cones regenerate from retinal stem cells sequestered in...
TRANSCRIPT

Cones Regenerate from Retinal Stem Cells Sequesteredin the Inner Nuclear Layer of Adult Goldfish Retina
David M. Wu,1 Todd Schneiderman,2,3 Jason Burgett,2,4 Parag Gokhale,2,5 Linda Barthel,6
and Pamela A. Raymond1,6
PURPOSE. To determine whether retinal progenitor cells in theinner nuclear layer give rise to regenerated cones after laserablation of photoreceptors in adult goldfish retina.
METHODS. Using a technique developed previously in this lab-oratory, photoreceptors in the retina of adult goldfish wereablated with an argon laser. The mitotic marker, bromode-oxyuridine, was used to label proliferating and regeneratedcells, which were identified with cell-specific markers.
RESULTS. Cells proliferating locally within lesion included mi-croglia, Muller glia, and retinal progenitors in the inner nuclearlayer (INL). The nuclei of both Muller glia and associated retinalprogenitors migrated from the inner to the outer nuclear layer.The proliferating retinal progenitors, which express Notch-3and N-cadherin, regenerated cone photoreceptors and thenrod photoreceptors.
CONCLUSIONS. Previous work has demonstrated that photore-ceptors in the goldfish retina regenerate selectively after laserablation, but the source of regenerated cones has not beenidentified. The results reported here provide support for theexistence of retinal stem cells within the adult fish retina thatare capable of regenerating cone photoreceptors. The data alsosupport the involvement of Muller glia in the production ofregenerated cones. (Invest Ophthalmol Vis Sci. 2001;42:2115–2124)
The neural retina in adult teleost fish contains several dis-tinct populations of proliferating retinal progenitor cells,
and neurogenesis continues in adult fish as part of an ongoinggrowth process.1,2 These retinal progenitor cells have beenidentified in the adult retina of several different species ofteleost fish,3–9 by labeling their nuclei with an antibody againstproliferating cell nuclear antigen (PCNA) or tagging them witha pulse-label of [3H]thymidine or the thymidine analogue bro-modeoxyuridine (BrdU). At the circumferential (ciliary) marginof the retina, a germinal zone consisting of multipotent retinalprogenitors produces annuli of new retina that are added
appositionally, similar to the rings of new growth in a treetrunk.1,10 The number of neurons added postembryonically tothe growing retina is substantial. For example, the retina gen-erated during embryonic stages in goldfish occupies less than5% of the retinal area present in a 2-year-old fish.11
Rod photoreceptors are the last retinal neurons to be gen-erated in the fish retina, and they are added in a second waveof mitotic activity that occurs in specialized rod precursors, notin the circumferential germinal zone, but located within theouter nuclear layer (ONL).3, 12–14 Rod precursors are mostabundant near the germinal zone, in the region of newlydifferentiated retina, but they are also found within the ONLand scattered across the entire differentiated retina. The pur-pose of interstitial addition of rod photoreceptors is to main-tain a constant planar density of rods as the retina enlarges bystretching.4,13,15 These mitotically active rod precursor cells inthe ONL have a restricted lineage, in that they give rise only torods in the intact retina.3,4,16 Given this readily available sourceof retinal progenitors dedicated to the production of rods, it isnot surprising that rods can regenerate when they are selec-tively destroyed in goldfish retina and that the selective regen-eration of rod photoreceptors is due to enhanced mitoticactivity of rod precursors in the ONL.17
Although it has been 20 years since proliferating rod pre-cursors were described in the outer nuclear layer in the adultgoldfish retina, the literature also contains repeated referencesto rare, mitotically active cells in the inner nuclear layer (INL)of the intact (undamaged) retina in larval or juvenile fish,7,13
but only recently have these cells been identified with cer-tainty in adult fish. Because they divide very slowly, they can bereliably labeled only by sustained exposure to mitotic markerssuch as BrdU.8,18,19 The mitotic progeny of these dividing cellsmigrate radially outward into the ONL, where they contributeto the ongoing neurogenesis in the adult retina by replenishingthe population of rod precursors.8 These dividing retinal pro-genitors sequestered in the INL represent a continuation, buton a slower time scale, of the final stages of neurogenesis in thelarval retina. Earlier studies in larval goldfish retina using lon-gitudinal analysis of [3H]thymidine-labeled cells and electronmicroscopic autoradiography with serial reconstructionshowed that rod precursors originate from residual multipo-tent progenitor cells in the INL, whose progeny migrate alongthe radial fibers of Muller glia to reach the ONL.20 Previousstudies have also shown that some retinal cells in the INLproliferate in response to mechanical damage or cytotoxicinsult, and they have often been observed in studies of retinaldamage and/or regeneration.21–27 Although some of these areMuller cells,28 the study of Julian et al.,8 which demonstratedthe existence of progenitor cells in the INL of the undamagedadult fish retina, suggests that some of these may be retinalstem cells.
The proliferating cells in the INL have many of the charac-teristics of stem cells found in other adult tissues.29,30 Forexample, they divide slowly to both self-renew and replenish apopulation of more specialized progenitor cells (rod precur-sors) that have a higher rate of mitotic activity, and theyrespond to tissue damage and cell loss by enhanced prolifera-
From the 1Neuroscience Graduate Program, the 2Department ofOphthalmology and Visual Science, and the 6Department of Cell andDevelopmental Biology, University of Michigan Medical School, AnnArbor.
3Present address: 20696 Bond Road NE, Poulsbo, WA.4Present affiliation: Burgett Eye Care, Lafayette, Indiana.5Present affiliation: Department of Ophthalmology, Medical Col-
lege of Georgia, Augusta.Pamela A. Raymond has published previously as Pamela R. Johns.Supported by National Institutes of Health Grant EY04318 (PAR).Submitted for publication January 5, 2001; revised March 9, 2001;
accepted March 29, 2001.Commercial relationships policy: N.The publication costs of this article were defrayed in part by page
charge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Corresponding author: Pamela A. Raymond, Department of Celland Developmental Biology, 4610 Medical Science II, University ofMichigan Medical School, Ann Arbor, MI [email protected]
Investigative Ophthalmology & Visual Science, August 2001, Vol. 42, No. 9Copyright © Association for Research in Vision and Ophthalmology 2115
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

tion, the purpose of which is to restore the lost cells. Anotherfeature characteristic of adult stem cells is their multipo-tency—that is, their ability to generate a diversity of specificcell types. However, the only certain fates of the progenitorcells in the INL of the intact, undamaged retina are that theygive rise to the rod precursors in the ONL and they self-renew.8
Although their existence was not known at the time of earlierstudies on retinal regeneration,17,21,22,25 and the rod precur-sors were originally suspected to be the retinal progenitorsresponsible for regenerating the neural retina, in retrospect,these putative retinal stem cells in the INL are the more likelycandidates to give rise to the regeneration blastema at thewound margin of the surgically lesioned retina and the neuro-genic clusters in retinas damaged by cytochemical toxins orlaser lesions.31,32
To test the hypothesis that the retinal progenitor cells in theINL have an important property expected in stem cells—thatis, that they can regenerate damaged retinal tissue by replacingretinal neurons—we challenged them in a selective regenera-tion paradigm, in which rods and cones in the adult goldfishretina were destroyed with an argon laser.17 Both cones androds regenerated in the lesioned area, and we asked whetherthe regenerated cone photoreceptors derive from mitotic cellsresident in the INL and whether these mitotic cells expressspecific markers characteristic of the multipotent retinal pro-genitor cells in the embryonic retina and in the germinal zoneat the ciliary margin.
METHODS
Animals
We obtained adult goldfish (Carassius auratus), with body lengths of6 to 8 cm and eye diameters of 5.0 to 6.5 mm, from a commercialfishery (Ozark Fisheries, Stoutland, MO). All procedures involvinganimals conformed to the ARVO Statement for the Use of Animals inOphthalmic and Vision Research.
Lentectomies
Because the goldfish lens is optically adapted to aquatic environments(and all the refractive power of the eye is in the lens), when the fish istaken out of the water, the retina is easier to visualize after lentec-tomy.17 Fish were anesthetized in 0.1% tricaine methane sulfonate(Sigma, St Louis, MO) and an incision was made in the nasal corneawith a 1.5-mm, 15° microscalpel (Becton-Dickinson, Franklin Lakes,NJ), and extended along a crescent by cutting the cornea on the dorsaland ventral margins with microscissors. The temporal edge of thecornea remained attached to the eye, and the cornea was reflectedaway, revealing the lens. The ligaments holding the lens in place werecarefully snipped with microscissors, and the lens was freed from itsattachments and extracted with forceps. The cornea was then foldedback over the pupil, and the fish was returned to water. Suturing wasnot necessary, and when attempted, it interfered with corneal clarity.A postoperative healing period of at least 2 weeks was allowed for thecornea to seal and clear.
