chapter 10 bacterial genetics. mutation and recombination, mutations and mutants
Post on 22-Dec-2015
227 views
TRANSCRIPT

CHAPTER 10Bacterial Genetics

Mutation and Recombination, Mutations and Mutants

• Mutation is a heritable change in DNA sequence that can lead to a change in phenotype.
•By definition, a mutant differs from its parental strain in genotype, the nucleotide sequence of the genome.
•hisC (1,2,3,…) mutants of E. coli
•Auxotroph – nutritional mutant rewiring a growth factors, amino acids (e.g. His-).
•Prototroph – wild type

• Selectable mutations are those that give the mutant a growth advantage under certain environmental conditions and are especially useful in genetic research.
•If selection is not possible, mutants must be identified by screening.

• Although screening is always more tedious than selection, methods are available for screening large numbers of colonies in certain types of mutations.
•Penicillin selection – pen kills growing but not mutant (non growing) cells in minimal medium
Nutritionally defective mutants can also be detected by the technique of replica plating (Figure 10.2).

Replica Plating

Molecular Basis of Mutation
• Mutations, which can be either spontaneous or induced, arise because of changes in the base sequence of the nucleic acid of an organism's genome.

• A point mutation, which results from a change in a single base pair, can lead to a single amino acid change in a polypeptide or to no change at all, depending on the particular codon involved (Figure 10.3).

Possible effects of bp substitution

• In a nonsense mutation, the codon becomes a stop codon and an incomplete polypeptide is made.
•In a missense mutation, the sequence of amino acids in the ensuing polypeptide is changed, resulting in an inactive protein or one with reduced activity.
•Temperature-sensitive mutations/
conditionally lethal mutations

• Deletions and insertions cause more dramatic changes in the DNA, including frameshift mutations, and often result in complete loss of gene function (Figure 10.4).


Table 10.1 lists various kinds of mutants.

Mutation Rates
• Different types of mutations can occur at different frequencies. For a typical bacterium, mutation rates of 10–7 to 10–11 per base pair are generally seen.

• Although RNA and DNA polymerases make errors at about the same rate, RNA genomes typically accumulate mutations at much higher frequencies than DNA genomes.

Mutagenesis
• Mutagens are chemical, physical, or biological agents that increase the mutation rate. Mutagens can alter DNA in many different ways, but such alterations are not mutations unless they can be inherited.

•Table 10.2 gives an overview of some of the major chemical and physical mutagens and their modes of action.

• There are several classes of chemical mutagens, one being the nucleotide base analogs (Figure 10.5).


• Several forms of radiation are highly mutagenic (Figure 10.6).


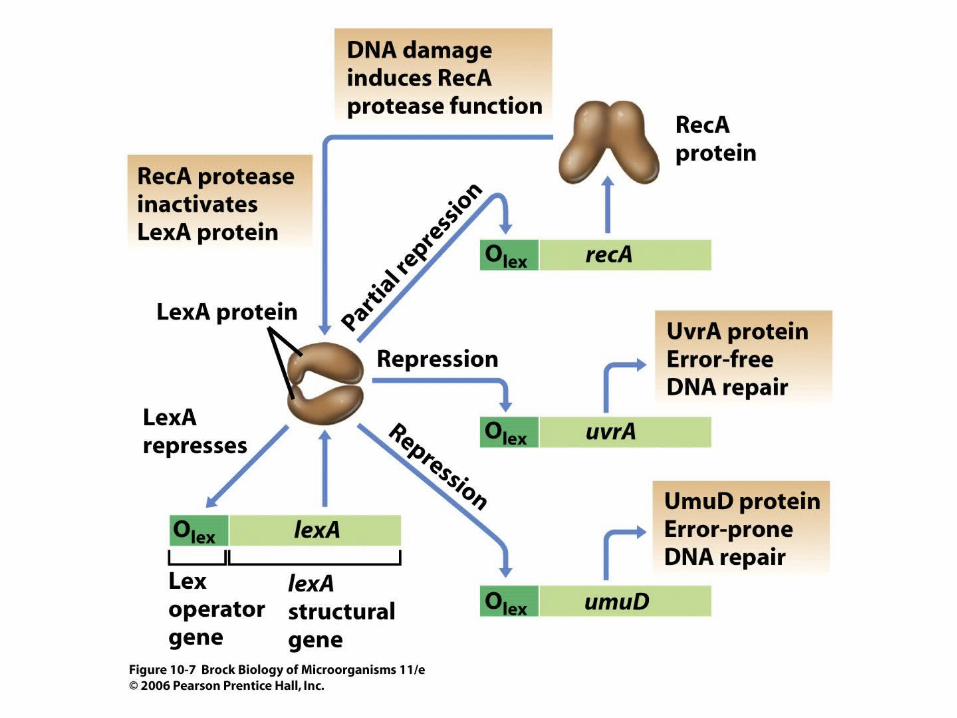
• Some DNA damage can lead to cell death if not repaired. A complex cellular mechanism called the SOS regulatory system is activated as a result of some types of DNA damage and initiates a number of DNA repair processes, both error-prone and high-fidelity (Figure 10.7).

