chapter 1 introduction - inflibnetshodhganga.inflibnet.ac.in/bitstream/10603/4972/6/06_chapter...
TRANSCRIPT

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
xiv
Chapter 1
Introduction
Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
1
1.1. Introduction:
Radiation is a ubiquitous energy. Radioactive materials and radiation have always
existed as a part of our environment. The increased use of radioactive materials,
which is a direct outgrowth of the current military and energy policies of the
developed world, poses a need for studying the health effects of radiation (Figure 1.1).
As of 2004, nuclear power provided 6.5% of the world's energy and 15.7% of the
world's electricity (1). As of 2007, the IAEA reported there are 439 nuclear power
reactors in operation in the world, operating in 31 countries (2).
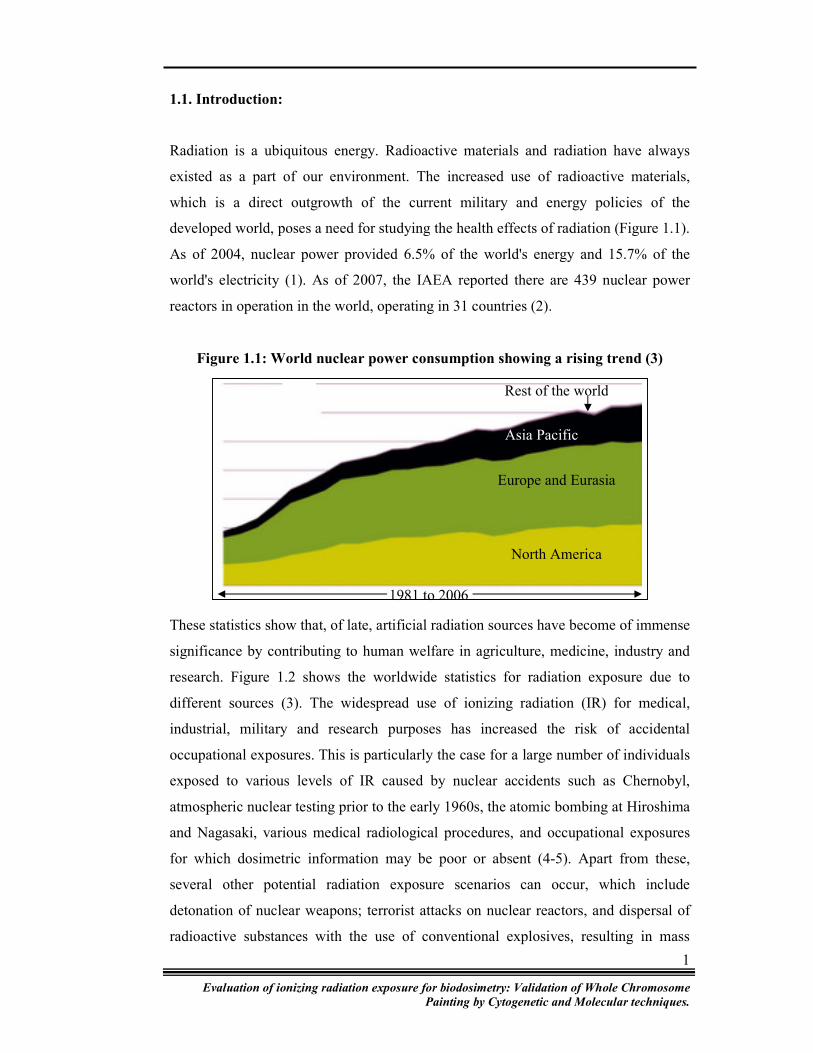
Figure 1.1: World nuclear power consumption showing a rising trend (3)
These statistics show that, of late, artificial radiation sources have become of immense
significance by contributing to human welfare in agriculture, medicine, industry and
research. Figure 1.2 shows the worldwide statistics for radiation exposure due to
different sources (3). The widespread use of ionizing radiation (IR) for medical,
industrial, military and research purposes has increased the risk of accidental
occupational exposures. This is particularly the case for a large number of individuals
exposed to various levels of IR caused by nuclear accidents such as Chernobyl,
atmospheric nuclear testing prior to the early 1960s, the atomic bombing at Hiroshima
and Nagasaki, various medical radiological procedures, and occupational exposures
for which dosimetric information may be poor or absent (4-5). Apart from these,
several other potential radiation exposure scenarios can occur, which include
detonation of nuclear weapons; terrorist attacks on nuclear reactors, and dispersal of
radioactive substances with the use of conventional explosives, resulting in mass
Asia Pacific
Europe and Eurasia
North America
Rest of the world
1981 to 2006

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
2
casualties. These disasters can result in different radiation exposure types: whole
body, localized or partial body, internal contamination, external contamination, and
contaminated burns and wounds (6).
Figure 1.2: World radiation exposure statistics due to various sources
There is a need for reliable methods to assess past exposure to clastogens and the
related risk. A systematic investigation of radiation-induced damage is of great
importance for the population of radiation workers at risk in the nuclear industry and
medical sector. Radiation accidents in foreign countries have pointed to the need for a
reliable methodology, allowing quick and accurate measurement of the radiation
burden received, not only for a restricted number of individuals but also for a sizeable
fraction of the population. At lower doses, exposure assessment is useful in evaluating
risk to late health effects. Furthermore, dose assessment may be used to reassure non
exposed persons that they have not received any significant exposure.
The onset, nature, severity, and duration of clinical symptoms following radiation
exposure are determined primarily by the casualty’s absorbed dose. But, these factors
are also influenced by the radiation field and quality, the dose rate, the individual’s
inherent radiosensitivity and general medical health status (7).
Dosimetry is hailed as the cornerstone of radiological protection. But, non-availability
or erroneous initial dose estimates, within hours to weeks after exposure, could result
18.9% exposure is due to artificial radiation sources used in the technological sector

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
3
in sub-optimal medical intervention. In all potential radiation exposure scenarios, it is
unlikely that physical dosimeters will be available for dose assessment to aid clinical
management of mass casualties. For early treatment of radiation victims, it is
recommended that medical personnel rely heavily on clinical signs and biological
dose assessments.
Since IR induces cellular and molecular changes, the most important cellular target of
radiation damage is the deoxyribonucleic acid (DNA). It is this characteristic that is
employed in biological dosimetry. The damage that is incurred by any living cell can
be observed by diverse techniques tailored for specific end points referred to as
biomarkers. The visualization and analysis of radiation associated damage forms the
basis and origin of these biodosimeters. The advances in cell and molecular biology
make it possible to screen human population for changes in biomarkers of exposure.
To be useful, a biomarker for exposure and risk assessment should employ an end
point that is highly quantitative, stable over time, and relevant to human risk. For any
biomarker to be considered as a biodosimeter, it needs to possess some basic features.
First, sampling should be easy, and the taken sample should represent the whole body.
Second, biomarkers must change as a function of exposure or dose enabling
establishment of in vitro dose response curves. Third, it is important for the assay to
be rapid and ultimately capable of automation if biomarkers are to be useful following
low-level exposures (8). Also, the biomarker needs to be present at a very low
background level and variability between individuals should be small (9). The
important characteristics that must be considered in the design of a radiation
biodosimeter assay system for field use are: hardenability, low maintenance,
portability, including low weight and footprint, operation by non-specialist personnel
and high-throughput.
One of the earliest and most direct methods of dose determination following radiation
exposure involves charting daily counts of different cell types circulating in the
peripheral blood. Total leukocyte counts decline rapidly in the first week following
radiation exposures in excess of about 1 Gy and the extent and duration of the decline
and subsequent recovery have been shown to correlate well with dose (10). Total

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
4
body irradiation doses of 1 Gy and higher can also be well estimated from peripheral
blood neutrophil counts. The dose estimates derived from these two methods agree
closely with each other and were widely used and confirmed following the Chernobyl
accident as well as other well documented accidents at research facilities in Russia
(11).
The hematopoietic system contains some of the most radiation sensitive and easily
sampled cells in the human body. This has been exploited by many of the
biodosimetry methods developed to date, including those based on cytogenetics,
somatic mutation and gene expression. The use of these biomarkers for biodosimetry
purpose has been discussed.
1.2. Cytogenetic Biodosimetry:
1.2.1. Chromosome aberrations – Dicentric chromosome (DC):
Double strand breaks induced by radiation in individual chromosomes can give rise to
exchanges resulting in an abnormal chromosome with two centromeres referred to as
the DC. Centric Rings, which are formed due to breaks and rejoining within the same
chromosome, are also considered as equivalents of DC chromosomes. Because of the
ease of identification and the low baseline frequency, the use of DC and ring
chromosomes became the standard for detection of past radiation exposure (12, 13).
Even today scoring of DC is considered as most specific for IR damage. Very few
clastogens can be confounding factors for this assay. Biodosimetry, based on the
analysis of DC chromosomes in circulating lymphocytes, is considered the “gold
standard” for estimating radiation injury, and is used to make informed decisions
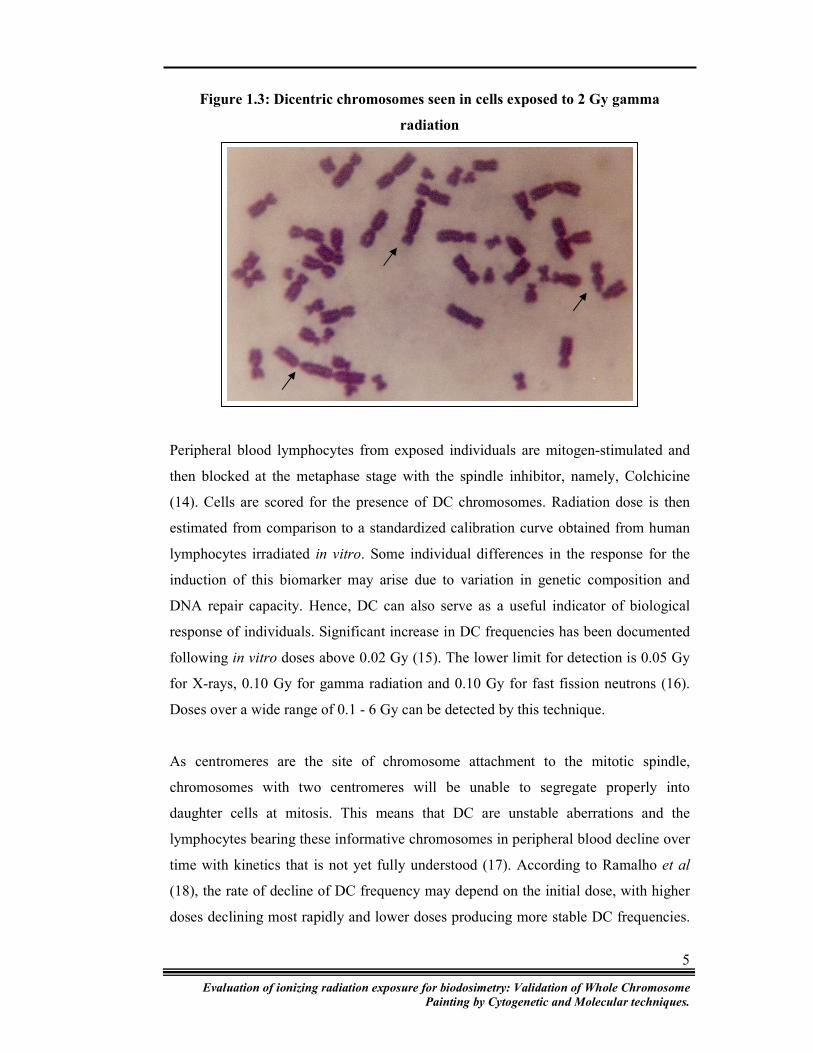
regarding the medical management of irradiated persons. Figure 1.3 shows dicentrics
induced by gamma radiation as observed by Giemsa staining.
Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
5
Figure 1.3: Dicentric chromosomes seen in cells exposed to 2 Gy gamma
radiation
Peripheral blood lymphocytes from exposed individuals are mitogen-stimulated and
then blocked at the metaphase stage with the spindle inhibitor, namely, Colchicine
(14). Cells are scored for the presence of DC chromosomes. Radiation dose is then
estimated from comparison to a standardized calibration curve obtained from human
lymphocytes irradiated in vitro. Some individual differences in the response for the
induction of this biomarker may arise due to variation in genetic composition and
DNA repair capacity. Hence, DC can also serve as a useful indicator of biological
response of individuals. Significant increase in DC frequencies has been documented
following in vitro doses above 0.02 Gy (15). The lower limit for detection is 0.05 Gy
for X-rays, 0.10 Gy for gamma radiation and 0.10 Gy for fast fission neutrons (16).
Doses over a wide range of 0.1 - 6 Gy can be detected by this technique.
As centromeres are the site of chromosome attachment to the mitotic spindle,
chromosomes with two centromeres will be unable to segregate properly into
daughter cells at mitosis. This means that DC are unstable aberrations and the
lymphocytes bearing these informative chromosomes in peripheral blood decline over
time with kinetics that is not yet fully understood (17). According to Ramalho et al
(18), the rate of decline of DC frequency may depend on the initial dose, with higher
doses declining most rapidly and lower doses producing more stable DC frequencies.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
6
Despite uncertainties in interpretation of DC frequencies obtained much later after
radiation exposure, this assay is still considered the most practical one shortly after
exposure. Measurement of DC yield has provided very reliable dose estimate during
several accidental scenarios (19, 20).
As with every technique, this assay also has some limitations. The need of a well-
experienced scorer is one of them. The scoring of DC is not only time consuming, but
also, not amenable to automation. Recent advances such as metaphase scanners and
centromeric painting by antikinetocore antibodies can enhance the scoring speed and
reduce errors in the identification of DC. Further, using the DC assay, the nature of
radiation exposure can be assessed by the distribution pattern of DC. In accidents
involving whole body exposures in the lethal range (3-6 Gy), scoring of just 25 - 50
metaphases is adequate for providing the preliminary information required for
medical management of victims. However, at higher doses most of the severely
damaged cells fail to go through cell division and it may be very difficult to find even
a few metaphases. At very low doses (0.10 to 0.20 Gy), where only a few DC are
involved, the error associated with dose estimates can be very large (16)
1.2.2. Chromosome aberrations: Translocations (TL)
Radiation induced unstable chromosomal exchanges like DC, rings and deletions are
eliminated from the body within 1-3 years depending on the exposure condition. As a
result, there is considerable uncertainty in this dosimetry for past exposures (21-22).
Scoring stable chromosomal exchanges such as TL, is a possible approach to
overcome this problem.
Studies of the Japanese A-bomb survivors and patients receiving radiotherapy have
shown TL to persist in peripheral blood lymphocytes many years after exposure and
repeated cytogenetic analyses have also indicated that the frequencies of cells with TL
remain unchanged (23-24). Thus, they are potentially a better indicator of cumulative
dose. This persistence reflects the induction of aberrations in stem cells with
subsequent constant replenishment of the mature lymphocyte pool (17).

