characterization of lipase producing …. r esults and d iscussion a. testing for lipase production...
TRANSCRIPT

Characterization of Lipase Producing
Rhodococcus sp. from Peninsular Malaysia
Jayesree Nagarajan, Norazah Mohammad Nawawi, and Abdul Latif Ibrahim Institute of BIO-IT Selangor, Universiti Selangor, Malaysia
Email: {jayesree_nagarajan, n_azahmn, alatifbio}@yahoo.com
Abstract—Rhodococcus species is recognized as an excellent
candidate for bioremediation due to its mass storage of
enzymes capacity. However, studies on lipase producing
ability by this actinomycete are far less being explored. Unit
Culture Collection of Institute Bio-IT Selangor preserves
and maintains Rhodococcus isolates which has been isolated
throughout Peninsular Malaysia. In this study, five
Rhodococcus isolates from various environmental niches
were identified for lipase production. The best isolate was
further assessed for lipase producing ability at different
grinding time interval. For better understanding on
behaviorism and adaptations of these isolates, further
characterization based on growth profiles and a few
biochemical tests were performed. Primary goal of this first-
stage study is to introduce new and potential source of lipase
from local resources while understanding the characteristics
of the isolates.
Index Terms—rhodococcus sp., bioremediation, lipase,
growth profile, biochemical tests.
I. INTRODUCTION
Enzyme or better known as protein molecule holds
fundamental tasks of accelerating chemical reactions in
living organisms. Precisely, this molecule serves as basic
tool for energy generation, growth, repair, cell
maintenance and formation of side products in every
biological system [1]. Among diverse available enzymes,
lipases are regarded as most pliable biocatalyst, ideal for
vast commercial applications [2]. Superiority of microbial
lipases is well represented by hiking number of research
articles and denotation as versatile biocatalyst.
Microbial lipases attained great preferability due to its
possibility for bulky production, consistent supply due to
absence in seasonal fluctuation, cheaper and highly stable.
In addition to these, lipase from microbes also exhibit all-
encompassing substrate selectivity, safer chemical
reactions while requiring mild growing conditions mostly.
Advantageously, development in the genetic engineering
also, promises use of recombinant gene technology in
order to increase the mass production of the cell, improve
efficiency of the enzyme either constitutively or by
inducing it and also introducing altered enzyme [3].
It was understood that, only 2% of world’s
microorganisms have been recognized as enzyme source
in which case, bacteria attained higher portion than yeasts
[4]. Diverse numbers of microorganisms have been
identified as supreme source for lipases including
Bacillus sp., Lactobacillus sp., Burkholderia sp. and
Candida sp. [5]. Based on the abundant studies of
bacterial lipases, further deep review revealed that
actinomycete or better known as branched filaments and
rather fragmentary bacteria are scarcely being studied on
enzyme producing ability [6].
Actinomycetes such as Rhodococcus sp. are also
pronounced for lipase producing ability however, detailed
evidences and analysis on this actinomycete are
inadequately explored [7]. This bacterium is popularly
known for its unique feature of producing natural oil
hence, arouses remarkable possibilities on its
triacylglycerol metabolism [8]. Potency of Rhodococcus
sp. in degrading hydrocarbon or crude oil extensively has
become an additional quality as potential source for
lipase enzyme as well [9]. However, the key factor for
this quality lipase, the fat degrading enzyme is far less
being explored.
Unit Culture Collection of Institute Bio-IT Selangor,
Malaysia maintains and preserves Rhodococcus isolates
which has been isolated throughout Peninsular Malaysia.
These isolates have been partially sequenced and further
compared with an established reference strain namely
Rhodococcus sp. 124. In this study, lipase producing
ability of these isolates was assessed. The strains were
further subjected for few characterization tests and
growth profile analysis for better understanding of
lipolytic Rhodococcus sp. behaviorism. This study
introduces new source for lipase enzyme among
diversified number of lipase sources ideal for applications.
II. MATERIALS AND METHODS
A. Bacteria and Seed Culture Preparation
Five Rhodococcus sp. isolated throughout Peninsular
Malaysia were kindly supplied by Culture Collection
Unit, Institute Bio-IT Selangor. Cultures from glycerol
stock were streaked on nutrient agar plate and incubated
at 30ºC. For seed culture, a loopful of bacteria from
nutrient agar plate was inoculated into 50 ml of nutrient
broth medium and left for shaking in incubator shaker
(Jeio Tech SI-600R, Korea) at 30 ºC at 160 rpm for
overnight. The details of the tested strains were tabulated
in Table I. As a reference strain for positive control
Bacillus subtilis (NCBI accession no: GU191916) from
122014 Engineering and Technology Publishingdoi: 10.12720/jolst.2.1.12-19
Manuscript received March 31, 2014 2014.; revised September 11,
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014
the culture collection were compared with the tested
isolates.
