chromosomallossandbreakageinmousebonemarrowandspleencells...
TRANSCRIPT

[CANCERRESEARCH54, 3533—3539,July 1, 1994)
ABSTRACT
Benzene Is a widely recognized human and animal carcinogen. In spiteof considerable research, relatively little Is known about the genotoxicevents that accompany benzene exposure in vivo.To gain Insinjitsinto themechanisms underlying the genotoxic effects of benzene, we have characterized the origin of the micronuclei that are formed in bone marrow
erythrocytes and splemc lymphocytes of benzene-treated mice using twomolecular cytogenetic approaches: (a) fluorescence in situ hybridizationwith a centromeric DNA probe; and (b) staining with the calcinosisRaynaud's phenomenon-esophageal dismoblllty-sclerodactyly-telangiectaxis syndrome of scleroderma (CREST) antibody, an antibody recognlz.
Ing a centromeric protein. Following the p.o. administration of benzene(220 or 440 mg/kg) to male CD-i mice, a significant increase in micronudel was observed in the bone marrow erythrocytes. In situ hybridizationwith a centromeric DNA probe and Immunofluorescent staining with theCREST antibody indicated that the inicronuclel in bone marrow erythrocytes were formed from both chromosome loss and breakage The
m*jorlty of the micronuclel originated from chromosome breakage. Adose-related increase in micronucleated cells was also observed in splenocyte cultures established from these benzene-treated animals. In contrast
to the bone marrow erythrocyte results, the mgjorlty of benzene-inducedniicronuclei in the cytokinesis-blocked splenocytes labeled with theCREST antibody indicating that these mlcronuclel were the result ofwhole chromosome loss. These data demonstrate that both aneuploidy andchromosomal breakage are early genotoxic events induced by benzene orIts metabolites in vivo and also indicate that the nature of the chromosomal alterations may vary depending on the target organ or cell type.
INTRODUCTION
Chronic exposure to benzene is associated with the development ofleukemia in humans and of tumors at multiple sites in animals (1, 2).In spite of considerable research, relatively little is known about thenature of the genotoxic events that accompany benzene exposure invivo (3—5).Extensive in vitro genotoxicity testing has indicated thatbenzene is weakly mutagenic or nonmutagenic in standard genemutation assays but is capable of inducing chromosomal breakage and
may interfere with chromosomal segregation (4, 5). In vivo studies onthe genotoxicity of this agent have also demonstrated that significantincreases in micronuclei and structural chromosomal aberrations areobserved in the bone marrow of rodents following benzene exposure(4—7).
Similar chromosomal damage has been observed in the peripheralblood lymphocytes of benzene-exposed workers. Cytogenetic studiesof individuals occupationally exposed to benzene have shown consistently an association between benzene exposure and elevated frequencies of structural chromosomal aberrations (8, 9). In addition, in
Received 11/12/93; accepted 5/4/94.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
I This work was supported by funds provided by the Cigarette and Tobacco Surtax
Fund ofthe State of California through the Tobacco-related Disease Research Program ofthe University of California, Grant 1KT36.
2 Present address: Office of Environmental Health Hazard Assessment, Pesticides and
Environmental Toxicology Section, 2151 Berkeley Way, Annex 11, Berkeley, CA 94704.3 To whom requests for reprints should be addressed, at Environmental Toxicology
GraduateProgram,Departmentof Entomology,Universityof California,Riverside,CA92521.
creased frequencies of numerical chromosomal aberrations have alsobeen observed in benzene-exposed workers (9—11). However, incontrast with most studies in which structural chromosomal aberrations have been observed in workers with current benzene exposure,the studies detecting aneuploidy have been performed generally onindividuals who exhibited previous bone marrow toxicity and wereinitiated some time after benzene exposure had ceased (10, 12—15).This suggests that the observed numerical chromosomal alterationsmay be an effect secondary to chronic myelotoxicity or aplasticanemia rather than a direct consequence of benzene exposure.
To determine whether aneuploidy is an early event accompanyingbenzene exposure and to gain insights into the mechanisms underlyingthe genotoxic effects of benzene, we have used two recently developed molecular cytogenetic approaches, fluorescence in situ hybridization with a centromeric DNA probe and immunofluorescent labeling with the CREST―antibody, to characterize the origin ofmicronuclei formed in bone marrow erythrocytes and splenic lymphocytes following the in vivo exposure of benzene to mice. Micronuclei are formed when either an entire chromosome or a chromosome fragment fails to segregate into the daughter cells during celldivision. To identify these two types of micronuclei, the presence orthe absence of centromeric DNA sequences or centromeric proteins ina micronucleus has been used to distinguish a micronucleus formed asa result of chromosome loss (centromere-containing) from a micronucleus formed as a result of chromosome breakage (centromerelacking). The presence of centromeric DNA sequences in the micronuclei of erythrocytes from mouse bone marrow preparations wasdetermined using a biotinylated murine major satellite probe (16).Recent studies by Miller et aL (17) have indicated that this approachis effective at detecting aneuploidy-inducing agents in mice. A similar
approach using an antibody isolated from the plasma of patients withthe autoimmune disease CREST scleroderma was used to detect thepresence of a kinetochore-associated centromeric protein in micronudci (18). The CREST staining technique has been shown to be highly
effective in distinguishing aneuploidy-inducing agents from clastogens in human and mouse cells (17, 19—21).In these experiments wehave used the CREST staining technique to confirm the results observed using the DNA probe in the micronucleated bone marrowerythrocytes; we have also used it to identify the origin of micronuclei
in splenocyte cultures established from benzene- and vincristinetreated animals. The induction of micronucleated cells in the splenocyte cultures was determined using the cytokinesis-blocked micronucleus assay developed by Fenech and Morley (22) and recentlymodified by Ren et a!. (23) for use with mouse splenocytes.
