ceresini florianopolis iiwbw_2016_recent_a
TRANSCRIPT

1
The recent emergence and evolution of the wheat blast species complex in Brazil!
!!
Paulo C. Ceresini and collaborators !

Prof. Paulo C. Ceresini UNESP ! ! !Dr. João L. N. Maciel Embrapa Wheat !!Dr. Vanina L. Castroagudín UNESP ! !Prof. Eduardo Alves UFLA Lavras!
! !!Dr. Silvino I. Moreira UFLA Lavras ! ! !Prof. Pedro Crous CBS The Netherlands !
!Dr. Anderson L. D. Danelli UPF Passo Fundo !Dr. Patrick Brunner ETH Zurich!
!Dr. Giselle Carvalho UNESP ! ! !Prof. Bruce A. McDonald ETH Zurich!
! ! ! ! ! ! ! ! ! ! !!! ! ! ! ! ! ! ! !Dr. Daniel Croll ETH Zurich!
!! ! ! ! ! ! ! ! !Dr. Ana Lídia Bonato Embrapa Wheat !
!! ! ! ! ! ! ! ! !Juliana T. A. Reges UNESP ! !!
Danilo A. S. Pereira ETH Zurich ! ! !Samanta C. Oliveira UNESP !! ! ! ! ! ! ! ! !Nadia Poloni UNESP !! ! ! ! ! ! ! ! !Adriano F. DoriganUNESP!
! ! ! ! ! ! ! ! !Matheus M. Dorigan UNESP!
! ! ! ! ! ! ! ! !Suellen SouzaUNESP!
Our Research Team

3
Wheat blast is caused by Pyricularia species closely related to the rice blast pathogen
Whe
at b
last
Ric
e bl
ast
Source: Danelli & Maciel
Source: Castroagudin, Moreira & Crous Source: Castroagudin, Moreira & Crous

A species with several names along its history
Magnaporthe oryzae: 2002 For both rice and wheat pathogens
Pyricularia grisea: 2007
Some resilience in accepting Magnaporthe oryzae
2014: Guess who’s back? Magnaporthe oryzae as the wheat blast pathogen

Adopting the new family name Pyriculariacea for the rice/wheat blast pathogen
“Species of Pyricularia s. str. belong to a monophyletic clade that includes all P. oryzae/P. grisea isolates tested, defining the Pyriculariaceae, which is sister to the Ophioceraceae, representing two novel families. These clades are clearly distinct from species belonging to the Gaeumannomyces pro parte / Magnaporthiopsis / Nakataea (syn. Magnaporthe) generic complex that are monophyletic and define the Magnaporthaceae.”

Pyricularia: From now on! Magnaporthe: No more.
“Magnaporthe oryzae (=M. salvinii), the type of Magnaporthe, forms a Nakataea asexual morph, and hence Luo & Zhang (2013) introduced the combination Nakataea oryzae for this fungus, as the name Nakataea (1939) is older than Magnaporthe (1972). This decision effectively reduced Magnaporthe to synonymy under Nakataea.“
“The majority of species formerly treated as Magnaporthe, fall in the Pyricularia complex (Murata et al. 2014).”

The recent origin and spread of wheat blast in South America
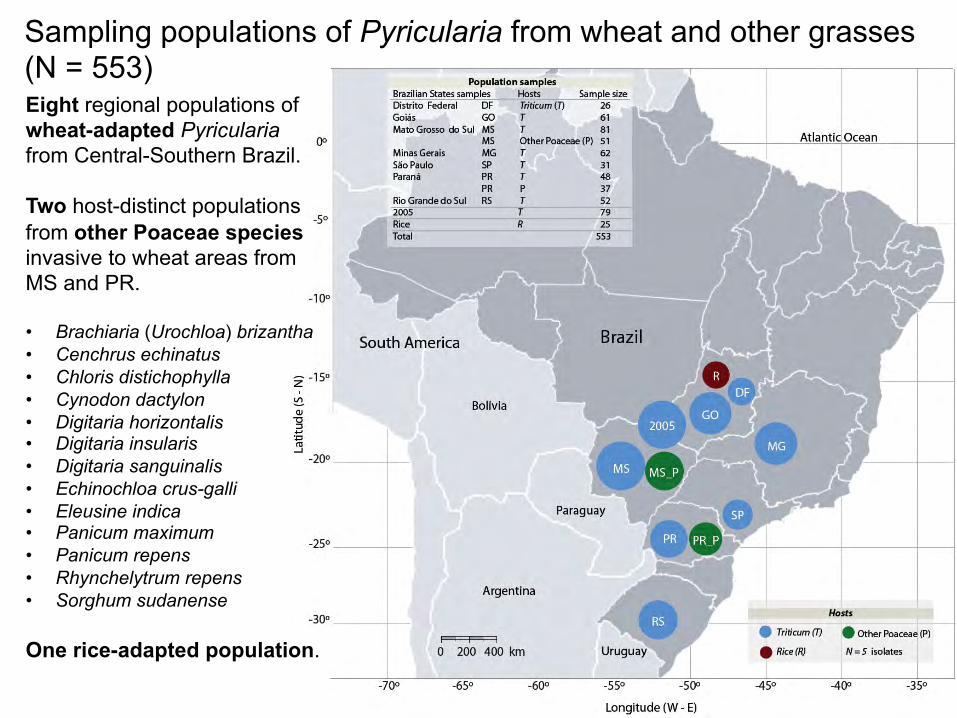
Sampling populations of Pyricularia from wheat and other grasses (N = 553) Eight regional populations of wheat-adapted Pyricularia from Central-Southern Brazil. Two host-distinct populations from other Poaceae species invasive to wheat areas from MS and PR. • Brachiaria (Urochloa) brizantha • Cenchrus echinatus • Chloris distichophylla • Cynodon dactylon • Digitaria horizontalis • Digitaria insularis • Digitaria sanguinalis • Echinochloa crus-galli • Eleusine indica • Panicum maximum • Panicum repens • Rhynchelytrum repens • Sorghum sudanense One rice-adapted population.