Laser Lesions
We used an argon laser (System 920; Coherent, Palo Alto, CA) toproduce photocoagulatory lesions in the retinas, as described previ-ously.17 Fish were anesthetized and manually held in position, to viewthe fundus with an ophthalmoscope. We made a series of four lesionsin the nasal–ventral quadrant of the retina, with the closest lesionpositioned less than 1 mm away from the optic disc. The lesions werearranged in a square with sides that were approximately four lesion-diameters in length. The parameters of the laser were set at a durationof 0.1 second, a spot size of 500 mm, and a power setting of 80, 100,or 130 mW. The goal was to produce lesions that destroyed photore-
ceptors but spared the inner retina (inner nuclear and ganglion celllayers).17 Most of the laser energy is absorbed by the melanin in thepigmented retinal epithelium, and the proximity of the photoreceptorsmakes them more vulnerable. We chose the laser power to use foreach eye after examining corneal clarity, because we had determinedempirically that greater powers were needed to produce the desiredlesion when the cornea was less transparent. In the ophthalmoscope,lesions appeared as white patches on the retina, and the more pow-erful burns produced larger and whiter patches. The actual tissuedamage caused by the laser, as assessed later by histology, varied withcorneal clarity, vitreal clarity, and other unidentified sources of indi-vidual variability. Retinas were selected for study when the retinaldamage from the laser was largely restricted to the ONL.
Intraocular Injections
To identify mitotically active cells and their progeny, the thymidineanalogue 5-bromo-29-deoxyuridine (BrdU; Sigma) was injected in-traocularly (1 mM in 0.9% NaCl). A small incision was made in the nasalcornea, and a blunt-tipped, 33-gauge needle attached to a microsyringe(Hamilton Company, Reno, NV) was inserted into the vitreal cavity.The amount injected was calculated to produce an approximate con-centration of 50 mM BrdU in the vitreous, based on estimates of eyevolume.22 Tissue was processed from 1 hour to 115 days after the BrdUinjections.
Tissue Processing
Fish were anesthetized and decapitated. The eyes were enucleated andprocessed for cryosectioning as described previously.33 For whole-mount preparations, fish were dark adapted for 2 hours before death tofacilitate separation of the neural retina from the pigmented retinalepithelium.
Immunocytochemistry
For immunofluorescence, cryosections (5-mm thickness) were pre-pared as described previously.17 Antibodies used included: RET1 (1:500), a monoclonal antibody generated against goldfish retina thatlabels an unidentified nuclear epitope found in cones (but not rods),horizontal cells, a subset of INL neurons, Muller glia, and ganglioncells34; NN2, a monoclonal antibody (1:1000) generated against gold-fish retina that labels an unknown cell-surface antigen on microglia andendothelial cells28; rabbit polyclonal antibodies against goldfish glialfibrillary acidic protein (FGP1, 1:500), a generous gift of MichalSchwartz (Weizmann Institute, Rehovot, Israel); rat anti-BrdU (1:20;Accurate Chemical, Westbury, NY); anti-glutamine synthetase (GS,1:50) a generous gift of Paul Linser (University of Florida, Gainesville);and the zpr1 monoclonal antibody (1:500) that recognizes an unchar-acterized surface epitope on double cones (from the zebrafish mono-clonal stock center at the University of Oregon Institute of Neuro-science, Eugene, OR).
After overnight incubation in primary antibody at 4°C, slides wererinsed and then incubated overnight at 4°C in secondary antibodiesconjugated to fluorochromes including 7-amino-4-methylcoumarin-3-acetic acid (AMCA; blue), fluorescein isothiocyanate (FITC; green), andCY3 (red-orange); secondary antibodies were from Jackson ImmunoRe-search Laboratories (Westgrove, PA). Alternatively, primary antibodieswere visualized with an ABC peroxidase kit (Vectastain; Vector Labo-ratories, Burlingame, CA), according to the manufacturer’s instruc-tions, except that incubation times were increased to overnight for thesecondary antibody and to 4 to 6 hours for the avidin-biotin complex.The enzyme substrate was 3,39-diaminobenzidine tetrahydrochloride(Sigma). For detection of BrdU in the nuclei, sections were labeledwith rat anti-BrdU antibody, as described previously,34 except thatsecondary antibodies were conjugated to CY3. Wholemount retinaswere also processed using this protocol, except that the primaryantibody concentrations were 1:20, primary incubations were 40 to 50hours in duration, secondary incubations were overnight, and rinsesteps were increased to at least 30 minutes.
2116 Wu et al. IOVS, August 2001, Vol. 42, No. 9
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

In some tissue sections, the number of cells expressing theseantibody markers was quantified. All cells designated as double-labeledwith two different antibody probes were verified by examination ofeach fluorescent signal under single channel illumination. An analysisof variance (ANOVA), with a Tukey-Kramer posttest (SAS Institute,Cary, NC) was used to compare data. A schematic drawing of a retinalsection (Fig. 1A) illustrates the criteria used for defining the bounds ofthe lesion (see the figure legend for more information). Labeled cellswere counted in 23 laser-lesioned eyes.
In Situ Hybridization
Cryosections were processed for in situ hybridization with digoxige-nin-labeled cRNA probes, as described previously,35,36 with proteinaseK treatment for 2 minutes and 45 seconds. The signals were visualizedby alkaline phosphatase histochemistry with 4-nitroblue tetrazoliumchloride and 5-bromo-4-chloro-3-indolyl phosphate solution (NBT-BCIP) as the substrate. The cRNA probes were generated according toprotocols in the instruction manual from Roche Molecular Biochemi-cals (formerly Boehringer-Mannheim Biochemicals; Indianapolis, IN).The zebrafish N-cadherin (cdh2) antisense probe was generated froma full-length cDNA in the plasmid pBS (the generous gift of BenjaminGeiger, Weizmann Institute, Israel), linearized with the restrictionenzyme HndIII, and transcribed with T7 RNA polymerase. The goldfishNotch-3 antisense cRNA probe was a mixture of two probes generatedfrom partial-length cDNA clones (Gotch TM and Gotch CDC), as de-scribed previously.37 Sense control probes were prepared for allclones.
RESULTS
Identification of the Lesioned Area
The argon laser permits the accurate placement of focusedphotocoagulation lesions that can be limited to the outer reti-nal layers.17,38,39 Figure 1B shows a wholemount preparationof a retina at 115 days after lesion, mounted vitreal surface up.Circular patches of BrdU immunoreactivity mark the four le-sions. In this preparation, the proliferating cells were labeledwith BrdU during the first week after the lesion. A highermagnification of one of the lesions, double immunolabeled forBrdU and glial fibrillary acidic protein (GFAP), is shown inFigure 1C. The end feet of the radial processes of the Mullerglia at the inner limiting membrane can be visualized with theFGP1 polyclonal antibodies prepared against goldfish GFAP.The BrdU-labeled cone nuclei, out of focus because they wereat a deeper focal plane, mark the lesion site. In the uninjuredretina surrounding the lesion, Muller glial end feet were dis-tributed in a regular array with uniform spacing, but within thelesion the end feet were disorganized. The radial processes ofthe Muller cells were pulled toward the center of the lesion, adistortion that is consistent with the defect in the outer retinathat resulted from the photocoagulation (Fig. 1A).
Proliferation of Cone Progenitorsafter Laser Lesion
To identify the progenitors of the regenerated cones, we firsthad to determine at what point the regenerated cones wereproduced after the lesion and then to systematically identifythe progenitor cells that were proliferating in the lesion areaduring that period and follow them at subsequent intervals todetermine which may give rise to cones. Our earlier study ofcone regeneration in laser-lesioned goldfish retina17 had shownthat BrdU injected within the first week after laser lesioning(3–8 days) is incorporated into cells that differentiate intocones. The results of the present study confirm the previousfindings. Figure 1D illustrates BrdU-labeled, regenerated conesfrom a retina in which BrdU was injected into the eye twice, at
5 and 8 days after laser lesion and the retina processed forimmunocytochemistry at 20 days, at which time, the regener-ating cones had begun to differentiate. The section was triple-labeled with RET1 (blue), anti-GFAP (green), and anti-BrdU(red). The RET1 monoclonal antibody labeled the nuclei ofsome retinal cells in all three cellular layers of the retina,including cones (but not rods), subsets of neurons in the INL,Muller glia, and retinal ganglion cells.34 The GFAP antibodylabeled the radial processes of the Muller glia, as described.Within the lesion, the nuclei of regenerated cones were dou-ble-labeled with RET1 and BrdU, which were both localized tothe nucleus, and the double-labeled nuclei were therefore pinkto pale violet (inset, Fig. 1D). Additional evidence that theRET1/BrdU-labeled nuclei were regenerated cones is that thesecells had begun to elaborate apical processes (inner and outersegments) that were characteristic of photoreceptors, butsmaller than the adjacent (surviving) cones (data not shown;see reference 17). In summary, both histologic and immuno-cytochemical observations suggest that regenerated coneswere produced by mitotic progenitors that were dividing dur-ing the first week after laser ablation.