Mutagenesis and Carcinogenesis: The Ames Test

• The Ames test employs a sensitive bacterial assay system for detecting chemical mutagens in the environment.

Genetic Recombination
• Homologous recombination arises when closely related DNA sequences from two distinct genetic elements are combined in a single element (Figure 10.9).


• Recombination is an important evolutionary process, and cells have specific mechanisms for ensuring that recombination takes place.

• Mechanisms of recombination that occur in prokaryotes involve DNA transfer during the processes of transformation, transduction, and conjugation (Figure 10.11).


Genetic Exchange in Prokaryotes
Transformation

• The discovery of transformation was one of the seminal events in biology because it led to experiments demonstrating that DNA is the genetic material (Figure 10.13).


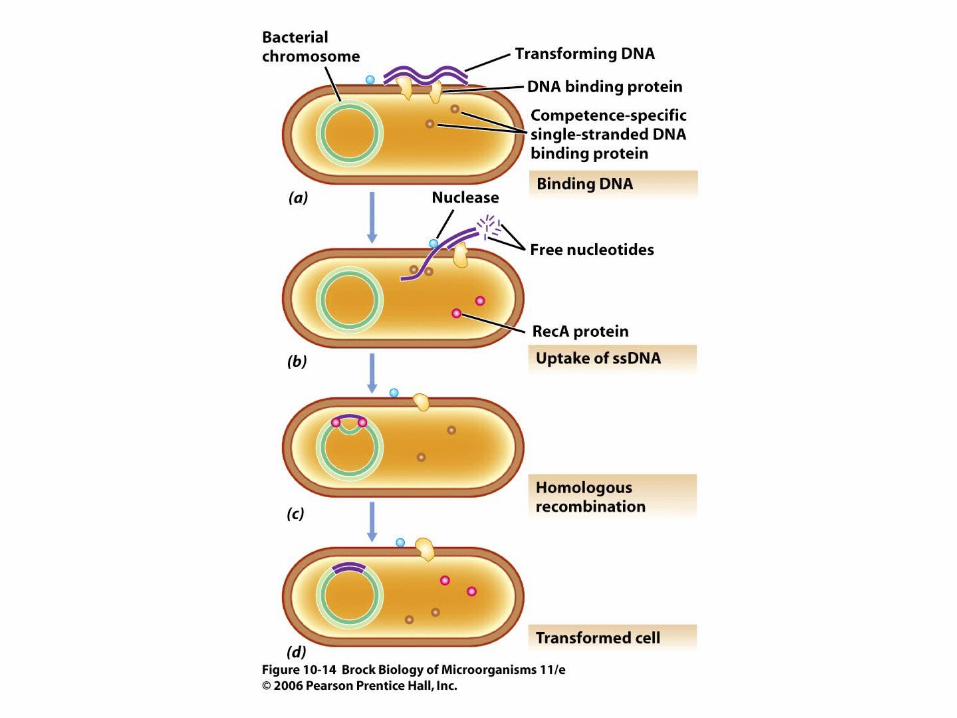
• Certain prokaryotes exhibit competence, a state in which cells are able to take up free DNA released by other bacteria.

• Incorporation of donor DNA into a recipient cell requires the activity of single-stranded binding protein, RecA protein, and several other enzymes. Only competent cells are transformable (Figure 10.14).

