department - applied and environmental microbiologyaem.asm.org/content/57/4/987.full.pdf · poona,...
TRANSCRIPT

Vol. 57, No. 4APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1991, p. 987-9920099-2240/91/040987-06$02.00/0Copyright © 1991, American Society for Microbiology
Production, Purification, and Characterization of 1-(1-4)-Endoxylanase of Streptomyces roseiscleroticus
ANTHONY C. GRABSKIt AND THOMAS W. JEFFRIES*Institute for Microbial and Biochemical Technology, Forest Products Laboratory, Forest Service,
U.S. Department of Agriculture, Madison, Wisconsin 53705-2398
Received 7 December 1990/Accepted 25 January 1991
Twelve species of Streptomyces that formerly belonged to the genus Chainia were screened for the productionof xylanase and cellulase. One species, Streptomyces roseiscleroticus (Chainia rosea) NRRL B-11019, producedup to 16.2 IU of xylanase per ml in 48 h. A xylanase from S. roseiscleroticus was purified and characterized.The enzyme was a debranching j0-(1-4)-endoxylanase showing high activity on xylan but essentially no activityagainst acid-swollen (Walseth) cellulose. It had a very low apparent molecular weight of 5,500 by native gelfiltration, but its denatured molecular weight was 22,600 by sodium dodecyl sulfate-polyacrylamide gelelectrophoresis. It had an isoelectric point of 9.5. The pH and temperature optima for hydrolysis ofarabinoxylan were 6.5 to 7.0 and 60°C, respectively, and more than 75% of the optimum enzyme activity wasretained at pH 8.0. The xylanase had a Km of 7.9 mg/ml and an apparent Vmax of 305 ,umol min- mg ofprotein-'. The hydrolysis rate was linear for xylan concentrations of less than 4 mg/ml, but significantinhibition was observed at xylan concentrations of more than 10 mg/ml. The predominant products ofarabinoxylan hydrolysis included arabinose, xylobiose, and xylotriose.
In recent years there has been an increasing interest inapplying xylanases to pulping processes. Particularly, theyhave been used to facilitate the bleaching of kraft pulps (5,10, 11, 31) or to improve fiber properties (16, 20). Endoglu-canase activity is unwanted in most cases because it de-grades cellulose fibers and destroys pulp properties. How-ever, cellulase is often present in crude extracellular broths,so it must be removed by purification (28) or inhibited (20).Cloned xylanase is highly specific (21), and when it is clonedand overexpressed by a multicopy plasmid vector, very highactivities can be obtained (3).
Streptomyces spp., being prevalent in soils and decom-posing vegetation, have been screened for xylanase (2, 8, 9).Species that have been identified as exceptional xylanaseproducers include Streptomyces lividans (13, 17), "Strepto-myces xylophagus" (12), and Chainia species (26), all ofwhich have been reclassified as Streptomyces species (7).The objective of this research was to screen a number of
former Chainia species as well as several other Streptomy-ces spp. for high xylanase titers with little or no cellulaseactivity. Streptomyces roseiscleroticus NRRL B-11019 pro-duced the highest xylanase activity but had very littlecellulase activity and was therefore selected for production,purification, and characterization of its xylanase.
MATERIALS AND METHODS
Microorganisms. The following organisms were obtainedfrom David P. Labeda, U. S. Department of AgricultureNorthern Regional Research Laboratory, Peoria, Ill.: Strep-tomyces sclerotialus (Chainia antibiotica) NRRL B-2317;Streptomyces sp. strain NRRL B-11017 (Chainia aurea);Streptomyces flaviscleroticus (Chainia flava) NRRLB-11018; Streptomyces fumigatiscleroticus (Chainia fumi-
* Corresponding author.t Present address: Protein Purification Facility, University of
Wisconsin Biotechnology Center, Madison, WI 53705-4087.
