chromosome territories, nuclear matrix filaments and inter...
TRANSCRIPT

Cremer, T., Dietzel, S., Eils, R., Lichter, P. and Cremer, C. Chromosome territories, nuclear matrix filaments and inter-chromatin channels: a topological view on nuclear architecture and function. In: P. E. Brandharn and M. D. Bennett (editors). Kew Chromosome Conference IV, pp. 63-81. Royal Botanic Gardens, Kew.
Chromosome territories, nuclear matrix filaments and inter-chromatin channels: a topological view on
nuclear architecture and function T. Cremer1, S. Dietzel1, R. EiIs2, R. Lichter3 and C. Cremer4 1. Institut für Humangenetik and Anthropologie, Universität Heidelberg, Germany. 2. Interdisziplinäres Zentrum rur wissenschaftliches Rechnen (MR), Universität Heidelberg, Germany. 3. Abteilung Organisation komplexer Genome, Deutsches Krebsfoschugszentrum, Heidelberg, Germany. 4. Institut fur Angewandte Physik,Universität Heidelberg, Germany. "There is no reason to doubt the tenet of molecular biologists that knowledge of structure leads to understanding of function; what we know about chromatin structure is simply not enough to permit the connection to be made." (Gary Felsenfeld 1978) Introduction Times are gone when a biochemist might have safely considered the nucleus as a bag in which extended chromatin fibres float around in the nuclear sap with few geometrical constraints. Instead, the nucleus has emerged in recent years as a highly compartmentalized structure. It contains chromosome territories (Cremer et al. 1993) and numerous multi-protein complexes which interact specifically with DNA or RNA and fulfill distinct functional roles in transcription, splicing, replication and repair (Spector 1993; Roth 1995). These nuclear machineries are often found in specific domains, resulting in typical punctuate or speckled distributions of nuclear antigens in immuno-fluorescent labeling experiments (van Driel et al. 1995). To avoid confusion with chromosomal domains (see below) we refer to the latter as non-chromatin domains. Studies of the topological relationships between non-chromatin domains and chromosome territories are still in their infancy (Zirbel et al. 1993). In this essay we will consider the possible role of in vivo networks of nuclear matrix filaments and interchromatin channels in nuclear compartmentalization with spezial reference to the question of their topological relationships with chromosomal domains and non-chromatin domains.
63

