chapter 5 enzymes jia-qing zhang ph.d biochemistry department medical school jinan university mar....
TRANSCRIPT

Chapter 5 Enzymes
Jia-Qing Zhang Ph.DBiochemistry department Medical school Jinan UniversityMar. 2007

What are enzymes?
Enzymes are proteins which act as catalysts
Catalyst :
A catalyst is something which by its very
nature increases the rate of a reaction and
remain uncharged at the end of reaction.

3
catalyst
enzyme

4
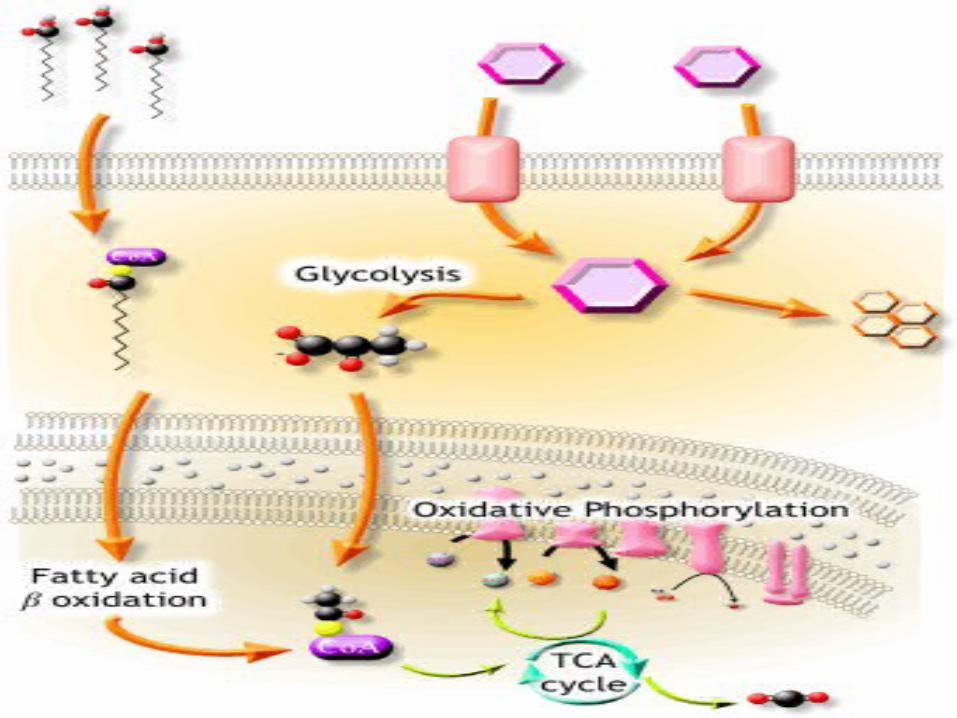
Enzymes control and regulate the
various metabolic activities inside living
cells.
5

In the absence of an enzyme:

whereas in its presence the rate can be
increased up to 107 fold.


The definition of enzymes?
Enzymes are powerful and highly effectual
biocatalyst produced by living tissues which
increase the rate of reactions that occur in the
tissue.

What are enzymes made up of ?
Almost all enzymes are make up of proteins

In 1982s, Thomas Cech discovered RNA possessing catalytic activity, were called ribozymes.
In 1995, Jack W. Szostak lab reported a DNA fragment with ligase function, termed Deoxyribozyme
So biocatalysts have enzyme, ribozymes, deoxyribozyme

13
Section 5.1 Structure and function of Enzymes
5.1.1 Structure of Enzymes
5.1.1.1 Composition of Enzymes Molecules Enzymes are claasified into
Simple enzyme (e.g. urease , protease , lipase etc.)
Conjugated enzyme. Most enzymes are conjugated.
Simple enzymes are all proteins,

Conjugated enzymes include protein component and
Non protein constituents,
Holoenzyme = apoenzyme + cofactorAn anpoenzyme is the protein part
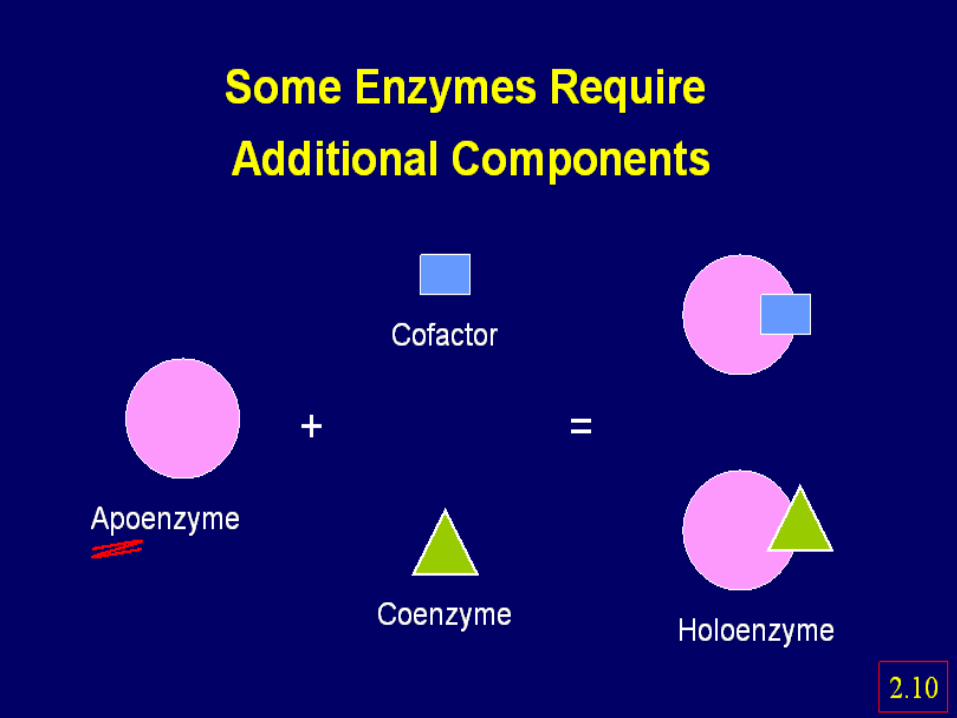
15

16
Cofactors commonly seen are 1. Metal ions ; iron, magnesium, cobalt manganese Metal ions tightly link to enzyme are known as
metalloenzyme
those link between E and S(substrate) are called metal activated enzyme(Table 5-2).
2. Low M.W. organic compounds. Vitamins often play their roles in H+ , electron
and other chemicals transfer when they take
place in metabolism as coenzyme ( Tab5-1 )

17
Cofactor
is divided into:
Coenzyme Prosthetic group

18
Differences between coenzyme and prosthetic group
1.Coenzyme linked to apoenzyme loosly and can be aparted from holoenzyme by dialysing,while prosthetic group linked tightly and can not be separated that way.

19
2.Coenzyme leave apoenzyme after catalyzing a reaction,
Prosthetic group can not depart from holoenzyme.