gata) NRRL B-3856; Streptomyces kunmingensis (Chainiakunmingensis) NRRL B-16240; Streptomyces niger (Chainianigra) NRRL B-3857; Streptomyces ochraceiscleroticus(Chainia ochracea) NRRL B-3041; Streptomyces olivacei-scleroticus (Chainia olivacea) NRRL B-2318; Streptomycespoonensis (Chainia poonensis) NRRL B-2319; Streptomycespurpurogeneiscleroticus (Chainia purpurogenea) NRRLB-12047; S. roseiscleroticus (Chainia rosea) NRRL B-11019;Streptomyces violens (Chainia violens) NRRL B-3484; S.lividans NRRL B-16148 (=John Innes Institute 1326);"Streptomyces xylophagus" NRRL B-12029 (=IFO 13845).Micromonospora echinospora subsp. ferruginea was ob-tained from Mala Rao at the National Chemical Laboratory,Poona, India.Growth media. Yeast extract-malt extract-xylose (YMX)
agar consisted of the following (in grams per liter): yeastextract, 4.0; malt extract, 10.0; xylose, 4.0; and agar, 15.0.The pH was adjusted to 7.3 prior to autoclaving. Definedxylan (DX) agar consisted of the following (in grams perliter): oat spelt xylan (Sigma Chemical Co., St. Louis, Mo.),10.0; K2HPO4, 1.0; NaCl, 1.0; (NH4)2SO4, 2.0; CaCO3, 2.0;and agar, 15. The pH was 7.0; the medium was sterilized byautoclaving. Basic formulations for these media were sug-gested by D. P. Labeda; we substituted xylose for glucose inYMX and xylan for starch in DX. Trypticase soy broth(TSB; Difco) was prepared according to the manufacturer'sinstructions. Xylanase production medium was formulatedaccording to the method of Morosoli et al. (17), except thata more-complete trace element solution was employed (29,33).
Cultivation. Lyophilized cultures were rehydrated in ster-ile YMX medium without agar. For stock cultivation, YMXagar was used. To obtain better sporulation, DX agar wasemployed. It was essential to subculture the organismsfrequently (every 2 weeks) to maintain viability. Stockinocula were preserved at an early stage by cutting colonizedplugs of agar from 1-week-old YMX agar plates and freezingthem at -70°C in 10% (wt/vol) sterile glycerol (4). In order to
987
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

988 GRABSKI AND JEFFRIES
obtain maximal xylanase production, it was necessary toprime the cultures in TSB as described by Morosoli et al.(17).
Seven-day-old (or sporulating) slants of each organismgrown on DX agar were washed with sterile TSB, and 10 mlof culture was used to inoculate 100 ml of TSB in 500-mlErlenmeyer flasks stoppered with cotton plugs. Cultureswere incubated at 37 to 38°C with shaking at 240 rpm for 24h. For screening, 10 ml of each culture was then used toinoculate 100 ml of xylanase production medium in a 500-mlErlenmeyer flask. All other cultivation conditions remainedunchanged. S. roseiscleroticus was scaled up for enzymepurification and characterization. Cultures were primed inthe same manner as before, but the full 110-ml inoculum wasadded to 500 ml of xylanase production medium in a 2,800-mlFernbach flask. Cultures were incubated at 37 to 38°C withshaking at 240 rpm for 48 h. Duplicate cultures were used forxylanase screenings and production, and the averages oftiters are reported here.
Assays. Reducing-sugar quantitations were routinely per-formed by Nelson's modification (19) of the Somogyi method(25) (NS method) using either D-xylose or D-glucose as astandard. For comparative purposes, the dinitrosalicylicacid (DNS) method was employed (15).
Samples (10 ml) were aseptically taken every 24 h, andcells were removed by centrifugation (10,000 x g, 15 min).Supernatant solutions were decanted and assayed for xyla-nase activity. Cellulase activity was assayed only at peakxylanase time points. Xylanase assays employed 0.25 ml of1% oat spelts xylan plus 0.25 ml of appropriately dilutedenzyme in 50 mM sodium phosphate buffer (pH 7.0; bufferA). Xylan was solubilized in 0.5 N NaOH, neutralized with1.0 N HCI, and diluted to 1% with buffer A. Reactions werestarted by the addition of substrate and incubated for 10 minat 60°C. Reactions were stopped by the addition of the NSmethod reagent C.
Cellulase assays were performed in the same manner asxylanase assays by replacing 1% xylan with 1% acid-swollencellulose (32) prepared from Solka-floc SW-40 wood pulpcellulose (Brown Co., Berlin, N.H.) as the substrate. Thecellulase assay temperature was reduced to 55°C (17).