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
7
Many laboratories initially explored the use of G-banding to identify stable
chromosomal aberrations – TL for biodosimetry purpose (25-28). Even in this modern
era of the fluorescent technology, some laboratories still prefer G-banding to FISH,
due to the cost viability of this technique (29).
Detection of TL by conventional block staining techniques will only identify those
with obvious length changes and is therefore inefficient and probably subject to scorer
bias (30). Analysis using G-banding, allows the identification of rearrangements
involving any chromosome in the genome and also the precise location of breakpoints
within each chromosome (Figure 1.4). But, it is more time consuming and tedious.
Automation of the same demands good quality of banded preparations.
Figure 1.4: Translocation between chromosomes 7 and 14 observed in peripheral
blood lymphocytes exposed to gamma radiation
Since the beginning of the 1990s, Fluorescent In Situ Hybridisation (FISH) has been
used as a cytogenetic tool for the detection of genome damage. The availability of
whole chromosome specific libraries has enabled painting of individual chromosomes
by fluorescence in situ hybridization technique (31). FISH using whole chromosome
paints enables the detection of TL involving selected chromosomes. Since this

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
8
technique does not necessarily require well-spread metaphases for analysis, it is
possible to increase the number of analyzable metaphases compared with the banding
technique.
Chromosome painting (FISH) is a simpler, more objective and practical method for
detecting chromosome rearrangements than conventional banding analyses. A number
of studies, using different combinations of painted chromosomes, have adopted the
approach of Lucas et al (32). The results point out that FISH for TL in as few as three
chromosomes, when combined with screening of numerous metaphases, provides
sensitivity comparable with that provided by G-banding, which covers the whole
genome (33-35). Figure 1.5 shows a reciprocal translocation between chromosomes 1
(WCP1 SpectrumGreen) and 3 (WCP3 SpectrumOrange) observed in a peripheral
blood lymphocyte metaphase exposed to gamma radiation.
Figure 1.5: Translocation observed in peripheral blood lymphocytes exposed to
gamma radiation by FISH painting of chromosomes 1 and 3
The translocation frequency obtained for the painted chromosomes is extrapolated to
the whole genome on the assumption that, radiation-induced exchanges are produced

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
9
randomly (36). Therefore, the data can be also examined for excesses and deficits in
the involvement of specific chromosomes or chromosome regions in rearrangements.
In principle, chromosomes from any cell can be subjected to FISH. However, the
method usually uses peripheral blood lymphocytes obtained from the individual to be
evaluated. The lymphocytes are cultured and metaphase spreads are deposited on
glass slides using standard cytogenetic methods. A cocktail of composite
chromosome-specific DNA probes can be used in conjunction with pancentromeric
probes to discriminate between TL and DC (37-39). TL involving exchange of parts
between the painted chromosomes and counterstained (unpainted) chromosomes are
visualized as bicoloured structures.
FISH assay not only makes the identification of TL very easy, but also increases the
sensitivity by its ability to score events, which the conventional banding may fail to
detect. In recent years many laboratories have explored the potential of FISH assay of
TL as a biological dosimeter (31, 36, 40-46). Since only a part of the genome is
painted (10-20%), the information for the whole genome is derived by extrapolation
of the response obtained for the painted fraction. As, it is likely that individual
chromosomes may differ in their radiosensitivity, there is a need to obtain calibration
curves with different cocktails of painted chromosomes. Some of the previous reports
suggest the usefulness of FISH assay for retrospective biological dosimetry of
radiation (34, 36, 47-51).
1.2.3. Whole genome painting: multicolour-FISH and Spectral Karyotyping
Until rather recently, it was usually assumed that virtually all chromosome exchanges
are simple, i.e. involve only two chromosome breaks. However, chromosome
"painting" techniques have now shown that complex aberrations, involving more than
two breaks in a single configuration, are common. Many whole-chromosome painting
techniques are based on FISH (31). More recent and sophisticated painting
techniques, such as mFISH or spectral karyotyping, employ combinatorial
hybridization schemes, allowing recognition of most exchanges between heterologous
chromosomes (52-56). Still further extensions of this approach allow better
recognition of exchanges between homologous chromosomes, better localization of

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
10
exchange breakpoints within a chromosome, and better recognition of inversions (57-
60). The intricate aberration spectra uncovered by mFISH/SKY give extra
information about the mechanisms and geometric aspects of radiation damage.
Whole genome analysis by SKY / mFISH is useful especially in the high dose range
to correct aberration data for complex exchange aberrations (61-62). m-FISH excites
and detects each of the five employed fluorochromes with narrow band pass filters
while SKY simultaneously excites multiple fluorophores separately with narrow
band-pass excitation/emission. These multicolor karyotyping technologies are being
used to detect subtle interchromosomal rearrangements that are otherwise below the
resolution of conventional banding methods (54). Pouzoulet et al (63) demonstrated
through their study that more TL were detected with m-FISH than with conventional
three colour FISH, and so m-FISH is expected to improve the accuracy of
chromosome aberration analyses in some situations. Figure 1.6 is a colourful picture
of chromosomal aberrations observed in peripheral blood lymphocytes exposed to
gamma radiation by mFISH whole genome painting of chromosomes.
Figure 1.6: Chromosomal aberrations observed in peripheral blood lymphocytes
exposed to gamma radiation by mFISH whole genome painting of chromosomes.
Despite their wide horizon of detection, their limitations are ill-defined. Multicolor
karyotyping results need to be interpreted with care. Structural rearrangements, which

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
11
juxtapose non-homologous chromosome material, frequently resulting in overlapping
fluorescence at the interface of translocated segments lead to “flaring.” Flaring
obscures or distorts the fluorescence pattern of adjacent chromatin, leading to
misinterpretation.
Szeles et al anticipate that radiation induced chromosomal aberrations may be more
complex than expected from conventional and single chromosome painting analyses
(64). While conventional Giemsa staining is generally accepted as the method of
choice for a triage situation, it is expected that extended mFISH / SKY analysis will
add to the knowledge of underlying mechanisms for irradiation associated
chromosomal aberrations. Some studies suggest that a 24-color FISH approach gives
a more complete picture of radiation-induced aberrations and aids in the detection and
quantification of genetically determined intrinsic radiosensitivity (65). The usefulness
of these whole genome scanning techniques can be understood while reviewing the
number of such studies being conducted these days and the explosive increase in
cytogenetic data, which, together with computer-assisted modeling, allows new
insights into the formation of radiation-induced chromosome aberrations. (53, 66-72).
1.2.4. Premature chromosome condensation (PCC)
Radiation damage can also be detected in interphase cells by the premature
chromosome condensation (PCC) assay (73). This method classically uses fusion of
the test cells with mitotic cells, which transmit a signal for dissolution of the nuclear
membrane and condensation of the interphase chromosomes as if in preparation for
mitosis. The 46 chromosomes have a single chromatid appearance (Figure 1.7). At
this stage, damage induced by radiation appears as excess breaks. Excess PCC
fragments have been shown to increase with increasing radiation exposure (74).
An important advantage of this method is that, information on the exposure can be
derived within 48 hours of obtaining the blood sample. Further, since the technique
does not involve cell division, technical factors associated with post irradiation
stimulation and progression through the cell cycle, do not interfere with the analysis.
Even in accidents involving exposure to doses in excess of 5 Gy, the cells can easily

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
12
undergo condensation (although they may fail to reach metaphase). PCC technique
would be very informative in such situations.
Figure 1.7: Chromosomal aberrations observed in peripheral blood lymphocytes
exposed to gamma radiation by the PCC assay.
The applications of this technique with respect to biodosimetry are diverse. Since
PCC is one of the techniques that allows visualization of initial DNA damage
incurred, studies pertaining to DNA break induction kinetics can be performed,
yielding useful information (75-76). The method is also suitable for studying radio-
sensitivity (77-78), partial body exposures (79), high dose exposures (80) and
improves detection of even low dose exposures (81-82). The use of PCC FISH can
simplify the analysis further by increasing the speed and accuracy of the assay (83-
85). In spite of the speed of analysis, this technique is in use in very few centres.
1.2.5. Micronucleus Assay (MN assay)
Much more than a century ago, MN have been described by many scientists. MN refer
to small nuclei formed from chromatin material, which fail to get incorporated in
either of the daughter nuclei formed during cell division. It is now known that
sometimes, even entire chromosomes can lag behind during cytokinesis and form MN
(86-87). The in vitro cytochalasin-B block methodology to score MN in cells, which
divided once in culture, became an important and useful tool to distinguish
clastogenic and aneugenic from each other. Exposure to clastogenic agents like
radiation results in a dose dependent increase in the frequency of MN.
Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
13
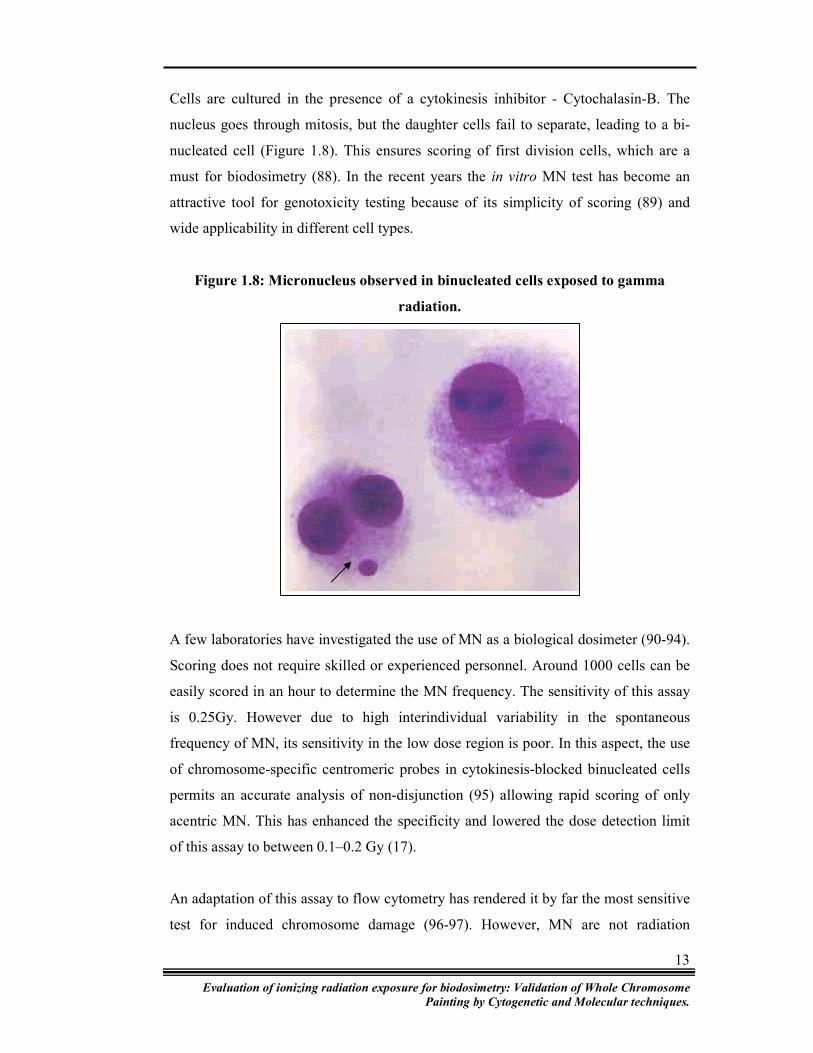
Cells are cultured in the presence of a cytokinesis inhibitor - Cytochalasin-B. The
nucleus goes through mitosis, but the daughter cells fail to separate, leading to a bi-
nucleated cell (Figure 1.8). This ensures scoring of first division cells, which are a
must for biodosimetry (88). In the recent years the in vitro MN test has become an
attractive tool for genotoxicity testing because of its simplicity of scoring (89) and
wide applicability in different cell types.
Figure 1.8: Micronucleus observed in binucleated cells exposed to gamma
radiation.
A few laboratories have investigated the use of MN as a biological dosimeter (90-94).
Scoring does not require skilled or experienced personnel. Around 1000 cells can be
easily scored in an hour to determine the MN frequency. The sensitivity of this assay
is 0.25Gy. However due to high interindividual variability in the spontaneous
frequency of MN, its sensitivity in the low dose region is poor. In this aspect, the use
of chromosome-specific centromeric probes in cytokinesis-blocked binucleated cells
permits an accurate analysis of non-disjunction (95) allowing rapid scoring of only
acentric MN. This has enhanced the specificity and lowered the dose detection limit
of this assay to between 0.1–0.2 Gy (17).
An adaptation of this assay to flow cytometry has rendered it by far the most sensitive
test for induced chromosome damage (96-97). However, MN are not radiation