MATERIALS AND METHODS
Chemicals. Benzene (+99%) was obtained from Aldrich Chemical Cornpany (Milwaukee, WI). Vincristine sulfate salt (98%), cytochalasin B, concanavalin A type IV, DNA type XIII (human placental DNA), herring sperm
4 The abbreviations used are: CREST, calcinosis-Raynaud's phenomenon-esophageal
dismobility-sclerodactyly-telangiectasia syndrome of sclerodenna; PBS, phosphate-buffered saline; SSC, standard saline-citrate; 2X SSC, 0.3 MNaCI plus 0.03 Msodium citrate,pH7.0; PCE,polychromaticerythrocyte;NCE,normochromaticerythrocyte;PNMbuffer,0.1 M phosphate buffer containing 5% powdered nonfat milk supernatant and 0.5%Nonidet P-40.
3533
Chromosomal Loss and Breakage in Mouse Bone Marrow and Spleen CellsExposed to Benzene in Vivo@
Hongwei Chen, Doppalapudi S. Rupa, Rajpal Tomar,2 and David A. Eastmond3
Environmental Toxicology Graduate Program, University of California at Riverside@ Riverside, California 92521
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Bone marrow cells were harvested 24 h after male mice were given vincristinesulfateandbenzene in vivo. The frequency of micronucleated PCE was determined byscoring1000
PCF)aniinal from acridine orange-stainedslides.Treatment
@a MNPCEbp@@CSaline―
5000 0.8 (0.8)e 1.6 (0.6)Vincristine―(0.125 mg/kg) 5000 53.2' (5.6) 1.2 (0.2)Olive@ 9000 0.56(0.7) 1.2 (0.2)Benzene@(220 mg/kg) 9000 18.0'@(6.4) 1.6―(0.3)
@ (440 mg/kg) 9000 14.9―(4.4) 1.8―(0.3)
CHROMOSOMALLOSSAND BREAKAGEINDUCEDBY BENZENE
DNA, and biotin-11-dUTP were purchased from Sigma Chemical Co. (St.Louis, MO). The CREST antibody (a centromere-positive control serum forthe antinuclear antibody test) was purchased from Chemicon International, Inc.(Temecula, CA). Fluorescent goat anti-human IgG was obtained from Antibodies, Inc. (Davis, CA). Fluoresceinated avidin DES, fluoresceinated rabbitanti-goat IgG, and biotinylated goat anti-avidin D were purchased from VectorLaboratories (Burlingarne, CA).
Treatment and Bone Marrow Harvest Conditions. Four-week-old maleCD-i mice were obtained from Charles River Laboratories (Raleigh, NC).Benzene was dissolved in olive oil and administered by p.o. gavage to6—8-week-old mice at doses of 220 and 440 mg/kg. Vincristine sulfate wasdissolved in 0.9% NaCI and administered by i.p. injection at a dose of 0.125mg/kg. Control mice received olive oil and 0.9% NaCI, respectively. Bonemarrow preparations were made 24 h following the administration of benzeneusing standard procedures (24). The bone marrow cells were smeared ontoslides and fixed in 90% methanol at —20°Cfor 20 miii. Slides were stored at—20°C within a nitrogen atmosphere until their use.
Isolation of Spleen Cells, Culture, and Harvest. Spleens from the controlor treated mice were removed, washed in Hanks' balanced salt solution (pH7.25) containing 0.5 mM L-glutamine, and the cells were mechanically teased
from the connective tissue using forceps. The splenocytes were then isolatedby Ficoll-Paque (Pharmacia, Piscataway, NJ) density gradient centrifugation,
washed in RPM!, and cultured in RPM! 1640 supplemented with 1 m@L-glutamine, 10% fetal calf serum, 50 @Wmlgentamicin sulfate, and 5 @g/mlconcanavalin A at an initial density of 1 X 10@ cells/ml. Cultures weremaintained in 96-well microtiter plates at 37°Cin a 5% CO2 atmosphere. Asrecommended by Ren et a!. (23), cultures for the micronucleus assay weretreated with cytochalasin B (5 @Wml)48 h after culture initiation. Cell harvestwas initiated at 64 h by centrifugation onto slides (Cytospin 2; Shandon,Pittsburg, PA) followed by fixation in 100% methanol for 10 mm at roomtemperature.
Conventional Bone Marrow Micronucleus Assay. Bone marrowpreparations were stained with acridine orange according to the method of Hayashici aL (25) with the following modifications. The slides were stained for 2 mmin the acridineorange solution L0.1%aqueousstock diluted 1:30with Sorensen'sphosphatebuffer (pH 6.8)] and rinsed twice for 3 mm each in phosphatebuffer.
Biotinylation of the Mouse Major Satellite Probe. A degenerate majorsatellite DNA probe, cloned into the HindIII site of the pBS vector, was kindlyprovided by Dr. Heinz-Ulrich Weier (University of California, San Francisco).This probe was amplified and biotinylated by polymerase chain reaction using
a Perkin Elmer CetsisThermacycler as described by Weier et al. (16). The twoprimers, WBS2 (5'CfCGAAATfAACCCFCACfAAAGG-3') and WBS4(5'-GAArfGTAATACGACTCACTATAG-3'), were synthesized by OperonLaboratories (Alameda, CA).
fluorescence in Situ Hybridization with the Major Satellite Probe.Bone marrow slides were immersed in 0.1 M sodium phosphate containing0.5% Nonidet P.40 for 15 mm and then refixed in 2% paraformaldehyde for30—455. The target DNA was denatured in 70% formamide-2X SSC for 4 mmat 72°C;chilled in ice-cold 70% ethanol for 4 mm; and dehydrated in 70, 85,and 100% ethanol solutions at room temperature. The biotinylated DNA probe(approximately 10 ng/slide) was mixed with sonicated human placental DNA(1 g.i.g/slide),herring sperm DNA (1 @tg/slide),and the hybridization cocktail(55% formamide-10% dextran sulfate-i X SSC). The hybridization mix wasdenatured at 72°Cfor 4 mm and chilled on ice. This mix (10 @l)was thenapplied to each slide and the slides were incubated overnight at 37°Cin ahumidified chamber. Following hybridization, the slides were washed in 50%formamide-2X SSC (pH 7.0) for 15 min at 42°C.To reduce nonspecific DNAbinding, the slides were then rinsed in PNM buffer for 15 mm at 37°C.Fluorescent staining of the hybridized probe was performed by alternatingincubations of fluoresceinated avidin and a biotinylated anti-avidin antibodyaccording to standard procedures (17, 26).