TABLE I. RHODOCOCCUS SP. AND ITS ISOLATED LOCATION
Rhodococcus
Strains
Location
R.NAM81 Palm oil Mill effluent, Kuala Selangor
R.NAM319 Palm oil Mill soil, Kuala Selangor
R.NAM350 Palm oil Mill effluent, Kuala Selangor
R.UKMP-5M Petroleum contaminated soil, Port Dickson
R.SeAG1 Oil contaminated soil, Serdang
B. Testing for Lipase Production
1) Production medium
Ten percentage of seed culture was inoculated into
production medium containing 0.1% (w/v) NH2SO4, 0.09%
(w/v) K2HPO4, 0.06% (w/v) KH2PO4, 0.02% (w/v)
MgSO4.7H2O, 0.01% yeast extract and 1% (v/v) olive oil.
2) Preparation of crude lysates
The culture was centrifuged for 15 minutes at 14000
rpm (Tomy MX350, Japan). Upon centrifugation, the
supernatant was discarded. The remaining cell pellet was
washed twice and 2 ml 0.05M potassium phosphate
buffer was added. Cell lysates was prepared by grinding
the cell pellet under liquid nitrogen with a pre-cooled
mortar and pestle for 10 minutes. The lysed sample was
centrifuged again for 15 minutes at 14000 rpm. The
yielded supernatant was used as crude extract for lipase
assay [10].
3) Lipase assay
Lipase activity was determined using titrimetric
method using olive oil as substrate. Emulsified substrate
was prepared using 5% (w/v) olive oil and gum arabic
incubated in water bath for homogenization. 1 ml of
extracted crude was added into 5 ml of emulsified
substrate. The mixture was later on incubated at 30 ºC for
2 hours. The reaction was terminated by addition of 10 ml
of 95% (v/v) ethanol solution into the mixture. Fatty acid
liberated was titrated using 0.05 M NaOH against 2-3
drops phenolphthalein as indicator. One unit of lipase
defined as amount of enzyme required to liberate one
micromole fatty acid under specified assay condition [11]. 4) Empirical study on lipase activity on UHT milk
Ultra-high treatment or UHT processed milk (Dutch
Lady) was used for this analysis. 5 ml of UHT milk was
prepared in a screw capped tube with addition of 2-3
drops of phenolphthalein. Later on, 1 ml of extracted
crude lysates from each isolate was added in the prepared
tubes each. The tubes were incubated at 37 ºC in water
bath and observed for any color changes after 30 minutes
[12].
5) Effect of different grinding time on cell lysates
with best lipase activity
Isolate which demonstrates the best lipase activity was
further cultured for six samples. Each sample was ground
from 10 minutes to 60 minutes with 10 minutes interval.
Lipase activity from each cell lysates was assessed. The
extracted crude enzyme was also subjected for SDS-gel
electrophoresis [13].
C. Characterization of the Isolates
1) Growth profile
a) Measurement for optical density (OD600nm)
The cultures were incubated in nutrient broth and
approximately 1ml of samples was withdrawn aseptically
for every 6 hours of intervals including 0 hour. Each
sample was withdrawn in triplicates aseptically. Prior to
quantification, the samples in cuvette were thoroughly
suspended and absorbance readings were obtained at
600nm wavelength against distilled water. The used
cuvettes were originated from Germany
(PLASTIBRAND), a semi-micro disposable cuvette with
the volume capacity of 1.5ml.
b) Measurement for dry cell weight (g/L)
Sterilized micro-centrifuge tubes with the capacity of
1.5 ml were oven-dried at 80 ºC for overnight. Prior to
use, the micro-centrifuge tubes were cooled down in
room temperature and the empty micro-centrifuge tubes
were weighed accordingly (Sartorious TE214S,
Germany). As previous, one ml of samples was pipetted
aseptically into the micro-centrifuge tubes in triplicates
for every interval of 6 hours including 0 hour of
incubation time (Mills & Lee, 1996). Later on, the
samples were subjected for centrifugation for 15 minutes
at 14000 rpm to obtain the cell pellet. Upon
centrifugation, the supernatant was gently discarded and
the cell pellets were re-suspended thoroughly with
autoclaved distilled water. Another three cycles of
centrifugation steps were repeated to eliminate possible
contaminants presence in the final cell pellet. Finally, the
micro-centrifuge tubes containing the washed cell pellets
were oven dried at 80 ºC for 24 hours. The micro-
centrifuge tubes were pre-cooled to room temperature
before being weighed and the readings were tabulates.
The calculated dry cell weight was expressed in (g/L).
2) Biochemical tests
The five isolates were subjected for few biochemical
tests. The chemicals used are shown in Table II.
