cellular and genetic basis for suppression of cytotoxic t cell generation by haloaromatic...
TRANSCRIPT

Cellular and Genetic Basis for Suppression of Cytotoxic T Cell Generation by Haloaromatic Hydrocarbons
David A. Clark, George Sweeney, Steven Safe, Elizabeth Hancock, Douglas G. Kilburn, and Jack Gauldie
Abstract: Generation of allospecific cytotoxic T cells in C57BI/6 mice is significantly impaired following exposure to TCDD at doses as low as 4 ng/kg. T helper activity, as assessed by the ability to produce Interleukin 2, and frequency of CTL precursors appear unaffected in treated animals and thus the TCCD-induced suppressor cells we have described are primarily responsible for the reduction in the CTL response. Suppression of CTL generation in DBA/2 mice requires a lO-lO0-fold greater dose than in C57B1/6, consistent with the observation that the Ah locus gene(s) of DBA/2 mice code for TCDD receptors with low binding affinity for TCDD. Other haloaromatic hydrocarbons (3,3'4,4'-tetrachlorobiphenyl and Aroclor 1254), capable of binding to the TCDD receptor, also suppress CTL generation, whereas the 2,2' ,4,4' ,6,6'-hex- achlorobiphenyl molecule that lacks affinity for the TCDD receptor does not suppress CTL. The immunotoxic effects of TCDD in C57B/6 and DBA/2 mice occur at dose levels below those required to induce mixed-function oxidase enzymes in the liver. Suppression of CTL by TCDD is associated with increased susceptibility to lethal herpes virus type II infection. These data suggest that low levels of TCDD may interact with cytoplasmic receptors for TCDD in the thymus and induce biologically significant immunosuppression through activation of suppressor cells.
Key Words. Suppressor cells; Cytotoxic T lymphocytes; TCDD; Haloaromatic hydrocarbons; Environmen- tal toxins; Herpes virus infection
I N T R O D U C T I O N
A variety of chemical agents can suppress immune responsiveness; recently, environmental contaminants such as 2, 3, 7, 8 tetrachlorodibenzo-p-dioxin (TCDD), polychorinated biphenyls (PCBs), and polybrominated byphenyls (PBBs) have been found to be immunosuppressive (Faith et al., 1978; Vos et al., 1980). The cellular and molecular basis of this immunotoxicity is unclear.
Received November 3, 1982; revised and accepted March 29, 1983. From the Departments of Medicine, Pathology, and Biochemistry, Host Resistance Program, McMaster
University, Hamilton, Ontario, Canada, L8N 4._19. (D.A.C.; G.S.; J.G.), the Department of Chemistry, University of Guelph, Guelph, Ontario, N I G 2W1 (S.S.), and the Department of Microbiology and Immunol- ogy, University of British Columbia, Vancouver, B.C., V6T lW5. (D.G.K.; E.H.).
Address reprint requests to Dr. David A. Clark, Department of Medicine and Host Resistance Program, McMaster University, 1200 Main Street West, Hamilton, Ontario L8N 3Z5 Canada.
© Elsevier Science Publishing Co., Inc., 1983 143 52 Vanderbilt Ave., New York, N.Y. lmmunopharmacology 6, 143-153 ( 1 9 8 3 ) 0162-3109/83/$03.00

144 D.A. Clark et al.
We have shown that low doses of TCDD (4 ng /kg- 0.4/.~g/kg) suppress the generation of cytotoxic T cells (CTL) by lymph node and spleen cells in male C57B1/6 mice while having little effect on delayed hypersensitivity, antibody responses, or thymus cellularity (Clark et al., 1981). Sup- pressor cells capable of blocking CTL response were found in the thymus of TCDD-treated mice following cumulative doses of TCDD as low as 4 ng/kg. In this paper we report that susceptibility to TCDD suppression in mice is strain-dependent, occurs at doses that have little effect on hepatic microsomal enzymes, and that other types of haloaromatic hydrocarbons can induce similar immune suppression provided they possess sufficient binding affinity for the genetically controlled TCDD receptor protein. These observations suggest a receptor-dependent mechanism for the stimulation of suppressor T cell activity by haloaromatic hydrocarbons.
MATERIALS AND METHODS
Mice
Male C57BI/6, DBA/2, and CBA strain mice were purchased from the Jackson Laboratory (Bar Harbor, ME), were housed 5 - 6 per cage and were allowed Purina chow and water ad libitum. C3H/HeJ x DBA/2JF I (C3D2FI) mice were bred in our own colony. Nude mice were obtained from the Ontario Veterinary College (Guelph, Ontario). P815 (H-2 d) and EL4 (H-2 b) tumor lines were maintained by serial ascites transfer in DBA/2 and C57BI/6 mice, respectively.
Haloaromatic Hydrocarbon Treatment
TCDD 99% pure, provided by the Dow Chemical Company (Sarnia, Ontario), was diluted in corn oil and 0.1 ml was injected i.p. into the mice at weekly intervals for a total of 4 doses. The treated mice were used 7 - 2 2 days after the last injection. Control animals received 0.1 ml corn oil alone i.p. weekly for 4 weeks. In some experiments, other haloaromatic hydrocarbons were used in place of TCDD. These included Aroclor 1254, a commercial PCB mixture previously rendered free of ch lor ina ted dibenzofurans; 3 , 3 ' , 4 , 4 ' - t e t r a c h l o r o b i p h e n y l (3 ,3 ' , 4 ,4 ' -TCB) ; and 2,2',4,4' ,6,6'-hexachlorobiphenyl (2,2',4,4',6,6'-HCB). The method of preparation and testing of these compounds has been described elsewhere (H utzinger et al., 1979; Parkinson et al., 1980).