A few BrdU-labeled nuclei were also located in the INL ofthese retinas at 20 days after laser lesioning (Fig. 1D). Some ofthese nuclei were double labeled with RET1 and were associ-ated with GFAP-positive fibers (Fig. 1D, and data not shown),suggesting that they were nuclei of Muller cells.28 However,other BrdU-labeled nuclei in the INL showed no RET1 immu-noreactivity, suggesting that they were not Muller glia. Thesecells may have been regenerated INL neurons, because wecannot exclude the possibility that the laser damaged cells inthe INL, some of which do not express the RET1 antigen(discussed later). Alternatively, they may have been undiffer-entiated INL progenitors, because one of the hallmarks of stemcells is a capacity to self-renew.
Some support for this latter suggestion comes from thefollowing observations. When BrdU was injected once at either3 or 5 days after laser lesion and the retina was processed at 25days after lesion, cone nuclei were also labeled with BrdU.However, the BrdU label was weaker than it was after twoBrdU injections, in that the label did not fill the nuclei but wasinstead granular in appearance (data not shown). This obser-vation suggests that the cells giving rise to cones at 3 to 5 daysunderwent several cell divisions before differentiating intocones. In contrast, some cells in the INL at 20 to 25 days hadstrong BrdU labeling after a single injection at 3 or 5 days,suggesting that they underwent one or at most only a fewmitotic divisions, which would be typical of stem cell self-renewal (data not shown).
Regeneration of Rods
Although most of the progeny of cells proliferating during thefirst week differentiated as cones, a few regenerated rods werealso labeled with BrdU, and the magnitude of cone genesisrelative to rod genesis decreased with time. For example,Figure 1E shows a retina processed at 30 days after the lesion,after BrdU injections at 14 and 17 days. Fewer cone than rodnuclei were BrdU labeled. The rods did not express the RET1epitope and were therefore not double labeled. They wereidentified by their position in the ONL, their small oval nuclei,and the absence of RET1 staining (inset, Fig. 1E). This temporalpattern of photoreceptor genesis during regeneration is remi-niscent of the order of cell production during normal develop-ment, in that cones are born before rods.17,20,40
Progenitors Giving Rise to Regenerated Cones
The foregoing results are consistent with our earlier study,17
which demonstrated that most regenerated cones are pro-
IOVS, August 2001, Vol. 42, No. 9 Cone Regeneration and Retinal Stem Cells 2117
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018
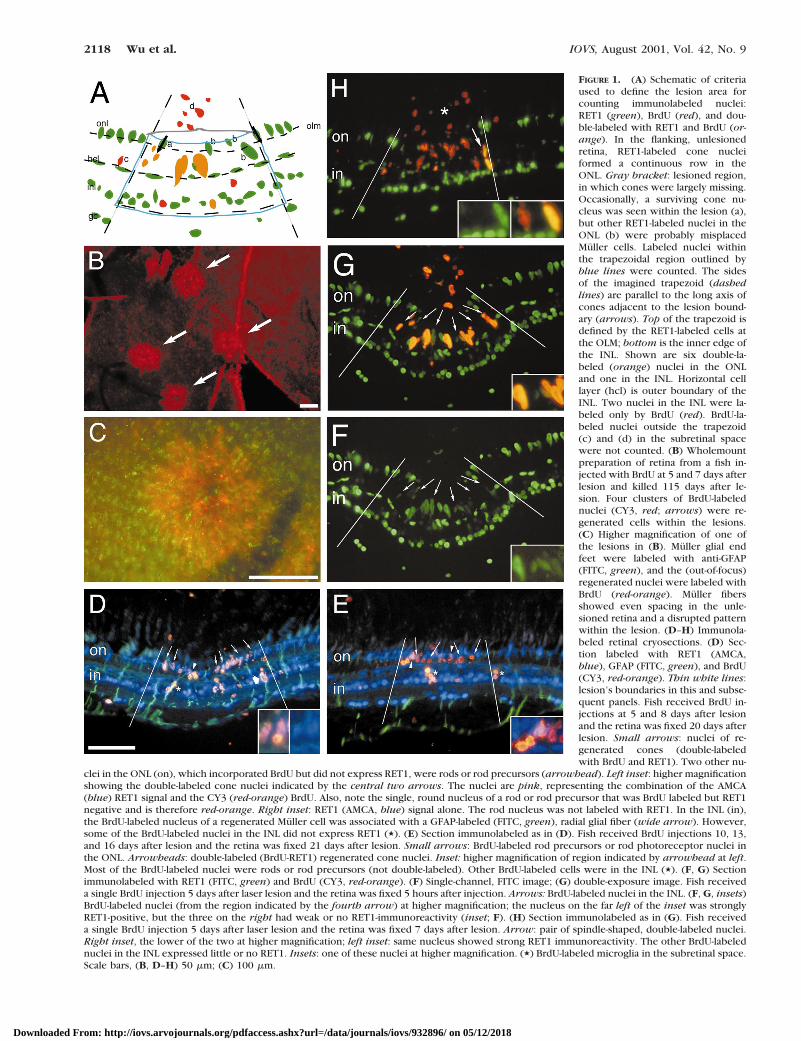
FIGURE 1. (A) Schematic of criteriaused to define the lesion area forcounting immunolabeled nuclei:RET1 (green), BrdU (red), and dou-ble-labeled with RET1 and BrdU (or-ange). In the flanking, unlesionedretina, RET1-labeled cone nucleiformed a continuous row in theONL. Gray bracket: lesioned region,in which cones were largely missing.Occasionally, a surviving cone nu-cleus was seen within the lesion (a),but other RET1-labeled nuclei in theONL (b) were probably misplacedMuller cells. Labeled nuclei withinthe trapezoidal region outlined byblue lines were counted. The sidesof the imagined trapezoid (dashedlines) are parallel to the long axis ofcones adjacent to the lesion bound-ary (arrows). Top of the trapezoid isdefined by the RET1-labeled cells atthe OLM; bottom is the inner edge ofthe INL. Shown are six double-la-beled (orange) nuclei in the ONLand one in the INL. Horizontal celllayer (hcl) is outer boundary of theINL. Two nuclei in the INL were la-beled only by BrdU (red). BrdU-la-beled nuclei outside the trapezoid(c) and (d) in the subretinal spacewere not counted. (B) Wholemountpreparation of retina from a fish in-jected with BrdU at 5 and 7 days afterlesion and killed 115 days after le-sion. Four clusters of BrdU-labelednuclei (CY3, red; arrows) were re-generated cells within the lesions.(C) Higher magnification of one ofthe lesions in (B). Muller glial endfeet were labeled with anti-GFAP(FITC, green), and the (out-of-focus)regenerated nuclei were labeled withBrdU (red-orange). Muller fibersshowed even spacing in the unle-sioned retina and a disrupted patternwithin the lesion. (D–H) Immunola-beled retinal cryosections. (D) Sec-tion labeled with RET1 (AMCA,blue), GFAP (FITC, green), and BrdU(CY3, red-orange). Thin white lines:lesion’s boundaries in this and subse-quent panels. Fish received BrdU in-jections at 5 and 8 days after lesionand the retina was fixed 20 days afterlesion. Small arrows: nuclei of re-generated cones (double-labeledwith BrdU and RET1). Two other nu-
clei in the ONL (on), which incorporated BrdU but did not express RET1, were rods or rod precursors (arrowhead). Left inset: higher magnificationshowing the double-labeled cone nuclei indicated by the central two arrows. The nuclei are pink, representing the combination of the AMCA(blue) RET1 signal and the CY3 (red-orange) BrdU. Also, note the single, round nucleus of a rod or rod precursor that was BrdU labeled but RET1negative and is therefore red-orange. Right inset: RET1 (AMCA, blue) signal alone. The rod nucleus was not labeled with RET1. In the INL (in),the BrdU-labeled nucleus of a regenerated Muller cell was associated with a GFAP-labeled (FITC, green), radial glial fiber (wide arrow). However,some of the BrdU-labeled nuclei in the INL did not express RET1 (,). (E) Section immunolabeled as in (D). Fish received BrdU injections 10, 13,and 16 days after lesion and the retina was fixed 21 days after lesion. Small arrows: BrdU-labeled rod precursors or rod photoreceptor nuclei inthe ONL. Arrowheads: double-labeled (BrdU-RET1) regenerated cone nuclei. Inset: higher magnification of region indicated by arrowhead at left.Most of the BrdU-labeled nuclei were rods or rod precursors (not double-labeled). Other BrdU-labeled cells were in the INL (,). (F, G) Sectionimmunolabeled with RET1 (FITC, green) and BrdU (CY3, red-orange). (F) Single-channel, FITC image; (G) double-exposure image. Fish receiveda single BrdU injection 5 days after laser lesion and the retina was fixed 5 hours after injection. Arrows: BrdU-labeled nuclei in the INL. (F, G, insets)BrdU-labeled nuclei (from the region indicated by the fourth arrow) at higher magnification; the nucleus on the far left of the inset was stronglyRET1-positive, but the three on the right had weak or no RET1-immunoreactivity (inset; F). (H) Section immunolabeled as in (G). Fish receiveda single BrdU injection 5 days after laser lesion and the retina was fixed 7 days after lesion. Arrow: pair of spindle-shaped, double-labeled nuclei.Right inset, the lower of the two at higher magnification; left inset: same nucleus showed strong RET1 immunoreactivity. The other BrdU-labelednuclei in the INL expressed little or no RET1. Insets: one of these nuclei at higher magnification. (,) BrdU-labeled microglia in the subretinal space.Scale bars, (B, D–H) 50 mm; (C) 100 mm.