Transduction
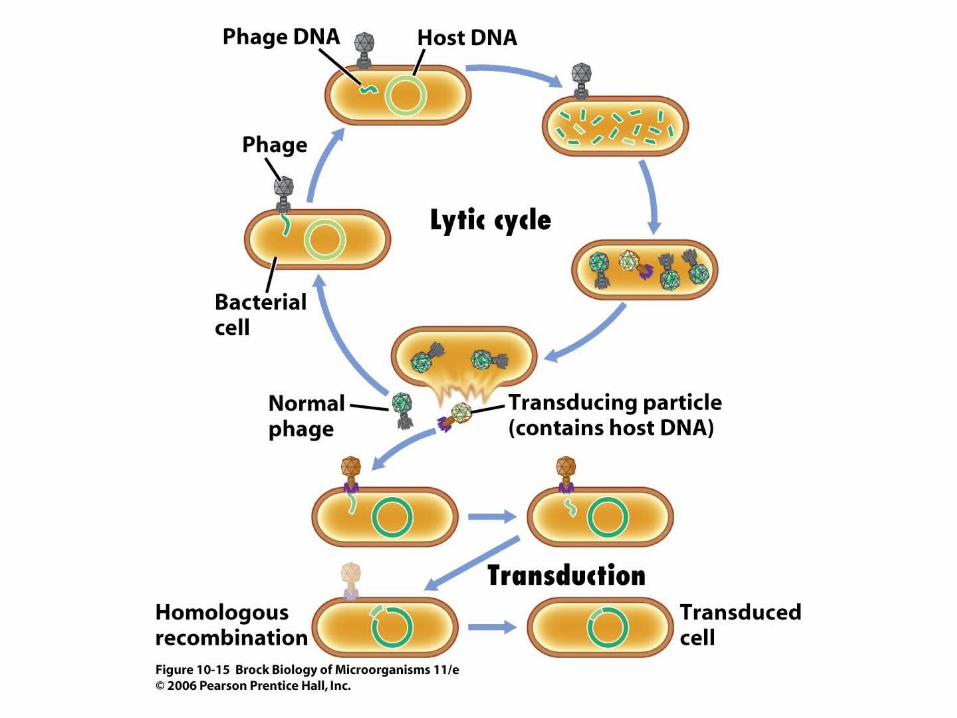
• Transduction involves the transfer of host genes from one bacterium to another by bacterial viruses.

• In generalized transduction (Figure 10.15), defective virus particles incorporate fragments of the cell's chromosomal DNA randomly, but the efficiency is low.

• In specialized transduction (Figure 10.16), the DNA of a temperate virus excises incorrectly and takes adjacent host genes along with it; transducing efficiency in this case may be very high.


Plasmids: General Principles
• Plasmids are small circular or linear DNA molecules that carry any of a variety of unessential genes. Although a cell can contain more than one plasmid, they cannot be closely related genetically.

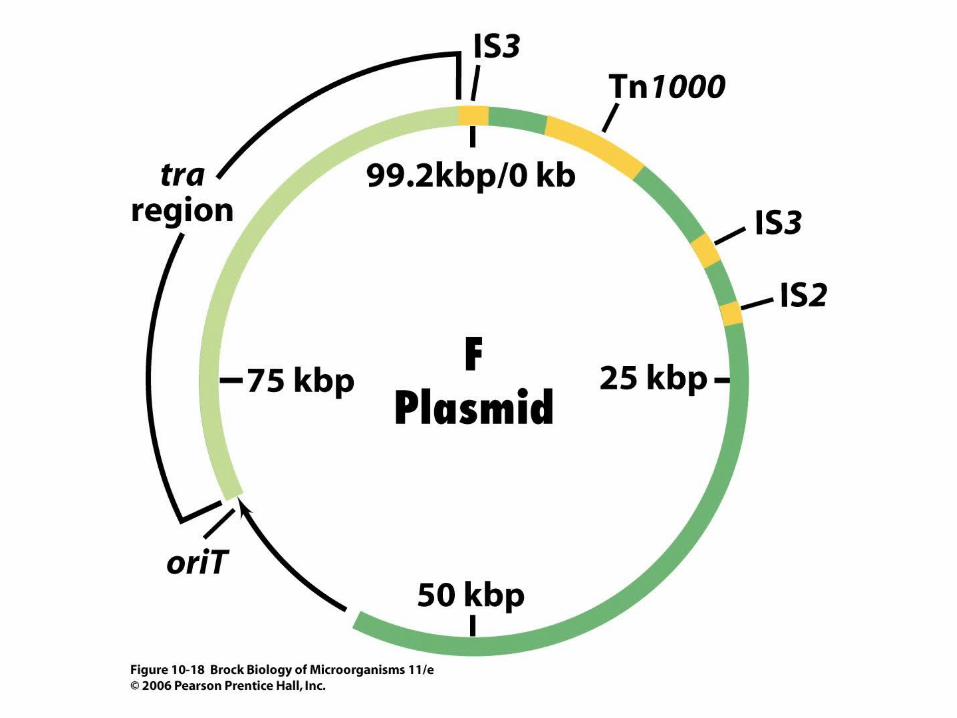
• Figure 10.18 shows a genetic map of the F (fertility) plasmid, a very well characterized plasmid of Escherichia coli.