CREST Antibody Labeling of Mouse Bone Marrow and Splenocytes.The CREST antibody labeling method described by Eastmond and Tucker (19,20) was used for the splenocyte studies. Similar procedures with the following
modifications were used for the bone marrow preparations. Briefly, the bonemarrow cells were fixed in 2% paraformaldehyde for 30—45s and rinsed inPBS containing0.01% Tween 20 for 5 min. The CRESTantibody(in a i:idilution with PBS) was applied to the slides for 75 rain. The slides were rinsedin 0.1% Tween 20 twice for 2 mm and then placed in PNM buffer for 5 mm.
The antibody was then labeled using fluoresceinated-goat anti-human IgCi(i:120 dilution in PBS) for 75 mm followed by the rinse in 0.1% Tween 20 andPNM. The signal was amplified by adding another layer of fluoresceinatedrabbit anti-goat IgO (1:120 dilution in PBS) and the nucleus was counterstained with propidium iodide (1 ,.@g/mlin antifade) (26).
Slide Scoring and Criteria. All slides were randomized and coded prior toscoring. For the analysis of bone marrow micronuclei, a minimum of i000micronucleated erythrocytes were scored per dose for each experiment. Onlythose micronuclei within erythrocytes exhibiting a clear plasma membraneboundary were scored. For the micronucleus analysis in splenocytes, a minimum of 1000 binucleated cells/animal were scored using previously publishedcriteria (19). For the acridine orange-stained slides, scoring was performedusing a blue filter (excitation at 475—495nm, emission at 520 nm). For theprobe-labeled bone marrow slides and the splenocyte slides, the micronucleiwere scored using a blue-violet filter (excitation at 530—585nm, emission at600 nm) which allowed the cell membrane as well as the nucleus to beobserved. Following the observationof a micronucleatedcell, the blue filterwas used to detect the presence of the fluorescein-labeled DNA probe or theCREST antibody. Photographs were taken using KOdak Ektachrome 400 orTmax 400 film.
Statistical Analyses Dose-related increases in the frequency of micronudci induced by benzene were determined using the Cochran-Armitage test for
trend in binomial proportions (27). Following a positive response in the trendtest, a 1-tailed Fisher exact test was used to compare each treatment with thecontrol. The Fisher exact test was also utilized to compare the frequency ofmicronuclei induced by vincristine sulfate with its corresponding control.Regression analysis and one-way analysis of variance was used to compare thenuclear division index and the PCE:NCE ratio data for the benzene-treatedanimals (28). Following a significant analysis of variance result, Scheffe's Ftest was used as a post hoc test to compare the results of the individual doses.Student's t test was used to compare the nuclear division index and thePCE:NCE data for the vincristine data. Critical values were determined usinga 0.05 probability of type I error.
RESULTS
Micronucleated Erythrocytes. Using conventional acridine orange staining, the frequency of micronucleated PCE in the bonemarrow was determined following the administration of benzene andvincristine sulfate to male CD-i mice (Table 1). Significant increasesin the frequency of micronucleated PCE were observed at both the
220- and 440-mg/kg doses of benzene and at the 0.125- mg/kg doseof vincristine sulfate. A comparison of the PCE:NCE ratios, a measureof the proliferation of the bone marrow erythrocytes, and an indirectmeasure of toxicity indicates that there was no significant decrease inthe PCE:NCE ratio between the control and the 220- and 440-mg/kgdoses of benzene (Table 1). Rather, a significant increase in the
Table 1 Induction ofmicronuclei in mouse bone marrow e,ythrocytes byvincristine sulfate and benzene
a Total number of polychromatic erythrocytes scored.M@ number of micronuclei/1000 polychromatic erythrocytes scored/animal.
C Mean of the polychromatic erythrocytes to normochromatic erythrocytes ratios per
animal.dAdministered by i.p. injection; 5 animals/treatment.e The SD is indicated in parentheses.‘Differssignificantly from the respective control (1-tailed Fisher exact test; P < 0.05).g Administered by p.o. gavage; 9 animals/treatment and dose.
h Exhibits a significant dose-related trend [Cochran-Armitage binomial trend test(MNPCE), or regression analysis (PCE/NCE); P < 0.05].
