cooperation in microbial biotechnology 2 - … · · 2017-04-12cooperation in microbial...
TRANSCRIPT

Cooperationinmicrobialbiotechnology
1
1
2
Cooperationinmicrobialcommunitiesandtheirbiotechnologicalapplications3
4
MatteoCavaliere1,SongFeng2,OrkunSoyer3andJoseI.Jimenez4*5
6
1SchoolofInformatics,BBSRC/EPSRC/MRCSyntheticBiologyResearchCentre,Universityof7Edinburgh8
2CenterforNonlinearStudies,TheoreticalDivision(T-6),LosAlamosNationalLaboratory9
3SchoolofLifeSciences,BBSRC/EPSRCWarwickIntegrativeSyntheticBiologyCentre,Universityof10Warwick11
4FacultyofHealthandMedicalSciences,UniversityofSurrey12
13
*Towhomcorrespondenceshouldbeaddressed:14
FacultyofHealthandMedicalSciences15
UniversityofSurrey16
Guildford,GU27XH17
UnitedKingdom18
Email:[email protected]
Phone:+440148368455720
21
Statementofsignificance:22
InthisreviewwesummariseadvancesinthefieldofEvolutionaryDynamicsappliedtomicrobial23
communitiesandtheirapplicationsinbiotechnology.Wediscussdifferentkindsofcooperative24
interactions,theirpotentialmechanisticoriginsandthefactorsthatcontributetotheirstability.We25
alsoanalysetheadvantagesofcooperativebehavioursinmicrobialpopulationsandevaluatetheir26
possibleusetodeveloprobustbiotechnologicalapplications.27
28

Cooperationinmicrobialbiotechnology
2
Abstract1
Microbialcommunitiesareincreasinglyutilisedinbiotechnology.Efficiencyandproductivityinmany2
oftheseapplicationsdependsonthepresenceofcooperativeinteractionsbetweenmembersofthe3
community.Twokeyprocessesunderlyingtheseinteractionsaretheproductionofpublicgoodsand4
metaboliccrossfeeding,whichcanbeunderstoodinthegeneralframeworkofecologicaland5
evolutionary(eco-evo)dynamics.Inthisreviewweillustratetherelevanceofcooperative6
interactionsinmicrobialbiotechnologicalprocesses,discusstheirmechanisticorigins,andanalyse7
theirevolutionaryresilience.Cooperativebehaviourscanbedamagedbytheemergenceof8
‘cheating’cellsthatbenefitfromthecooperativeinteractionsbutdonotcontributetothem.Despite9
this,cooperativeinteractionscanbestabilizedbyspatialsegregation,bythepresenceoffeedbacks10
betweentheevolutionarydynamicsandtheecologyofthecommunity,bytheroleofregulatory11
systemscoupledtotheenvironmentalconditionsandbytheactionofhorizontalgenetransfer.12
Cooperativeinteractionsenrichmicrobialcommunitieswithahigherdegreeofrobustnessagainst13
environmentalstressandcanfacilitatetheevolutionofmorecomplextraits.Therefore,the14
evolutionaryresilienceofmicrobialcommunitiesandtheirabilitytoconstraintdetrimentalmutants15
shouldbeconsideredinordertodesignrobustbiotechnologicalapplications.16
17

Cooperationinmicrobialbiotechnology
3
Evolutionarydynamicsandcooperationinmicrobialpopulations1
Thedesignandoptimizationofmicroorganismsforbiotechnologicalpurposesoftenconsiderscells2
inisolation.Whilethisreductionistapproachaimstothriveforsimplicityintheprocess,itcreatesa3
situationthatrarelytakesplaceinNature.Intheirnaturalenvironmentmicroorganismsthrivein4
complexcommunitiesinwhichthefitnessofasinglecelldependsontheinteractionswithother5
cellsinthepopulation(Westetal.,2006).Thisscenarioalsoappliestobioprocessesinwhichthe6
efficiencyoftheprocessiscoupledtotheproductionofshared(public)goodsthatallowcellsto7
performtasksina‘cooperative’manner(Lindemannetal.,2016):agoodexampleofsharedgoods8
arethecellulasessecretedintheproductionofcellulosicethanol(ZomorrodiandSegrè,2016).9
Thepresenceofcooperativeinteractionshasasignificantimpactontheevolutionary10
dynamicsofmicrobialcommunities,representedbythechangeinthefrequenciesofcellsand11
speciesthatimplementdifferentphysiologicalstrategies(suchasproductionofpublicgoodsvs.12
not).Thus,cooperativetraitsneedtobetakenintoaccountwhenusinganevolutionaryapproach13
foroptimisingagivenbioprocess.Itispossiblethatsimpleselectionschemestargetingabioprocess-14
relatedtrait(e.g.growthrate)willnotalignwiththeselectionforthecooperativetrait(e.g.15
productionofcostlyextracellularenzymes)ultimatelyresultinginthelossofthetrait.Indeed,16
tradeoffsbetweentheoptimizationofso-calledhigh-rateandhigh-yieldarefrequentlyobservedin17
controlledevolutionaryexperiments(Bachmannetal.,2013).Thus,weadvocateconsideringthe18
interactionsbetweenthecellsandthefunctioningofcooperativetraitswhendesigningevolutionary19
optimisationandstabilisationofbioprocesses.Achievingthiswouldrequireconsideringhow‘social’20
interactionsshapemicrobialprocesses,ratherthansimplyfocusingsolelyonindividualistictraits21
suchasgrowthrate.22
Thissituationmayconfronttheintuitiveideathat‘evolutionimpliesimprovement’(i.e.the23
averagefitnessofthecommunityisexpectedtoincreaseovergenerationsasitwouldbeexpected24
formonocultures).Thekeypointisthatthepresenceofinteractionsbetweenthespeciesgivesrise25
toamorecomplicatedevolutionarypictureinwhichthefitnessofacelldependsnotonlyonits26
phenotypebutalsoontheoverallcompositionofthepopulation.Thespreadingofagiven27
phenotypictraitmaythuschangethefitnessofothermembersofthecommunityandthesechanges28
mayinturnfeedbackonthefitnessoftheindividualcells(Westetal.,2006).Theseintertwined29
selectionmechanismsareexpectedtooperateinanymicrobialpopulationwherethereispossibility30
ofdifferentcellsimplementingdifferentstrategieswithrespecttotheirphysiology,asisthecaseof31
phenotypicheterogeneity.32
Phenotypicheterogeneityariseseveninmonoculturesandsimplebioprocessesdueto33
differentreasons,suchastheuseofnon-homogenouscultureconditions,stochasticityingene34

Cooperationinmicrobialbiotechnology
4
expressionanddifferentialepigeneticcontrol(Enforsetal.,2001;Avery,2006;Mülleretal.,2010).1
Suchheterogeneitydoesnotrepresentastaticpicture–cellscommunicate,competeandcooperate2
andthesuccessofatraitmaybeconsequenceoftheinteractionwiththeothertraitsandofthe3
specificecologicalcontext(Carlquistetal.,2012).Therefore,itisnotsufficientforatraittobe4
successfulinonespecificsettingbutrather,itneedstobesuccessfulgiventhepresenceofother5
traitsandtheassociatedecologicalcontext.Moreover,thedilutionofatraitmayleadtochangesin6
thecommunity(bothecologicaland/orinthefrequencyofothertraits)thatcouldfeedbackonthe7
evolutionarydynamicsofthetraititself.Forinstance,atraitmaybefavouredbynaturalselection8
onlywhenrareinacomplexpopulation,becomingdisfavouredwhenitismorefrequent.These9
complexevolutionaryandecologicaldynamics,whereessentiallythesuccessofatraitdependson10
thecompositionofthecommunity,canbemathematicallyanalysedwithevolutionarygametheory11
(NowakandSigmund,2004;Frey,2010).12
Evolutionarygametheoryisamathematicalframeworkthatcomesfromclassicalgame13
theoryusedtodescribethebehaviourofrationalplayers.Classicalgametheorytriestoanalysethe14
behaviourinconflictsineconomicandsocialsettingsinwhichthesuccessofanindividualstrategy15
dependsonthestrategiesemployedbytheotherplayers.Awell-studiedexampleingametheoryis16
theprisoner’sdilemmainwhichthechoicestoeitherconfessorremainsilentdeterminewhether17
twosuspectsareconsideredguilty(Axelrod,1990).Inevolutionarygametheory,thestrategiesare18
notassociatedtorationalandcognitivechoices,butaretraitsencodedintoinheritedprogramsthat19
canbepassedtotheoffspring(forthisreason,thetermstraitandstrategiesareusedinan20
indistinguishablemanner).Traitssuchastheusageofmetabolicpathwaysortheexpressionof21
certainenzymescanbethenregardedasstrategiesandasuccessfulstrategyisthenselectedfor.22
Inamicrobialcommunitycomposedofspeciesthatcompeteusingdifferentstrategies,each23
oftheindividualcellspossessesafitnessthatdependsonitsstrategyandonthestrategyofthe24
individualswithwhomitinteracts.Individualsthatusemoresuccessfulstrategieshavehigher25
chancestopropagateandtheirfrequencyinthecommunitywillincrease.Althoughthedynamicsof26
anevolutionarygametheorymodelcanbestudiedanalyticallywhenthesetofstrategiesissmall,27
duetothelargenumberofinteractionstakingplaceinmicrobialcommunitiesmanyauthorsprefer28
tosimulatethedynamicsofthecommunityusingagent-basedmodelling.Inthesemodels,the29
replicationanddeathofindividualcells(agents)areexplicitlysimulatedusingasystemupdatedbya30
seriesofdiscreteevents(Adamietal.,2016).Thesetypesofmodelsalsoincludethepossibilityof31
addingmutationsthatcanintroducenovelstrategiesnotyetpresentinthespecies,whichcanbe32
usedtosimulaterandomevolutionofmembersofthecommunity(ErikssonandLindgren,2005).33