• Lateral transfer in the process of conjugation can transfer plasmids (Figure 10.19).


Types of Plasmids and Their Biological Significance

• The genetic information that plasmids carry is not essential for cell function under all conditions but may confer a selective growth advantage under certain conditions.

• Examples include antibiotic resistance (Figure 10.20), enzymes for degradation of unusual organic compounds, and special metabolic pathways. Virulence factors of many pathogenic bacteria are often plasmid-encoded.
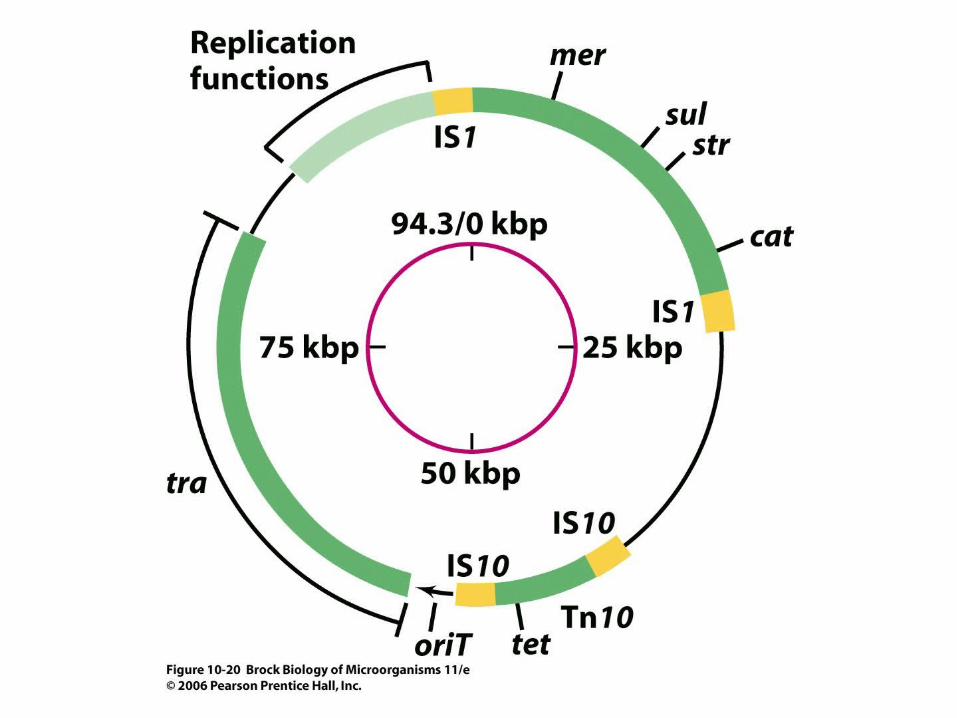

• Table 10.3 lists some phenotypes that plasmids confer on prokaryotes.


Conjugation: Essential Features
• Conjugation is a mechanism of DNA transfer in prokaryotes that requires cell-to-cell contact.