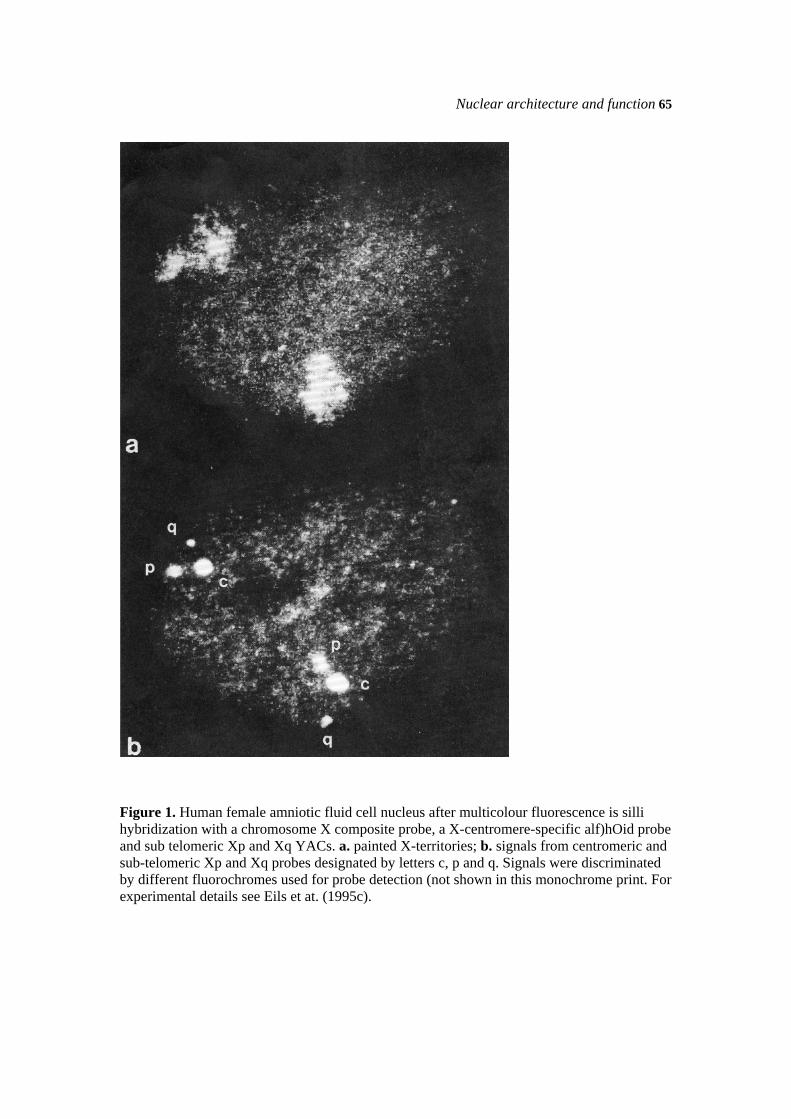
64 T. Cremer et al. Evidence for organized chromosome territories The hypothesis that each interphase chromosome occupies an exclusive nuclear territory was first put forward by Carl Rabi (1885) and Theodor Boveri (1909). In the 1960s and 1970s the view was popular that the euchromatic parts of all chromosomes strongly de-condense and intermingle during interphase, while heterochromatin remains tightly condensed throughout the cell cycle (for reviews see Comings 1968; Wischnitzer 1973; Vogel & Schroeder 1974). Direct evidence for the existence of chromosome territories in the cell nucleus of a variety of species from the animal and plant kingdoms was obtained by in situ hybridization with complex probes delineating entire chromosomes (Manuelidis 1985; Schardin et al. 1985; Cremer et al. 1988; Lichter et al. 1988; Pinkel et al. 1988; Leitch et al. 1990). For this procedure Pinkel and his colleagues coined the now widely-used term "chromosome painting". The possible penetration of chromatin fibres from neighbouring chromosome territories is limited to territory surfaces (Eils et al. 1995c). The same study and two other studies (Bischoff et al. 1993; Rinke et al. 1995) contradict the prevalent view that the inactive X chromosome (Xi) should retain a very compact organization during the entire cell cycle, while the active X chromosome (Xa) should become largely de-condensed during interphase. In a series of human female amniotic fluid cell nuclei, the painted X-territories showed similar volumes, but differed significantly in surface and most conspicuously in shape, as judged by the calculation of a roundness factor (RF, for details see Eils et al. 1995c). The mean RFXi/RFXa ratio obtained for 54 pairs of X-territories was 2.03 ± 0.74 and in 51 nuclei the territory with the larger RF was unequivocally identified as Xi by its co-localization with the Barr body. The reasons for these structural differences are not understood (Eils et al. 1995a,c). As an example, Fig. la shows an overlay of light optical sections through a female amniotic fluid cell nucleus with two painted X-territories. In addition, Xp- and Xq-subtelomeric YAC clones and an X-specific alphoid probe were used to visualize the chromosome ends and the centromeric heterochromatin (Fig. ib). Fig. 2 shows a three-dimensional reconstruction of the same X-territories viewed from different angles. A growing body of evidence indicates that chromosome territories are partitioned into discrete chromosomal compartments or domains, such as chromosome arm and band-like domains, centromeric and telomeric regions, chromatin loop domains or single genes (Manuelidis 1990; Lawrence et al. 1993; Schedl & Grosveld 1995; Zhao et al. 1995; Eils et al. 1995a, 1995b; Kurz et al. 1995, and our unpublished data). Centromeric and telomeric regions, as well as a number of active and inactive genes were observed at the surface of the X- and several autosome chromosome territories (Cremer et al. 1993; Eils et al. 1995c; Kurz et al. 1995). Three genes studied so far, i.e. the dystrophin gene, ß-myosin HC and ß-globin, were
Nuclear architecture and function 65
Figure 1. Human female amniotic fluid cell nucleus after multicolour fluorescence is silli hybridization with a chromosome X composite probe, a X-centromere-specific alf)hOid probe and sub telomeric Xp and Xq YACs. a. painted X-territories; b. signals from centromeric and sub-telomeric Xp and Xq probes designated by letters c, p and q. Signals were discriminated by different fluorochromes used for probe detection (not shown in this monochrome print. For experimental details see Eils et at. (1995c).
66 T. Cremer et al.
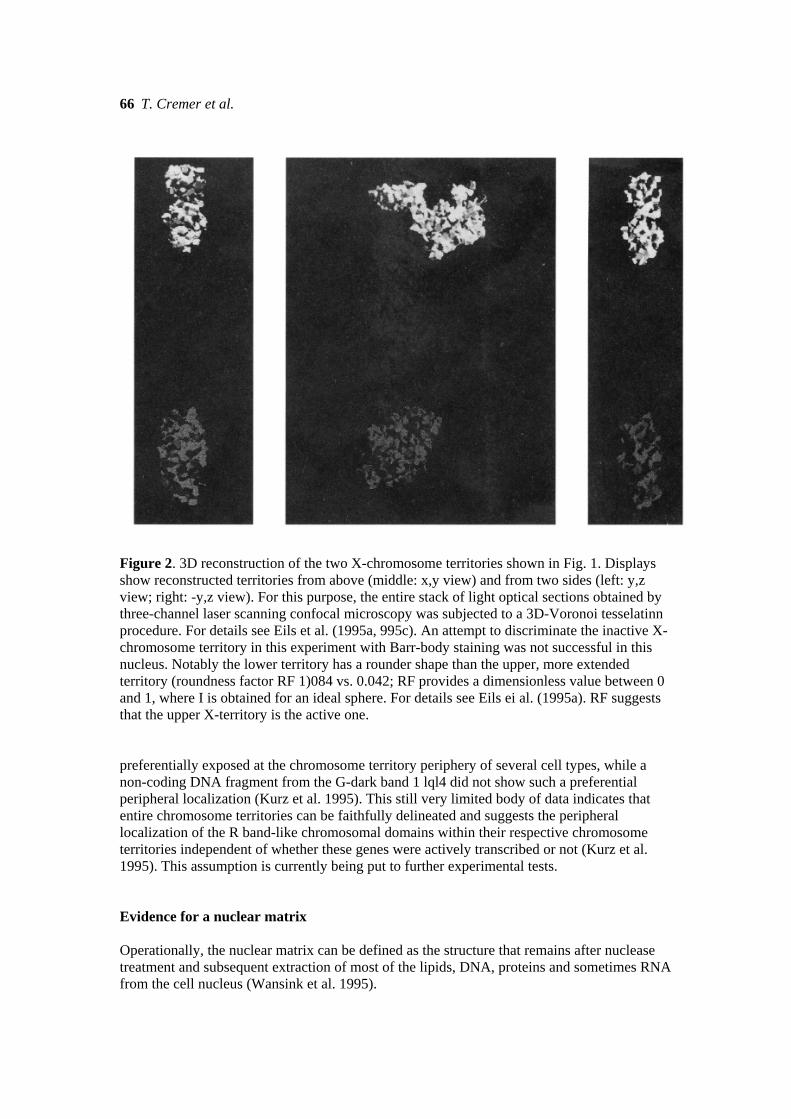
Figure 2. 3D reconstruction of the two X-chromosome territories shown in Fig. 1. Displays show reconstructed territories from above (middle: x,y view) and from two sides (left: y,z view; right: -y,z view). For this purpose, the entire stack of light optical sections obtained by three-channel laser scanning confocal microscopy was subjected to a 3D-Voronoi tesselatinn procedure. For details see Eils et al. (1995a, 995c). An attempt to discriminate the inactive X-chromosome territory in this experiment with Barr-body staining was not successful in this nucleus. Notably the lower territory has a rounder shape than the upper, more extended territory (roundness factor RF 1)084 vs. 0.042; RF provides a dimensionless value between 0 and 1, where I is obtained for an ideal sphere. For details see Eils ei al. (1995a). RF suggests that the upper X-territory is the active one. preferentially exposed at the chromosome territory periphery of several cell types, while a non-coding DNA fragment from the G-dark band 1 lql4 did not show such a preferential peripheral localization (Kurz et al. 1995). This still very limited body of data indicates that entire chromosome territories can be faithfully delineated and suggests the peripheral localization of the R band-like chromosomal domains within their respective chromosome territories independent of whether these genes were actively transcribed or not (Kurz et al. 1995). This assumption is currently being put to further experimental tests. Evidence for a nuclear matrix Operationally, the nuclear matrix can be defined as the structure that remains after nuclease treatment and subsequent extraction of most of the lipids, DNA, proteins and sometimes RNA from the cell nucleus (Wansink et al. 1995).