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
14
specific and the background frequency seems to be much larger and variable. Hence
care needs to be taken in interpretation of observed frequency in terms of radiation
dose. Overall, the simplicity, accuracy, multi-potentiality and large tissue applicability
of the MN technology make it an attractive assay of choice.
1.2.6. Comet Assay
The “Comet Assay” (single cell gel electrophoresis) is a sensitive, rapid and relatively
inexpensive method for measuring DNA damage in individual cells (98). Single cells
are embedded in agarose on microscope slides, lysed to remove majority of the
proteins, electrophoresed and stained with a fluorescent dye in order to visualize the
DNA. When visualized using a fluorescent microscope, DNA of undamaged cells
appears as a spherical mass occupying the cavity formed by the lysed cell. Following
radiation damage, the smaller the fragment size and the greater the number of
fragments of DNA, the greater the percentage of DNA that it is able to migrate in an
electric field, forming a comet image (Figure 1.9).
Figure 1.9: DNA damage observed in peripheral blood lymphocytes exposed to
gamma radiation by the Comet assay.
The assay can be performed under alkaline conditions to examine DNA single strand
breaks (SSB), or in non-denaturing (neutral) conditions to measure double strand
breaks (DSB) in individual cells. The advantages of the technique include: collection

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
15
of data at the level of individual cells, making it possible to identify different
populations of cells within the same sample; the need for small numbers of cells per
sample; sensitivity for detecting DNA damage and that virtually any eukaryotic cell
population is amenable to analysis (99).
Many variant versions of this assay exist. One such evolved version is the halo-comet
assay. The halo-comet assay uses whole blood and does not require post sampling
incubation and is characterized as having a short processing time requirement,
yielding a potentially good slide processing profile. The persistence of the halo-comet
assay can accommodate its use in biodosimetry. (85)
As contradistinct from the normal comet assay, the halo-comet uses nuclear
suspensions. Samples are analyzed after a sufficient interval of time such that initial
strand break repairs have been substantially completed. The halo-comet analysis
exhibits persistent DNA conformational effects.
In the halo assay permeabilised cells have their nuclear protein extracted with high
salt. The DNA remains within a residual nucleus-like structure called a nucleoid. If
the nucleoid DNA contains breaks, a halo of DNA extends around the original form
of the nucleus. The presence of breaks in DNA relaxes supercoiling and loops. The
loops are free to extend outside the nuclear matrix. Measurements of the radius of the
halo give an indication of the size of the loops. In the halo-comet assay, sufficient
time is given, prior to scoring, to eliminate the transient damage repair (single strand-
double strand breaks) component and arrive at a persistent radiation dose effect
(approximately 24 hours). The benefit of cytogenetic halo-comet assay complemented
by Flow Cytometry lies in its ability to estimate partial body exposure effects.
The comet assay is suitable for in vivo human biomonitoring, especially in cases of
incidental exposure to IR (100), as a predictor of radiosensitivity (101) and a
biomarker in assessment of DNA damage during occupational exposures (99) and
predictor of radiotherapy (102-103).
1.3. Somatic Mutation Assays

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
16
Somatic cell mutations are a major cause of cancer formation and hence their
evaluation can be an important tool in predicting cancer risk for radiation-exposed
populations. Repair of DNA damage caused by radiation in hematopoietic stem cells
can result in somatic mutations in certain loci that can be monitored as biological
indicators of dose. Mutations in several different loci have been exploited for
detection of radiation exposure, including expression of glycophorin A (GPA)
variants in erythrocytes and mutations at the T-cell receptor (TCR) or hypoxanthine
guanine phosphoribosyltransferase (HPRT) loci in T-lymphocytes. These assays may
be used for individual assessment of long-term health consequences after the
irradiation, because persons with elevated frequencies of mutant cells may represent a
group at high risk in respect to oncological diseases. A drawback common to these
somatic mutation end-points is their relative lack of specificity for radiation exposure
as other environmental exposures or physiological states can also increase the
observed mutant frequencies in vivo.
1.3.1. GPA variants
The Glycophorin-A (GPA) assay in erythrocytes has been widely used for
biodosimetry. Glycophorin A is a sialoglycoprotein found exclusively and abundantly
in the red-cell membrane. The gene for glycophorin A in humans has two equally
prevalent alleles, M and N, whose gene products differ by two amino acids and are
the basis for the M and N blood types. The Glycophorin A somatic-mutation test is
limited to the 50% of humans who are MN by blood type and, hence, normally
express the M and N gene products on the surface of every red cell.
The test uses flow cytometry and differently labelled fluorescent monoclonal
antibodies to the M and N products to measure their content on the surface of each of
several million red cells. Three mutant types are scored: the M0 and N0, which are
those cells respectively lacking expression of either the N or M product but with
normal amounts of the counterproduct; and the MM, which are those cells lacking the
N product but with double amounts of the M product. M0 and N0 are interpreted as
simple loss-of-function mutations in which the Glycophorin A gene product from one
chromosome 4 has been modified (lost or changed) to the point of being
Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
17
unrecognizable by the antibody, whereas the gene product from the other
chromosome 4 is expressed normally. The M0 and N0 mutational events are
statistically independent but otherwise identically likely events and can be averaged
into a common estimate of the gene-loss endpoint. The MM appears to have two
functioning and identical genes and probably represents reduction to homozygosity
through some type of recombinational event. The background mutant frequencies are
in the range of 5–20 per million cells for each of the three endpoints, depending on
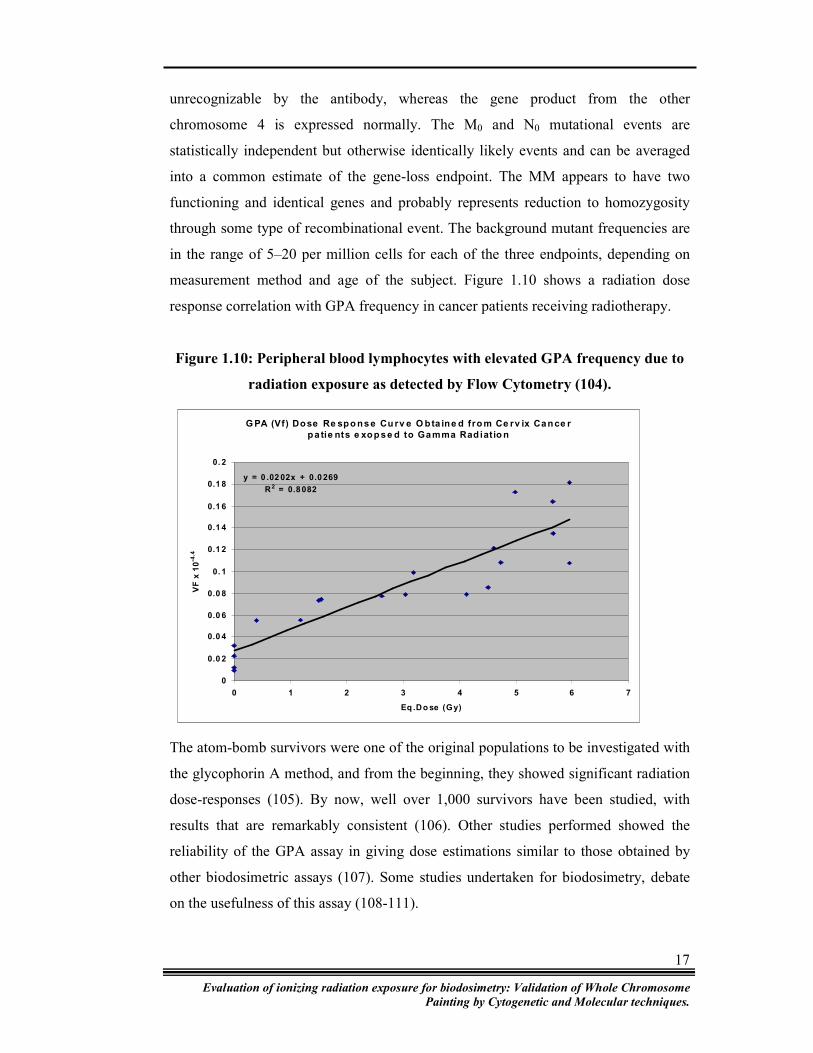
measurement method and age of the subject. Figure 1.10 shows a radiation dose
response correlation with GPA frequency in cancer patients receiving radiotherapy.
Figure 1.10: Peripheral blood lymphocytes with elevated GPA frequency due to
radiation exposure as detected by Flow Cytometry (104).
The atom-bomb survivors were one of the original populations to be investigated with
the glycophorin A method, and from the beginning, they showed significant radiation
dose-responses (105). By now, well over 1,000 survivors have been studied, with
results that are remarkably consistent (106). Other studies performed showed the
reliability of the GPA assay in giving dose estimations similar to those obtained by
other biodosimetric assays (107). Some studies undertaken for biodosimetry, debate
on the usefulness of this assay (108-111).
G PA (Vf) Dose Re spo nse Cu rv e O bta ine d fro m Ce rv ix Cance r
pa tie nts e xop se d to Gamma Rad iatio n
y = 0 .02 02x + 0.0 269
R2 = 0.8 082
0
0.0 2
0.0 4
0.0 6
0.0 8
0. 1
0.1 2
0.1 4
0.1 6
0.1 8
0. 2
0 1 2 3 4 5 6 7
Eq .Do se (Gy)
VF
x 1
0-4
.4

