carotid baroreceptor function in dogs with chronic...
TRANSCRIPT

745
Carotid Baroreceptor Function in Dogs WithChronic Norepinephrine Infusion
Jie Wang, Manuel Ochoa, Mrugesh B. Patel, Irving H. Zucker,
Alden V. Loud, Guillermo A. Zeballos, and Thomas H. Hintze
Carotid baroreceptor function, the compliance of the carotid sinus wall, and the structure of thecarotid artery were examined in dogs with elevated plasma norepinephrine (2,000-4,000 pg/ml)for 28 days. The dogs with high norepinephrine were normotensive (100±4.0 versus 98±4.0mm Hg;/7>0.05) with bradycardia (65±4.0 versus 87±16 beats/min; p<0.05) compared withnormal dogs in the conscious state. However, after pentobarbital anesthesia blood pressure wassignificantly higher in dogs with chronic norepinephrine infusion (165 ±6 mm Hg) comparedwith normal dogs (132±6 mm Hg). To assess baroreceptor sensitivity, multiunit carotidbaroreceptor activity was recorded from the right carotid sinus nerve, and the carotid sinus wallcompliance (sonomicrometers) was measured during nitroglycerin and phenylephrine injec-tions. The threshold and saturation pressures increased from 96±3.9 to 117±4.2 mm Hg andfrom 145±4J to 171±5.7 mm Hg, respectively, in the normal dogs compared with the highnorepinephrine dogs. The most striking differences were the marked increases in sensitivity ofcarotid baroreceptors (0.47±0.05 versus 1.99±0.45 spikes • sec"1 • mm Hg"1; p<0.01) andmaximum firing frequency of the baroreceptors (24±3.1 versus 48±4.4 spikes/sec; /?<0.01),whereas the carotid sinus wall compliance was unchanged (0.014 ±0.003 versus 0.012 ±0.002mm/mm Hg; /?>0.05). Similar alterations were observed using single fiber recordings, that is,an increase in threshold and saturation pressures and slope of baroreceptor units in dogs withelevated norepinephrine. The wall thickness and area of the carotid artery were determined.Both increased significantly (0.77±0.06 versus 1 J0±0.12 mm and 9.0±0.8 versus 11.9±0.9mm1; p<0.05) in dogs chronically infused with norepinephrine while the dry weight-to-wetweight ratio of left carotid artery tissue also increased from 26.0 ±0.73% to 29.0 ±0-57%. Thesestudies indicate that 1) one of the possible mechanisms responsible for bradycardia in theconscious dogs with high norepinephrine is enhanced sensitivity of carotid baroreceptors; 2).theenhanced sensitivity of carotid baroreceptors is not due to a change in compliance of the carotidsinus wall; and 3) chronic elevation of norepinephrine causes hypertrophy or hyperplasia of thewall of the common carotid artery. {Hypertension 1991;17:745-754)
Chronic elevations in plasma arterial catechol-amine concentrations are observed in sev-eral diseases including mitral valve prolapse,1
chronic myocardial failure,2"4 and pheochromocy-toma.3 These diseases may exhibit sustained or epi-sodic hypertension, dysautonomia, hyperadrenergicstates, and hypervagotonia.6-8 Previous studies from
From the Departments of Physiology (J.W., M.O., M.B.P.,G.A.Z., T.H.H.) and Pathology (A.V.L.), New York MedicalCollege, Valhalla, N.Y., and Department of Physiology and Bio-physics (I.H.Z.), University of Nebraska College of Medicine,Omaha, Neb.
Supported by grant HL-34167, HL-36264, and HL-33448 fromthe National Institutes of Health.
Address for correspondence: Thomas H. Hintze, PhD, Depart-ment of Physiology, New York Medical College, Valhalla, NY10595.
Received May 23, 1989; accepted in revised form February 20,1991.
this laboratory9 have shown that despite chronicallyhigh levels of norepinephrine (NE) (2,000-4,000 pg/ml), conscious dogs were normotensive due to brady-cardia and low cardiac output. These data suggest thatreflex buffering mechanisms are responsible for themaintenance of systemic arterial pressure, especiallysince hexamethonium or atropine in these dogs causedtachycardia and hypertension. We hypothesized thatthe arterial baroreceptor reflex plays a key role as abuffering mechanism in this model. Heymans et al10
demonstrated in 1953 that local application of NEonto the carotid sinus induced stimulation and sensiti-zation of the baroreceptors, leading to intense activa-tion of the baroreceptor reflex in dogs.10 Blacket et aJn
showed that intravenous infusion of catecholamines for8 days caused only transient hypertension in rabbits andthat thereafter the rabbits were normotensive withbradycardia. As yet, there have been no studies docu-
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from

746 Hypertension Vol 17, No 6, Part 1 June 1991
meriting the effects of chronic elevation of NE oncarotid sinus baroreceptors (CSBs). The goal of thepresent experiments was to determine if chronic eleva-tion of NE altered CSB function and whether thebaroreceptors might be responsible for normalizingsystemic arterial pressure in awake dogs with chroni-cally elevated NE.
MethodsA total of 37 mongrel dogs of either sex, weighing
20-34 kg, were used in four sets of experiments. Alldogs were anesthetized with sodium pentobarbital(30 mg/kg i.v.). Supplemental doses of anestheticwere given as necessary throughout the experiment.The protocol and methods were approved by theLACUC of New York Medical College and of theUniversity of Nebraska Medical School and conformto the Guiding Principles for the Use and Care ofLaboratory Animals of the American PhysiologicalSociety and the National Institutes of Health.
Effects of Chronic Elevation in PlasmaNorepinephrine on Carotid SinusBaroreceptor Function
Dogs in group 1 were used to determine the CSBfunction in six dogs with chronic NE infusion and infive normal dogs. A Tygon (Norton Plastics, Akron,Ohio) catheter was implanted in the descendingthoracic aorta and secured with a purse-string suturethrough a thoracotomy in the fifth left intercostalspace using sterile surgical techniques. The chest wasclosed in layers and the pneumothorax reduced. Thedogs were allowed 2-3 weeks to recover. The meanarterial blood pressure and heart rate were measuredfrom the implanted catheter connected to a strain-gauge transducer (Statham P23ID, Oxnard, Calif.)with the dogs in the conscious state. The procedurewas performed for dogs with chronic NE infusion asdescribed by us previously.9 In brief, two Alzet os-motic pumps (model 2ML4, Palo Alto, Calif.) con-taining NE (equivalent to 0.5 ^g/kg/min for 28 days)were implanted subcutaneously in the dorsal aspectof the neck using local anesthesia with 2% lidocaine.After 10 days, another osmotic pump containing 30%of the original concentration of NE was implanted tomaintain a NE plasma level of 2,000-4,000 pg/ml.Arterial pressure and heart rate were measured atthe first, second, and third week after the elevation ofplasma NE. At the 28th day of high NE, meanarterial pressure and heart rate were measured firstin the conscious state. Then, in both normal dogs anddogs with high NE, CSB activity was recorded fromthe carotid sinus nerve (CSN) after pentobarbitalanesthesia.