2118 Wu et al. IOVS, August 2001, Vol. 42, No. 9
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018
duced within the first 2 weeks after the lesion. To identify thecells that were dividing at the beginning of cone genesis, weinjected BrdU at 2 to 5 days after the laser lesion and processedthe retinas 1 to 5 hours after the injection. This paradigmallowed sufficient time for incorporation of BrdU, but notenough time for complete cell division or differentiation, and ittherefore labeled the cells that were proliferating at the time ofinjection. To trace the origin of the regenerated cones, weinjected BrdU during the first week after the lesion and thenobserved the locations of the labeled cells at intervals over thenext 2 weeks. The BrdU-labeled nuclei in each layer werecounted, and the proportion that expressed the RET1 antigenwas determined.
From previous studies we knew that retinal damage pro-duced by a number of different causes (e.g., mechanicaltrauma, chemical toxins, ischemia) produces a progressive
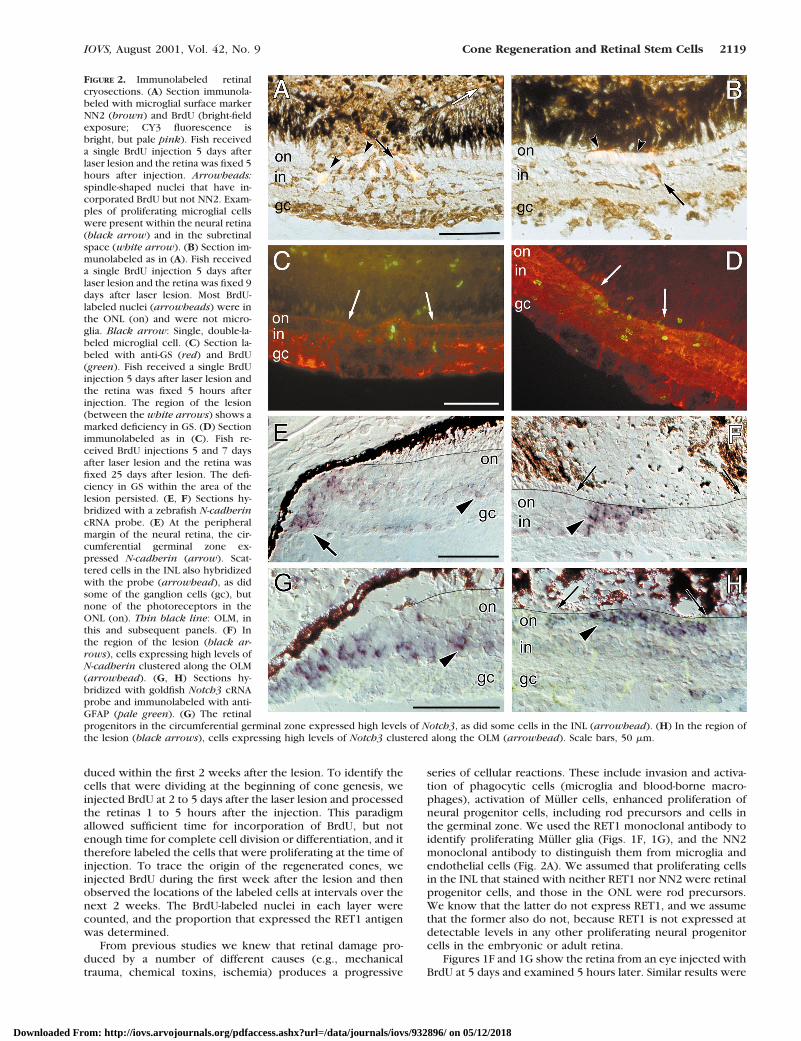
series of cellular reactions. These include invasion and activa-tion of phagocytic cells (microglia and blood-borne macro-phages), activation of Muller cells, enhanced proliferation ofneural progenitor cells, including rod precursors and cells inthe germinal zone. We used the RET1 monoclonal antibody toidentify proliferating Muller glia (Figs. 1F, 1G), and the NN2monoclonal antibody to distinguish them from microglia andendothelial cells (Fig. 2A). We assumed that proliferating cellsin the INL that stained with neither RET1 nor NN2 were retinalprogenitor cells, and those in the ONL were rod precursors.We know that the latter do not express RET1, and we assumethat the former also do not, because RET1 is not expressed atdetectable levels in any other proliferating neural progenitorcells in the embryonic or adult retina.
Figures 1F and 1G show the retina from an eye injected withBrdU at 5 days and examined 5 hours later. Similar results were
FIGURE 2. Immunolabeled retinalcryosections. (A) Section immunola-beled with microglial surface markerNN2 (brown) and BrdU (bright-fieldexposure; CY3 fluorescence isbright, but pale pink). Fish receiveda single BrdU injection 5 days afterlaser lesion and the retina was fixed 5hours after injection. Arrowheads:spindle-shaped nuclei that have in-corporated BrdU but not NN2. Exam-ples of proliferating microglial cellswere present within the neural retina(black arrow) and in the subretinalspace (white arrow). (B) Section im-munolabeled as in (A). Fish receiveda single BrdU injection 5 days afterlaser lesion and the retina was fixed 9days after laser lesion. Most BrdU-labeled nuclei (arrowheads) were inthe ONL (on) and were not micro-glia. Black arrow: Single, double-la-beled microglial cell. (C) Section la-beled with anti-GS (red) and BrdU(green). Fish received a single BrdUinjection 5 days after laser lesion andthe retina was fixed 5 hours afterinjection. The region of the lesion(between the white arrows) shows amarked deficiency in GS. (D) Sectionimmunolabeled as in (C). Fish re-ceived BrdU injections 5 and 7 daysafter laser lesion and the retina wasfixed 25 days after lesion. The defi-ciency in GS within the area of thelesion persisted. (E, F) Sections hy-bridized with a zebrafish N-cadherincRNA probe. (E) At the peripheralmargin of the neural retina, the cir-cumferential germinal zone ex-pressed N-cadherin (arrow). Scat-tered cells in the INL also hybridizedwith the probe (arrowhead), as didsome of the ganglion cells (gc), butnone of the photoreceptors in theONL (on). Thin black line: OLM, inthis and subsequent panels. (F) Inthe region of the lesion (black ar-rows), cells expressing high levels ofN-cadherin clustered along the OLM(arrowhead). (G, H) Sections hy-bridized with goldfish Notch3 cRNAprobe and immunolabeled with anti-GFAP (pale green). (G) The retinalprogenitors in the circumferential germinal zone expressed high levels of Notch3, as did some cells in the INL (arrowhead). (H) In the region ofthe lesion (black arrows), cells expressing high levels of Notch3 clustered along the OLM (arrowhead). Scale bars, 50 mm.