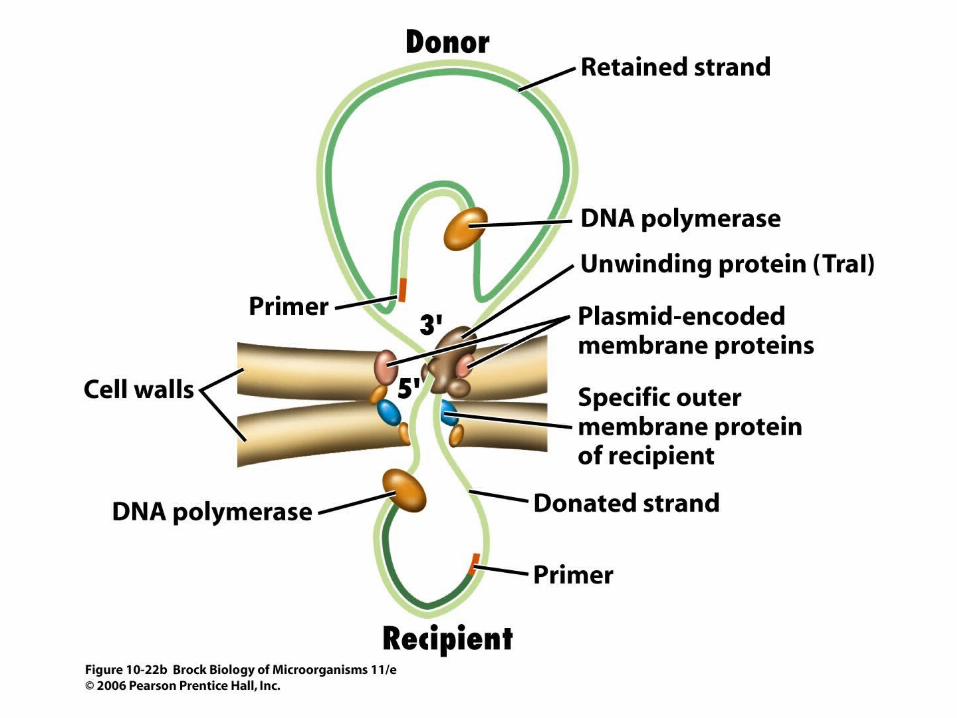
• Genes carried by certain plasmids (such as the F plasmid) control conjugation, and the process involves transfer of the plasmid from a donor cell to a recipient cell (Figure 10.22). Plasmid DNA transfer involves replication via the rolling circle mechanism.


10.12 The Formation of Hfr Strains and Chromosome
Mobilization, p. 279

• The donor cell chromosome can be mobilized for transfer to a recipient cell. This requires that the F plasmid integrate into the chromosome to form the Hfr phenotype. Transfer of the host chromosome is rarely complete but can be used to map the order of the genes on the chromosome.

• F' plasmids are previously integrated F plasmids that have deintegrated and excised some chromosomal genes.

• Integration of the F plasmid into the host chromosome can occur at several specific sites, called IS (for insertion sequence) sites (Figure 10.23). These sites are regions of DNA sequence homology between chromosomal and F plasmid DNA.


Complementation
• If a cell is treated so that it contains two copies of a region of its genome, complementation tests can determine if two mutations are in the same or different genes (Figure 10.28).
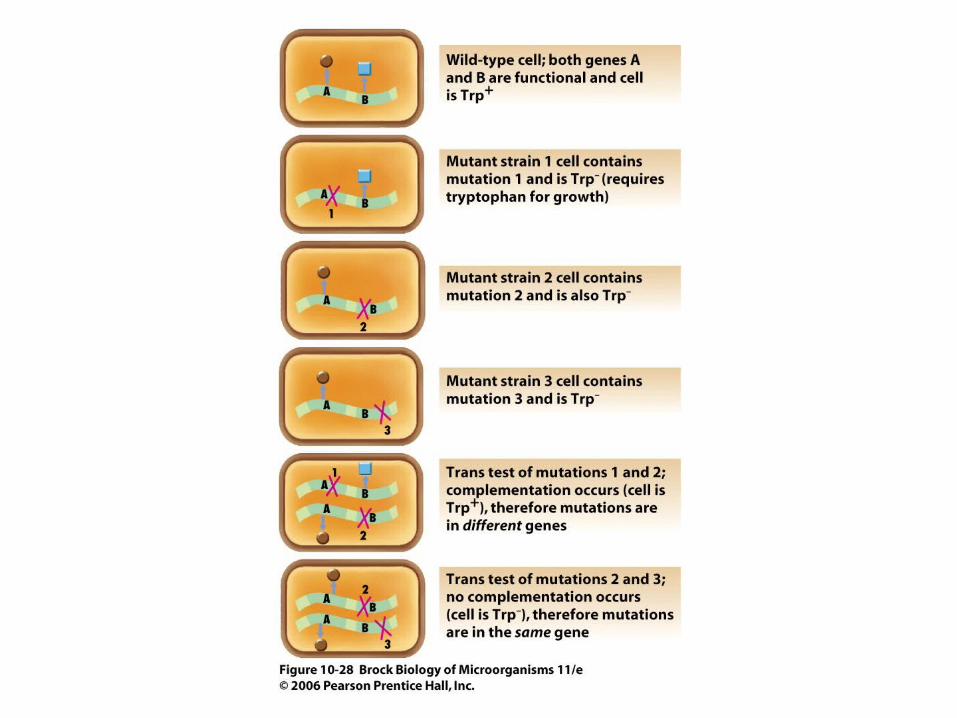