Characterization of Pyricularia from wheat and other grasses
Genotypes 11 SSRs
DNA sequences Genomic data 10 housekeeping genes ~434’000SNPs
cytB and cyp51 (mtDNA) from 17 entire genomes AvrCO39 .
PCR assays
AvrCO39 AvrPITA
Mating types
Phenotypes Pathogenicity to barley, oat, rice, signalgrass and wheat Virulence on leaves (7 wheat cultivars + barley + rice)
Virulence on ears (7 wheat cultivars + barley) Sensitivity to QoI (strobilurins) and triazole fungicides

Sympatric samples: Wheat-derived isolates 76 Other Poaceae 26 Rice 23 Pyricularia grisea 1 P. pennisetigena 1
3301 bp sequenced 552 polymorphic sites 30 fixed mutations among groups
Genes ACT (actin) BAC6 βT-1 (beta-tubulin) CAL (calmodulin) CH7-BAC7 CH7-BAC9 CHS1 (chitin synthase 1) EF-1α (translation elongation factor 1-alpha) MPG1 (hydrophobin) NUT1 (nitrogen regulatory protein 1)
Comparisons of 10 genes in 125 Pyricularia strains show divergence between strains infecting
wheat, other Poaceae, or rice

% divergence (only fixed mutations within a particular group) Pgt vs. PoT = 0.50% Pgt vs. PoO = 0.53% PoT vs. PoO = 0.16% Isolates causing wheat blast could be separated into two distinct phylogenetic clusters: Cluster I = Pgt and II = PoT, with a high posterior probability (P= 0.99). Cluster II is further subdivided in two clades (1 and 2), distinct from the rice blast pathogen PoO.
0.0060
12�1�127
364
12�1�186
706
12�0�535i
12�1�236
12�1�217
12�1�183
12�1�146
12�0�543i
12�1�035
12�1�078
12�1�037
12�1�015
10659
12�1�315
284
12�1�058
12�0�366
12�1�169
12�1�048i
12�1�010i
421
12�1�213
8847
12�1�234
12�1�311
12�1�050i
678
12�0�346
12�0�347
12�1�116
12�1�001
12�1�191
8844
12�1�149
12�1�132
12�1�032i
12�1�139
8762
12�1�109
12�1�207
641
12�1�002
323
658
12�0�625i
12�1�193
8772
12�0�613i
12�1�204
12�1�153
12�0�555i
12�1�225
12�1�19412�1�197
12�1�002i
8763
12�1�089
10877
12�1�021i
10783
12�1�087
12�1�180
12�1�179
12�0�655i
12�1�1�8
12�0�051i
704
97
12�0�194
12�1�174
12�0�321
12�1�075
12�1�085
12�1�009
12�1�24112�1�135
12�1�097
12�0�368
12�1�291
12�1�051i
12�1�187
12�1�005i
12�0�326
12�1�053i
12�1�205
12�0�038i
12�1�019i
10879
12�1�288
10880
12�1�147
12�1�107
695
12�0�607i
12�1�148
674
12�1�10012�0�009i
12�1�034i
P.grisea Brazil
12�1�052i
12�0�578i