In brief, a midline cervical incision was used toexpose the right carotid sinus. The dissection wascarefully continued to define the glossopharyngealnerve and the CSN. The CSN was cleaned of sur-rounding connective tissue and cut at its junctionwith the glossopharyngeal nerve. With the aid of a
dissecting microscope, the sheath of the peripheralend of the CSN was removed, and the nerve wasplaced on platinum bipolar electrodes. The nerveimpulses were preamplified using a P15D AC pream-plifier (Grass Instruments, Inc., Waltham, Mass.)(gain of 1,000; 30-Hz to 3-KHz bandpass); nerveactivity was displayed on an oscilloscope (type 565,Tektronix Inc., Portland, Ore.) and simultaneouslytransferred into a window discriminator (model 120,W-P Instruments Inc., Hamden, Conn.). The lowerwindow was set above the noise level, and the upperlevel window was set high enough to incorporate allthe moderate and large spike activity that was visibleon the oscilloscope. Only a few very large spikeswhose amplitude was higher than the upper windowand only those spikes that became silent after injec-tion of nitroglycerin were counted. The rastor/step-per (rate meter) was set to count the discriminatedspikes and then reset after counting 16 spikes. Inreality, we counted only a few large fibers in eachpreparation. The CSN was covered by warm mineraloil to prevent dehydration of the nerve. The phasicand mean arterial pressures, original nerve dis-charge, discriminator output, and rate meter outputwere recorded on a strip-chart recorder (1000ES,Gould, Rahway, N.J.).
The arterial pressure was decreased and increasedafter intravenous bolus injections of nitroglycerin(25-100 /ig/kg) and phenylephrine (5-10 /ig/kg),respectively. Nitroglycerin was always administeredfirst. Phenylephrine was given when the arterialpressure, the original nerve discharge, and the dis-criminated spikes had returned to control values. Thedoses of nitroglycerin that we used were large enoughto completely inhibit all nerve activity. The protocolwas then repeated but with phenylephrine given first.The pressure at which the CSB firing disappearedwas designated as threshold pressure and the pres-sure at which the CSB activity no longer increased,even though arterial pressure continued to increase,was designated as saturation pressure. The pressure-discharge data for nitroglycerin and phenylephrinewere combined and a function curve (see below) ofCSB discharge versus arterial pressure was con-structed and the peak gain of the curve was calcu-lated. In addition to recording multiunit activity, wealso obtained threshold and saturation pressure andgain in five single units from two dogs with high NEand five single fibers from four normal dogs.
Effects of Chronic Elevation in PlasmaNorepinephrine on Carotid Sinus Wall Functionand the Maximum Firing Rate of CarotidSinus Baroreceptor
The second group (group 2) was designed tomeasure the diameter of the carotid sinus and themaximum firing rate of CSB in the same dogs. Tendogs were used. Six dogs were given chronic NEinfusion for 28 days. Another four dogs served ascontrols. A Millar microtip pressure transducer
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Wang et al Baroreceptor Function and Norepinephrine 747
(Millar Instruments, Houston, Tex.) was advancedinto the right carotid sinus through the thyroid arteryto permit high fidelity measurement of the intrasinuspressure. Two ultrasonic dimension transducers at-tached to a Dacron backing were sutured to thebottom and the top of the carotid sinus to givecontinuous measurements of carotid sinus diameter.Arterial blood pressure was changed by intravenousinjections of nitroglycerin and phenylephrine. Thereceived ultrasonic signal was monitored on an oscil-loscope during placement and during the experimentand the output was displayed on a strip-chart re-corder. Dimensions were measured using a TransitTime ultrasonic dimension gauge. The relation be-tween intrasinus systolic pressure and carotid sinusdiameter, as well as between carotid sinus diameterand CSN discharge, was determined and the slopesof these curves were calculated. In these experi-ments, the original CSN discharge was also sent to aHeathkit counter (model IM-4100, Benton Harbor,Mich.) to quantify spike frequency.
Because of potential interference between the sys-tem for measuring carotid wall diameter and nerverecording, we were careful to ground between thediameter crystals and the recording electrode. In all ofthe animals, nitroglycerin and phenylephrine weregiven while measuring pressure and diameter and thengiven again while measuring pressure and spike activity.The order of these studies was then reversed. Pressurewas measured each time and thus assured that eachinjection had similar effects. In three of the dogs, twowith NE and one normal dog, we were able to recordspike activity and diameter simultaneously.
Determination of Morphological Changes in CarotidSinus Wall
The third group of dogs (group 3) was used todetermine the external and internal diameters, area,and wall thickness of the right carotid sinus by aplanimetric technique using a Videoplan Image Ana-lyzer (Zeiss, Hawthorne, N.Y.). The right commoncarotid artery including the carotid sinus was perfusedwith formalin at 100 mm Hg in 10 dogs (high NE, n=4;normal, n=6) and then the carotid artery was cuttransversely into at least three rings just adjacent to thecarotid sinus for planimetric measurements. Since thecarotid artery bifurcates at the level of the carotid sinus,if these rings were cut at the carotid sinus there wouldhave been a septum making it impossible to measurethe luminal diameter. Mean values for these data wereobtained in each dog and then averaged in dogs withhigh NE and in control dogs. The left carotid artery wascollected in four dogs with high NE and in sevencontrol dogs. The dry weight-to-wet weight ratio of theartery was examined and compared between dogs withand without high NE.
Data AnalysisFor multifiber preparation, the CSB function curve
was fitted using a Sigmoid computer program based onthe five parameter logistic equations by Ishikawa et al12
TABLE 1. Influences of Pentobarbital Anesthesia and ChronicallyElevated Norepinephrine on Mean Arterial Pressure and HeartRate in Dogs
Parameter
Mean arterialpressure (mm Hg)
Heart rate(beats/min)
Condition
Control
High NE
Control
HighNE
Conscious
98±4.0
(*=5)
100±4.0(«=5)87±16(n=5)
65±4.0t
(«=5)
Anesthetized
132±6.0*
("=6)
165±6.0*f("=5)
143±16'(n=6)
185±14*t(n=5)
NE, norepinephrine.'p<0.05 compared with conscious state.t/><0.05 compared with control.
and was plotted using MULPLT (Decus, Marlboro,Mass.) on a PDP 11/73 computer. The maximum gainwas determined by calculating the first derivative of thiscurve. Strain was calculated as radius minus unstressedradius divided by radius.13-14 Unstressed diameter wastaken at a pressure of 40 mm Hg. Statistical analysiswas performed using Student's / test to evaluate thesignificance of differences between two means fromdogs with high NE and control dogs. Statistical signifi-cance was determined at the/?<0.05 level. The resultsare expressed as mean±SEM.
ResultsTable 1 shows the influence of barbiturate anes-
thesia on mean arterial pressure and heart rate bothin dogs with high NE and in normal dogs. In theconscious state, there was no significant difference inmean arterial pressure between normal dogs andthose with high NE. However, dogs with high NE hada marked bradycardia. After anesthesia, mean arte-rial pressure and heart rate rose more in the dogswith high NE than in control dogs (p<0.05).