• This is often necessary because mutations in different genes in the same pathway may give the same phenotype. Complementation tests do not involve recombination.

Transposons and Insertion Sequences
• Transposons and insertion sequences are genetic elements that can move from one location on a chromosome to another by a process called transposition, a type of site-specific recombination (Figure 10.30).


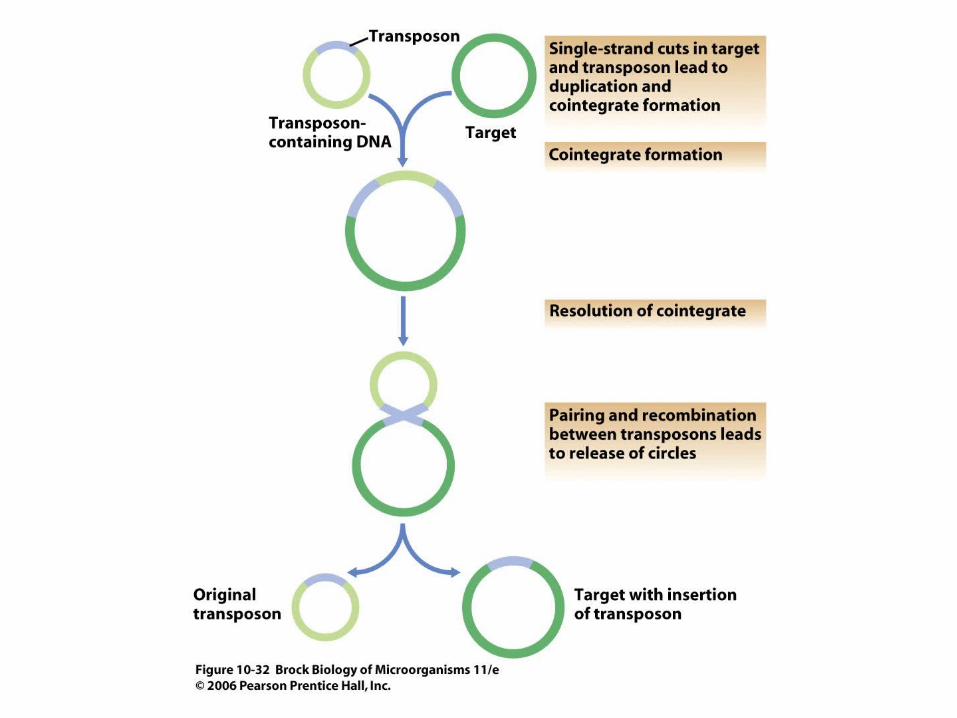
• Transposition is linked to the presence of special genetic elements called transposable elements. Transposition can be either replicative or conservative (Figures 10.31, 10.32).


• Transposons often carry genes encoding antibiotic resistance, and they can be used as biological mutagens (Figure 10.33).


