chemoselective chemical ligations of biological … · are the second least common amino acid in...
TRANSCRIPT

Chemoselective Chemical Ligations of Biological Relevance
"Bioconjugate Techniques"
2nd Edition, Greg T. Hermanson, Academic Press, 2008.
Hermanson says that a 3rd edition is underway.

Some Recent Reviews • "Functionalizing Nanoparticles with Biological Molecules: Developing Chemistries that Facilitate
Nanotechnology", Sapsford, K. E.; Algar, W. R.; Berti, L.; Gemmill, K. B.; Casey, B. J.; Oh, E.; Stewart, M. H.; Medintz, I. L. Chem Rev 2013, 113, 1904–2074.
• "Through the Looking Glass – a New World of Proteins Enabled by Chemical Synthesis", Kent, S.; Sohma, Y.; Liu, S.; Bang, D.; Pentelute, B.; Mandal, K. J. Pept. Sci. 2012, 18, 428–436.
• "From Mechanism to Mouse: A Tale of Two Bioorthogonal Reactions", (1)Sletten, E. M.; Bertozzi, C. R. Acc Chem Res 2011, 44, 666-676.
• "Bioorthogonal Chemistry for Site-Specific Labeling and Surface Immobilization of Proteins", Chen, Y.-X.; Triola, G.; Waldmann, H. Acc Chem Res 2011, 44, 762–773..
• "Bioconjugation with Strained Alkenes and Alkynes ", Debets, M. F.; van Berkel, S. S.; Dommerholt, J.; Dirks, A. T. J.; Rutjes, F. P. J. T.; van Delft, F. L. Acc Chem Res 2011, 805-815.
• "Staudinger Ligation as a Method for Bioconjugation", van Berkel, S. S.; van Eldijk, M. B.; van Hest, J. Angew Chem Int Ed Engl 2011, 50, 8806–8827..
• "Advances in Bioconjugation", Kalia, J.; Raines, R. T. Current Organic Chemistry 2010, 14, 138–147.
• "Chemoselective Ligation Techniques. Modern Applications of Time-Honored Chemistry", Tiefenbrunn, T. K. T.; Dawson, P. E. P. Biopolymers 2010, 94, 95–106.
• "Chemoselective Ligation and Modification Strategies for Peptides and Proteins", Hackenberger, C. P. R.; Schwarzer, D. Angew. Chem. Int. Ed 2008, 47, 10030–10074.
• "New Methods for Protein Bioconjugation", Francis, M. B. In Chemical Biology; Schreiber, S. L.; Kapoor, T. M.; Wess, G., Eds. Wiley-VCH Verlag GmbH: Weinheim, Germany, 2007; Vol. 1–3, pp. 593–634.
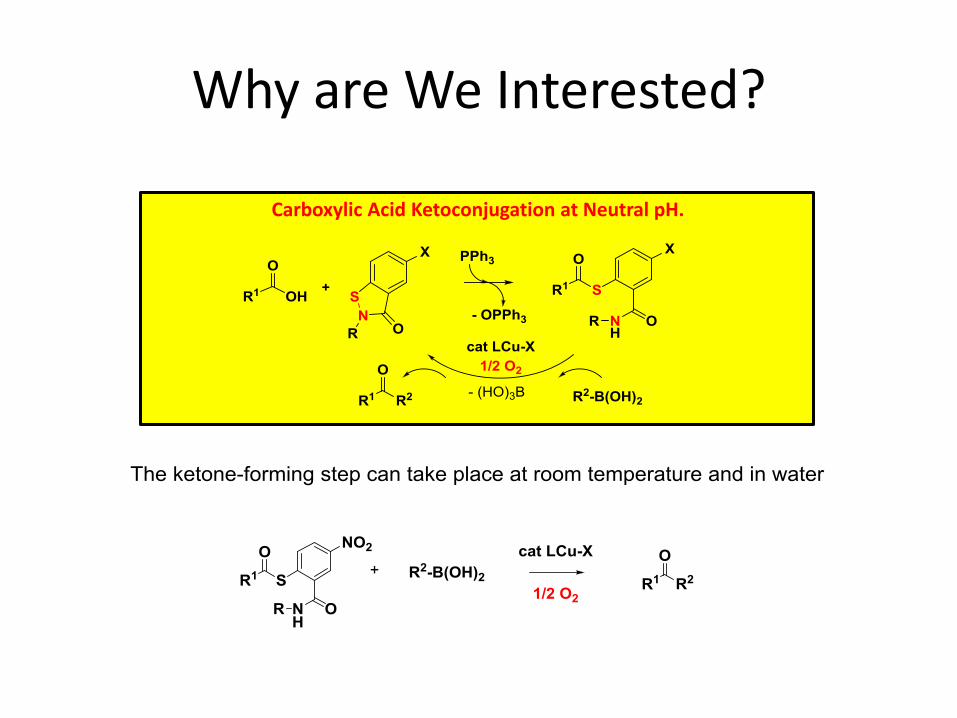
Why are We Interested?
Carboxylic Acid Ketoconjugation at Neutral pH.
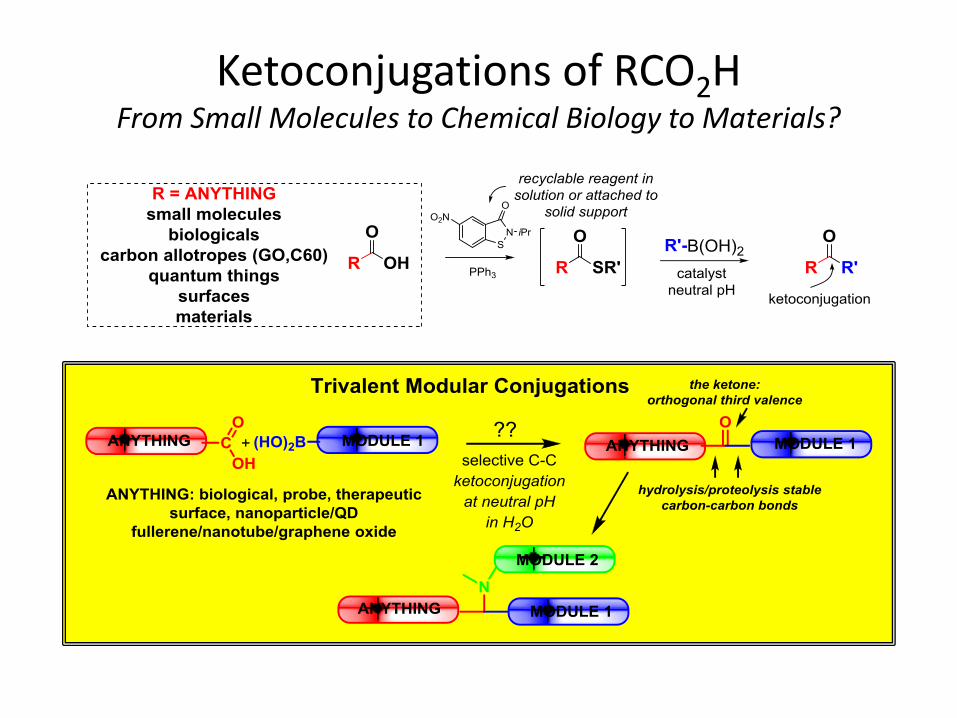
Ketoconjugations of RCO2H From Small Molecules to Chemical Biology to Materials?

Chemoselective Bioconjugation
• The chemoselective coupling of one reactive functional group specifically with another reactive functional group without side reactions:
– Between biomolecules and other biomolecules, surfaces, or particles
– Often in aqueous solutions or biological buffers and in the presence of biological material (proteins, DNA, RNA, complex carbohydrates).
– A pair of specifically "tuned" chemoselective reactants is built into the design of cross-linkers, labeling reagents, modified biomolecules, and surfaces/particles.
• Requires the rapid and selective reaction of substrates
• Reactants must be stable near neutral pH and in aqueous environments
"Advances in Bioconjugation", Kalia, J.; Raines, R. T. Current Organic Chemistry 2010, 14, 138–147.
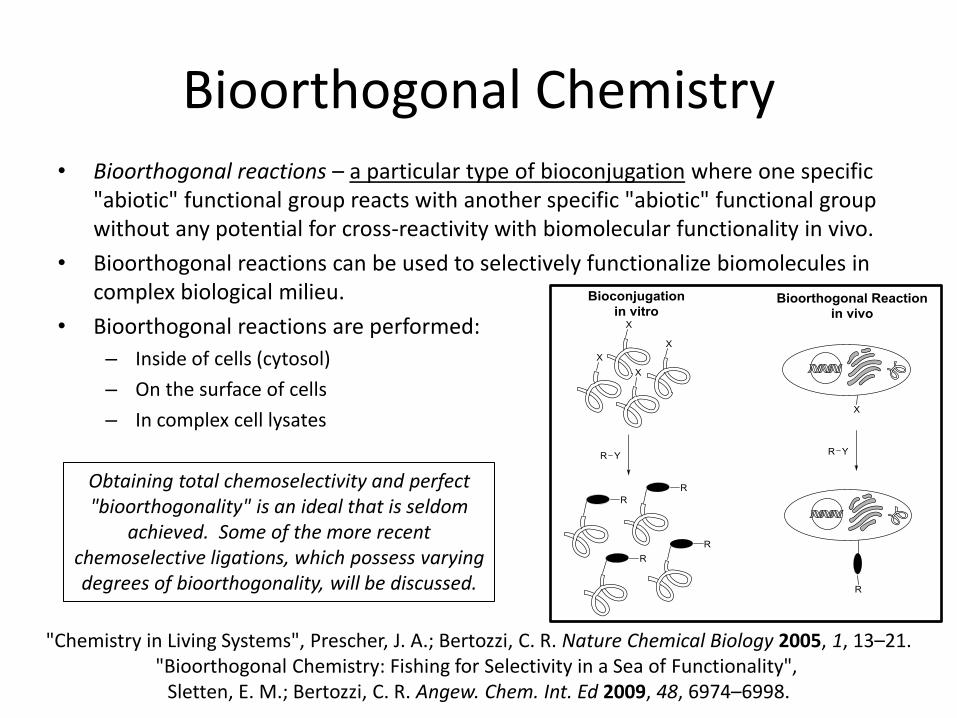
Bioorthogonal Chemistry • Bioorthogonal reactions – a particular type of bioconjugation where one specific
"abiotic" functional group reacts with another specific "abiotic" functional group without any potential for cross-reactivity with biomolecular functionality in vivo.
• Bioorthogonal reactions can be used to selectively functionalize biomolecules in complex biological milieu.
• Bioorthogonal reactions are performed:
– Inside of cells (cytosol)
– On the surface of cells
– In complex cell lysates
Obtaining total chemoselectivity and perfect "bioorthogonality" is an ideal that is seldom
achieved. Some of the more recent chemoselective ligations, which possess varying degrees of bioorthogonality, will be discussed.
"Chemistry in Living Systems", Prescher, J. A.; Bertozzi, C. R. Nature Chemical Biology 2005, 1, 13–21. "Bioorthogonal Chemistry: Fishing for Selectivity in a Sea of Functionality",
Sletten, E. M.; Bertozzi, C. R. Angew. Chem. Int. Ed 2009, 48, 6974–6998.