Nuclear architecture and function 67 Evidence for such an operationally-defined nuclear matrix was first provided by the pioneering work of Berezney & Coffey (1974) and has been confirmed for many cell types and species. In electron micrographs a lamina-pore complex located at the nuclear periphery, residual nucleolar structures and an anastomosing intra-nuclear network of 10 to 13 nm core filaments was discriminated (for reviews see Mattem et al. 1995; Penman 1995). It has been proposed that the lamina-pore complex, together with the internal nuclear matrix, plays an essential role in chromatin organization and in maintaining the size and shape of the original nucleus after extensive digestion of the DNA and subsequent extraction of chromatin and all loosely-bound proteins. Important functional roles have also been ascribed to the nuclear matrix as attachment sites for the protein machineries involved in transcription, splicing, DNA replication and repair and for the nucleo-cytoplasmic transport of macromolecules (Berezney & Coffey 1975; Zehnbauer & Vogelstein 1985; Mullenders et al. 1988; Berezney 1991a, 1991b; Cook 1991; Getzenberg et al. 1991; Boulikas 1992; Hassan et al. 1994). Many proteins with roles in transcription, splicing, replication and repair have been isolated from nuclear matrix preparations, but the constitution, in vivo presence and functional role(s) of the core filaments seen in EM preparations have not been defined to date (Mattem et al. 1995; Penman 1995). Standard protocols of nuclear matrix preparations, originally introduced by Berezney & Coffey (1974) and modified by numerous other authors, include high salt extraction of nuclei, but it should be noted that nuclear matrix preparations can also be isolated under isotonic conditions employing lithium 3.5-di-iodosalicylate (US) in the millimolar range (Mirkovitch et al. 1984), or after removal of nuclease-digested chromatin by electrophoresis from nuclei embedded in agarose beads (Jackson et al. 1988). Differences in the protocols applied by various authors may have contributed to the variability in the composition of nuclear matrix preparations. Still, it is difficult to rule out the possibility that these procedures favoured the entanglement and cross-linking of protein/nucleic acid complexes (Cook 1988). In order to provide unequivocal evidence for a nuclear matrix in vivo in the specific sense of an anastomosing three-dimensional intra-nuclear network of matrix core filaments, it is necessary to isolate the proteins which constitute the respective homo- or hetero-polymers and to demonstrate that these polymers are indeed present in the nucleus of the living cell. The role of several candidate proteins is under investigation at present (for details see Mattem et al. 1995).

68 T. Cremer et al. Chromosome territories and the nuclear matrix: structural, functional and dynamic aspects Structural and functional aspects For metaphase chromosomes Saitoh & Laemmli (1993) have presented a loop/scaffold model. The principal feature of their model (quoted from Saitoh & Laemmli 1993) is a "hypothetical, unfolded, 'chromatid fibre' which contains a central, longitudinal region called the AT-queue and two different types of loops, the shorter Q-loops and the longer R-loops. The ATqueue, an optical signal, is proposed to arise from a line-up of highly AT-rich SARs as mediated by the scaffold." It has been argued that the chromosome scaffold is not a solid rod-like sub-structure along the chromatid axis, but is made up of many discrete anchoring complexes (Earnshaw 1988). One pertinent question concerns the fate of the mitotic scaffold when a chromatid is transformed into a chromosome territory in late telophase/ early GI. The most simple view implies that the scaffold is retained throughout interphase and forms an essential internal structure of chromosome territories as well. The possibility of a different scaffold topology in mitotic and interphase chromosomes should nevertheless also be taken into consideration, e.g. one scaffold structure may be located in the interior of mitotic chromosomes, but it becomes at least partially exposed at chromosome territory surfaces during interphase. Alternatively, the mitotic scaffold may be replaced by a different interphase structure. Chai & Sandberg (1988) have proposed that the nuclear matrix plays an essential role in forming individual chromosomal "microcosms" in the nucleus and confining them to given nuclear locations. One would like to know to which extent structurally and topologically defined chromosomal domains at various hierarchical levels represent functionally autonomous units. In transgenic experiments the function of an autonomous unit should be entirely independent of the site of integration into the host genome. To the extent that function is based on certain topological requirements the sequence information contained in an autonomous unit should be sufficient to meet such needs. The definition of domain boundaries, which allow the differential compaction of higher-order chromatin structures and restrict the activity of regulatory elements to the chromosomal domain in which they reside, provides a major focus of ongoing research. Scaffold/matrix-associated regions (SARs/MARs) have been implicated in the provision of such boundary functions, but regions other than SARs/MARs apparently play a role as boundaries as well (Bonifer et al. 1991; Eissenberg & Elgin 1991; Schedl & Grosveld 1995). What we know at present about chromosomal domain structure and topology and the possible involvement of a nuclear matrix is simply not enough to permit the functional connections to be made.