Bacterial Genetics and Gene Cloning
Essentials of Molecular Cloning

• A plasmid or virus is used as the cloning vector to isolate a specific gene or region of a chromosome by molecular cloning (Figure 10.35).
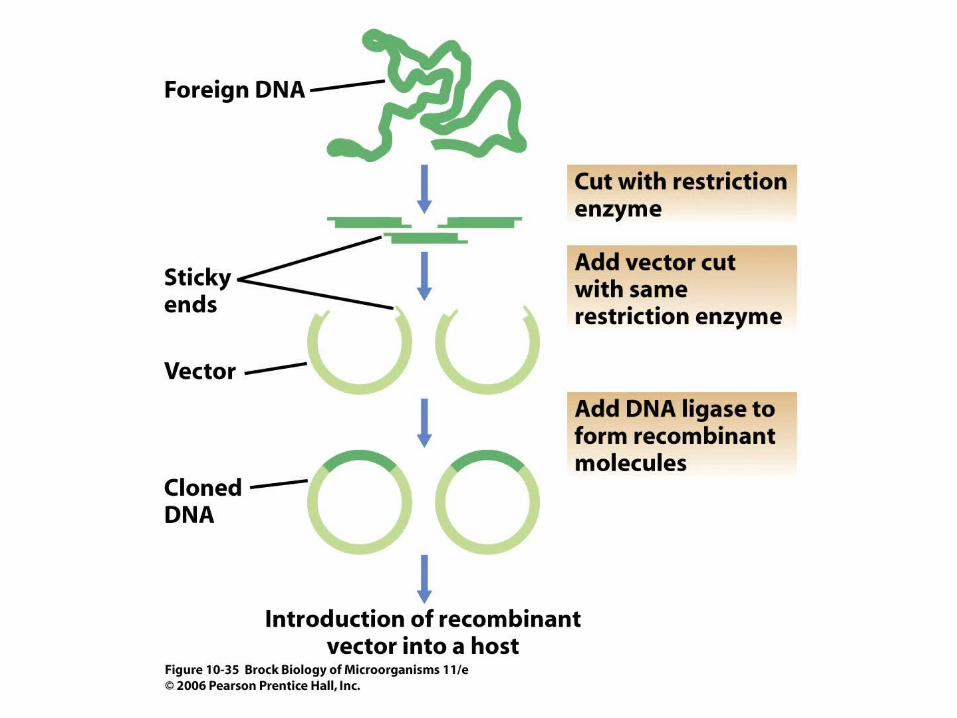

• An in vitro recombination procedure uses restriction enzymes and DNA ligase to produce the hybrid DNA molecule. Once introduced into a suitable host, the cloning vector can control production of large amounts of the target DNA.

• Making a gene library by cloning random fragments of a genome is called shotgun cloning, and it is a widely practiced technique in gene cloning and genomic analyses.

Plasmids as Cloning Vectors
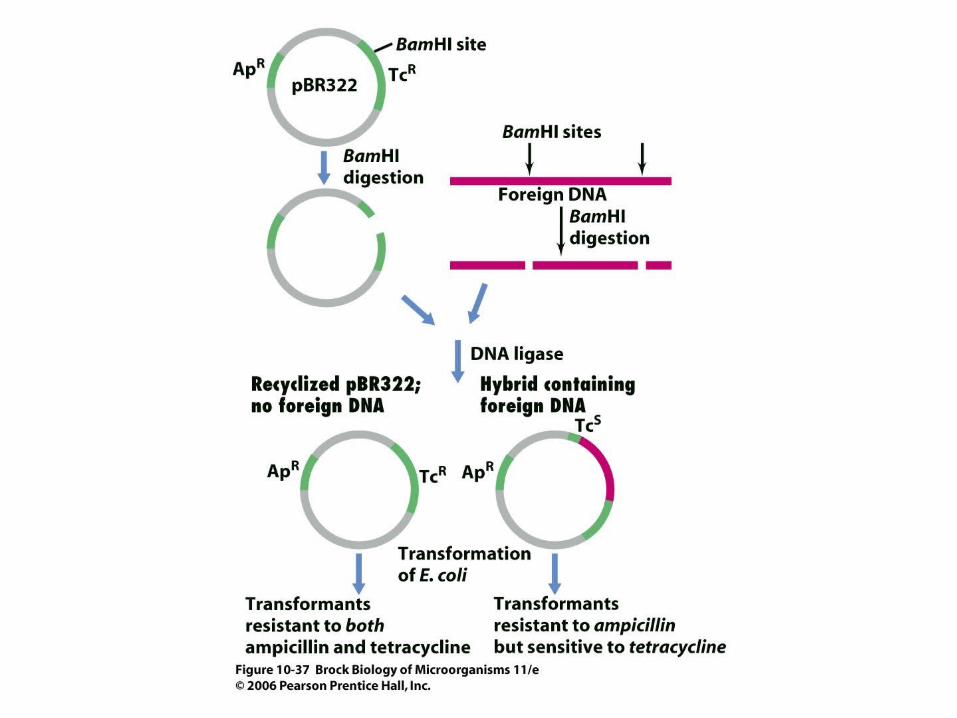
• Plasmids are useful cloning vectors (Figure 10.36) because they are easy to isolate and purify and can multiply to high copy numbers in bacterial cells.