Unless otherwise stated, all references to internationalunits (e.g., 1 Rmol/min) use the NS method to quantitatereducing sugars.Enzyme purification. Cells were harvested by centrifuga-
tion (11,000 x g, 30 min). The dark red supernatant solutionwas filtered (Whatman no. 41 filter), and 500 ml of filtratewas concentrated by ultrafiltration to approximately 100 ml(Amicon YM-5; 5,000-molecular-weight cutoff). Proteins inthe crude concentrate were precipitated by using (NH4)2SO4saturation at 0°C. The 0 to 30% saturation precipitate wasremoved by centrifugation (11,000 x g, 30 min) and dis-carded. The supernatant solution was precipitated again with30 to 50% (NH4)2SO4 saturation. The pellet was resuspendedin 50 mM sodium acetate buffer (pH 4.0; buffer B) and eitherdialyzed or diafiltered against the same buffer for approxi-mately 12 h. Dialysis employed standard cellulose-baseddialysis tubing having a nominal molecular weight cutoff of10,000 (Viscase Corp., Chicago, Ill.). Diafiltration employedAmicon YM-5 polymeric membranes with nominal molecu-lar weight cutoffs of .5,000. The characterization and actionpattern studies reported here were performed with thedialyzed preparation. The dialyzed sample was centrifuged,and the supernatant was purified by cation-exchange fastprotein liquid chromatography (FPLC). Supernatant wasapplied to a Pharmacia C 10/10 column (Piscataway, N.J.)
packed with carboxymethyl Bio-Gel A cation-exchangeresin (Bio-Rad, Richmond, Calif.) coupled to a PharmaciaFPLC. The flow rate was 0.50 ml/min; 0.5-ml fractions werecollected. Unbound protein was washed from the columnwith buffer B. Bound protein was eluted by using a linear50-ml gradient of 0 to 0.4 M NaCl in buffer B. Elution wasmonitored for protein at 280 nm. Fractions containing thexylanase activity were dialyzed against buffer B for approx-imately 5 h. The dialyzed sample was applied to a PharmaciaMono-S cation-exchange column and chromatographed asdescribed above. The purified xylanase was stored at -70°Cin buffer A.
Gel filtration. Apparent molecular weights of undenaturedxylanases were determined by gel filtration using a Superose12 column coupled to a Pharmacia FPLC. Proteins were
eluted isocratically with buffer A, and elution was monitoredat 280 nm. The following Sigma molecular weight markerswere used: glucose oxidase (150,000), bovine albumin(66,000), egg albumin (45,000), trypsinogen, (24,000), andvitamin B12 (1,350). Blue dextran (average molecular weight,2,000,000) was used to measure the column void volume(VO).
Electrophoresis. Purity and apparent molecular weights ofdenatured xylanases were examined by using a PharmaciaPhast system. A 10 to 15% gradient polyacrylamide gel wasrun and stained with Coomassie brilliant blue according tothe manufacturer's instructions. Sigma sodium dodecyl sul-fate (SDS) molecular weight markers were the follow-ing: lysozyme (14,300), trypsinogen (24,000), egg albumin(45,000), and bovine albumin (66,000).
pl determination. Isoelectric point determination was byanalytical isoelectric focusing using Serva (Westbury, N.Y.)Precote gel (pH 3 to 10) and the Serva violet stainingprocedure. Serva protein test mixture 9 contained the fol-lowing pH marker proteins: amyloglucosidase (3.5), ferritin(4.4), albumin bovine (4.7), P-lactoglobulin (5.34), conalbu-min (5.9), horse myoglobin (7.3), whale myoglobin (8.3),RNase (9.45), and cytochrome c (10.65).pH effect. Enzyme was incubated for 20 min at room
temperature (21°C) in the following buffers: pH 4.0 to 5.5,sodium acetate; pH 6 to 8, sodium phosphate. All bufferswere 50 mM. The enzyme was then assayed for xylanaseactivity in the same buffer.Temperature effect. Enzyme and substrate were equili-
brated separately for 5 min at each test temperature and thencombined and assayed for xylanase activity at the same testtemperature.Km and Vmax. The Km and Vmax were determined by NS
method assay at pH 7.0 and 60°C with 0.10 ,ug of purifiedxylanase in each assay and an increasing substrate concen-tration of 0.10 to 16.0 mg of oat spelts xylan per ml. Datawere plotted by the method of Lineweaver-Burk.