3534
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
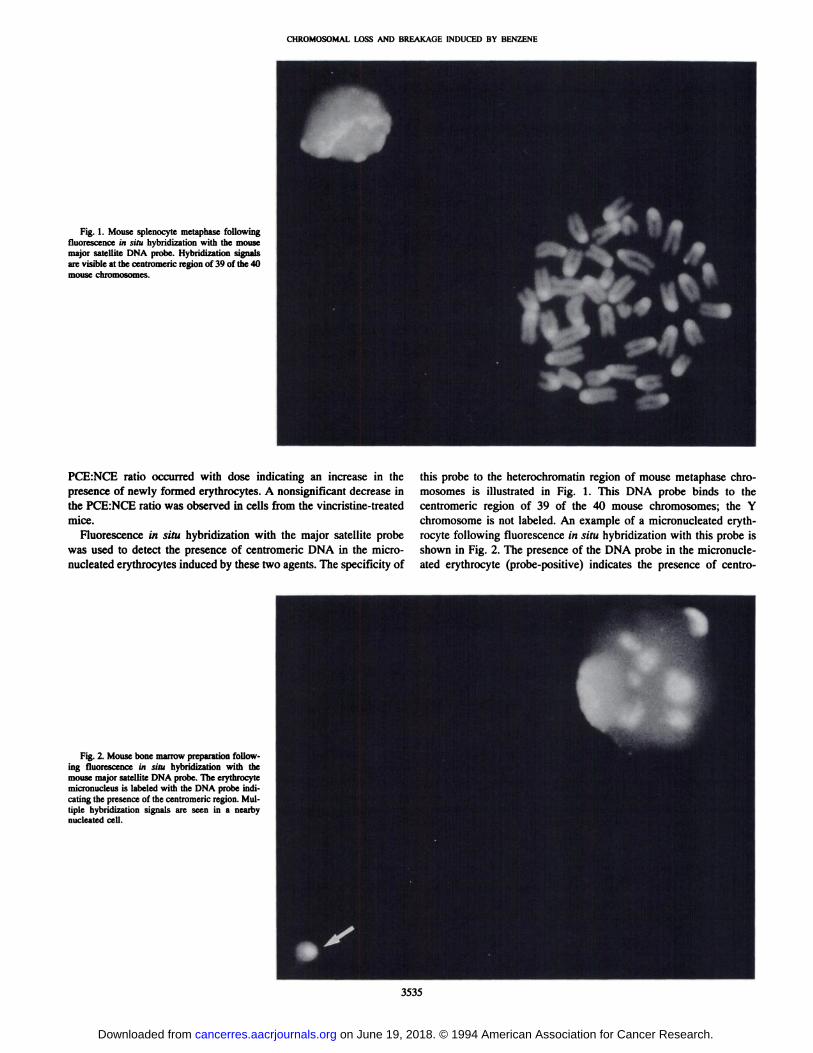
CHROMOSOMAL LOSS AND BREAKAGE INDUCED BY BENZENE
Fig. 1. Mouse splenocyte metaphase followingfluorescencein situ hybridizationwiththe mousemajor satellite DNA probe. Hybridization signalsare visible at the centromenc region of39 ofthe 40mousechromosomes.
PCE:NCE ratio occurred with dose indicating an increase in thepresence of newly formed erythrocytes. A nonsignificant decrease inthe PCE:NCE ratio was observed in cells from the vincristine-treatedmice.
Fluorescence in situ hybridization with the major satellite probewas used to detect the presence of centromeric DNA in the micronucleated erythrocytes induced by these two agents. The specificity of
Fig. 2. Mouse bone marrow preparation following fluorescencein situ hybridizationwith themousemajorsateffiteDNAprobe.Theerythrocytemicronucleus is labeled with the DNA probe mdicaring the presence of the centromeric region. Multiple hybridization signals are seen in a nearbynucleated cell.
this probe to the heterochromatin region of mouse metaphase chromosomes is illustrated in Fig. 1. This DNA probe binds to thecentromeric region of 39 of the 40 mouse chromosomes; the Y
chromosome is not labeled. An example of a micronucleated erythrocyte following fluorescence in situ hybridization with this probe isshown in Fig. 2. The presence of the DNA probe in the micronucleated erythrocyte (jrobe-positive) indicates the presence of centro
3535
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Saline
animals, a 43-fold increase. A statistically significant increase inprobe-lacking micronucleated erythrocytes was observed in the yin
cristine-treated animals. These micronuclei increased from 0.67/1000
erythrocytes in the control animals to 5.0/1000 in the treated mice.To confirm these results using the major satellite probe, we used
bone marrow slides from the same experiments and applied theCREST antibody procedure. Using this technique, the proportions ofcentrornere-containing and centromere-lacking micronuclei observedwere similar to those obtained using the DNA probe technique (Fig.5). Significant increases in CREST-positive and -negative micronuclei
were observed in mice that were given doses of both benzene andvincristine sulfate.
Micronucleated Splenocytes. The micronucleus assay was performed on splenocytes isolated from the benzene- and vincristinesulfate-treated mice and cultured in vitro for 64 h after isolation.Immunofluorescent labeling using the CREST antibody was performed to determine the origin of the induced micronuclei in cytokinesis-blocked binucleated cells. An illustration of a cytokinesisblocked splenocyte in which the micronucleus is labeled with theCREST antibody is shown in Fig. 6. Using this approach, significantdose-related increases in micronucleated cells were observed in thecultured splenocytes isolated from the benzene- and vincristinetreated mice (Fig. 7). CREST staining indicated that splenocytesisolated from the benzene- and vincristine-treated animals exhibitedsignificant increases in CREST-positive micronuclei (Fig. 7). In contrast to the bone marrow, the majority of benzene-induced micronucleiin the splenocytes were labeled with the CREST antibody indicatingthat they originated from lagging chromosomes. The frequency of
3536
CHROMOSOMAL LOSS AND BREAKAGE INDUCED BY BENZENE
meric DNA in the micronucleus. Therefore, the micronucleus wasmost likely formed by the failure of an entire chromosome to segregate properly during mitosis (Fig. 2). The absence of centromericDNA sequences in a micronucleated erythrocyte (probe-negative)indicates that the micronucleus was formed from an acentric chromosome fragment or the Y chromosome (Fig. 3). Nucleated bone marrowcells, each containing numerous centromeric regions labeled with thecentromeric DNA probe, are visible in Figs. 2 and 3.