Nuclear architecture and function 71 Prediction 1: Formation of a three-dimensional channel network lined by chromosome territory and chromosomal domain surfaces An extended, three-dimensional in vivo network of nuclear channels supposedly starts at nuclear pores (Fig. 3d) and expands between chromosome territory surfaces (Zachar et al. 1993; Zirbel et al. 1993). We noted the possibility that the surface of chromosome territories could be greatly enhanced by infoldings (Cremer et al. 1993). Accordingly, branches of the channel network may lead from the chromosome territory periphery to chromosomal domains located in the chromosome territory interior (Cremer et al. 1995). We call the entire hypothetical channel network together with its lining chromatin surfaces the inter-chromosomal domain (lCD) compartment and the interior of the entire channel network the lCDspace. This definition leaves the question of the possible extent of channel ramifications into the territory interior open for future studies. Interchromatin channels have been proposed by other authors as well (Blobel 1985; Chai & Sandberg 1988; Spector 1990; Kramer et al. 1994; Wansink et al. 1994; 1995; Razin & Gromova 1995). The lCD-compartment model, however, provides a possible physico-chemical explanation for the formation and topology of a channel network based on the following rationale: We assume that chromosome territories and chromosomal domains are negatively charged under physiological conditions. This results in a Donnan potential between the interior and exterior of these structures and leads to repulsive electric forces between opposite surfaces of chromosome territories or chromosomal domains. In addition to a fraction of negatively charged phosphate groups, which are not completely neutralized by histones, negatively charged, chromatin-bound non-histone proteins may increase the Donnan potential further. We predict that a Donnan potential in the range of a few mV is sufficient to maintain lCD channels in the range of a few um width. In transcriptionally inactive chromatin (see below) the channels may become empty and very narrow, although they should never collapse completely as long as the range of repulsive forces between negatively charged, opposite chromosomal domains exceeds the very short range of attractive forces (Fig. 3a). The channel width can be modulated by local differences in the size of repulsive forces, by Brownian movements of chromosomal domain surfaces, and quite extensively by the inclusion of macromolecules and macromolecule complexes with negative electric net charge, e.g. by proteins entering the lCD-space via the nuclear pores or by RNA transcribed and released at chromosomal domain surfaces (Wansink et al. 1994) (Fig. 3b,d). The existence of an intra-nuclear channel network has not been proved by direct experimental evidence. However, micro-electrode measurements support theassumption that chromatin in the nucleus is in fact negatively charged (in the range of a few millivolts) (Loewenstein & Kanno 1963;
72 T. Cremer et al.

Giulian & Diacumakos 1977; Oberleitner et al. 1993; R.H.A. Fink & H. Koester, University of Heidelberg, pers. comm.). Model calculations based on the Debye-Hückel and Donnan potential theory performed under highly simplified assumptions (C. Cremer, unpublished) suggest that the predicted physical effects seem plausible enough to justify further theoretical and experimental efforts. Prediction 2: Facilitated diffusion, enrichment of macromolecules and preferential formation of macromolecule complexes within lCD channels The assumed physical properties of the lCD-compartment permit the enrichment and preferential transport of proteins and RNAs within the lCDspace, either by channelled diffusion or via matrix filaments (see below for the lCD-space as a potential site for the preferential formation of matrix core filaments). A different distribution of macromolecules between the lCDspace and the chromosomal domain interior is expected for both electrical and mechanical reasons. Due to electric exclusion from the interior of negatively charged chromosomal domains, macromolecules with negative electric net charge should become enriched to a certain extent within the lCD-space, while the opposite effect is expected for macromolecules with positive electric net charge (e.g. histones). In addition, we assume that the size and shape of macromolecules, as well as the extent of chromosomal domain compaction, affect their diffusion into the interior of the domains. Experimental data to provide reasonable estimates of expected enrichment factors are not available at present. It should neverthiess be noted that a moderate enrichment of individual components of a macromolecule complex within the lCD-space compared to concentrations in the chromosomal domain interior could largely increase the probability that complexes involved in transcription, splicing, DNA replication and repair assemble within the lCD-space. Large macromolecule aggregates may become immobilized between opposite chromosomal domain surfaces at sufficiently narrow sites of the lCD channels without the need for a filamentous nuclear matrix. The observation that large accumulations of splicing snRNPs, and certain RNA species were located at the surface of chromosome territories (Zirbel et al. 1993) is compatible with the idea that channels between chromosome territories can expand to an extent necessary to harbour aggregates of a size that is visible easily in the light microscope. Prediction 3: lCD-channels and chromosomal domain surfaces play an essential role in gene regulation Let us consider the following scenario: a number of factors specifically required to trigger the transcriptional activity of a so far silent gene enter the lCD-channel network via the nuclear pores. These factors have to scan the
Nuclear architecture and function 73