Effects of Chronic Elevation in PlasmaNorepinephrine on Carotid SinusBaroreceptor Function
After injection of nitroglycerin and phenyleph-rine, the threshold and saturation pressures ofCSBs were measured. Figure 1 shows an originalrecording from one dog with high NE. The windowdiscriminator pulse in this figure clearly shows thatonly a few of the larger units were counted. As canbe seen during nitroglycerin administration, athreshold pressure is reached at which dischargeceases. We found that the average threshold pres-sure and saturation pressure of CSB were signifi-cantly elevated in dogs with high NE. Figure 2illustrates typical curves from one control dog andone dog with high NE. Although both curves followa characteristic sigmoid relation, threshold pres-sure, saturation pressure, and the slope of thelinear portion of the curve are all elevated in thedog with high NE. The mean data for gain and
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from
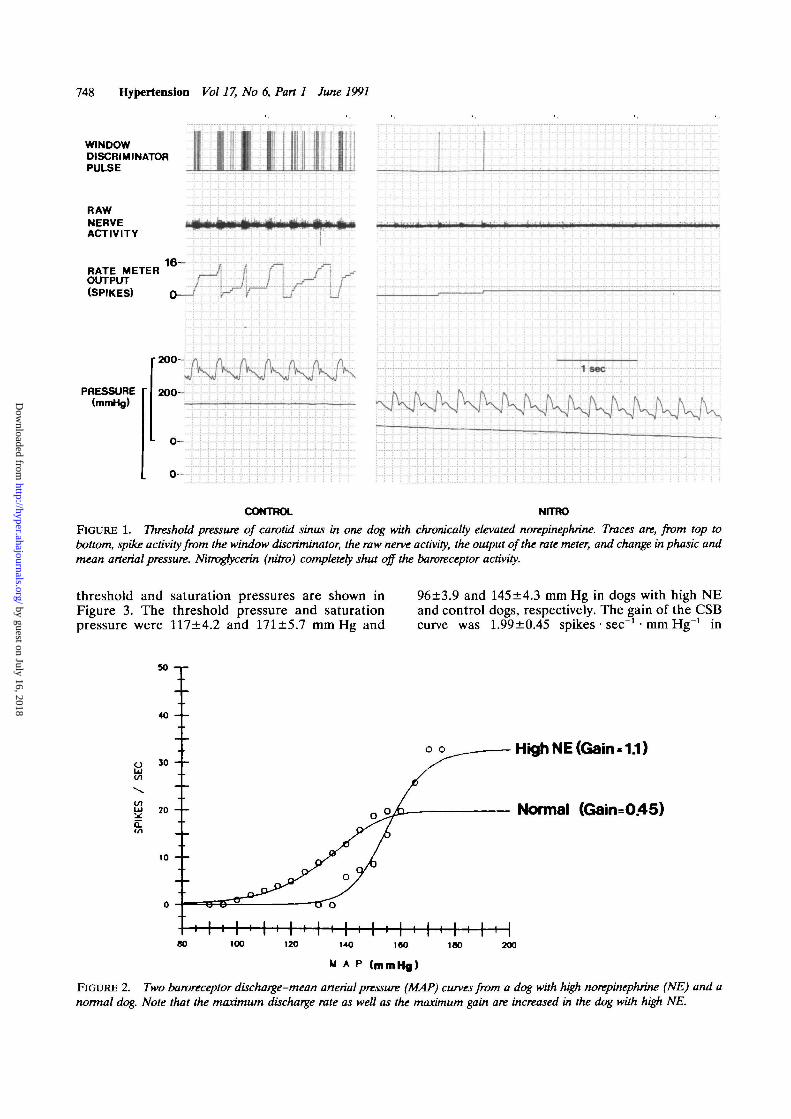
748 HjtKJrtension Vol 17, No 6, Part 1 June 1991
WINDOWDISCRIMINATORPULSE
RAWNERVEACTIVITY
RATE METEROUTPUT(SPIKES)
PRESSURE(mtnHg)
CONTROL NITRO
FIGURE 1. Threshold pressure of carotid sinus in one dog with chronically elevated norepinephrine. Traces are, from top tobottom, spike activity from the window discriminator, the raw nerve activity, the output of the rate meter, and change in phasic andmean arterial pressure. Nitrogtycerin (nitro) completely shut off the baroreceptor activity.
threshold and saturation pressures are shown inFigure 3. The threshold pressure and saturationpressure were 117±4.2 and 171±5.7 mm Hg and
96±3.9 and 145±4.3 mm Hg in dogs with high NEand control dogs, respectively. The gain of the CSBcurve was 1.99±0.45 spikes • sec"1 • mm Hg"1 in
High NE (Gain-1.1)
Normal (Gain=0.45)
I ' I ' I • I ' I i I i I ' I ' I100 120 140 100 100 200
M A P ( m m H g )
FIGURE 2. Two baroreceptor discharge-mean arterial pressure (MAP) curves from a dog with high norepinephrine (NE) and anormal dog. Note that the maximum discharge rate as well as the maximum gain are increased in the dog with high NE.
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from
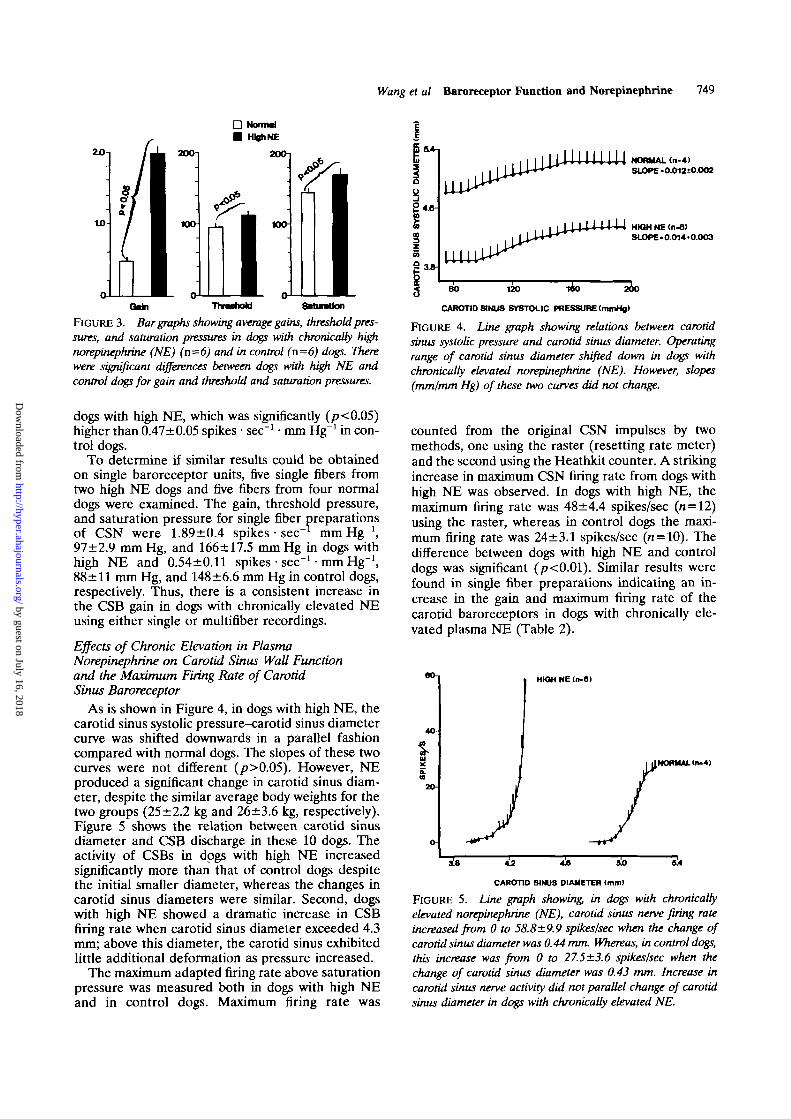
Wang et al Baroreceptor Function and Norepinephrine 749
Qafci ThrMhoU SMuraUon
FIGURE 3. Bar graphs showing average gains, threshold pres-sures, and saturation pressures in dogs with chronically highnorepinephrine (NE) (n=6) and in control (n=6) dogs. Therewere significant differences between dogs with high NE andcontrol dogs for gain and threshold and saturation pressures.
dogs with high NE, which was significantly (/?<0.05)higher than 0.47±0.05 spikes • sec"1 • mm Hg"1 in con-trol dogs.