Cooperationinmicrobialbiotechnology
5
Incellularpopulations,acooperativetraitisoftencharacterisedbythepresenceofashared1
publicgood,whichisafiniteresource,producedbycooperativecellsandthatisfreelyavailableto2
allothercells.Thepresenceofapublicgoodisalwaysassociatedwiththeriskofcheatingcells,3
whichexploitthepublicgoodwithoutprovidinganycontributiontoitandwhichcanspreadinthe4
population–duetotheirimprovedfitnessarisingfromnotinvestingthecostsassociatedwithpublic5
goodproduction.Althoughinthisreviewwefocusonmicrobialpopulations,thisisaverygeneral6
issueinthesustainabilityofmanyorganismsatdifferentscalesincludinghumans,justifyingwhythe7
evolution(andresilience)ofcooperationisconsideredoneofthemajoropenquestionsinbiology8
(Pennisi,2009).9
Evolutionaryconflictsbetweencooperativeandcheatingcellshavebeenstudiedinavariety10
ofmicrobialscenarios,includingtheconversionofsucroseintoglucosebytheyeastSaccharomyces11
cerevisiae(Goreetal.,2009),theproductionoftheshareableiron-scavengingsiderophore12
pyoverdineinPseudomonasaeruginosa(Kümmerlietal.,2009)andtheformationoffruitingbodies13
inMyxobacteria(VelicerandVos,2009).Giventhepotentialsimilaritieswithcelluloseandother14
polymersbiodegradation,theexamplefromyeastisworthexplainingfurther.Inthiscase,15
cooperativeandcheatingcellsonlydifferbytheproductionoftheenzymeinvertasethatconverts16
sucroseintoglucoseandfructose.Bothmonosaccharidescaneventuallydiffuseawayfromthe17
producingcellandbecomeavailabletoneighbouringcells.Inotherwords,theybecomepublic18
goods:cooperators—thecellsthat‘feed’themselvesandtheirneighboursattheexpenseof19
expressingtheenzyme—canbeexploitedbycheaters,cellsthatdonotexpresstheenzymeand20
relyoncooperatorstomakefood(Fig.1A).Inascenariolikethis,itwouldbeexpectedthatcheaters21
couldtakeoverthepopulation.However,thefitnessofthecellsisanon-linearfunctionofthe22
glucoseconcentrationand,forcertainvaluesofglucoseuptakeandmetaboliccostofenzyme23
production,itispossibletoobservetheco-existenceofthetwospeciesasanticipatedbyan24
evolutionarygametheorymodel(Goreetal.,2009).Infact,inacomplexcommunitycomposedof25
multitudeofspeciesitislikelythatsuchmechanisticpropertiesrelatingtotheimplementationof26
thedifferentstrategies,suchasregulatorymechanismscontrollingtheproductionofapublicgood,27
willaffecttheevolutionaryandecologicaldynamicsofthestrategiesandthusthewholecommunity.28
Beforediscussingfurtherthesepotentialmechanismsthatcanstabilisecooperativeinteractions,we29
willfirstdescribetypesofcooperativeinteractionsinmicrobialpopulations.30
31
Microbialcooperationsbasedonpublicgoods32
Shared(public)goodsaremoleculesproducedbycertainindividualsandcanbenefittheentire33
population(Westetal.,2007).Asexplainedabove,thesemoleculesaresynthesisedatacostand,34

Cooperationinmicrobialbiotechnology
6
therefore,aresusceptibletobeexploitedbycheatercellsthatcanbenefitfromthembutdonot1
contributetotheirproduction–henceacquiringafitnessadvantageovercooperators.Thistypeof2
cooperationisbasedonalargevarietyofsharedmolecules:siderophores,enzymes,biosurfactants,3
componentsofbiofilmmatrix,quorumsensingmolecules,bacteriocins(proteinssecretedbyone4
straintoinhibitthegrowthofacloselyrelatedstrain)andtoxinsassummarizedin(Westetal.,5
2007).Giventheirinterestinmicrobialbiotechnology,inthisreviewwewillfocusonsecretionof6
degradatoryenzymes.7
Microorganismsdigestlargemacromolecules,whicharepoorlysoluble,throughthe8
secretionofextracellularenzymes.Themacromoleculesaretypicallypolymersofbiologicalor9
syntheticorigin,suchasstarch,celluloseandpolyesters,whichconstituteanabundantsourceof10
nutrientsforbacteria,fungiandothereukaryoticmicroorganisms(Allison,2005;Richardsand11
Talbot,2013).Thesepolymersalsoconstituteaveryinterestingsubstrateforindustrialbioprocesses,12
astheyareinexpensive,biodegradableatsomeextentandoftenobtainedfromrenewablesources13
(GrossandKalra,2002).Theenzymessecretedbymicroorganismsactbydegradingthe14
macromoleculesintosimplerandsmallercomponentsthatcanthenbeassimilatedbythemicrobial15
community(Burns,2010).Inthisscenario,thedynamicsofthecooperatingandcheating16
populationsdependonparameterssuchasthecostofproducingtheenzymesandtheirdiffusibility17
(Allison,2005).18
Cellulasesandoxidativeenzymessecretedtocleavecellulosesuchascellobiase19
dehydrogenasescanbeconsideredasinstancesof‘publicgoods’(Dimarogonaetal.,2012)andare20
foundinthegenomeofmostwood-degradingmicrobialcommunities(Zamockyetal.,2006).Similar21
tocellulases,amylasescapableofdegradingtheglycosidiclinkagesofstarchesalsoplayan22
importantroleaspublicgoodsandhavebeenidentifiedinmanybacteriaandfungi,suchasBacillus23
subtilis(ColemanandElliott,1962),Thermomyceslanuginosus(Arnesenetal.,1998),Penicillium24
expansum(Doyleetal.,1998),andseveralspeciesofStreptomyces(El-Fallaletal.,2012).Similarly,25
enzymesresponsibleforthedigestionofothermacromoleculessuchasextracellularlipasesand26
proteasesarealsoexamplesofpublicgoods,andtheirproductioninacomplexmicrobialcommunity27
isinfluencedbytheinteractionsbetweenitsmembers(WillseyandWargo,2015).Collectively28
producedenzymesarealsoresponsibleforthedegradationofoil-derivedplasticpolymerssuchas29
poly-ethylenterephthalate(PET).Theidentificationofbacterialspeciesproducingenzymescapable30
ofPETdepolymerisation,thereforegeneratingmoleculesthatcanthenbeassimilatedbythe31
microbialcommunityinthatniche(Chenetal.,2010;Yoshidaetal.,2016)pavesthewayforthe32
remediationofPETwasteanditsuseasabioprocessingsubstrate(Wierckxetal.,2015).33
34

Cooperationinmicrobialbiotechnology
7
Microbialcooperationsbasedonmetabolicinteractions1
Metabolicexchangeisanotherwayinwhichmicroorganismscaninteractcooperatively.Metabolic2
interactionsarewidespreadinnaturalmicrobialcommunitiesandarisefrommetabolitesfromone3
speciesbeingusedasenergysourcesorbuildingblocksbyotherspecies(Pacziaetal.,2012;Cooper4
andSmith,2015;Fioreetal.,2015).Theformerscenarioleadstocross-feeding,whereasthelatter5
canleadtoemergenceofauxotrophies(anorganismfullyrelyingontheenvironmentalprovisionof6
certaincompoundsrequiredforitsgrowth)(Fig1B).Themetabolitesreleasedintotheenvironment7
canbeexplainedbyeitherpassiveoractivemeans,i.e.organismsnotbeingabletomaintaincertain8
compoundsduetoleakageissuesoractivelysecretingthosecompoundsduetosomefunctional9
benefits.Whiletheformerexplanationcouldariseduetosomefundamentalbiophysicallimitations10
onbiologicalmembranes,thesecond(functional)explanationisdifficulttorationwithinasimplistic11
viewoforganismalfitness.Onecouldnaivelyarguethatsinceotherorganismsusethesecreted12
metabolitesasaresource,evolutionshouldhaveallowedthe‘secretingorganism’alsotoinnovate13
thatcapacityofusingthismetabolite(asanenergysourceorbuildingblock)ratherthansecretingit.14
Thisnaïveview,however,ignoreslimitationsarisingfromcellulartradeoffsandthermodynamics.15
16
Metabolicinteractionsemergingfromthermodynamiclimitations17
Inprinciple,cross-feedingandauxotrophicinteractionscouldbeseenasanextremeformof18
cooperation(i.e.,‘altruism’)astheybenefitonlythereceivingorganisms.Undercertainconditions,19
however,secretionofinternalmetabolitescanalsobenefittheproducerleadingtoamutually-20
beneficialinteraction:iftheproductsreleasedhaveaninhibitoryeffectontheproducer,the21
presenceofanadditionalspeciesthatwouldassimilatetheseproductswouldleadtomoremild22
formsofcooperativeinteractionratherthanastraight‘altruistic’actonbehalfoftheproducer(Lilja23
andJohnson,2016).Morespecifically,thistypecross-feedinginteraction,involvingreleaseof24
inhibitionarisingfrombyproductsofmetabolismofoneorganismbyanotherisoftenreferredtoas25
syntrophy(Fig2A).Themost-wellknownexampleistheH2-mediatedsyntrophicinteractions26
betweensecondarydegradersandmethanogens(Schink,1997).Intheseinteractions,theinhibition27
ofthedegradingspeciesarisesduetoitsgrowth-supportingmetabolicreactionreachingtowards28
thermodynamicequilibriumasH2accumulates(Schink,1997;GroßkopfandSoyer,2016).This29
‘thermodynamicinhibition’isrelievedbytheconsumingofH2bythesyntrophicpartners(McInerney30
andBryant,1981;Seitzetal.,1988;ScholtenandConrad,2000),creatingasituationinwhich31
continuedgrowthisonlypossiblewhenthetwopartnersco-exist.Manyofthebiodegradation32
processesconsistofindividualsyntrophicandcross-feedinginteractionsamongdifferentspecies33
(Schink,1997),withexamplesincludingthedegradationofmonoaromaticandpolyaromatic34

Cooperationinmicrobialbiotechnology
8
compoundsinsyntrophywithmethanogens(KnollandWinter,1989;Berdugo-Clavijoetal.,2012;1
Morrisetal.,2013).Syntrophicinteractionsarealsoimportantinoil-degradingmicrobial2
communities,althoughtheexactrolesofmanyindividualmembersinthesecommunitiesareless3
clear.Ithasbeenreported,forinstance,thatsyntrophicinteractionsbetweenDesulfatibacillum4
alkenivoransandMethanospirillumhungateiarenecessarytodegraderefractoryhydrocarbons5
(Westerholmetal.,2011;Callaghanetal.,2012).6
Theseexamplesillustratehowubiquitousandessentialsyntrophicinteractionsarefor7
completedegradationoforganiccompounds.Therefore,forfullybeingabletooptimize8
bioprocessesandbiotechnologiesaroundorganicdegradationandtransformationsweneedabetter9
understandingoftheemergenceandmaintenanceofmetaboliccooperations. Itisimportantto10
notethatsyntrophicandcross-feedinginteractionsareshowntoaltercellularmetabolicfluxes11
withinindividualspecies,aswellasinsimplecommunitiessuchthatthepresenceofadownstream12
syntrophicpartnercanresultinchangesinthemetabolicby-productsandyieldsfromupstream13
producermicroorganisms(McInerneyandBryant,1981;Seitzetal.,1988;Schink,1997;Scholten14
andConrad,2000).Inotherwords,organisms’preferredmetabolicroutes(or‘strategies’)would15
changewithlocalsubstrate/productavailabilities(aswellasinternalconstraintssuchasonuptake16
ratesorcofactoravailabilities),buttheseinturnwoulddependonwhatotherorganismswould17
choosetodometabolically.Fromatheoreticalperspective,thissituationcannotbeanalysed18
assumingasimpleindividualfitnessoptimizationunderconstantselectionpressure,butwould19
requireinsteadthecombinationofevolutionarygametheoryandecologyinordertodevelop20
theoreticalframeworksandexperimentalmodelsystemsaccountingforthedescribedcomplex21
interplays.22
Theinclusionofthermodynamicsinmodelsofmicrobialgrowthandmetabolismcould23
contributetounraveltheemergenceofmetabolicinteractions.Takingintoaccountthe24
thermodynamicconstraintsofgrowth-supportingmicrobialbiochemicalreactionswouldenable25
bettercapturingchangesintheconcentrationsofdifferentcompoundsintheenvironmentandthus26
allowdirectlinkagebetweenecologyandindividualgrowthrates.Therehavebeenseveralrecent27
attemptsinthisdirection,andmodelsincludingthethermodynamicsofmetabolicreactionshave28
beensuccessfullyemployedtodescribethedynamicsofsomebiodegradationprocesses,suchasthe29
fermentationofglucoseandthereductionofnitrate(González-Cabaleiroetal.,2013,2015;Cueto-30
Rojasetal.,2015),toexplainmicrobialdiversity(GroßkopfandSoyer,2016),aswellastomodel31
individualspeciesgrowth(HohandCord-Ruwisch,1996;JinandBethke,2007).Additionalworksin32
thisdirectionwillallowbetterpredictivemodelstoexplainevolutionaryandecologicaldynamicsof33