Action pattern. Hydrolysis products were determined byincubating a 1.0-ml mixture in a shaking water bath at 38°C.The hydrolysis mixture contained 25 mg of xylan, buffer A,and enzyme (-5 ,ug of purified enzyme or -20 ,ug of crudeextract). Hydrolysates were centrifuged (16,000 x g, 5 min),and the supernatant was examined by paper chromatographyusing a butanol-pyridine-water (6:4:3) solvent system andsilver nitrate staining (30). Authentic xylooligosaccharidestandards xylobiose and xylotriose were obtained from R. B.Hespell, U.S. Department of Agriculture Northern RegionalResearch Laboratory. D-Xylose and L-arabinose standardswere obtained from Sigma.
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

XYLANASE OF S. ROSEISCLEROTICUS 989
TABLE 1. Comparison of peak xylanase and cellulase activities from the 15 organisms screened
Enzymatic activity(IU/ml)"cXylanase/ Production ratebStrain cellulase ( h
Xylanase Cellulase ratio (lU/h)
S. sclerotialus NRRL B-2317 8.4 0.12 70 0.23Streptomyces sp. strain NRRL B-11017 1.4 0.0 0.07S. flaviscleroticus NRRL B-11018 8.2 0.10 82 0.27S. fumigatiscleroticus NRRL B-3856 8.6 0.02 430 0.04S. kunmingensis NRRL B-16240 3.7 0.03 123 0.03S. niger NRRL B-3857 8.4 0.03 280 0.35S. ochraceiscleroticus NRRL B-3041 4.2 0.02 210 0.14S. olivaceiscleroticus NRRL B-2318 4.6 0.02 230 0.19S. poonensis NRRL B-2319 5.4 0.16 34 <0.01S. purpurogeneiscleroticus NRRL B-12047 1.5 0.01 150 0.03S. roseiscleroticus NRRL B-11019 16.2 0.21 77 0.57S. violens NRRL B-3484 3.8 0.01 38 0.11S. lividans NRRL B-16148 (1326) 3.1 0.04 78 0.01"S. xylophagus" NRRL B-12029 6.4 0.03 213 0.08M. echinospora subsp. ferruginea 4.5 0.03 150 0.16
a Peak xylanase activities reached at 48 h with the exceptions of NRRL B-11017 and NRRL B-3857 (24 h), NRRL B-2318 and NRRL B-12029 (72 h), NRRLB-3041 (94 h), and NRRL B-2319 and NRRL B-12047 (96 h).
b Xylanase production rate determined at 24 h.
RESULTS AND DISCUSSION
Screening. Table 1 summarizes the maximum xylanaseactivities obtained with each strain, cellulase activities de-termined at that time point, ratios of xylanase to cellulase,and rate of xylanase production. The ratios of xylanase tocellulase activity in the crude broth ranged from 34 (S.poonensis) to 430 (S. fumigatiscleroticus). One strain did notproduce detectable cellulase activity (Streptomyces sp.
strain NRRL B-11017), but it also formed very little xyla-nase. S. roseiscleroticus formed 16.2 IU of xylanase per mlafter 48 h and had a rapid rate of enzyme production (0.57IU/h). The screening experiments showed that four Strepto-myces strains produced more than 8 IU of xylanase per mland had significantly higher titers than "S. xylophagus," M.echinospora, or the other Streptomyces species screened forxylanase activity.
S. lividans NRRL B-16148 formed considerably less xyla-nase than previously reported by Kluepfel et al. (13). Weemployed a derivative of the S. lividans strain, used essen-
tially the same medium (with the exception of the traceelement solution), and followed their protocol for inoculumpreparation. Our enzyme assay employed the same sub-strate, but where they used the DNS method to determinereducing sugars, we routinely used the NS method. Inseparate studies, we have found that depending on the stageof substrate hydrolysis, the DNS method gives 3- to 10-fold-higher values than the NS method when used to assay forxylanase activity. We have assayed the clarified crude brothfrom S. roseiscleroticus by both methods. Using oat speltsxylan as substrate, the NS method gave 11.7 IU/ml, whereasthe DNS method gave 35 IU/ml. Similar discrepancies havebeen reported for the determination of the chain lengths ofmaltodextrins (23) and for the determination of xylanaseactivity (24). The DNS reagent is apparently capable ofhydrolyzing the glycosidic linkages of maltodextrins andxylan and hence results in more reducing groups than are
observed with the NS reagent. This factor accounts for mostof the difference between our findings and those previouslyreported for xylanase activity in S. lividans.