Native and Non-Native Methods
• Bioconjugation methods rely on a variety of specific functional group interactions to achieve selectivity. They include, but are not limited to:
– Carbon-sulfur conjugates
– Hydrazide-aldehyde and related -C=N- conjugates
– Phenylboronic acid (PBA)-salicylhydroxamic acid (SHA) conjugates
– 4+2 and 3+2 dipolar cycloadditions
– Staudinger reaction between an azide and a phosphine
• Native bioconjugation methods use naturally-occurring biological processes, such as the formation of a peptide bond between a C-terminal thiol ester and an N-terminal cysteine.
• Many bioconjugation methods, including those based on non-covalent interactions, are described in "Bioconjugate Techniques" 2nd Edition, Greg T. Hermanson, Academic Press, 2008.

Carbon—Sulfur Linkages
• Technique relies on cysteine residues, which are the second least common amino acid in natural proteins (represent 1.7% of AA residues in natural proteins) .
• Therefore, somewhat selective bioconjugation using cysteine residues can sometimes be achieved.
• Common ligating functional groups for thiols are A a-iodoamides (alkylation), B maleimides (conjugate addition), and C disulfides (disulfide exchange).
Other non-selective linkages used for bioconjugation are based on the formation of C=N bonds from hydrazides and oximes reacting with aldehyde and ketone functional groups.
See: Curr. Org. Chem. 2010, 14, 138; Bioconjugate Chem 2008, 19, 2543–2548.
Thiol linkages at Cys represent one of the oldest methods of bioconjugation.
Traditional protein conjugation chemistries have exploited the reactivity of surface-exposed nucleophilic amino acids, such as cysteine (SH) or lysine (NH2).
As a result, these methods typically result in heterogeneous mixtures of products.
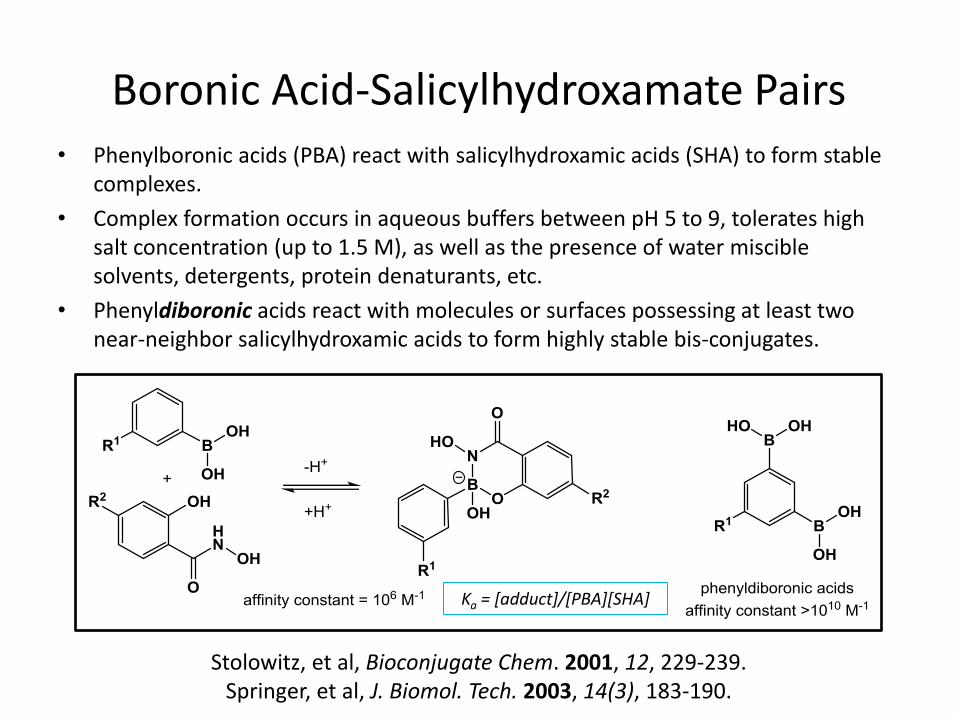
Boronic Acid-Salicylhydroxamate Pairs • Phenylboronic acids (PBA) react with salicylhydroxamic acids (SHA) to form stable
complexes.
• Complex formation occurs in aqueous buffers between pH 5 to 9, tolerates high salt concentration (up to 1.5 M), as well as the presence of water miscible solvents, detergents, protein denaturants, etc.
• Phenyldiboronic acids react with molecules or surfaces possessing at least two near-neighbor salicylhydroxamic acids to form highly stable bis-conjugates.
Stolowitz, et al, Bioconjugate Chem. 2001, 12, 229-239. Springer, et al, J. Biomol. Tech. 2003, 14(3), 183-190.
Ka = [adduct]/[PBA][SHA]
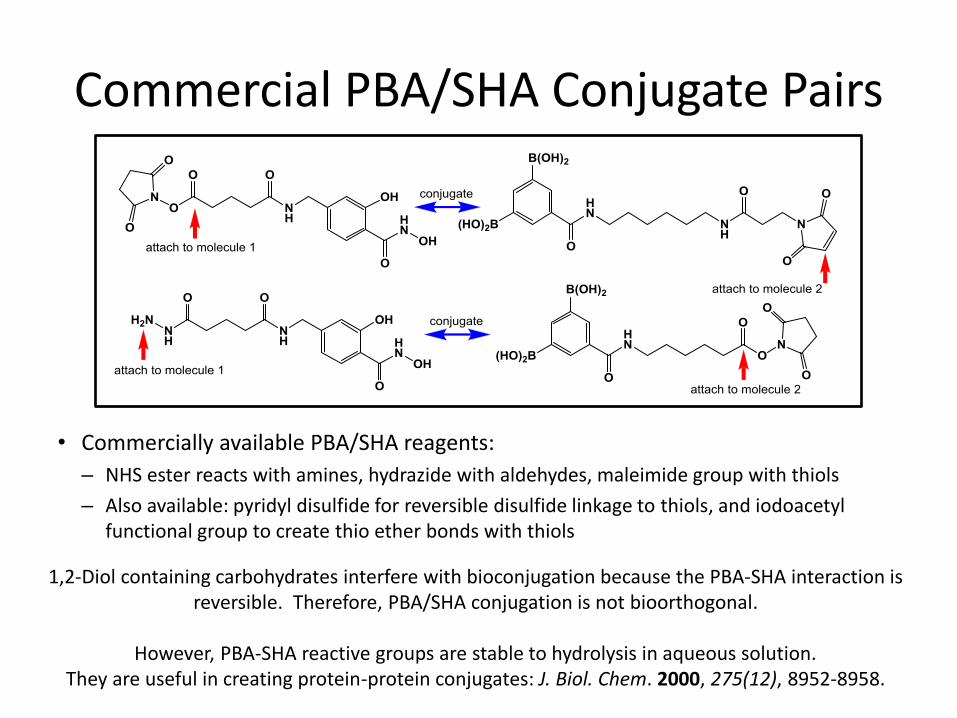
Commercial PBA/SHA Conjugate Pairs
• Commercially available PBA/SHA reagents:
– NHS ester reacts with amines, hydrazide with aldehydes, maleimide group with thiols
– Also available: pyridyl disulfide for reversible disulfide linkage to thiols, and iodoacetyl functional group to create thio ether bonds with thiols
1,2-Diol containing carbohydrates interfere with bioconjugation because the PBA-SHA interaction is reversible. Therefore, PBA/SHA conjugation is not bioorthogonal.
However, PBA-SHA reactive groups are stable to hydrolysis in aqueous solution.
They are useful in creating protein-protein conjugates: J. Biol. Chem. 2000, 275(12), 8952-8958.
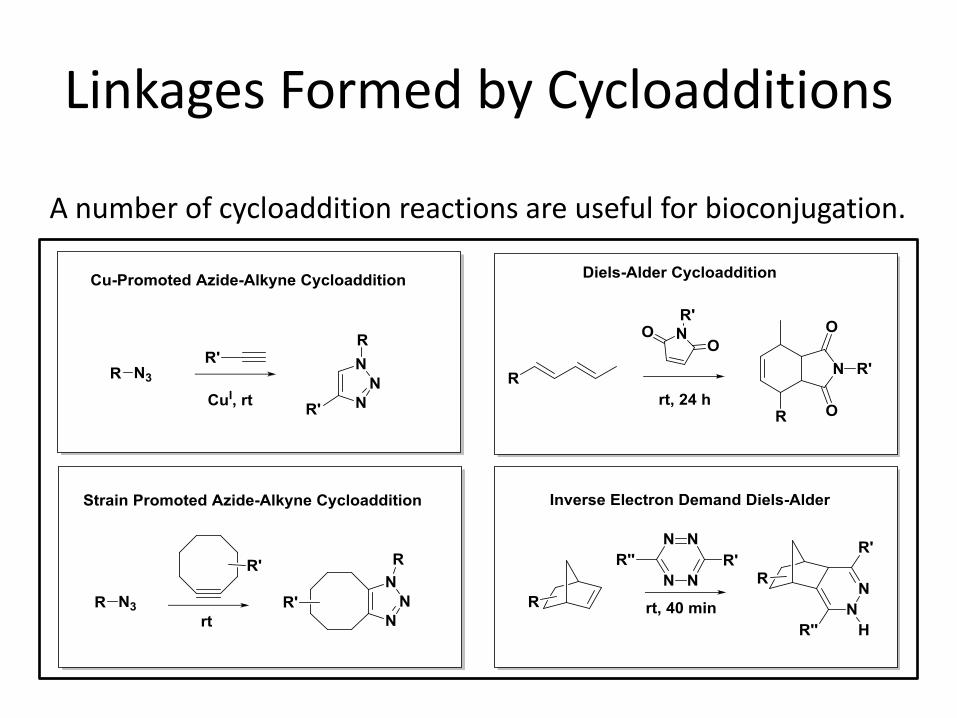
Linkages Formed by Cycloadditions
A number of cycloaddition reactions are useful for bioconjugation.
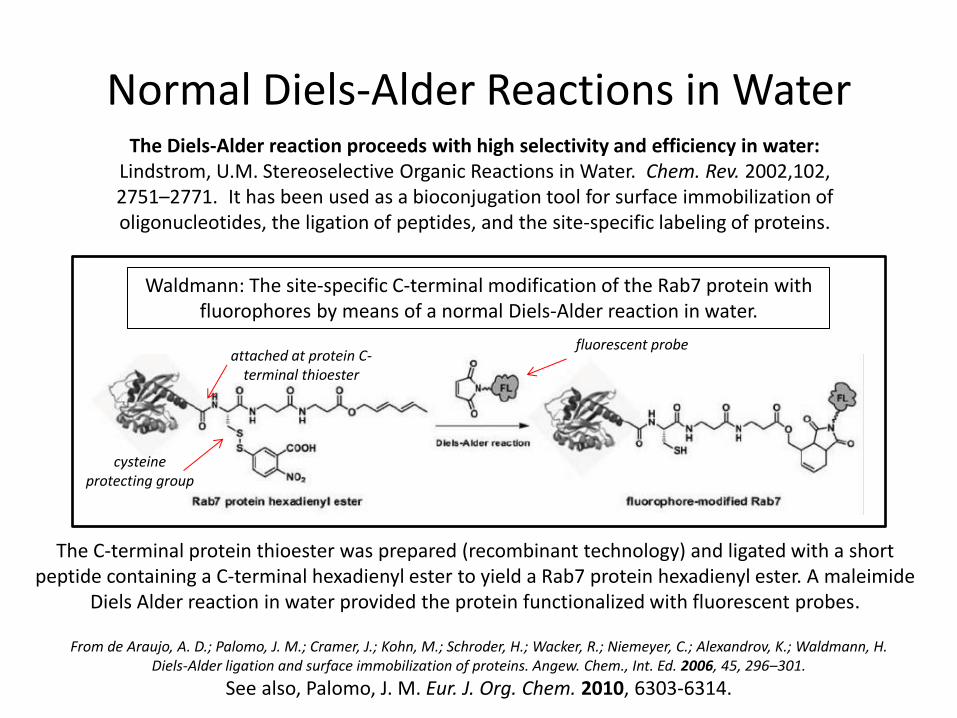
Normal Diels-Alder Reactions in Water The Diels-Alder reaction proceeds with high selectivity and efficiency in water:
Lindstrom, U.M. Stereoselective Organic Reactions in Water. Chem. Rev. 2002,102, 2751–2771. It has been used as a bioconjugation tool for surface immobilization of oligonucleotides, the ligation of peptides, and the site-specific labeling of proteins.