IOVS, August 2001, Vol. 42, No. 9 Cone Regeneration and Retinal Stem Cells 2119
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

obtained with survival times of 1 hour and/or injections at 3days, although proportionately more of the labeled nuclei werein the INL than in the ONL at 3 days than at 5 days (discussedlater). In the region of the lesion, the laser had typically de-stroyed all the photoreceptors and, we presume, all or most ofthe rod precursors (photoreceptors and rod precursors are theonly nuclei normally resident in the ONL). Within the lesionthere were many proliferating cells (Fig. 1G), in contrast to thesurrounding undamaged retina, in which only rare BrdU-la-beled nuclei were seen. The proliferating cells (BrdU-labelednuclei) that were present at 3 to 5 days in the INL, in the ONL,and between these two layers were heterogeneous in level ofexpression of RET1 immunoreactivity. Some were clearly RET1positive (consistent with an identity as Muller glia), but somehad little or no RET1 expression (insets, Figs. 1F, 1G).
During the first week after the lesion (3–7 days), the major-ity of BrdU-labeled nuclei in the INL expressed RET1 (Table 1,column 7). The RET1-positive, BrdU-labeled nuclei were pre-sumed to be Muller cells (Figs. 1F, 1G). Many of these cells hadirregular, spindle-shaped nuclei, consistent with their designa-tion as Muller cells.28,41 Subsequent studies with Muller-spe-cific antibodies supported this inference and demonstratedadditional cytological responses to the damage produced bythe laser lesion. Muller cells in the goldfish retina expressed abasal level of GFAP, which was upregulated in the lesionedarea, as we had observed previously.17 Antibodies to GS canalso be used to demonstrate Muller cell processes in the reti-na.42 By 5 days after the lesion, the level of expression of GS inMuller cells in the region of the lesion was downregulated (Fig.2C), and this local decrease in expression levels was stillapparent at 25 days (Fig. 2D). These alterations in expressionlevels of GS are consistent with previously observed behaviorof Muller cells that have lost their neuronal contacts.42,43
Many other proliferating cells in the INL during the firstweek after the lesion were RET1 negative. To determinewhether any of these were putative neural progenitors (i.e.,neither RET1-labeled Muller cells nor NN2-labeled microglia),we examined lesioned retinas labeled with BrdU at 2, 3, or 5days after the lesion and processed them for double immuno-cytochemistry after a 5-hour survival with a cocktail of RET1and NN2 monoclonal antibodies. At each time point, we wereable to find substantial numbers of BrdU-labeled nuclei in theINL that were not labeled with the RET1 and NN2 antibodies,suggesting that these cells were neither Muller glia nor micro-glia. Quantification of these data showed that proliferatingneural progenitor cells in the INL were more frequent inretinas at 5 days than at 2 days after lesion: Of the BrdU-labelednuclei in the INL at 5 days, 44% (2 lesions, 3 sections counted,3–9 BrdU-labeled nuclei per section) were RET1-NN2–negativecompared with only 6% (4 lesions, 8 sections counted, 2 to 17BrdU-labeled nuclei per section) at 2 days. This observation is
consistent with the delay in mitotic activation that would beexpected of stem cells, which are rare and cycle slowly in theundamaged tissue.29,30 In contrast, mitotic activation of micro-glia and Muller glia in damaged adult goldfish retina is knownto be rapid.28
Movement of Proliferating Cells from the INL tothe ONL
We next examined changes in the spatiotemporal distributionof proliferating retinal cells during the first 2 weeks of regen-eration. During the first week we found a tendency for BrdU-labeled nuclei in the lesioned area to increase in the ONL anddecrease in the INL. Of the BrdU-labeled nuclei, 41% were inthe INL at 3 days but only 21% at 9 days (weighted average of19% and 27%; Table 1, column 5). Over the same interval, theproportion of BrdU-labeled nuclei in the ONL increased from59% to 78% (weighted average of 81% and 72%; Table 1,column 6). The biggest incremental change occurred between3 and 5 days (Table 1, columns 5 and 6). These changes in thedistribution of BrdU-labeled nuclei are consistent with a shift inthe distribution of proliferating cells from INL to ONL (al-though the apparent changes in percentages between 3 and 9days were not statistically significant). However, Muller nuclei(identified as those double-labeled with BrdU and RET1)showed a significant (P , 0.01) shift in distribution. At 5 days,71% of the BrdU-labeled nuclei in the ONL expressed RET1(weighted average of 75% and 61%), compared with only 16%at 3 days (Table 1, column 8). In contrast, the fraction ofBrdU-labeled nuclei in the INL that expressed RET1 between 3and 7 days was more constant (range: 51%–72%, P . 0.05;Table 1, column 7). These data are consistent with migration ofMuller nuclei from the INL to the ONL.
If the Muller cells proliferating in the region of the lesionproduced progeny that survived, a local increase would beexpected in the density of end feet in the regenerated region.To test this, we counted the glial end feet in the four lesionsseen in Figure 1A and compared these data to counts in anadjacent region of comparable area, in which end feet wereregularly spaced. Four separate counts were made of eachlesion and control area, and the ratios were averaged. Thenumber of end feet within the lesions ranged from 50 to 104.There were fewer end feet within the regenerated region in allfour lesions (18%, 26%, 31%, and 37%), suggesting that thelesion-induced proliferation of Muller cells does not result inpersistent gliosis—that is, the increase in number of Mullercells in the lesioned area was transient.
Figure 1H shows the retina from a fish that was injectedwith BrdU at 5 days and fixed at 7 days. Most of the nuclei thathad incorporated BrdU at 5 days were in the ONL (Table 1,column 6). They were not labeled with NN2, although occa-
TABLE 1. Quantification of BrdU- and RET1-Labeled Nuclei
1 2 3 4 5 6 7 8 9 10
BrdUInjections*
SurvivalTime*
Numberof
Sections
Total BrdUNuclei† per
SectionINL BrdU‡/Total BrdU*
ONL BrdU‡/Total BrdU*
INL Double§/INL BrdU‡
ONL Double§/ONL BrdU‡
INL Double§/Total BrdU*
ONL Double§/Total BrdU*
3 3 8 2–29 0.41 0.59 0.62 0.16 0.30 0.125 5 12 8–27 0.26 0.74 0.72 0.75 0.22 0.565 5 5 14–31 0.33 0.67 0.51 0.61 0.21 0.415 7 8 28–64 0.23 0.77 0.56 0.55 0.13 0.415 9 8 19–61 0.19 0.81 0.33 0.22 0.05 0.186, 7 9 4 25–52 0.27 0.73 0.29 0.27 0.08 0.17
* Days after laser treatment.† BrdU-labeled nuclei in the ONL and INL within the lesion (Fig. 1A).‡ BrdU-labeled nuclei in the INL (or ONL) within the lesion.§ Double-labeled nuclei (BrdU- and RET1-positive).
2120 Wu et al. IOVS, August 2001, Vol. 42, No. 9
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

sional BrdU-NN2–labeled microglia persisted in the INL in theregion of the lesion (Fig. 2B). Although RET1-BrdU double-labeled nuclei were still found in the INL (inset, Fig. 1H; Table1, column 7), the intensity of the RET1 labeling in most of theproliferating cells within the lesion at 7 days (Fig. 1H) was notas strong as it was at 5 days (Figs. 1F, 1G), consistent withdownregulation in levels of RET1 expression in the proliferat-ing Muller glia and/or increased representation of the RET1-negative proliferating progenitor cells after the first week. Only24% of the total BrdU-labeled nuclei that had been dividing at5 to 7 days expressed RET1 at 9 days (sum of the weightedaverages of 5% and 8% in the INL and 18% and 17% in the ONL),compared with 54% at 7 days (13% in the INL and 41% in theONL), and 73% at 5 days (sum of the weighted average of 22%and 21% in the INL and 56% and 41% in the ONL; Table 1,columns 9 and 10). Although these counts show a consistentdecrease in the overall proportion of double-labeled nucleiwith increased time after lesion, the differences were notstatistically significant. It is unlikely that the apparent loss ofRET1 expression in proliferating nuclei in the ONL is entirelyaccounted for by migration of Muller glial nuclei back into theINL, because the proportion of RET1-BrdU double-labeled nu-clei in the INL decreased, rather than increased, during thisperiod (Table 1, column 9).