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
18
As any other biodosimeter, the GPA assay has its limitations. First there is the
problem of individual variability with the glycophorin A assay. The assay is
reasonably stable for repeat measurements on an individual but its standard deviation
is only slightly larger than expected for counting errors. A target population of
approximately 105 cells is needed to score an average of one mutation, and well over
106 cells to estimate a reasonable frequency of mutation. A human, particularly one
who has just been irradiated, has barely enough hematopoietic stem cells to measure a
continuous response with this test. In short, the glycophorin A test, useful as it is for
large, ordered populations, cannot function as an individual dosimeter. The only other
potentially helpful strategy is to combine the test with other mutational endpoints into
a composite estimate per person. However, this possibility is yet to be explored (112).
1.3.2. T-Cell Receptor
The T-cell receptor (TCR) mutation assay for in vivo somatic mutations is a sensitive
indicator of exposure to IR. In this light, the T-cell receptor mutation assay is known
to be a sensitive indicator of IR. This assay requires only a small volume of blood and
can be performed in a few hours.
For the TCR to be expressed on the T cell surface, the complete TCR/CD3 complex is
required. Any defect in any of the components of the TCR results in the loss of CD3
expression on the T cell surface. Therefore cell surface expression of CD3 can be
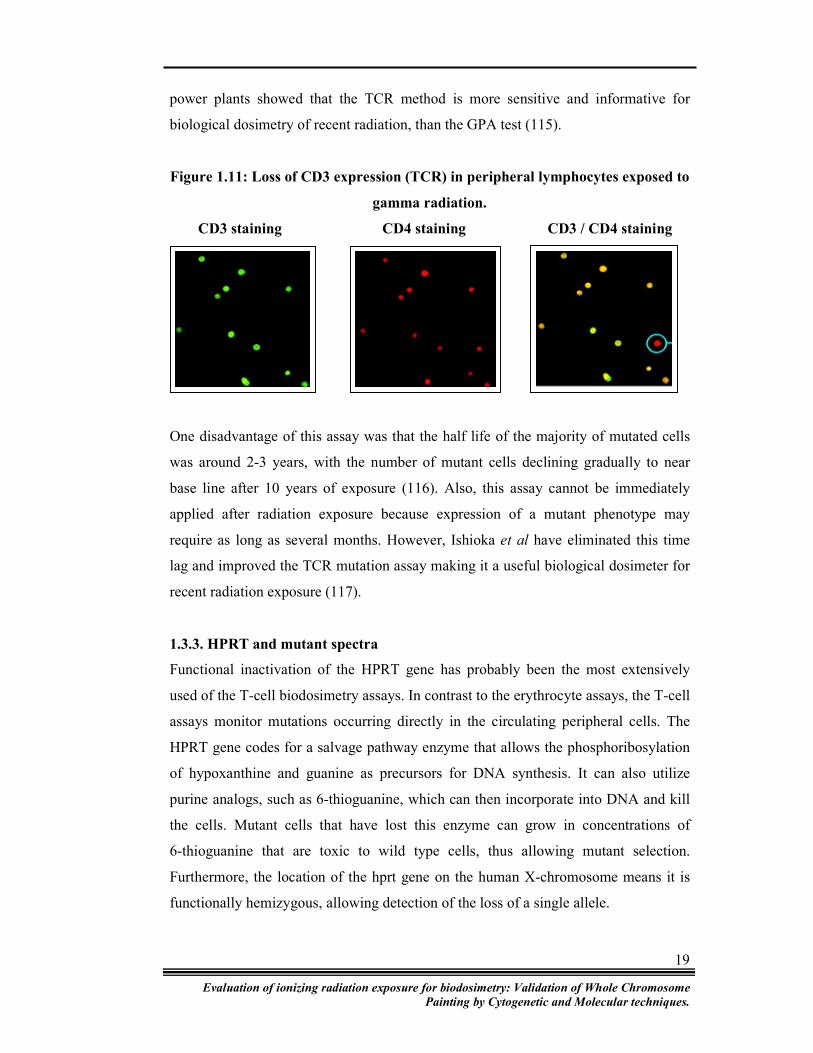
used as a marker of TCR mutation rate. Figure 1.11 shows loss of CD3 expression in
peripheral lymphocytes exposed to gamma radiation.
Flow cytometric detection of TCR mutations has made this assay easily amenable to
automation and hence numerous studies involving the TCR have been carried out for
biodosimetry purposes. In one report, the TCR mutation assay appeared to be a useful
biological dosimeter even after a period of 40 years since radiation exposure, where
internal radiation exposure was suspected (113). Smirnova et al suggest the usefulness
of the TCR assay in revealing the nature of radiation exposure (to be internally
localized) (114). Another study undertaken to determine TCR and GPA mutation
frequencies in persons exposed professionally to IR or a result of accidents at nuclear
Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
19
power plants showed that the TCR method is more sensitive and informative for
biological dosimetry of recent radiation, than the GPA test (115).
Figure 1.11: Loss of CD3 expression (TCR) in peripheral lymphocytes exposed to
gamma radiation.
CD3 staining CD4 staining CD3 / CD4 staining
One disadvantage of this assay was that the half life of the majority of mutated cells
was around 2-3 years, with the number of mutant cells declining gradually to near
base line after 10 years of exposure (116). Also, this assay cannot be immediately
applied after radiation exposure because expression of a mutant phenotype may
require as long as several months. However, Ishioka et al have eliminated this time
lag and improved the TCR mutation assay making it a useful biological dosimeter for
recent radiation exposure (117).
1.3.3. HPRT and mutant spectra
Functional inactivation of the HPRT gene has probably been the most extensively
used of the T-cell biodosimetry assays. In contrast to the erythrocyte assays, the T-cell
assays monitor mutations occurring directly in the circulating peripheral cells. The
HPRT gene codes for a salvage pathway enzyme that allows the phosphoribosylation
of hypoxanthine and guanine as precursors for DNA synthesis. It can also utilize
purine analogs, such as 6-thioguanine, which can then incorporate into DNA and kill
the cells. Mutant cells that have lost this enzyme can grow in concentrations of
6-thioguanine that are toxic to wild type cells, thus allowing mutant selection.
Furthermore, the location of the hprt gene on the human X-chromosome means it is
functionally hemizygous, allowing detection of the loss of a single allele.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
20
An assay using T-cell cloning and hprt mutant fraction determination has been used to
show a strong relationship between dose and induced mutations in atomic bomb
survivors and patients receiving high doses of radiation therapy (118). An increase in
hprt mutant fractions may also be detectable following lower dose exposures but these
results seem more variable depending on the time of sampling (119). Albertini et al
show that radiation quality affects both the efficiency of induction and the molecular
spectrum of HPRT mutations in human T lymphocytes both in vitro and in vivo. The
mutational spectrum may be relatively specific for radiations of different quality and
thus allow a more precise measurement of the induction of somatic gene mutations
resulting from individual exposures to radiation, thereby providing more sensitive
assessments of health risks (120). The results of the study conducted by Thomas et al
illustrate the sensitivity of HPRT somatic mutation as a biomarker for populations
with low dose radiation exposure, and the dependence of this sensitivity on time
elapsed since radiation exposure (121). Moreover, the HPRT mutational assay has
also been shown to reveal dose rate differences and hence serves as a useful parameter
for risk estimation in radiation protection (122).
1.4. Molecular biodosimetry
1.4.1. Molecular profiling by gene expression
Mutation induction in cells directly exposed to radiation is currently regarded as the
main component of the genetic risk of IR for humans. Recent technological advances
may allow an additional exploitation of the molecular responses of cells to IR.
Radiation dose, dose rate, radiation quality, and elapsed time since exposure result in
variations in the response of stress genes suggesting that gene expression signatures
may be informative markers of radiation exposure. Irradiation initiates a plethora of
signal transduction cascades responsible for maintaining cellular homeostasis and
promoting interactions with neighboring cells. Large-scale changes in gene expression
have also been found after irradiation, and microarrays have helped discern these
subsequent transcriptional alterations.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
21
Exposure of cells to DNA-damaging agents elicits a highly complex molecular
response, much of which is mediated through changes in gene expression. A
transcriptional response to genotoxic stress, estimated to involve 1% or more of the
genome, was initially identified in yeast and similar complex transcriptional responses
were soon confirmed in mammalian cells (123-124). The stress response pathways
responding to different environmental and physiological stresses have many
overlapping components, including growth factors, cytokines, oncogenes and genes
involved in cell cycle, apoptosis, signaling pathways and DNA repair. The recent
development of functional genomic approaches to simultaneously quantify expression
of thousands of genes in a single experiment may allow the determination of
expression signatures indicative of exposure to IR or other environmental toxins.
Although earlier it was in the speculative realm, this approach is highly attractive as it
is amenable to rapid, even automated, non-invasive analysis and may additionally
have the potential to discern competing effects from incidents involving different
quality radiations or mixed chemical and physical exposure components.
A number of high-throughput gene expression measurement methods are currently
available, including serial analysis of gene expression (SAGE), oligonucleotide arrays
and cDNA arrays (125-126). The premise is developed that stress gene responses can
be employed as molecular markers for radiation exposure using a combination of
informatics and functional genomics approaches (17). A generalised post-exposure
profile may identify exposed individuals within a population, while more specific
fingerprints may reveal details of a radiation exposure. Changes in gene expression in
human cell lines occur after as little as 0.02 Gy rays, and in peripheral blood
lymphocytes alter as little as 0.2 Gy (127).
Accumulated evidence also implies that the biological effects of low-dose and high-
dose IR are not linearly distributed. According to the study done by Ding et al the
predominant functional groups responding to low-dose radiation are those involved in
cell-cell signaling, signal transduction, development and DNA damage response
(128). At high dose, the responding genes are involved in apoptosis and cell
proliferation. Interestingly, several genes, such as cytoskeleton components ANLN
and KRT15 and cell-cell signaling genes GRAP2 and GPR51, were found to respond

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
22
to low-dose radiation but not to high-dose radiation. Pathways that are specifically
activated by low-dose radiation are also evident. These quantitative and qualitative
differences in gene expression changes may help explain the non-linear correlation of
biological effects of IR from low dose to high dose.
While some studies have focused on low dose-rate experiments, others have analyzed
the gene expression response of IR compared to other DNA damaging agents. Very
few genes have been found to be consistently up-regulated by IR, but that set includes
GADD45, CDKN1A, connexin-43 and genes associated with the nucleotide excision
repair pathway (129-130)
Low doses of radiation have an identifiable biosignature in human tissue, irradiated in
vivo with normal intact three-dimensional architecture, vascular supply, and
innervation. The ability to detect a distinct radiation response pattern following low
dose IR exposure has important implications for risk assessment in both therapeutic
and national defense contexts (131).
1.5. Protein biodosimetry
1.5.1. Gamma h2ax assay
The need to detect DNA damage by radiation requires specific markers that can be
easily seen and quantified, and gamma-h2ax foci formation is one such event that can
be used in this scenario. An early event after introduction of DSB is the
phosphorylation of a special form of histone 2A, denoted h2ax that is part of 10% of
all nucleosomes in the cell (88, 132-133). Histone 2AX contains a distinct C-terminal
extension, with a consensus phosphorylation at serine 139. It is known that h2ax
phosphorylation is specific to sites of DNA damage and is also indicative of amount
of DNA damage (134). However, in order to use gamma-h2ax as a quick screening
tool, it must be optimized for sensitivity and rapidity.
Gamma-h2ax can be tagged with a fluorescently labeled antibody, and can then be
detected with excellent sensitivity using in situ image analysis (Figure 1.12). IR is an
efficient inducer of DSB and most of the early research on gamma-h2ax has been

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
23
done with IR (135-136). Both DSB and thus gamma-h2ax are formed linearly with
dose from very low to extremely high (> 10 Gy) doses (135). It has also been shown
that exposure to 10-3Gy of X-rays, induces a significant elevation in h2ax
phosphorylation in human fibroblasts (137), making it more sensitive to low doses.
The gamma-h2ax system well complements the MN system as a radiation
biodosimeter (138), requiring much shorter processing times as the cells do not have
to be cultured. Furthermore, the gamma-h2ax foci reach their maximum value within
about 30 minutes of irradiation (138), decaying over 24-36 hours post-exposure (139).
This is contrasted with MN that appear about 24 hours post-exposure and decay over
months or years (140).
Figure 1.12: Fluorescent antibody tagged detection of gamma h2ax: each signal
represents a double strand break.
The local formation of gamma-h2ax allows microscopical detection of distinct foci by
fluorescent gamma-h2ax-specific antibodies that most likely represent a single DSB
(137-138, 141). The potential to detect a single focus within the nucleus makes this
the most sensitive method currently available for detecting DSB in cells. This method
is, however, labor-intensive and will be difficult to adapt in clinical practice. In
contrast, flow cytometry allows simple detection of gamma-h2ax in a large number of
cells (142). Several reports show that the level of gamma-h2ax as detected by flow