Following hybridization with the major satellite probe, the ftcquency of probe-containing and probe-lacking micronuclei was determined in the bone marrow erythrocytes for both benzene andvincristine sulfate (Fig. 4). Since PCE and NCE cannot be easilydistinguished following either the DNA hybridizations or the CRESTstaining, these results represent the frequency of micronuclei cornbined from these two types of erythrocytes. Significant increases ofboth probe-containing and probe-lacking micronucleated erythrocyteswere observed at each dose of benzene. The major proportion (approximately 84%) of the induced micronuclei were probe lackingindicating that they were formed from chromosome breakage andacentric fragments. The frequency of probe-negative micronuclei increased from 1.6/i000 erythrocytes in the control animals to 11.7 and10.9/1000 erythrocytes in animals treated with the 220- and 440-mg/kg doses of benzene, respectively. This represents an approximately 7-fold increase over the control frequency. Sixteen percent ofthe micronuclei induced by benzene labeled with the centromericprobe. The frequency of these micronuclei increased from 0.22/1000erythrocytes in the control animals to 2.3 and 2.0/1000 erythrocytes in
animals treated with the 220- and 440-mg/kg doses of benzene, a9—10-foldincrease. In contrast, the majority (73%) of the micronucleated erythrocytes induced by the spindle-disrupting agent vincristinesulfate consisted of probe-positive micronuclei indicating that thesemicronuclei were formed from lagging chromosomes (Fig. 4). Thefrequency of probe-containing micronuclei increased from 0.33/1000erythrocytes in the control animals to 14.3 in the vincristine-treated
0
U0
0
000
a,
z
Majorprobe-lackingUN
I Majorprobe-containingMN
25
20
15
10
5
Saline Vincristine Olive oil Benzene Benzene(0.125 mg/kg) (220 mg/kg) (440 mg/kg)
Fig. 4. Induction of micronuclei (MN) in the bone marrow erythrocytes following theadministration ofvarious doses ofbenzene and vincristine sulfate to male CD-i mice. Thefrequency of micronuclei labeling or failing to label with the centromeric mouse majorsatellite DNA probe is shown.
U)a,>.U0
a,000
a,a,.z
0 Centromere-lackingim(CREST.)U Centrornare-containingMN(CREST+)U iei unscorableforCRESTstalntng
Vincristine Olive oil Benzene Benzene
(0.125 mg/kg) (220 mg/kg) (440 mg/kg)
Fig. 5. Induction of micronuclei (MN) in bone marrow erythrocytes staining with thecentromeric CREST antibody following the administration of various doses of benzeneand vincristine sulfate to male CD-i mice. Micronuclei in which the CREST staining wasinadequate for scoring are also indicated.
Fig. 3 Mouse bone marrow preparation following fluorescence in situ hybridizationwith the mouse major satellite DNA probe. An erythrocyte micronucleus (arrow) is seenwhich is not labeled with the DNA probe indicating that this micronucleus was formedfroman acentricfragment.Multiplehybridizationsignalsare also seen in the nearbynucleated cell.
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Table 2 Mouse splenocyte nuclear divisionindexSplenocyteswere isolated from mice given benzene or vincristine sulfate invivo.Cultures
wereestablished,cytochalasinBwasaddedat48h, andthecellswereharvestedat64 h of culture. The percentage of binucleated cells and the nuclear division indexweredetermined
by scoring 200 cells/animal.Binucleated
%binucleatedTreatmentcells° cells―NDICSaline'
3000 50.0 (18) 1.8 (0.4)Vincristine―(0.125 mg/kg) 3000 45.7 (22) 1.7(0.5)Olive
oile 9000 45.9 (5.1) 1.7 (0.2)Benzenee (220 mg/kg) 9000 52.I'(9.i) l.8@(0.2)Benzenee (440 mg/kg) 9000 56.@/(5.5) l.9'@(0.1)
S
CHROMOSOMAL LOSS AND BREAKAGE INDUCED BY BENZENE
CREST-positive micronuclei increased from approximately 3/1000binucleated cells in the controls to 11/1000 cells in animals treatedwith benzene at 440 mg/kg. A very similar increase was seen in thesplenocytes of the vincristine sulfate-treated mice. These results indicate that chromosomal loss accounted for approximately 80% of themicronuclei induced in both the benzene- and vincristine-treated animals. Significant increases in CREST-negative micronuclei wereobserved only in the splenocytes isolated from the benzene-treatedanimals. The frequency of CREST-negative micronuclei increasedfrom 1.3/1000 binucleated cells in the olive oil-treated mice to 3.0/1000 in mice given the highest dosage of benzene.
The nuclear division index, a measure of cell growth and nuclear
0 Centromere-iacking MN (CREST-)U Centromere-contalningMN (CREST+)
a)@ 15@
U@@ - _
@0@@
I10@ i@ _
:@@
Saline Vlncrlstine Olive oil Benzene Benzene(0.125 mg/kg) (220 mg/kg) (440 mg/kg)
Fig. 7. Induction of CREST-containing and CREST-lacking micronucleated splenocytes (MN) in cultured splenocytes isolated from CD-i mice treated in vivo with variousdosesof benzeneandvincristinesulfate.
‘a'
a woo binuclested cells were scored per animal for micronuclei.
b Mean percentage and SD of 200 binucleated cells scored per animal.C Mean and SD of nuclear division index per animal. Nuclear division index [Ml +
(2 X M2) + (3 x M3) + (4 X M4)]/N; MI—M4,number of cells with 1—4nuclei; N, totalnumber of cells scored.
d Administered by i.p. injection; 3 animals/treatment.e Administered by p.o. gavage; 9 animals/treatment and dose.tExhibits a significant dose-related trend (regression analysis; P < 0.05).
. a
bFig. 6. Cytokinesis-blocked splenocyte with one micronucleus (arrow) photographed
using (a) a blue-violet filter setting and (b) a blue filter setting. The labeling of thecentromeric CREST antibody to multiple regions within both the main nuclei and themicronucleus is visible in b.
division, for the splenocytes isolated from the controls and the treatedanimals is presented in Table 2. A significant dose-related increase inthe nuclear division index was also observed in the benzene-treatedanimals when compared with control animals.
DISCUSSION
In these studies, two new molecular cytogenetic techniques havebeen used to characterize the origin of micronuclei formed in bonemarrow erythrocytes and cultured splenic lymphocytes following thein vivo administration of benzene to male CD-i mice. Hybridizationwith a centromeric DNA probe or staining with the CREST antibodyindicated that the induced micronuclei resulted from both chromosome breakage and chromosomal loss. These results indicate that bothclastogenicity and aneuploidy are early genotoxic effects that accompany benzene exposure.