12�0�073
12�1�007
12�1�209
12�0�534i
12�1�045i
12�1�119
12�1�049i
12�1�014i
P. pennisetigena Brazil
12�1�181
12�1�21912�1�228
12�1�112
12�1�061
12�1�020i
12�0�345
12�1�014
611
12�1�117
12�1�243
12�1�182
1.00
1.00
1.00
1.00
0.99
0.75
0.99
0.99
0.98
0.64
0.85
1.00
3\ULF
XODULD�JUDPLQLV�WULWLFL�VS��QRY��
)LJ����3K\ORJHQHWLF�DQDO\VLV�
&ODGH���
&ODGH���
3\ULF
XODULD�RU\]DH�
SDWKRW\SH�2U\]D�
3\ULF
XODULD�RU\]DH�SDWKRW\SH�7ULWLFX
P�
I
II
I
&ODGH���
Pgt
PoT
PoO
Three independent emergences for the wheat blast in the Brazilian agroecosystem.
0.98
1.00
1.00
1.00
1.00
0.99
0.99
0.99
BPP
BPP = Bayesian Posterior Probability
Castroagudin et al. (2016)
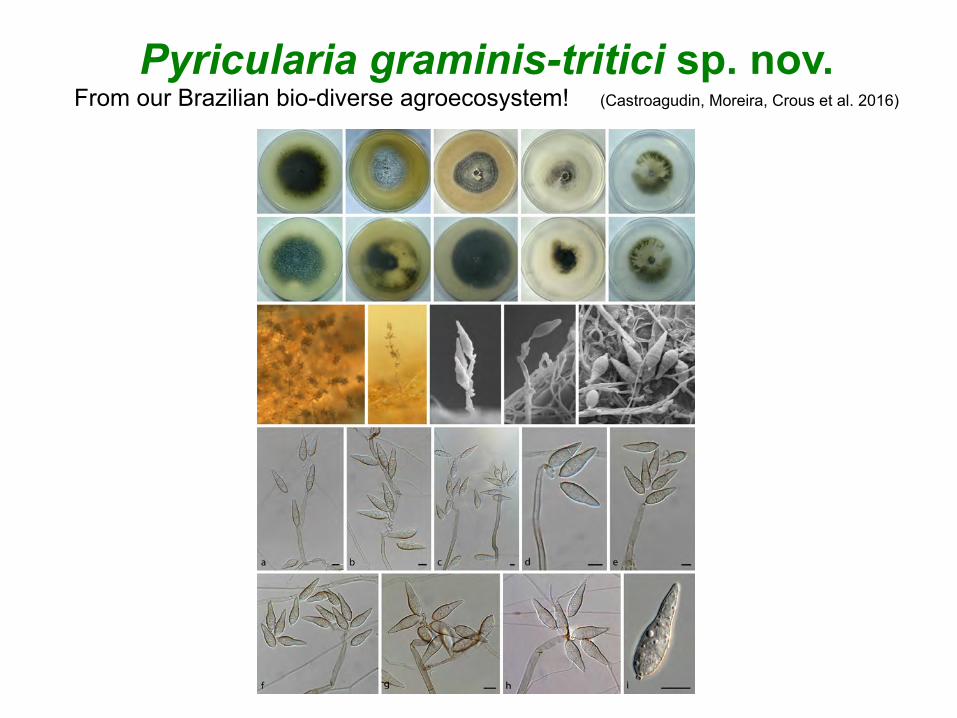
Pyricularia graminis-tritici sp. nov. From our Brazilian bio-diverse agroecosystem! (Castroagudin, Moreira, Crous et al. 2016)

Pyricularia graminis-tritici sp. nov. From our Brazilian bio-diverse agroecosystem! (Castroagudin, Moreira, Crous et al. 2016)
To our knowledge, P. graminis-tritici sp. nov. is still restricted to Brazil, but obviously represents a serious threat for the cultivation of wheat globally.

Pathogenicity spectra of Pyricularia graminis-tritici (Pgt) and P. oryzae pathotypes Triticum (PoT) and Oryza (PoO)
Castroagudin et al. (2016)

Pathogenicity spectra of Pyricularia graminis-tritici (Pgt) and P. oryzae pathotypes Triticum (PoT) and Oryza (PoO)
Castroagudin et al. (2016)
Barley Oat Signal grass Wheat Rice
Pgt + + + + - PoT + + + (1)/ -(2) + - (clades 1 and 2)
PoO + + - + + Wheat blast isolates from Pgt or PoT do not infect rice,
but they all infect barley, oat and signal grass (except PoT clade 2). Rice blast isolates (PoO) infect barley, oat, and wheat, but not signal grass.
The hypothesis of ”host-specific forms” does not hold for the wheat blast pathogens.
The distribution of the wheat blast pathogens varies according to a temperature cline
Prevalent in warmer Central Brazil.
RS SP MS PR Rice
Mato Grosso do Sul
MS
Minas GeraisMG
GoiásGO
São PauloSP
ParanáPR
Rio Grande do Sul
RS
Distrito Federal
DF
BRAZIL
Weeds - derived
0.8
0.6
0.4
0.2
Prop
ortio
n
Pyricularia graminis-tritici sp. nov.
N = 146on wheat
Central Brazil
DF + GO
MG
MS
SP
PRRS
Distribution towards
A
B
t test p value = 0.0061
SouthernBrazil
43%