From de Araujo, A. D.; Palomo, J. M.; Cramer, J.; Kohn, M.; Schroder, H.; Wacker, R.; Niemeyer, C.; Alexandrov, K.; Waldmann, H. Diels-Alder ligation and surface immobilization of proteins. Angew. Chem., Int. Ed. 2006, 45, 296–301.
See also, Palomo, J. M. Eur. J. Org. Chem. 2010, 6303-6314.
Waldmann: The site-specific C-terminal modification of the Rab7 protein with fluorophores by means of a normal Diels-Alder reaction in water.
The C-terminal protein thioester was prepared (recombinant technology) and ligated with a short peptide containing a C-terminal hexadienyl ester to yield a Rab7 protein hexadienyl ester. A maleimide
Diels Alder reaction in water provided the protein functionalized with fluorescent probes.
cysteine protecting group
fluorescent probe attached at protein C-
terminal thioester
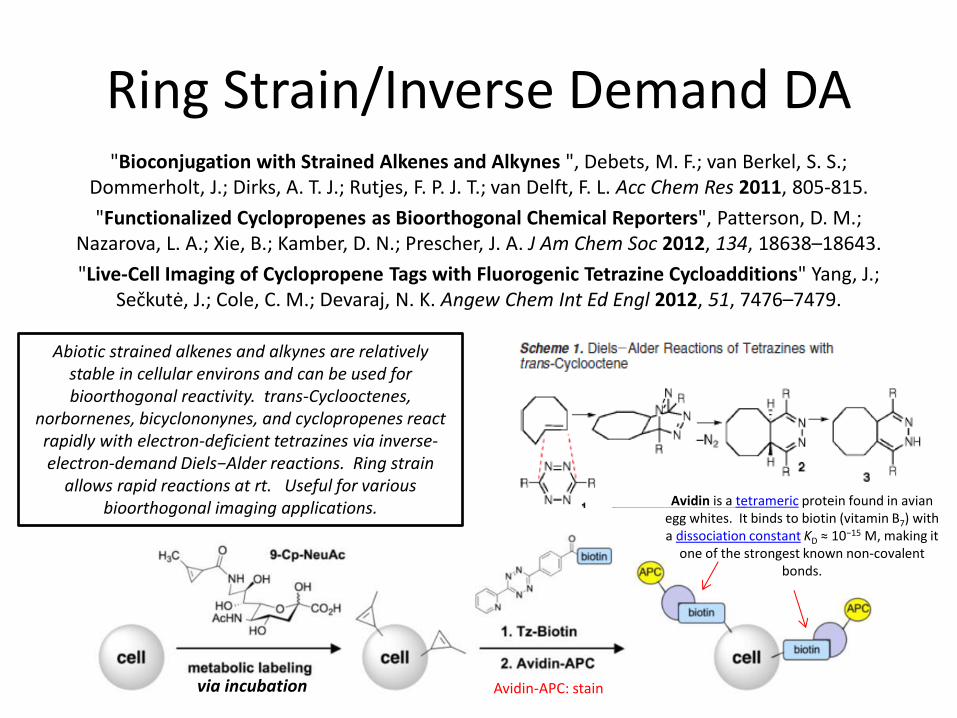
Ring Strain/Inverse Demand DA "Bioconjugation with Strained Alkenes and Alkynes ", Debets, M. F.; van Berkel, S. S.;
Dommerholt, J.; Dirks, A. T. J.; Rutjes, F. P. J. T.; van Delft, F. L. Acc Chem Res 2011, 805-815.
"Functionalized Cyclopropenes as Bioorthogonal Chemical Reporters", Patterson, D. M.; Nazarova, L. A.; Xie, B.; Kamber, D. N.; Prescher, J. A. J Am Chem Soc 2012, 134, 18638–18643.
"Live-Cell Imaging of Cyclopropene Tags with Fluorogenic Tetrazine Cycloadditions" Yang, J.; Sečkutė, J.; Cole, C. M.; Devaraj, N. K. Angew Chem Int Ed Engl 2012, 51, 7476–7479.
Abiotic strained alkenes and alkynes are relatively stable in cellular environs and can be used for bioorthogonal reactivity. trans-Cyclooctenes,
norbornenes, bicyclononynes, and cyclopropenes react rapidly with electron-deficient tetrazines via inverse-electron-demand Diels−Alder reactions. Ring strain
allows rapid reactions at rt. Useful for various bioorthogonal imaging applications.
Avidin-APC: stain via incubation
Avidin is a tetrameric protein found in avian egg whites. It binds to biotin (vitamin B7) with a dissociation constant KD ≈ 10−15 M, making it
one of the strongest known non-covalent bonds.
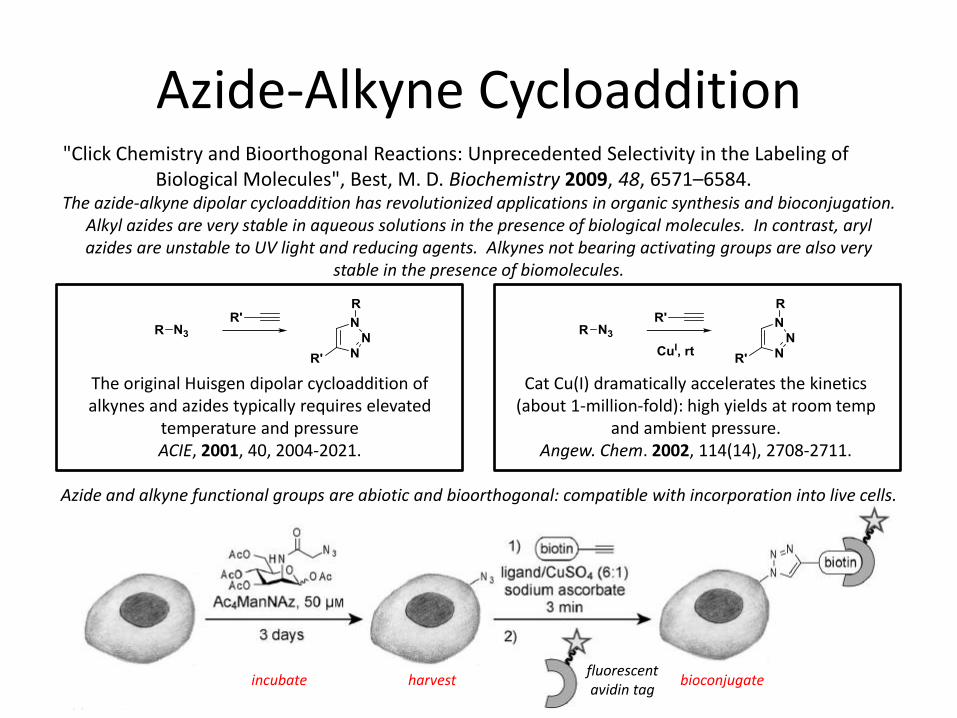
Azide-Alkyne Cycloaddition
Besanceny-Webler, C.; Jiang, H.; et al. Ang. Chem. Int. Ed. 2011, 50, 8051.
The original Huisgen dipolar cycloaddition of alkynes and azides typically requires elevated
temperature and pressure ACIE, 2001, 40, 2004-2021.
The azide-alkyne dipolar cycloaddition has revolutionized applications in organic synthesis and bioconjugation. Alkyl azides are very stable in aqueous solutions in the presence of biological molecules. In contrast, aryl azides are unstable to UV light and reducing agents. Alkynes not bearing activating groups are also very
stable in the presence of biomolecules.
Cat Cu(I) dramatically accelerates the kinetics (about 1-million-fold): high yields at room temp
and ambient pressure. Angew. Chem. 2002, 114(14), 2708-2711.
Azide and alkyne functional groups are abiotic and bioorthogonal: compatible with incorporation into live cells.
fluorescent avidin tag
"Click Chemistry and Bioorthogonal Reactions: Unprecedented Selectivity in the Labeling of Biological Molecules", Best, M. D. Biochemistry 2009, 48, 6571–6584.
incubate harvest bioconjugate
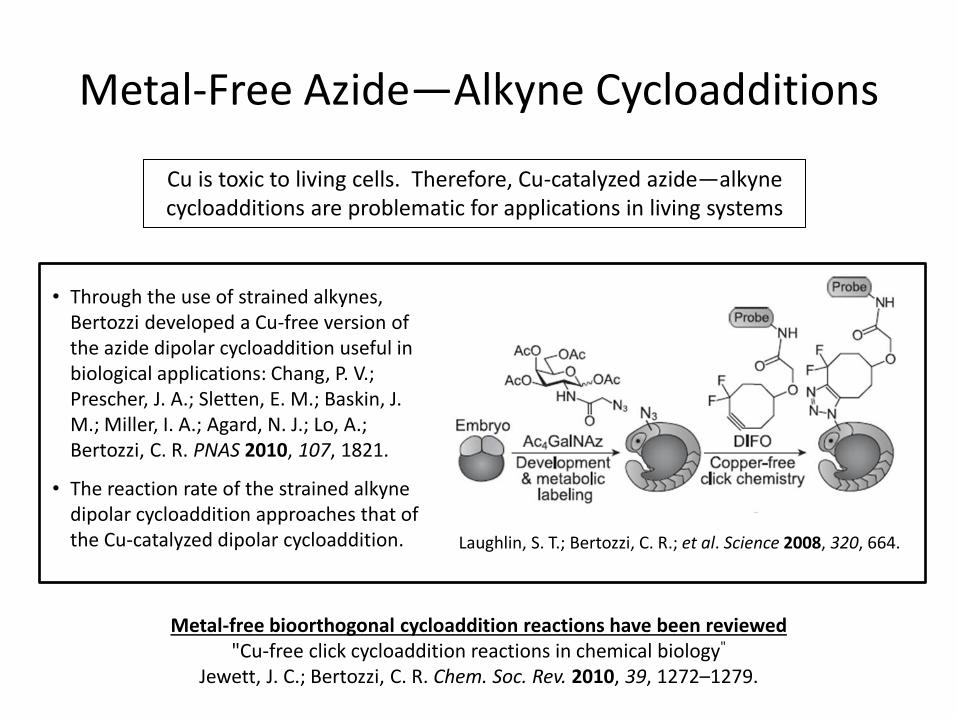
Metal-Free Azide—Alkyne Cycloadditions
• Through the use of strained alkynes, Bertozzi developed a Cu-free version of the azide dipolar cycloaddition useful in biological applications: Chang, P. V.; Prescher, J. A.; Sletten, E. M.; Baskin, J. M.; Miller, I. A.; Agard, N. J.; Lo, A.; Bertozzi, C. R. PNAS 2010, 107, 1821.
