:43–50.gev R, Yeheskely-Hayon D, Katzir H, Eytan GD. Transport of an-acyclines and mitoxantrone across membranes by a flip-flopchanism. Biochem Pharmacol 2005;70:161–9.ou TC, Talalay P. Quantitative analysis of dose-effect relation-ips: the combined effects of multiple drugs or enzyme inhibitors.v Enzyme Regul 1984;22:27–55.en Y, Volrath SL, Weatherly SC, Elich TD, Tong L. A mechanismthe potent inhibition of eukaryotic acetyl-coenzyme A carboxyl-by soraphen A, a macrocyclic polyketide natural product. Mol
ll 2004;16:881–91.igner HP, Hermetter A. Oxidized phospholipids: emerging lipiddiators in pathophysiology. Curr Opin Lipidol 2008;19:289–94.binovich AL, Ripatti PO. On the conformational, physical proper-s and functions of polyunsaturated acyl chains. Biochim Biophysta 1991;1085:53–62.hneider C, Porter NA, Brash AR. Routes to 4-hydroxynonenal: fun-mental issues in the mechanisms of lipid peroxidation. J Biolem 2008;283:15539–43.st JD, Ji C, Duncan ST, et al. Induction of apoptosis in colorectalrcinoma cells treated with 4-hydroxy-2-nonenal and structurallyated aldehydic products of lipid peroxidation. Chem Res Toxicol04;17:453–62.ng DG, La E, Kern J, Kehrer JP. Fatty acid oxidation and signalingapoptosis. Biol Chem 2002;383:425–42.wirth GO, Hermetter A. Mediation of apoptosis by oxidized phos-olipids. Subcell Biochem 2008;49:351–67.pertuis YM, Meguid MM, Pichard C. Colon cancer therapy: newrspectives of nutritional manipulations using polyunsaturated fattyids. Curr Opin Clin Nutr Metab Care 2007;10:427–32.llwell W, Wassall SR. Docosahexaenoic acid: membrane proper-s of a unique fatty acid. Chem Phys Lipids 2003;126:1–27.vies CL, Loizidou M, Cooper AJ, Taylor I. Effect of γ-linolenic acidcellular uptake of structurally related anthracyclines in human
g sensitive and multidrug resistant bladder and breast cancer cells. Eur J Cancer 1999;35:1534–40.Cancer Research
0. © 2010 American Association for Cancer

2010;70:8117-8126. Published OnlineFirst September 28, 2010.Cancer Res Evelien Rysman, Koen Brusselmans, Katryn Scheys, et al. Saturationand Chemotherapeutics by Promoting Membrane Lipid
Lipogenesis Protects Cancer Cells from Free RadicalsDe novo
Updated version
10.1158/0008-5472.CAN-09-3871doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2010/09/24/0008-5472.CAN-09-3871.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/70/20/8117.full#ref-list-1
This article cites 43 articles, 13 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/70/20/8117.full#related-urls
This article has been cited by 32 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/70/20/8117To request permission to re-use all or part of this article, use this link
Research. on April 16, 2020. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 28, 2010; DOI: 10.1158/0008-5472.CAN-09-3871