The distribution of the wheat blast pathogens varies according to a temperature cline
Predominant in Southern Brazil.
0.8
0.6
0.4
0.2
Pyricularia oryzae pathotype
Triticum
Central Brazil
SouthernBrazil
Distribution towards
DF + GO
MG
MS
SP
PRRS
A
B
t test p value = 0.006
Mato Grosso do Sul
MSMinas GeraisMG
GoiásGO
São PauloSP
ParanáPR
Rio Grande do Sul
RS
Distrito Federal
DF
BRAZIL
57%Pr
opor
tion
N = 193 on wheat

Major Interpretation 1
Blast on wheat and other invasive Poaceae hosts from the Brazilian agroecosystem
is a complex disease caused mainly by two Pyricularia species:
Pyricularia graminis-tritici sp. nov. (Pgt)
Pyricularia oryzae pathotype Triticum (PoT).
…with broader host range within the Poaceae
but with ecotype differences.

Two other Pyricularia species cause blast on wheat, oat and Urochloa
Pyricularia species associated with blast on several poaceous hostsinvasive of wheat fields
Pyricularia graminis-tritici
Pyricularia oryzae
pathotypeTriticum
Other Pyricularia
species
N = 76
*Identified phylogenetically according to Klaubauf et al. 2015, Studies in Mycology.
N = 38
N = 13
N = 25
Pyricularia zingibericola *
Pyricularia spp.
Pyricularia pennisetigena *
Pyricularia grisea
Cenchrus echinatus
Urochloa spp.
Panicum maximum
Urochloa spp.
Panicum maximum
Chloris distichophyllaAvena sativa
Urochloa spp.Panicum maximum
Digitaria sanguinalis
2014

Pp = Pyricularia pennisetigena Pz = Pyricularia zingibericola
Two other Pyricularia species cause blast on wheat, oat and Urochloa
SNA. res.
(2014)Studiesinmycology,79,85-120.

Pg = Pyricularia grisea Pgt = Pyricularia graminis-tritici Pp = Pyricularia pennisetigena Pz = Pyricularia zingibericola
Pyricularia species
Diseased leaf area (%)
Two other Pyricularia species caused blast on wheat, oat and Urochloa
N=3639

Sympatric samples from Brazil Wheat-derived isolates 08 Other Poaceae 09
Phylogeny reconstruction • Using RAxML
• ~434,000 SNPs from the entire genome • The dataset was analyzed using alignments against
the 70-15 reference genomes (Bowtie2 aligner) • Used only SNPs genotyped among at least
90% of all isolates (i.e., very closely related isolates may be slightly more differentiated
due to SNPs only callable within a subgroup)
Compilation of available Pyricularia genomic data
• Darren Soares / Exeter • NCBI draft assemblies (several) • Chiapello et al. (GBE 2015) / Montpellier
Genomic data comparisons of SNPs in 17 Pyricularia strains confirms the divergence between strains infecting
wheat, other Poaceae, and rice