Lymphocyte Cultures
Peripheral lymph nodes (axillary + brachial + inguinal) (PLN) and spleens were aseptically removed and single cell suspensions prepared in tissue culture medium as described elsewhere (Clark et al., 1981). Culture medium consisted of a-MEM supplemented to a concentration of 5 x 10-SM 2-mercaptoethanol, 100 IU/ml penicillin, 100 ps/ml streptomycin, and 10% v/v fetal bovine serum (Grand Island Biol. Co., Grand Island, NY). Two types of mixed lymphocyte cultures (MLC) were used to generate CTL. One million to 3 x 106 responding spleen or lymph node cells were cultured with 1 x 106 allogeneic stimulator cells (inactivated with 1500 R 6°Co) in 3.2 ml medium in 17 x 100 mm polystyrene tubes at 37°C in 6 - 7 % CO 2. After 5 days, the cell pellet in the cultures was resuspended in 1 ml medium, serially diluted and added to fiat-bottom 96-well microtrays together with 1 x 10451Cr-labeled target cells. Cytotoxic activity (equivalent to lytic units) was calculated as described elsewhere (Clark et al., 1977). Limiting dilution cultures to
Abbreviations. TCDD: 2,3,7,8 tetrachlorodibenzo-p-dioxin; PBB: polybrominated biphenyl; CTL: cytotoxic T lymphocytes; Con A: concanavalin A; C3D2FI: C3H x DBA 2 F1; i.p.: intraperitoneal; IU: International units; ERR: ethoxyresorufin-O-deethylase; PFU: plaque-forming units; MLC: mixed lymphocyte culture; PLN: peripheral lymph nodes; CTLP: precursors of CTL; MFO: mixed-function oxidase; MEM: minimum essential medium; TCB: tetrachlorobiphenyl; HCB: hexachlorobiphenyl.

CTL Suppression by Haloaromatic Hydrocarbons 145
determine the frequency of precursors of CTL were performed in V-bottom microtrays with 2 × 105 irradiated stimulator cells and 2 × 10 s nu/nu helper cells. After 6 days, the presence of CTL in individual culture wells was tested by adding 2 x 103 SlCr-labeled targets (Teh et al., 1977). The frequency of CTL precursors was calculated from the Poisson formula using a computer program to determine the maximum likelihood estimate, as described by Porter and Berry (1964).
The ability of spleen cells to produce Interleukin 2 (costimulator) was determined by culturing 1 x 107spleen cells in Linbro 24-well plates in 2 ml RMP11640 medium containing antibiotics, 5 x 10-SM 2 mercaptoethanol, and 10% v/v fetal bovine serum and 4 pJg/ml con A for 18 hr at 37 ~C in 5% CO 2. Cell-free supernatants were harvested and added at different dilutions to triplicate 200/~I cultures in 96-well fiat-bottom plates containing I x los CBA/J thymocytes and 2/.tg/ml con A. After 3 days incubation at 37°C, 0.5 / ~ i tritiated thymidine was added; 9 hr later, isotope incorporation was measured by liquid scintillation. Under the conditions of the assay, significant stimulation of isotope incorporation by con A only occurred when supernatant containing co- stimulator was present.
Enzyme Assays The microsomal fraction was isolated from freshly obtained livers by ultracentrifugation. The activity of the mixed function oxidase ethoxyresorufin-O-deethylase (ERR) was measured as described elsewhere (Burke and Mayer, 1974).
Herpes Virus Challenge Herpes simplex type lI strain 333 was grown in VERO cells and the number of infectious units/ml was determined by plaque information on VERO cells (Rawls, 1979). Preliminary dose-response curves showed a 5 0 - 8 0 % mortality when 5 - 8 x l0 s PFU was given i.p. to C57B1/6 male mice. The infected mice began to die after 7 days and deaths ceased by 16 days, at which point the number of survivors was determined.
RESULTS
Inhibition of In Vitro Generation of CTL by TCDD Treatment
Previous studies demonstrated that exposure to 4 ng /kg-400 ~g/kg TCDD significantly impaired the generation of alloantigen-stimulated CTL from PLN and splenic lymphocytes of C57BI/6 male mice in vitro (Clark et al., 1981). Figure I illustrates typical titration curves from the 51Cr-release assay done on CTL generated in in vitro MLC tube cultures containing 3 x 106 lymph node cells (PLN) stimulated by C3D2F I splenocytes. It can be readily seen that PLN from mice given 4 - 4 0 ng/kg TCDD generated suboptimal cytotoxicity and that this impairment disappeared when the dose was reduced to 0.4 ng/kg. The cellular basis for the defect in CTL generation is further explored in the experiment shown in Table I. It can be seen that the frequency of CTL precursors (CTLP) was not reduced by TCDD treatment but, rather, the ability of CTLP to develop into CTL in the tube MLC cultures was impaired. The magnitude of this impairment was gauged by calculating the yield of CTL per CTLP added to the tube MLC cultures.