genome until they find their specific target site (s). Let us further assume that most chromosomal domains are organized in a way that their interior is inaccessible for genome scanning by these factors. In this scenario genome scanning would be limited to the lining surfaces of lCD-channels. A successful and rapid search would be facilitated if target DNA sequences were exposed at chromosomal domain surfaces. Similarly, the shift of regulatory sequences into the chromosomal domain interior could help to shut off the transcription of certain genes permanently, for example in terminally differentiated cell types. The binding of a certain transcription factor at its specific DNA-target site could trigger a sequence of events resulting in a locally restricted, functionally "open" (and nuclease sensitive) chromatin configuration. The genetically active configuration could be provided (i) by the formation of a micro-domain comprising the active transcription complex together with the respective regulatory and coding DNA-segments at the surface of a larger chromosomal domain, which retains an inaccessible interior structure (Fig. 3b,d) or (ii) by the opening (de-condensation) of an entire chromosomal domain to an extent that transcription complexes can now enter the domain interior (a possibility not indicated in Fig. 3). The active transcription complex may move along the coding sequence entirely exposed in the lCD-space or the coding sequence may pass the fixed transcription complex (for the possible involvement of matrix core filaments, see below). The chromosomal domain/lCD-channel organization suggested here could limit genome scanning to a minor fraction of the entire genome. This would provide an advantage in case the number of certain factors is limited to an extent that the time needed for scanning of the entire genome becomes prohibitively long, or in case the specificity of factor binding is strongly impaired by the number of unspecific binding sites provided by the entire genome. It is not known to which extent these potential advantages play a role in gene regulation, if any. The potential advantage of nuclear compartmentalization should become less obvious if the nuclear chromatin is compartmentalized into an exceedingly large number of small chromosomal domains. In such a case the total surface of chromosomal domains would increase markedly and most of the genome would become exposed at this surface. Alternatively, chromosomal domains may not have any specific 3D-structure leading to the preferential exposure of certain sequences at their surface, but may be dynamic (for example due to Brownian movements of chromatin fibres) to an extent that regulatory sequences become exposed randomly at chromosomal domain surfaces. In this case a factor would miss its target site if it passes the respective chromosomal domain surface at an inappropriate time. It remains a completely open question to which extent the 3D-structure and topology of higher order chromatin structures above the level of single chromatin domains with autonomous genetic functions matter in the 74 T. Cremer et al.

functional architecture of the cell nucleus. It has been noted that nuclei of differentiated cell types vary greatly in the amount and distribution of apparent heterochromatic and euchromatic regions (Manuelidis 1990), ranging from nuclei which appear to be entirely heterochromatic and transcriptionally inactive (e.g. nuclei of later-stage normoblasts during erythropoiesis) to almost euchromatic nuclei with high transcriptional complexity (e.g. nuclei of some neuronal cell types). A narrow width of channels in heterochromatic nuclear regions as compared to a large width in euchromatic regions could help to direct factors to nuclear compartments with high transcriptional activity. Prediction 4: DNA-replication and DNA-repair are topologically related to chromosomal domain surfaces Similar to the transcription and splicing complexes, the machineries for DNA replication and repair should also form preferentially within the lCDspace. DNA sequences (genes) exposed at lining chromosomal domain surfaces should replicate early. Findings that repair is preferentially observed for DNA sequences associated with nuclear matrix preparations (for review see Boulikas 1992) are compatible with the view that nuclear matrix preparations reflect the content of the lCD-compartment (see below). lCD-channels and nuclear matrix networks The biophysical properties of the lCD-space described above would favor the formation of nuclear matrix core filaments within lCD channels in vivo as soon as proteins (and RNA components) with homo- or hetero-polymerforming capabilities become abundant in this space. Intra-channel matrix core filaments may then serve as attachment sites for macromolecules and macromolecule complexes for the various nuclear functions (Fig. 3b). The line of reasoning presented here for the potential formation of intrachannel matrix core filaments is based on specific physico-chemical properties of chromosomal domains, which have already formed for other reasons (see below). While we may attribute a secondary role to such an lCD matrix in the organization and spatial fixation of chromosomal domains and entire chromosome territories, the lack or breakdown of this matrix should be tolerated without a breakdown of the basic nuclear chromatin organization. High salt conditions and/or the removal of fixed negative electric net charges of chromosomal domains by DNase digestion, two treatments often performed in nuclear matrix preparations could reduce the repulsive forces which normally exist between negatively-charged macromolecule aggregates and negatively charged chromatin surfaces, to a critical extent. As a result the complex content of the lCD-space would agglomerate including, if present,
Nuclear architecture and function 75

the matrix core filaments, as well as individual proteins and protein complexes and associated DNA or RNA (Fig. 3c,e). Figure 4 presents an overview on possible topological relationships between nuclear matrix networks, chromosomal domains and lCD-channels (models A-D). In model A we distinguish two hypothetical in vivo matrix networks which serve different purposes and have distinctly different
Figure 4. Hypothetical topological relationships between inter-chromosomal domain (lCD) channels, matrix core filaments and chromosomal domains. Model A. five chromosomal domains are organized by a continuous domain-organizing matrix which extends from a continuous chromosomal scaffold. The scaffold is located in the interior of a chromosome territory, while the outer parts of the chromosomal domains are exposed towards the territory surface (for simplicity helical turns/foldings of this scaffold are not indicated). The domains are separated by the lCD-space and shown in a "closed" state (i.e. with its interior inaccessible to the macromolecule complexes contained in the lCD-space). The lCD-space contains another continuous matrix, called the ICD matrix, presumably serving for the attachment/ transport of macromolecules in this space (compare Fig. Sb). In spite of their supposedly different in eivo topologies the two matrices may approach very close or even interact at some sites and may become topologically indistinguishable in matrix preparations obtained after extensive DNA digestion and extraction. Model B. Like model A, this model assumes a continuous scaffold, but the ICD matrix is disintegrated and reflected only by a few discontinuous ICD fibres (or it may be completely lacking). Numerous macromolecule aggregates contained in the lCD-space are symbolized by small dots (compare Fig. 3). Model C. Here, the chromosome territory and chromosomal domain organizing structure is discontinuous, while a continuous ICD matrix is present as in model A. The chromosome territory/ chromosome domain organization is maintained by specific protein-DNA and protein-protein interactions (symbolized by black patches). Model IL In extreme contrast to model A both the nuclear scaffold and the lCDmatrix are discontinuous, or even entirely lacking. 76 T. Cremer et al.