0.2
Mo401.4
K96-07
PY35.3
Mo4603.4
MoK84-01
BM1-24
K93-16K96-11
PY6025
PY86.1
12.0.009i
P131
BR32
12.1.205
MoCQ11
Y34
K88-24
12.0.368
K91-13
Mo2303.1
12.1.053i
TH12-rn
MoJS25
12.1.127
Mo1801.4
MoKJ201_3
12.0.012i
PY5035
PH14-rn
MoHB12
PY5002
363
Mo1106.2
TH16
12.0.345i
12.1.169
HN19311
PY0925
Mo1836.3
4091 -5 -8
GY11
PY36.1
MoZJ15
MoGX01
12.0.534i12.0.642i
IT10
K98-02
PY5003
Mo4403.2
FJ81278
12.1.117
US71
BR29
12.0.326
MoAH06
MoSC05
PY6017
CA205
K88-07K91-30
12.0.555i
12.1.032i
FR13
B157MG01
PR72
98 -06
GOV41
PY6045
PY5033
70 -15
PY5010
PY6047PY25.1
12.0.007i
12.1.204
K98-10
Mo903.4
MoGD22
CD156
K91-10
MoHN06
MoJL10
MoNX37
12.1.037
Unknown host (Brazil)
Oryza sativa
Setaria italica
Digitaria sanguinalis
Triticum aestivum
Triticum aestivum
Triticum aestivum
Cenchrus echinatus
Eleusine indica
Triticum aestivum
Triticum aestivumTriticum aestivumTriticum aestivumTriticum aestivum
Urochloa ssp.Urochloa ssp.Urochloa ssp.
Digitaria sanguinalis
Triticum aestivum
Urochloa ssp.Echinochloa crusgalliAvena sativa
Triticum aestivumTriticum aestivumTriticum aestivum
Eleusine indica
Weeping lovegrass isolateCross between K76-79 (host: weeping lovegrass)and WGG-FA40 (host: finger millet)(Valent et al. 1986)
Chiapello et al. (GBE 2015)Ceres in i / McDonald / Croll
NCBI (various)Soares / Exeter
Data source
Triticum aestivumTriticum aestivumTriticum aestivumTriticum aestivum
Pgt
Pgt
PoT
PoT
PoT
Pgt
PoO
Pyricularia grisea ( Brazil)
Whole-genome phylogenetic tree built using RAxML with ~434,000SNPs. *The effect of genotyping rates per SNP was tested and found not significantly influencing the branching of the tree.
Wheat-infecting (and other poaceous hosts-) strains
Rice-infecting strains
PoO
The hypothesis of ”host-specific forms” does not hold for the wheat blast species complex.

Major Interpretation 2
Wheat blast and related strains (Pgt and PoT) found on different hosts showed overall much more polymorphism than the rice-infecting strains (PoO). No strict host specialization among the different clusters. Wheat blast might have originated from multiple sources and/or maintains significant genetic variability.

Evidence supporting the scenario of one disease, two major species: Pgt and PoT
RST values
0.24 0.24
RST values
0
0.25
0.50
1.00
0.75
completly
N = 208 MLMS genotypes
PoT0.843** 0.846**
Pgt PoT PoO
RST values in bold colored p values at _ = 0.01 (**)
PoO0.057**
Pgt -
-
-
rectangle
Current subdivision between Pgt and PoT
Subdivision among the two blast pathogens and PoO

High historical asymmetrical migration rate from Pgt to PoT
A coalescent-based genealogical analyses of population size and migration rates (Beerli and Felsenstein, 2001, Proc. Natl. Acad. Sci. 98:4563-4568) using SSR markers.
MIGRATION RATE AND POPULATION SIZE ESTIMATION
Migrate-n version 3.6.11 [June-18-15] Compiled for a PARALLEL COMPUTER ARCHITECTURE One master and 66 compute nodes are available.
Markov chain settings: Long chainNumber of chains 1
Recorded steps [a] 8334Increment (record every x step [b] 500Number of concurrent chains (replicates) [c] 6Visited (sampled) parameter values [a*b*c] 25002000Number of discard trees per chain (burn-in) 500Forcing percentage of new genealogies 0.10
Multiple Markov chains:Static heating scheme
5 chains with temperatures100000.00 10000.00 1000.00 100.00 1.00
Swapping interval is 10.0
20.0
40.0
60.0
80.0
100.0
120.0
Pgt
PoT PoO
0.0
20.0
40.0
60.0
80.0
100.
0
120.
0
0.0
20.0
40.0
60.0
80.0
100.0
120.0
Number of migrants exchanged/generation��e2Nm)
Historical migration estimates using the coalescent modelEvidence that the PoT population from wheat may be derived from an original Pgt population.

No differentiation between populations of Pgt from wheat and other poaceous hosts in Brazil
RST values
0.24 0.24
RST values
0
0.25
0.50
1.00
0.75
completly
N = 79 MLMS genotypes
Pgt
RST value is non-significant ( p = 0.32)
0.004Pgt (wheat) NS
(other poaceous hosts)
Populations from the two host groups had the same genetic structure.
Other poaceous hosts for Pgt: Avena sativa, Cenchrus echinatus, Cynodon spp.,
Digitaria sanguinalis, Elionurus candidus, Echinochloa crusgalli, Eleusine indica, Rhynchelytrum repens, and Urochloa spp.