In the bone marrow erythrocytes, chromosomal breakage cornprised the majority (approximately 85%) of the observed geneticdamage induced by benzene. However, in cultured splenocytes isolated from the same animals, chromosomal loss was the predominanteffect accounting for approximately 80% ofthe observed micronuclei.This different profile of chromosomal changes occurring in the bonemarrow and spleen was somewhat surprising and may be related tocertain organ-specific aspects of benzene-induced genotoxicity suchas the unique metabolic capabilities of the bone marrow or a selectiveaccumulation of benzene metabolites in this organ (29, 30). There are,however, a number of significant differences between these two assaysystems which may also contribute to this different profile of geno
I
a,
3537
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CHROMOSOMAL WS5 AND BREAKAGE INDUCED BY BENZENE
toxicity. For example, the assay on bone marrow samples was performed on erythroid cells, an actively proliferating cell lineage,whereas the assays on the spleen were performed on splenocytes, cellsin the nondividing G0 state. The higher percentage of chromosomalbreakage observed in the bone marrow may also be due in part to theselective extrusion of large micronuclei containing whole chromosomes which may occur as erythrocytes extrude their main nucleiduring the maturation process. Previous studies have shown that asignificant proportion of micronuclei containing entire chromosomescan be lost through this mechanism (31). The observed differences inthe bone marrow and spleen may also reflect different inherent susceptibifities of these cell types to genetic damage. The frequency andorigin of spontaneous rnicronuclei observed in the control animals differed considerably between the two cell types. The spontaneous micronucleus frequency in the bone marrow erythrocytes was approximately0.0008 of which only 18% contained the centromeric region. In contrast,in the cultured splenocytes the spontaneous micronucleus frequency was0.0046 ofwhich 67% contained the centromeric region. The spontaneousmicronucleus frequencies observed in these studies are consistent withthe those observed by previous investigators (17, 23, 24).
The requirement for splenocyte culturing is another differencebetween the bone marrow and spleen preparations that may accountfor some of the differential effects observed in these two cell types.Since micronucleus formation requires nuclear division, splenocyteswhich are not actively dividing under normal conditions must bestimulated to undergo at least one cell division for micronuclei tobe expressed. Coils containing significant chromosome breakagemay be unable to undergo cell division, divide at an altered rate, orlose fragment-containing micronuclei during culture. The recovery ofa high proportion of chromosome-containing micronuclei observed inthis study may reflect a unique nature of the cells harvested at the 64-hharvest time. However, the contribution of such effects to our resultsis believed to be relatively minor: (a) the splenocyte harvest timeselected in this study was developed to optimize the recovery ofmicronuclei following the in vivo administration of cyclophosphamide, a potent clastogen (23). The effectiveness of these proceduresfor detecting ciastogen-induced micronuclei in cultured splenocyteshas been demonstrated for a number of clastogenic agents (32, 33);(b) the high recoveryof binucleatedcellsobservedat the64-h harvesttime (>50% in cultures from the benzene-treated mice; Table 2)allowed the majority of the cells in the culture to be evaluated formicronucleus formation. These cells represent an accumulation ofcells dividing over a relatively long period of cell culture (16 h); and(c) in the first of the two experiments, cytochalasin B was added at 24h to the splenocyte cultures and these cells were harvested 16 h laterto determine whether a selective loss of micronuclei had occurred atan earlier time in culture. Although an adequate number of binucleated cells was recovered in cultures from control and vincristinetreated animals, an inadequate number of binucleated cells was recovered for scoring in the 40-h cultures from the benzene-treatedanimals. Apparently cell division was significantly delayed in thecultured splenocytes from these animals because a high yield ofbinucleated cells was obtained from parallel cultures from these sameanimals at the 64-h harvest. These results indicate that a significantloss of micronuclei was unlikely to have occurred at early time pointsbecause relatively little cell division was occurring in splenocytes
from the benzene-treated animals.One unique aspect of these data is an apparent stimulation of cell
growth that was observed following benzene exposure. After exposurea significant dose-related increase in the bone marrow PCE:NCE ratiowas observed (Table 1). The biological significance of this observation may be questioned inasmuch as a relatively high PCE:NCE ratiowas observed in the saline-treated controls in the vincristine experi
3538
ment. However, increased PCE:NCE ratios were also observed inpreliminary experiments with benzene. Further, the PCE:NCE ratioobserved at the 440-mg/kg dose of benzene exceeds that of historicalcontrol frequencies from this laboratory. These results suggest thatwithin 24 h following benzene administration to these CD-i mice,cellular regeneration had already commenced within the erythrocytecompartment of the bone marrow. A similar dose-related increase inthe nuclear division index was also observed in the cytokinesisblocked splenocytes isolated from the benzene-treated animals (Table2). However, rather than indicating that cellular regeneration wasoccurring, we believe that this more likely reflects a shift in thekinetics of cell division in the benzene-treated splenocytes sincerelatively little nuclear division was observed in cells harvested at40 h of culture.
The significant increases in chromosomal loss observed in bothbone marrow erythrocytes and splenic lymphocytes following theadministration of a single dose of benzene to mice indicates thataneuploidy is an early genotoxic event that accompanies benzeneexposure. Aneuploidy has been shown to play an important role inneoplastic development resulting from exposure to chemical andphysical agents as well as that resulting from congenital and familialpredispositions to cancer (34—36).The induction of aneuploidy observed in these experiments, combined with the observations of numerical aberrations in patients at various stages of benzene-inducedbone marrow dysfunction such as aplastic anemia, preleukemia, andacute nonlymphocytic leukemia (9, 15, 37, 38), indicates that theinduction of aneuploidy can be a lasting consequence of benzeneexposure and one factor which may contribute to leukemogenesis.Patients with myelodysplastic syndromes who exhibit elevated frequencies of chromosomal alterations have increased risks of developing acute nonlymphocytic leukemia (39, 40). Furthermore, the nonrandom loss or gain of chromosomes 5, 7, and 8 in leukemia cells isfrequently observed in patients with acute nonlymphocytic leukemiasassociated with chemical and solvent exposures (41—43).