Susceptibility of Resistant DBA/2 Strain Mice to TCDD Impairment of CTL Generation
There is strong evidence that TCDD exerts its toxic effects on a variety of tissues by interaction with a specific cytoplasmic receptor (Poland and Knutson, 1982; Knutson and Poland, 1982). The binding affinity between the receptor and TCDD appears to be genetically controlled, and there are several mouse strains which have low or undetectable levels of receptor, as determined by

146 D. A. Clark et al.
5 0 -
i,u
UJ ,-I I.k/ n- n- r.
w a. (/) I - z UJ
I,U el.
40
30
20
10
/ / /
I l l l~
/ / / / / /
/ /" !" z/ /I .,,
/ , , /--......" II/" / / ._...'"" / / ./ ........
/ / o,°" /L..-"" ~ 7 . / .....""
~ J " . / . .." . , , . 1 1 - / / . / . . . ' "
/ v f .0o ,°
i ~ / / , ~ "..~.~." ~ooOO°
~:...-
O - I I i / 1:8 1:4 1:2 1:1
CULTURE DILUTION TESTED
Figure I Cytotoxic activity generated in tube MLC cultures in 4 hr 51Cr-release assay. 3 x 106 lymph node cell from C5 7131/6 control mice (0) or mice given total TCDD doses of O. 4 ng/kg (~), 4 ng/kg (x), or 40 ng/kg (~) were cultured with 106 irradiated C3D2F 1 spleen cells.
binding of radiolabeled TCDD (Mason and Okey, 1982). Low binding affinity is associated with reduced sensitivity to the teratogenic effects of TCDD, with greater resistance to induction of hepatic mixed-function oxidase enzymes (MFO), such as ethoxyresorulin-o-deethylase (ERR), by TCDD (Poland and Knutson, 1982, Poland and Glover, 1980; Knutson and Poland, 1982), and with greater resistance to the hepatotoxic effects of TCCD (Jones and Sweeney, 1980).
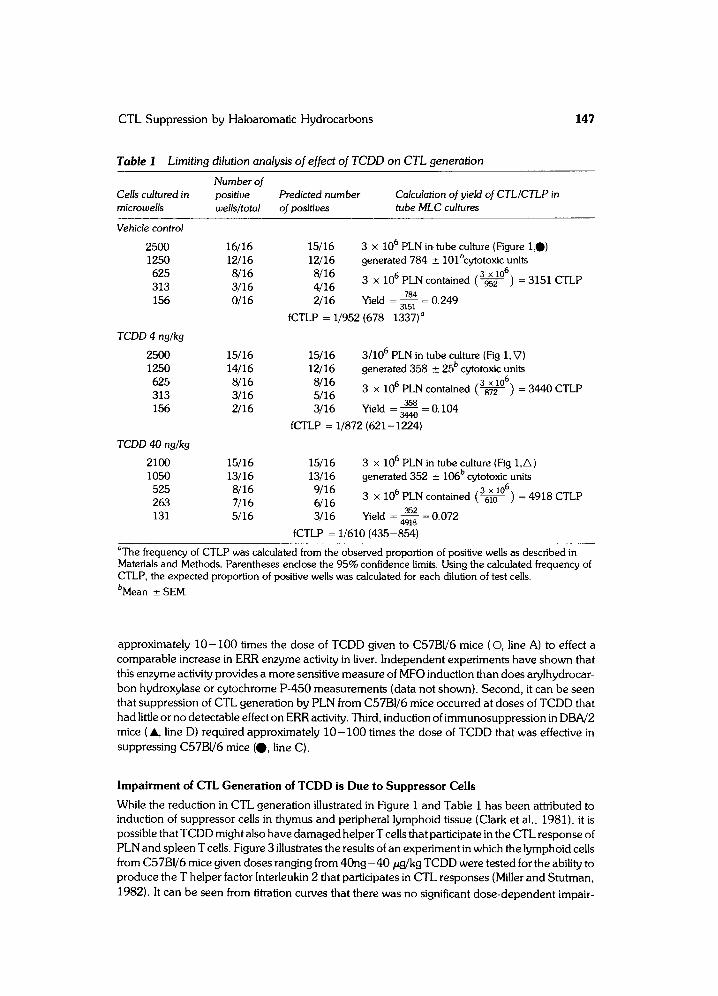
Figure 2 illustrates the difference in TCDD sensitivity between the TCDD-susceptible C57BI/6 and resistant DBA/2 strains. It can be readily seen that DBA/2 recipients (& line B) required
CTL Suppression by Haloaromatic Hydrocarbons 147
Table I Limiting dilution analysis of effect of TCDD on CTL generation
Number of Cells cultured in pos i t i ve Predicted number microwells wells~total of positives
Calculation of yield of CTL/CTLP in tube MLC cultures
Vehic~ control
2500 16/16 1250 12/16 625 8/16 313 3/16 156 0/16
TCDD 4 ng/kg
2500 15/16 1250 14/16 625 8/16 313 3/16 156 2/16
TCDD 40 ng/kg
2100 15/16 1050 13/16 525 8/16 263 7/16 131 5/16
15/16 3 × 106 PLN in tube culture (Figure 1,O) 12/16 generated 784 ± 101%ytotoxic units 8/16 106 / ~ I N
4/16 3 x PLN contained ~3~5 ----~°6) = 3151 CTLP 784
2/16 Yield = 3~- = 0.249
fCTLP = 1/952 (678-1337) °
15/16 3/106 PLN in tube culture (Fig i, V) 12/16 generated 358 + 25 b cytotoxic units 8/16 106 f x l ~ 5/16 3 x PLN contained ( 3 ~ ---~06) = 3440 CTLP
358 3/16 Yield = ~ = 0.104
fCTLP = 1/872 (621-1224)
15/16 3 x 106 PLN in tube culture (Fig 1,A) 13/16 generated 352 ± 106 b cytotoxic units 9/16 106 6/16 3 × PLN contained ~3~°6~ = 4918 CTLP
352 3/16 Yield = ~ = 0.072
fCTLP = 1/610 (485-854)
aThe frequency of CTLP was calculated from the observed proportion of positive wells as described in Materials and Methods. Parentheses enclose the 95% confidence limits. Using the calculated frequency of CTLP, the expected proportion of positive wells was calculated for each dilution of test cells. bMean ~ SEM.