To determine if similar results could be obtainedon single baroreceptor units, five single fibers fromtwo high NE dogs and five fibers from four normaldogs were examined. The gain, threshold pressure,and saturation pressure for single fiber preparationsof CSN were 1.89±0.4 spikes • sec"' mm Hg"1,97±2.9 mmHg, and 166+17.5 mm Hg in dogs withhigh NE and 0.54±0.11 spikes • sec"1 • mm Hg"1,88±11 mm Hg, and 148±6.6 mm Hg in control dogs,respectively. Thus, there is a consistent increase inthe CSB gain in dogs with chronically elevated NEusing either single or multifiber recordings.
Effects of Chronic Elevation in PlasmaNorepinephrine on Carotid Sinus Wall Functionand the Maximum Firing Rate of CarotidSinus Baroreceptor
As is shown in Figure 4, in dogs with high NE, thecarotid sinus systolic pressure-carotid sinus diametercurve was shifted downwards in a parallel fashioncompared with normal dogs. The slopes of these twocurves were not different (/>>0.05). However, NEproduced a significant change in carotid sinus diam-eter, despite the similar average body weights for thetwo groups (25±2.2 kg and 26±3.6 kg, respectively).Figure 5 shows the relation between carotid sinusdiameter and CSB discharge in these 10 dogs. Theactivity of CSBs in dogs with high NE increasedsignificantly more than that of control dogs despitethe initial smaller diameter, whereas the changes incarotid sinus diameters were similar. Second, dogswith high NE showed a dramatic increase in CSBfiring rate when carotid sinus diameter exceeded 4.3mm; above this diameter, the carotid sinus exhibitedlittle additional deformation as pressure increased.
The maximum adapted firing rate above saturationpressure was measured both in dogs with high NEand in control dogs. Maximum firing rate was
I &A-,
!4JB-
1J 1 NORMAL (n-4)SLOPE -0.012tO.OO2
HtQH NE (n-6)Sl_OPE-0.014»0.0O3
BO 120 260
CAROTID SINUS SYSTOLIC PRESSURE (mmHg)
FIGURE 4. Line graph showing relations between carotidsinus systolic pressure and carotid sinus diameter. Operatingrange of carotid sinus diameter shifted down in dogs withchronically elevated norepinephrine (NE). However, slopes(mm/mm Hg) of these two curves did not change.
counted from the original CSN impulses by twomethods, one using the raster (resetting rate meter)and the second using the Heathkit counter. A strikingincrease in maximum CSN firing rate from dogs withhigh NE was observed. In dogs with high NE, themaximum firing rate was 48 ±4.4 spikes/sec (n = 12)using the raster, whereas in control dogs the maxi-mum firing rate was 24±3.1 spikes/sec (n = 10). Thedifference between dogs with high NE and controldogs was significant (/?<0.01). Similar results werefound in single fiber preparations indicating an in-crease in the gain and maximum firing rate of thecarotid baroreceptors in dogs with chronically ele-vated plasma NE (Table 2).
so-,
40
20-
HKJH NE(n>8)
NOmUL(n-4)
&4
CAROTID SINUS DIAMETER (mm)
FIGURE 5. Line graph showing, in dogs with chronicallyelevated norepinephrine (NE), carotid sinus nerve firing rateincreased from 0 to 58.8±9.9 spikes/sec when the change ofcarotid sinus diameter was 0.44 mm. Whereas, in control dogs,this increase was from 0 to 27.5±3.6 spikes/sec when thechange of carotid sinus diameter was 0.43 mm. Increase incarotid sinus nerve activity did not parallel change of carotidsinus diameter in dogs with chronically elevated NE.
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from
750 Hypertension Vol 17, No 6, Part 1 June 1991
TABLE 2. Increases in Gain and Maximum Firing Rate of CarotidBaroreceptors in Dogs With Elevated Plasma Norepinephrine
Measure
Singlefiber
Windowdis
Heathkitcounter
Conditions
ControlHigh NE
ControlHigh NE
ControlHighNE
Gain(spikes/sec-mm Hg)
0.54 ±0.111.89±0.40*
0.47 ±0.051.99 ±0.45*
Maximumfiring rate
(impulses/sec)
30±2.158±4.2*
24±3.1*48±4.4*
497±17989 ±29
Single fibers control n=5, norepinephrine n=5; window discrim-inator control n=5, norepinephrine n=6; Heathkit counter controln=4, norepinephrine n=6. NE, norepinephrine; dis, discriminator.
*p<0.05 compared with control.
To determine whether there may be a change inthe mechanical function of the carotid sinus in dogswith chronically elevated plasma NE, we calculatedwall strain and plotted this as a function of barore-ceptor activity. Figure 6 shows the relation betweenwall strain and spike activity as arterial pressurechanged using an injection of nitroglycerin and phen-ylephrine. At low-to-intermediate wall strains, therelation between strain and spike activity was shiftedto the right in dogs with chronically elevated plasmaNE; however, at high wall strains baroreceptor activ-ity increased dramatically.