20
5.1.1.2 Monomeric enzymes,
Oligomeric enzymes and Mult
ienzyme complex

21
monomeric sarcosineoxidase
Monomeric enzymes
Monomeric enzymes only contains tertiary structure
trypsin
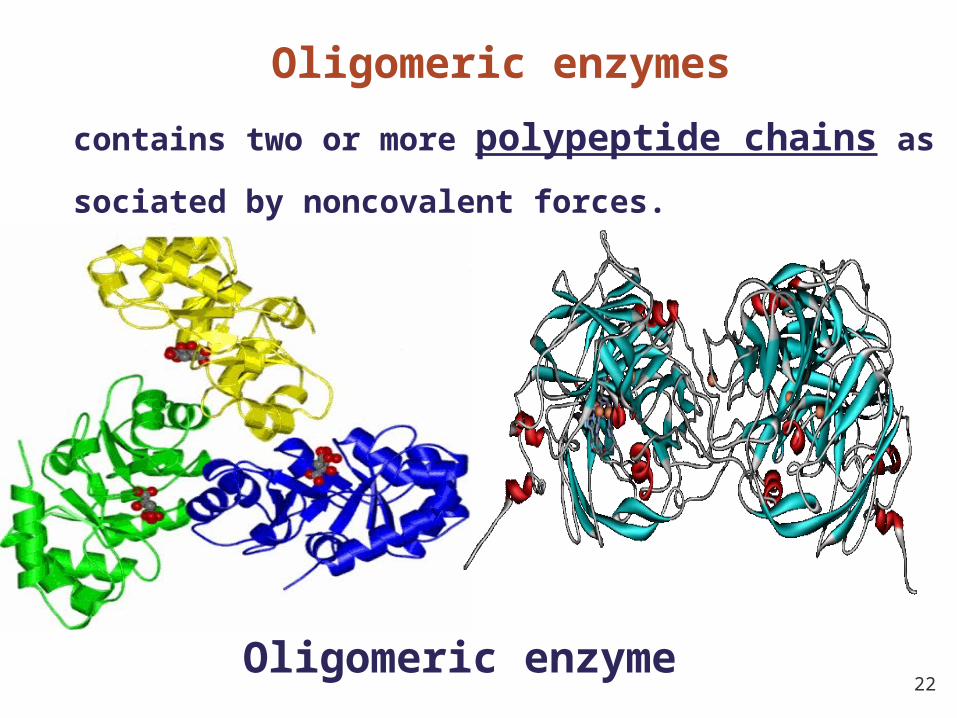
22Oligomeric enzyme
Oligomeric enzymes
contains two or more polypeptide chains associated b
y noncovalent forces.

23
Multienzyme complex is that different enzymes
catalyze sequential reactions in the same path
way are bound together. PDC
Multienzyme complex

24
5.1.2 Active site of an enzyme
Active center: the region of an enzyme that contains some chemical groups for binding substrate and for catalyzing the conversion of substrates to products.
All groups in active site are termed the essential groups.
Essential group: some chemical groups connected to activity of enzyme.They are classified into binding group and catalytic group.

25
底 物 活性中心以外的必需基团
结合基团
催化基团
活性中心 目 录
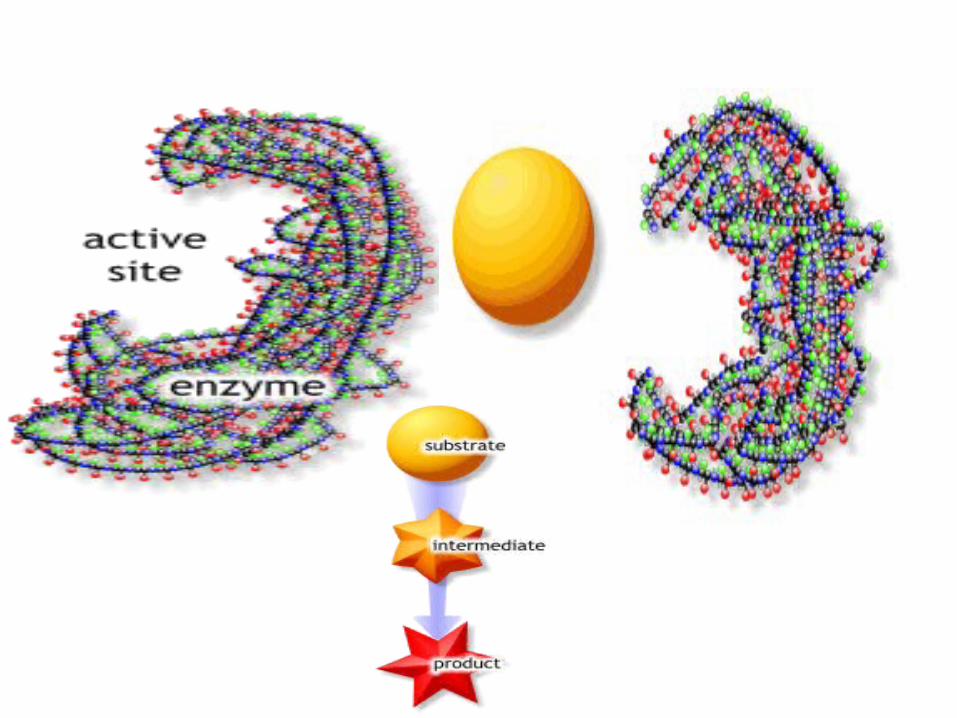

Active sites are
usually located in
clefts between
domains or
subunits or
indentations on
surface of proteins

28
5.1.3 Structure and function of enzymes
5.1.3.1 The primary structure of enzymes and Its function
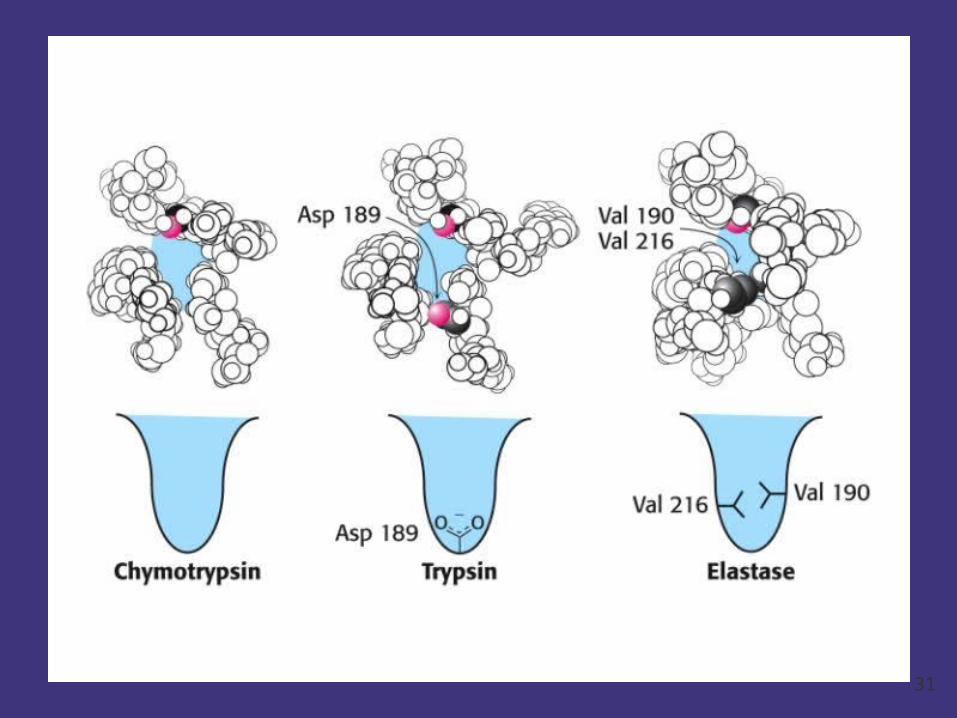
Simliar catalyzed function means structure homology.
eg. Chymotrypsin trypsin and elastase