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
24
cytometry correlates well with the number of DNA strand breaks, to the level of cell
death and radiosensitivity (135, 143-144).
1.6. Electron paramagnetic resonance
Electron paramagnetic resonance (EPR) is a physical measurement of absorbed dose
that can be applied to biological material. Although samples of fingernails and
clothing have been used in ESR determinations of dose shortly after exposure, dental
enamel is the most widely used material.
1.6.1. Electron paramagnetic resonance of dental enamel
Tooth enamel as a detector for in vivo dosimetry has been known for more than three
decades (145). The usefulness of enamel for dosimetry results from its high content of
hydroxyapatite (146). Carbonate impurities, which are incorporated into or attached to
the surface of hydroxyapatite crystals during formation, are converted to CO2 radicals
through absorption of IR (147). The concentration of radicals increases with absorbed
dose. The intensity of the resultant EPR absorption is a measure for the absorbed
dose. Examples of the use of EPR in dose reconstruction include the dose evaluation
of survivors of the atomic bomb explosions in Hiroshima and Nagasaki (148-149),
nuclear workers in the South Urals (150), residents of the Techa river basin (151), the
populations of contaminated areas in the Urals (152), the population living near the
Chernobyl nuclear reactor (153) and workers in the Chernobyl Sarcophagus (154).
Finally, EPR dosimetry was applied to a population from an uncontaminated area in
Russia (155) demonstrating the potential to estimate the absorbed dose from natural
background radiation.
International comparisons on EPR tooth dosimetry were carried out (156-160). These
comparisons were designed to check the consistency and reliability of EPR dose
reconstruction among different laboratories. These comparisons led to critical
revisions and improvements to the different variations of the EPR dosimetry method
applied by the participants. Moreover, the capability of EPR dosimetry to measure
low doses in the range of 100 mGy was demonstrated. Today, EPR dosimetry with
tooth enamel is a leading method for retrospective dosimetry of individual radiation
exposures.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
25
Teeth that are extracted for health reasons are readily available for dose reconstruction
and can be archived for a prolonged period before examination. EPR dosimetry has
the capability to measure the volume of samples required for epidemiological studies
(159). Dose reconstruction can be applied to the distinctive tissues that comprise a
tooth, namely enamel, and dentine. Tooth enamel is preferred in retrospective
dosimetry because this tissue is completely formed in childhood and once formed, is
never remodelled, even after abrasion. Therefore, the accumulated concentration of
radiation-induced radicals in the exposed enamel is preserved. At 25°C, a lifetime of
107 years was determined for the CO2 radicals in fossil tooth enamel (161). Hence,
EPR dosimetry with tooth enamel is suitable for dose reconstruction after long periods
of exposure and for many years after the exposure. The complementary measurement
of the absorbed dose in dentine offers the possibility to measure the dose resulting
from ingested radionuclides deposited in dentine, in addition to the dose from external
sources. This technique has been applied to the measurement of the accumulated dose
resulting from the intake of the bone-seeking radionuclide strontium (151).
There are several strong indications that EPR dosimetry gives correct and accurate
dose assessment even long after the exposure event. Among them are the results of
two international comparisons (162-163), several blind comparisons of the results of
EPR dose reconstruction, and data of operational personal monitoring for nuclear
workers (150-152, 164).
However, there are also certain shortcomings of EPR biodosimetry (165). It is not
always possible to obtain extracted teeth from all individuals in the study group. For
bone-seeking radionuclides (e.g. Sr-90) the reconstruction of the individual dose is
complicated and in certain cases impossible (166). EPR dose reconstruction
procedures are also considered to be time-and labour-consuming. For these reasons, at
present, EPR biodosimetry will likely not be used as the sole method applied to large
cohorts, but will remain invaluable for validation purposes.
Eva
lua
tion
of
ion
izin
g r
ad
iati
on
exposu
re f
or
bio
do
sim
etry
: V
ali
da
tio
n o
f W
hole
Ch
rom
oso
me
Pain
tin
g b
y C
yto
gen
etic
an
d M
ole
cula
r te
chn
iqu
es.
26
Co
mp
aris
on
of
som
e b
iod
osi
metr
y m
eth
od
s fo
r r
ad
iati
on
exp
osu
re
Meth
od
olo
gy
Ass
ay
Base
lin
e
varia
bil
ity
Ap
pli
cab
le
ra
nge (
Gy)
Op
tim
al
test
perio
d a
fter
exp
osu
re
Va
lid
ity (
Tim
e)
Can
be
au
tom
ate
d
Cy
togen
eti
c
Dic
entr
ics
Mod
erat
e 0.
1 -
5 D
ays
- w
eeks
S
ever
al y
ears
N
o
PC
C
Mod
erat
e 0.
1 -
20
Hou
rs -
day
s D
ays
- w
eeks
Y
es
Mic
ronu
clei
M
oder
ate
0.3
- 5
Day
s -
wee
ks
1 ye
ar h
alf-
life
Y
es
FIS
H
Mod
erat
e 0.
1 -
5 R
etro
spec
tive
Y
ears
N
o
Com
et a
ssay
M
oder
ate
0.02
- 1
0 H
ours
- d
ays
Yea
rs
Yes
So
mati
c
mu
tati
on
GP
A b
y fl
ow
cyto
met
ry
Mod
erat
e 0.
3 -
5 R
etro
spec
tive
In
defi
nite
(ye
ars)
Y
es
HP
RT
– T
-cel
l cl
onin
g M
oder
ate
1 -
2 R
etro
spec
tive
T
rans
ient
(<
1 ye
ar)
No
T-c
ell
rece
ptor
by
flow
cyt
omet
ry
Mod
erat
e 0.
4 -
2 R
etro
spec
tive
F
ew y
ears
Y
es
Mole
cu
lar
Gen
e ex
pres
sion
L
ow
0.2
- 8
Hou
rs -
day
s A
t le
ast
72 h
Y
es
Prote
in
Gam
ma
h2ax
by
flow
cyt
omet
ry
Mod
erat
e 0.
1 -
10
Hou
rs -
day
s T
rans
ient
Y
es
E
SR
T
o 3-
fold
>
0.5
R
etro
spec
tive
In
defi
nite
(ye
ars)
N
o

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
27
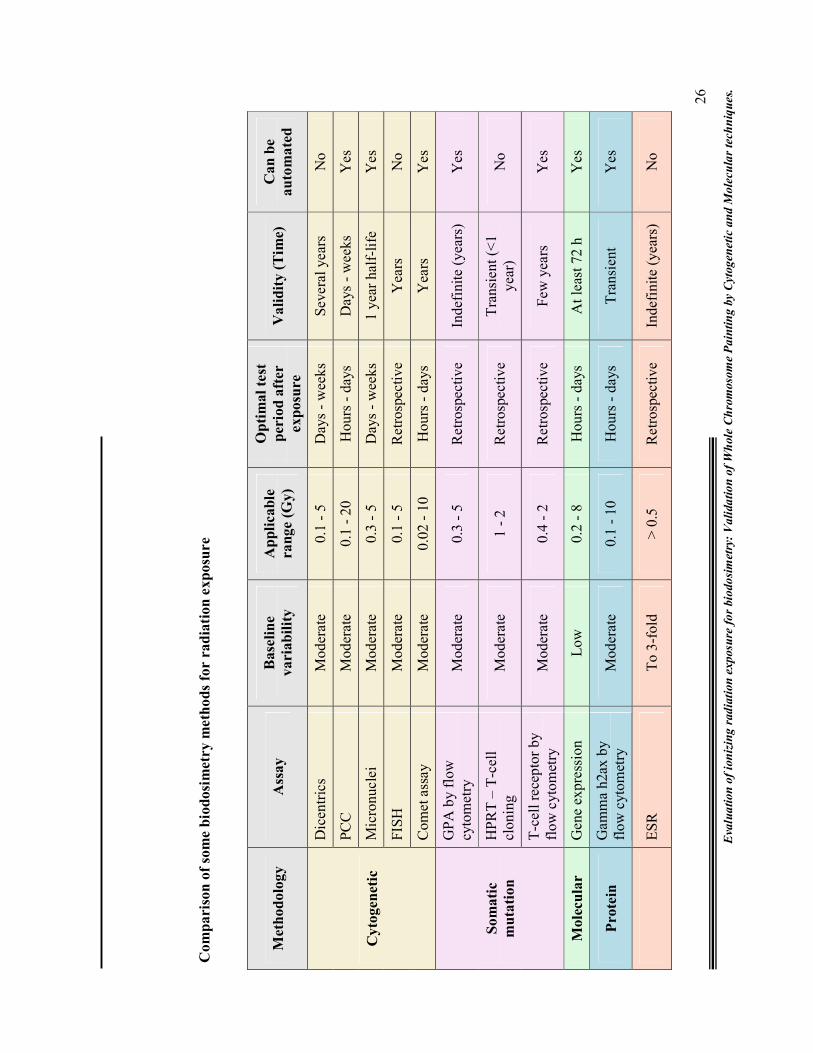
1.7. Biodosimetry - Which assay is reliable and future trends?
In response to radiation exposure, rapid and reliable dose estimates are crucial for risk
assessments during investigation of real or suspected exposed victims. The
employment of biodosimetry can represent more than a complementary methodology
to physical dosimetry in individual monitoring. Quantification of the biologically
relevant dose is required for the establishment of cause-and-effect between radiation
dose and important biological outcomes. Most epidemiological studies of
unanticipated radiation exposure fail to establish cause and effect because of an
inability to construct a valid quantification of dose for the exposed population.
Knowledge about the quantity of absorbed dose, a number together with its unit, is
certainly not sufficient to evaluate the risks associated with radiation exposure. In
addition, the comparison between the changes in biological indicators as a result of an
irradiation, with the same alterations caused by other physicochemical agents may be
important for better understanding of radiation hazards and the risks associated with
them. This will aid professionals, as well as lay people, in a better observance of
radioprotection practices.
With a host of assays available, it is difficult to say which is the best one. No single
biodosimetric technique (biophysical or biological) meets all the requirements of an
ideal dosimeter and thus qualifies as a "gold standard." An approach of using
combined dosimetry from the most appropriate methods in a given situation has been
advocated. However cytogenetic methods of dose determination, have dominated the
biodosimetry arena due to their precise and increasingly informative nature, in the
time immediately following exposure. It should be noted, however, that some of these
more complex analyses may require a high degree of expertise and so may be more
useful for research purposes than in the field (17).
Several attempts have already been made to automate biodosimetric assays (167).
During the last decade many engineers and scientists have teamed-up in the quest for
designing automated systems for medical applications featuring higher and higher
throughputs. Many innovations and new developments with motorized microscopes,

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
28
automated slidescanning workstations, digital imaging, and the associated computer
software programs have made the cytogenetic biodosimetry process more efficient
and less time consuming. For example, automated systems can quickly scan a
microscope slide and locate the scoreable cells in minutes compared to the hours
required using manual methods. Several prototypes have been designed (168).
The search for biomarkers of effective dose and the early effects of ionizing radiation
exposure in both humans and experimental animals has a history spanning several
decades and is still an ongoing process. Blood cells and serum have proven to be
abundant sources of human radiation biomarkers, including those of DNA damage
and repair, chromosomal aberrations, serum proteomic profiles, and gene expression
profiles determined by both microarrays and RT-PCR. The use of global profiling
technologies has contributed substantially to the understanding of the radiation
cellular stress response and has contributed to the elucidation of many of the complex
biological networks associated with gene expression and signal transduction. Now,
identification of radiation-induced metabolic changes is under study for development
of reliable metabolomic markers to assess radiation exposure and extent of injury.
However, it is unlikely that the more established, tested and well proven
biodosimetric assays, such as cytogenetic biodosimetry will become outdated.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
29
1.8. Aims and objective of the study:
As nuclear activity in India is on the rise due to the immense potential of nuclear
energy, radiation monitoring and safety bodies like the Atomic Energy Regulatory
Board (AERB), India, are preparing various centers all across the country for
biodosimetry purpose. Hence, the current study served as a good attempt in
establishing biodosimetry through the construction and validation of the in vitro dose
response curves. As many biodosimetry assays are available, there is a need to
validate the usefulness of these assays. Cytogenetic assays have been extensively used
for biodosimetry purpose since the 1960’s. The cytogenetic Fluorescent In Situ
Hybridization assay (FISH) is a powerful tool aiding in the identification and
enumeration of both stable and unstable chromosomal aberrations. In light of this, the
present study aimed to validate and establish its usefulness for biodosimetry purpose.
Hence the specific objectives of the study comprised of:
a) Construction of in vitro dose response curves for Co-60 gamma rays by the
FISH assay using dicentrics and translocations as endpoints.
b) Validation of in vitro FISH based dicentric yields using:
i) In vitro dicentric yields for Cobalt-60 gamma rays by Fluorescence Plus
Giemsa assay.
ii) In vitro micronucleus yields for Cobalt-60 gamma rays.
c) Validation of in vitro FISH based translocation yields using:
i) In vitro translocation yields for Cobalt-60 gamma rays by G-banding.
ii) In vitro translocation yields for Cobalt-60 gamma rays by mFISH.
iii) To check if radiation induced DNA damage is random or non random.
d) Validation of in vivo FISH based translocation frequency using G-banding
e) Impact of radiation on certain genes and search for newer biodosimetry
techniques at the gene level.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
30
1.9. References
1. Key World Energy Statistics (PDF). International Energy Agency; 2006.
2. Nuclear power plants information. IAEA; 2005.
3. World nuclear power reactors 2005-2006. Australian Uranium Information
Centre; 2006.
4. Awa AA, Honda T, Sofuni T, Neriishi S, Yoshida MC, Matsui T. Chromosome-
aberration frequency in cultured blood cells in relation to radiation dose of A-
bomb survivors. Lancet. 1971; 2(7730): 903-905.
5. Granath F, Darroudi F, Auvinen A, Ehrenberg L, Hakulinen T, Natarajan A,
Rahu M, Rytomaa T, Tekkel M, Veidebaum T. Retrospective dose estimates in
Estonian Chernobyl clean-up workers by means of FISH. Mutat Res. 1996;
369(1-2): 7-12.
6. Mettler Jr FA, Voelz GL. Current concepts: Major radiation exposure - What to
expect and how to respond. N Engl J Med. 2002; 346: 1554-1561.
7. Prasanna PGS, Muderhwa JM, Miller AC, Grace MB, Salter CA, Blakely WF.
Diagnostic Biodosimetry Response for Radiation Disasters: Current Research
and Service Activities at AFRRI. 1994; RTO-MP-HFM-108.
8. Brooks A. Biomarkers of exposure, sensitivity and disease. 1999; Int J Radiat
Biol. 75(12): 1481-1503
9. Pala FS, Köksal G.
http://kutuphane.taek.gov.tr/internet_tarama/dosyalar/cd/3881/Aplication/Applic
ation-76.PDF.
10. Baranov AE, Guskova AK, Nadejina NM, Nugis VY. Chernobyl.experience:
biological indicators of exposure to ionizing radiation. Stem Cells. 1995; 13 (1):
69-77
11. Vorobiev AI. Acute radiation disease and biological dosimetry in 1993. Stem
Cells. 1997; 15(2): 269–274.
12. Bender MA, Gooch PC. Types and rates of X-ray-induced chromosome
aberrations in human blood irradiated in vitro. Proc Natl Acad Sci. 1962; 48:
522-532.
13. Bender MA, Awa AA, Brooks AL, Evans HJ, Groer PG, Gayle-Littlefield L,
Pereira C, Preston RJ Wachholz BW. Current status of cytogenetic procedures to