In summary, we have utilized two novel molecular cytogeneticapproaches, immunofluorescent labeling with the CREST antibodyand fluorescence in situ hybridization with a centromeric DNA probe,to characterize the origin of micronuclei formed in bone marrowerythrocytes and in cultured splenic lymphocytes following benzeneexposure. These studies have demonstrated that both chromosomebreakage and chromosome loss are induced in the bone marrow andspleen as a consequence of benzene exposure. Chromosome breakagerepresented the majority of the alterations in the bone marrow erythrocytes whereas chromosome loss comprised the majority of thealterations observed in the cultured splenocytes. Studies are continuing in our laboratory to investigate the in vivo clastogenic and aneuploidy-inducing effects of the various benzene metabolites as well asto understand the mechanisms underlying these genotoxic effects.
REFERENCES
1. International Agency for Research on Cancer. Benzene. IARC Monographs on theEvaluation of Carcinogenic Risks to Humans. Overall Evaluations of Carcinogenicity: An Updating of IARC Monographs 1—42,Suppl. 7, pp. 120—122.Lyons, France:InternationalAgencyfor Researchon Cancer,1987.
2. Huff, J. E., Haseman, J. K.. DeMarini, D. M., Eustis, S., Maronpot, R. R., Peters,A. C., Persing, R. L, @hrisp,C. E., and Jacobs, A. C. Multiple-site carcinogemcityof benzene in Fischer 344 rats and B6C3F1 mice. Environ. Health Perspect., 82:125—163,1989.
3. international Agency for Research on Cancer. Benzene. IARC Monogr. Eval. Carcinog. Risks Chem. Hum., 29: 93-148, 1982.
4. Dean, B. J. Recent findings on the genetic toxicology of benzene, toluene, xylenesand phenols. Mutat. Rca., 154: 153—181,1985.
5. Waters, M. D., Stack, H. F., Brady, A. L, Lohman, P. H. M., Haroun, L, and Vainio,H. Use of computerized data listings and activity profiles of genetic and relatedeffects in the review of 195 compounds. Mutat. Res., 205: 295—312,1988.
6. Ciranni, R., Barale, R., and Adler, I. D. Dose-related clastogenic effects induced by
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CHROMOSOMAL LOSS AND BREAKAGE INDUCED BY BENZENE
benzene in bone marrow cells and in differentiating spermatogonia of Swiss CD1mice. Mutagenesis, 6: 417—421,1991.
7. Tice, R. R., Costa, D. L., and Drew, R. T. Cytogenetic effects of inhaled benzene inmurine bone marrow: induction of sister chromatid exchanges, chromosomal aberrations, and cellular proliferation inhibition in DBA/2 mice. Proc. Nail. Acad. Sci.USA,77: 2148—2152,1980.
8. Sarto, F., Cominato, I., Pinton, A. M., Brovedani, P. G., Merler, E., Peruzzi, M.,Bianchi, V., and Levis, A. G. A cytogenetic study on workers exposed to lowconcentrations of benzene. Carcinogenesis (Lond.), 5: 827—832,1984.
9. Aksoy, M. Benzene carcinogenicity. In: M. Aksoy (ed), Benzene Carcinogenicity,pp. 113-148. Boca Raton, FL: CRC Press, Inc., 1988.
10. Pollini, G., and Biscaldi, 0. P. Persistenza dde alterazioni del cariotipo nei IinfOCitidisoggetti benzolici a dieci anrn dalla intossicazione [Persistence ofkaryotype alterations inlymphocytes ten years after benzene poisoning]. Med. Lay., 67: 465—472,1976.
11. Eastmond, D. A. Induction of micronuclei and aneuploidy by the quinone-formingagents benzene and o-phenylphenol. Toxicol. Left. (AmsL), 67: 105—118, 1993.
12. Pollini, G., Biscaldi, 0. P., and Robustelli Della Curia, G. Le alterazioni chromosomiche dci linfociti rilevate dope cinque anni in soggetti gia affeui da emopatiabenzolica [Chromosome changes in lymphocytes five years after benzene hemopathy]. Med. Lxv., 60: 743—758,1969.
13. Forni, A., Cappellini, A., Pacifico, E., and Vigliani, E. C. Chromosome changes andtheir evolution in subjects with past exposure to benzene. Arch. Environ. Health, 23:385—391,1971.
14. Liniecki, J., Bajerska, A., and Gluszczowa, M. Analiza kariologiczna limfocytowkiwi obwodowej u osob z przebytym przewleklym zatruciem benzenem [The effectsof passed chronic benzene poisoning upon the karyotype of peripheral blood lymphocytes].Med.Pr.,22: 187—193,1971.
15. Ding, X., Li, Y., Ding, Y., and Yang, H. Chromosome changes in patients withchronic benzene poisoning. Chin. Med. J. (EngI. Ed.), 96: 681—685,1983.
16. Weier, H., Zitzelsberger, H. F., and Gray, J. W. Non-isotopical labeling of murineheterochromatin in situ by hybridization with in vitro-synthesized biotinylated gamma(major)satelliteDNA.Biotechniques,10: 498—505,1991.
17. Miller, B. M., Zitzelsberger, H. F., Weier, H., and Adler, I. D. Q@ification ofmicronuclei in murine erythrocytes: immunofluorescent staining using CREST antibodies compared to in situ hybridization with biotinylated gamma satellite DNA.Mutagenesis, 6: 297—302,1991.