29
Chymotypsin and trypsin

30
5.1.3.2 The spatial structure of enzymes and its function
The catalyzed activity of enzymes also depends on the conformation of enzymes.
31

Section 5-2 Nomenclature and Classification of Enzyme
In Naming an enzyme substrates are stated first
ending –ase is affixed
Enzymes are grouped into six classes according to the nature of the reactions



4) Peroxidases
5) reductase





Catalyze cleavege of bonds by addition of water



Catalyxe racemization of optical or geometric
Isomers and intramolecular oxidation-reduction
reactions


Ligases are usually referred to as synthetases

Carboxylase

5.3 Properities and catalytic mechanisms of enzymes
5.3.1 properities of enzyme catalytic reactions
5.3.2 catalytic mechanisms of enzymes

50
Properities of enzyme catalytic reactions
1).High catalytic activity of enzymes:
(a) Enzymes can decrease activation energy i.e.
molecules are activated using activation energy.
By decreasing activation energy,enzymes promotes
chemical reactions
(fig 5-5) .

51

52
(b) After Product has formed, free Enzymes are released and reused.
(c) Enzyme can make a reaction 103-107 times faster than uncatalyzed,

53
2) Highly specificity of enzymes:
Specificity refers to the ability of an enzyme to discriminate between two competing substrates.
The specificity can be divided into 3 types:
(a)Absolute specificity: the extreme selectivity of E that
allows it to catalyze only a single S.
e.g chymotrypsin hydrolysis peptide bond having aroma
tic AA, trypsin catalyse those having basic AA.

54
(b) Group specificity(relative specificity):
Enzymes atc on a group of related substrates
e.g. Hexokinase catalyzes the phosphorylation of
glucose, mannose, fructose, glucosamine
Esterase catalyse the break down of ester
bonds .

55
(c) Optical(stereo) specificity:
e.g. LDH hydrolyzes L-lactic acid, but not
D- lactic acid

56
3). Under control :enzyme can be regulated
Regulated in different levels: biosynthesis, allo
steric regulation covalent modification isoenzy
mes.

57
5.3.2 catalytic mechanisms of enzymes
Enzymes increase the rate of reaction by lowering the
activation energy?
Then, how to decrease the energy?

58
Formation of ES complex and Induced-fit hypothesis
The enzyme reversibly combines with its substrate
to form an ES complex, that subsequently break down
to product, regenerating the free enzyme ;
E + S ----- ES ------- P + E
P

59

60
Lock and Key Theory:
The specific action of an enzyme with a single substrate
can be explained using a Lock and Key analogy first po
stulated in 1894 by Emil Fischer. In this analogy, the lock
is the enzyme and the key is the substrate. Only the corr
ectly sized key (substrate) fits into the key hole (active
site) of the lock (enzyme).

61

62

63
The following experimental evidence show the lock
and key theory is rigid model and can not explain the
ES complex well. For this reason a modification
called the induced-fit theory has been proposed

64
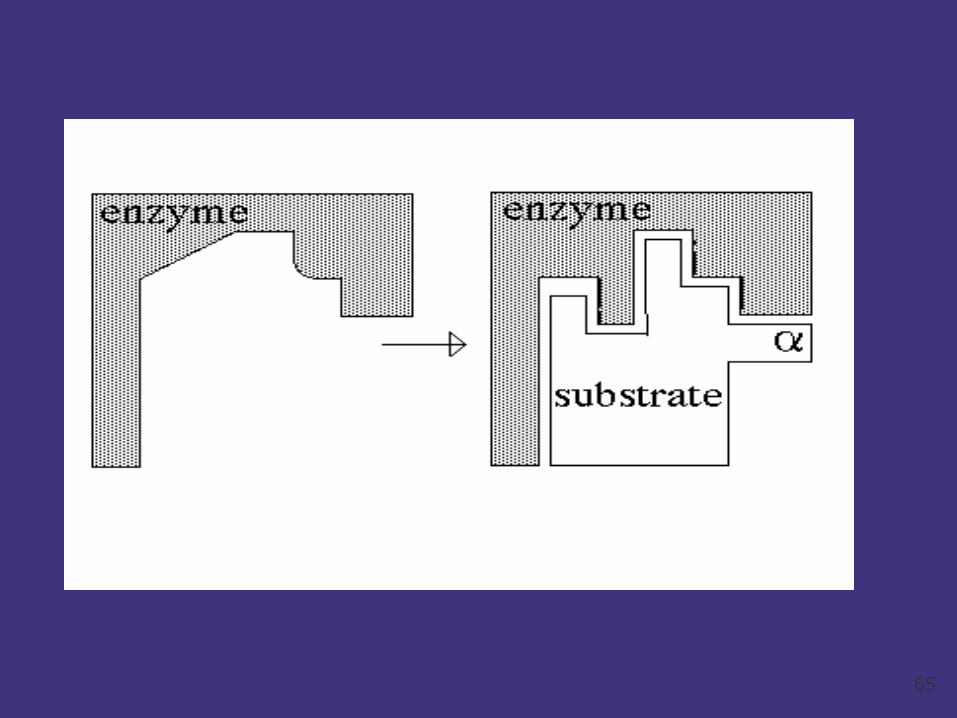
Induced Fit Theory:
The induced-fit theory assumes that the substrate
plays a role in determining the final shape of the
enzyme and that the enzyme is partially flexible. I
65

66
summaryenzyme\enzyme_binding.swf

67
5.3.2.3 several factors contribute to enzyme catalysis
2. Electrostatic effects
3. Acid-base catalysis
4. Covalent Catalysis
1. Proximity effects and orientation arrange:

68
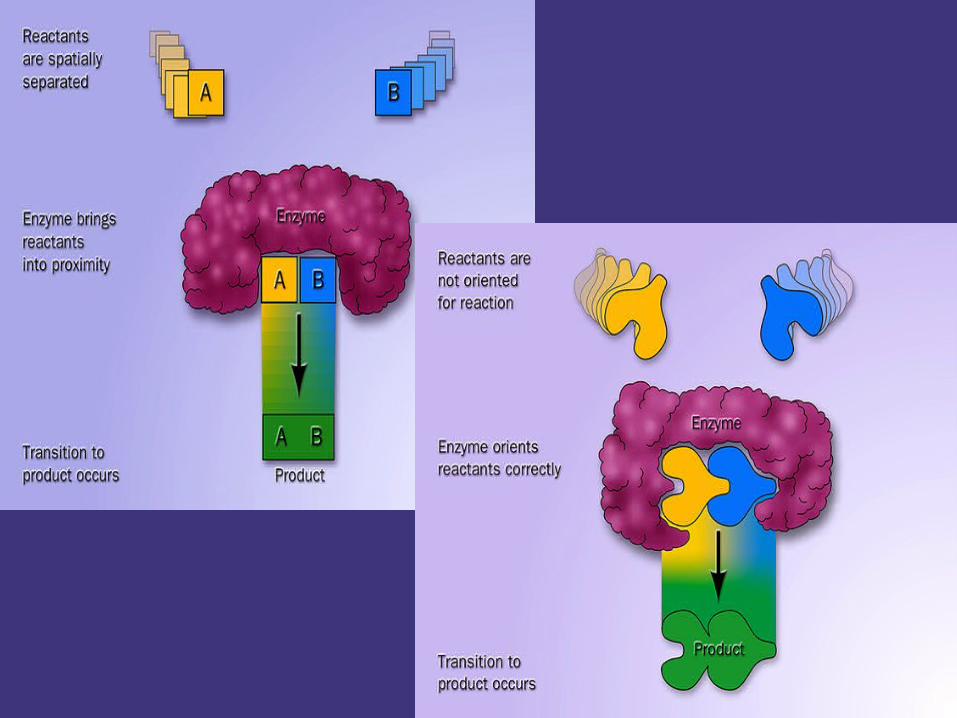
1. Proximity effects and orientation arrange:
Chemical reactions are based on molecular
collision,proximity and orientation lead to correct
direction for collision.
69

70

71
2. Electrostatic effects The strength of electrostatic interactions can reduce
the attractive forces and increase the chemical
reactivity of the substrate.
3. Acid-base catalysis
Enzymes is usually ampholyte, can provide acid or basi
c enviorment, partial proton tranfer from an acid or to a
base lowers the activation energy. the rate of reaction c
an often be promoted by adding or removing a proton.


4. Covalent Catalysis
Accelerates reaction rates through the transient formation
of a catalyst-substrate covalent bond


Section 5.4 Kinetics of Enzyme-catalyzed reactions
KineticsThe branch of chemistry that is concerned with the
rates of change in the concentration of reactants in a
chemical reaction.
Enzyme kinetics
Study of the rate of change of reactants to products and factors by which rate of reaction is influenced, such as substrates, activators, inhibitors, pH and temperature.
75

Enzyme activity can be expressed by kinetic properties:
Internation units(IU) one IU is defined as the amount of enzyme that produces 1umole of product per minute.
1 katal(kat)1 katal(kat) is equal to the amount of enzyme that converts one mole of substrate to product per second.

5.4.1
The effect of substrate on the rate of e
nzyme-catalyzed reactions

5.4.1.1
The basic conditions for discussing the
effect of substrate on the rate of enzyme-
catalyzed reactions

79
5.4.1.2 Rectangular hyperbola plot of initial
velocity V0 versus substrate concentration
[S]


[S][S]
VVVmaxVmax
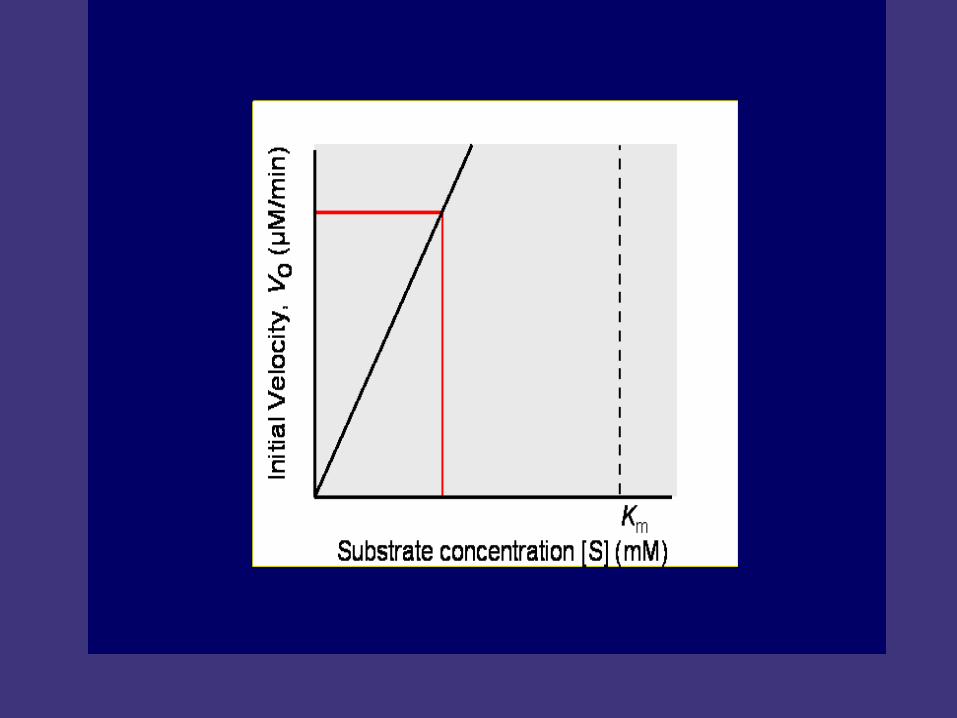
At low [s], V0 increases linearly with increase in [S]
[S][S]
VVVmaxVmax
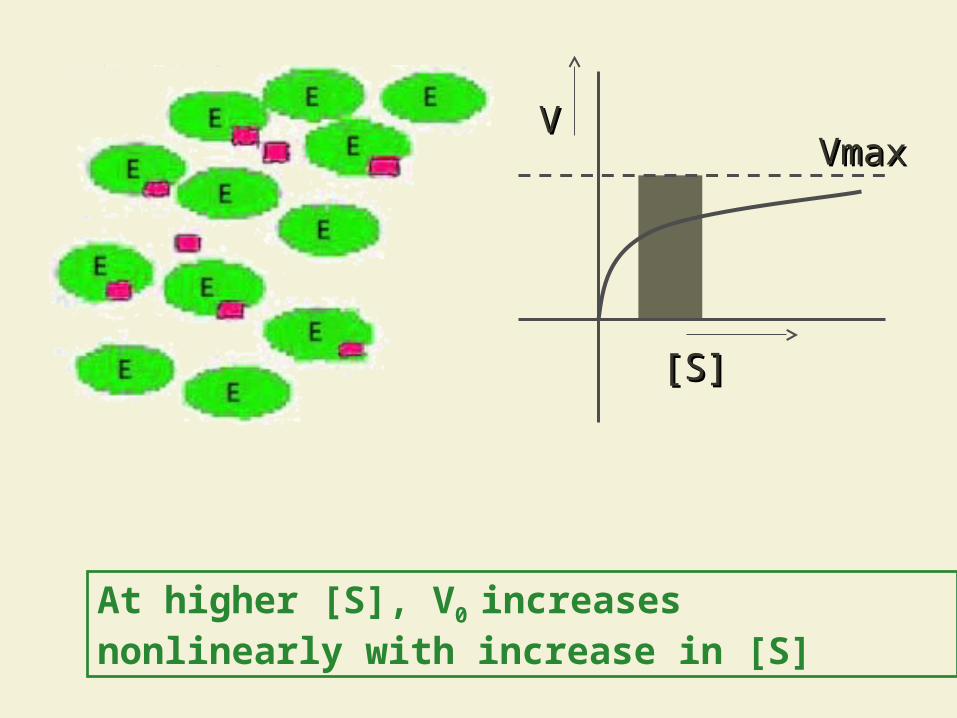
At higher [S], V0 increases nonlinearly with increase in [S]
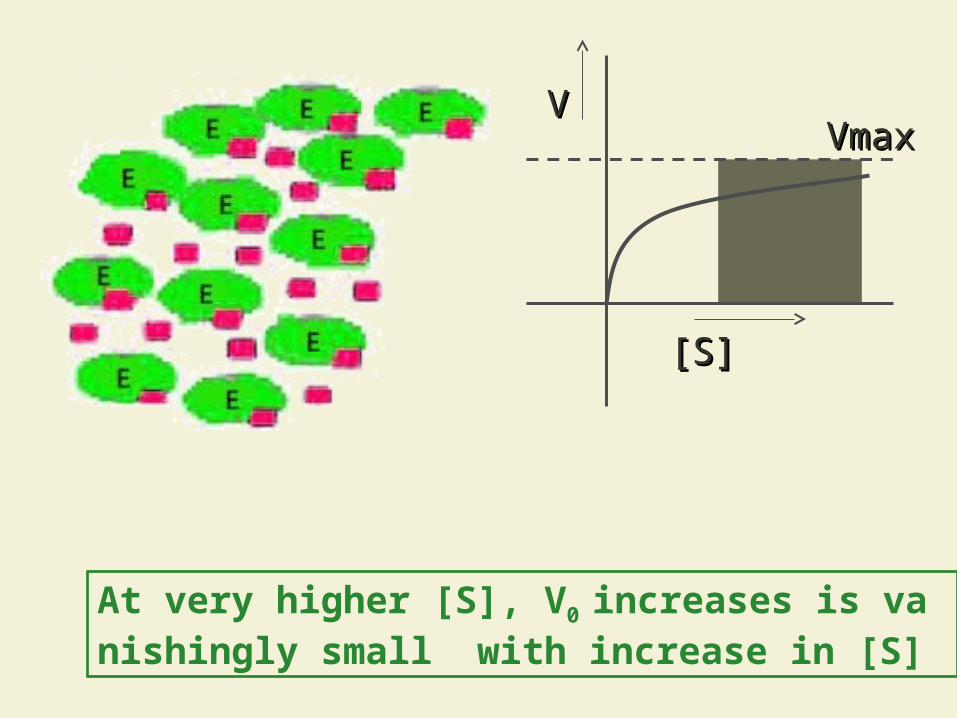
[S][S]
VVVmaxVmax
At very higher [S], V0 increases is vanishingly small with increase in [S]