approximately 1 0 - I00 times the dose of TCDD given to C57BI/6 mice ( ©, line A) to effect a comparable increase in ERR enzyme activity in liver. Independent experiments have shown that this enzyme activity provides a more sensitive measure of MFO induction than does arylhydrocar- bon hydroxylase or cytochrome P-450 measurements (data not shown). Second, it can be seen that suppression of CTL generation by PLN from C57BI/6 mice occurred at doses of TCDD that had little or no detectable effect on ERR activity. Third, induction of immunosuppression in DBA/2 mice ( A, line D) required approximately 10 -100 times the dose of TCDD that was effective in suppressing C57BI/6 mice (O, line C).
Impairment of CTL Generation of TCDD is Due to Suppressor Cells
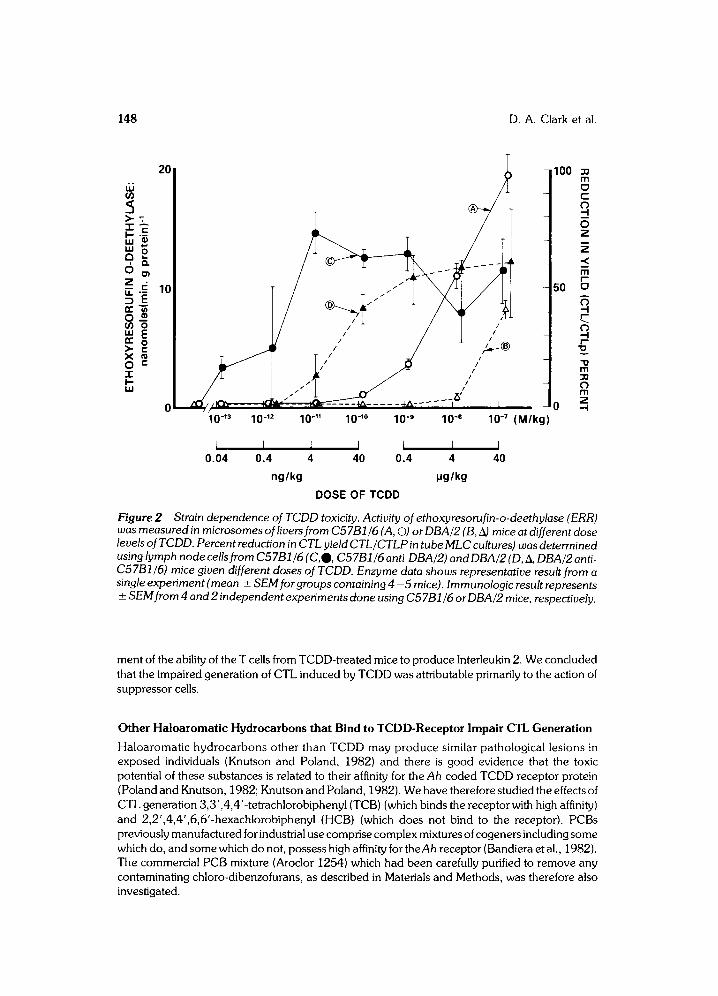
While the reduction in CTL generation illustrated in Figure 1 and Table 1 has been attributed to induction of suppressor cells in thymus and peripheral lymphoid tissue (Clark et al., 1981), it is possible that TCDD might also have damaged helper T cells that participate in the CTL response of PLN and spleen T cells. Figure 3 illustrates the results of an experiment in which the lymphoid cells from C57BI/6 mice given doses ranging from 40ng - 40 pg/kg TCDD were tested for the ability to produce the T helper factor Interleukin 2 that participates in CTL responses (Miller and Stutman, 1982). It can be seen from titration curves that there was no significant dose-dependent impair-
D. A. Clark et al.
4
"T
0 ~ Z •
O _ • 0
×~ 0 z I,--
2 0 -
10 -13
148
10 -12 10"" 10 -1° 10 .9 10 "8
- 100
50
0 10 -7 (M/kg)
:;0 I"11
C C~ "-I
Z
-<
r -
"-I I '--
--I I " "0
"0 m
¢3 1'13
Z --I
• I J I I I I 0.04 0.4 4 40 0.4 4 40
ng/kg pg/kg
DOSE OF TCDD
Figure 2 Strain dependence of TCDD toxicity. Activity of ethoxyresorufin-o-deethylase (ERR) was measured in microsomes of livers from C5 7131/6 (A, ©) or DBA/2 (B, ~ mice at different close levels of TCDD. Percent reduction in CTL yield CTL /CTLP in tube ML C cultures) was determined using lymph node cells from C5 7131/6 (C,0, C5 7131/6 anti-DBA/2) and DBA/2 ( D, A, DBA /2 anti- C57B1/6) mice given different doses of TCDD. Enzyme data shows representative result from a single experiment (mean +_ SEM for groups containing 4 -5 mice). Immunologic result represents +_ SEM from 4 and 2 independent experiments done using C5 7B1/6 or DBA/2 mice, respectively.