When fish were injected with BrdU at 6 and 7 days andexamined at 12 days, we found faintly RET1-labeled nuclei inthe ONL that appeared to be young, regenerating cones, someof which were labeled with BrdU (data not shown). We con-firmed this inference by labeling these immature (regenerated)cone photoreceptors with a cone-specific marker, the zpr1monoclonal antibody (data not shown).
In summary, both the qualitative and quantitative data sug-gest several concurrent events. First, RET1-positive Muller cellsin the INL began to proliferate within a few days of the laserlesion. At 3 days, the majority of dividing cells in the INLexpressed RET1, and the majority of RET1-BrdU double-labelednuclei resided in the INL. The high percentage of double-labeled nuclei in the BrdU-labeled population within the INLup to 7 days suggests that Muller cells continued to divideduring the first week after lesion. Second, RET1-BrdU double-labeled Muller nuclei quickly migrated into the ONL within thefirst few days after the lesion. Radially elongated, ectopic nu-clei with varying levels of RET1 expression spanned the INL–ONL boundary within the first week, concomitant with a sud-den increase in RET1-BrdU double-labeled cells in the ONL(between 3 and 5 days), and a gradual decline in double-labeledcells in the INL (between 3 and 9 days). Third, although thedisplaced Muller cell nuclei disappeared from the ONL duringthe second week, they did not migrate back into the INL.Fourth, proliferating, RET1-NN2–negative, retinal progenitorsin the ONL and the INL increased sharply in number from 5 to10 days. Taken together, these data are consistent with injury-induced proliferation of RET1-positive Muller cells and (slightlydelayed) of RET1-negative neural progenitor cells and migra-tion of nuclei of both cell types from the INL to the ONL duringthe period of cone regeneration within the first week after thelaser lesion.
Expression of Cell Surface Markers Characteristicof Retinal Stem Cells
The foregoing results showed that cone photoreceptors regen-erated from proliferating retinal progenitors whose nuclei mi-grated from the INL into the “gap” in the ONL that wasproduced by the photocoagulation lesion. We next askedwhether these retinal progenitors express markers character-istic of the multipotent retinal progenitors known to generatecones—that is, the progenitors in the embryonic retina and in
the circumferential germinal zone of the adult retina. Twodevelopmentally regulated genes that code for cell surfacereceptors involved in cell adhesion and cell–cell signaling—N-cadherin (cdh2) and Notch-3—are strongly expressed by theembryonic and germinal zone retinal progenitors, as demon-strated by in situ hybridization with cRNA probes: zebrafishcdh2 (Fig. 2E) and goldfish G-Notch-336,37 (Fig. 2G). The gold-fish cdh2 gene has not been cloned, but the zebrafish cdh2probe showed specific hybridization in goldfish retina consis-tent with the pattern in zebrafish retina (Liu Q, Barthel LK,Raymond PA, et al., unpublished observations, 1999). In addi-tion to the strong level of expression of both these genes in thegerminal zone, we found that scattered cells in the INL, usuallyin or near the amacrine cell stratum on the inner side of theINL, also expressed both cdh2 (Fig. 2E) and Notch-336 (Fig.2G). In the ONL outside the region of damage, there was noexpression of cdh2 or Notch-3 in differentiated photoreceptorsor in rod precursors37 (Figs. 2E–H). However, within the le-sion, the expression of both genes was very strong in progen-itor cells that had migrated into the ONL (Figs. 2F, 2H).
DISCUSSION
The present results confirm and extend previous work fromthis laboratory demonstrating cone regeneration in the adultgoldfish retina.17 Mitotic divisions that occurred in neural pro-genitor cells within the first 2 weeks after laser ablationsproduced progeny that differentiated preferentially into cones,whereas regenerated rod photoreceptors appeared later, andthey were produced by specialized rod precursors.
During the first week after laser lesion, many different typesof proliferative cells infiltrated the ONL in response to theablation of photoreceptors. Some of these were vascular-de-rived cells (microglia) recruited to dispose of the cellular de-bris. These were identified with a specific cell surface markerNN2. Others were Muller cells, identified by cell-specific anti-body markers, including RET1, which recognizes an unknownnuclear epitope not specific to Muller glial markers and theglial-specific markers, GFAP and GS. It was surprising, how-ever, that proliferation of Muller cells did not generate extraglial cells in the region of the lesion, nor was there evidence ofMuller cell proliferation or turnover in the uninjured adultgoldfish retina. For example, a recent study41 found that thelevel of GS immunoreactivity in the retina remained constant inadult cichlid fish as the retina grew in size, whereas the densityof Muller cells themselves declined. These data suggest thatMuller cells in teleost fish respond to growth-related retinalexpansion by increasing their size, rather than by adding newcells interstitially. In the present study, we found that the levelof GS was reduced locally in the region of the lesion for up to25 days, and the density of Muller cells was lower in theregenerated retina than in the surrounding, intact retina, de-spite evidence of substantial proliferation of Muller cells in theregion of the lesion.
The fate of new cells produced by the proliferating Mullercells is unclear. Because they did not produce extra Mullercells, they must have either died or transformed into anothercell type. We cannot exclude the possibility that the progenyof the dividing Muller cells dedifferentiated or transdifferenti-ated into cone progenitors, but we have no direct evidence forthis. The gradual loss of RET1 immunoreactivity that we ob-served in the proliferating progenitors in the ONL, where theMuller nuclei migrated, could reflect their dedifferentiation.The Muller glial cell phenotype is unstable, at least in cellculture, where Muller cells can transdifferentiate into lentoidcells that express crystalline proteins.44 However, in the mam-malian retina, Muller cells also proliferate in response to vari-
IOVS, August 2001, Vol. 42, No. 9 Cone Regeneration and Retinal Stem Cells 2121
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

ous pathologic conditions, including photocoagulation le-sions,45–47 but they are not able to restore lost neurons andphotoreceptors. Instead, the Muller cell response is associatedwith a pathologic, reactive gliosis.38,48,49
Although there was no evidence of transformation of Mullercells into neuronal progenitors in the injured mammalian ret-ina, the possibility that this might occur in the fish retina isworthy of serious consideration, especially given surprisingnew findings about neural stem cells in the adult mammalianbrain. Two recent reports have suggested that neural stem cellsin the adult mammalian brain are actually a subclass of glialcell—either specialized astrocytes in the subventricular zone50
or ependymal cells at the ventricular surface.51 Both of theseglial cell types express the intermediate filament protein,GFAP.30,52 Muller cells are a specialized type of radial glia,53
and radial glial cells in the developing cerebral cortex in mam-mals can also behave as neural progenitors, even as they con-tinue to express glial-specific markers such as GFAP.54 In ad-dition, lineage-tracing studies in developing brain have shownclones of neurons and glia associated with a single radial glialcell,55,56 and in the retina, similar studies have shown thatMuller cells and retinal neurons, especially rod photoreceptors,are produced from a common retinal progenitor.57,58
The results reported herein show that presumptive retinalprogenitors in the INL responded to the loss of photoreceptorswith behavior similar to Muller cells: increased mitotic activityand migration of their nuclei to the ONL. Julian et al.8 havedemonstrated conclusively that slowly cycling retinal progen-itor cells persist in the INL of juvenile rainbow trout (On-choryncus mykiss). They used a mitotic labeling paradigmoptimized to detect slowly cycling cells, in which the thymi-dine analogues IdU (iododeoxyuridine) and BrdU were admin-istered continuously for up to 10 days. By exposing fish se-quentially to IdU and then BrdU, they showed that the labelednuclei moved from the INL to the ONL and that the purpose ofthis migration was to replenish the rapidly cycling rod precur-sor pool in the ONL, which eventually differentiated into rods.The radially elongated shape of these cells, and their frequentposition straddling the plexiform layer between the INL andthe ONL, are also consistent with outward radial migration. Wehad previously described the analogous behavior of retinalprogenitor cells in the INL of the larval goldfish retina, and wealso showed that these cells that originated in the INL were thesource of the rod precursors, which did not appear in the ONLuntil early larval stages.20
Both our earlier study20 and the more recent work8 havedemonstrated that the only progeny of the retinal progenitorsin the INL of the uninjured retina are the rod precursors, andthe only progeny of rod precursors are rod photoreceptors.The results of the present study demonstrate that in the le-sioned adult goldfish retina, the rate of proliferation was locallyupregulated in the INL progenitors at the time when coneswere being generated and that their nuclei migrated into theONL, in association with the Muller cell nuclei. These resultsare consistent with the proposal that the INL progenitors arenot, in fact, restricted to the rod lineage, but can also generatecone photoreceptors. Previous studies have reported regener-ation of the complete array of retinal neurons in adult goldfishfrom clusters of proliferating cells with behavior and morphol-ogy similar to that described here.31,32,59 We therefore con-clude that the neural progenitor cells in the INL are likely to bemultipotent retinal stem cells, with a capacity similar to that ofthe primitive, multipotent retinal progenitor cells in the ger-minal zone at the retinal margin.