5.4.1.3 Formulation of the Michaelis-menten equation
Michaelis-menten equation, a mathematical equ
ation expressing the hyperbolic relationship between
the initial velocity, Vo, and the substrate concentratio
n, [S].




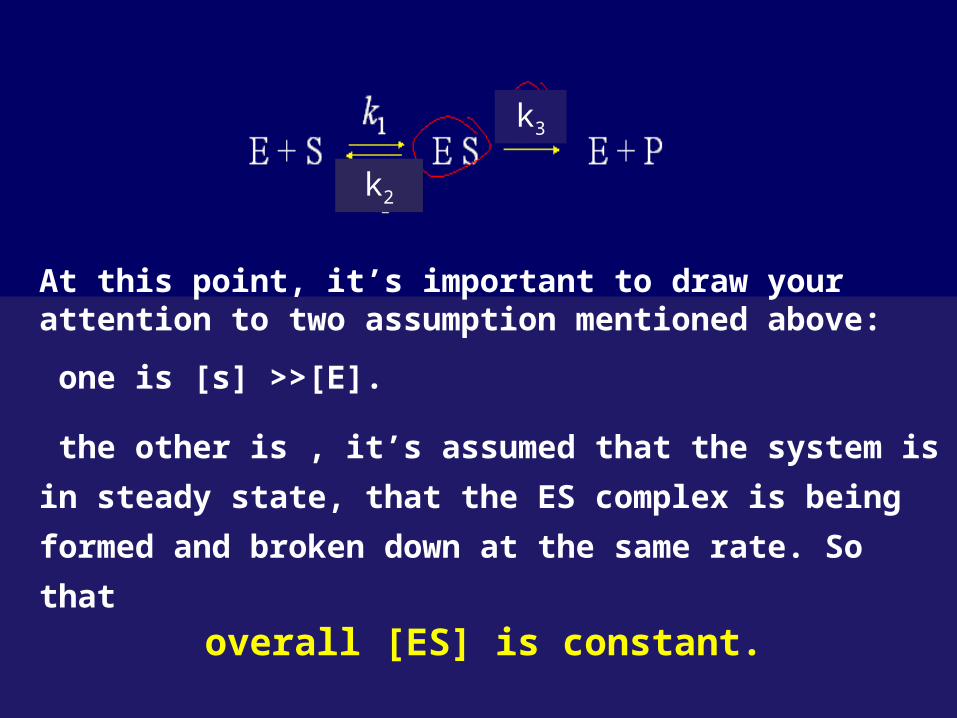
The enzymes-catalyzed reaction process may be
summarized as follow:
k2
k3
k4
,so k4 ingored

k2
k3

k3
k3
k2
[1]
k3 [1]
93
k3
k3
k2
[1]
At this point, it’s important to draw your attention to two assumption mentioned above:
one is [s] >>[E].
the other is , it’s assumed that the system is in steady
state, that the ES complex is being formed and broken
down at the same rate. So that
overall [ES] is constant.

94
Formation equal to breakdown, then
K1[E][S] = (k2 + k3) [ES]
The formation of ES will depend on the k1 and the
availability of E and S.
So
Rate of ES formation = k1 [E][S]
The breakdown of [ES] can occur in two ways, either the conversion of S to P, or non-reactive dissociation of S from ES complex.
Rate of ES breakdown = (k2 + k3) [ES]

95
[ES]
k1
K2 + k3=
[E][S]
By rearranging
K1[E][S] = (k2 + k3) [ES]
[2]

96
[ES]=
[E][S]
As the name implies, these 3 rate terms k are constants, so M
ichaelis actually combine them into one term, this new consta
nt is termed the michaelis constant and is written Km
k1
K2 + k3=Km
Substituting the Km into equation [2]
[2]
Km
[ES]
k1
K2 + k3=
[E][S]

97
The total amount of enzyme in the system must be the same throughout the experiment, but it can either be free (unbound) E or in complex with substrate, ES. If we term the total enzyme Et, this relationship can be written out:
t
[3]
Substituting this definition of [E] back into equation [3] gives us:

98
t
t
[E] = [Et] –[ES]

99
First of all, open the bracket so that the [E] and [ES] are separately multiplied by [S]
t
Next, multiply each side by KM, this gives us:
t
t