Cooperationinmicrobialbiotechnology
9
microbialcommunitiesunderconditionswherethermodynamics-drivenmetabolicinteractions1
dominate.2
3
Metabolicinteractionsemergingfromcellulartradeoffs4
Asdiscussedabove,fitnessoptimizationisacomplexfunctionofmultipletraitsanditissubjectto5
intrinsictradeoffsthatcouldreadilyexplainmetabolicsecretions.Inparticular,theoptimizationof6
ATP-generatingpathwaysunderlimitationsonenzymeinvestmentandinternalmetabolic7
concentrationsisshowntoleadtotheevolutionofimpartialpathwaysandmetaboliteexcretion8
(PfeifferandBonhoeffer,2004).Similarly,limitationsonmembranespaceandinternalresources9
suchasenzymesandconservedmoietiescancausetradeoffsinsubstrateuptakeratesandinternal10
metabolicfluxes,resultingindifferentgenotypesthatdifferentiallyutilizerespiratory(i.e.pathways11
endingwithinorganicterminalelectronacceptors)andfermentation(i.e.pathwaysendingwith12
organicterminalelectronacceptors)pathways(MajewskiandDomach,1990;Vemurietal.,2006;13
Molenaaretal.,2009;Zhuangetal.,2011;vanHoekandMerks,2012;Flamholzetal.,2013;Basan14
etal.,2015).Sincetheendproductsoffermentativepathwaysareusuallystillabletosustainfurther15
microbialgrowth,thiscouldagainexplainthefirststageofformationofmetabolicinteractions16
throughmetabolicexcretions.Subsequently,limitationsonsubstrateuptakearepredictedtoactas17
aforcetodrivemetabolicspecializationonsuchexcretedcompounds(Doebeli,2002;Spenceretal.,18
2007).19
Theideaofcellulartradeoffsdrivingtheemergenceofmetaboliccross-feedinghasrecently20
beenevaluatedinacombinedinsilicoandexperimentalevolutionstudy(Großkopfetal.,2016).In21
thatstudy,theauthorshaveincorporatedtradeoffsinastoichiometricmetabolicmodelofE.coliby22
imposingglobalconstraintsonthetotaluptakerates.Thismodelwasthensimulatedusing23
dynamicalfluxbalanceanalysis,whichallowsmodellingofbothmicrobialgrowthandenvironmental24
substrateconcentrations,andmutations,whichcanalterthedistributionoftotaluptakefluxamong25
differentsubstrates.Inotherwords,thisapproachcombinedsimulationofecologicaland26
evolutionarydynamicsatthesametime;startingfromasinglemodel,theinsilicosimulationscan27
leadtoalterationsbothintheenvironmentalconditionsandmutantmodels(Fig.2B).The28
applicationofthisapproachtothemodellingoftheexperimentallong-termevolutionofEscherichia29
colirevealedthatthecombinationoftradeoffsandecological/evolutionarydynamicsresultsinthe30
emergenceoftwodominantmodels(Fig.2C).Thesetwomodelshavedistinctuptakefluxes31
suggestiveofacross-feedinginteraction;onemodelhadincreasedglucoseuptakeandacetate32
excretionrateandtheotherhadincreasedacetateuptakerate(Großkopfetal.,2016).Further33
experimentalanalysesrevealedthatthetwomodelsshowmetabolicfluxpatternsthatqualitatively34

Cooperationinmicrobialbiotechnology
10
matchexperimentallyobservedgenotypesinonelineageofthelong-termexperiments,indicating1
thatthisapproachmightprovideusefulinsightsintohowecologicalandevolutionarydynamicscan2
shapemetabolicsystems.Indeed,anemergingtrendintheanalysisofcommunitydynamicsisto3
increasinglycombinemulti-speciesecologicalsimulationswithstoichiometricmodelsdescribingthe4
metabolismofthoseinteractingspeciesinanattempttogenerateinsightsintoecology–5
evolutionaryinterplay(LoucaandDoebeli,2015;Widderetal.,2016;ZomorrodiandSegrè,2016).6
7
Factorscontributingtothestabilizationofcooperativeinteractionsinmicrobialpopulations8
9
Structuredenvironments10
Oneofthebasicmechanismsthataffecttheresilienceofcooperationisthepresenceofspatial11
structure.Structurewouldultimatelyfacilitatetheresilienceofcooperationasitallowsthe12
‘segregation’ofcooperativefromcheatingcells(Nowak,2006)(i.e.,cooperativecellscanthenshare13
theproducedpublicgoodwiththesimilartrait,excludingcheatingcells)(Fig3A).14
Thereareseveraltheoreticalstudiesandexperimentalevidencesofspatialsegregationin15
cellularpopulations(VanDykenetal.,2013),withbiofilmsbeingaparadigmaticexampleofbacterial16
communitiesexhibitingstablecooperationduetothesegregationinstructuredenvironments17
(Nadelletal.,2009).Thestructureandcompositionofbiofilmscanfeedbackonthehighlydynamic18
competitionbetweensub-populationsofcooperators(i.e.,contributingtothebiofilmassembly)and19
cheaters.Inthesecircumstances,thespatialarrangementsofthedistinctgenotypescruciallyaffect20
thedegreeofcooperationandcompetitionpresentinthebiofilm(Nadelletal.,2016).21
Abroadernotionofstructurecanalsorefertothecaseofhavingapopulationdistributed22
intodifferentheterogeneoussub-populationsthatmaybespatiallysegregated(e.g.forming23
colonies).Inthiscasethestructureofthepopulationcanleadtoacharacteristicissueofmulti-level24
selectionknownasSimpson’sparadox.Simpson’sparadoxisastatisticalphenomenomthatcan25
emergewhencomparinggroupsofdata;groupscandisplayatrendwhenanalysingthem26
individually,butthistrendisreversedwhenthegroupsarecombined.Afamousexampleof27
Simpson’sparadoxistheonebehindthegenderdiscriminationaccusationagainsttheUniversityof28
Berkeleyinearly1970s.Inthatcase,44%ofthetotalmaleapplicationstothegraduateschoolwere29
acceptedagainstthe35%ofthefemaleapplicantssuggestingabiasagainstfemaleapplicants.30
Lookingintohowtheapplicationsweredistributedamongthedifferentdepartments,however,it31
becameclearthattherewasnobias,andthedifferencesintheratesweretheresultofamajorityof32
womenhavingappliedtothemostcompetitivedepartments,whichdecreasedthesuccessrateof33
thefemaleapplicants.Inotherwords,theapparentbiasisonlytheresultofthewaysthe34

Cooperationinmicrobialbiotechnology
11
applicationsareaggregatedtogether(Bickeletal.,1975).Inthecontextofmicrobialcommunities,1
Simpson’sparadoxisshowntoemergewhenthedifferentsub-groupsaresufficiently2
heterogeneousintheircompositiontoguaranteethatintheaggregatepopulationthecooperative3
individualshaveanadvantageoverthecheatingcells(despiteineachofthecolonies–the4
disaggregatedpopulation–cheatersarefavoured)(Chuangetal.,2009).Thisfindingsuggeststhat5
theopportunedesignoftheorganizationofamicrobialcommunityinsub-populations(and6
subsequentcoalescenceofthosesub-populations)maybeusefultoimproveitsresilienceto7
detrimentalmutants.Ingeneral,othermorecomplexnotionsofstructuredpopulationsfrom8
ecology(e.g.,meta-populationdynamics)couldalsoberelevanttounderstandandcontrolthe9
evolutionarydynamicsofcooperativeinteractions(Dattaetal.,2013).10
11
Interplaybetweenecologicalandevolutionarydynamics12
Anotherstabilisinganddrivingfactorbeyondcooperativeinteractionsinmicrobialcommunitiesis13
theinterplaybetweenecologicalandevolutionarydynamicsthatresultsinchangesinthe14
compositionofthecommunityovertime.Thishappenswhen,duetotheinteractionsina15
community,certaintraits(suchascheatingandcooperation)areselectedfororagainst,resultingin16
rapidchangesinthefrequencyoftheindividualscarryingthetraitthataffecttheecologyofthe17
globalcommunity.Thechangesintheecologycanthenfeed-backontheselectiveadvantageofthe18
differenttraits(asdiscussedabove),leadingtoaneco-evolutionaryfeedback(Fig.3B)(Lennonand19
Denef,2015).Thisaspecthasbecomeofrecentinterestduetoseveraltheoreticalandexperimental20
studiesshowingthenon-trivialeffectsofthetime-scalesoverlapbetweenecologyandevolutionin21
whatarecalledeco-evofeedbacks(Schoener,2011).Thereareseveralexamplesofeco-evo22
feedbacksinmicrobialpopulationsinvestigatedexperimentally(FiegnaandVelicer,2003;Ross-23
Gillespieetal.,2009;Moreno-Fenolletal.,2017)withthemostknownexamplebeingtheinterplay24
betweenpopulationdensityandfitness(SanchezandGore,2013).Forinstance,intheyeast25
communitiesdiscussedabove,cooperativecellshavehigherfitnessthancheatingcellsonlyatlower26
populationdensity.This,coupledtothefactthatcheatersleadtolowerpopulationgrowth,27
facilitatestheobservedco-existencebetweenthetwotraits,i.e.thestabilisationofcooperation28
(SanchezandGore,2013).Eco-evofeedbackscanbemodelledbyaddingnotionsofpopulation29
dynamicstoevolutionarygametheory,leadingtotheframeworkofecologicalpublicgoodgames30
(Hauertetal.,2008)thatextendthestandardevolutionarygametheory(inwhich,usually,thefocus31
oftheanalysisisthechangeinfrequencyofacertaintrait).Combinationofpopulationdynamics32
withmetabolicmodelsatthelevelofindividualspeciesorgenotypes(Harcombeetal.,2014)with33