TABLE II. LIST OF CHEMICAL/REAGENTS INVOLVED FOR EACH
BIOCHEMICAL TESTS
Biochemical Tests Chemical/ Reagents
Gram staining Safranin, iodine, crystal violet,
decolorizing (Merck), Iodine Solution
(PC Laboratory Reagent)
MacConkey MacConkey agar (Pronadisa Conda) Agar-agar (R&M chemicals)
Simon Citrate
utilization
Simon Citrate Agar (Pronadisa Conda),
Agar-agar (R&M chemicals)
Catalase test Hydrogen Peroxide
MR-VP test MR-VP broth (Merck)
Methyl Red (Sigma Aldrich)
Starch Hydrolysis Starch Agar (Pronadisa Conda)
Agar-agar (R&M chemicals) Iodine Solution (PC Laboratory
Reagent)
Kovacs’s indole test Trypthophan Peptone Broth (Oxoid) Methyl Red (Sigma Aldrich)
Antibiotic test Kanaamycin, Gentamicin, Ampicillin,
Tetracycline, Vancomycin (BioBasic, Inc) Antimicrobial susceptibility disks
(Oxoid)
132014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014

III. RESULTS AND DISCUSSION
A. Testing for Lipase Production
1) Lipase assay
As shown in Fig. 1, the tested isolates projected lipase
activity from the range of 4.7 U/ml to 9 U/ml. This
outcome emphasizes that Rhodococcus isolates exist as
possible source for lipase production. R.NAM 319
displays highest lipase activity, potent to be most
promising isolate among other isolates. Interestingly, this
isolate even displayed higher enzyme activity compared
to the tested positive culture. R.NAM81, R.NAM350 and
R.SeAG1 on the other hand, exhibited lipase production
almost in similar level. In this assay, lipase hydrolytic
activity was evaluated based on oleic acid (C18), a long
carbon chain [10]. As a hydrolytic enzyme, lipases would
disrupt the major chemical bond of the saturated
hydrocarbons and results into harmless fatty acid droplets.
This mechanism is been influenced by level of lipase
activity (U/ml) occurring on the boundary lipid-water
interface [14].
Figure 1. Titration analysis on tested Rhodococcal isolates
2) Empirical study on lipase activity on UHT milk
The objective of this test is to evaluate efficacy of
Rhodococcal lipase action on milk. UHT milk is known
as long life product; however trace action of lipolysis on
this milk would affect the life span of the product mostly,
6 months without refrigeration [15]. Fig. 2 illustrates the
extent of lipase contamination occurred in the tested UHT
processed milk by different lipase source.
Figure 2. Observation on colour changes on UHT processed milk due to lipase activity.
Upon degradation of the milk and excessive yield of
free fatty acid, the colour of UHT milk changes from pink
(alkaline) to colourless (acidic). This phenomenon
indicates the presence high amount of free fatty acid thus,