ment of the ability of the T cells from TCDD-treated mice to produce |nterleukin 2. We concluded that the impaired generation of CTL induced by TCDD was attributable primarily to the action of suppressor cells.
Other Haloaromat ic Hydrocarbons that Bind to TCDDoReceptor Impair CTL Generation
Haloaromatic hydrocarbons other than TCDD may produce similar pathological lesions in exposed individuals (Knutson and Poland, 1982) and there is good evidence that the toxic potential of these substances is related to their affinity for the Ah coded TCDD receptor protein (Poland and Knutson, 1982; Knutson and Poland, 1982). We have therefore studied the effects of CTL generation 3,3',4,4'-tetrachlorobiphenyl (TCB) (which binds the receptor with high affinity) and 2,2',4,4',6,6'-hexachlorobiphenyl (HCB) (which does not bind to the receptor). PCBs previously manufactured for industrial use comprise complex mixtures of cogeners including some which do, and some which do not, possess high affinity for theAh receptor (Bandiera et al., 1982). The commercial PCB mixture (Aroclor 1254) which had been carefully purified to remove any contaminating chloro-dibenzofurans, as described in Materials and Methods, was therefore also investigated.

CTL Suppression by Haloaromatic Hydrocarbons 149
LI. O l- z
O
60
50
40
30
20
m
1 0 -
0 0
I 1/2 1/4 1/8 1/16 1/32 1/64
DILUTION OF TEST SUPERNATANT
Effect of TCDD on production of helper factor. Different dilutions of supematants Figure 3 prepared by con A - stimulation of spleen cells from C57B1/6 control mice (0) or mice given 40 ng/kg TCDD (0), 0.4 Ixg/kg TCDD ( A) , 4 p~g/kg TCDD (El), or 40 p.g/kg ( • TCDD were added) to CBA/J thymocytes together with con A.The stim ulation of thymidine incorporation ( CPM with con A minus CPM without) is plotted on the abscissa. Significant incorporation occurred only in the presence of both con A and supematant.
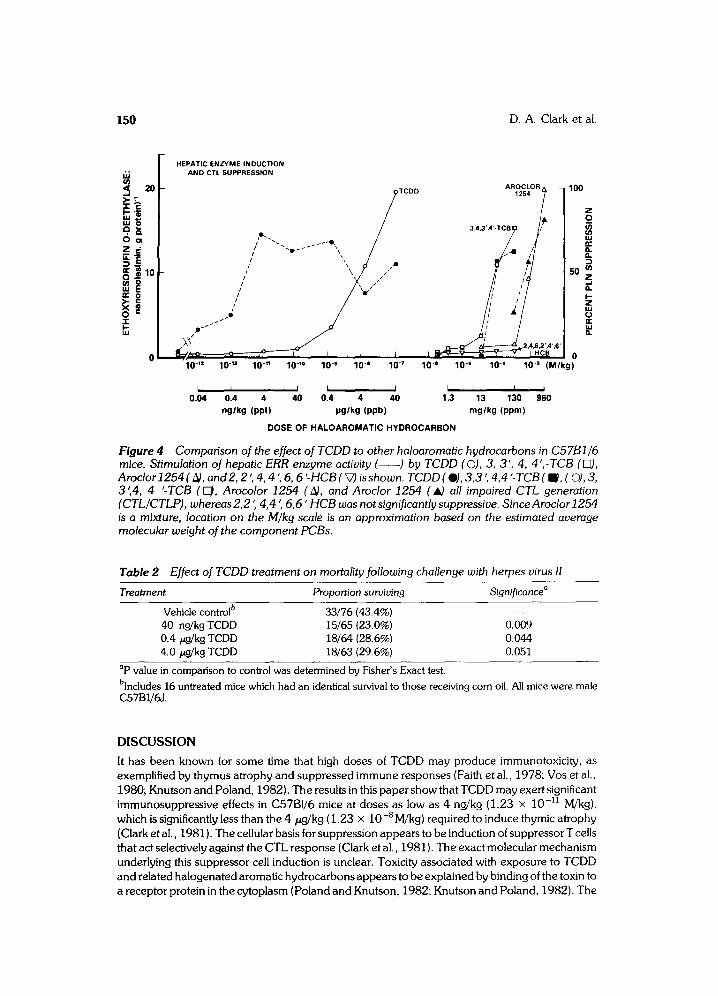
Figure 4 compares the effect of different doses of these compounds with the effect of TCDD in C57BI/6 mice. Aroclor 1254, as well as 3,3', 4,4'-TCB, induced hepatic ERR and suppressed CTL generation. In contrast, 2,2' ,4,4' , 6,6'-HCB did not induce either a detectable ERR response or produce suppression. Taken together with the data in Figure 2, we concluded that the immu- nosuppressive effects of TCDD and related haloaromatic hydrocarbons, like the induction of MFO activity, are probably mediated through interaction with receptor protein coded by the Ah locus.