The regeneration of cones was associated with migration ofthe nuclei of retinal progenitors to establish contact with theouter limiting membrane (OLM), which represents the apical(ventricular) surface of the retinal epithelium. We have shown
previously that in other experimental paradigms, regenerationof neurons in the adult fish retina takes place only whenproliferating retinal progenitors reach the OLM.17,22,34,60 Thiscellular arrangement mimics the cytological organization of theprimitive retinal neuroepithelium and the circumferential ger-minal zone at the retinal margin, and it may reflect the pres-ence of obligatory environmental factors that perhaps regulatethe multipotent capacity of the retinal progenitors—that is,their ability to produce multiple types of retinal neurons. Anassociation with the apical surface of the neuroepithelium maybe a fundamental feature of stem cells. Neural stem cells in theembryonic and adult mammalian brain are also associated withthe ventricular surface.61,62
Although there are no known markers specific for retinalstem cells, the proliferating retinal progenitors within the le-sioned retina expressed high levels of two developmentallyregulated genes that have been implicated in retinal neurogen-esis: N-cadherin (cdh2), a calcium-dependent, homophilic ad-hesion molecule that is expressed by multipotent neural pro-genitor cells, including those in the retina,63,64 and G-Notch-3,a member of the Notch family of cell surface receptors thatregulate contact-mediated, lateral inhibitory interactions andcell fate in developing tissues,65,66 including the retina.37,67
Rod precursors express neither of these genes, and their up-regulation in association with regeneration of cone photore-ceptors suggests that the expression of Notch and N-cadherinmay be associated with multipotent retinal stem cells. Neuralstem cells derived from the ependymal layer of the brain alsoexpress Notch, and antibodies to this cell surface marker havebeen used to enrich neural stem cells from the adult mamma-lian brain.30,51 However, under some circumstances, activationof the Notch signaling pathway promotes gliogenesis,68 includ-ing differentiation of Muller glia.69
If the INL progenitors are multipotent retinal stem cells,they would be expected to express other developmentallyregulated genes that are characteristic of the primitive retinalprogenitors in the embryonic retina and in the germinal zoneat the retinal margin.70,71 This includes transcriptional regula-tors in the homeodomain family (e.g., Rx, Pax6, Chx10/vsx1,Six3) and others in the basic helix-loop-helix family (atonaland achaete scute homologues), all of which have been impli-cated in retinal development.67,72 The multipotent retinal pro-genitors in the regeneration blastema at the edge of a surgicallesion in adult goldfish retina express both vs.x1 and pax6,73,74
and recently, Otteson et al.18,19 showed that the slowly prolif-erating, INL progenitors in the undamaged adult goldfish retinacould be immunostained with antibodies directed against re-combinant zebrafish pax6.1 protein. Efforts are under way todetermine what other genes are expressed by the retinal stemcells in the adult fish retina.
Acknowledgments
The authors thank the Kellogg Eye Center of the Department ofOphthalmology and Visual Sciences at the University of MichiganMedical School for the use of the argon laser and Kathleen Welch,University of Michigan Center for Statistical Consultation and Research,for help with statistical analysis.
References
1. Johns PR. Growth of the adult goldfish eye. III: source of the newretinal cells. J Comp Neurol. 1977;176:343–358.
2. Easter SS. Postnatal neurogenesis and changing connections.Trends Neurosci. 1983;6:53–56.
3. Sandy JM, Blaxter JHS. A study of retinal development in larvaeherring and sole. J Mar Biol Assoc (UK). 1980;60:59–71.
2122 Wu et al. IOVS, August 2001, Vol. 42, No. 9
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

4. Johns PR. The formation of photoreceptors in larval and adultgoldfish. J Neurosci. 1982;2:179–198.
5. Munk O, Jørgensen JM. Mitoses in the retina of two deep-seateleosts. Vidensk Meddr Dansk Naturh Foren. 1983;144:75–81.
6. Mansour-Robaey S, Pinganuad G. Quantitative and morphologicalstudy of cell proliferation during morphogenesis in the trout visualsystem. J Hirnforsch. 1990;31:495–504.
7. Hagedorn M, Fernald R. Retinal growth and cell addition duringembryogenesis in the teleost, Haplochromis burtoni. J CompNeurol. 1992;321:193–208.
8. Julian D, Ennis K, Korenbrot JI. Birth and fate of proliferative cellsin the inner nuclear layer of the mature fish retina. J Comp Neurol.1998;394:271–282.
9. Negishi K, Stell WK, Takasaki Y. Early histogenesis of the tel-eostean retina: studies using a novel immunochemical marker,proliferating cell nuclear antigen (PCNA/cyclin). Dev Brain Res.1990;55:121–125.
10. Muller H. Bau und Wachstum der Netzhaut des Guppy (Lebistesreticulatus). Zool Jb. 1952;63:275–324.
11. Raymond PA. Movement of retinal terminals in goldfish optictectum predicted by analysis of neuronal proliferation. J Neurosci.1986;6:2479–2488.
12. Johns PR, Fernald RD. Genesis of rods in teleost fish retina. Nature.1981;293:141–142.
13. Raymond PA. The unique origin of rod photoreceptors in theteleost retina. Trends Neurosci. 1985;8:12–17.
14. Fernald RD. Retinal rod neurogenesis. In: Finlay BL, Sengelaub DR,eds. Development of the Vertebrate Retina. New York: Plenum;1989:31–42.
15. Johns PR, Easter SS. Growth of the adult goldfish eye. II: increasein retinal cell number. J Comp Neurol. 1977;176:331–341.
16. Mack AF, Fernald RD. Cell movement and cell cycle dynamics inthe retina of the adult teleost, Haplochromis burtoni. J CompNeurol. 1997;388:435–443.
17. Braisted JE, Essman TF, Raymond PA. Selective regeneration ofphotoreceptors in goldfish retina. Development. 1994;120:2409–2419.
18. Otteson DC, Hitchcock PF. Neurogenesis in the retina of thegoldfish: long-term systemic exposure to BrdU reveals slowly di-viding cells in the inner nuclear layer [ARVO Abstract]. InvestOphthalmol Vis Sci. 2000;41(4):S848. Abstract nr 4505.
19. Otteson DC, D’Costa AR, Hitchcock PF. Putative stem cells and thelineage of rod photoreceptors in the mature retina of the goldfish.Devel Biol. 2001;232:62–76.
20. Raymond PA, Rivlin PK. Germinal cells in the goldfish retina thatproduce rod photoreceptors. Dev Biol. 1987;122:120–138.
21. Maier W, Wolburg H. Regeneration of the goldfish retina afterexposure to different doses of ouabain. Cell Tissue Res. 1979;202:99–118.
22. Raymond PA, Reifler MJ, Rivlin PK. Regeneration of goldfish retina:rod precursors are a likely source of regenerated cells. J Neurobiol.1988;19:431–463.
23. Negishi K, Stell WK, Teranishi T, Karkhanis A, Owusu-Yaw V,Takasaki Y. Induction of proliferating cell nuclear antigen (PCNA)-immunoreactive cells in goldfish retina following intravitreal injec-tion with 6-hydroxydopamine. Cell Mol Neurobiol. 1991;11:639–659.
24. Negishi K, Sugawara K, Shinagawa S, Teranishi T, Kuo CH,Takasaki Y. Induction of immunoreactive proliferating cell nuclearantigen (PCNA) in goldfish retina following intravitreal injectionwith tunicamycin. Dev Brain Res. 1991;63:71–83.
25. Hitchcock PF, Lindsey Myhr KJ, Easter SS, Mangione-Smith R, JonesDD. Local regeneration in the retina of the goldfish. J Neurobiol.1992;23:187–203.
26. Negishi K, Shinagawa S. Fibroblast growth factor induces prolifer-ating cell nuclear antigen-immunoreactive cells in goldfish retina.Neurosci Res. 1993;18:143–156.
27. Negishi K. 5-fluorouracil reduces proliferating cell nuclear antigenimmunoreactive cells in goldfish retina. Neurosci Res. 1994;19:21–29.