Determination of Morphological Changes in CarotidSinus Wall
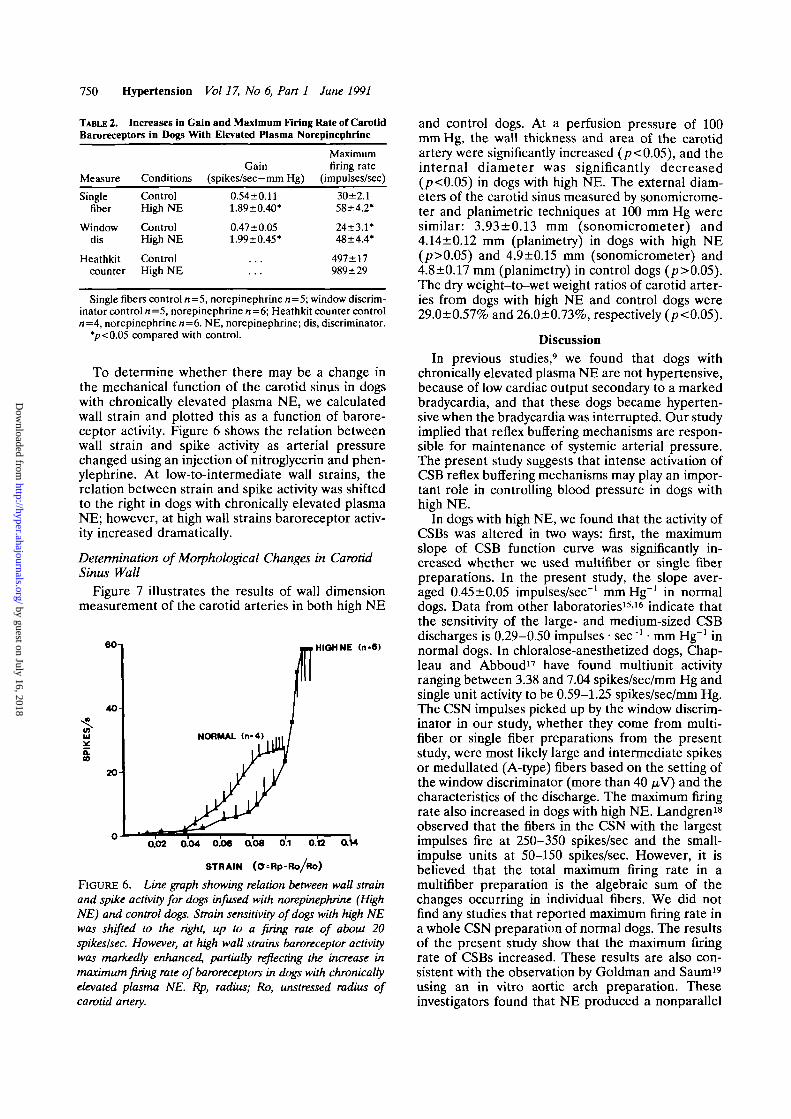
Figure 7 illustrates the results of wall dimensionmeasurement of the carotid arteries in both high NE
eo-i HIGHNE (n-8)
o)ui
0.02 O04 0.06 0.08 0.1 Ott O14
STRAIN (OCRP-
FIGURE 6. Line graph showing relation between wall strainand spike activity for dogs infused with norepinephrine (HighNE) and control dogs. Strain sensitivity of dogs with high NEwas shifted to the right, up to a firing rate of about 20spikes/sec. However, at high wall strains baroreceptor activitywas markedly enhanced, partially reflecting the increase inmaximum firing rate of baroreceptors in dogs with chronicallyelevated plasma NE. Rp, radius; Ro, unstressed radius ofcarotid artery.
and control dogs. At a perfusion pressure of 100mm Hg, the wall thickness and area of the carotidartery were significantly increased (p<0.05), and theinternal diameter was significantly decreased(p<0.05) in dogs with high NE. The external diam-eters of the carotid sinus measured by sonomicrome-ter and planimetric techniques at 100 mm Hg weresimilar: 3.93±0.13 mm (sonomicrometer) and4.14±0.12 mm (planimetry) in dogs with high NE(p>0.05) and 4.9+0.15 mm (sonomicrometer) and4.8±0.17 mm (planimetry) in control dogs (p>0.05).The dry weight-to-wet weight ratios of carotid arter-ies from dogs with high NE and control dogs were29.0+0.57% and 26.0+0.73%, respectively (p<0.05).
DiscussionIn previous studies,9 we found that dogs with
chronically elevated plasma NE are not hypertensive,because of low cardiac output secondary to a markedbradycardia, and that these dogs became hyperten-sive when the bradycardia was interrupted. Our studyimplied that reflex buffering mechanisms are respon-sible for maintenance of systemic arterial pressure.The present study suggests that intense activation ofCSB reflex buffering mechanisms may play an impor-tant role in controlling blood pressure in dogs withhigh NE.
In dogs with high NE, we found that the activity ofCSBs was altered in two ways: first, the maximumslope of CSB function curve was significantly in-creased whether we used multifiber or single fiberpreparations. In the present study, the slope aver-aged 0.45+0.05 impulses/sec"1 mmHg"1 in normaldogs. Data from other laboratories1516 indicate thatthe sensitivity of the large- and medium-sized CSBdischarges is 0.29-0.50 impulses • sec"1 • mm Hg"1 innormal dogs. In chloralose-anesthetized dogs, Chap-leau and Abboud17 have found multiunit activityranging between 3.38 and 7.04 spikes/sec/mm Hg andsingle unit activity to be 0.59-1.25 spikes/sec/mm Hg.The CSN impulses picked up by the window discrim-inator in our study, whether they come from multi-fiber or single fiber preparations from the presentstudy, were most likely large and intermediate spikesor medullated (A-type) fibers based on the setting ofthe window discriminator (more than 40 (JLV) and thecharacteristics of the discharge. The maximum firingrate also increased in dogs with high NE. Landgren18
observed that the fibers in the CSN with the largestimpulses fire at 250-350 spikes/sec and the small-impulse units at 50-150 spikes/sec. However, it isbelieved that the total maximum firing rate in amultifiber preparation is the algebraic sum of thechanges occurring in individual fibers. We did notfind any studies that reported maximum firing rate ina whole CSN preparation of normal dogs. The resultsof the present study show that the maximum firingrate of CSBs increased. These results are also con-sistent with the observation by Goldman and Saum19
using an in vitro aortic arch preparation. Theseinvestigators found that NE produced a nonparallel
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Wang et al Baroreceptor Function and Norepinephrine 751
OO 4.14lO.12mmID1.47±aO9mm
WTl30±0.12mmAREA 11.9±0.9iW!
ID/WT 1.17JO.12
NOREPINEPHRINE
OO 4.8010.17mmID 3.3610.15mmWT 0.77 ±006 mmAREA 9.010.8mm2
NORMAL
FIGURE 7. Schematic showing external ca-rotid artery diameter, internal diameter, wallthickness, and mass of carotid artery weresignificantly different in dogs with chronicallyelevated norepinephrine (High NE) and con-trol dogs. External carotid artery diameter andinternal diameter decreased (p<0.05), andwall thickness and mass increased (p<0.05)in high NE dogs. Data are from six normaldogs and four dogs with chronic NE infusion.
shift in the aortic baroreceptor function curve and anincrease in the maximum firing frequency. The in-crease in slope of CSB function curve in the high NEdogs suggests that the augmented CSB activity entersthe brain stem (especially as pressure rises), and theincrease in maximum firing rate also signifies that themaximum "gain"20 of the CSBs was elevated. Theincreased gain may result in enhanced baroreceptorcontrol of cardiac autonomic activity, may inhibitperipheral sympathetic nervous activity, and facili-tate vagal outflow to promote the reflex lowering ofarterial pressure.
Additional evidence for the elevated vagal toneresulting from the arterial baroreceptor reflex in dogswith high NE was the increase in arterial pressureand tachycardia after pentobarbital anesthesia, asshown in Table 1. Pentobarbital anesthesia wasfound to depress the arterial baroreceptor reflex ofthe dog.21-22 Korner and collaborators demonstrat-ed23 that pentobarbital specifically blocked dience-phalic regions influencing vagal efferents. Becausethe vagal efferent component was completely blockedby pentobarbital anesthesia, the dogs with high NEhad significantly greater increases in mean arterialpressure and heart rate than normal dogs simplybecause of the anesthesia. In other words, consciousdogs with high NE had very high vagal tone. Despitethe relatively small bradycardia in dogs with chronicNE infusion, at a constant stroke volume of about 45ml a reduction in heart rate from 87 to 65 beats/minwould result in a calculated reduction of cardiacoutput from 3,915 to 2,925 ml/min or 25%. Thus, thisincrease in vagal tone can significantly reduce cardiacoutput. Minimum heart rates in dogs without A-Vblock are 40-50 beats/min. These estimates of car-diac output are similar to our previously publishedstudy.9 Another way of describing the level of vagaltone would be to subtract the heart rate in theconscious state from that in the same group afteranesthesia. This difference is 120 beats/min in the NEgroup compared with only 56 beats/min in the controlgroup. This provides an additional estimate of thedegree of vagal tone in dogs with chronic elevation ofplasma NE and of the important role of the barore-ceptor reflexes in the control of arterial pressurethrough altering heart rate and cardiac output.