Major Interpretations 3 • Pgt populations from wheat and other poaceous hosts had
the same genetic structure. • Among these grass species Urochloa is a widely grown
pasture grass occupying more than 90 million ha in Brazil, we propose that Urochloa provides a major source of wheat blast inoculum.
…and may be the preferred host for pathogen recombination.
Urochloa brizantha cv. Piatã

A mixed reproductive system Some clonality but gametic equilibrium in most of the locus pairs
Ratio MAT1-1 : MAT1-2
Pgt PoT
QoI-R cytB H1 15 : 1 18 : 1QoI-S cytB H9 1 : 1 1 : 5
Selection for QoI resistance increased the frequency of MAT1-1 / QoI-R strains
Despite the high selection against MAT1-2 QoI-S on wheat, the pathogens have potential for sexual reproduction on non-sprayed grasses.
Skewed mating types ratios

Phalaris canariensis (bird seed)
Oryza sativa cv. Primavera
Oryza sativa cv. Relâmpago
Oryza sativa cv. Yin Lu(Red rice)
Urochloa Mulato hybrid(signal grass)
Hordeum vulgare cv. BR Elis
(barley)Hordeum vulgare
cv. MN73Secale cereale
cv. BR1 (rye)
Avena strigosa cv. Embrapa 29 Garoa
(black oat)
Setaria italica(foxtail millet)
Triticum aestivumBRS 264 (wheat)
Triticum aestivumMGS Brilhante
XTriticum secalecv. IAC Canindé
(triticale)
Ascocarps abundantly
formed in senescent
tissues from wheat and other poaceous hosts inoculated with
both fungal mating types.
Source: S. I. Moreira
Co-inoculation of MAT1-1 x MAT1-2 lineages Pyricularia graminis-tritici

Major Interpretations 4
The genetic structure consistent with a mixed reproductive system includes regular cycles of recombination followed by clonal expansion. The fungal biology observations reinforces the population genetics evidence in favor of a mixed reproductive system.
Rapid local adaptation in populations of the wheat blast pathogens They have become resistant to QoI fungicides in ~10 years
Haplotype % isolates Host
H1 (QoI-R) 71% (Pgt and PoT) Wheat / other Poaceae
H2-H4 (QoI-R) 2% (Pgt and Pg) Other Poaceae
H5-H8 (QoI-R) 7% (Pgt , Pp, Pz, and Pg) Other Poaceae H9 (QoI-S) 20% (Pgt and PoT) Wheat / Other Poaceae
H7H6
H5H1
H3
H9H8
H4
H2
Other Poaceae
Rice-S
Triticum and other Poaceae
2005
DF
GO
MG
MSPR
RS
SP
Other Poaceae
PRMG
PRMS
MS
2005
DF MG
MS PR
Oth
er P
oace
ae
MS
PR
QoI Resistant
QoI Sensitive
cyt B haplotypes
PR
MSMSPR
Haplotypic variation at the citB gene
Castroagudin et al. 2015. Phytopathology 105:284-294
Sampling year % resistance isolates 2005 ~ 36 % 2012 ~ 80%
Resistance to azoxystrobin (100 ppm)

Progressive local adaptation in the wheat blast pathogens They have become quantitatively resistant
to azole fungicides in ~25 years
Freq
uenc
y
n= 44 MS
n= 26 DF/GO
n= 19 RS
n= 27
n= 28
n= 36
MG
SP
PR
EC 50 (ppm) to tebuconazole
0.6 0.8 1.0 1.2 1.4 0.6 0.8 1.0 1.2 1.4
20
15
10
5
0
15
10
5
0
10
8
6
4
2
0
1086420
1086420
15
10
5
0
H1PR, MS, RS,
MG, SP, DF_GO
165 95.9
H2 MG 1 0.6H3 PR,SP 1 0.6H4 SP, DF_GO 1 0.6H5 DF_GO 1 0.6H6 DF_GO 1 0.6H7 RS 1 0.6H8 RS 1 0.6
Total 6 172 100
LocationHaplotype N %N % N %
64 35.6 101 56.1
1 0.61 0.6
1 0.61 0.61 0.61 0.6
1 0.669 38.3 103 57.2
Pgt PoT
doses tebuconazol (ppm)
tebuconazole (ppm )
1210
48i
1211
30
0
20
40
60
80
100
Rela
tive
grow
th (
%)
doses of tebuconazole (pp
MO121048i
MO121130
Populations
cyp51 A
Poloni et al. 2016
Level of tebuconazole tolerance = 35 – 70 X higher than the labeled field dose Only few PoO isolates from rice were sensitive!
Several non-synonymous mutation detected in the cyp51A gene.