Cooperationinmicrobialbiotechnology
12
evolutionarydynamics(Großkopfetal.,2016)isanotherpromisingroutetowardscapturingeco-1
evolutionarydynamics,especiallywhencooperativeinteractionsinvolvemetabolitesecretions.2
3
Regulatorymechanisms4
Anotherpotentialfactorforthestabilisationofcooperationthathasrecentlyattractedattentionis5
cellularregulatorymechanisms.Animals,includinghumans,havedevelopedcomplexsocial6
strategiestocontrolcheaters,andthereisgreatinterestindeterminingtowhichextentsinglecell7
organismscouldemploysimilarmechanismstofightdetrimentalmutants(TravisanoandVelicer,8
2004).9
Oneoftheseregulatorymechanismsisknownas‘reciprocity’.Inthiscasetheamount10
contributedofapublicgooddependsontheenvironmentalconditions,whichinturnmaydepend11
onthecontributionsmadebyothers.ThisisforinstancethecaseofironuptakeinP.aeruginosa12
whereironscavengingsiderophores(thepublicgood)arereleasedingreaterorsmallerquantities13
dependingontheamountofironintheenvironment(Kümmerlietal.,2009).Recentexperiments14
usingthissystemhaveconfirmedthatcellsuseatypeof‘reciprocity’thatfacilitatesthecontrolof15
cheaters:thecellulardecisionofproducingpublicgoodismadeonlyinanenvironmentwithmany16
producers.Inotherwords,thecellsseemtoimplementarulestating‘cooperatewhensurrounded17
bymostlycooperators’.Coupledtoquorumsensing,thisruleallowsbacteriatomatchtheir18
investmentatlowerlevelsofpopulationstructuringanditisaneffectivewaytorepresscheaters19
(Allenetal.,2016).Inyeast,asimilarmechanismhappensintheproductionofinvertase.Another20
regulatorymechanismthatcouldbeinterpretedasafunctional‘decision’tolimitthespreadof21
cheatersistoincreasethenoiseintheexpressionofgenesencodingforpublicgoods(Goreetal.,22
2009).Thisisthecaseofself-destructivecooperation,inwhichcooperativecellsdiewhilehelping23
others,forexample,asithappensduringthesecretionoftoxinsthatenhancethecolonizationof24
tissuesbycertainbacterialpathogens(Ackermannetal.,2008).Sincethetoxinisgenetically25
encoded,itisonlyexpressedbyafractionofthepopulationorthewholemicrobialpopulation26
woulddie.The‘decision’onwhichcellsmaketheultimatesacrificeisgivenbythestochastic27
expressionofthegeneencodingthetoxin.Similarly,cell-cellvariabilityintheproductionofother28
kindsofpublicgoodsmayallowcooperativecellstotemporarilyswitchofftheproductionofapublic29
good,thereforelimitingitscostandallowingforenhancedcompetitionagainstthecheatingcells.30
Thesetypesofcellulardecision-makingmechanismscaninterplaywithanunderlyingeco-31
evodynamics(HarringtonandSanchez,2014)andcruciallyaffecttheresilienceofcooperation,as32
shownintheoreticalmodels(CavaliereandPoyatos,2013)(Fig.3C).Thus,itisplausibletopropose33
thecontrolofpublicgoodproductionforsuccessfulbioprocesses(suchasthedescribedcellulose34

Cooperationinmicrobialbiotechnology
13
degradation)throughexistinggeneregulatorymechanismsorbyengineeringsuchmechanismsde1
novo.2
3
Horizontalgenetransferofcooperativetraits4
Mobilegeneticelements(plasmids,bacteriophages,transposons,etc.)transmittedviahorizontal5
genetransferareoneofthemainfactorscontributingtoshapingmicrobialevolution.Apartfromthe6
genesessentialforthereplicationandtransmissionessentialforthemobileelements,theyoften7
carrymultipletraitstheenablesocialinteractionsinmicrobialcommunitiesandmakethemactive8
agentsdefiningtheevolutionarydynamicsofthesecommunities(Rankinetal.,2011).9
Cooperativetraitssuchaspublicgoodproducingexoenzymesarecommonlyacquireddueto10
thetransferenceofmobileelements.Infact,agenomicanalysisinsomebacterialspeciesshowthat11
thefrequencyofgenesencodingextracellularproteinsissignificantlyhigherinchromosomal12
locationsknowntobetransferred(e.g.transposons)comparedtoregionsthatarenot,andthe13
frequencyisevenhigherinplasmids,whichwerethemostmobileelementspresentintheanalysis14
(Nogueiraetal.,2009).Horizontalgenetransferisalsoresponsibleforthetransmissionof15
exoenzymesineukaryoticmicroorganisms,asrevealedbyasimilaranalysiscarriedoutin16
osmotrophicfungi,inwhichitbecameevidentthatnotonlytheenzymes,butalsothetransporters17
requiredfortheuptakeoftheproductsresultingfromtheactivityoftheenzymesonlargepolymers,18
wereencodedinmobilegeneticelements(RichardsandTalbot,2013).19
Theseobservationsareconsistentwiththeideaofmobileelementsenablingcooperationin20
acommunityowingtotheinvasionofmobileelementstransmittingcooperativetraits.However,the21
mobileelementsalsogenerateacosttothecellsharbouringthemand,therefore,canpotentiallybe22
lostoroutcompetedby‘cheat’geneticelements(Rankinetal.,2011).Recentexperimental23
evidencesshowneverthelessthathorizontalgenetransferhelpstomaintaintheproductionof24
publicgoodsdespitethepotentialpresenceofnon-cooperativeorganismsandnon-cooperative25
mobileelements(Dimitriuetal.,2015)owing,amongotherfactors,totheincreaseingenetic26
relatednessduetothepresenceofthemobileelements(McGintyetal.,2013).Inotherwords,27
transmissiblemobileelementsallowforthelocalenrichmentincooperativeinteractions,which28
may,inthelongterm,leadtothespecializationofsub-populationsincooperativenichesspeciallyin29
thepresenceofstrongstructure(Niehusetal.,2015).30
31
Therelevanceofcooperationforbiotechnologicalapplications32
Thepresenceofcooperativeinteractionsfacilitatesthedevelopmentofcomplexfunctionsthat33
wouldbeotherwisedifficultorimpossible(Nowak,2006).34

Cooperationinmicrobialbiotechnology
14
Cooperativemicrooorganismscanexhibitdistributionoflabour:alargecollectionofdistinct1
phenotypicbehaviours,organizedinsubpopulations,cancoordinatetofulfilsomecomplextasksina2
collectiveway(Fig4A).Shareddiffusiblemoleculesallowcellstocommunicateandspatially3
distributethelabour.Examplesofcomplextasksrangefromthecontrolledgrowthofbiofilms4
dependingonenvironmentalconditions(Liuetal.,2015;Kimetal.,2016)tothedistributed5
computationofBooleanfunctions(Regotetal.,2011).6
Thistypeofinteractioniscommonlyobservedinbiodegradativeprocessescarriedoutby7
interspeciesbiofilms.Forinstance,thepresenceofaalgaeinamicrobialconsortiumwithmorethan8
ninebacterialspeciesenhancesthedegradationofthepesticidediclofopmethyl(Wolfaardtetal.,9
1994).Anotherinterestingexampleisthesyntrophicinteractionbetweenthenon-cellulolytic10
speciesTreponemabryantii,andthecellulolyticspeciesRuminococcusflavefaciens,toenhancethe11
rateofcellulosedegradation.TheslowlygrowingculturesofR.flavefaciensbenefitsfromT.bryantii12
removingthecellulolyticproduct,whichresultsinhigherpopulationdensityanddegradationrates13
(Jamesetal.,1995).14
Distributionoflabouris,however,notrestrictedtospatiallystructuredpopulationsor15
populationscomposedbymorethanonespecies,butcanalsoapplytootherbiologicalprocesses16
likethebiochemicalpathwaysforthedegradationofaromaticsinpopulationscomposedofone17
strain(Nikeletal.,2014).Thesepathwaysaresometimesorganisedintotwodistinctgeneoperons,18
oneencodingfortheactivitiesrequiredtofunnelthearomaticsubstrateintoamoreaffordable19
aromaticcarbonsourceandasecondrequiredtotransformthisaromaticcompoundintocentral20
metabolites.Forinstance,theTOLpathwayofPseudomonasputidaresponsiblefortolueneand21
xylenedegradationcontainsan‘upper’partthatconvertstolueneintobenzoate,anda‘lower’22
segmentresponsibleforthedegradationofbenzoate(Franklinetal.,1981).Inprinciple,itwouldbe23
expectedthatallcellsexpressbothoperonswhenaclonalpopulationofP.putidaisculturedinthe24
presenceoftoluenebut,surprisingly,manyofthecellsdisplayanearbimodaldistribution25
expressingeitheroneoperonortheother(Nikeletal.,2014).Themechanisticexplanationofthis26
behaviourisunknownalthoughaplausibleexplanationofthephenotypicdistributionmayarise27
fromtheintricatetranscriptionalcontroloftheoperons(Silva-RochaanddeLorenzo,2012).28
Distributionoflabouralsoappearsintheanaerobicmetabolismofaromaticcompoundsin29
Rhodopseudomonaspalustris.Monoculturesofthisspeciesorganiseinthreedifferent30
subpopulationswhenusingp-coumarateorbenzoateasthecarbonsource.Eachofthese31
subpopulationsisresponsiblefortheutilizationofeitherthearomaticcompound,CO2andH2or,32
whengrowingonbenzoate,N2andformate,formingasyntrophicconsortiadefactocomposedofa33
singlespecies(Karpinetsetal.,2009).However,whetherthisparticulartypeofcooperativecross-34