the milk turns acidic. As per theory, in case spoilage
although the lipase producing microbe has been destroyed
yet the activity of lipase still remains determining the life
span of the dairy product. As the milk portion has been
degraded to free fatty acid, it tends to develop bitter –off
flavors by free fatty acid yielded [16]. Therefore, this first
stage outcome proves that the extracted crude lipase
practically applicable in food products. 3) Effects of different grinding time on cell lysates
with best lipase activity
Apart from analysis on bacteria growth conditions and
nutrients, another essential aspect influencing the enzyme
analysis would be the grinding time required. Being the
most potential lipase producer, demonstration in Fig 3
shows analysis on crude lysates of R.NAM391 obtained
at different grinding time. Rhodococcus sp. evidently has
been proven to own rigid cell envelope, where the
internal osmotic pressure in the microorganism cytoplasm
regarded as another resistance to disintegrate cell wall
from the intracellular content [17]. Hence, evaluation on
level of grinding for high operable quality of lipase
enzyme esteemed to be necessary.
Figure 3. Effect of grinding time on lipase activity of R.NAM319
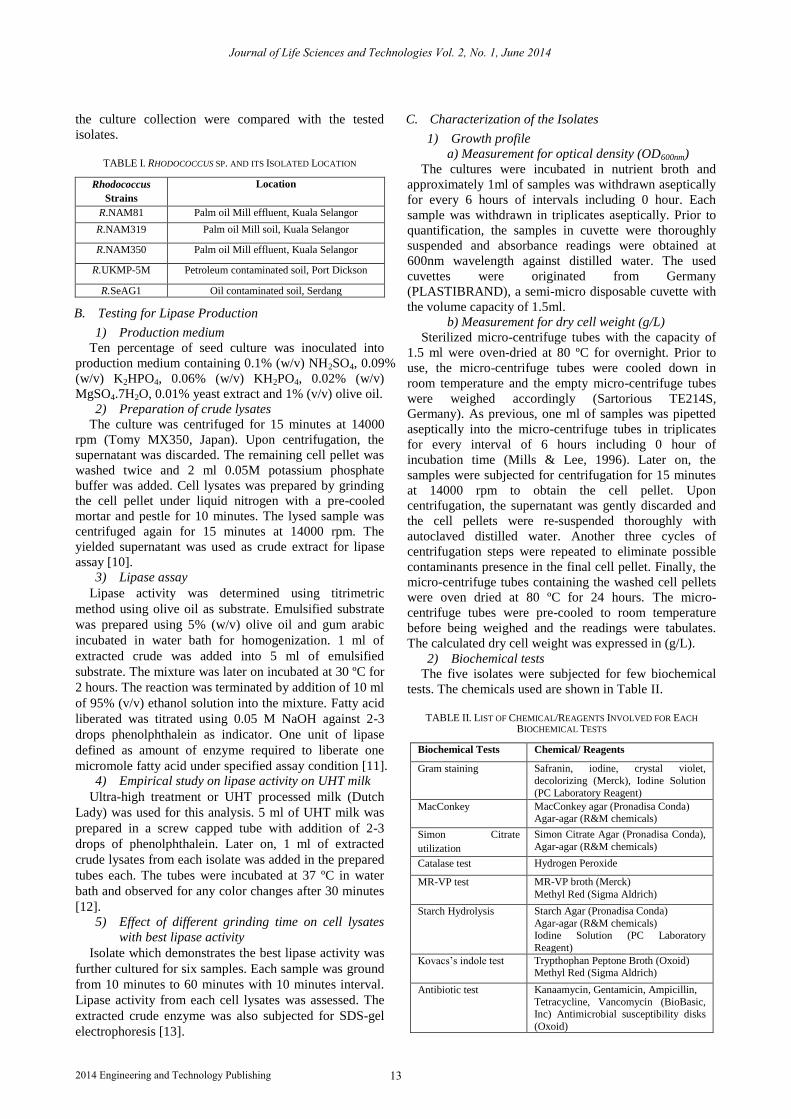
Cryogenic grinding persistently earned great protein
content when Rhodococcus cell membrane is disrupted. It
was understood that constant non-stop force from
grinding will overcome barrier like “capsular slime”
properties produced by this bacteria. Convincingly,
structure of lipase enzyme made of α/β hydrolase folds, is
also recognised as a strong robust catalyst with greater
capacity to withstand harsh conditions [18]. Hence,
possibilities for denaturation or damage on this specified
protein during consistent grinding can be least expected. Extracted protein content proportionally increases by
grinding time for R.NAM319. Therefore, lipase activity
noticeably increases as time grows, indicating increasing
level amount of hydrolysis end product from prolong
lysis. In agreement to the previous finding, extracted cell
lysates resulted in visible protein bands for every 10
minutes of interval grinding. Due to the increasing
protein recovery for each grinding intervals, darker
protein bands were observed from 1st lane to 6
th lane as
shown in Fig 4.
142014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014
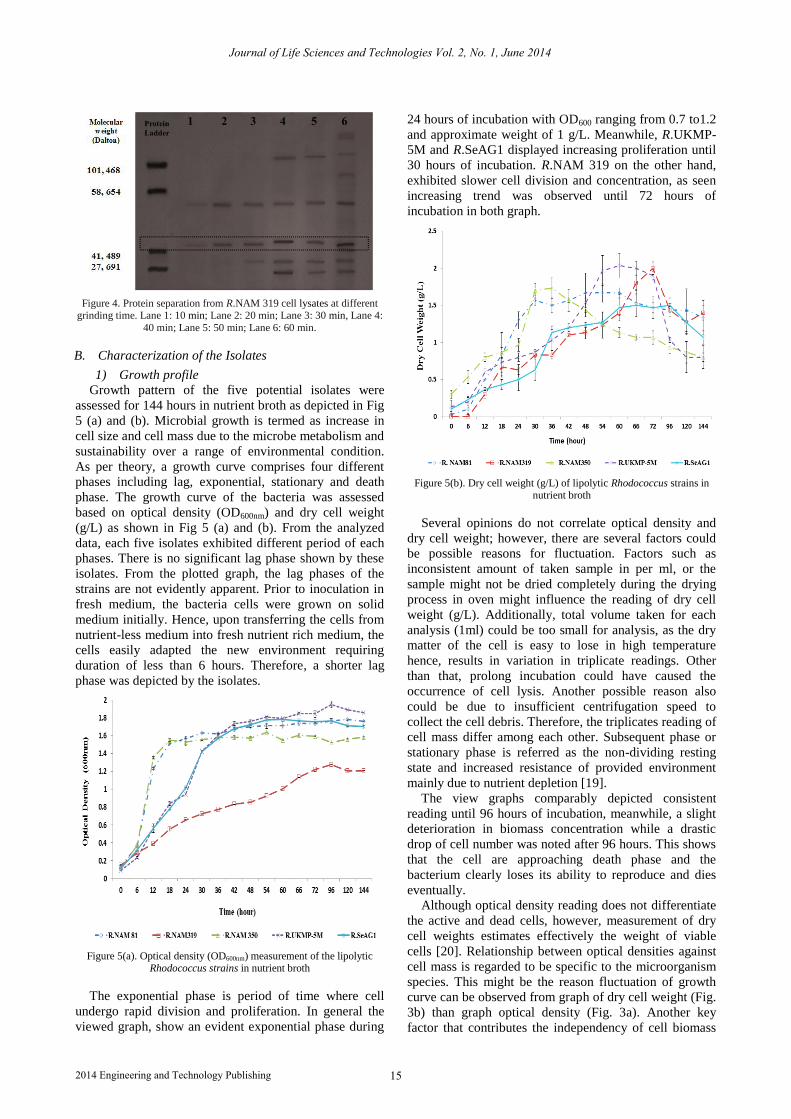
Figure 4. Protein separation from R.NAM 319 cell lysates at different grinding time. Lane 1: 10 min; Lane 2: 20 min; Lane 3: 30 min, Lane 4:
40 min; Lane 5: 50 min; Lane 6: 60 min.