Emergence of 10 - 26 pathotypes that show cultivar specialization and host tissue specialization
Source: Cristina Boaretto, Anderson Danelli, João L. Nunes Maciel
Maciel et al. 2014. Phytopathology 104:95-107
Rapid local adaptation in populations of the wheat blast pathogens They have become host specialized in ~25 Years

Rapid local adaptation in populations of the wheat blast pathogens:
Latit
ude
(S -
N)
Longitude (W - S)
0 200 400 kmUruguay
Argentina
Paraguay
Bolivia
South America
Atlantic Ocean
Brazil
-3
0°
-2
5°
-20°
-1
5°
-
10°
-5
°
0°
-70° -65° -60° -55° -50° -45° -40° -35°
Pyricularia graminis-tritici (Pgt) 90 Pyricularia oryzae pathotype Triticum (PoT) 55
Total 145
MS = 38
RS = 15
MG = 22
SP = 5
DF_GO = 36
PR = 29 N
Sampling strategy for virulence grouping

High “race” diversity based on seedling reaction. Race A is virulent on all wheat + oat cultivars.
Complexity of virulence groups varied.
Source: Cristina Boaretto, Anderson Danelli, João L. Nunes Maciel / Maciel et al. 2014. Phytopathology 104:95-107
Rapid local adaptation in populations of the wheat blast pathogens
ABCDEFGHIJKLMNOPQRSTUVWXYZ
Virulence
group
Seedling reaction
Wheat Oat Isolates
S S S S S S S S S S S 32 16
S S S S S S S S R S S 4 2
S S S S S S S S S S R 2 1
S S S S S S S S R S R 1 0.5
S S S S S S S R S S S 6 3
S S S S S S S S R R S 5 2.5
S S S S S S S R R R S 15 7.5
S S S S S S R R R R R 9 4.5
S S S S S S S S S R S 2 1
S S S S S S S R S R S 3 1.5
S S S S S S S R R S S 2 1
S S S S S S R R R R S 87 43.5
S S S S S S R R R S S 3 1.5
S S S S S S R R S R S 2 1
S S S S S S R R S S S 1 0.5
S S S S S S S S S R R 1 0.5
S S S S S S S R R R R 2 1
S S S R S R R R R R S 1 0.5
S S S R S S R R R R S 1 0.5
S R R R R R R R R R S 6 3
S S S S S S R S S S S 1 0.5
S R R R S R R R R R S 6 3
S R R R S S R R R R S 4 2
S S R S S S R R R R S 1 0.5
S R R S S S R R R R S 1 0.5
R R R R R R R R R R S 2 1
Total 200 100
BRS BuritiBRS 229 Renan BRS 234PFC
2010123
Anahuac
75BR 24
MGS 3
Brilhante
BR 18
TerenaBRS 220 CNT 8 Number Freq. (%)
Emergence of 26 pathotypes that show cultivar specialization and host tissue specialization.
Rapid local adaptation in populations of the wheat blast pathogens
Pyricularia graminis-tritici (Pgt) 90 Pyricularia oryzae pathotype Triticum (PoT) 55
Total 145
N
Frequency distribution of virulence groups (races) in Brazil
Seedling reaction La
titu
de
(S
- N
)
Longitude (W - S)
0 200 400 kmUruguay
Argentina
Paraguay
Bolivia
South America
Atlantic Ocean
Brazil
-
30° -
25° -2
0° -
15
° -1
0° -5
°
0°
-70° -65° -60° -55° -50° -45° -40°
-35°
Pyricularia graminis-tritici (Pgt) 90 Pyricularia oryzae pathotype Triticum (PoT) 55
Total 145
MS = 38
RS = 15
MG = 22
SP = 5
DF_GO = 36
!"
Pyricularia pathotype Triticum (P-T)Pyricularia graminis-tritic(Pgt)
PR = 29 N
Sampling strategy for virulence grouping

Lower “race” diversity based on ear reaction.
Race A and C are virulent on all wheat + oat cultvars.
Rapid local adaptation in populations of the wheat blast pathogens
ABCD
AABBCCDDEET
Virulence group
Ear reactionWheat Oat Isolates
S S S S S S S S S S S 162 80.6S S S S S S S S R S S 27 13.4S S S S S S S S S S R 2 1.0S S S S S S S S R S R 3 1.5S R S S S S S S S S S 2 1.0S R S S S S S R R S S 1 0.5S S S S S S S S R R R 1 0.5S S R S S R S S R R R 1 0.5S S R R R S R R R R S 1 0.5S R R R R R R R R R S 1 0.5
Total 201 100.0
Anahuac 75
BR 24 MGS 3 Brilhante
BR 18 Terena
BRS 220 CNT 8 BRS 229 BRS Buriti Renan BRS 234 PFC 2010123
Number Freq. (%)
Source: Cristina Boaretto, Anderson Danelli, João L. Nunes Maciel / Maciel et al. 2014. Phytopathology 104:95-107