Cooperationinmicrobialbiotechnology
15
feedinginteractionisadvantageoustopreventwasteofresourcesoraccumulationoftoxic1
intermediatesisanopenquestion.2
Distributionoflabourcanalsobeengineeredtogetherwithcooperativetraitsin‘synthetic’3
communities(Fig.4B).Thisisthecaseofco-culturingengineeredstrainsofthebacteriumE.coliand4
theyeastS.cerevisiaethatareartificiallymutualistic.Eachofthesestrainsismodifiedtoexpressone5
moduleofthebiosyntheticpathwayofanantitumoralcompoundofinterest(theacetylateddiol6
paclitaxelprecursor).Thecooperationbetweenthesespeciesallowsproductionoftaxaneswith7
higheryieldsthanusingE.colialone.ThemixedculturecombinesthecapabilitiesofE.colifor8
producingtheintermediatetaxadienewiththesuperiorpropertiesofS.cerevisiaecomparedtoE.9
colitocatalysetheoxygenationreactionsrequiredtorenderthefinalcompound(Zhouetal.,2015).10
Syntheticconsortiacanbeusedinbioprocessesevenintheabsenceofmutualismasexplainedin11
theprevioussections(e.g.ifeco-evofeedbackstakeplace).Thisisthecaseofanartificial12
communitydesignedtoproduceisobutanolfromcellulosicbiomasscomposedbythefungus13
TrichodermareeseiandanengineeredstrainofE.coli.InthisconsortiumT.reseeiactsasa14
cooperatorsecretingcellulasesrequiredtodegradelignocellulosicpolymersandtheresulting15
saccharidesareusedtofeedtheE.colistrainthatdeliversthefinalproduct(Mintyetal.,2013).16
Syntheticcommunitiescanalsoimprovebiodegradationprocessescomparedtomonocultures.17
Degradationofcrudeoilisagoodexampleinwhichmicrobialcommunitiescanexhibitcooperative18
interactionsinNatureincludingmetaboliccross-talkandsharedgoodsthatmaycontributetothe19
formationofinterspeciesbiofilms(McGenityetal.,2012).Moreover,theseinteractionscanbe20
harnessedtoproduceartificialcommunitieswithenhanceddegradationcapabilitiessuitableforoil21
removal(Gallegoetal.,2007).Anotherexampleisthedesulphurizationofdibenzothiophene(DBT)22
toformsulphur-free2-hydroxybiphenyl.Inarecentwork,DBTdesulphurizationwascarriedout23
usingeitheranengineeredP.putidastrainexpressingallthedszABCDgenesrequiredintheprocess,24
oramixedcultureofthesamestrainexpressingonlysomeofthegenes.Inthisexperiment,25
desulphurationofDBTwashigherwhencombiningmultiplecells‘specialising’inonestepofthe26
biochemicalpathwaycomparedtothecaseofhavingallreactionstakingplaceinthesameorganism27
(Martínezetal.,2016).28
Cooperativeinteractionsinmicrobialcommunitiescanalsoleadtohigherresistanceto29
environmentalandecologicalstress.Empiricalobservationsusingartificialcommunitiesofyeast30
showthatthisresistancetakesplaceoverawiderangeofconditions(Goreetal.,2009).Inaddition,31
experimentscarriedoutwithengineeredpopulationsofBacillussubtilislackingtheabilitytoform32
biofilmsshowthattheyneverthelesstendtoformclustersthat,althoughcanhavereducedgrowth33
duetolimitedmobility,allowthecellstoendureharshenvironmentalconditions(RatzkeandGore,34

Cooperationinmicrobialbiotechnology
16
2016).Inthiscase,cooperativeindividualstendtoaggregateleadingtothe‘privatization’ofpublic1
goodsandtotheexclusionofcheatingindividuals(Pandeetal.,2016).Ontheotherhand,thelossof2
cooperationmakescellularcommunitiesmorefragile(SanchezandGore,2013)andmorevulnerable3
tocompositionalshiftsarising,forexample,fromantibiotictreatments(Liuetal.,2015).Thefact4
thatthesebehavioursareobservedinexperimentswithdifferentmanipulatedspeciessuggeststhat5
thesemechanismsaregeneralandcouldbecommonplaceinNature.6
Thepresenceofmechanismsthatfacilitatecooperationcanalsoleadtocomplexco-7
evolutionarydynamicswiththeconsequentemergenceofnovelsocialinteractions.Themost8
significantexampleinthisrespectisthemechanismofquorumsensing(QS)thatisinvolvedin9
controllingtheinvestmentin‘publicgoods’(Allenetal.,2016).AlthoughtheoriginalroleofQSis10
unknown,itsabilitytofacilitatethe(beneficial)presenceofcooperativeinteractionsmayhaveledto11
theselectionofcomplexfunctionalities,e.g.,coordinatingtheexpressionofgenesinvolvedin12
multiplecooperativestrategies,oftenco-evolvingwiththem(Popatetal.,2015).Thisexample13
suggeststhepossibilityofusingthepresenceofcooperativeinteractionstodirecttheevolutionof14
thecommunitiestowardsotherpropertiesofinterest.15
16
Conclusion17
Thekeypointofevolutionarygametheoryisthatthefitnessofindividualsdependsnotonlyonthe18
environmentbutalsoonothermembersinthepopulation.Thistheoryprovidesaframeworkto19
understandthedynamicsofmanybioprocessesinvolvingcomplexmicrobialpopulations(natural20
andsynthetic)inwhichthefitnessofanindividualcellisinfactaffectedbytheenvironmentandby21
thepresenceofothercells.Aparticularcaseofthisscenarioconcernsthepresenceofcooperative22
interactionsbasedonpublicgoodsandmetabolicinteractionsandthathavebeenthemainfocusof23
thisreview.Wehavealsodiscussedsomeofthefactorsshapingtheseinteractionssuchascellular24
andthermodynamicconstraints,aswellasfactorsstabilisingthemsuchasstructuredenvironments,25
feedbacksarisingfromtheecologyofthepopulation,cellularregulatorymechanismsimplementing26
certainbehaviouralstrategiesandtheroleofmobilegeneticelements.Thesepropertiesendow27
cooperativemicrobialpopulationswiththepossibilitytoresistcheatersinvasionsandthecapability28
ofperformingmoresophisticatedtasks.29
Despiteitsgrowingusetostudytheevolutionofcooperation,evolutionarygametheoryhas30
hadsofaraverylimitedimpactinfieldorindustrialbiotechnologicalapplicationsinwhichthe31
environmentalconditionsaregenerallynotwell-definedandmayaffectthemicrobialcommunities32
(Bouchezetal.,2000;SaylerandRipp,2000;CasesanddeLorenzo,2005).Infact,wehave33
presentedseveralexamplessuggestingthatcooperativeinteractionsbasedoncross-feedingand34

Cooperationinmicrobialbiotechnology
17
publicgoodsareatthecoreofmanyprocessesrelevantforindustrialbiotechnologyincludingfood,1
energyandenvironmentalapplicationsofmicroorganisms.2
Therefore,theyaresuitableofimprovementbyincorporatingthemechanismsinvestigated3
inthelargeliteratureoftheevolutionofcooperation.Aswehavediscussed,populationscouldbe4
manipulatedbasedonthermodynamicconstrainstopromotecertainmetabolic(cooperative)5
interactions.Similarly,bioprocesses,includingbioreactordesign,couldbeengineeredtoaccount6
(andexploit)foreco-evofeedbacksandspatialorganizations.7
Understandinghowsyntrophyandcooperationendowthemicrobialpopulationswith8
resistanceandresilienceagainstecologicalandenvironmentaldisturbanceslikecompositionalshifts9
intheenvironmentorantibioticshockscouldbeusedtoengineerrobustmicrobialcommunities10
withenhancedperformanceandpredictabledynamics(BrionesandRaskin,2003;Allisonand11
Martiny,2008;Sözenetal.,2014).Overall,webelievethatthemigrationofresultsand12
methodologiesfromtheareaofevolutionarygametheoryintothedesignofmicrobialconsortia13
wouldfacilitatetheengineeringofevolutionaryresilientcommunitieswithabetterperformancein14
awiderangeofbiotechnologicalapplications.15
16
Acknowledgements17
JJwouldliketoacknowledgethesupportreceivedfromtheEuropeanUnion'sHorizon2020research18
andinnovationprogrammeundergrantagreementno.633962fortheprojectP4SBandthesupport19
fromtheBiotechnologyandBiologicalSciencesResearchCouncil(BBSRC)(grantBB/M009769/1).20
OSSacknowledgessupportfromBBSRC(grantsBB/K003240/1andBB/M017982/1).M.C.21
acknowledgesthesupportfromtheEngineeringandPhysicalSciencesResearchCouncil(EPSRC)22
grantEP/J02175X/1andfromUKResearchCouncils'SyntheticBiologyforGrowthprogramme.S.F.23
acknowledgesthesupportbyLaboratoryDirectedResearch&Development(LDRD)grant24
XWJX00/3000FENGfromLosAlamosNationalLaboratory.Theauthorsdonotahaveconflictof25
interesttodeclare.26
27

Cooperationinmicrobialbiotechnology
18
1
Figure1.(A)Interactionsbasedonsharedpublicgoods.Somecells(cooperators,showninblack2
edge)produceanenzymerequiredtosplitasubstrateintodigestibleproducts.Othercells(cheats,3
showningrey),donotproducetheenzymebuttakeadvantageofthepublicgoodsproducedbythe4
others.(B)Interactionsbasedoncross-feeding.Somecellsinthecommunityexcretemetabolites5
thatcanbetakenupbyothercellsgivingrisetoawebofinteractions.6
7