topologies. One network is supposedly built up within lCD-channels and is involved in essential nuclear functions, such as transcription and splicing (see above and Fig. 3b), while the other resides in the interior of chromosomal domains and fulfills a basic role in the compartmentalization and organization of nuclear chromatin. Models B and C indicate the possibility that only one of the two matrices is present as a continuous network. In model B the chromatin-organizing nuclear matrix is retained, while only discontinuous matrix core filaments or inter-chromatin fibres are contained within the lCD-space. In model C the opposite situation is indicated. A continuous lCD-matrix exists, but chromosome territories and chromosomal domains are organized by large numbers of discrete, specifically spaced protein-DNA and protein-protein interactions. In model D a continuous matrix network is entirely lacking, both in the chromosomal domain interior and in the lCD-space. In this case artificial cross-linking in nuclei subjected to matrix preparation procedures would be required to explain how the nuclear shape and the positioning of intra-nuclear structures, such as nucleoli and centromeres, can be retained after removal of most of the chromatin. The situation depicted in models A and C might interfere with the selective movement of a given chromosome territory, in the case that the continuous lCD-matrix is permanent and connects this territory tightly with adjacent ones. The same consideration holds true for movements of a given chromosomal domain with respect to neighbouring domains within a dynamically-organized chromosome territory. Such restrictions of chromosome territory/chromosomal domain movements (e.g. by an actin/myosin machinery located in the lCD-compartment) are eliminated when the lCD-matrix becomes discontinuous or entirely absent (models B and D). The flexibility of chromosome territory shape is another requirement for complex and independent movements. In essence, model A apparently best serves the need to retain a given three-dimensional nuclear chromatin organization, while its extreme counterpart (model D) seems particularly suited to serve the needs of the nucleus as a dynamic structure. Models A-D may not necessarily be mutually exclusive, but may represent features of a continuum of transient, functional states reflecting a highly dynamic in vivo nuclear matrix organization. The search for the principle (s) of chromosome territory organization and nuclear compartmentalization When considering different models of nuclear compartmentalization, it seems helpful to remember the view of Francois Jacob that evolution has acted as a tinker trying any available opportunity to solve a problem. In his summary of the 1993 Cold Spring Harbor Symposium on "DNA and Chromosomes" entitled "Genetic tinkering - local problems, local solutions.", the late Harold Weintraub reinforced this view. Solutions
Nuclear architecture and function 77

provided by genetic tinkering may not be exclusive, i.e. exploiting a nuclear matrix or channels, but the ingenious combination of both opportunities. While some may argue that much more experimental data should be collected before further attempts to present and defend models of nuclear compartmentalization become worthwhile, we argue the other way round. The formulation of models with specific predictions is indispensable to provoke new insights and decisive experiments, including functional assays. For example, the lCD-compartment model prompts attempts to visualize the predicted lCD-channel network and to see whether channel width varies in heterochromatic and euchromatic nuclear regions. Functional tests could involve attempts to manipulate the in vivo width of lCD-channels experimentally, e.g. by cytoplasmic micro-injection of macromolecules properly targeted for nuclear import. We would predict that highly negatively charged macromolecules should become specifically trapped in the lCD-space. It should also become possible to demonstrate different diffusion kinetics of molecules within the lCD-space as compared to the chromosomal domain interior, exploiting the reasoning that the kinetics of macromolecules should be strongly affected by their size and net charge. New technologies, which allow the fluorescent in vivo labelling of proteins (Chalfie et al. 1994; Wang & Hazelrigg 1994) and possibly in vivo targeting of specific nucleic acids, are urgently required to study the dynamics of nonchromatin domain and chromosomal domain topology. One should note in this context that the resolution of present 3D light microscopic approaches is not sufficient to study the 3D structure of small chromosomal domains, such as compacted chromatin loop domains, comprising some 100 kb of DNA or less. This limitation should be overcome by new types of far field light microscopy with considerably enhanced resolution (Cremer & Cremer 1978; Hell et al. 1994a, 1994b; Hänninen et al. 1995). Acknowledgments We are indebted to our colleagues Constanze Bonifer, Luitzen de Jong, Renato Paro, Roel van Driel and Daniele Zink for stimulating discussions and comments on various versions of this manuscript. While they deserve credit for their advice to improve aspects of the manuscript, we retain full responsibility for its inadequacies. Essential features of the lCD-compartment model were presented at the Kew Chromosome Conference 1V. The line of reasoning presented here also takes into account lectures and discussions at the Keystone Symposium "The nuclear matrix: involvement in replication, transcription, gene splicing, and cellular regulation", which was held from 410 April 1995 at Hilton Head Island, South Carolina. Our work has been supported by grants from the Deutsche Forschungsgemeinschaft and the European Community.
78 T. Cremer et al.