28. Wagner EC, Raymond PA. Muller glial cells of the goldfish retinaare phagocytic in vitro but not in vivo. Exp Eye Res. 1991;53:583–589.
29. Morrison SJ, Shah NM, Anderson DJ. Regulatory mechanisms instem cell biology. Cell. 1997;88:287–298.
30. Momma S, Johansson CB, Frisin J. Get to know your stem cells.Curr Opin Neurobiol. 2000;10:45–49.
31. Raymond PA, Hitchcock PF. Retinal regeneration: common prin-ciples but a diversity of mechanisms. Adv Neurol. 1997;72:171–184.
32. Raymond P, Hitchcock P. How the neural retina regenerates. In:Fini M, ed. Vertebrate Eye Development. Heidelberg: Springer;2000:197–218.
33. Barthel LK, Raymond PA. Improved method for obtaining 3-microncryosections for immunocytochemistry. J Histochem Cytochem.1990;38:1383–1388.
34. Braisted JE, Raymond PA. Regeneration of dopaminergic neuronsin goldfish retina. Development. 1992;114:913–919.
35. Barthel LK, Raymond PA. Subcellular localization of alpha-tubulinand opsin mRNA in the goldfish retina using digoxigenin-labeledcRNA probes detected by alkaline phosphatase and HRP histo-chemistry. J Neurosci Methods. 1993;50:145–152.
36. Barthel LK, Raymond PA. In situ hybridization studies of retinalneurons. Methods Enzymol. 2000;316:579–590.
37. Sullivan SA, Barthel LK, Largent BL, Raymond PA. A goldfishNotch-3 homologue is expressed in neurogenic regions of embry-onic, adult, and regenerating brain and retina [published correc-tion appears in Dev Genet. 1997;21:175–176] Dev Genet. 1997;20:208–223.
38. Ishigooka H, Hirata A, Kitaoka T, Ueno S. Cytochemical studies onpathological Muller cells after argon laser photocoagulation. InvestOphthalmol Vis Sci. 1989;30:509–520.
39. Humphrey MF, Chu Y, Mann K, Rakoczy P. Retinal GFAP and bFGFexpression after multiple argon laser photocoagulation injuriesassessed by both immunoreactivity and mRNA levels. Exp Eye Res.1997;64:361–369.
40. Raymond PA. Cytodifferentiation of photoreceptors in larvalgoldfish: delayed maturation of rods. J Comp Neurol. 1985;236:90–105.
41. Mack AF, Germer A, Janke C, Reichenbach A. Muller (glial) cells inthe teleost retina: consequences of continuous growth. Glia. 1998;22:306–313.
42. Linser P, Moscona AA. Induction of glutamine synthetase in em-bryonic neural retina: localization in Muller fibers and dependenceon cell interactions. Proc Natl Acad Sci USA. 1979;76:6476–6480.
43. Lewis G, Erickson P, Guirin J, Anderson D, Fisher S. Changes in theexpression of specific Muller cell proteins during long-term retinaldetachment. Exp Eye Res. 1989;49:93–111.
44. Degenstein L, Moscona AA. Retinoic acid inhibits conversion ofdissociated Muller glia into lens-like cells. Exp Eye Res. 1986;43:93–102.
45. Fisher SK, Erickson PA, Lewis GP, Anderson DH. Intraretinal pro-liferation induced by retinal detachment. Invest Ophthalmol VisSci. 1991;32:1739–1748.
46. Puro DG. Growth factors and Muller cells. Prog Retinal Eye Res.1995;15:89–101.
47. Dyer MA, Cepko CL. Control of Muller glial cell proliferation andactivation following retinal injury. Nat Neurosci. 2000;3:873–880.
48. Sarthy V. Reactive gliosis in retinal degenerations. In: Anderson RE,Hollyfield JG, LaVail MM, eds. Retinal Degenerations. Boca Raton,FL: CRC Press; 1991:109–116.
49. Xiao M, Sastry SM, Li ZY, et al. Effects of retinal laser photocoag-ulation on photoreceptor basic fibroblast growth factor and sur-vival. Invest Ophthalmol Vis Sci. 1998;39:618–630.
50. Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla A.Subventricular zone astrocytes are neural stem cells in the adultmammalian brain. Cell. 1999;97:703–716.
51. Johansson CB, Momma S, Clarke DL, Risling M, Lendahl U, FrisenJ. Identification of a neural stem cell in the adult mammaliancentral nervous system. Cell. 1999;96:25–34.
52. Barres BA. A new role for glia: generation of neurons! Cell. 1999;97:669–670.
IOVS, August 2001, Vol. 42, No. 9 Cone Regeneration and Retinal Stem Cells 2123
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018

53. Reichenbach A, Reichelt W. Postnatal development of radial glial(Muller) cells of the rabbit retina. Neurosci Lett. 1986;71:125–130.
54. Misson JP, Edwards MA, Yamamoto M, Caviness VS Jr. Mitoticcycling of radial glial cells of the fetal murine cerebral wall: acombined autoradiographic and immunohistochemical study.Brain Res. 1988;466:183–190.
55. Halliday AL, Cepko CL. Generation and migration of cells in thedeveloping striatum. Neuron. 1992;9:15–26.
56. Gray GE, Sanes JR. Lineage of radial glia in the chicken optictectum. Development. 1992;114:271–283.
57. Turner DL, Cepko CL. A common progenitor for neurons and gliapersists in rat retina late in development. Nature. 1987;328:131–136.
58. Reh TA, Levine EM. Multipotential stem cells and progenitors inthe vertebrate retina. J Neurobiol. 1998;36:206–220.
59. Hitchcock PF, Raymond PA. Retinal regeneration. Trends Neuro-sci. 1992;15:103–108.
60. Braisted JE, Raymond PA. Continued search for the cellular signalsthat regulate regeneration of dopaminergic neurons in goldfishretina. Dev Brain Res. 1993;76:221–232.
61. Morshead CM, Reynolds BA, Craig CG, et al. Neural stem cells inthe adult mammalian forebrain: a relatively quiescent subpopula-tion of subependymal cells. Neuron. 1994;13:1071–1082.
62. Shen Q, Qian X, Capela A, Temple S. Stem cells in the embryoniccerebral cortex: their role in histogenesis and patterning. J Neu-robiol. 1998;36:162–174.
63. Matsunaga M, Hatta K, Takeichi M. Role of N-cadherin cell adhe-sion molecules in the histogenesis of neural retina. Neuron. 1988;1:289–295.
64. Rutishauser U. N-Cadherin: a cell adhesion molecule in neuraldevelopment. Trends Neurosci. 1989;12:275–276.
65. Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fatecontrol and signal integration in development. Science. 1999;284:770–776.
66. Weinmaster G. Notch signal transduction: a real rip and more.Curr Opin Genet Dev. 2000;10:363–369.
67. Perron M, Harris WA. Determination of vertebrate retinal pro-genitor cell fate by the Notch pathway and basic helix-loop-helix transcription factors. Cell Mol Life Sci. 2000;57:215–223.
68. Wang S, Barres BA. Up a notch: instructing gliogenesis. Neuron.2000;27:197–200.
69. Furukawa T, Mukherjee S, Bao ZZ, Morrow EM, Cepko CL.rax, Hes1, and notch1 promote the formation of Mullerglia by postnatal retinal progenitor cells. Neuron. 2000;26:383–394.
70. Perron M, Kanekar S, Vetter ML, Harris WA. The genetic sequenceof retinal development in the ciliary margin of the Xenopus eye.Dev Biol. 1998;199:185–200.
71. Perron M, Harris W. Molecular recapitulation: the growth of thevertebrate retina. Int J Dev Biol. 1998;42:299–304.
72. Cepko CL. The roles of intrinsic and extrinsic cues and bHLHgenes in the determination of retinal cell fates. Curr Opin Neuro-biol. 1999;9:37–46.
73. Levine EM, Hitchcock PF, Glasgow E, Schechter N. Restrictedexpression of a new paired-class homeobox gene in normal andregenerating adult goldfish retina. J Comp Neurol. 1994;348:596–606.
74. Hitchcock PF, Macdonald RE, VanDeRyt JT, Wilson SW. Anti-bodies against pax6 immunostain amacrine and ganglion cellsand neuronal progenitors, but not rod precursors, in the normaland regenerating retina of the goldfish. J Neurobiol. 1996;29:399 – 413.
2124 Wu et al. IOVS, August 2001, Vol. 42, No. 9
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932896/ on 05/12/2018