A question arises concerning whether the differ-ence in arterial pressure between control dogs anddogs with high NE is responsible for the increasedslope that we observed. Tomomatsu and Nishi24 usedthe same concentration of NE (10~9 M) as in ourstudy and found that NE might lower, have no effect,or elevate the threshold pressure of rabbit CSBs invitro. Although an increase in pressure may limit theevaluation of the threshold pressure and saturationpressure of the CSBs, the slope of CSB functioncurve was not altered by the elevated mean arterialpressure. When baroreceptors are reset, the functioncurve is shifted in a parallel fashion with no consis-tent change in gain. Increases in the slope of thefunction curve only occurred at 15 minutes of sus-tained pressure elevation.25 The threshold pressureand slope of the baroreceptor function curve may beindependently regulated and resting blood pressuredoes not affect the receptor gain.13 In the presentstudy, the delay between inducing anesthesia andrecording nerve activity was always longer than 2hours, so there is no reason to suspect a significantinfluence of blood pressure on the slope of CSBfunction curve.
The enhanced sensitivity and maximum firing fre-quency of CSBs could be due to either the directeffect of NE on the CSBs or the indirect effect of NEon the smooth muscle of the carotid sinus wall. Onepossible conclusion of the present study is that NEhad direct biochemical effects on the CSN endings.The parallel downward shift of the carotid sinussystolic pressure-carotid sinus systolic diametercurve in dogs with high NE suggests that the compli-ance of carotid sinus was not affected by NE. Thisimplies that baroreceptors are equally stretched bythe mechanical deformation of carotid sinus wall dueto increasing intrasinus pressure in dogs with NE andnormal dogs. These results are consistent with thereport by Tomomatsu and Nishi24 in the rabbitcarotid sinus where the compliance of the carotidsinus did not change when perfused with the sameconcentration of NE (10~9 M) as we used in ourexperiment (5,000 pg/mlxl pmol/267 pgx 1,000 ml/1=19x10"' M). Carotid sinus diameter decreased inour study, which might be partially due to the vaso-constrictor effect of NE.
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from
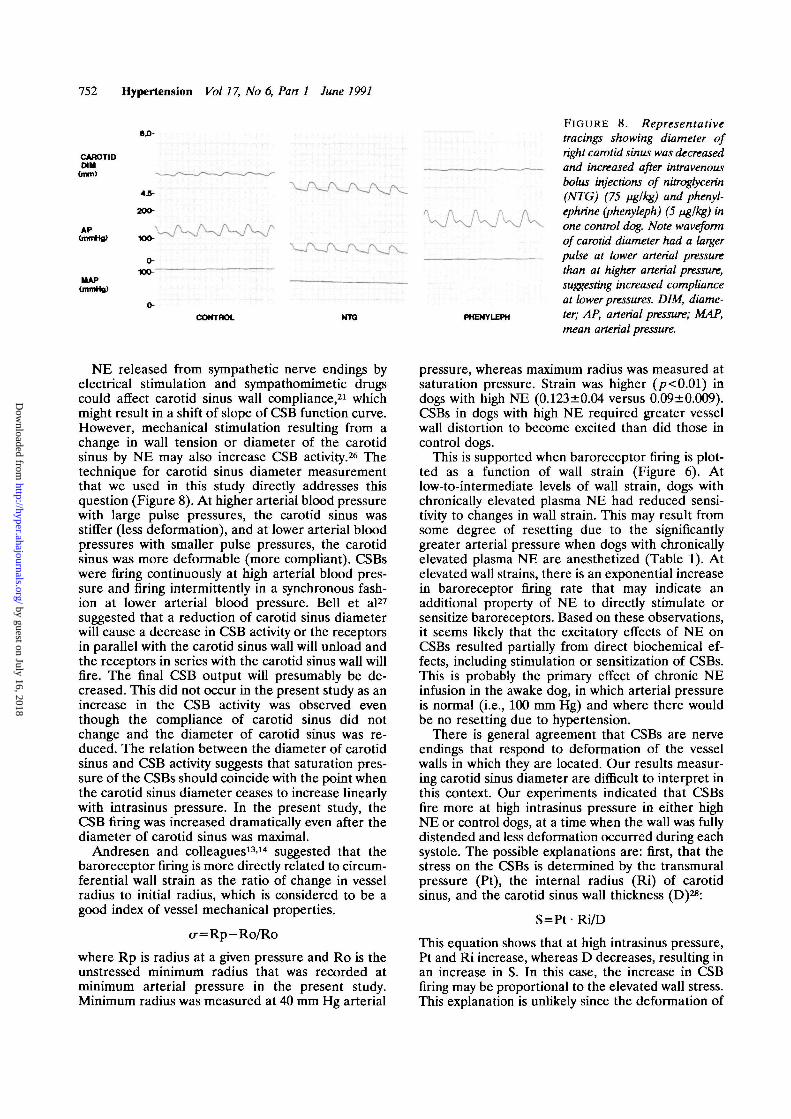
752 Hypertension Vol 17, No 6, Part 1 June 1991
CAROTIDMM
(mm)
AP(jltTTI rQj
MAP
6jO-
200-
100-
o-wo-
0*CONTROL NTQ
F I G U R E 8. Representativetracings showing diameter ofright carotid sinus was decreasedand increased after intravenousbolus injections of nitrogtycerin(NTG) (75 fig/kg) and phenyl-ephrine (phenyleph) (5 fig/kg) inone control dog. Note waveformof carotid diameter had a largerpulse at lower arterial pressurethan at higher arterial pressure,suggesting increased complianceat lower pressures. DIM, diame-ter; AP, arterial pressure; MAP,mean arterial pressure.
NE released from sympathetic nerve endings byelectrical stimulation and sympathomimetic drugscould affect carotid sinus wall compliance,21 whichmight result in a shift of slope of CSB function curve.However, mechanical stimulation resulting from achange in wall tension or diameter of the carotidsinus by NE may also increase CSB activity.26 Thetechnique for carotid sinus diameter measurementthat we used in this study directly addresses thisquestion (Figure 8). At higher arterial blood pressurewith large pulse pressures, the carotid sinus wasstiffer (less deformation), and at lower arterial bloodpressures with smaller pulse pressures, the carotidsinus was more deformable (more compliant). CSBswere firing continuously at high arterial blood pres-sure and firing intermittently in a synchronous fash-ion at lower arterial blood pressure. Bell et al27
suggested that a reduction of carotid sinus diameterwill cause a decrease in CSB activity or the receptorsin parallel with the carotid sinus wall will unload andthe receptors in series with the carotid sinus wall willfire. The final CSB output will presumably be de-creased. This did not occur in the present study as anincrease in the CSB activity was observed eventhough the compliance of carotid sinus did notchange and the diameter of carotid sinus was re-duced. The relation between the diameter of carotidsinus and CSB activity suggests that saturation pres-sure of the CSBs should coincide with the point whenthe carotid sinus diameter ceases to increase linearlywith intrasinus pressure. In the present study, theCSB firing was increased dramatically even after thediameter of carotid sinus was maximal.