References Ashkin, A. and J.M. Dzied.zic 1989. Internal cell manipulation using infra-red laser traps. Proc. Nail. Acad. Sci. USA 86, 7914-7918. Berezney, R. 1991a. The nuclear matrix: a heuristic model for investigating genomic organization and function in the cell nucleus. j Gell Biochem. 47, 109-123. Berezney, R. 1991b. Visualizing DNA replication sites in the cell nucleus. Semin. Gell Biol. 2, 103-115. Berezney, R. and D.S. Coffey 1974. Identification of a nuclear protein matrix. Biochem. Biophys. Res. Gommun. 60,1410-1417. Berezney, R. and D.S. Coffey 1975. Nuclear protein matrix: Association with newly synthesized DNA. Science 189, 291-293. Bischoff, A., J. Albers, I. Kharboush, E. Stelzer, T. Cremer, and C. Cremer 1993. Differences in size and shape of active and inactive X-chromosome domains in human amniotic fluid cell nuclei. Microsc. Res. Tech. 25, 68-77. Blobel, G. 1985. Gene gating: A hypothesis. Proc. Natl. Acad. Sei. USA 82, 8527-8529. Bonifer, C., A. Hecht, H. Saueressig, D.M. Winter and A.E. Sippel 1991. Dynamic chromatin: the regulatory domain organization of eukaryotic gene li. J. Gell Biochem. 47, 99-108. Boulikas, T.C. 1992. Evolutionary consequences of non-random damage and repair of chromatin domains. j Mol. EvoL 35, 156-180. Boveri, T. 1909. Die Blastomerenkerne von Ascaris megalocephala und die Theorie der Chromosomenindividualitãt. Arch. Zellforschung 3, 181-268. Chai, L.S. and A.A. Sandberg 1988. Chromosomes and their relationship to nuclear components during the cell cycle in Chinese hamster cells. Gell Tissue Res. 251, 197-204. Chalfie, M., Y. Tu, G. Euskirchen, W.W. Ward and D.C. Prasher 1994. Green fluorescent protein as a marker for gene expression. Science 263, 802-805. Comings, D.E. 1968. The rationale for an ordered arrangement of chromatin in the interphase nucleus. Am. j Hum. Genet. 20, 440-460. Cook, P.R. 1988. The nucleoskeleton: Artefact, passive framework or active site?j Gell Sei. 90, 1-6. Cook, P.R. 1991. The nucleoskeleton and the topology of replication. Gell 66, 627-635. Cremer, C. and T. Cremer 1978. Consideration on a laser-scanning-microscope with high resolution and depth of field. Microscopica Acta 81, 31-44. Cremer, T., P. Lichter,J. Borden, D.C. Ward and L. Manuelidis 1988. Detection of chromosome aberrations in metaphase and interphase tumor cells by in situ hybridization using chromosome-specific library probes. Hum. Genet. 80, 235-246. Cremer, T., A. Kurz, R. Zirbel, S. Dietzel, B. Rinke, E. Schröck, M.R. Speicher, U. Mathieu, A. Jauch, P. Emmerich, H. Scherthan, T. Ried, C. Cremer and P. Lichter 1993. The role of chromosome territories in the functional compartm
Nuclear architecture and function 79

Eils, R., S. Dietzel, E. Bertin, M. Granzow, E. Schröck, M. R. Speicher, T. Voim, T. Ried, M. Robert-Nicoud, C. Cremer and T. Cremer 1995c. Three-dimensional reconstruction of painted human interphase chromosomes: active and inactive X-chromosome territories have similar volumes but differ in surface and shape. Submitted. EissenbergJ.C. and SCR. Elgin 1991. Boundary functions in the control of gene expression. TIG. 7:335-340. Felsenfeld, G. 1978. Chromatin. Nature 271, 115-122. Getzenberg, RH., KJ. Pienta, W.S. Ward and D.S. Coffey 1991. Nuclear structure and the threedimensional organization of DNA. J Cell Biochem. 47, 289-299. Giulian, D. and E.G. Diacumakos 1977. The electrophysiological mapping of compartments within a mammalian cell. Exp. Cell Res. 72, 86-103. Hãnninen, P.E., S.W. Hell, J. Salo, E. Soini and C. Cremer 1995. Two-photon excitation Vi confocal microscope: Enhanced axial resolution microscope for biological research. AppI. Phys. Lett. 66, 1698-1700. Hassan, A.B., R.J. Errington, N.S. White, D.A. Jackson and P.R. Cook 1994. Replication and transcription sites are co-localized in human cells. J. Cell Sei. 107, 425-434. He, D., D. Turner, B. Scott and BR. Brinkley 1995. The nucleoskeleton and chromatin organization: movements, alignment and association of centromeres in interphase involves the nuclear matrix core filaments. J. Cell. Biochem. Suppi. 21B, 138 (abstract). Hell, SW., S. Lindek, C. Cremer and E.H.K. Stelzer 1994a. Measurement of the 4Pi-confocal point spread function proves 75 um axial resolution. Appl. Phys. Lett. 64, 1335-1337. Hell, SW., E.H.K. Steizer, S. Lindek and C. Cremer 1994b. Confocal microscopy with an increased detection aperture: Type-B 4Pi confocal microscopy. Optics Letters 19, 222-224. Ingber, D.E. 1993. Cellular integrity: defining new rules of biological design that govern the cytoskeleton.J. Cell Sei. 104, 613-627. Ingber, D.E. and A.J. Mariotis 1995. Demonstration of mechanical continuity between cell surface integrins, cytoskeletal filaments and nuclei in living cells. J. Cell Biochem. Sup-pt. 21B, 116 (abstract). Jackson D.A., J. Yuan and P.R. Cook 1988. A gentle method for preparing cyto- and nucleoskeleton and associated chromatin. j Cell Sd. 90, 365-378. Kramer, J., Z. Zachar and P.M. Bingham 1994. Nuclear pre-mRNA metabolism: Channels and tracks. Trends in Cell Biol. 4, 35-37. Kurz, A., S., Lampel,J., Nikolencko,J., Bradl, A., Benner, K., Zirbel, T., Cremer, and P. Lichter 1995. Active and inactive genes localize in the periphery of chromosome territories. Submitted. Lawrence, J.B., K.C. Carter and X. Xing 1993. Probing functional organization within the nucleus: Is genome structure integrated with RNA metabolism? Cold Spring Harb. Symp.Quant. Biol. 58, 807-818. Leitch, AR., W. Mosgoller, T. Schwarzacher, M.D. Bennett and J.S. Heslop-Harrison 1990. Genomic in situ hybridization to sectioned nuclei shows chromosome domains in grass hybrids. j Cell Sei. 95, 335-341. Lichter, P., T. Cremer, J. Borden, L. Manuelidis and D.C. Ward 1988. Delineation of individual human chromosomes in metaphase and interphase cells by in situ suppression hybridization using recombinant DNA libraries. Hum. Genet. 80, 224-234. Loewenstein, W.R. and Y. Kanno 1963. The electrical conductance and potential across the membrane of some cell nuclei. J. Cell. Biol. 16, 421-425. Manuelidis L. 1985. Individual intephase chromosome domains revealed by in situ hybridization. Hum. Genet. 71, 288-293. Manuelidis, L. 1990. A view of interphase chromosomes. Science 250, 1533-1540. Mattem, -A., R. van Driel and L. de Jong 1995. Composition and structure of the internal nuclear matrix. In Nuclear structure and gene expression, R.C. Bird et al., eds. Academic Press, in press. 80 T. Cremer et al.