• The reaction rate of the strained alkyne dipolar cycloaddition approaches that of the Cu-catalyzed dipolar cycloaddition. Laughlin, S. T.; Bertozzi, C. R.; et al. Science 2008, 320, 664.
Metal-free bioorthogonal cycloaddition reactions have been reviewed "Cu-free click cycloaddition reactions in chemical biology"
Jewett, J. C.; Bertozzi, C. R. Chem. Soc. Rev. 2010, 39, 1272–1279.
Cu is toxic to living cells. Therefore, Cu-catalyzed azide—alkyne cycloadditions are problematic for applications in living systems

The Classical Staudinger Reaction • The Staudinger reaction (1919): the reduction of azides to amines via an
iminophosphorane.
• The intermediate iminophosphorane also reacts with a variety of electrophiles to provide amines, imines, and related compounds:
van Berkel, S. S.; van Eldijk, M. B.; van Hest, J. Angew Chem Int Ed 2011, 50, 8806–8827. Kohn, M.; Breinbauer, R. Angew. Chem. Int. Ed. 2004, 43, 3106.
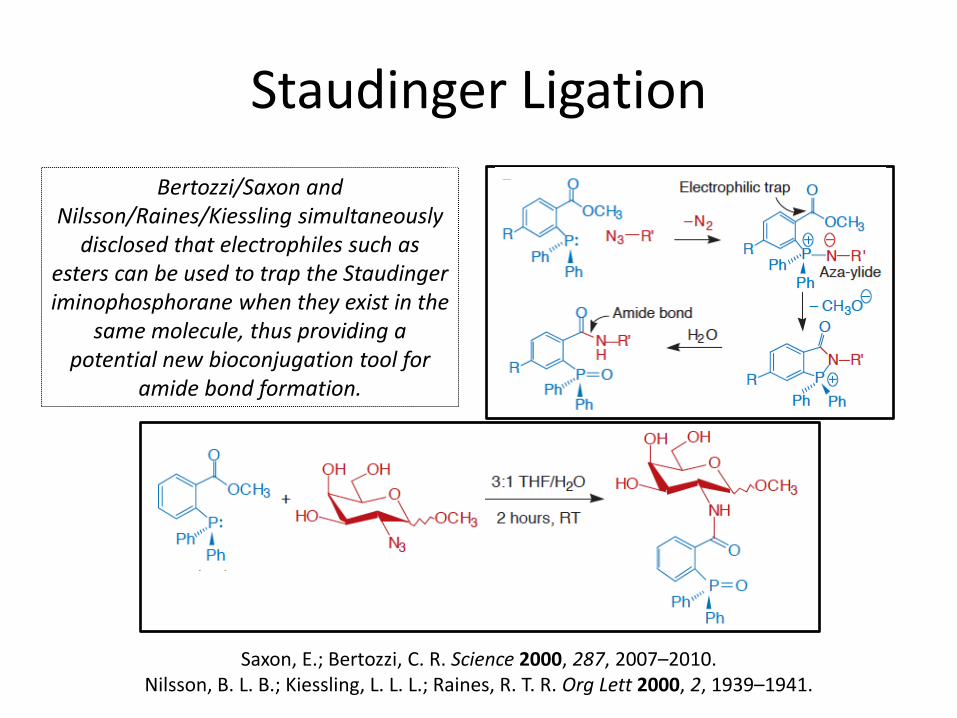
Staudinger Ligation
Bertozzi/Saxon and Nilsson/Raines/Kiessling simultaneously
disclosed that electrophiles such as esters can be used to trap the Staudinger iminophosphorane when they exist in the
same molecule, thus providing a potential new bioconjugation tool for
amide bond formation.
Saxon, E.; Bertozzi, C. R. Science 2000, 287, 2007–2010. Nilsson, B. L. B.; Kiessling, L. L. L.; Raines, R. T. R. Org Lett 2000, 2, 1939–1941.
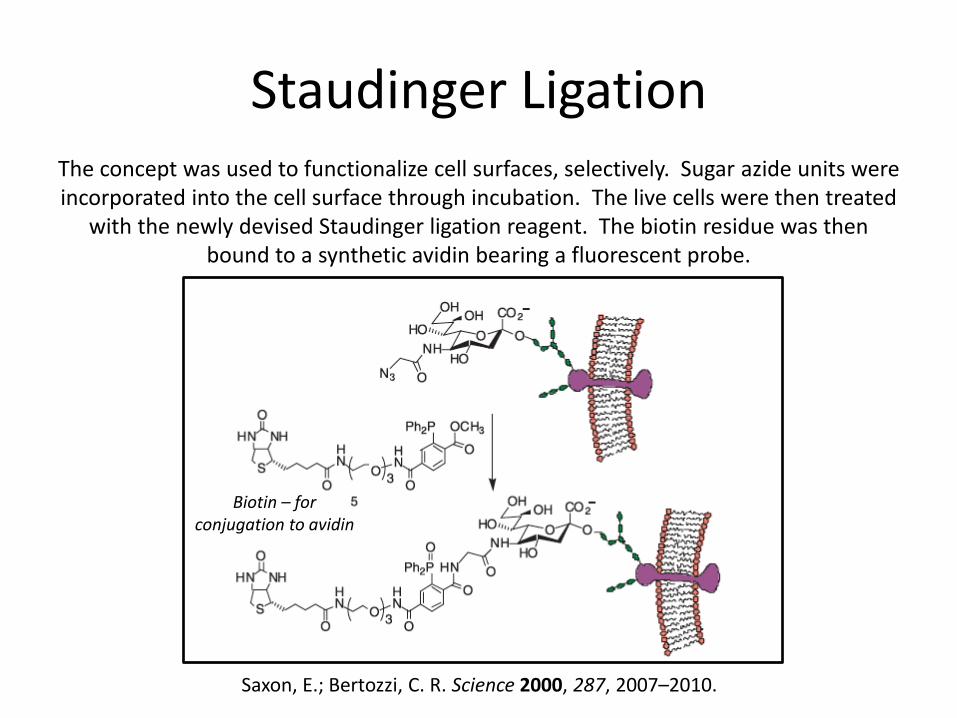
Staudinger Ligation The concept was used to functionalize cell surfaces, selectively. Sugar azide units were incorporated into the cell surface through incubation. The live cells were then treated
with the newly devised Staudinger ligation reagent. The biotin residue was then bound to a synthetic avidin bearing a fluorescent probe.
Saxon, E.; Bertozzi, C. R. Science 2000, 287, 2007–2010.
Biotin – for conjugation to avidin
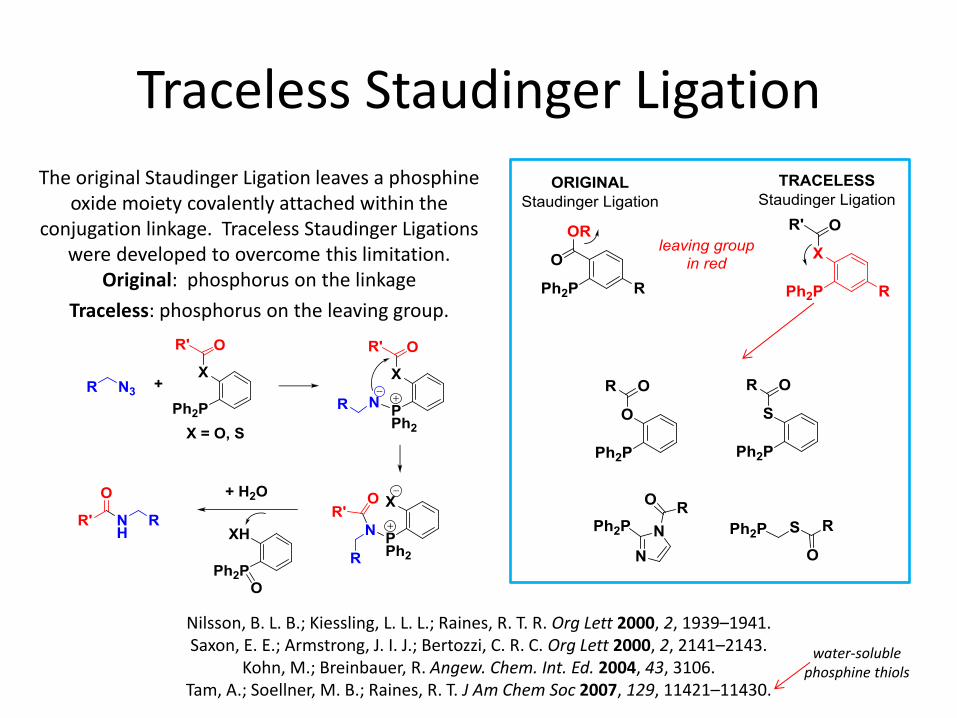
Traceless Staudinger Ligation
The original Staudinger Ligation leaves a phosphine oxide moiety covalently attached within the
conjugation linkage. Traceless Staudinger Ligations were developed to overcome this limitation.
Original: phosphorus on the linkage
Traceless: phosphorus on the leaving group.
Nilsson, B. L. B.; Kiessling, L. L. L.; Raines, R. T. R. Org Lett 2000, 2, 1939–1941. Saxon, E. E.; Armstrong, J. I. J.; Bertozzi, C. R. C. Org Lett 2000, 2, 2141–2143.
Kohn, M.; Breinbauer, R. Angew. Chem. Int. Ed. 2004, 43, 3106. Tam, A.; Soellner, M. B.; Raines, R. T. J Am Chem Soc 2007, 129, 11421–11430.
water-soluble phosphine thiols

Ligations Using Native Functionality
Native Chemical Ligation
Expressed Chemical Ligation
For an interesting historical review of peptide coupling strategies that preceded Native Chemical Ligation, see "Peptide segment coupling by prior ligation and proximity-
induced intramolecular acyl transfer", Coltart, D. M. Tetrahedron 2000, 56, 3449–3491.

Limitations of Solid Phase Peptide Synthesis
• Automated solid phase peptide synthesis (SPPS) allows the routine synthesis of peptides up to 50 amino acids in length, the size of small protein domains.
• Generating larger proteins is limited by the insolubility of protected peptide segments, by racemization that occurs in solution-phase synthesis, and by the statistical accumulation of error peptide byproducts in stepwise solid phase synthesis.
• This fundamental limitation in the length of synthetic peptides was overcome by the development of a new chemoselective ligation reactions where two or more fully unprotected peptides are linked together: Native Chemical Ligation.

Native Chemical Ligation
Dawson, P. E.; Muir, T. W.; Clark-Lewis, I.; Kent, S. B. Synthesis of proteins by native chemical ligation. Science 1994, 266, 776–779.