B. Characterization of the Isolates
1) Growth profile
Growth pattern of the five potential isolates were
assessed for 144 hours in nutrient broth as depicted in Fig
5 (a) and (b). Microbial growth is termed as increase in
cell size and cell mass due to the microbe metabolism and
sustainability over a range of environmental condition.
As per theory, a growth curve comprises four different
phases including lag, exponential, stationary and death
phase. The growth curve of the bacteria was assessed
based on optical density (OD600nm) and dry cell weight
(g/L) as shown in Fig 5 (a) and (b). From the analyzed
data, each five isolates exhibited different period of each
phases. There is no significant lag phase shown by these
isolates. From the plotted graph, the lag phases of the
strains are not evidently apparent. Prior to inoculation in
fresh medium, the bacteria cells were grown on solid
medium initially. Hence, upon transferring the cells from
nutrient-less medium into fresh nutrient rich medium, the
cells easily adapted the new environment requiring
duration of less than 6 hours. Therefore, a shorter lag
phase was depicted by the isolates.
Figure 5(a). Optical density (OD600nm) measurement of the lipolytic Rhodococcus strains in nutrient broth
The exponential phase is period of time where cell
undergo rapid division and proliferation. In general the
viewed graph, show an evident exponential phase during
24 hours of incubation with OD600 ranging from 0.7 to1.2
and approximate weight of 1 g/L. Meanwhile, R.UKMP-
5M and R.SeAG1 displayed increasing proliferation until
30 hours of incubation. R.NAM 319 on the other hand,
exhibited slower cell division and concentration, as seen
increasing trend was observed until 72 hours of
incubation in both graph.
Figure 5(b). Dry cell weight (g/L) of lipolytic Rhodococcus strains in nutrient broth
Several opinions do not correlate optical density and
dry cell weight; however, there are several factors could
be possible reasons for fluctuation. Factors such as
inconsistent amount of taken sample in per ml, or the
sample might not be dried completely during the drying
process in oven might influence the reading of dry cell
weight (g/L). Additionally, total volume taken for each
analysis (1ml) could be too small for analysis, as the dry
matter of the cell is easy to lose in high temperature
hence, results in variation in triplicate readings. Other
than that, prolong incubation could have caused the
occurrence of cell lysis. Another possible reason also
could be due to insufficient centrifugation speed to
collect the cell debris. Therefore, the triplicates reading of
cell mass differ among each other. Subsequent phase or
stationary phase is referred as the non-dividing resting
state and increased resistance of provided environment
mainly due to nutrient depletion [19].
The view graphs comparably depicted consistent
reading until 96 hours of incubation, meanwhile, a slight
deterioration in biomass concentration while a drastic
drop of cell number was noted after 96 hours. This shows
that the cell are approaching death phase and the
bacterium clearly loses its ability to reproduce and dies
eventually.
Although optical density reading does not differentiate
the active and dead cells, however, measurement of dry
cell weights estimates effectively the weight of viable
cells [20]. Relationship between optical densities against
cell mass is regarded to be specific to the microorganism
species. This might be the reason fluctuation of growth
curve can be observed from graph of dry cell weight (Fig.
3b) than graph optical density (Fig. 3a). Another key
factor that contributes the independency of cell biomass
152014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014

and optical density is the robust wavelength [21]. Various
range of pigment profile displayed by the cell in the
medium lead to diverse absorption spectra. Hence,
suspension samples with similar absorption ability results
in reduced deviation during optical density analysis.
Figure 6. Morphology of Rhodococcus colonies (a) R.NAM81, (b) R.NAM319, (c) R.NAM350, (d) R.UKMP-5M, (e) R.SeAG1, (f)
Positive culture.
2) Biochemical test
a) Gram staining
Cell envelope of Rhodococcus cell is being capsulated
with large branch chain lipid known as mycolic acids.
Additionally, this bacterium is well-denoted for its tough
arabinogalactan (AG) cell wall polysaccharide anchoring
the formed mycolic acids [22]. This tough and rigid
layers cell envelope retains the crystal violet dye stained
initially during this test though stages of decolorizing and
washing has been done subsequently. Thus, the stained
cells appear to be purple under microscope.
Figure 7. Gram staining observation under microscope 4 x100
b) Mac conkey agar test
As both selective and differential medium,
composition of Mac Conkey agar includes mixture of bile
salt and crystal violet which inhibits growth of gram
positive bacteria. In agreement to gram staining result,
the streaked bacteria could not tolerate bile salts and
crystal violet in the agar, thus do not exhibits any growth
on the agar. This confirms that the streaked strains belong
to gram positive category.
c) Simmon citrate agar test
This differential medium is composed of sodium
citrate as the sole carbon source and ammonium ion as
the only nitrogen source. Bromothymol blue indicator has
been incorporated in the agar, where the medium remains
green for pH <7 but changes to blue when pH >7.6.