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
31
detect and quantify previous exposures to radiation. Mutat. Res. 1988; 196: 103-
159.
14. Moorhead PS, Nowell PC, Mellman WJ, Battips DM, Hungerford DA.
Chromosome preparations of leukocytes cultured from human peripheral blood.
Exp Cell Res. 1960; 20: 613-616.
15. Lloyd DC, Edwards AA, Leonard A Deknudt GL, Verschaeve L, Natarajan AT,
Darroudi F, Obe G, Palitti F, Tanzarella C, Tawn EJ. Chromosomal aberrations
in human lymphocytes induced in vitro by very low doses of X-rays. Int J Radiat
Biol 1992; 61: 335-343.
16. Rao BS. Biological indicators of absorbed radiation and biological dosimetry.
Bhabha Atomic Research Centre (BARC) Newsletter. 2002; 224.
17. Amundson SA, Bittner M, Meltzer P, Trent J, Fornace AJ Jr. Biological
indicators for the identification of ionizing radiation exposure in humans. Expert
Rev Mol Diagn 1(2). 2001; 211-219.
18. Ramalho AT, Costa ML, Oliveira MS. Conventional radiation-biological
dosimetry using frequencies of unstable chromosome aberrations. Mutat Res.
1998; 404(1-2): 97-100.
19. Sevankaev AV, Lloyd DC, Edward AA, Moiseenko VV. High exposure to
radiation received by workers inside the Chernobyl Sarcophagus. Radiat Prot
Dosimetry 1995; 59: 85-91.
20. Ramahlo AT, Nascimento ACH, Natarajan AT. Dose assessment by cytogenetic
analysis in the Goiania (Brazil) radiation accident. Radia Prot Dosimetry.1998;
25: 97-100.
21. Biological Dosimetry: Chromosome aberration analysis for dose assessment.
1986; STI/PUB/10/260 (Vienna: IAEA).
22. Sreedevi B, Rao BS, Bhatt B. Radiation-Induced Chromosome Aberration
Yields Following an Accidental Non-Uniform Exposure. Radiat Prot Dosimetry.
1993; 50: 45-49.
23. Ohtaki K. G-banding analysis of radiation-induced chromosome damage in
lymphocytes of Hiroshima A-bomb survivors. Jpn J Hum Genet. 1992; 37(4):
245-262.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
32
24. Ohtaki K, Sposto R, Kodama Y, Nakano M, Awa AA. Aneuploidy in somatic
cells of in utero exposed A-bomb survivors in Hiroshima. Mutat Res. 1994;
316(1): 49-58.
25. Awa A. Analysis of chromosome aberrations in atomic bomb survivors for dose
assessment: studies at the Radiation Effects Research Foundation from 1968 to
1993. Stem Cells. 1997; 15(2): 163-173.
26. Kodama Y, Ohtaki K, Awa AA, Nakano M, Itoh M, Nakamura N. The F value
for chromosome aberrations in atomic bomb survivors does not provide evidence
for a primary contribution of neutrons to the dose in Hiroshima. Radiat Res.
1999; 152(5): 558-562.
27. Tawn EJ, Whitehouse CA, Holdsworth D, Morris S, Tarone RE. Chromosome
analysis of workers occupationally exposed to radiation at the Sellafield nuclear
facility. Int J Radiat Biol. 2000; 76(3): 355-365.
28. Ankina MA, Zavitaeva TA, Semenova TG, Sevan'kaev AV, Baranov AE,
Gus'kova AK, Nugis VIu. Analysis of stable chromosome aberrations using G-
banding stain in lymphocyte of patients with high dose radiation injury. Radiats
Biol Radioecol. 2001; 41(1): 43-47.
29. Tawn EJ, Whitehouse CA. Chromosome intra and inter-changes determined by
G-banding in radiation workers with in vivo exposure to plutonium. J Radiol
Prot. 2005; 25(1): 83-88.
30. Nakano M, Kodama Y, Ohtaki K, Itoh M, Delongchamp R, Awa AA, Nakamura
N. Detection of stable chromosome aberrations by FISH in A-bomb survivors:
comparison with previous solid Giemsa staining data on the same 230
individuals. Int J Radiat Biol. 2001; 77(9): 971-977.
31. Pinkel D, Gray J, Trask B, van den Engh G, Fuscoe J, van Dekken H.
Cytogenetic analysis by in situ hybridization with fluorescently labeled nucleic
acid probes. Cold Spring Harbor Symposia on Quantitative Biology. 1986; 51:
151-157.
32. Lucas JN, Tenjin T, Straume T, Pinkel D, Moore D, Litt M, Gray J. Rapid
human chromosome aberration analysis using fluorescence in situ hybridization.
Int J Radiat Biol 1989; 56(1): 35-44.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
33
33. Obukhova TN, Domracheva EV. Registration of stable aberrations in peripheral
blood lymphocytes using G-differential chromosome staining and fluorescence
in situ hybridization method. Radiats Biol Radioecol. 1998; 38(6): 793-799.
34. Sevan'kaev AV, Golub EV, Khvostunov IK, Potetnia OI, Shkavrova TG,
Skvortsov VG, Ivannikov AI, Tikunov DD, Sidorov OS, Amiev GN.
Retrospective dose estimation in remote period after exposure using different
biological methods. Radiats Biol Radioecol. 2004; 44(6): 637-652.
35. Kodama Y, Ohtaki K, Nakano M, Hamasaki K, Awa AA, Lagarde F, Nakamura
N. Clonally expanded T-cell populations in atomic bomb survivors do not show
excess levels of chromosome instability. Radiat Res. 2005; 164(5): 618-626.
36. Lucas JN, Awa A, Straume T, Poggensee M, Kodama Y, Nakano M, Ohtaki K,
Weier HU, Pinkel D, Gray J, Littlefield G. Rapid translocation frequency
analysis in humans decades after exposure to ionizing radiation. Int J Radiat
Biol. 1992; 62: 53-63.
37. Bauchinger M, Salassidis K, Braselmann H, Volzilova A, Pressl S, Stephan G,
Snigiryova G, Kozheurov VP. FISH-based analysis of stable translocation in a
Techa river population. Int J Radiat Biol. 1998; 73: 605-612.
38. Fomina J, Darroudi F, Boei J, Natarajan A. Discrimination between complete
and incomplete chromosome exchanges in X irradiated human lymphocytes
using FISH with pan-centromeric and chromosome specific DNA probes in
combination with telomeric PNA probe. Int J Radiat Biol. 2000; 76: 807-813.
39. Benkhaled L, Barrios L, Mestres M, Caballin MR, Ribas M, Barquinero JF.
Analysis of gamma-rays induced chromosome aberrations: a fingerprint
evaluation with a combination of pan-centromeric and pan-telomeric probes. Int
J Radiat Biol. 2006; 82(12): 869-875.
40. Tucker JD, Ramsey MJ, Lee DA, Minkler. Validation of chromosome painting
as a biological dosimeter in human peripheral blood lymphocytes following
acute exposure to ionizing radiation in vitro. Int J Radiat Biol. 1993; 64: 27-37.
41. Natarajan AT, Balajee AS, Boei JJ, Chatterjee S, Darroudi F, Grigorova M,
Noditi M, Oh HJ, Slijepcevic P, Vermeulen S. Recent developments in the
assessment of chromosomal damage. Int J Radiat Biol. 1994; 66: 615-623.
42. Tucker JD. Fish cytogenetics and the future of radiation biodosimetry. Radiat
Prot Dosimetry. 2001; 97(1): 55-60.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
34
43. Tucker JD. Sensitivity, specificity, and persistence of chromosome
translocations for radiation biodosimetry. Mil Med. 2002; Feb; 167(2): 8-9.
44. Terzoudi GI, Pantelias GE. Cytogenetic methods for biodosimetry and risk
individualisation after exposure to ionising radiation. Radiat Prot Dosimetry.
2006; 122(1-4): 513-520.
45. Chaizhunusova N, Yang TC, Land C, Luckyanov N, Wu H, Apsalikov KN,
Madieva M. Biodosimetry study in Dolon and Chekoman villages in the vicinity
of Semipalatinsk nuclear test site. J Radiat Res (Tokyo). 2006; 47: A165-A169.
46. Fucic A, Znaor A, Strnad M, van der Hel O, Aleksandrov A, Miskow S, Grah J,
Sedlar M, Jazbec AM, Ceppi M, Vermeulen R, Boffetta P, Norppa H, Bonassi S.
Chromosome damage and cancer risk in the workplace: the example of
cytogenetic surveillance in Croatia. Toxicol. Lett. 2007; 172(1-2): 4-11.
47. Rao BS, Natarajan AT. Retrospective biological dosimetry of absorbed
radiation. Rad. Prot Dosimetry. 2001; 95: 17-23.
48. Maznyk NA, Vinnikov VA. Possibilities and limitations of fluorescence in situ
hybridization technique in retrospective detection of low dose radiation exposure
in post-chernobyl human cohorts. Tsitol Genet. 2005; 39(4): 25-31.
49. Moore DH, Tucker JD. Biological dosimetry of Chernobyl clean-up workers:
Inclusion of age and smoking data provide improved radiation dose estimates.
Radiat Res. 1999; 152:655-664.
50. Cigarran S, Barquinero JF, Barrios L, Ribas M, Egozcue J, Caballin MR.
Cytogenetic analyses by fluorescence in situ hybridization (FISH) in hospital
workers occupationally exposed to low levels of ionizing radiation. Radiat Res.
2001; 155: 417-423.
51. Stephan G, Pressl S. Chromosome aberrations in human lymphocytes analysed
by fluorescence in situ hybridization in radiation workers 11 years after an
accidental radiation exposure. Int. J. Radiat. Biol. 1997; 71: 293-299.
52. Schrock E, Padilla-Nash H. Spectral karyotyping and multicolor fluorescence in
situ hybridization reveal new tumor-specific chromosomal aberrations. Semin
Hematol. 2000; 37: 334-347.
53. Cornforth MN, Greulich-Bode KM, Loucas BD, Arsuaga J, Vázquez M, Sachs
RK, Brückner M, Molls M, Hahnfeldt P, Hlatky L, Brenner DJ. Chromosomes

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
35
are predominantly located randomly with respect to each other in interphase
human cells. J Cell Biol. 2002; 159(2): 237-244.
54. Greulich KM, Kreja L, Heinze B, Rhein AP, Weier HG, Bruckner M, Fuchs P,
Molls M. Rapid detection of radiation-induced chromosomal aberrations in
lymphocytes and hematopoietic progenitor cells by mFISH. Mutat. Res. 2000;
452(1): 73-81.
55. Speicher MR, Ballard SG, Ward DC. Karyotyping human chromosomes by
combinatorial multi-fluor FISH. Nat Genet.1996; 12: 368-375.
56. Lee C, Gisselsson D, Jin C, Nordgren A, Ferguson DO, Blennow E, Fletcher JA,
Morton CC. Limitations of chromosome classification by multicolor
karyotyping. Am J Hum Genet. (2001); 68: 1043-1047.
57. Fauth C, Speicher MR. Classifying by colors: FISH-based genome analysis.
Cytogenet Cell Genet. 2001; 93: 1-10.
58. Saracoglu K, Brown J, Kearney L, Uhrig S, Azofeifa J, Fauth C, Speicher MR,
Eils R. New concepts to improve resolution and sensitivity of molecular
cytogenetic diagnostics by multicolor fluorescence in situ hybridization.
Cytometry. 2001; 44: 7-15.
59. Karhu R, Ahlstedt-Soini M, Bittner M, Meltzer P, Trent JM, Isola J.
Chromosome arm-specific multicolor FISH. Genes Chromosomes Cancer. 2001;
30: 105-109.
60. Wiegant J, Bezrookove V, Rosenberg C, Tanke HJ, Raap AK, Zhang H, Bittner
M, Trent JM, Meltzer P. Differentially painting human chromosome arms with
combined binary ratio-labeling fluorescence in situ hybridization. Genome Res.
2000; 10: 861-865.
61. Braselmann H, Kulka U, Baumgartner A, Eder C, Muller I, Figel M,
Zitzelsberger H. SKY and FISH analysis of radiation-induced chromosome
aberrations: a comparison of whole and partial genome analysis. Mutat Res.
2005; 578(1-2): 124-133.
62. Anderson RM, Tsepenko VV, Gasteva GN, Molokanov AA, Sevan'kaev AV,
Goodhead DT. mFISH analysis reveals complexity of chromosome aberrations
in individuals occupationally exposed to internal plutonium: a pilot study to
assess the relevance of complex aberrations as biomarkers of exposure to high-
LET alpha particles. Radiat Res. 2005; 163(1): 26-35.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
36
63. Pouzoulet F, Roch-Lefèvre S, Giraudet AL, Vaurijoux A, Voisin P, Buard V,
Delbos M, Bourhis J, Voisin P, Roy L. Monitoring translocations by m-FISH
and three-color FISH painting techniques: a study of two radiotherapy patients. J
Radiat Res (Tokyo). 2007; 48(5): 425-434.
64. Szeles A, Joussineau S, Lewensohn R, Lagercrantz S, Larsson C. Evaluation of
spectral karyotyping (SKY) in biodosimetry for the triage situation following
gamma irradiation. Int J Radiat Biol. 2006; 82(2): 87-96.
65. Kuechler A, Neubauer S, Grabenbauer GG, Claussen U, Liehr T, Sauer R,
Wendt TG. Is 24-color FISH detection of in-vitro radiation-induced
chromosomal aberrations suited to determine individual intrinsic
radiosensitivity. Strahlenther Onkol. 2002; 178(4): 209-215.
66. Sumption ND, Goodhead DT, Anderson RM. No increase in radiation-induced
chromosome aberration complexity detected by m-FISH after culture in the
presence of 5'-bromodeoxyuridine. Mutat Res. 2006; 594(1-2): 30-38.
67. Vazquez M, Greulich-Bode KM, Arsuaga J, Cornforth MN, Brückner M, Sachs
RK, Hlatky L, Molls M, Hahnfeldt P. Computer analysis of mFISH chromosome
aberration data uncovers an excess of very complicated metaphases. Int J Radiat
Biol. 2002; 78(12): 1103-1115.
68. Matsui S, Sait S, Jones CA, Nowak N, Gross KW. Rapid localization of
transgenes in mouse chromosomes with a combined Spectral Karyotyping/FISH
technique. Mamm Genome. 2002; 13(12): 680-685.
69. Anderson RM, Stevens DL, Goodhead DT. M-FISH analysis shows that
complex chromosome aberrations induced by alpha -particle tracks are
cumulative products of localized rearrangements. Proc Natl Acad Sci U S A.
2002; 99(19): 12167-12172.
70. Kakazu N, Abe T. Multicolor banding technique, spectral color banding
(SCAN): new development and applications. Cytogenet Genome Res. 2006;
114(3-4): 250-256.
71. Liehr T, Starke H, Weise A, Lehrer H, Claussen U. Multicolor FISH probe sets
and their applications. Histol Histopathol. 2004; 19(1): 229-237.
72. Zitzelsberger H, Bruch J, Smida J, Hieber L, Peddie CM, Bryant PE, Riches AC,
Fung J, Weier HU, Bauchinger M. Clonal chromosomal aberrations in simian