Dawson, P. E.; Kent, S. B. Synthesis of native proteins by chemical ligation. Annu. Rev. Biochem. 2000, 69, 923–960.
Kent, S.; Sohma, Y.; Liu, S.; Bang, D.; Pentelute, B.; Mandal, K. Through the looking glass – a new world of proteins enabled by chemical synthesis. J. Pept. Sci. 2012, 18, 428–436.
• Kent and coworkers developed native chemical ligation (NCL) as one solution to overcome SPPS limitations.
• In NCL, one unprotected peptide with an N-terminal cysteine reacts with a second unprotected peptide possessing a C-terminal thioester to form a peptide bond.
• Native Chemical Ligation proceeds via a reversible trans-thioesterification, which is followed by an irreversible nucleophilic attack of an internally positioned amine.
• Critical feature of NCL: the avoidance of any protecting groups on either fragment.
reversible irreversible
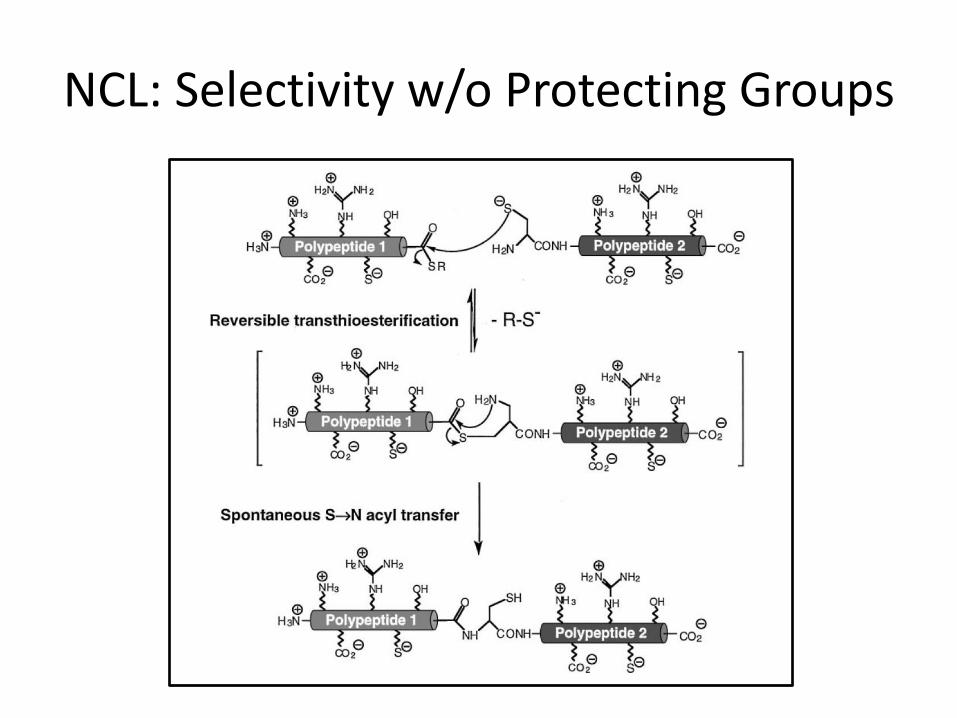
NCL: Selectivity w/o Protecting Groups
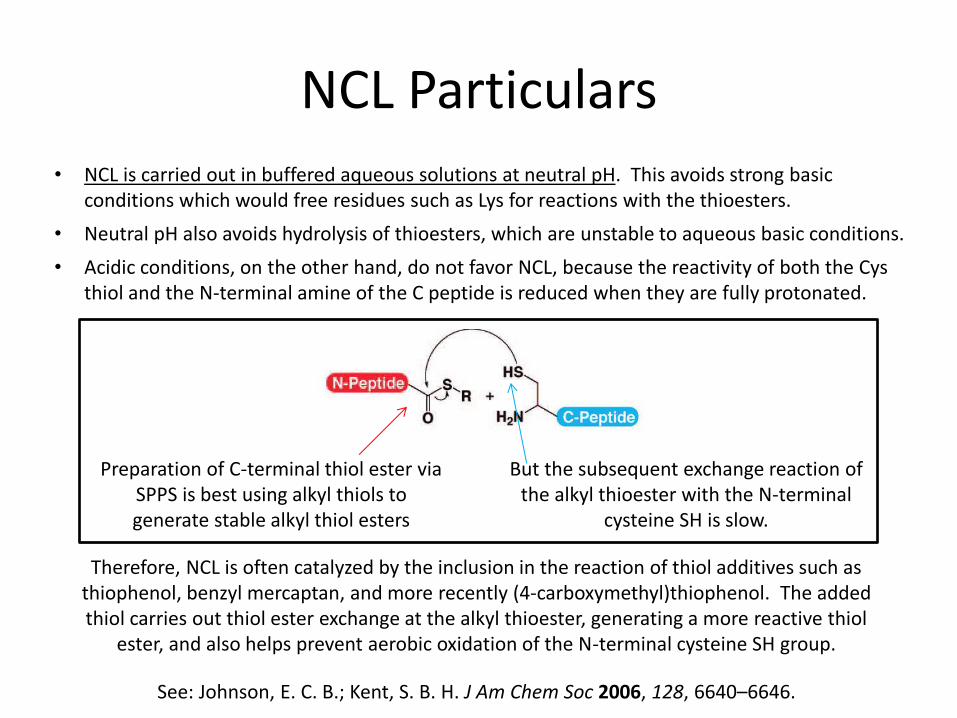
NCL Particulars
Preparation of C-terminal thiol ester via SPPS is best using alkyl thiols to generate stable alkyl thiol esters
But the subsequent exchange reaction of the alkyl thioester with the N-terminal
cysteine SH is slow.
Therefore, NCL is often catalyzed by the inclusion in the reaction of thiol additives such as thiophenol, benzyl mercaptan, and more recently (4-carboxymethyl)thiophenol. The added thiol carries out thiol ester exchange at the alkyl thioester, generating a more reactive thiol
ester, and also helps prevent aerobic oxidation of the N-terminal cysteine SH group.
See: Johnson, E. C. B.; Kent, S. B. H. J Am Chem Soc 2006, 128, 6640–6646.
• NCL is carried out in buffered aqueous solutions at neutral pH. This avoids strong basic conditions which would free residues such as Lys for reactions with the thioesters.
• Neutral pH also avoids hydrolysis of thioesters, which are unstable to aqueous basic conditions.
• Acidic conditions, on the other hand, do not favor NCL, because the reactivity of both the Cys thiol and the N-terminal amine of the C peptide is reduced when they are fully protonated.
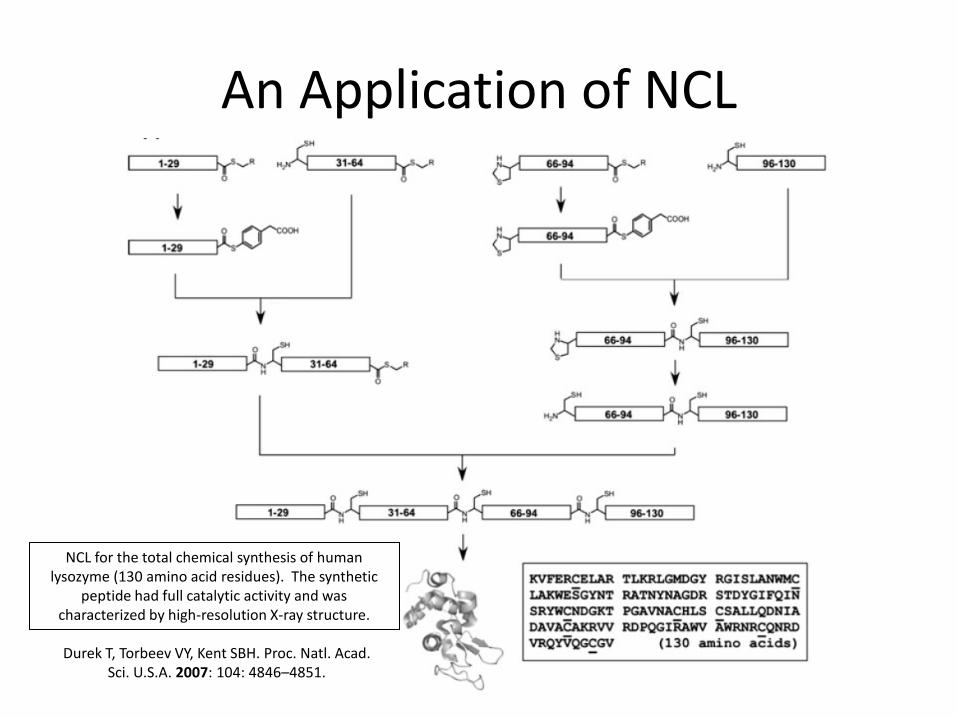
An Application of NCL
NCL for the total chemical synthesis of human lysozyme (130 amino acid residues). The synthetic
peptide had full catalytic activity and was characterized by high-resolution X-ray structure.
Durek T, Torbeev VY, Kent SBH. Proc. Natl. Acad. Sci. U.S.A. 2007: 104: 4846–4851.
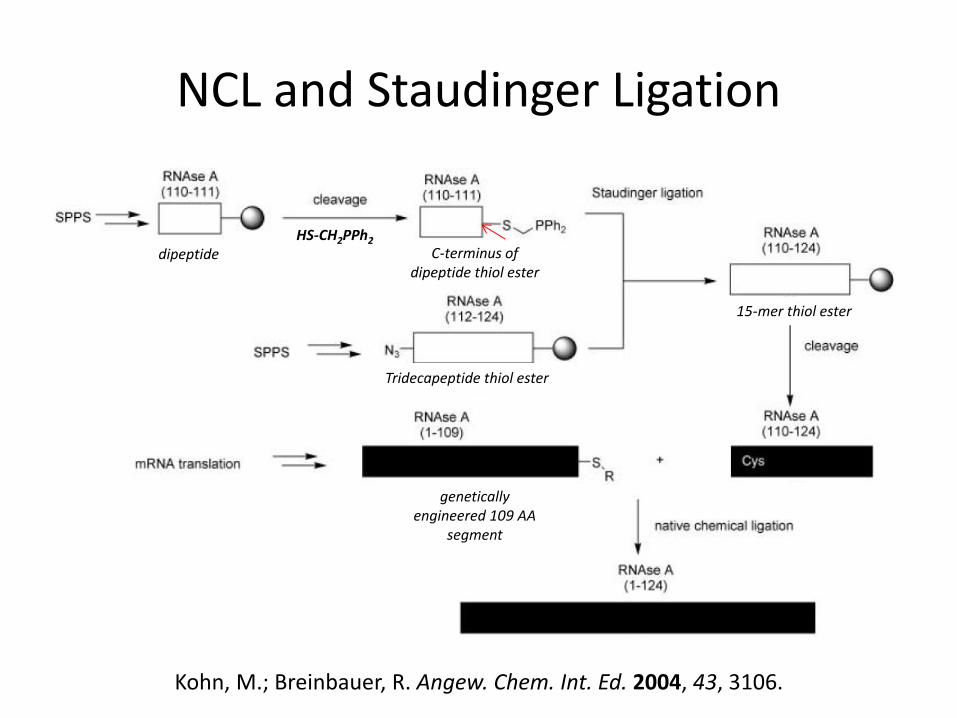
NCL and Staudinger Ligation
Kohn, M.; Breinbauer, R. Angew. Chem. Int. Ed. 2004, 43, 3106.
dipeptide C-terminus of dipeptide thiol ester
Tridecapeptide thiol ester
15-mer thiol ester
HS-CH2PPh2
genetically engineered 109 AA
segment

Reactivity Issues with NCL
Dawson and co-workers: Proc. Natl. Acad. Sci. USA 1999, 96, 10068 – 10073.