Mirkovitch, J., M.-E. Mirault and U.K. Laemmli 1984. Organization of the higher-order chromatin loop: specific DNA attachment sites on nuclear scaffold. Cell 39, 223-232. Mullenders, L.H.F., A.C. van Kesteren van Leeuwen, A.A. van Zeeland and AT. Natarajan 1988. Nuclear matrix associated DNA is preferentially repaired in normal human fibroblasts exposed to a low dose of ultraviolet light but not in Cockayne's syndrome fibroblasts. Nuci. Acid. Res. 16, 10607-10622. Oberleitner, H., B. Schuricht, S. Wünsch, S. Schneider and B. Püschel 1993. Role of H+ ions in volume and voltage of epithelial cell nuclei. Ear j Physiol. 423, 88-96. Penman, S. 1995. Re-thinking cell structure. Proc. Natl. Acad. Sei. USA 92, 5251-5257. Pinkel, D., J. Landegent, C. Collins, J. Fuscoe, R. Segraves, J. Lucas and J.W. Gray 1988. Fluorescence in situ hybridization with human chromosome-specific libraries: detection of trisomy 21 and translocations of chromosome 4. Proc. Nail. Acad. Sci. USA 85, 9138-9142. Rabl, C. 1885. Über Zelltheilung. Morphol. Jahrbuch 10, 214-330. Rann, S.V. and 1.1. Gromova 1995. The channels model of nuclear matrix structure. BioEssays 17, 443-450. Rinke, B., A. Bischoff, M.C. Meffert, R. Scharschmidt, M. Hausmann, E.H.K. Stelzer, T. Cremer and C. Cremer 1995. Volume ratios of painted chromosome territories 5, 7 and X in female human cell nuclei studied with confocal laser microscopy and the Cavalieri estimator. Bioimaging3, 1-11. Roth, M.B. 1995. Spheres, coiled bodies and nuclear bodies. Curr. Opin. Cell Biol. 7, 325-328. Saitoh, Y. and U. K. Laemmli 1993. From the chromosomal loops and the scaffold to the classic bands of metaphase chromosomes. Cold Spring Harbor Symp. Quant. Biol. 58, 755-765. Schardin, M., T. Cremer, H.D. Hager and M. Lang 1985. Specific staining of human chromosome position in chinese hamster x man hybrid cell lines demonstrates inter-phase chromosome territorries. Hum. Genet. 71, 281-287. Schedl, P., and F. Grosveld 1995. Domains and boundaries. In Chromatin structure and gene expression, C.R. Elgin, cd. Oxford University Press, pp. 172-196. Spector, D.L. 1990. Higher order nuclear organization: Three-dimensional distribution of small nuclear ribonucleoprotein particles. Proc. Natl. Acad. Sei. USA 87, 147-151. Spector, D.L. 1993. Macromolecular domains within the cell nucleus. Ann. Rev. Cell Biol. 9, 265-315. van Driel, R., D.G. Wansink, B. van Steensel, M.A. Grande, W. Schul and L. de Jong 1995. Nuclear domains and the nuclear matrix. Int. Rev. CytoL, in press. Vogel, F. and T.M. Schroeder 1974. The internal order of the interphase nucleus. Humangenetik25,265-297. Wang, S. and T. Hazelngg 1994. Implications for bcd mRNA localization from spatial distribution of exu protein in Drosophila oogenesis. Nature 369, 400-403. Wansink, D.G., L. de Jong and R. van Driel 1995. Transcription by RNA polymerase II and nuclear architecture. In Nuclear structure and gene expression, R.C. Bird ei al., eds. Academic Press, in press. Wansink, D.G., R. van Driel and L. dc Jong 1994. Organization of (pre) mRNA metabolism in the cell nucleus. Mol. Biol. Reports 20, 45-55. Weintraub, H. 1993. Genetic tinkering - local problems, local solutions. Cold Spring Harbor Symp.Quant. Biol. 58, 819-836. Weimer, R., T. Haaf, M. Poot and M. Schmid 1992. Characterization of centromere arrangements and test for random distribution in GO, GI, S, G2, Gi' and early 5' phase in human lymphocytes. Hum. Genet. 88, 673-682. Wischnitzer, 5. 1973. The submicroscopic morphology of the interphase nucleus. Int. Rev. Cytol.34,1-48. Zachar, Z., J. Kramer, I.P. Mims and P.M. Bingham 1993. Evidence for channeled diffusion of pre-mRNAs during nuclear RNA transport in metazoans. j Cell Biol. 121, 729-742.
Nuclear architecture and function 81

Zehnbauer, B.A. and B. Vogelstein 1985. Supercoiled loops and the organization of replication and translocation in eukaryotes. BioEssays 2, 52-54. Zhao, K., C. M. Hart and U.K Laemmli 1995. Visualization of chromosomal domains with boundary element-associated factor beaf-32. Cell 81, 879-889. Zirbel, R.M., U.R. Mathieu, A. Kurz, T. Cremer and P. Lichter 1993. Evidence for a nuclear compartment of transcription and splicing located at chromosome domain boundaries. Chromosome Res. 1,93-106.