Rapid local adaptation in populations of the wheat blast pathogens
La
titu
de
(S
- N
)
Longitude (W - S)
0 200 400 kmUruguay
Argentina
Paraguay
Bolivia
South America
Atlantic Ocean
Brazil
-
30° -
25° -2
0° -
15
° -1
0° -5
°
0°
-70° -65° -60° -55° -50° -45° -40°
-35°
Pyricularia graminis-tritici (Pgt) 90 Pyricularia oryzae pathotype Triticum (PoT) 55
Total 145
MS = 38
RS = 15
MG = 22
SP = 5
DF_GO = 36
PyrpatTriPyrgra(Pg
!"
Pyricularia oryzae pathotype Triticum (Po-T)Pyricularia graminis-tritici (Pgt)
PR = 29 N
Sampling strategy for virulence grouping
Ear reaction
Pyricularia graminis-tritici (Pgt) 90 Pyricularia oryzae pathotype Triticum (PoT) 55
Total 145
N Frequency distribution of virulence groups (races) in Brazil
Emergence of 10 pathotypes that show cultivar specialization and host tissue specialization

Can we correlate diversity of “avirulence” genes with phenotypic diversity for virulence?
MS PR RS SP MS PR Rice
Mato Grosso do Sul
MS
Minas GeraisMG
GoiásGO
São PauloSP
ParanáPR
Rio Grande do Sul
RS
Distrito Federal
DF
BRASIL
Haplotype network and distribution of AVR1-CO39*
126 isolates
H1Other poaceous hosts (PR and MS)
Triticum (DF)GO
MSMGSPPR
RS
2005N
105
11
1
8
H2 Other poaceous hosts (PR and MS)Triticum (MG)
MSRS
2005
H3 Triticum (PR)
H5Triticum (2005)
Other poaceous hosts (MS)Triticum (GO)
MS
MGSP
RS
H4
* Haplotypes network were constructed by statistical parsimony with the program TCS 1.21 (Clement et 2000. Mol. Ecol. 6:1657-1659).
Haplotypes are connected by line segments representing a single mutational step each.
The unshaded squares represent unsampled haplotypes.
Mutations (sites) 59122222223 78902336692 463366791HaplotypesH1 CTAGAGGATACH2 .....T.....H3 ....GT...G.H4 GCTA..AGA..H5 ..........T
Total: 330 bp
Pgt 76.9 %PoT 54.7 %
Pgt -PoT 25.0 %
Pgt -PoT 1,6 %
Pgt 7.7 %PoT 4.7%
Pgt - %PoT 1.6 %

Major Interpretations 5
Since its emergence the wheat blast pathogens have evolved rapidly to become resistant to QoI and azole fungicides and specialized to include at least 10 pathotypes (26 based on seedling reaction!)

Summary (if there is time left!) Blast disease on wheat and other invasive Poaceae hosts from the Brazilian agroecosystem is caused by multiple Pyricularia species. Pyricularia graminis-tritici and P. oryzae pathotype Triticum are wheat pathogens that likely emerged de novo in Brazil ~30 years ago Pyricularia graminis-tritici did not emerge through adaptive evolution from the rice blast pathogen. Pyricularia oryzae pathotype Triticum may have emerged from an ancestor Pyricularia graminis-tritici population. Several grasses, but mainly Urochloa, may be the preferred host of Pyricularia graminis-tritici, a place for sexual recombination and a reservoir of sexual and asexual inoculum (new focus for control efforts?) Pyricularia graminis-tritici and P. oryzae pathotype Triticum appear to have a high evolutionary potential and poses a risk of becoming an invasive pathogen on wheat crops on other continents

43!!
Funding institutions and grants in Brazil CNPq - Brazilian National Research Council, PPq grant 307361/2012-8 EMBRAPA / Monsanto Research Macro-program FAPESP - São Paulo State Foundation for Research Advancement, grants 2013/10655-4 and 2015/10453-8 Research scientists sponsored in Brazil: Dr. João Maciel (PI/Embrapa Monsanto Research Macroprogram’s grant) Dr. Paulo Ceresini (PI/CNPq and FAPESP’s grants) Dr. Ana Lídia Bonato (EMBRAPA Wheat) Post-Docs: Dr. Vanina Castroagudin (UNESP), CNPq/PDJ grant 150490/2013-5 and FAPESP/PD grant 2014/25904-2 Dr. Giselle Carvalho (UNESP), CAPES/PNPD grant Students: Juliana Reges (PhD/UNESP), Samanta de Oliveira, Nadia Poloni and Danilo Pereira (MSc/UNESP), Adriano Dorigan (undergrad/UNESP): scholarships from CAPES, CNPq or FAPESP.
Acknowledgements