Cooperationinmicrobialbiotechnology
19
1Figure2.(A)Metabolicinteractionsthatcantakeplaceinapopulation.Cellscanexchangemetabolitesthat2
arerequiredtosupporteachother’sgrowthinamutualisticinteraction(left).Oneofthecellscanusea3
metaboliteexcretedbyanothercell,favouringinthiswaythemetabolismoftheproducerthroughthe4
pathwaysleadingtotheexcretion(centre).Whenthemetabolitesexcretedhaveaninhibitoryeffectonthe5
producer(e.g.becausetheyleadtothermodynamicequilibrium),therelationshipwithadegradercellofthe6
inhibitorymetaboliteismutuallybeneficialandknownassyntrophy(right).(B)Dynamicmodellingofthe7
evolutionofFBAmodels.Cellscanbemodelledasmetabolicnetworksexchangingmetaboliteswithother8
cellsinthepopulation.InthisabstractioneachcellisrepresentedbyaFluxBalanceAnalysismodel.These9
modelscanreplicateovertimeandalsoevolve,producingpopulationscomposedbymodelswithdifferent10
constrainsforuptakeandsecretionofmetabolites.(C)Dynamicanalysisofmodelgenealogy.Thefrequency11
ofeachmodelinthepopulationchangesovertimebeingthedarkestbarsthemostabundantmodels.Dueto12
mutations,newmodelsariseandtheyarerepresentedasnewbranchesinthephylogeny.Plotredrawnfrom13
(Großkopfetal.,2016).14
15
Mutualism Cross-Feeding Synthrophy
A
B
FBA1
FBA2FBA3
FBA4
Generation n
FBA1
FBA1*
Generation IGenerations
Mod
els
C
Cooperationinmicrobialbiotechnology
20
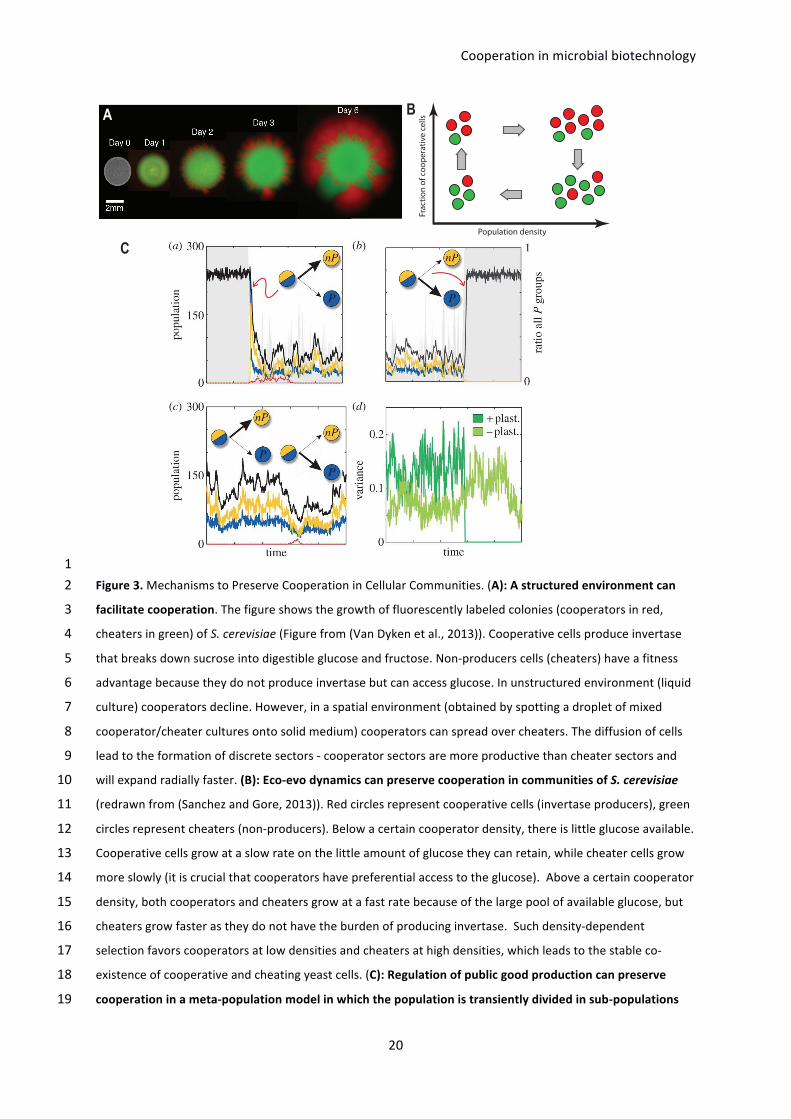
1Figure3.MechanismstoPreserveCooperationinCellularCommunities.(A):Astructuredenvironmentcan2
facilitatecooperation.Thefigureshowsthegrowthoffluorescentlylabeledcolonies(cooperatorsinred,3
cheatersingreen)ofS.cerevisiae(Figurefrom(VanDykenetal.,2013)).Cooperativecellsproduceinvertase4
thatbreaksdownsucroseintodigestibleglucoseandfructose.Non-producerscells(cheaters)haveafitness5
advantagebecausetheydonotproduceinvertasebutcanaccessglucose.Inunstructuredenvironment(liquid6
culture)cooperatorsdecline.However,inaspatialenvironment(obtainedbyspottingadropletofmixed7
cooperator/cheaterculturesontosolidmedium)cooperatorscanspreadovercheaters.Thediffusionofcells8
leadtotheformationofdiscretesectors-cooperatorsectorsaremoreproductivethancheatersectorsand9
willexpandradiallyfaster.(B):Eco-evodynamicscanpreservecooperationincommunitiesofS.cerevisiae10
(redrawnfrom(SanchezandGore,2013)).Redcirclesrepresentcooperativecells(invertaseproducers),green11
circlesrepresentcheaters(non-producers).Belowacertaincooperatordensity,thereislittleglucoseavailable.12
Cooperativecellsgrowataslowrateonthelittleamountofglucosetheycanretain,whilecheatercellsgrow13
moreslowly(itiscrucialthatcooperatorshavepreferentialaccesstotheglucose).Aboveacertaincooperator14
density,bothcooperatorsandcheatersgrowatafastratebecauseofthelargepoolofavailableglucose,but15
cheatersgrowfasterastheydonothavetheburdenofproducinginvertase.Suchdensity-dependent16
selectionfavorscooperatorsatlowdensitiesandcheatersathighdensities,whichleadstothestableco-17
existenceofcooperativeandcheatingyeastcells.(C):Regulationofpublicgoodproductioncanpreserve18
cooperationinameta-populationmodelinwhichthepopulationistransientlydividedinsub-populations19
A
C
B
Population density
Frac
tion
of c
oope
rativ
e ce
lls

Cooperationinmicrobialbiotechnology
21
(figurefrom(CavaliereandPoyatos,2013)).In-silicosimulationspresenttwopossiblesuccessfultypesof1
regulationagainstcheaters:positiveplasticity(toprow)inwhichcooperatorsconstraintcheatersbystopping2
theproductionofpublicgoodwhencheatersappear(a)andfullyrestartingonlywhencheatershave3
disappeared(b)andnegativeplasticity(bottomrow)inwhichcooperatorsproducepermanentlylow4
amountsofpublicgoodwhichhelpscontrollingcheatersinvasion(c).Thickarrowsdenotethecellulardecision5
toproduce(P)ornotproduce(nP)thepublicgood.Thesuccessoftheregulationiscoupledtothe6
heterogeneity(variance)ofthesub-populations,i.e.,positiveplasticitytransientlymodifiesthevariancewhile7
negativeplasticitykeepsarelativelyconstantheterogeneity(varianceshownin(d)correspondto8
trajectories(b)and(c),respectively).9
10

Cooperationinmicrobialbiotechnology
22
12
Figure4.(A)Divisionoflabourinmicrobialpopulations.ColoniesofPseudomonasfluorescensP0-1are3
composedbycellswithtwodifferentmorphologiesknownasmucoidanddrythatcanevolvefromeachother4
duetoasinglemutation(leftpicture).Coloniescomposedbyamixtureofthetwophenotypesexpandfaster5
allowingcellstocoloniselargerregionsinshorterperiodsoftimecomparedtocoloniescomposedbyeachof6
theindividualphenotypes.Thetwomorphotypesoccupydifferentregionsofthecolonyasshownwhen7
labelledwithfluorescentreporters(centre).Drycells(inred)exhibitaradialdistributiongrowingontopofthe8
mucoid(ingreen).Confocalmicroscopyrevealsthattheedgeofthecolony(rightpicture)displaysadistinct9
spatialorganizationinwhichmucoidcellsformathinstripattheveryedge.Thedifferentiationandspatial10
segregationallowsthedistributionoflabourinthepopulation:Mucoidcellsproducealubricantpolymerat11
theedge,whereasdrycellssitbehindandpushbothofthemalong.Thecooperationofthesetwophenotypes12
resultsinafastgrowingcolony.Pictureshavebeenreproducedfrom(Kimetal.,2016).(B)Engineered13
populationscanimprovebioprocesses.Twostrainsarecombinedtocarryoutthesynthesisofaproductof14
interest(redpentagons)thatcannotbeproducedusingeachofthestrainsindividually.Theprocessinvolves15
thatoneofproducesanintermediate(theyellowpentagon)thatisusedbytheothertosynthesizethefinal16
product.Ifthetwocellscompeteforthesameresources(e.g.carbonsourceshownbythebluehexagon;left17
panel)thepopulationwiththelowerfitnessunderthoseconditionswilleventuallycollapse.However,when18
thetwopopulationsareengineeredsothatonegrowsattheexpensesoftheother(e.g.throughcross-feedor19
syntrophyshownbythepurpletriangle),thetwopopulationscooperate(centrepanel)andthesynthesisof20
theproductofinteresttakesplaceforalongerperiodoftimeresultinginhigheryields(rightpanel).Panels21
inspiredby(Zhouetal.,2015).22
time
A
B
Competition Cooperationprod
uct
competitioncooperation

Cooperationinmicrobialbiotechnology
23
Bibliography1
2
Ackermann,M.,Stecher,B.,Freed,N.E.,Songhet,P.,Hardt,W.-D.,andDoebeli,M.(2008)3Self-destructivecooperationmediatedbyphenotypicnoise.Nature454:987–990.4
Adami,C.,Schossau,J.,andHintze,A.(2016)Evolutionarygametheoryusingagent-based5methods.PhysLifeRev19:1–26.6
Allen,R.C.,McNally,L.,Popat,R.,andBrown,S.P.(2016)Quorumsensingprotectsbacterial7co-operationfromexploitationbycheats.ISMEJ10:1706–1716.8
Allison,S.D.(2005)Cheaters,diffusionandnutrientsconstraindecompositionbymicrobial9enzymesinspatiallystructuredenvironments.EcolLett8:626–635.10
Allison,S.D.andMartiny,J.B.(2008)Resistance,resilience,andredundancyinmicrobial11communities.ProcNatlAcadSciUSA105Suppl1:11512–11519.12
Arnesen,S.,HavnEriksen,S.,Olsen,J.andJensen,B.(1998)Increasedproductionofα-13amylasefromThermomyceslanuginosusbytheadditionofTween80.Enzyme14MicrobTechnol23:249–252.15
Avery,S.V.(2006)Microbialcellindividualityandtheunderlyingsourcesofheterogeneity.16NatRevMicrobiol4:577–587.17
Axelrod,R.M.(1990)Theevolutionofcooperation,PenguinBooks,London.18
Bachmann,H.,Fischlechner,M.,Rabbers,I.,Barfa,N.,BrancodosSantos,F.,Molenaar,D.,19andTeusink,B.(2013)Availabilityofpublicgoodsshapestheevolutionofcompeting20metabolicstrategies.ProcNatlAcadSciUSA110:14302–14307.21
Basan,M.,Hui,S.,Okano,H.,Zhang,Z.,Shen,Y.,Williamson,J.R.,andHwa,T.(2015)22OverflowmetabolisminEscherichiacoliresultsfromefficientproteomeallocation.23Nature528:99–104.24
Berdugo-Clavijo,C.,Dong,X.,Soh,J.,Sensen,C.W.,andGieg,L.M.(2012)Methanogenic25biodegradationoftwo-ringedpolycyclicaromatichydrocarbons.FEMSMicrobiolEcol2681:124–133.27
Bickel,P.J.,Hammel,E.A.,andO’connell,J.W.(1975)Sexbiasingraduateadmissions:data28fromBerkeley.Science187:398–404.29
Bouchez,T.,Patureau,D.,Dabert,P.,Juretschko,S.,Doré,J.,Delgenès,P.,etal.(2000)30Ecologicalstudyofabioaugmentationfailure.EnvironMicrobiol2:179–190.31
Briones,A.andRaskin,L.(2003)Diversityanddynamicsofmicrobialcommunitiesin32engineeredenvironmentsandtheirimplicationsforprocessstability.CurrOpin33