• Antibiotic resistance genes of the plasmid are used to identify bacterial cells containing the plasmid (Figure 10.37).

Bacteriophage Lambda as a Cloning Vector
• Bacteriophages such as lambda have been modified to make useful cloning vectors (Figures 10.38, 10.39).

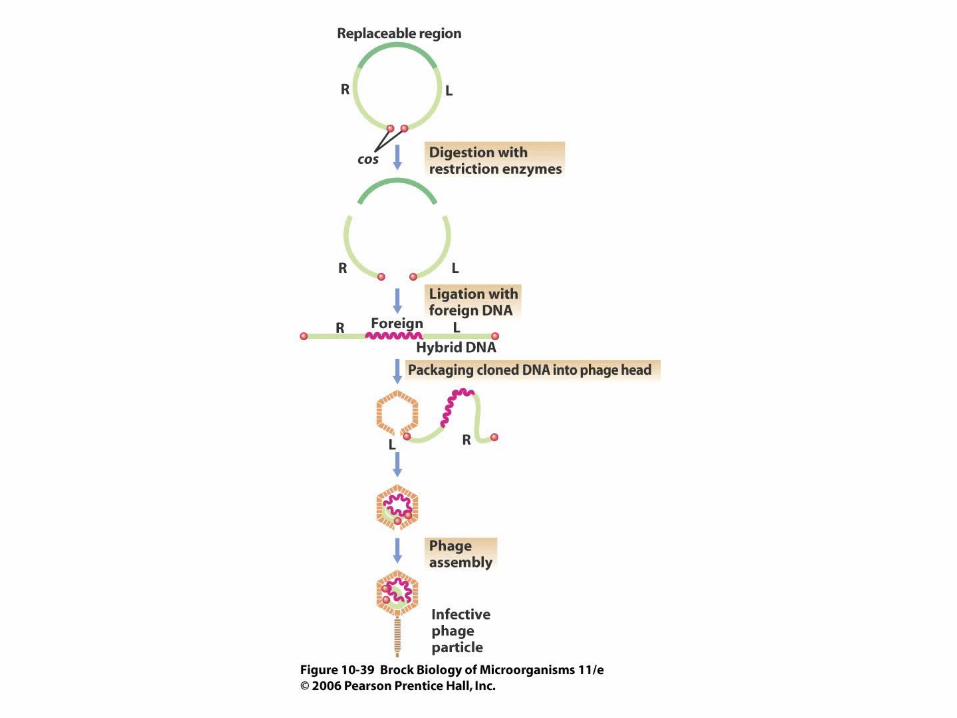

• Larger amounts of foreign DNA can be cloned with lambda than with many other plasmids. In addition, the recombinant DNA can be packaged in vitro for efficient transfer to a host cell. Plasmid vectors containing the lambda cos sites are called cosmids, and they can carry a large fragment of foreign DNA.

In Vitro and Site-Directed Mutagenesis
• Site-directed mutagenesis allows synthetic DNA molecules of desired sequence to be made in vitro and used to construct a mutated gene directly or to change specific base pairs within a gene (Figure 10.40).


• Inserting DNA fragments, called cassettes, into genes can also cause gene disruption (Figure 10.41). The inserted cassette eliminates the function of the wild-type gene while conferring a new, and usually selectable, phenotype on the cell.


The Bacterial Chromosome Genetic Map of the Escherichia
coli Chromosome

• The Escherichia coli chromosome has been mapped using conjugation, transduction, molecular cloning, and sequencing (Figure 10.42).


• E. coli has been a useful model organism, and a considerable amount of information has been obtained from it, not only about gene structure but also about gene function and regulation.