Purification. Following the results of the initial screen, we
scaled up production and purified the xylanase from S.roseiscleroticus NRRL B-11019. However, S. fumigatiscle-
roticus might be of future interest because of its highxylanase-to-cellulase ratio. The results of the initial purifi-cation are shown in Table 2.
Cultivation of S. roseiscleroticus on a larger scale resultedin slightly lower xylanase activity (13.3 versus 16.2 IU/ml).This decrease could be due to decreased oxygenation orvariability in the primer inoculum. The first step of thepurification concentrated the proteins to facilitate ammo-nium sulfate precipitation. Concentration increased the totalactivity by 13%. However, concentration was slow becauseof fouling of the ultrafiltration membrane by red pigments.The red pigments must be removed prior to the Mono-Schromatography step in order to prevent possible damage tothe column.Ammonium sulfate precipitation removed most of the red
pigments and resulted in an 11-fold increase in purity.However, this precipitation step was extremely inefficient.Low yield can be attributed to xylanase lost in the 20 to 30%saturation ammonium sulfate cut. When 20 to 50% saturationis used instead of 30 to 50% saturation, a recovery of 83%was obtained, but the specific activity was lower, andpigments remained.The final steps in the purification were successive cation-
exchange chromatography columns. Carboxymethyl Bio-Gel A agarose cation exchange removed the remainingpigments and acidic proteins. Xylanase proteins adsorbed tothe column and eluted as a single broad peak. Fractions fromthis peak were pooled, dialyzed, and applied to the Mono-S
TABLE 2. Purification of a ,B-(1-4)-endoxylanase fromS. roseiscleroticus grown on liquid xylan substrate
ActivityTotal protein Fold
Purlficatstep (mg) Total (IU) Specific purificationPurification Total ~(IU)(U/mg)Crude culture filtrate 1,556 6,670 4.3 1.0Concentration 830 7,705 9.3 2.2Ammonium sulfate 53.9 2,552 47.4 11.0Bio-Gel A 3.5 245 70.1 16.3chromatography
Mono-S chromatography 0.1 19.4 194.0 45.1
VOL. 57, 1991
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from
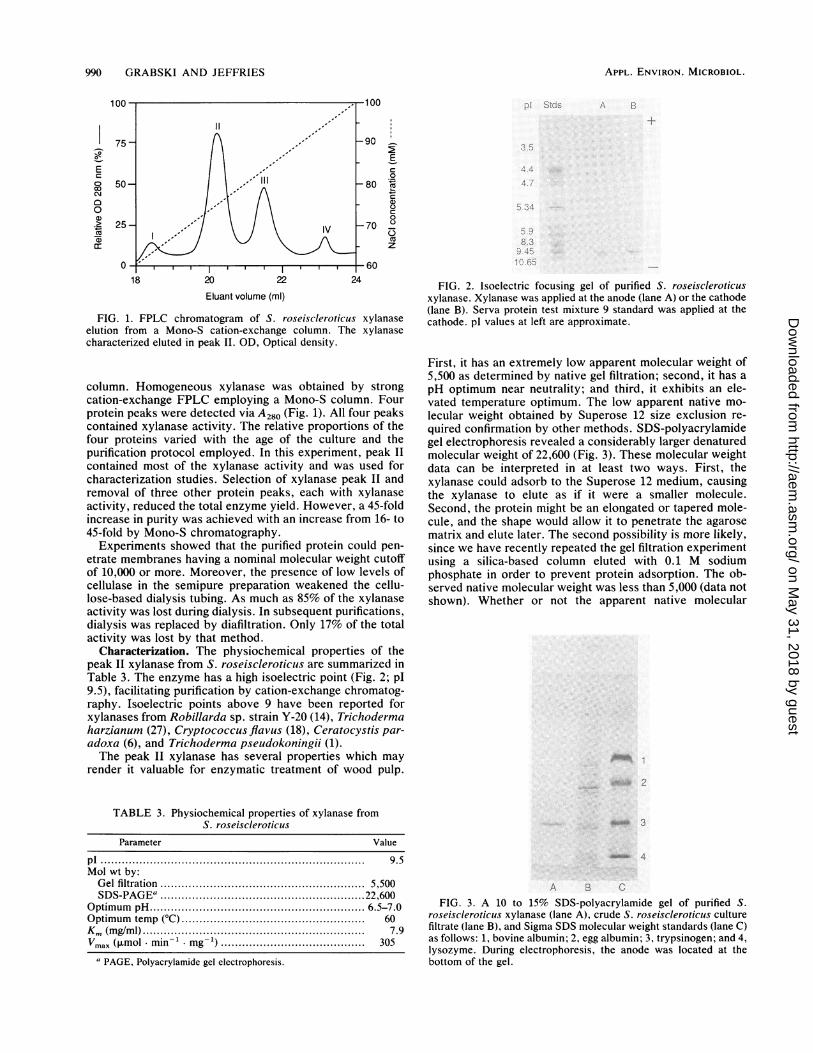
990 GRABSKI AND JEFFRIES
Ec0
coc._00a)
a)
Ec0
co
a0C00
az
Eluant volume (ml)
FIG. 1. FPLC chromatogram of S. roseiscleroticus xylanaseelution from a Mono-S cation-exchange column. The xylanasecharacterized eluted in peak IL. OD, Optical density.