Therefore, upon utilization of citrate carbon source for
energy, an alkaline by- product formed will initiate colour
changes from green to blue. In accordance to Table III,
few lipolytic Rhodococcus isolates namely R.NAM81,
R.NAM319 and R.SeAG1 changes colour to blue,
indicating their ability to utilize citrate as the carbon
source, though few other Rhodococcus isolates resulted
no changes. This indicates although same species, their
adaptability can differs according to the metabolism of
the strain [23].
Figure 8. Observation on simmon citrate agar
d) Catalase test
In this test, the ability of tested microorganism to
produce catalase enzyme were tested. From this test, the
specimen can be known either living in aerobic or
anaerobic condition. Biologically, hydrogen peroxide is
the by- product of respiration and in case of accumulation,
it endangers the cell. Therefore, to prevent cell damages,
any cell that uses oxygen or lives in presence of oxygen,
produces catalase enzyme to breakdown hydrogen
peroxide into water and oxygen. From Fig. 9, it was clear
that the lipolytic Rhodococcus isolates are aerobic
bacteria meanwhile; the tested positive control is an
anaerobic microorganism.
Figure 9. Catalase test on microscope slide
e) MR-VP test
MR-VP broth is a differential medium, which
differentiate specimen based on acid or acetylmethyl
162014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014

production. The broth composed of peptone, buffers,
glucose or dextrose. In the process of converting the
dextrose or glucose to pyruvate, the cultured strain uses
mixed acidic pathway which yields acidic end such as
lactic acid, formic acid and acetic. Pertaining Table III, it
was evident that all the cultured strains do not produce
acidic product in their metabolism, thus conversion of
dextrose to pyruvate do not occur in these bacteria
metabolism.
TABLE III. RESULTS OF DIFFERENT BIOCHEMICAL TESTS CONDUCTED ON EACH RHODOCOCCUS ISOLATE
Isolate Biochemical Test
Gram
staining
Mac
Conkey
Simon
Citrate
Catalase
test
MR-VP test Starch
hydrolysis
Kovacs
indole
R.NAM81 + - + + - - -
R.NAM319 + - + + - - -
R.NAM350 + - - + - - -
R.UKMP-5M + - - + - - -
R.SeAG1 + - + + - - -
Positive control + - - - - - - “+” indicates positive changes occurred during the test; “-” indicates no changes have been detected during the test. Positive control= Bacillus subtilis
f) Starch hydrolysis
This test is used to differentiate the ability of specimen
to produce enzyme known as α-amylase or amino-1,6-
glucosidase. Starch is a macro nutrient which is hard to
be absorbed into the cell. Hence, in order to breakdown
starch into smaller subunits, the specimen produces
exoenzymes to initiate breakdown. A clearing around the
streaked colony after addition of iodine on the plate
indicates starch has been hydrolysed. However, none of
the streaked strains could produce exoenzymes
effectively thus, no changes seen on starch agar.
g) Kovac’s indole test
This test reveals ability of a specimen to degrade the
amino acid tryptophanane produce indole as the end
product. This is the chemical reaction involved:
Tryptophan+water=indole + pyruvic acid +ammonia
In presence of indole after the cultivation, top of the
broth will form a “cherry-red” ring upon addition of
Kovac’s reagent. As shown in the equation, formation of
indole is the result of reductive deamination from
tryptophan via the intermediate molecule indole pyruvic
acid. However, the cultivated isolates resulted in negative
observation, indicating these strains are not able to
breakdown tryptophan amino acids [24].
h) Antibiotic resistance test
Disk diffusion method was applied to test the
susceptibility of the bacteria on few antibiotics on
Mueller Hinton agar plate. Table IV represents the mean
of inhibition zone diameter measurement in mm for
tested antibiotics discs.
This test would determine the most effective antibiotic
for treatment against particular type of microbe infection.
Size of inhibition zone represents degree of resistance,
and can be predicted for resistance mechanism and
resistance gene involved. Rhodococcus sp. relevantly, is
known as uncommon opportunistic pathogen in human
especially for immunocompromised patient [25].
Formation of an inhibition zone cannot be automatically
concluded as susceptible yet; the diameters of zone need
to be compared to standard zone inhibition chart [26].
Table V are the subsequent interpretations of Table IV.