Side chains significantly affect the NCL rates of the 20 different amino acid thioesters.
Fastest: Gly thioester reacts quantitatively in less than two hours.
Slowest: thioesters containing b-branched amino acids or proline did not result in a quantitative conversion after two days.
Sterically hindered thioesters however can be ligated by using more reactive thiols.

Influence of the Thioester on NCL
Johnson, E. C. B.; Kent, S. B. H. J Am Chem Soc 2006, 128, 6640–6646.
• Alkyl thioesters are less reactive than aryl thioesters.
• Alkyl thioesters are good for SPPS synthesis and handling, but NCL requires a rapid thiol–thioester exchange, which is much more efficient with aryl thioesters.
• Therefore, peptide thioesters are commonly synthesized as their alkyl derivatives, often with sodium-2-mercaptoethanesulfonate (MESNA), and are then converted into the corresponding aryl thioesters in situ by the addition of an excess of an aryl thiol, such as the water-soluble (4-carboxymethyl)thiophenol.
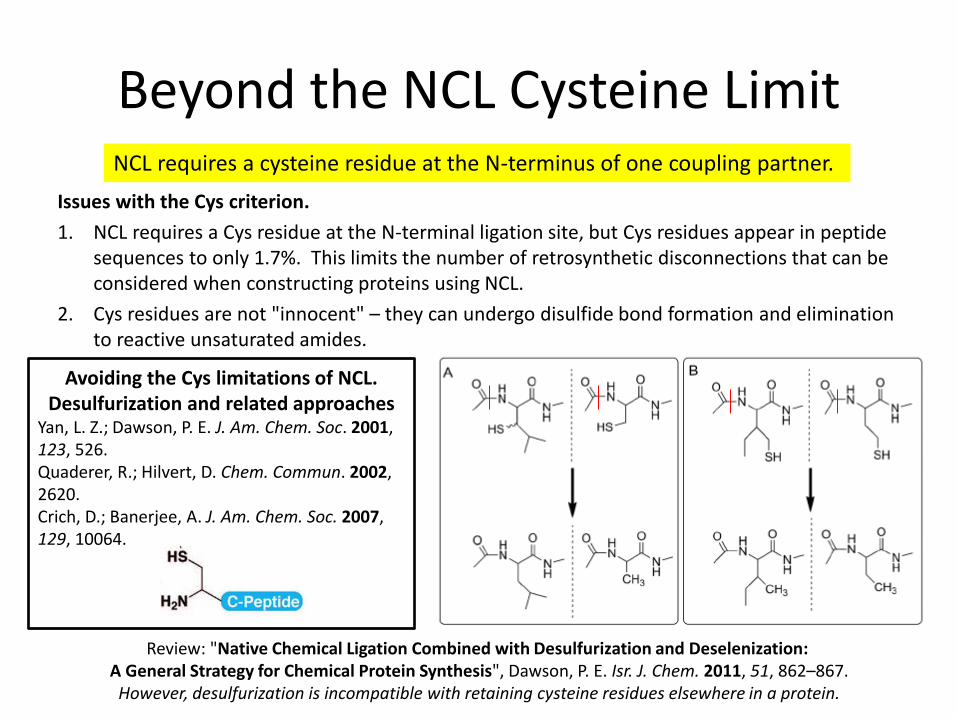
Beyond the NCL Cysteine Limit
Issues with the Cys criterion.
1. NCL requires a Cys residue at the N-terminal ligation site, but Cys residues appear in peptide sequences to only 1.7%. This limits the number of retrosynthetic disconnections that can be considered when constructing proteins using NCL.
2. Cys residues are not "innocent" – they can undergo disulfide bond formation and elimination to reactive unsaturated amides.
NCL requires a cysteine residue at the N-terminus of one coupling partner.
Avoiding the Cys limitations of NCL. Desulfurization and related approaches
Yan, L. Z.; Dawson, P. E. J. Am. Chem. Soc. 2001, 123, 526. Quaderer, R.; Hilvert, D. Chem. Commun. 2002, 2620. Crich, D.; Banerjee, A. J. Am. Chem. Soc. 2007, 129, 10064.
Review: "Native Chemical Ligation Combined with Desulfurization and Deselenization: A General Strategy for Chemical Protein Synthesis", Dawson, P. E. Isr. J. Chem. 2011, 51, 862–867.
However, desulfurization is incompatible with retaining cysteine residues elsewhere in a protein.
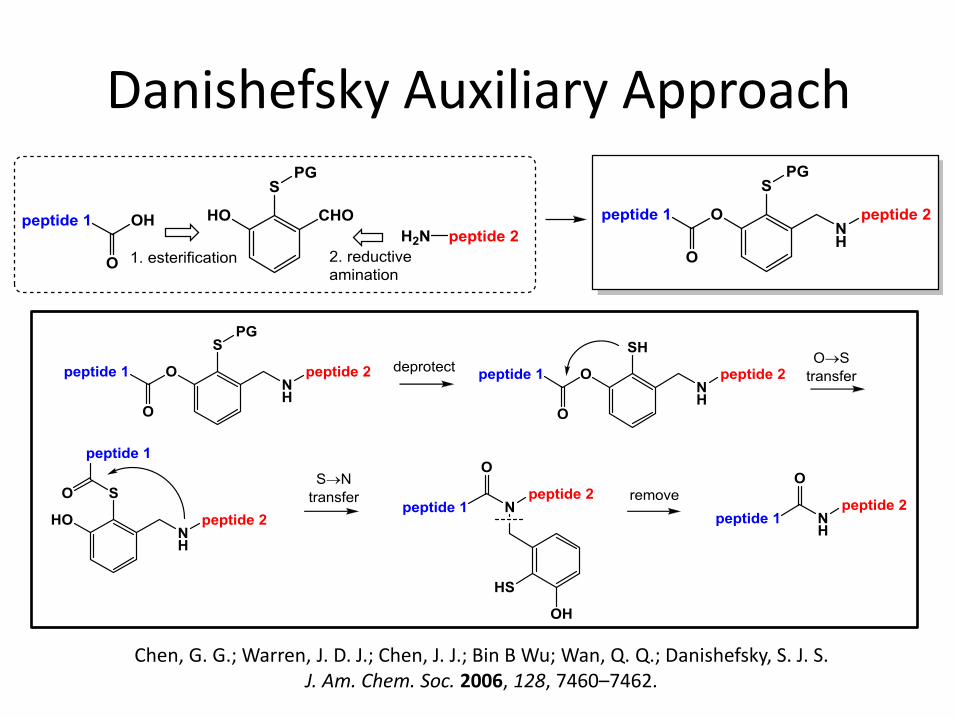
Danishefsky Auxiliary Approach
Chen, G. G.; Warren, J. D. J.; Chen, J. J.; Bin B Wu; Wan, Q. Q.; Danishefsky, S. J. S. J. Am. Chem. Soc. 2006, 128, 7460–7462.
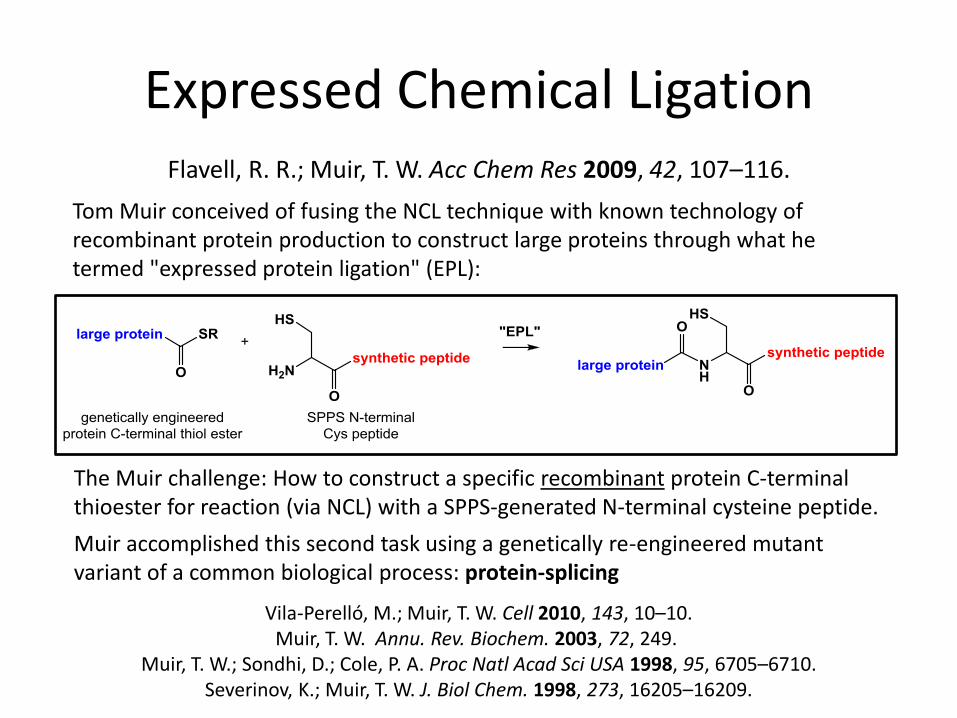
Expressed Chemical Ligation
Vila-Perelló, M.; Muir, T. W. Cell 2010, 143, 10–10. Muir, T. W. Annu. Rev. Biochem. 2003, 72, 249.
Muir, T. W.; Sondhi, D.; Cole, P. A. Proc Natl Acad Sci USA 1998, 95, 6705–6710. Severinov, K.; Muir, T. W. J. Biol Chem. 1998, 273, 16205–16209.
Flavell, R. R.; Muir, T. W. Acc Chem Res 2009, 42, 107–116.
Tom Muir conceived of fusing the NCL technique with known technology of recombinant protein production to construct large proteins through what he termed "expressed protein ligation" (EPL):
The Muir challenge: How to construct a specific recombinant protein C-terminal thioester for reaction (via NCL) with a SPPS-generated N-terminal cysteine peptide.