Andresen and colleagues1314 suggested that thebaroreceptor firing is more directly related to circum-ferential wall strain as the ratio of change in vesselradius to initial radius, which is considered to be agood index of vessel mechanical properties.
o-=Rp-Ro/Ro
where Rp is radius at a given pressure and Ro is theunstressed minimum radius that was recorded atminimum arterial pressure in the present study.Minimum radius was measured at 40 mm Hg arterial
pressure, whereas maximum radius was measured atsaturation pressure. Strain was higher (p<0.0l) indogs with high NE (0.123±0.04 versus 0.09±0.009).CSBs in dogs with high NE required greater vesselwall distortion to become excited than did those incontrol dogs.
This is supported when baroreceptor firing is plot-ted as a function of wall strain (Figure 6). Atlow-to-intermediate levels of wall strain, dogs withchronically elevated plasma NE had reduced sensi-tivity to changes in wall strain. This may result fromsome degree of resetting due to the significantlygreater arterial pressure when dogs with chronicallyelevated plasma NE are anesthetized (Table 1). Atelevated wall strains, there is an exponential increasein baroreceptor firing rate that may indicate anadditional property of NE to directly stimulate orsensitize baroreceptors. Based on these observations,it seems likely that the excitatory effects of NE onCSBs resulted partially from direct biochemical ef-fects, including stimulation or sensitization of CSBs.This is probably the primary effect of chronic NEinfusion in the awake dog, in which arterial pressureis normal (i.e., 100 mm Hg) and where there wouldbe no resetting due to hypertension.
There is general agreement that CSBs are nerveendings that respond to deformation of the vesselwalls in which they are located. Our results measur-ing carotid sinus diameter are difficult to interpret inthis context. Our experiments indicated that CSBsfire more at high intrasinus pressure in either highNE or control dogs, at a time when the wall was fullydistended and less deformation occurred during eachsystole. The possible explanations are: first, that thestress on the CSBs is determined by the transmuralpressure (Pt), the internal radius (Ri) of carotidsinus, and the carotid sinus wall thickness (D)28:
S=Pt • Ri/D
This equation shows that at high intrasinus pressure,Pt and Ri increase, whereas D decreases, resulting inan increase in S. In this case, the increase in CSBfiring may be proportional to the elevated wall stress.This explanation is unlikely since the deformation of
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Wang et al Baroreceptor Function and Norepinephrine 753
carotid sinus wall during each systole was reduced athigh pressure in our study. Alternatively, a change instrain may be responsible for the enhanced CSBfiring at high intrasinus pressures. Since unstresseddiameter is lower in dogs with chronic NE infusion,maximum strain or the change in strain at largediameters will also be greater leading to the genera-tion of increased CSB activity.
Second, the increased firing rate might be relatedto the arrangement of the CSB within the carotidsinus wall. Presumably, the majority of CSBs are inparallel with the carotid sinus wall and are activatedat high pressure. Only the receptors in series with thecarotid sinus wall keep firing within the low pressurerange. In this regard, Landgren et al29 suggested thatCSBs were "in parallel" with the smooth muscle ofcarotid sinus wall, whereas Bergel et al26 believedthat the receptors were "in series" with the carotidsinus wall. A third reason for the increased firing ratewas suggested by Kirchheim,28 who divided CSNfibers into two groups. The first fiber group firingcoincides with the "high-distensibility region" inwhich the receptor-carrying vessel transforms trans-mural pressure into circumferential stretch. Thesestretch receptors transmit the dynamic componentsof blood pressure: dP/dt. The firing range of thesecond fiber group, according to Kirchheim, coin-cides with the "low-distensibility region" of the re-ceptor-bearing vessel. These units are likely to sensemainly mean pressure. Kirchheim concluded thatbaroreceptor activity is attributed to the mechanicaldeformation of the vessel wall no matter how onedivides the baroreceptor activity. Whether the en-hanced sensitivity in the present study results fromgreater stress, strain, or some specific anatomic ar-rangement of CSB cannot be determined.
Historically, Landgren et al,29 Heymans et al,10 anda number of other investigators have demonstratedthat NE or other catecholamines affect the activity ofbaroreceptors in various species19-24 and in differentexperimental preparations including local adminis-tration of NE on the outside of the carotid sinus28
and perfusion of the carotid sinus with a solutioncontaining NE in either in vivo23 or in vitro18 studies.The results of these observations are consistent withthe hypothesis that NE increases the activity of CSBsdue to either an effect on smooth muscle of thecarotid sinus wall or the direct excitatory effects onCSBs. More recent studies have supported the laterconclusion.19'24-26 Kunze et al30 demonstrated thedirect excitatory effects of NE on the receptor activitywhen the aortic arch was superfused with NE atconcentrations of 10"7 to 10"5 M. Goldman andSaum19 reported similar results (i.e., aortic barore-ceptor activity was significantly increased by 10~7 to10"5 M NE). Similar findings were reported by To-momatsu and Nishi24 who used very low concentra-tions of NE (10~9 M). The concentration of NE usedin the present study was lfT9 M.
Bell et al27 suggested that the physiological signif-icance of the effects of NE on CSBs might reduce the
sympathetic efferent activity and modulate bloodpressure during hemorrhagic hypotension. This hy-pothesis is supported by the present study. NE hastwo opposing effects on modulation of arterial pres-sure, namely increases in myocardial contractility,heart rate, and total peripheral resistance, leading toan increase in arterial pressure. On the other hand,NE also stimulates and sensitizes CSBs resulting inreflex decreases in heart rate, myocardial contractil-ity, and total peripheral resistance due to high vagaltone and the reduction of sympathetic efferent activ-ity. The final arterial pressure will be determined bythe interaction of these two opposing actions and thedegree and duration of hemorrhage. In the dogs withchronically elevated plasma NE, reflex modulationappears to be dominant in maintaining normal arte-rial pressure. This hypothesis may provide a morereasonable explanation for the changes in arterialpressure in some pathological states that are charac-terized by high catecholamine levels. The presentstudy suggests that this feedback system may play animportant role in the regulation of blood pressure,perhaps through the excitatory action on arterialbaroreceptors during high catecholamine states. Theconcept of CSB resetting may also be modified by thepresent study, in that the working range of CSBs isnot only rapidly adjustable to the prevailing pressurebut also that the effects of chronic NE on CSBsprevent systemic hypertension. The influences ofchronic elevation of NE on central nervous systemwere not studied so that some potential effects ofhigh NE on cardiovascular centers could not be ruledout.
Finally, the present study clearly demonstrates thatthe chronic elevation of NE causes morphologicalchanges in the common carotid artery withoutchanges in compliance of the carotid sinus. Catechol-amines can stimulate the growth of vascular smoothmuscle in cell culture.31 Bevan32 reported that sym-pathetic nerves had influences on the development ofnormal and hypertrophied vessels through an arterialpressure-independent action. Casals-Steenzel et al33
and Lever et al34 found that chronic elevation ofplasma NE in rats caused hypertrophy of large arter-ies. The results from our study indicate that increasesin wall thickness and in the area of the commoncarotid artery wall by the chronic elevation of NEwere due to hypertrophy or hyperplasia.