TABLE IV. RESULTS FOR ANTIBIOTIC RESISTANCE TEST ON CHOSEN LIPOLYTIC RHODOCOCCUS STRAINS
Isolate Mean of inhibition zone (mm)
Tet (30μg) Vanco (30μg) Gent (10μg) Kana (30 μg) Amp (10μg)
R.NAM81 25.17a ± 0.76 18.33b ± 0.76 13.17c ± 1.04 14.83d ± 0.29 13.67e ± 0.76
R.NAM319 20.83a ± 0.29 15.5c ± 0.5 12d ± 0.86 15.67 b ± 0.76 7.33e ± 0.58
R.NAM350 16 ± 0.5 13.83 ± 0.29 13.33 ± 0.58 14.17 ± 0.29 9 ± 0.5
R.UKMP-5M 27.5 ± 0.86 24.83 ± 0.76 13.83 ± 0.29 19.5 ± 0.5 17.83± 0.29
R.SeAG1 22.83 ± 0.29 25.67 ± 0.58 24.17 ± 0.29 20.67 ± 0.58 25.67 ± 0.58
+ve control 17.17 ± 0.76 15.5 ± 0.5 12.5 ± 0.5 16.7 ± 0.58 9.33 ± 0.76
TABLE V. INTERPRETATION ON ZONE OF INHIBITION AS PER THE STANDARD CHART
Isolate Mean of inhibition zone (mm)
Tet (30μg) Vanco (30μg) Gent (10μg) Kana (30 μg) Amp (10μg)
R.NAM81 S S I I I
R.NAM319 S S R I R
R.NAM350 I I I I R
R.UKMP-5M S S I S S
R.SeAG1 S S S S S
+ve control I I I I R
S= susceptible, I= Intermediate; R= resistant
172014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014

It was understood that the formed inhibition zone
largely depends on several factors such as degree of
susceptibility of bacteria and rate of growth of the
inoculum. Evident to few studies of antibacterial
activities on Rhodococcus sp., the obtained outcome was
in agreement to the reported results. As an example,
generally most of Rhodococcus sp. is reported to be
susceptible to vancomycin [25]. There is also a report
stated that Rhodococcus equi potrays moderate
susceptibility to ampicillin, tetracycline and gentamicin
[24]. Meanwhile in current finding, susceptibility of
different Rhodococcus isolates seems to be different for
each tested antibiotics, suggesting variable anti-microbial
properties displayed by different Rhodococcus isolates.
IV. CONCLUSION
This initial stage of study confirms that locally isolated
Rhodococcus isolates are potent to be lipase producers.
As discussed, R.NAM 391 being the most promising
isolate, reveal that as grinding time of the cell increases,
lipase activity of the cell lysates would increase. Since
the tested isolates are tentatively characterized,
biochemical tests resembles that the tested isolates
resemble characteristics of Rhodococcus sp. as per
analysed in literature. This initial stage of study serves as
an introductory study on locally available isolates, which
has not been fully exploited for lipase producing
capacities.
ACKNOWLEDGMENT
The authors would like to express gratitude to Ministry
of Science, Technology and Innovation (MOSTI),
Malaysia (3090104000(G)) and the Selangor State
Government, Malaysia for the financial assistance. We
would like to acknowledge Institute Bio-IT, Universiti
Selangor, for providing us necessary lab facilities
throughout this investigation.
REFERENCES
[1] C. O. Ibrahim, “Development of applications of industrial
enzymes from Malaysian indigenous microbial sources,”
Bioresource technology, vol. 9, pp. 4572-4582, 2008. [2] R. Aravindan, P. Anbumathi, and T. Viruthagiri, “Lipase
applications in food industry,” Indian Journal of Biotechnology,
vol. 6, pp. 141-158, 2007. [3] F. Hasan, A. Shah, and A. Hameed, “Industrial applications of
microbial lipases,” Enzyme and Microbial Technology, vol. 39, pp.
235-251, 2006. [4] B. Andualema and A. Gessesse, “Microbial lipases and their
industrial applications: Review,” Biotechnology, vol. 3, pp. 100-
118, 2012. [5] P. Boonmahome and W. Mongkoltharanuk, “Lipase producing
bacterium and its enzyme,” Journal of Life Sciences and
Technology, vol. 1, pp. 196-200, 2013. [6] V. Suneetha and Z. A. Khan, “Actinomycetes: Sources for soil
enzymes,” Soil Enzymology, vol. 22, pp. 259-269, 2011.
[7] M. A. Hernandez, W. M. Mohn, E. Martinenz, E. Rost, A. F. Alvarez, and H. M. Alvarez, “Biosynthesis of storage compounds
by rhodococcus jostii RHA1and global identification of genes
involved in their metabolism,” BMC Genomics, vol. 9, pp. 1-14, 2008.
[8] M. Kosa and A. J. Ragauskas, “Bioconversion of lignin model
compounds with oleaginous rhodococci,” Applied Microbioloy and Biotechnology, vol. 93, pp. 891-900, 2012.
[9] E. H. Lee, J. Kim, K. S. Cho, Y. G. Ahn, and G. S. Hwang,
“Degradation of hexane and other recalcitrant hydrocarbons by a
novel isolate, Rhodococcus sp. EH831,” Environmental Science
and Pollution Research International, vol. 17, pp. 64-77, 2010. [10] J. Nagarajan, N. M. Nawawi, and A. L. Ibrahim, “Rhodococcus
UKMP-5M, an endogenous lipase producing actinomycete from
Peninsular Malaysia,” Biologia, vol. 69, pp. 123-132, 2014. [11] K. Selvam, B. Vishnupriya, and S. C. Bose, “Screening and
quantification of marine actinomycetes producing industrial
enzymes amylase, cellulose, lipase from south coast of India,” International Journal of Pharmaceutical and Biological Archieves,
vol. 2, pp. 1481-1485, 2010.