Effect of TCDD on Susceptibility to Herpes Simplex II Virus
It has been proposed that CTLs play a role in defense against certain types of viruses, such as herpes simplex and cytomegalovirus (Quinan et al, 1982). To test for an in vivo effect of immunosuppression by low doses of TCDD, we examined the effect of TCDD treatment on susceptibility to lethal herpes virus type II infection. Table 2 shows that mortality was significantly increased by the 40 ng/kg dose. A less pronounced, but still significant, effect was seen at 0.4 and 4pg/kg dose. These data suggest that low doses of TCDD might increase susceptibility to certain types of viral infections.
150 D. A. Clark et al.
¢/) <~ 20
HEPATIC ENZYME INDUCTION AND CTL SUPPRESSION
~ \ ~ T C D D C~
3,4,3',4'-T ,=.
/ ", "J ~ ~~~H,I I I i/ i / ~,/J' .... A ' ~--t t ' : -0 , I I I
":I0"" 10 -'z 10"11 10 4o 10-' I 0 " I 0 " 10"'
AROCLOR 100 1254 z O
UJ O. O.
50 ca z a . i,- z
Ix:
1',6 0
10 " 10-" 10 "z (M/kg)
I I I I I i I I I I J 0.04 0.4 4 40 0.4 4 40 1.3 13 130 960
nglkg (ppt) pg/kg (ppb) mg/kg (ppm)
DOSE OF HALOAROMATIC HYDROCARBON
Figure 4 Comparison of the effect of TCDD to other haloaromatic hydrocarbons in C57B1/6 mice. Stimulation of hepatic ERR enzyme activity ( ) by TCDD (©), 3, 3', 4, 4 ',-TCB ([E]), Aroclor 1254 (~), and2, 2 ', 4, 4 ', 6, 6 '-HCB (V) is shown. TCDD (0), 3,3 ', 4,4 '- TCB (m), ((3), 3, 3',4, 4 '-TCB ( ~ , Arocolor 1254 ( ~), and Aroclor 1254 ( A) all impaired CTL generation (CTL/CTLP), whereas2,2 ', 4,4 ', 6,6 ;HCB was notsignificantlysuppressive. SinceAroclor J254 is a mixture, location on the M/kg scale is an approximation based on the estimated average molecular weight of the component PCBs.
Table 2 Effect of TCDD treatment on mortality following challenge with herpes virus II
Treatment Proportion surviving Significance a
Vehicle control b 33/76 (43.4%) - - 40 ng/kg TCDD 15/65 (23.0%) 0.009 0.4 k~g/kg TCDD 18/64 (28.6%) 0.044 4.0/~g/kg TCDD 18/63 (29.6%) 0.051
ap value in comparison to control was determined by Fisher's Exact test. blncludes 16 untreated mice which had an identical survival to those receiving corn oil. All mice were male C57B1/6J.
D I S C U S S I O N
It has been known for some time that high doses of TCDD may produce immunotoxicity, as exemplified by thymus atrophy and suppressed immune responses (Faith et al., 1978; Vos et al., 1980; Knutson and Poland, 1982). The results in this paper show that TCDD may exert significant immunosuppressive effects in C57BI/6 mice at doses as low as 4 ng/kg (1.23 × I0 -11 M/kg), which is significantly less than the 4/~g/kg (1.23 × 10-8 M/kg) required to induce thymic atrophy (Clark et al., 1981). The cellular basis for suppression appears to be induction of suppressor T cells that act selectively against the CTL response (Clark et al., 1981). The exact molecular mechanism underlying this suppressor cell induction is unclear. Toxicity associated with exposure to TCDD and related halogenated aromatic hydrocarbons appears to be explained by binding of the toxin to a receptor protein in the cytoplasm (Poland and Knutson, 1982; Knutson and Poland, 1982). The

CTL Suppression by Haloaromatic Hydrocarbons 151
level or affinity of this receptor is controlled in mice by a gene or genes at the Ah locus, and the cells in thymus and liver appear to contain high concentrations of the receptor (Mason and Okey, 1982). Our studies indicate that the DBA/2 strain that is genetically more resistant to TCDD toxicity, as measured by teratogenesis and MFO induction (Poland and Knutson, 1982~ Poland and Glover, 1980~ Knutson and Poland, 1982), is also more resistant to TCDD-induced immu- nosuppression. Furthermore, the ability of other haloaromatics to induce immunosuppression appears to relate to their ability to bind the Ah-coded cytoplasmic TCDD receptor molecule (Poland and Knutson, 1982~ Knutson and Poland, 1982~ and Figure 4). One must note that susceptibility and resistance are relative terms and a sufficient dose of haloaromatic may induce toxicity. For example, TCDD-resistant mice that lack detectable cytosolic receptors possess nuclear receptors for TCDD (Okey et al., 1979; Mason and Okey, 1982). The latter may explain the immunotoxicity achieved in DBA/2 mice given TCDD in Figure 2. More precise data on the role of the Ah locus in immunotoxicity will be obtained by studies of Ah d congenic C57BI/6 mice and B6D2F I × D2 backcross progeny.