100
Then collect the two [ES] terms together on the same side. This gives:
t
Then because both terms on the right-hand side are multiplied by [ES] we can collect them together into a bracket:
Dividing both sides by (KM + [S]) now gives us:
t
t

101
t
Substituting this left-hand side into Equation 1 in place of [ES] results in:
3
t0 [4]

102
The maximum rate, which we can call Vmax, would be achieved when all of the enzyme molecules have substrate bound. Under conditions when [S] is much greater than [E], it is fair to assume that all E will be in the form ES. Therefore
[Et] = [ES]
Notice that k3[Et] was present in equation 4, so we can replace this with Vmax, giving a final equation:
Thinking again about Equation 1, we could substitute the term Vmax for v and [Et] for [ES]. This would give us:
Vmax = k3[Et]

103
0
This final equation is actually called the Michaelis-Menten equation.
3t
0 [4]

104
5.4.1.4 The Significance of Km and Vmax
Let us consider the case when V is exactly half of Vmax. Under those circumstances, the Michaelis-Menten equation could be written:
On dividing both sides by Vmax this becomes:

105
Multiplying both sides by (KM + [S]) gives:
And then multiplying both sides by 2 further resolves the equation to:

106
[S] on the right-hand side is the same as [S] + [S], so we
can take away one [S] from each side. Thus when the
rate of the reaction is half of the maximum rate:
If we now reconsider the graph that present at
the start of this class it could be written:

107
Significance of Km:
1. Km is an important constant depends on E , S structure and reaction environment.
2. Km is equal to the [S] at which the reaction occurs at half of the Vmax rate.

108
3. Sometime(When k2 is much greater than k3), km c
an indicates the affinity of the ES complex.
Some enzymes have more than 2 substrates, higher K
m, weaker binding of S and E.
e.g. Glucose and Fructose are substrates of hexokina
se , Glu has less Km , higher affinity to hexokinase, in c
ontrast, Fru. has higher Km and less affinity.
If E has more than 2 substrates, the least Km is th
e optimum

Significance of Vmax:
Vmax, the maximal initial velocity, is obtained only whe
n all the enzymes is in the form of the ES complex, from
which it follows that:
From above equation, Vmax = k3 [Et]
3
2
3
2

110
K3 is the turnover number of enzymes.
Turnover number of an enzyme
is the number of substrate molecules converted into
product by an enzyme molecule in a unit of time when
the enzyme is fully saturated with substrate.

, Vmax


113
Double reciprocal plot ( Lineweaver-Burk plot)
In order to obtain accurate Km, the Michaelis formul
a can be turned into a linear formula by up-side-dow
n the numerator / denominator so as to obtain 1/ Vm
ax at y axis and 1/Km at x axis.

0
up-side-down the numerator / denominator, give us

1/v0 : y
1/[s]: x




119
The temperature at which enzymes operate at
maximal efficiency
Optimum temperature



122
At a pH above this optimum, the enzyme's activity will be reduced and therefore the reaction rate will be lowered; at a pH below this optimum, the enzyme's activity again will be reduced and lower reaction rates result


5.4.5 The effect of inhibitors on the rate of enzyme-catalyzed reactions
Inhibitor (I)
An agent that can decrease enzyme activity without
causing denaturation of enzyme.
Inhibitor can be classified into:
(1)Irreversible inhibitors:
(2) Reversible inhibitors: Competitive, un-competitive, non-competitive

Irreversible inhibition
Irreversible inhibitors bond and destroy a functional
group in an enzyme that is essential for the enzyme
activity

126
•In most cases forms a covalent link with the enzyme
•Permanently renders the protein inactive

127
AChE
Inhibitor--Organic phosphorus
Inhibition of AChE by organic phosphorus
PAM
Organic phosphorus (public Hazards in a
gricultural drugs) : usually binds to active sit
e of acetylcholine esterase.
ACh

-noncompetitive

129
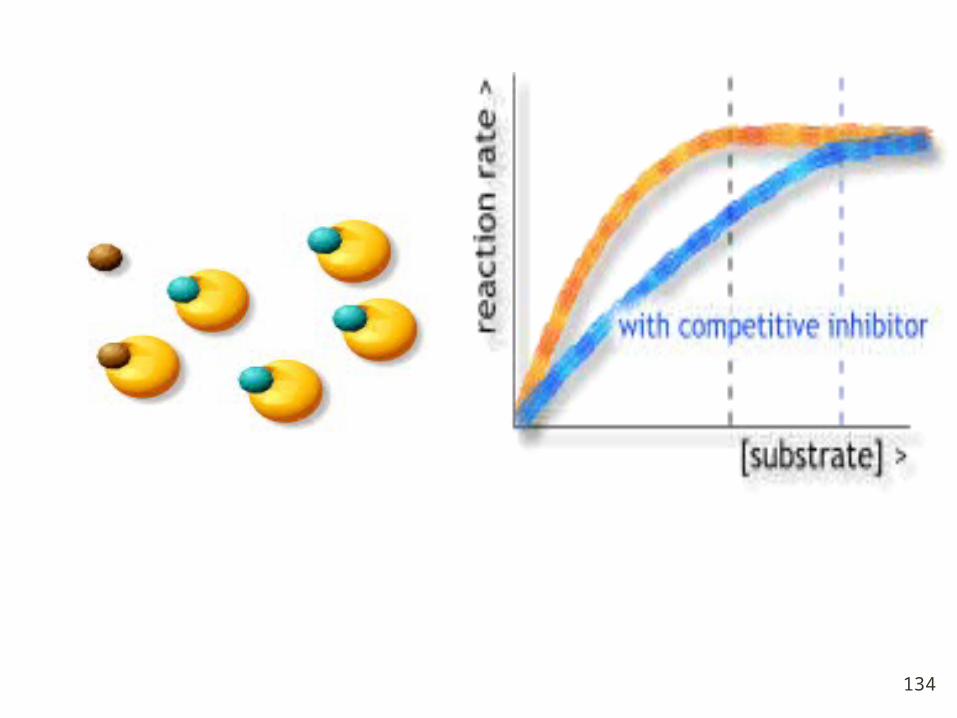
Chemicals that resemble an enzyme's normal substrate an
d compete with it for the active site. Block active site fro
m the substrate.
If reversible, the effect of these inhibitors can be overco
me by increased substrate concentration.
Competitive inhibition

130


132
In competitive inhibition, both inhibitor and substrate can
bind to enzyme and form two independent complexes.
Only ES degrades to products: EI is considered a 'dead-
end'. Because the inhibitor binds to the active site, the
substrate cannot (and vice versa), so there cannot be
an ternary ESI complex.