Cooperationinmicrobialbiotechnology
24
Biotechnol14:270–276.1
Burns,R.G.(2010)Howdomicrobialextracellularenzymeslocateanddegradenaturaland2syntheticpolymersinsoil.In,Xu,J.andHuang,P.M.(eds),MolecularEnvironmental3SoilScienceattheInterfacesintheEarth’sCriticalZone.SpringerBerlinHeidelberg,4Berlin,Heidelberg,pp.294–297.5
Callaghan,A.V.,Morris,B.E.L.,Pereira,I.A.C.,McInerney,M.J.,Austin,R.N.,Groves,J.T.,et6al.(2012)ThegenomesequenceofDesulfatibacillumalkenivoransAK-01:ablueprint7foranaerobicalkaneoxidation.EnvironMicrobiol14:101–113.8
Carlquist,M.,Fernandes,R.L.,Helmark,S.,Heins,A.-L.,Lundin,L.,Sørensen,S.J.,etal.9(2012)Physiologicalheterogeneitiesinmicrobialpopulationsandimplicationsfor10physicalstresstolerance.MicrobCellFact11:94.11
Cases,I.anddeLorenzo,V.(2005)Geneticallymodifiedorganismsfortheenvironment:12storiesofsuccessandfailureandwhatwehavelearnedfromthem.IntMicrobiol8:13213–222.14
Cavaliere,M.andPoyatos,J.F.(2013)Plasticityfacilitatessustainablegrowthinthe15commons.JRSocInterface10:20121006.16
Chen,S.,Su,L.,Billig,S.,Zimmermann,W.,Chen,J.,andWu,J.(2010)Biochemical17characterizationofthecutinasesfromThermobifidafusca.JournalofMolecular18CatalysisB:Enzymatic63:121–127.19
Chuang,J.S.,Rivoire,O.,andLeibler,S.(2009)Simpson’sparadoxinasyntheticmicrobial20system.Science323:272–275.21
Coleman,G.andElliott,W.H.(1962)Studiesonalpha-amylaseformationbyBacillussubtilis.22BiochemJ83:256–263.23
Cooper,M.B.andSmith,A.G.(2015)Exploringmutualisticinteractionsbetweenmicroalgae24andbacteriaintheomicsage.CurrOpinPlantBiol26:147–153.25
Cueto-Rojas,H.F.,vanMaris,A.J.A.,Wahl,S.A.,andHeijnen,J.J.(2015)Thermodynamics-26baseddesignofmicrobialcellfactoriesforanaerobicproductformation.Trends27Biotechnol33:534–546.28
Datta,M.S.,Korolev,K.S.,Cvijovic,I.,Dudley,C.,andGore,J.(2013)Rangeexpansion29promotescooperationinanexperimentalmicrobialmetapopulation.ProcNatlAcad30SciUSA110:7354–7359.31
Dimarogona,M.,Topakas,E.,andChristakopoulos,P.(2012)Cellulosedegradationby32oxidativeenzymes.ComputStructBiotechnolJ2:e201209015.33

Cooperationinmicrobialbiotechnology
25
Dimitriu,T.,Misevic,D.,Lindner,A.B.,andTaddei,F.(2015)Mobilegeneticelementsare1involvedinbacterialsociality.MobGenetElements5:7–11.2
Doebeli,M.(2002)Amodelfortheevolutionarydynamicsofcross-feedingpolymorphisms3inmicroorganisms.Populationecology44:59–70.4
Doyle,E.,Noone,A.,Kelly,C.,Quigley,T.,andFogarty,W.(1998)Mechanismsofactionof5themaltogenicα-amylaseofByssochlamysfulva.EnzymeMicrobTechnol22:612–6616.7
VanDyken,J.D.,Müller,M.J.I.,Mack,K.M.L.,andDesai,M.M.(2013)Spatialpopulation8expansionpromotestheevolutionofcooperationinanexperimentalPrisoner’s9Dilemma.CurrBiol23:919–923.10
El-Fallal,A.,Abou,M.,El-Sayed,A.,andOmar,N.(2012)StarchandMicrobialα-Amylases:11FromConceptstoBiotechnologicalApplications.In,Chang,C.-F.(ed),Carbohydrates-12ComprehensiveStudiesonGlycobiologyandGlycotechnology.InTech.13
Enfors,S.O.,Jahic,M.,Rozkov,A.,Xu,B.,Hecker,M.,Jürgen,B.,etal.(2001)Physiological14responsestomixinginlargescalebioreactors.JBiotechnol85:175–185.15
Eriksson,A.andLindgren,K.(2005)Cooperationdrivenbymutationsinmulti-person16Prisoner’sDilemma.JTheorBiol232:399–409.17
Fiegna,F.andVelicer,G.J.(2003)Competitivefatesofbacterialsocialparasites:persistence18andself-inducedextinctionofMyxococcusxanthuscheaters.ProcBiolSci270:1527–191534.20
Fiore,C.L.,Longnecker,K.,KidoSoule,M.C.,andKujawinski,E.B.(2015)Releaseof21ecologicallyrelevantmetabolitesbythecyanobacteriumSynechococcuselongatus22CCMP1631.EnvironMicrobiol17:3949–3963.23
Flamholz,A.,Noor,E.,Bar-Even,A.,Liebermeister,W.,andMilo,R.(2013)Glycolytic24strategyasatradeoffbetweenenergyyieldandproteincost.ProcNatlAcadSciUS25A110:10039–10044.26
Franklin,F.C.,Bagdasarian,M.,Bagdasarian,M.M.,andTimmis,K.N.(1981)Molecularand27functionalanalysisoftheTOLplasmidpWWOfromPseudomonasputidaandcloning28ofgenesfortheentireregulatedaromaticringmetacleavagepathway.ProcNatl29AcadSciUSA78:7458–7462.30
Frey,E.(2010)Evolutionarygametheory:Theoreticalconceptsandapplicationstomicrobial31communities.PhysicaA:StatisticalMechanicsanditsApplications389:4265–4298.32
Gallego,J.L.R.,García-Martínez,M.J.,Llamas,J.F.,Belloch,C.,Peláez,A.I.,andSánchez,J.33(2007)Biodegradationofoiltankbottomsludgeusingmicrobialconsortia.34

Cooperationinmicrobialbiotechnology
26
Biodegradation18:269–281.1
González-Cabaleiro,R.,Lema,J.M.,Rodríguez,J.,andKleerebezem,R.(2013)Linking2thermodynamicsandkineticstoassesspathwayreversibilityinanaerobic3bioprocesses.EnergyEnviron.Sci.6:3780.4
González-Cabaleiro,R.,Ofiţeru,I.D.,Lema,J.M.,andRodríguez,J.(2015)Microbialcatabolic5activitiesarenaturallyselectedbymetabolicenergyharvestrate.ISMEJ9:2630–62641.7
Gore,J.,Youk,H.,andvanOudenaarden,A.(2009)Snowdriftgamedynamicsandfacultative8cheatinginyeast.Nature459:253–256.9
Großkopf,T.,Consuegra,J.,Gaffé,J.,Willison,J.C.,Lenski,R.E.,Soyer,O.S.,andSchneider,10D.(2016)Metabolicmodellinginadynamicevolutionaryframeworkpredicts11adaptivediversificationofbacteriainalong-termevolutionexperiment.BMCEvol12Biol16:163.13
Großkopf,T.andSoyer,O.S.(2016)Microbialdiversityarisingfromthermodynamic14constraints.ISMEJ.15
Gross,R.A.andKalra,B.(2002)Biodegradablepolymersfortheenvironment.Science297:16803–807.17
Harcombe,W.R.,Riehl,W.J.,Dukovski,I.,Granger,B.R.,Betts,A.,Lang,A.H.,etal.(2014)18Metabolicresourceallocationinindividualmicrobesdeterminesecosystem19interactionsandspatialdynamics.CellRep7:1104–1115.20
Harrington,K.I.andSanchez,A.(2014)Eco-evolutionarydynamicsofcomplexsocial21strategiesinmicrobialcommunities.CommunIntegrBiol7:e28230.22
Hauert,C.,Wakano,J.Y.,andDoebeli,M.(2008)Ecologicalpublicgoodsgames:cooperation23andbifurcation.TheorPopulBiol73:257–263.24
vanHoek,M.J.A.andMerks,R.M.H.(2012)Redoxbalanceiskeytoexplainingfullvs.partial25switchingtolow-yieldmetabolism.BMCSystBiol6:22.26
Hoh,C.Y.andCord-Ruwisch,R.(1996)Apracticalkineticmodelthatconsidersendproduct27inhibitioninanaerobicdigestionprocessesbyincludingtheequilibriumconstant.28BiotechnolBioeng51:597–604.29
James,G.A.,Beaudette,L.,andCosterton,J.W.(1995)Interspeciesbacterialinteractionsin30biofilms.JIndMicrobiol15:257–262.31
Jin,Q.andBethke,C.M.(2007)Thethermodynamicsandkineticsofmicrobialmetabolism.32AmJSci307:643–677.33

Cooperationinmicrobialbiotechnology
27
Karpinets,T.V.,Pelletier,D.A.,Pan,C.,Uberbacher,E.C.,Melnichenko,G.V.,Hettich,R.L.,1andSamatova,N.F.(2009)Phenotypefingerprintingsuggeststheinvolvementof2single-genotypeconsortiaindegradationofaromaticcompoundsby3Rhodopseudomonaspalustris.PLoSONE4:e4615.4
Kim,W.,Levy,S.B.,andFoster,K.R.(2016)Rapidradiationinbacterialeadstoadivisionof5labour.NatCommun7:10508.6
Knoll,G.andWinter,J.(1989)Degradationofphenolviacarboxylationtobenzoatebya7defined,obligatesyntrophicconsortiumofanaerobicbacteria.ApplMicrobiol8Biotechnol30.:9
Kümmerli,R.,Jiricny,N.,Clarke,L.S.,West,S.A.,andGriffin,A.S.(2009)Phenotypicplasticity10ofacooperativebehaviourinbacteria.JEvolBiol22:589–598.11
Lennon,J.T.andDenef,V.J.(2015)Evolutionaryecologyofmicroorganisms:Fromthetamed12tothewild.In,Pillai,S.D.,Nakatsu,C.H.,Miller,R.V.,andYates,M.V.(eds),Manualof13EnvironmentalMicrobiology,4thEdition.AmericanSocietyofMicrobiology,pp.144.1.2–1–4.1.2–12.15
Lilja,E.E.andJohnson,D.R.(2016)Segregatingmetabolicprocessesintodifferentmicrobial16cellsacceleratestheconsumptionofinhibitorysubstrates.ISMEJ10:1568–1578.17
Lindemann,S.R.,Bernstein,H.C.,Song,H.-S.,Fredrickson,J.K.,Fields,M.W.,Shou,W.,etal.18(2016)Engineeringmicrobialconsortiaforcontrollableoutputs.ISMEJ10:2077–192084.20
Liu,J.,Prindle,A.,Humphries,J.,Gabalda-Sagarra,M.,Asally,M.,Lee,D.D.,etal.(2015)21Metabolicco-dependencegivesrisetocollectiveoscillationswithinbiofilms.Nature22523:550–554.23
Louca,S.andDoebeli,M.(2015)Calibrationandanalysisofgenome-basedmodelsfor24microbialecology.elife4:e08208.25
Majewski,R.A.andDomach,M.M.(1990)Simpleconstrained-optimizationviewofacetate26overflowinE.coli.BiotechnolBioeng35:732–738.27
Martínez,I.,Mohamed,M.E.-S.,Rozas,D.,García,J.L.,andDíaz,E.(2016)Engineering28syntheticbacterialconsortiaforenhanceddesulfurizationandrevalorizationofoil29sulfurcompounds.MetabEng35:46–54.30
McGenity,T.J.,Folwell,B.D.,McKew,B.A.,andSanni,G.O.(2012)Marinecrude-oil31biodegradation:acentralroleforinterspeciesinteractions.AquatBiosyst8:10.32
McGinty,S.É.,Lehmann,L.,Brown,S.P.,andRankin,D.J.(2013)Theinterplaybetween33relatednessandhorizontalgenetransferdrivestheevolutionofplasmid-carried34