[12] W. S. Versaw, S. L. Cuppet, D. D. Winters, and L. E. Williams, “An improved calorimetric assay for bacterial lipase in nonfat dry
milk,” Journal of Food Science, vol. 6, pp. 1557-1559, 1989.
[13] U. K. Laemmli, “Cleavage of structure proteins during the assembly of head,” Nature, vol. 227, pp. 680-685, 1970.
[14] C. S. Karigar and S. S. Rao, “Role of microbial enzymes in
bioremediation of pollutants: A review,” Enzyme Research, 2011. [15] P. Andrewes et al., “Detection of lipase in skim and whole milk
powders using triheptanoin as a substrate,” International Dairy
Journal, vol. 17, pp. 587-595, 2007. [16] C. W. Fox, G. L Chrisope, and R. T. Marshall, “Incidence and
identification of phospholipases C-producing bacteria in fresh and
spoiled homogenization milk,” Journal of Dairy Science, vol. 59, pp. 1857-1864, 1976.
[17] Y. Chisti and M. M. Young, “Disruption of microbial cells for
intracellular products,” Enzyme Microbe Technology, vol. 8, pp. 195-204, 1986.
[18] F. Hefner and T. Norin, “Molecular modelling of lipase catalysed
reactions prediction of enantioselectivities,” Chemical and Pharmaceutical Bulletin, vol. 47, pp. 591-600, 1999.
[19] P. K. Herman, “Stationary phase in yeast,” Current Opinion in
Microbiology, vol. 5, pp. 602-607, 2002. [20] [20] B. E. G. Peters, M. W. Reiji, H. Joosten, L. G. Gorris, and M.
H. Zwietering, “Comparison of two optical-density- based
methods and a plate count method for estimation of growth parameters of bacillus cereus,” Applied and Environmental
Microbiology, vol. 76, pp.1399-1405, 2010. [21] J. A. Myers, B. S. Curtis, and W. R. Curtis, “ Improving accuracy
of cell and chromophore concentration measurements using
optical density,” BMC Biophysics, 2012.
[22] I. C. Sutcliffe, “Characterisation of a lipomannan lipoglycan from
the mycolic acid containing actinomycete dietzia maris,” Antonie
van Leeuwenhoek, vol. 78, pp. 195-201, 2000. [23] S. Harisha, An Introduction to Practical Biotechnology, 1st ed.
India: LP, 2006, pp .208.
[24] H. D. Isenberg and L. H. Sundeihm, “Indole reactions in bacteria,” Journal of Bacteriology, vol. 75, pp. 682-692, 1958.
[25] P. A. Conlin, J. Kevin, K. J. Willingham, and J. W. Oliver,
“Rhodococcus bacterimia in immunocompetent patient,” Microbiology, vol. 31, pp. 263-265, 2000.
[26] A. W. Baur, D. M. Perry, and W. M. M. Kirby, “Single-disk
antibiotic-sensitivity testing of staphylococci,” Jama internal medicine, vol. 104, pp. 208-215, 1959.
[27] T. Buckley, E. M. Manamon, and S. Standbridge, “Resistance
studies of erythromycin and rifampin for rhodococcus equi over a 10-year period,” Irish Veterinary Journal, vol. 60, pp. 728-730,
2007.
Jayesree Nagarajan is a postgraduate student in Institute Bio-IT Selangor,
Universiti Selangor, Malaysia. She obtained
her Bachelor of Science (Hons) Biotechnology from University Tunku Abdul
Rahman in 2011. Her majoring was on mycology and lipase studies during her
degree. Her current project highlights on
Rhodococcal lipases and its practical applications in industry under the supervision
of Prof. (E) Dato’ Dr. Abdul Latif Ibrahim. Recently she has published
a full length research article entitled “Rhodococcus UKMP-5M, an endogenous lipase producing from actinomycete from Peninsular
Malaysia” under Biologia, Springer publication.
182014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014

Norazah Mohammad Nawawi is a senior lecturer in Universiti
Selangor. Earlier she pursued her Masters in Universiti Kebangsaan
Malaysia and currently continuing her PhD in Universiti Putra Malaysia.
Her expertise field is on Microbiology and Molecular biology. Her current project focuses on Rhodococcus sp. and phenol hydroxylase
enzyme.
Prof. (E) Dato’ Dr. Abdul Latif Ibrahim is a senior professor in
Universiti Selangor since 2003. He started his career as a Veterinary
Assistant at the Veterinary Office in Selangor. Later on, he pursues a
Bachelor Degree in Veterinary Medicine at the East Pakistan. He furthered his studies at the University of Hawaii for an Msc in
microbiology. And then he successfully graduated with a PhD from
University of California USA. He also developed a vaccine for Newcastle disease during his service at Universiti Putra Malaysia. He
received Personality Academic Award from Ministry of Education in
2012.
192014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 1, June 2014