We observed some interesting differences between the effects of TCDD and other haloaroma- tic hydrocarbons, illustrated by Figure 4. The immunosuppressive effects of TCDD, unlike that produced by 3,3 ', 4,4 '-TCB and Aroclor 1254, occurred at doses substantially lower than those required to stimulate ERR enzyme levels in liver. This observation has not yet been explained, but could be caused by differences in organ distribution between TCDD and PCBs or by differences in their half lives. For example, our preliminary studies indicate that the onset of immunosuppression following a single injection of TCDD occurs after a delay of at least 7 days (unpublished data), whereas induction of MFO in liver mediated by the A/a-coded cytosolic receptor occurs within 2 days (Poland et al., 1974). One might expect that a haloaromatic with a short half life in thymus and liver would cause less of an effect on thymus than on liver. Precise measurements concerning the half life of 3,3 ',4,4'-TCB in our mice are not known, and data on half life cannot be provided for mixtures such as Aroclor 1254 to allow comparison to TCDD. A second factor that must be considered in interpreting Figure 4 is the observation that induction of MFO enzymes, such as AHH, in a wide variety of cells is not necessarily associated with observable toxicity (Knutson and Poland, 1982). These investigators have therefore proposed that the toxic effects of TCDD and related haloaromatics may occur by activation of other genes (linked to the murine Ah locus) which regulate cellular differentiation. These latter effects show considerable tissue and species specificity, and this observation could explain the remarkable sensitivity of the thymus of C57BI/6 mice to TCDD even though the level of MFO induction in the thymus has been reported to be only 0.1% that detected in liver tissue (Mason and Okey, 1982). We have recently studied sensitivity to the 4 ng/kg dose of TCDD in bone marrow chimeras in which genetically TCDD-resistant lymphomyeloid DBA/2 cells were used to reconstitute genetically TCDD-susceptible C57BI/6 hosts, and vice versa. We observed that susceptibility to TCDD is determined by the radioresistant cells of the recipient (submitted for publication). It seems likely, therefore, that TCDD may act through receptors in thymic epithelium, a minor subpopulation of cells in thymus that regulate T cell differentiation, rather than by a direct effect on lymphoid cells. This hypothesis is consistent with the low levels of MFO activity induced by TCDD in mouse thymus when this measurement is expressed in terms of whole thymus. Furthermore, as suggested by Nebert et al. (1981), it is likely that the TCDD receptor serves a physiological function and interacts with an endogenous regulatory ligand in the body. Experiments to study further the role of thymic epithelium in TCDD immunotoxicity and to identify the endogenous ligand acting through the TCDD receptor in thymus are currently in progress.
The consequences to the host of haloaromatic hydrocarbon-induced immune suppression remains unclear. Human populations exposed to TCDD or the related PBBs have shown little evidence for a major increase in susceptibility to infections (Pocchiari et al., 1979; Hay, 1982) in spite of evidence of impaired T cell immunity in vitro (Bekesi et al., 1979). However, one should note that patients subjected to total lymphoid irradiation for treatment of lymphornas or arthritis

152 D.A. Clark et al.
develop a profound long-term suppression of both cellular and humoral immunity due to sup- pressor T cells, and yet these patients do not suffer from a high incidence of serious infection (Kotzin et al., 1981). A slight increase in frequency of nonfatal herpes virus infections seems to be the only notable feature in these patients. Our data is in agreement with this observation. TCDD treatment in C57BI/6 mice produced a marginal but detectable increase in susceptibility to lethality following challenge with herpes simplex type II. Imanishi et al. (1980) have reported that oral feeding of mice with the PCB, Kanechlor 500, increased susceptibility to lethal herpes virus infection. We estimate that this effect was achieved with a total dose of approximately 3 × I0 -s M/kg. Taken together with the data shown in Figure 4, the observations of Imanishi et al. suggest that haloaromatic hydrocarbons may produce biologically significant immunosuppression. The observation that ultra-low doses of TCDD can suppress murine CTL responses in mice suggests a need for further study of the effects of low-dose haloaromatic hydrocarbon exposure on the human immune system.
David A. Clark is a Scholar of the Medical Research Council of Canada. This work was supported by the Ontario Ministry of the Environment.
REFERENCES
Bandiera S, Safe S, Okey AB (1982) Binding of polychlorinated biphenyls classified as either phenobarbitone-, 3-methylcholanthrene- or mixed-type inducers to cytosolic Ah receptor. Chern Biol Interactions 39:259.
Bekesi JG, Anderson HA, Roboz JP, Fischbein A, Selikoff IJ, Holland JF (1979) Immunologic dysfunction among PBB-exposed Michigan dairy farmers. Ann New York Acad Sci 320: 717.
Burke MD, Mayer RT (1974) Ethoxyresorufin: direct flourimetric assay of a microsomal o- dealkylation which is preferentially inducible by 3-methylcholanthrene. Drug Metab & Disposi- tion 2:583.
Clark DA, Phillips RA, Miller RG (1977) Characterization of the cells that suppress the cytotoxic activity of T lymphocytes. II. Physical properties, specificity, and developmental kientics of the splenic inhibitory non-T cell population. Cell Irnmunol 34:25.