In a recent study, Nejima et al35 reported that dogswith chronic infusion of NE had a slight, but statis-tically insignificant, bradycardia. However, after gan-glionic blockade in all groups of dogs that wereinfused with NE, heart rate increased to a largerdegree compared with control (before pumps). In agroup of cardiac-denervated dogs with chronic NEinfusion, heart rate increased compared with control(before pumps). These studies indicate that somereflex mechanism is responsible for preventing tachy-cardia during NE infusion. Our data suggest that thebaroreceptors are responsible.
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from

754 Hypertension Vol 17, Mo 6, Part 1 June 1991
References1. Pasternac A, Tubau JF, Puddu PE, Krol RB, de Champlain J:
Increased plasma catecholamine levels in patients with symp-tomatic mitral valve prolapse. Am J Med 1982;73:787-790
2. Chidsey CA, Braunwald E, Morrow AG: Catecholamineexcretion and cardiac stores of norepinephrine in congestiveheart failure. Am J Cardiol 1965;39:422-451
3. Thomas JA, Marks BH: Plasma norepinephrine in congestiveheart failure. Am J Cardiol 1978;41:233-243
4. Bristow MR: The adrenergic nervous system in heart failure.N Engl J Med 1984;311:850-851
5. Cryer PE: Physiology and pathophysiology of the humansympathoadrenal neuroendocrine system. N Engl J Med 1980;303:436-444
6. Coghlan HC, Phares P, Cowley M, Copley D, James TN:E>ysautonomia in mitral valve prolapse. Am J Med 1979;67:236-244
7. Goldsmith SR, Francis GS, Levine TB, Cohn JN: Regionalblood flow response to orthostasis in patients with congestiveheart failure. J Am Coll Cardiol 1983;1:1391-1395
8. Gaffney FA, Karlsson ES, Campbell W, Schutte JE, Nixon JV,Willerson JT, Blomqvist CG: Autonomic dysfunction inwomen with mitral valve prolapse syndrome. Circulation 1979;59:894-901
9. King BD, Sack D, Kichuk MR, Hintze TH: Absence ofhypertension despite chronic marked elevations in plasmanorepinephrine in conscious dogs. Hypertension 1987;9:582-590
10. Heymans C, Delaunois AL, Van Den Heuvel-Heymans G:Tension and distensibility of carotid sinus wall, pressoceptorsand blood pressure regulation. Ore Res 1953;l:3-7
11. Blacket BB, Pickering GW, Wilson GM: The effects of pro-longed infusion of noradrenaline and adrenaline on the arte-rial pressure of rabbit. Clin Sci 1950;9:247-257
12. Ishikawa N, Kallman CH, Sagawa K: Rabbit carotid sinusreflex under pentobarbital, urethan, and chloralose anesthe-sia. Am J Physiol 1984;246:H696-H701
13. Andresen MC: Short- and long-term determinants of barore-ceptor function in aged normotensive and spontaneouslyhypertensive rats. Ore Res 1984^4:750-759
14. Andresen MC, Kurawska S, Brown AM: Baroreceptor func-tion and changes in strain sensitivity in normotensive andspontaneously hypertensive rats. Ore Res 1980;47:821-828
15. Diamond J: Observations on the excitation by acetylcholineand by pressure of sensory receptors in the cat's carotid sinus.J Physiol (Lond) 1955;130:513-532
16. Frang GN, Scher AM, Ito CS: Small signal characteristic ofcarotid sinus baroreceptors of rabbits. JAppI Physiol 1971 ;30:527-535
17. Chapleau MW, Abboud FM: Contrasting effects of static andpulsatile pressure on carotid baroreceptor activity in dogs. OreRes 1987;61:648-658
18. Landgren S: On the excitation mechanism of the carotidbaroreceptors. Ada Physiol Scand 1952;26:l-35
19. Goldman WF, Saum WR: A direct excitatory action of cate-cholamines on rat aortic baroreceptors in vitro. Ore Res1984^5:18-30
20. Sagawa K: Concerning "gain." Am J Physiol 1978;235:H117-H121
21. Peiss CN, Manning JW: Effects of sodium pentobarbital onelectrical and reflex activation of the cardiovascular system.Ore Res 1964;14:228-235
22. Vatner SF, Braunwald E: Cardiovascular control mechanismsin the conscious state. N Engl J Med 1975;293:970-976
23. Korner PI: Integrative neural cardiovascular control. PhysiolRev 1971^1:312-367
24. Tomomatsu E, Nishi K; Increased activity of carotid sinusbaroreceptors by sympathetic stimulation and norepinephrine.Am J Physiol 1981;240:H650-H658
25. Undesser KP, Lynn MP, Bishop VS: Rapid resetting of aorticnerves in conscious rabbits. Am J Physiol 1984;246:H302-H305
26. Bergel DH, Anand IS, Brooks DE, MacDermott AJ, PevelierRC, Robinson JL, Sleight P: Carotid sinus wall mechanics andbaroreceptor function in the dog, in Sleight P (ed): ArterialBaroreceptors and Hypertension. New York, Oxford UniversityPress, 1980, pp 1-4
27. Bell LB, Seagard JL, Zuperku EJ, Kampine JP: Mechanicaleffects of vasoactive drugs on carotid sinus. Am J Physiol1986;250:R1074-R1080
28. Kirchheim HR: Systemic arterial baroreceptor reflex. PhysiolRev 1976^6:100-175
29. Landgren S, Neil E, Zotterman Y: The response of the carotidbaroreceptors to the local administration of drugs. Acta PhysiolScand 1952;26:24-37
30. Kunze DL, Krauhs JM, Orlea CJ: Action of norepinephrineon aortic baroreceptor of rat adventia. Am J Physiol 1984;247:H811-H816
31. Blaes N, Boissel JP: Growth-stimulating effect of catechol-amines on rat aortic smooth muscle cells in culture. J CellPhysiol 1983;116:167-172
32. Bevan RD: Trophic effects of peripheral adrenergic nerves onvascular structure. Hypertension 1984;6(suppl III):III-27-III-32
33. Casals-Steenzel J, Tree M, Brown JJ, Fraser R, Lever AF,Millar JA, Morton JJ, Robertson JIS, Reid JL, Hamilton C:Effects of prolonged and brief infusions of noradrenaline onarterial pressure and on the plasma concentrations of activerenin, angiotensin II, aldosterone and potassium. J Hypertens1983;l:27-35
34. Lever AF: Slow pressor mechanisms in hypertension: A rolefor hypertrophy of resistance vessels. / Hypertens 1986;4:515-524
35. Nejima J, Vemura N, Vatner DE, Homay CJ, Hintze TH,Vatner SF: Role of intact cardiac nerves and reflex mecha-nisms in desensitization to catecholamines in conscious dogs. /Clin Invest 1990;86:2046-2053
KEYWORDS • baroreceptors • norepinephrine • hypertrophy• compliance • dog studies
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from

J Wang, M Ochoa, M B Patel, I H Zucker, A V Loud, G A Zeballos and T H HintzeCarotid baroreceptor function in dogs with chronic norepinephrine infusion.
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 1991 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.17.6.745
1991;17:745-754Hypertension.
http://hyper.ahajournals.org/content/17/6_Pt_1/745World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer process is available in the
Request Permissions in the middle column of the Web page under Services. Further information about thisOffice. Once the online version of the published article for which permission is being requested is located, click
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertension Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 16, 2018http://hyper.ahajournals.org/
Dow
nloaded from