Muir accomplished this second task using a genetically re-engineered mutant variant of a common biological process: protein-splicing
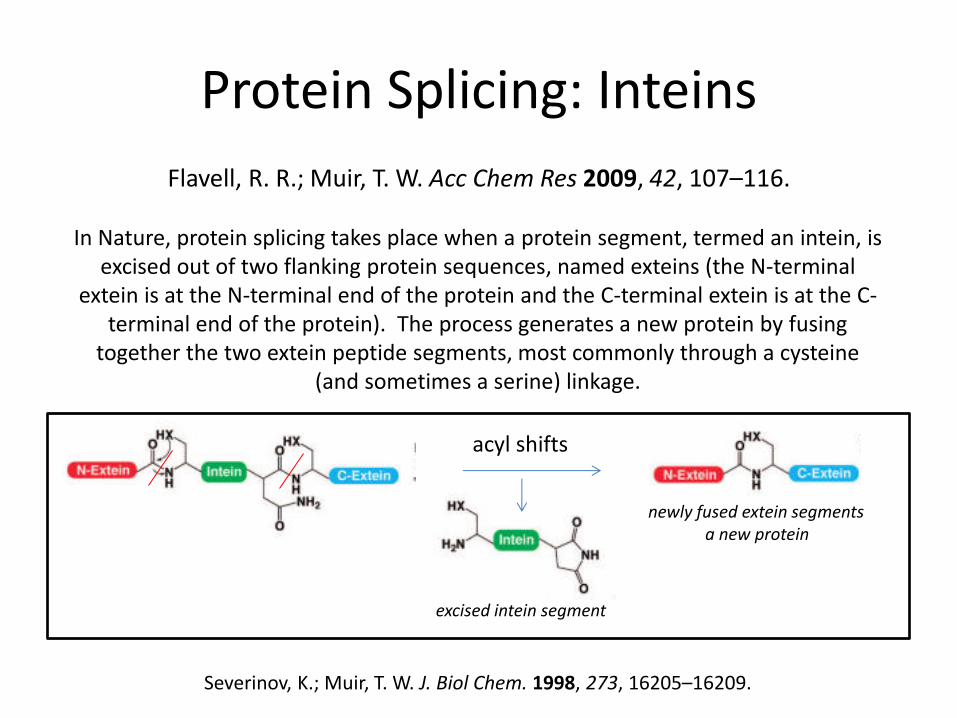
Protein Splicing: Inteins
Severinov, K.; Muir, T. W. J. Biol Chem. 1998, 273, 16205–16209.
Flavell, R. R.; Muir, T. W. Acc Chem Res 2009, 42, 107–116.
In Nature, protein splicing takes place when a protein segment, termed an intein, is excised out of two flanking protein sequences, named exteins (the N-terminal
extein is at the N-terminal end of the protein and the C-terminal extein is at the C-terminal end of the protein). The process generates a new protein by fusing
together the two extein peptide segments, most commonly through a cysteine (and sometimes a serine) linkage.
acyl shifts
excised intein segment
newly fused extein segments a new protein
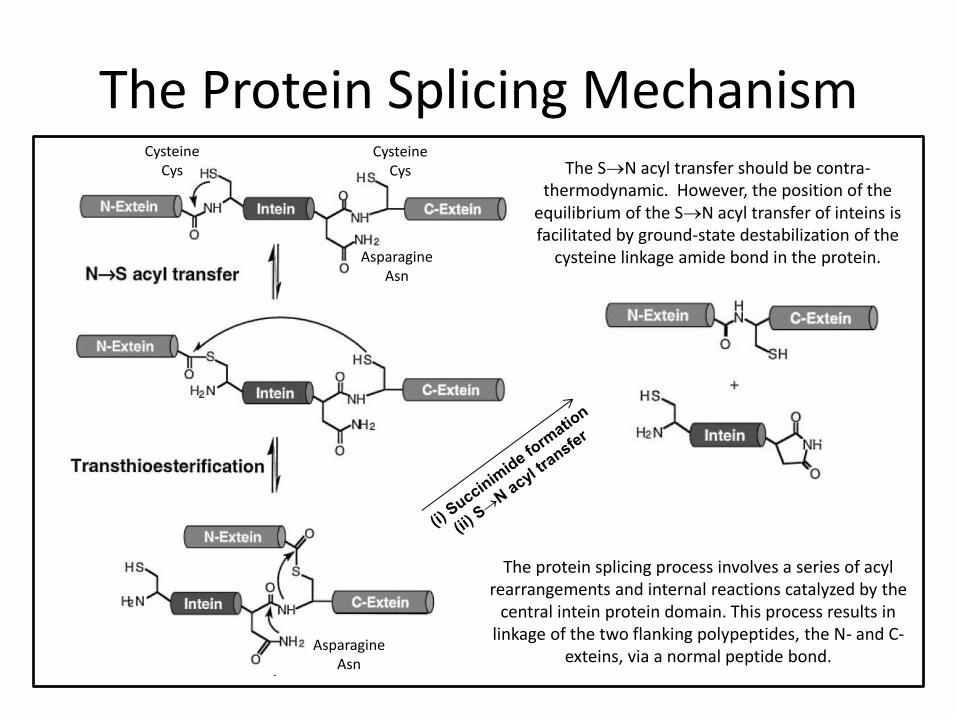
The Protein Splicing Mechanism
The protein splicing process involves a series of acyl rearrangements and internal reactions catalyzed by the
central intein protein domain. This process results in linkage of the two flanking polypeptides, the N- and C-
exteins, via a normal peptide bond.
Asparagine Asn
Asparagine Asn
The SN acyl transfer should be contra-thermodynamic. However, the position of the
equilibrium of the SN acyl transfer of inteins is facilitated by ground-state destabilization of the
cysteine linkage amide bond in the protein.
Cysteine Cys
Cysteine Cys
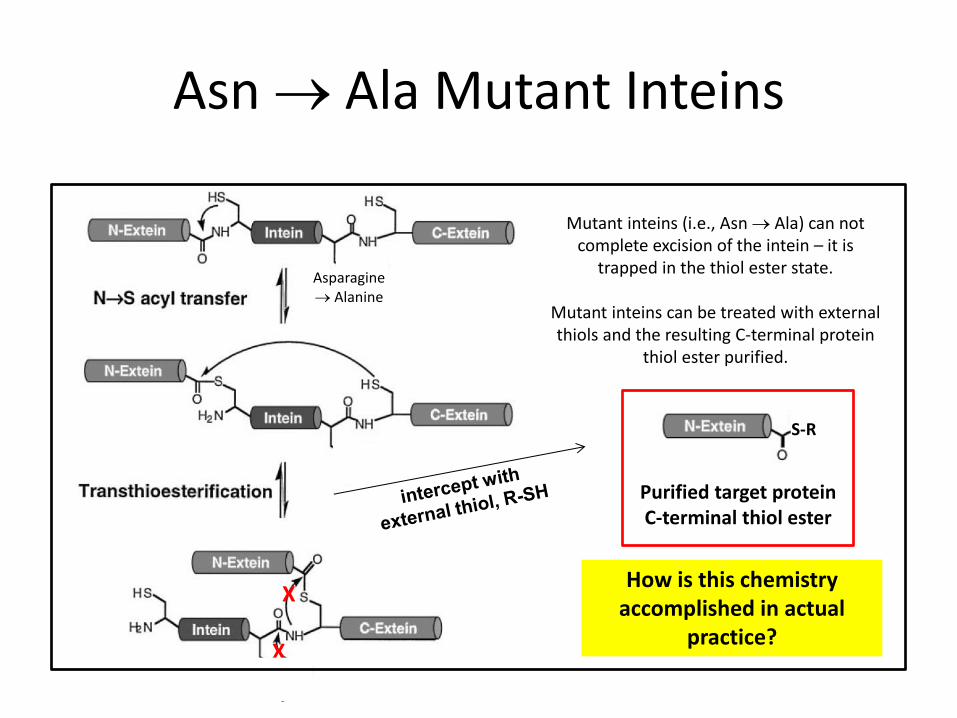
Asn Ala Mutant Inteins
Mutant inteins (i.e., Asn Ala) can not complete excision of the intein – it is
trapped in the thiol ester state.
Mutant inteins can be treated with external thiols and the resulting C-terminal protein
thiol ester purified.
Asparagine Alanine
X
X
S-R
Purified target protein C-terminal thiol ester
How is this chemistry accomplished in actual
practice?
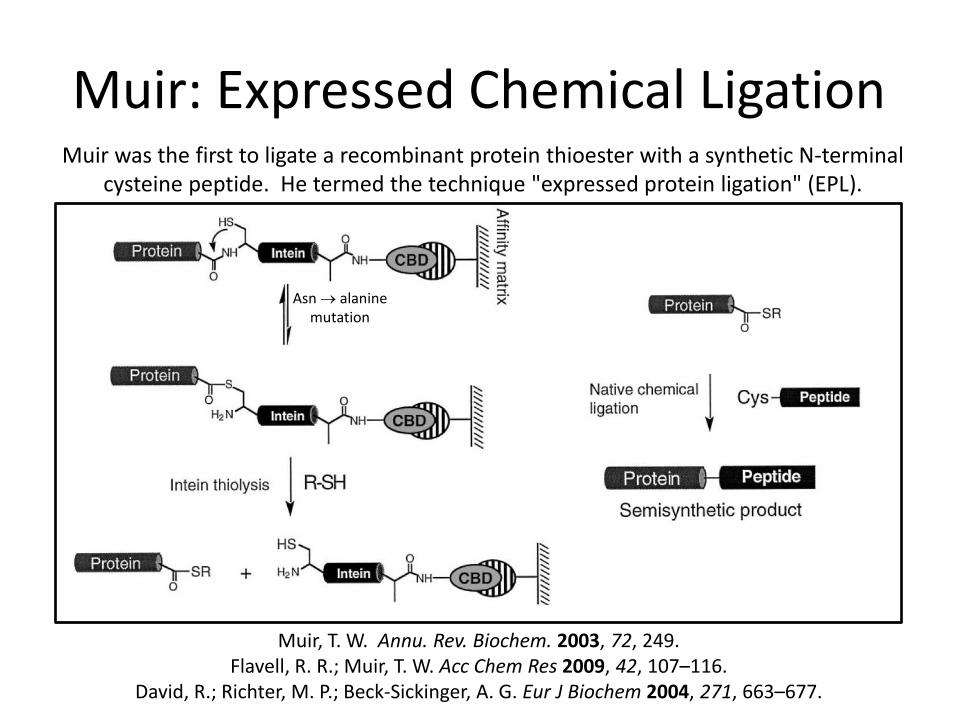
Muir: Expressed Chemical Ligation
Asn alanine mutation
Muir, T. W. Annu. Rev. Biochem. 2003, 72, 249. Flavell, R. R.; Muir, T. W. Acc Chem Res 2009, 42, 107–116.
David, R.; Richter, M. P.; Beck-Sickinger, A. G. Eur J Biochem 2004, 271, 663–677.
Muir was the first to ligate a recombinant protein thioester with a synthetic N-terminal cysteine peptide. He termed the technique "expressed protein ligation" (EPL).