133
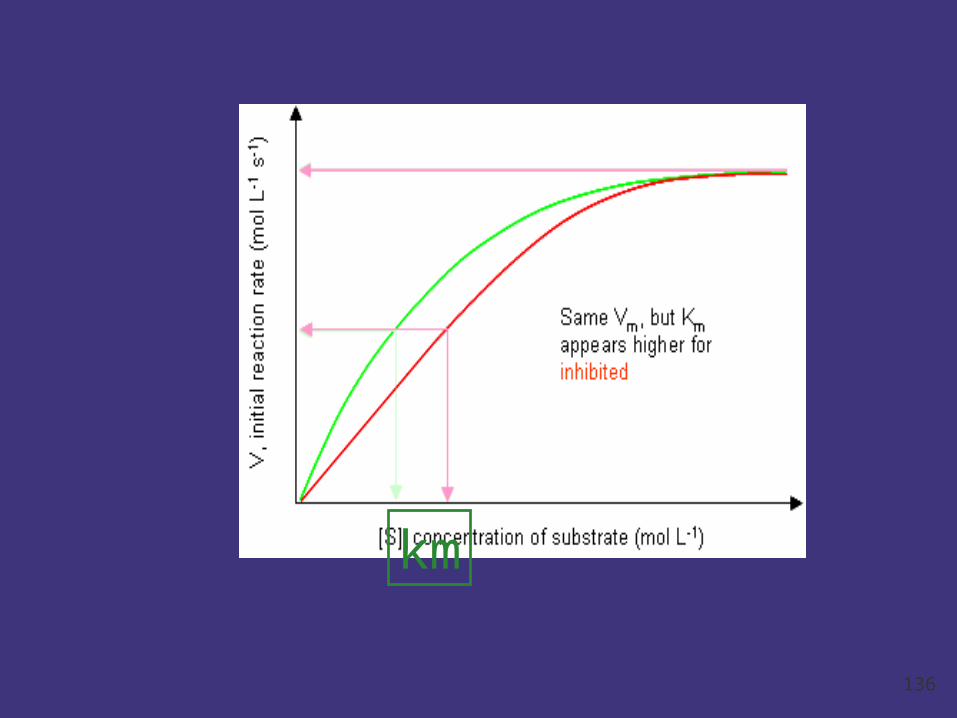
Mechanism and Features:
1. I and S compete the active site;
2. Which one occupies the active site depends
on the affinity of S / I affinity to E and concentr
ations between I and S.
3. Km increase ( affinity decrease), Vmax unc
hanged
134

Mathematical analysis
For those who are interested in Maths
only
136
km

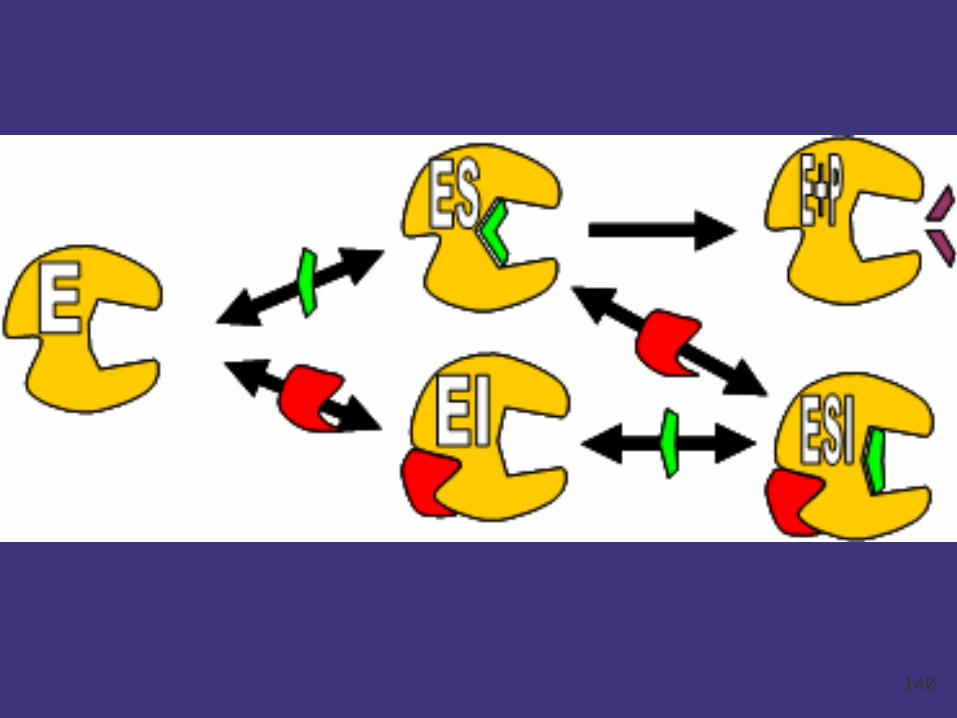
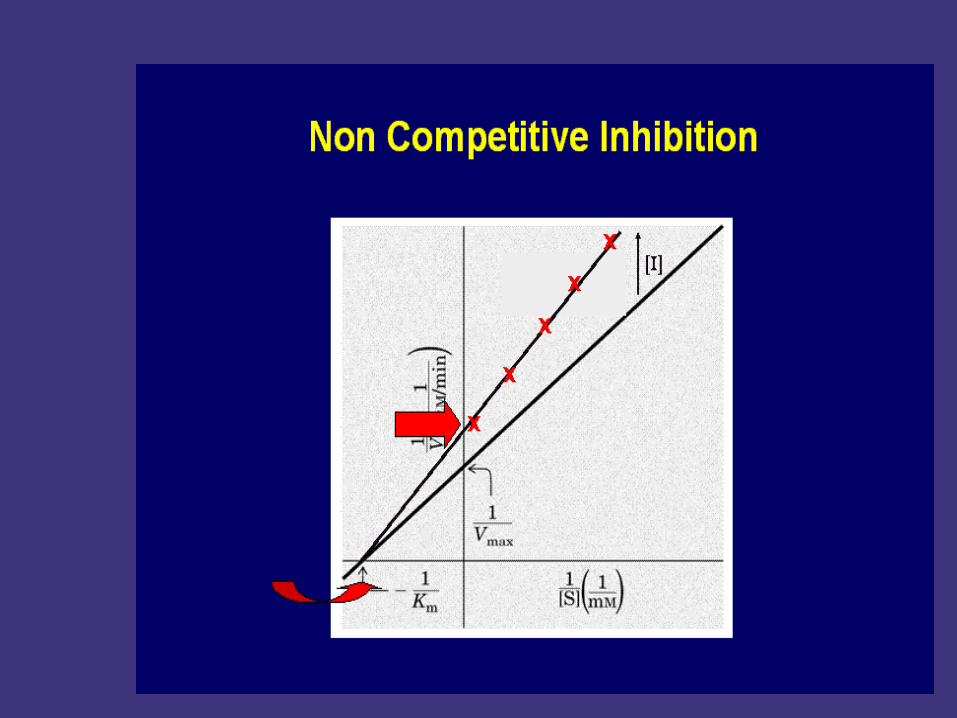
Noncompetitive inhibition
Noncompetitive inhibitors :Enzyme inhibitors that do
not enter the enzyme's active site, but bind to another
part of the enzyme molecule. Causes enzyme to
change its shape so the active site cannot bind
substrate.

138

139
140

141
The Vmax is decreased without a change in the K
m for the substrate
The inhibition cannot be overcome by increaseing
the substrate concentration.
Features of non-competitive inhibition:

142

144
Uncompetitive inhibition
Uncompetitive inhibitors bind only to the ES
complex and not the free enzyme.



Features:
Decrease of Km and Vmax.
summary

148
5.4.6 Activators of enzymes
Activator: Agents increase the activity of enzyme
or make the inactive form become active form are c
alled Activator of enzyme.
They are mostly metal ions(Mg2+,K+,Mn2+),also som
e anion(Cl-)and organic compounds.
Most anions are necessary to enzyme activity,they
are called essensial activator.Of course there are n
on-essential activator,Cl- is so to ptyalin .