Cooperationinmicrobialbiotechnology
28
publicgoods.ProcBiolSci280:20130400.1
McInerney,M.J.andBryant,M.P.(1981)Anaerobicdegradationoflactatebysyntrophic2associationsofMethanosarcinabarkeriandDesulfovibriospeciesandeffectofH2on3AcetateDegradation.ApplEnvironMicrobiol41:346–354.4
Minty,J.J.,Singer,M.E.,Scholz,S.A.,Bae,C.-H.,Ahn,J.-H.,Foster,C.E.,etal.(2013)Design5andcharacterizationofsyntheticfungal-bacterialconsortiafordirectproductionof6isobutanolfromcellulosicbiomass.ProcNatlAcadSciUSA110:14592–14597.7
Molenaar,D.,vanBerlo,R.,deRidder,D.,andTeusink,B.(2009)Shiftsingrowthstrategies8reflecttradeoffsincellulareconomics.MolSystBiol5:323.9
Moreno-Fenoll,C.,Cavaliere,M.,Martínez-García,E.,andPoyatos,J.F.(2017)Eco-10evolutionaryfeedbackscanrescuecooperationinmicrobialpopulations.SciRep7:1142561.12
Morris,B.E.L.,Henneberger,R.,Huber,H.,andMoissl-Eichinger,C.(2013)Microbial13syntrophy:interactionforthecommongood.FEMSMicrobiolRev37:384–406.14
Müller,S.,Harms,H.,andBley,T.(2010)Originandanalysisofmicrobialpopulation15heterogeneityinbioprocesses.CurrOpinBiotechnol21:100–113.16
Nadell,C.D.,Drescher,K.,andFoster,K.R.(2016)Spatialstructure,cooperationand17competitioninbiofilms.NatRevMicrobiol14:589–600.18
Nadell,C.D.,Xavier,J.B.,andFoster,K.R.(2009)Thesociobiologyofbiofilms.FEMS19MicrobiolRev33:206–224.20
Niehus,R.,Mitri,S.,Fletcher,A.G.,andFoster,K.R.(2015)Migrationandhorizontalgene21transferdividemicrobialgenomesintomultipleniches.NatCommun6:8924.22
Nikel,P.I.,Silva-Rocha,R.,Benedetti,I.,anddeLorenzo,V.(2014)Theprivatelifeof23environmentalbacteria:pollutantbiodegradationatthesinglecelllevel.Environ24Microbiol16:628–642.25
Nogueira,T.,Rankin,D.J.,Touchon,M.,Taddei,F.,Brown,S.P.,andRocha,E.P.C.(2009)26Horizontalgenetransferofthesecretomedrivestheevolutionofbacterial27cooperationandvirulence.CurrBiol19:1683–1691.28
Nowak,M.A.(2006)Fiverulesfortheevolutionofcooperation.Science314:1560–1563.29
Nowak,M.A.andSigmund,K.(2004)Evolutionarydynamicsofbiologicalgames.Science30303:793–799.31
Paczia,N.,Nilgen,A.,Lehmann,T.,Gätgens,J.,Wiechert,W.,andNoack,S.(2012)Extensive32exometabolomeanalysisrevealsextendedoverflowmetabolisminvarious33

Cooperationinmicrobialbiotechnology
29
microorganisms.MicrobCellFact11:122.1
Pande,S.,Kaftan,F.,Lang,S.,Svatoš,A.,Germerodt,S.,andKost,C.(2016)Privatizationof2cooperativebenefitsstabilizesmutualisticcross-feedinginteractionsinspatially3structuredenvironments.ISMEJ10:1413–1423.4
Pennisi,E.(2009)Origins.Ontheoriginofcooperation.Science325:1196–1199.5
Pfeiffer,T.andBonhoeffer,S.(2004)Evolutionofcross-feedinginmicrobialpopulations.Am6Nat163:E126–35.7
Popat,R.,Cornforth,D.M.,McNally,L.,andBrown,S.P.(2015)Collectivesensingand8collectiveresponsesinquorum-sensingbacteria.JRSocInterface12.:9
Rankin,D.J.,Rocha,E.P.C.,andBrown,S.P.(2011)Whattraitsarecarriedonmobilegenetic10elements,andwhy?Heredity106:1–10.11
Ratzke,C.andGore,J.(2016)Self-organizedpatchinessfacilitatessurvivalinacooperatively12growingBacillussubtilispopulation.Naturemicrobiology1:16022.13
Regot,S.,Macia,J.,Conde,N.,Furukawa,K.,Kjellén,J.,Peeters,T.,etal.(2011)Distributed14biologicalcomputationwithmulticellularengineerednetworks.Nature469:207–15211.16
Richards,T.A.andTalbot,N.J.(2013)Horizontalgenetransferinosmotrophs:playingwith17publicgoods.NatRevMicrobiol11:720–727.18
Ross-Gillespie,A.,Gardner,A.,Buckling,A.,West,S.A.,andGriffin,A.S.(2009)Density19dependenceandcooperation:theoryandatestwithbacteria.Evolution63:2315–202325.21
Sanchez,A.andGore,J.(2013)Feedbackbetweenpopulationandevolutionarydynamics22determinesthefateofsocialmicrobialpopulations.PLoSBiol11:e1001547.23
Sayler,G.S.andRipp,S.(2000)Fieldapplicationsofgeneticallyengineeredmicroorganisms24forbioremediationprocesses.CurrOpinBiotechnol11:286–289.25
Schink,B.(1997)Energeticsofsyntrophiccooperationinmethanogenicdegradation.26MicrobiolMolBiolRev61:262–280.27
Schoener,T.W.(2011)Thenewestsynthesis:understandingtheinterplayofevolutionary28andecologicaldynamics.Science331:426–429.29
Scholten,J.C.andConrad,R.(2000)Energeticsofsyntrophicpropionateoxidationindefined30batchandchemostatcocultures.ApplEnvironMicrobiol66:2934–2942.31
Seitz,H.J.,Schink,B.,andConrad,R.(1988)Thermodynamicsofhydrogenmetabolismin32

Cooperationinmicrobialbiotechnology
30
methanogeniccoculturesdegradingethanolorlactate.FEMSMicrobiolLett55:119–1124.2
Silva-Rocha,R.anddeLorenzo,V.(2012)StochasticityofTOLplasmidcatabolicpromoters3setsabimodalexpressionregimeinPseudomonasputidamt-2exposedtom-xylene.4MolMicrobiol86:199–211.5
Sözen,S.,Çokgör,E.U.,Başaran,S.T.,Aysel,M.,Akarsubaşı,A.,Ergal,I.,etal.(2014)Effectof6highloadingonsubstrateutilizationkineticsandmicrobialcommunitystructurein7superfastsubmergedmembranebioreactor.BioresourTechnol159:118–127.8
Spencer,C.C.,Bertrand,M.,Travisano,M.,andDoebeli,M.(2007)Adaptivediversificationin9genesthatregulateresourceuseinEscherichiacoli.PLoSGenet3:e15.10
Travisano,M.andVelicer,G.J.(2004)Strategiesofmicrobialcheatercontrol.Trends11Microbiol12:72–78.12
Velicer,G.J.andVos,M.(2009)SociobiologyoftheMyxobacteria.AnnuRevMicrobiol63:13599–623.14
Vemuri,G.N.,Altman,E.,Sangurdekar,D.P.,Khodursky,A.B.,andEiteman,M.A.(2006)15OverflowmetabolisminEscherichiacoliduringsteady-stategrowth:transcriptional16regulationandeffectoftheredoxratio.ApplEnvironMicrobiol72:3653–3661.17
Westerholm,M.,Dolfing,J.,Sherry,A.,Gray,N.D.,Head,I.M.,andSchnürer,A.(2011)18Quantificationofsyntrophicacetate-oxidizingmicrobialcommunitiesinbiogas19processes.EnvironMicrobiolRep3:500–505.20
West,S.A.,Diggle,S.P.,Buckling,A.,Gardner,A.,andGriffin,A.S.(2007)Thesociallivesof21microbes.Annu.Rev.Ecol.Evol.Syst.38:53–77.22
West,S.A.,Griffin,A.S.,Gardner,A.,andDiggle,S.P.(2006)Socialevolutiontheoryfor23microorganisms.NatRevMicrobiol4:597–607.24
Widder,S.,Allen,R.J.,Pfeiffer,T.,Curtis,T.P.,Wiuf,C.,Sloan,W.T.,etal.(2016)Challenges25inmicrobialecology:buildingpredictiveunderstandingofcommunityfunctionand26dynamics.ISMEJ10:2557–2568.27
Wierckx,N.,Prieto,M.A.,Pomposiello,P.,deLorenzo,V.,O’Connor,K.,andBlank,L.M.28(2015)Plasticwasteasanovelsubstrateforindustrialbiotechnology.Microb29Biotechnol8:900–903.30
Willsey,G.G.andWargo,M.J.(2015)Extracellularlipaseandproteaseproductionfroma31modeldrinkingwaterbacterialcommunityisfunctionallyrobusttoabsenceof32individualmembers.PLoSONE10:e0143617.33

Cooperationinmicrobialbiotechnology
31
Wolfaardt,G.M.,Lawrence,J.R.,Robarts,R.D.,andCaldwell,D.E.(1994)Theroleof1interactions,sessilegrowthandnutrientamendmentsonthedegradativeefficiency2ofamicrobialconsortium.CanJMicrobiol40:331–340.3
Yoshida,S.,Hiraga,K.,Takehana,T.,Taniguchi,I.,Yamaji,H.,Maeda,Y.,etal.(2016)A4bacteriumthatdegradesandassimilatespoly(ethyleneterephthalate).Science351:51196–1199.6
Zamocky,M.,Ludwig,R.,Peterbauer,C.,Hallberg,B.M.,Divne,C.,Nicholls,P.,andHaltrich,7D.(2006)Cellobiosedehydrogenase--aflavocytochromefromwood-degrading,8phytopathogenicandsaprotropicfungi.CurrProteinPeptSci7:255–280.9
Zhou,K.,Qiao,K.,Edgar,S.,andStephanopoulos,G.(2015)Distributingametabolic10pathwayamongamicrobialconsortiumenhancesproductionofnaturalproducts.11NatBiotechnol33:377–383.12
Zhuang,K.,Vemuri,G.N.,andMahadevan,R.(2011)Economicsofmembraneoccupancy13andrespiro-fermentation.MolSystBiol7:500.14
Zomorrodi,A.R.andSegrè,D.(2016)Syntheticecologyofmicrobes:Mathematicalmodels15andapplications.JMolBiol428:837–861.16
17