Clark DA, Gauldie J, Szewczuk MR, Sweeney G (1981) Enhanced suppressor cell activity as a mechanism of immunosuppression by 2,3,7,8-tetrachlorodibenzo-p-dioxin (41275). Proc Soc Exp Biol Med 168:290.
Faith RE, Luster MI, Moore JA (1978) Chemical separation of helper cell function and delayed hypersensitivity responses. Cell Immunol 40:275.
Hay AWM (1982) Exposure to TCDD: The Health Risks. In Chlorinated Dioxins and Related Compounds. Eds., O Hutzinger, RW Frei, E Merian, and F Pocchiari. New York: Pergamon Press, pp. 589-600.
Hutzinger O, Safe S, Zitko V (1971) Polychlorinated biphenyls: synthesis of some individual chlorobiphenyls. Bull Environ Contain Toxicol 6:209.
Imanishi J, Nomura H, Matsurbara M, Masakazu K, Won S J, Mizutani T, Kishida T (1980) Effect of polychlorinated biphenyl on viral infections in mice. Infection and Immunity 29:275.
Jones KG, Swenney GD (1980) Dependence of the porphyrogenic effect of 2,3,7,8- tetrachlorodibenzo-p-dioxin upon inheritance of aryl hydrocarbon hydroxylase responsive- ness. Toxicol Appl Pharm 53:42.
Knutson J, Poland A (1982) Response of routine epidermis to 2,3,7,8-tetrachloro-dibenzo-p- dioxin: interaction of the Ah and hr loci. Cell 30:225.
Kotzin BL, Strober S, Engleman EG, Calin A, Hoppe RT, Kanss GS, Terrell CP, Kaplan HS (1981) Treatment of intractable rheumatoid arthritis with total lymphoid irradiation. New Engl J Med 305:969.

CTL Suppression by Haloaromatic Hydrocarbons 153
Mason ME, Okey AB (1982) Cytosolic and nuclear binding of 2,3,7,8-tetra-chorodibenzo-p- dioxin to the Ah receptor in extra-hepatic tissues of rats and mice. EurJ Biochem 123:209.
Miller RA, Stutman O (1982) Use of positively selected Lyt-2 + mouse splenocytes to examine interleukin-2 secretion in response to alloantigen and to TNP-modified syngenic cells. Cell Immunol 68:114.
Nebert DW, Eisen HJ, Negishi M, Lang MA, Hjelmeland LM (1981) Genetic mechanisms controlling the induction of poly-substrate monooxygenase (P-450) activities. Ann Rev Phar- macol Toxicol 21:431.
Okey AB, Bondy GP, Mason ME, Kahl GF, Eisen HJ, Guenther TM, Nebert DW (1979) Regulatory gene product of the Ah locus. Characterization of the cytsolic inducer-receptor complex and evidence for its nuclear translocation. J Biol Chem 254:11636.
Parkinson A, Robertson L, Safe L, Safe S (1980) Polychlorinated biphenyls as inducers of hepatic microsomal enzymes: structure-activity rules. Chem Biol Interact 30:271.
Pocchiari F, Silvano V, Sampieri A (1979) Human health effects from accidental release of tetrachlorodibenzo-p-dioxin (TCDD) at Seveso, Italy. Ann New York Acad Sci 320:311.
Poland AP, Glover E, Robinson JR, Nebert DW (1974) Genetic expression of aryl hydrocarbon hydroxylase activity. Induction of mono oxygenase activities and cytochrome P-450 formation by 2,3, 7,8-tetrachlorodibenzo-p-dioxin in mice genetically "nonresponsive" to other aromatic hydrocarbons. J Biol Chem 249:5599.
Poland A, Glover E (1980) 2,3,7,8-tetrachlorodibenzo-p-dioxin: segregation of toxicity with the Ah locus. Mol Pharm 17:86.
Poland A, Knutson JC (1982) 2,3,7,8-tetrachlorodibenzo-p-dioxin and related halogenated aromatic hydrocarbons. Examination of the mechanism of toxicity. Ann Rev Pharmacol Toxicol 22:517.
Porter EH, Berry RJ (1964) The efficient design of transplantable tumor assays. Brit J Cancer 17:583
Quinnan G, Santos GW, Saral R, Burns WH (1982) Cytotoxic T cells in cytomegalovirus infection: HLA-restricted T-lymphocyte and Non-T-lymphocyte mediated cytotoxic responses correlate with recovery from cytomegalovirus infection in bone-marrow-transplant recipients. New Engl J Med 307: 7.
Rawls WE (1979) Herpes simplex virus types 1 and 2 and herpes virus simiae. In Diagnostic Procedures for Viral, Rickettsial and Chlamydial lnfections. 5th Edition. Eds., EH Lennette and NJ Schmidt. Washington: American Public Health Association, pp. 309-373.
Teh HS, Harley E, Phillips RA, Miller RG (1977) Quantitative studies of the precursors of cytotoxic lymphocytes. I. Characterization of a clonal assay and determination of the size of clones derived from a single precursor. J Immunol 118:1049.
Vos JG, Faith RE, Luster MI (1980) Immune alterations. In Halogenated biphenyls, terphenyls, naphthalenes, dibenzodioxins and related products. Ed., R Kimbrough. Amsterdam: Elseviel'/North-Holland Biomedical Press, pp. 241-266.