Section 5.5 Regulation of Enzyme Activity
•Allosteric regulation
•Covalent modification
There are two major strategies for regulating enzymes:


151
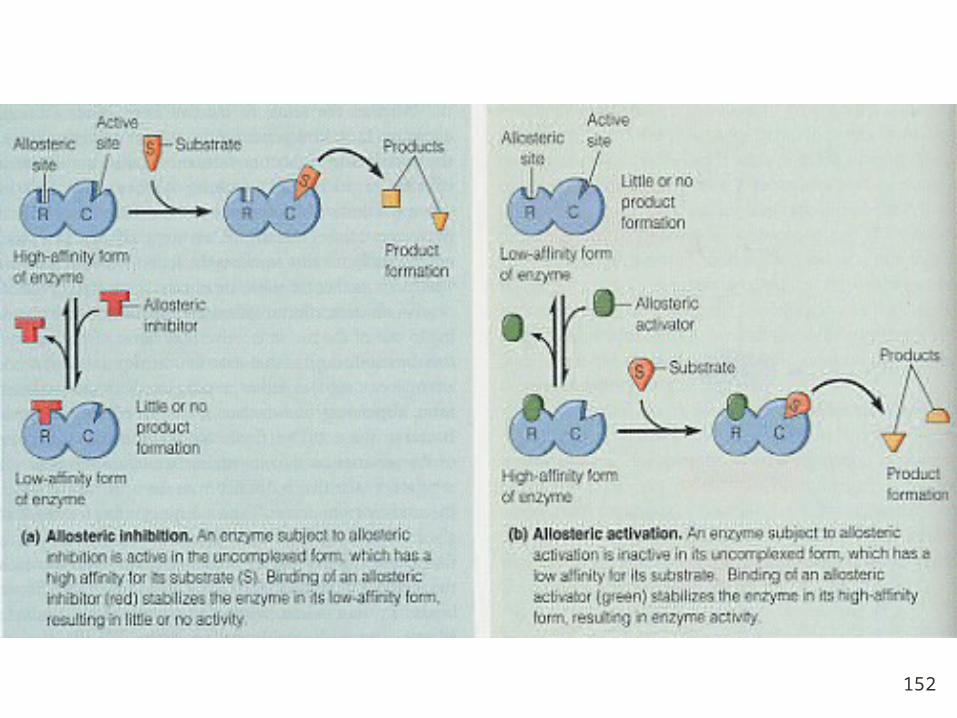
The mechanism of Allosteric regulation
152

153
General properties of allosteric enzymes
1. The activities of allosteric enzymes are charged by metabolic activators and inhibitors, which seldom resemble the substrates or products. So allosteric regulation is not inhibition.
2. Modulators bind noncovalently to enzymes.
3. Allosteric enzymes almost possess quaternary structure.
4. Allosteric enzymes often display sigmoid plots of the the reaction V versus S

154

155
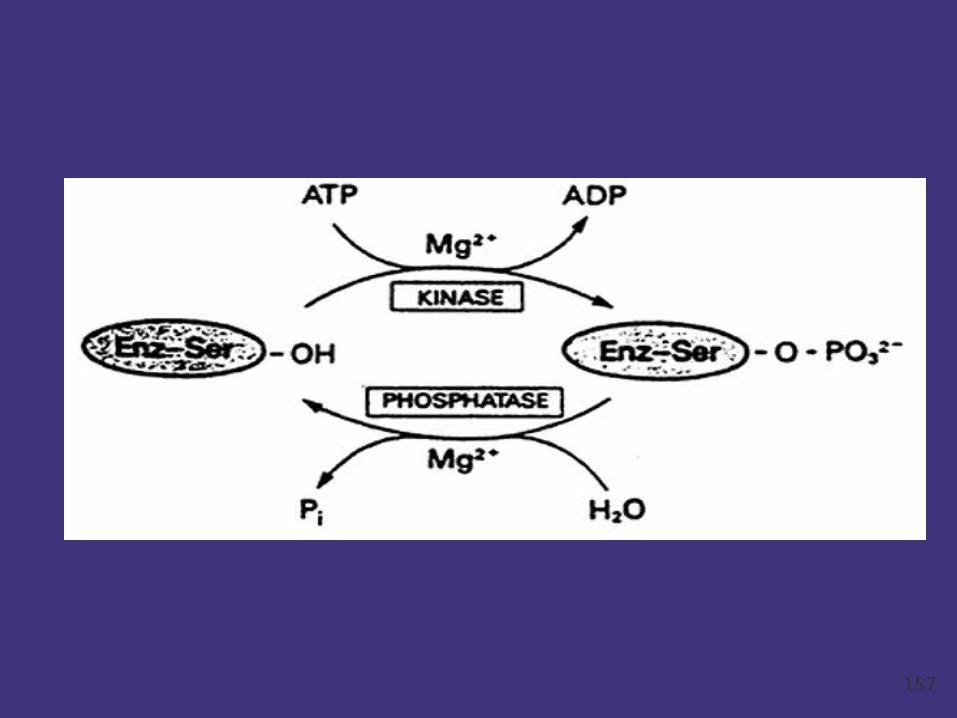
Covalent modification
The covalent attachment of another molecule can
modify the activity of enzymes and many other
proteins.
In these instances, a donor molecule provides a
functional moiety that modifies the properties of
the enzyme.
Most modifications are reversible.

156
Phosphorylation and dephosphorylation are the
most common but not the only means of covalent
modification.
157

158

159
Other forms of modification:
Acetylation
Methylation
nucleoside modification.
Significance of modification :
Rapidly change between the active and inactive form

160

161
Zymogen
Several enzymes are produced and stored as
inactive precursors called zymogens

162

163



166
e.g. LDH(lactate dehydrogenase) LDH
1,2,3,4,5 are HHHH, HHHM, HHMM, HM
MM and MMMM . i.e. LDH isozymes are
tetramers formed by 2 sets of subunits.

167

168
Significance :
In the heart, LDH1 (4 H) H=heart, catalyzes Lactate convert to pyruvate for energy supply
In muscles, LDH5 ( 4M ) M=muscle, convert pyr to Lact. For energy storage.
Clinical diagnosis using isozyme. E.g. when heart attack(infarction) happens, enzymes release from injured cells to the blood showed different enzyme(isozyme ) pattern.
Isozyme pattern: different isozymes appear as a peak sooner or later followed by the progress of the disease.

169
Summary
1.Definition:
active site, allosteric regulation, Covalent modification,
Zymogen, Isoenzymes, Coenzyme, prothetic group,
2. Explain the kinetic significance of Km and Vmax
3. What would affect the activity of an enzyme, and how?
4. What are differences betweeen competitive ,
noncometitive and uncompetitive inhibition?
6. How does the value of Km and Vmax change when adding competitive, noncompetitive and uncompetitive inhibitor respectively?