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
37
virus 40-transfected human thyroid cells and in derived tumors developed after
in vitro irradiation. Int J Cancer. 2001; 96(3): 166-177.
73. Pantelias GE, Maillie HD. Direct analysis of radiation-induced chromosome
fragments and rings in unstimulated human peripheral blood lymphocytes by
means of premature chromosome condensation technique. Mutat Res. 1985; 139:
67-72.
74. Prasanna PGS, Escalada ND, Blakely WF. Induction of premature chromosome
condensation by a phosphatase inhibitor and a protein kinase in unstimulated
human peripheral blood lymphocytes: a simple and rapid technique to study
chromosome aberrations using specific whole-chromosome DNA hybridization
probes for biological dosimetry. Mutat. Res. 2000; 466: 131-141.
75. Vral A, Thierens H, Baeyens A, De Ridder L. Induction and disappearance of
G2 chromatid breaks in lymphocytes after low doses of low-LET gamma-rays
and high-LET fast neutrons. Int J Radiat Biol. 2002; 78(4): 249-257.
76. Stoilov LM, Mullenders LH, Darroudi F, Natarajan AT. Adaptive response to
DNA and chromosomal damage induced by X-rays in human blood
lymphocytes. Mutagenesis. 2007; 22(2): 117-122.
77. Suzuki M, Tsuruoka C, Nakano T, Ohno T, Furusawa Y, Okayasu R. The PCC
assay can be used to predict radiosensitivity in biopsy cultures irradiated with
different types of radiation. Oncol Rep. 2006; 16(6): 1293-1299.
78. Wang ZZ, Li WJ, Zhang H, Yang JS, Qiu R, Wang X. Comparison of
clonogenic assay with premature chromosome condensation assay in prediction
of human cell radiosensitivity. World J Gastroenterol. 2006; 12(16): 2601-2605.
79. Pouget JP, Laurent C, Delbos M, Benderitter M, Clairand I, Trompier F,
Stéphanazzi J, Carsin H, Lambert F, Voisin P, Gourmelon P. PCC-FISH in skin
fibroblasts for local dose assessment: biodosimetric analysis of a victim of the
Georgian radiological accident. Radiat Res. 2004; 162(4): 365-376.
80. Lamadrid AI, García O, Delbos M, Voisin P, Roy L. PCC-ring induction in
human lymphocytes exposed to gamma and neutron irradiation. J Radiat Res
(Tokyo). 2007; 48(1): 1-6.
81. Okada M, Saito S, Okayasu R. Facilitated detection of chromosome break and
repair at low levels of ionizing radiation by addition of wortmannin to G1-type
PCC fusion incubation. Mutat Res. 2004; 562(1-2): 11-17.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
38
82. M'kacher R, Violot D, Aubert B, Girinsky T, Dossou J, Béron-Gaillard N, Carde
P, Parmentier C. Premature chromosome condensation associated with
fluorescence in situ hybridisation detects cytogenetic abnormalities after a CT
scan: evaluaton of the low-dose effect. Radiat Prot Dosimetry. 2003; 103(1): 35-
40.
83. Coco-Martin JM, Begg AC. Detection of radiation-induced chromosome
aberrations using fluorescence in situ hybridization in drug-induced premature
chromosome condensations of tumour cell lines with different radiosensitivities.
Int J Radiat Biol. 1997; 71: 265-273.
84. Kanda R, Eguchi-Kasai K, Hayata I. Phosphatase inhibitors and premature
chromosome condensation in human peripheral lymphocytes at different cell
cycle phases. Somat Cell Mol. Genet. 1999; 25: 1-8.
85. Prasanna PGS, Muderhwa JM, Miller AC, Grace MB, Salter CA, Blakely WF.
Diagnostic Biodosimetry Response for Radiation Disasters: Current Research
and Service Activities at AFRRI. 2004; RTO-MP-HFM-108.
86. Vral A, Thierens H, De Ridder L. In vitro micronucleus-centromere assay to
detect radiation-damage induced by low doses in human lymphocytes. Int J
Radiat Biol. 1997; 71: 61-68.
87. Thierens H, Vral A, Barbe M, Aousalah B, De Ridder L. A cytogenetic study of
nuclear power plant workers using the micronucleus-centromere assay. Mutat
Res.1999; 445: 105-111.
88. Cytogenetic analysis for radiation dose assessment. Technical Report Series:
405, IAEA Vienna. 2001.
89. Hatayoglu SE, Orta T. Relationship between radiation-induced dicentric
chromosome aberrations and micronucleus formation in human lymphocytes. J
Exp Clin Cancer Res. 2007; 26(2): 229-234.
90. Nakamura N, Miazawa C, Sawada S, Akiyama M, Awa AA. A close correlation
between electron spin resonance (ESR) dosimetry from tooth enamel and
cytogenetic dosimetry from lymphocytes of Hiroshima atomic bomb survivors.
Int J Radiat Biol. 1998; 73: 619-627.
91. Prossor JS, Moquet JE, Lloyd DC, Edwards AA. Radiation induction of
micronuclei in human lymphocytes. Mutat Res. 1988; 199: 37-45.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
39
92. Sreedevi B, Rao BS. Assay of micronuclei in peripheral blood lymphocytes as a
biological indicator of radiation dose. Radiat Prot Dosimetry.1994; 51: 41-45.
93. Le Roux J, Slabbert J, Smit B, Blekkenhorst G. Assessment of the micronucleus
assay as a biological dosimeter using cytokinesis-blocked lymphocytes from
cancer patients receiving fractionated partial body-radiotherapy. Strahlenther
Onkol. 1998; 174(2): 75-81.
94. Song EY, Rizvi SM, Qu CF, Raja C, Yuen J, Li Y, Morgenstern A, Apostolidis
C, Allen BJ. The cytokinesis-block micronucleus assay as a biological dosimeter
for targeted alpha therapy. Phys Med Biol. 2008; 53(2): 319-328.
95. Decordier I, Kirsch-Volders M. The in vitro micronucleus test: from past to
future. Mutat Res. 2006; 607(1): 2-4.
96. Wagner ED, Anderson D, Dhawan A, Rayburn AL, Plewa MJ. Evaluation of
EMS-induced DNA damage in the single cell gel electrophoresis Comet assay
and with flow cytometric analysis of micronuclei. Teratog Carcinog Mutagen.
2003; 23(S2): 1-11.
97. Bryce SM, Bemis JC, Avlasevich SL, Dertinger SD. In vitro micronucleus assay
scored by flow cytometry provides a comprehensive evaluation of cytogenetic
damage and cytotoxicity. Mutat Res. 2007; 630(1-2): 78-91.
98. Olive PL. DNA damage and repair in individual cells: applications of the comet
assay in radiobiology. Int J Radiat Biol 1999; 75(4): 395-405.
99. Kopjar N, Garaj-Vrhovac V. Assessment of DNA damage in nuclear medicine
personnel – comparative study with the alkaline comet assay and the
chromosome aberration test. Int J Hyg Environ Health. 2005; 208: 179-191
100. Garaj-Vrhovac V, Kopjar N, Raem D, Veki B, Miljani S, Ranogajec-Komor M.
Application of the alkaline comet assay in biodosimetry: assessment of in vivo
DNA damage in human peripheral leukocytes after a gamma radiation incident.
Radiat Prot Dosimetry. 2002; 98: 407-415.
101. Shahidi M, Mozdarani H, Bryant PE. Radiation sensitivity of leukocytes from
healthy individuals and breast cancer patients as measured by the alkaline and
neutral comet assay. Cancer Lett. 2007; 257(2): 263-273.
102. McKeown SR, Robson T, Price ME, Ho ETS, Hirst DG, McKelvey-Martin VJ.
Potential use of the alkaline comet assay as a predictor of bladder tumour
response to radiation. Br J Cancer. 2003; 89: 2264-2270.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
40
103. Gamulin M, Garaj-Vrhovac V, Kopjar N. Evaluation of DNA damage in
radiotherapy-treated cancer patients using the alkaline comet assay. Coll
Antropol. 2007; 31(3): 837-844.
104. Ravi M, Solomon Paul FD. A rapid biodosimetric technique at the human
Glycophorin-A locus. Int J Hum Genet. 2002; 2(4): 251-254.
105. Kyoizumi S, Kusunoki Y, Hayashi T. Flow cytometric measurement of mutant T
cells with altered expression of TCR: detecting somatic mutations in humans and
mice. Methods Mol Biol. 2005; 291:197-204.
106. Akiyama M, Kyoizumi S, Hirai Y, Hakoda M, Nakamura N, Awa AA. Studies
on chromosome aberrations and HPRT mutations in lymphocytes and GPA
mutation in erythrocytes of atomic bomb survivors. Prog Clin Biol Res. 1990;
340C: 69-80.
107. Kleinerman RA, Romanyukha AA, Schauer DA, Tucker JD. Retrospective
assessment of radiation exposure using biological dosimetry: chromosome
painting, electron paramagnetic resonance and the glycophorin A mutation
assay. Radiat Res. 2006; 166(1 Pt 2): 287-302.
108. Lindholm C, Murphy BP, Bigbee WL, Bersimbaev RI, Hultén MA, Dubrova
YE, Salomaa S. Glycophorin A somatic cell mutations in a population living in
the proximity of the Semipalatinsk nuclear test site. Radiat Res. 2004; 162(2):
164-170.
109. Tawn JE, Whitehouse CA, Paul Daniel C, Tarone RE, Bothwell AM, Fisher A.
Somatic cell mutations at the glycophorin A locus in erythrocytes of radiation
workers from the Sellafield nuclear facility. Radiat Res. 2003; 159(1): 117-122.
110. Jones IM, Tucker JD, Langlois RG, Mendelsohn ML, Pleshanov P, Nelson DO.
Evaluation of three somatic genetic biomarkers as indicators of low dose
radiation effects in clean-up workers of the Chernobyl nuclear reactor accident.
Radiat Prot Dosimetry. 2001; 97(1): 61-7.
111. Jones IM, Galick H, Kato P, Langlois RG, Mendelsohn ML, Murphy GA,
Pleshanov P, Ramsey MJ, Thomas CB, Tucker JD, Tureva L, Vorobtsova I,
Nelson DO. Three somatic genetic biomarkers and covariates in radiation-
exposed Russian cleanup workers of the chernobyl nuclear reactor 6-13 years
after exposure. Radiat Res. 2002; 158(4): 424-442.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
41
112. Mendelsohn ML, Peeters JP, Normandy MJ. Biomarkers and occupational
health: Progress and perspectives. 1995; 9-19.
113. Taooka Y, Takeichi N, Noso Y, Kawano N, Apsalikov KN, Hoshi M. Increased
T-cell receptor mutation frequency in radiation-exposed residents living near the
Semipalatinsk nuclear test site. J Radiat Res (Tokyo). 2006; 47(A): A179-A181.
114. Smirnova SG, Zamulaeva IA, Orlova NV, Saenko AS, Sevan'kaev AV, Gasteva
GN, Nugis VIu, Molokanov AA. Comparative investigation of structural and
gene somatic mutations in workers of nuclear chemical plants. II. Frequency of
lymphocytes mutant in T-cell receptor loci. Radiats Biol Radioecol. 2005;
45(2):162-167.
115. Saenko AS, Zamulaeva IA, Smirnova SG, Orlova NV, Selivanova EI, Nugis VI,
Nadezhina NM. Determination of mutation frequency at loci of glycophorin A
and T-cell receptors: informativeness for biological dosimetry of acute and
prolonged irradiation. Radiats Biol Radioecol. 1998; 38(2):171-180.
116. Kyoizumi S, Akiyama M, Hirai Y, Kusunoki Y, Tanabe K. Spontaneous loss
and alteration of antigen receptor expression in mature CD4+ T cells. J Exp Med.
1990; 171: 1981-1999.
117. Ishioka N, Umeki, Hirai S, Akiyama Y, Kodama M, Ohama TK, Kyoizumi S.