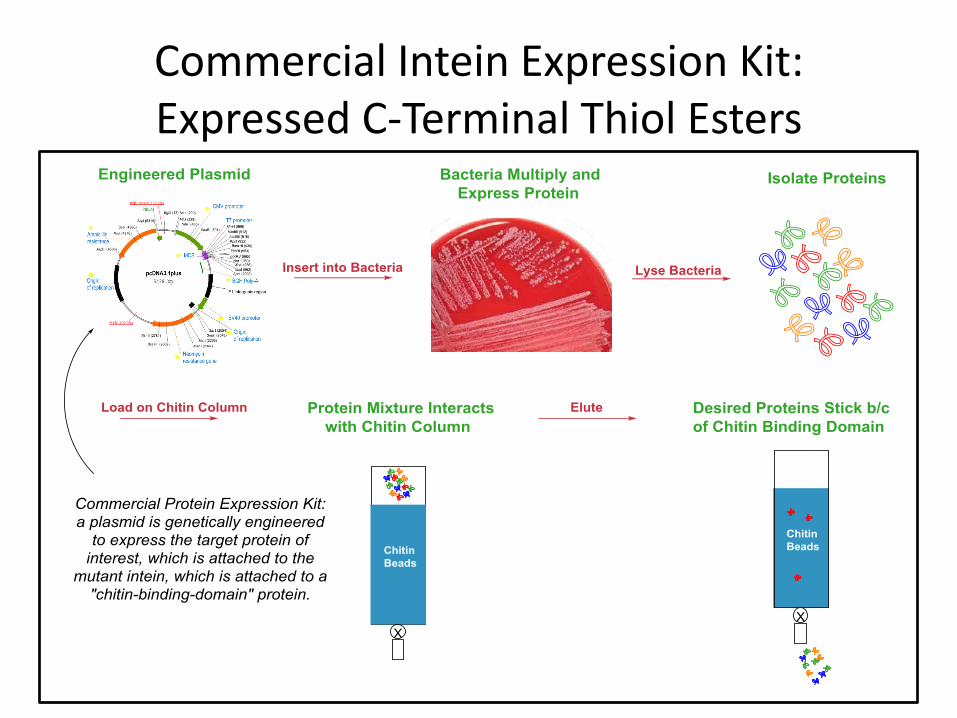
Commercial Intein Expression Kit: Expressed C-Terminal Thiol Esters
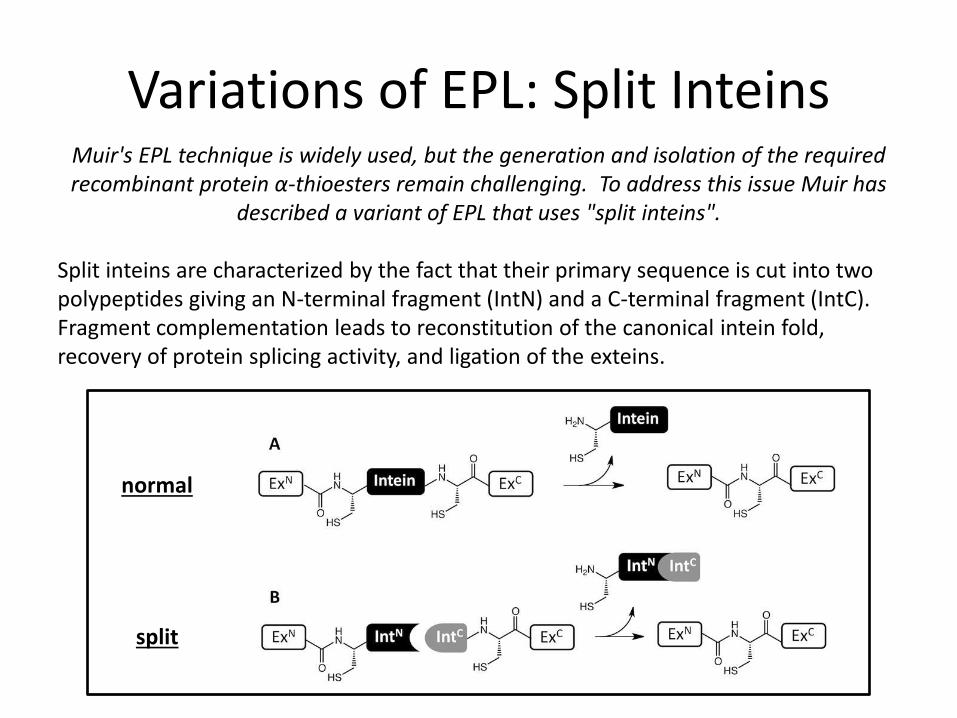
Variations of EPL: Split Inteins Muir's EPL technique is widely used, but the generation and isolation of the required recombinant protein α-thioesters remain challenging. To address this issue Muir has
described a variant of EPL that uses "split inteins". Split inteins are characterized by the fact that their primary sequence is cut into two polypeptides giving an N-terminal fragment (IntN) and a C-terminal fragment (IntC). Fragment complementation leads to reconstitution of the canonical intein fold, recovery of protein splicing activity, and ligation of the exteins.
normal
split
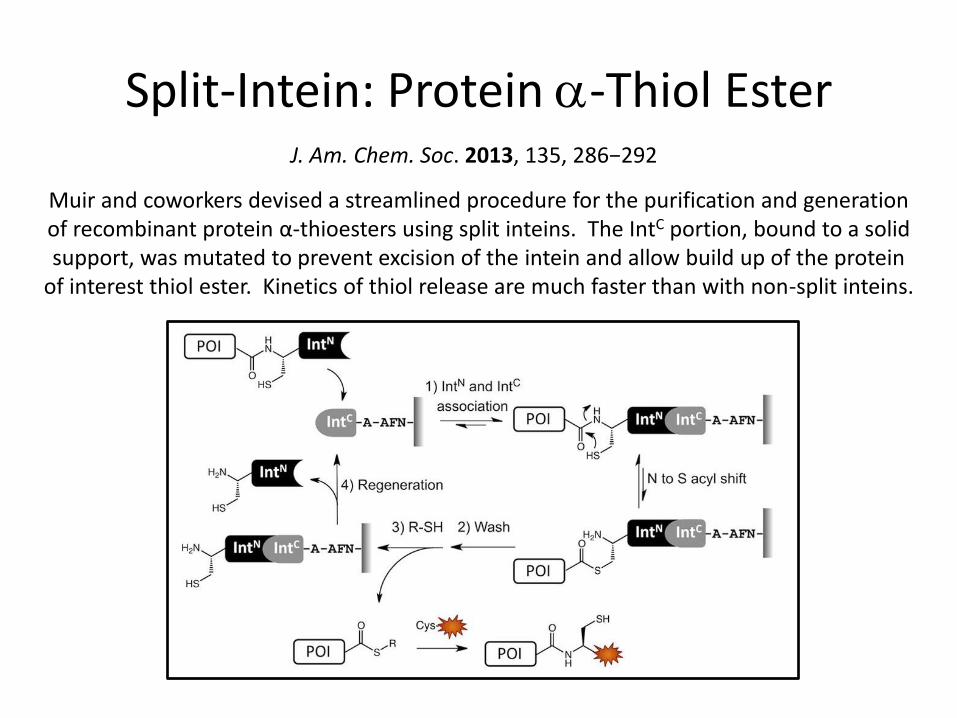
Split-Intein: Protein a-Thiol Ester J. Am. Chem. Soc. 2013, 135, 286−292
Muir and coworkers devised a streamlined procedure for the purification and generation of recombinant protein α-thioesters using split inteins. The IntC portion, bound to a solid support, was mutated to prevent excision of the intein and allow build up of the protein
of interest thiol ester. Kinetics of thiol release are much faster than with non-split inteins.
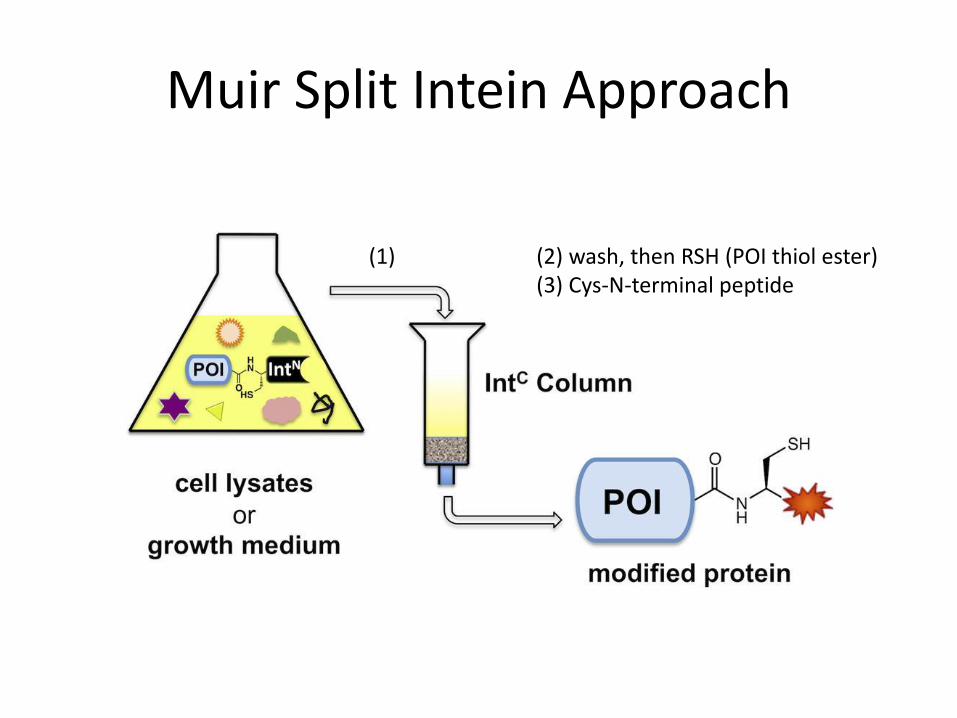
Muir Split Intein Approach
(1) (2) wash, then RSH (POI thiol ester) (3) Cys-N-terminal peptide
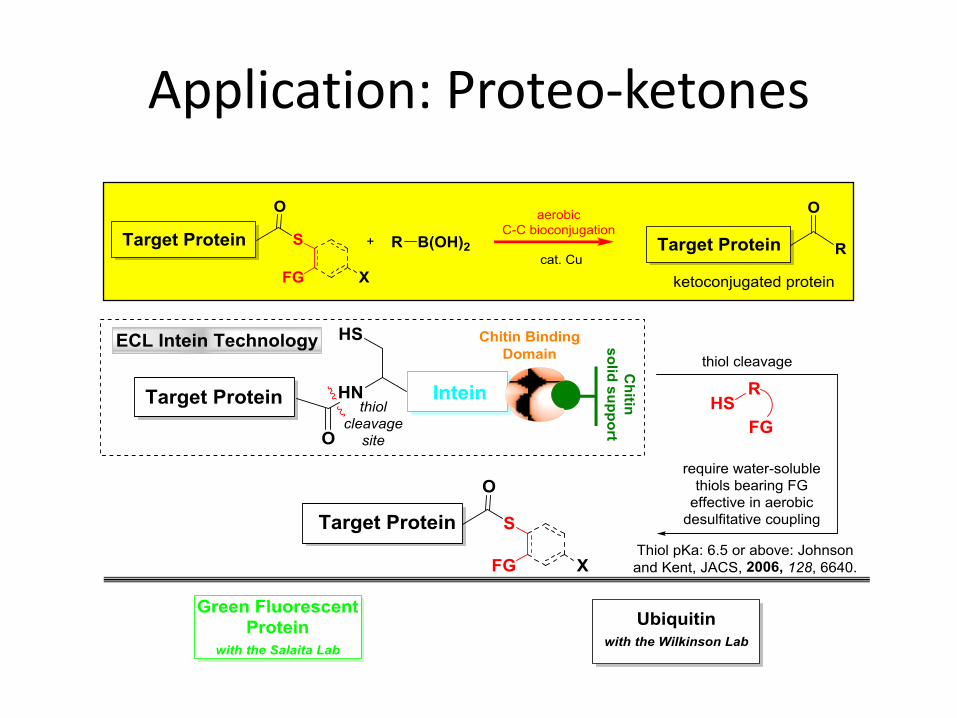
Application: Proteo-ketones