18. Rattner, J. B. The structure of the mammalian centromere. Bioessays, 13: 51—56,1991.
19. Eastmond, D. A., and Tucker, J. D. Identification of aneuploidy-inducing agents usingcytokinesis-blocked human lymphocytes and an antikinetochore antibody. Environ.Mol. Mutagen., 13: 34—43,1989.
20. Eastmond, D. A., and Tucker, J. D. Kinetochore localization micronucleated cytokinesis-blockeda@mnesehamsterovarycells:a new and rapidassayfor identifyinganeuploidy-inducing agents. Mutat. Rca., 224: 517—525,1989.
21. Gudi, R., Xu, J., and Thilagar, A. Assessment of the in vivo aneuploidy/micronucleusassay in mouse bone marrow cells with 16 chemicals. Environ. Mol. Mutagen., 20:106—116,1992.
22. Fenech, M., and Morley, A. Measurement of micronuclei in lymphocytes. Mutat.Res., 147: 29—36,1985.
23. Ren, L, Yang, J., and Zhang, H. Use of the cytokinesis-block micronucleus methodin mouse splenocytes. Mutat. Res., 262: 119—124,1991.
24. MacGregor, J. T., Heddle, J. A., Elite, M., Margolin, B. H., Ramel, C., Salamone,M. F., Tice, R. R., and Wild, D. Guidelines for the conduct of micronucleus assays
in mammalian bone marrow erythrocytes. Mutat. Res., 189: 103—112, 1987.25. Hayashi, M., Sofuni, T., and Ishidate, M., Jr. An application of acridine orange
fluorescent staining to the micronucleus test. Mutat. Res., 120: 241—247,1983.26. Trask, B., and Pinkel, D. Fluorescence in situ hybridization with DNA probes.
Methods Cell Biol., 33: 383—400,1990.27. Margolin, B. H., and Risko, K. J. The statistical analysis of in vivo genotoxicity data:
case studies of the rat hepatocyte UDS and mouse bone marrow micronucleus assays.In: J. Ashby, F. J. de Serres, M. D. Shelby, B. H. Margolin, M. Ishidate, Jr., andG. C. Backing(eds.), Evaluationof Short-TermTests for Carcinogens,Vol. 1,pp. 1.29—1.42.Cambridge, United Kingdom: Cambridge University Press, 1988.
28. Abacus Concepts. Berkeley, CA: StatView II Statistical Software, 1987.29. Sawahata, T., Rickert, D. E., and Greenlee, W. F. Metabolism of benzene and its
metabolites in bone marrow, In: R. D. Irons (ed), Toxicology of the Blood and BoneMarrow, pp. 141—148.New York: Raven Press, 1985.
30. Rickert, D. E., Baker, T. S., Bus, J. S., Barrow, C. S., and Irons, R. D. Benzenedisposition in the rat after exposure by inhalation. Toxicol. AppI. Pharmacol., 49:417—423,1979.
31. Parton, J. W., Garriott, M. L., and Beyers, J. E. Expulsion of demecolcine-inducedmicronuclei from mouse bone marrow polycttromatic erythrocytes. Environ. Mol.Mutagen., 17: 79—83, 1991.
32. Ren, L., Zhang, H., Yang, J., and Zhang, Z. A sequential study on the use ofcytokinesis-block micronucleus method in mouse splenocytes. Mutat. Res., 301:223—227,1993.
33. Farooqi, Z., Darroudi, F., and Natarajan, A. T. The use of fluorescence in situhybridization for the detection of aneugens in cytokinesis-blocked mouse splenocytes.Mutagenesis, 8: 329—334, 1993.
34. Sasaki, M. Current status of cytogenetic studies in animal tumors with specialreference to nonrandom chromosome changes. Cancer Genet. Cytogenet., 5: 153—172,1982.
35. Oshimura, M., and Barrett, J. C. Chemically induced aneuploidy in mammalian cells:mechanisms and biological significance in cancer. Environ. Mutagen., 8: 129—159,1986.
36. Hansen, M., and Cavenee, W. Genetics of cancer predisposition. Cancer Res., 47:5518—5527,1987.
37. Forni, A., and Moreo, L. Cytogenetic studies in a case of benzene leukaemia. Eur. J.Cancer, 3: 251—255,1967.
38. Erdogan, 0., and Aksoy, M. Cytogenetic studies in thirteen patients with pancytopenia and leukaemia associated with long-term exposure to benzene. New IstanbulContrib. Clin. Sci., JO: 230—247, 1973.
39. Appelbaum, F. R., Barrall, J., Storb, R., Ramberg, R., Doney, K., Sale, G. E., andThomas, E. D. Clonal cytogenetic abnormalities in patients with otherwise typicalaplastic anemia. Exp. Hematol. (NY), 15: 1134—1139, 1987.
40. Beris, P. Primary clonal myelodysplastic syndromes. Semin. Hematol., 26: 216—233,1989.
41. Yunis, J. J. The chromosomal basis of human neoplasia. Science (Washington DC),221: 227—236,1983.
42. Rowley, J. D., and La Beau, M. M. Cytogenetic and molecular analysis of therapyrelated leukemia. Ann. NY Acad. Sci., 567: 130—140,1989.
43. Cuneo, A., Fagioli, F., Pam, I., Tallarico, A., Previta, R., Piva, N., Gretel Carli, M.,Balboni, M., and Castoldi, G. Morphologic, immunologic and cytogenetic studies inacute myeloid leukemia following occupational exposure to pesticides and organicsolvents. Leuk. Res., 16: 789—796,1992.
3539
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1994;54:3533-3539. Cancer Res Hongwei Chen, Doppalapudi S. Rupa, Rajpal Tomar, et al.
in VivoSpleen Cells Exposed to Benzene Chromosomal Loss and Breakage in Mouse Bone Marrow and
Updated version
http://cancerres.aacrjournals.org/content/54/13/3533
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/54/13/3533To request permission to re-use all or part of this article, use this link
on June 19, 2018. © 1994 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from