Stimulated rapid expression in vitro for early detection of in vivo T-cell receptor
mutations induced by radiation exposure. Mutat Res. 1997; 390(3): 269-82.
118. Hakoda M, Akiyama M, Kyoizumi S, Awa AA, Yamakido M, Otake M.
Increased somatic cell mutant frequency in atomic bomb survivors. Mutat
Res.1988; 201: 39-48.
119. Messing K, Bradley WE. In vivo mutant frequency rises among breast cancer
patients after exposure to high doses of γ- radiation. Mutat Res.1985; 152: 107-
112.
120. Albertini RJ, Clark LS, Nicklas JA, O'Neill JP, Hui TE, Jostes R. Radiation
quality affects the efficiency of induction and the molecular spectrum of HPRT
mutations in human T cells. Radiat Res. 1997; 148(5): S76-S86.
121. Thomas CB, Nelson DO, Pleshanov P, Jones IM. Induction and decline of HPRT
mutants and deletions following a low dose radiation exposure at Chernobyl.
Mutat Res. 2002; 499(2): 177-187.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
42
122. Kumar PR, Mohankumar MN, Hamza VZ, Jeevanram RK. Dose-rate effect on
the induction of HPRT mutants in human G0 lymphocytes exposed in vitro to
gamma radiation. Radiat Res. 2006; 165(1): 43-50.
123. Herrlich P, Ponta H, Rahmsdorf HJ. DNA damage induced gene expression:
signal transduction and relation to growth factor signaling. Rev Physiol Biochem
Pharm. 1992; 119: 187–223.
124. Holbrook NJ, Liu Y, Fornace AJ. Signaling events controlling the molecular
response to genotoxic stress. EXS. 1996; 77: 273-88.
125. Miller G, Fuchs R, Lai E. Image cDNA clones, UniGene clustering and ACeDB:
an integrated resource for expressed sequence information. Genome Res. 1997;
7: 1027–1032.
126. Schena M, Shalon D, Heller R, Chai A, Brown PO, Davis RW. Parallel human
genome analysis: microarray-based expression monitoring of 1000 genes. Proc
Natl Acad Sci. USA 1996; 93: 10614–10619.
127. Amundson SA, Fornace AJ Jr. Gene expression profiles for monitoring radiation
exposure. Radiat Prot Dosimetry. 2001; 97(1): 11-16.
128. Ding LH, Shingyoji M, Chen F, Hwang JJ, Burma S, Lee C, Cheng JF, Chen DJ.
Gene expression profiles of normal human fibroblasts after exposure to ionizing
radiation: a comparative study of low and high doses. Radiat Res. 2005; 164(1):
17-26.
129. Snyder AR, Morgan WF. Gene expression profiling after irradiation: clues to
understanding acute and persistent responses. Cancer Metastasis Rev. 2004;
23(3-4): 259-68.
130. Azzam EI, de Toledo SM, Little JB. Expression of Connexin43 is highly
sensitive to ionizing radiation and other environmental stresses. Cancer Res.
2003; 63(21): 7128-7135.
131. Goldberg Z, Rocke DM, Schwietert C, Berglund SR, Santana A, Jones A,
Lehmann J, Stern R, Lu R, Hartmann Siantar C. Human in vivo dose-response to
controlled, low-dose low linear energy transfer ionizing radiation exposure. Clin
Cancer Res. 2006; 12(12): 3723-3729.
132. Durante M. Potential applications of biomarkers of radiation exposure in nuclear
terrorism events. Phisica Medica. 2003; 19(3): 191-212.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
43
133. Fenech M, Morley AA. Measurement of micronuclei in lymphocytes. Mutat Res.
1985; 147(1-2): 29-36.
134. Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM. DNA Double-
stranded Breaks Induce Histone h2ax Phosphorylation on Serine 139. J Biol
Chem. 1998; 273(10): 5858-5868.
135. MacPhail SH, Banath JP, Yu TY, Chu EH, Lambur H, Olive PL. Expression of
phosphorylated histone h2ax in cultured cell lines following exposure to X-rays.
Int J Radiat Biol. 2003; 79: 351–358.
136. Rogakou EP, Boon C, Redon C, Bonner WM. Megabase chromatin domains
involved in DNA double-strand breaks in vivo. J Cell Biol. 1999; 146: 905-916.
137. Rothkamm K, Lobrich M. Evidence for a lack of DNA double-strand break
repair in human cells exposed to very low x-ray doses. Proc Natl Acad Sci USA.
2003; 100: 5057–5062.
138. Pilch DR, Sedelnikova OA, Redon C, Celeste A, Nussenzweig A, Bonner WM.
Characteristics of gamma-h2ax foci at DNA double-strand breaks sites. Biochem
Cell Biol. 2003; 81: 123–129.
139. Banath JP, Macphail SH, Olive PL. Radiation sensitivity, h2ax phosphorylation,
and kinetics of repair of DNA strand breaks in irradiated cervical cancer cell
lines. Cancer Research. 2004; 64: 7144-7149.
140. Da Cruz AD, McArthur AG, Silva CC, Curado MP, Glickman BW. Human
micronucleus counts are correlated with age, smoking and cesium-137 dose in
the Goiânia (Brazil) radiological accident. Mutat Res. 1994; 313: 57-68.
141. Sedelnikova OA, Rogakou EP, Panyutin IG, Bonner WM. Quantitative detection
of I-125 induced DNA doublestrand breaks with gamma-h2ax antibody. Radiat
Res. 2002; 158: 486–492.
142. Olive PL. Detection of DNA damage in individual cells by analysis of histone
h2ax phosphorylation. Methods Cell Biol. 2004; 75: 355–373.
143. Olive PL, Banath JP. Phosphorylation of histone h2ax as a measure of
radiosensitivity. Int J Radiat Oncol Biol Phys. 2004; 58: 331–335.
144. Banath J.P, Olive PL. Expression of phosphorylated histone h2ax as a surrogate
of cell killing by drugs that create DNA double-strand breaks. Cancer Res. 2003;
63: 4347–4350.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
44
145. Brady JM, Aarestad NO, Swartz HM. In vivo dosimetry by electron spin
resonance spectroscopy. Med Phys. 19681; 5: 43–47.
146. Driessens FCM. The mineral in bone, dentine and tooth enamel. Bull Soc Chem
Belg. 1980; 89: 663–689.
147. Moens P, De Volder P, Hoogewijs R, Callens F, Verbeeck R. Maximum-
likelihood common-factor analysis as a powerful tool in decomposing
multicomponent EPR powder spectra. J Magn Reson. 1993; 101: 1–15.
148. Ikeya M, Miyajima J, Okajima S. ESR dosimetry for atomic bomb survivors
using shell buttons and tooth enamel. Jpn J Appl Phys. 1984; 23: 697-699.
149. Nakamura N, Miyazawa C, Akiyama M, Sawada S, Awa AA. A close
correlation between electron spin resonance (ESR) dosimetry from tooth enamel
and cytogenetic dosimetry from lymphocytes of Hiroshima atomic-bomb
survivors. Int J Radiat Biol. 1998; 73: 619–627.
150. Romanyukha AA, Ignatiev EA, Vasilenko EK, Drozhko EG, Wieser A, Jacob P,
Keriim-Markus IB, Kleschenko ED, Nakamura N, Miyazawa C. EPR dose
reconstruction for Russian nuclear workers. Health Phys. 2000; 78(1): 15-20.
151. Wieser A, Romanyukha A, Degteva M, Kozheurov V, Petzoldt G. Tooth enamel
as a natural beta dosimeter for bone seeking radionuclides. Radiat Prot
Dosimetry. 1996; 65: 413–416.
152. Romanyukha AA, Degteva MO, Kozheurov VP, Wieser A, Ignatiev EA,
Vorobiova MI, Jacob P. Pilot study of the population of the Ural Region by EPR
tooth dosimetry. Radiat Envir Biophys. 1996; 35: 305-310.
153. Chumak V, Sholom S, Pasalkaya L. Application of high precision EPR
dosimetry with teeth for reconstruction of doses to Chernobyl populations.
Radiat Prot Dosimetry. 1999; 84: 515-520.
154. Chumak VV, Sholom SV, Likhtarev IA. Some results of retrospective dose
reconstruction for selected groups of exposed population in Ukraine. In: The
Radiological Consequences of the Chernobyl Accident. Report EUR 16544 EN.
1996; 1059-1062.
155. Ishii H, Ikeya M, Okano M. ESR dosimetry of teeth of residents close to
Chernobyl reactor accident. J Nucl Sci Technol.1990; 27: 1153-1155.
156. Tatsumi-Miyajima J, Okajima S. ESR dosimetry using human tooth enamel. In:
ESR dating and dosimetry (Ikeya M, Miki T), Tokyo. 1985.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
45
157. Sevan’kaev AV, Lloyd DC, Edwards AA, Moiseenko VV. High exposures to
radiation received by workers inside the Chernobyl Sargophagus. Radiat Prot
Dosimetry. 1995; 59: 85-91.
158. Ivannikov AI, Skvortsov VG, Stepanenko VF, Tikunov DD, Fedosov IM,
Romanyukha AA, Wieser A. Wide scale EPR retrospective dosimetry. Results
and problems. Radiat Prot Dosimetry. 1997; 71: 175-180.
159. Bailiff IK, Stepanenko BV. Final report, Experimental Collaboration Project
ECP10. Retrospective dosimetry and dose reconstruction. 1996; European
Commission. EUR 165540.
160. Chumak V, Likhtarev I, Sholom S, Meckbach R, Krjuchkov V. Chernobyl
experience in field of retrospective dosimetry: reconstruction of doses to the
population and liquidators involved in the accident. Radiat Prot Dos. 1998; 77:
91-95.
161. Kvortsov V, Stepanenko V, Vaher Ü. International intercomparison of dose
measurements using EPR spectrometry of tooth enamel. In: The Radiological
Consequences of the Chernobyl Accident. Report EUR 16544 EN, European
Commission, Brussels. 1996; 957-964.
162. Wieser A, Chumak, Bailiff I, Baran N, Bougai A, Brik A, Dubrovsky S, Finin V,
Haskell E, Hayes R, Ivannikov A, Kenner G, Kirillov V, Kolesnik S, Liidj G,
Lippmaa E, Maksimenko V, Matyash M, Meijer A, Minenko V, Pasalskaya L,
Past J, Pavlenko J, Puskar J, Radchuk V, Scherbina O, Sholom S.
163. Chumak, V, Bailiff I, Baran N, Bugai A, Dubovsky S, Fedosov I, Finin V,
Haskell E, Hayes R, Ivannikov A, Kenner G, Kirillov V, Khamidova L,
Kolesnik S, Liidja G, Likhtarev I, Lippmaa E, Maksimenko V, Meijer A,
Minenko V, Pasalskaya L, Past J, Puskar J, Radchuk V, Sholom S, Skvortzov V,
Stepanenko V, Vaher U, Wieser A. The first international intercomparison of
EPR-dosimetry with teeth: first results. Appl Radiat Isot. 1996; 47: 1281-1286.
164. Wieser A, Romanyukha A, Onori S, Vasilenko E, Bayankin S, Fattibene P,
Ignatiev E, Knyazev V, Jacob P. External exposures of nuclear workers at
Mayak: Comparison of occupational film dosimetry and EPR measurements
with tooth enamel. Health Phys. 2002.

Evaluation of ionizing radiation exposure for biodosimetry: Validation of Whole Chromosome
Painting by Cytogenetic and Molecular techniques.
46
165. Vanhaelewyn G, Amira S, Debuyst R, Callens F, Glorieuxleloup G, Thierens
HA. Critical discussion of the 2nd intercomparison on electron paramagnetic
resonance dosimetry with tooth enamel. Radiat Meas. 2001; 33: 417-426.
166. Romanyukha AA, Nagy V, Sleptchonok O, Desrosiers MF, Jiang J, Heiss A.
Individual biodosimetry at the natural radiation background level. Health Phys.
2001; 80: 71-73.
167. Pellmar TC, Rockwell S. Priority list of research areas for radiological nuclear
threat countermeasures. Radiat Res. 2005; 163: 115-123.
168. Hayata I, Tabuchi H, Furukawa A, Okabe N, Yamamoto M, Sato K. Robot
system for preparing lymphocyte chromosome. J Radiat Res (Tokyo). 1992; 33:
231-241.