column. Homogeneous xylanase was obtained by strongcation-exchange FPLC employing a Mono-S column. Fourprotein peaks were detected via A280 (Fig. 1). All four peakscontained xylanase activity. The relative proportions of thefour proteins varied with the age of the culture and thepurification protocol employed. In this experiment, peak IIcontained most of the xylanase activity and was used forcharacterization studies. Selection of xylanase peak II andremoval of three other protein peaks, each with xylanaseactivity, reduced the total enzyme yield. However, a 45-foldincrease in purity was achieved with an increase from 16- to45-fold by Mono-S chromatography.
Experiments showed that the purified protein could pen-etrate membranes having a nominal molecular weight cutoffof 10,000 or more. Moreover, the presence of low levels ofcellulase in the semipure preparation weakened the cellu-lose-based dialysis tubing. As much as 85% of the xylanaseactivity was lost during dialysis. In subsequent purifications,dialysis was replaced by diafiltration. Only 17% of the totalactivity was lost by that method.
Characterization. The physiochemical properties of thepeak II xylanase from S. roseiscleroticus are summarized inTable 3. The enzyme has a high isoelectric point (Fig. 2; p19.5), facilitating purification by cation-exchange chromatog-raphy. Isoelectric points above 9 have been reported forxylanases from Robillarda sp. strain Y-20 (14), Trichodermaharzianum (27), Cryptococcus flavus (18), Ceratocystis par-adoxa (6), and Trichoderma pseudokoningii (1).The peak II xylanase has several properties which may
render it valuable for enzymatic treatment of wood pulp.
TABLE 3. Physiochemical properties of xylanase fromS. roseiscleroticus
Parameter Value
pI ...................................................... . . . . . . . 9.5Mol wt by:Gel filtration ................... 5,500SDS-PAGEa ................... 22,600
Optimum pH................... 6.5-7.0Optimum temp (°C) ................... 60K,, (mg/ml)................... 7.9Vmax (,umol min-' mg-') ..... .............. 305
a PAGE, Polyacrylamide gel electrophoresis.
pI Stds A B
3.5
4.4
4.7
5.34
5.9
8.3*
9.45
10.65
FIG. 2. Isoelectric focusing gel of purified S. roseiscieroticus
xylanase. Xylanase was applied at the anode (lane A) or the cathode
(lane B). Serva protein test mixture 9 standard was applied at the
cathode. pl values at left are approximate.
First, it has an extremely low apparent molecular weight of
5,500 as determined by native gel filtration; second, it has a
pH optimum near neutrality; and third, it exhibits an ele-
vated temperature optimum. The low apparent native mo-
lecular weight obtained by Superose 12 size exclusion re-
quired confirmation by other methods. SDS-polyacrylamide
gel electrophoresis revealed a considerably larger denatured
molecular weight of 22,600 (Fig. 3). These molecular weight
data can be interpreted in at least two ways. First, the
xylanase could adsorb to the Superose 12 medium, causing
the xylanase to elute as if it were a smaller molecule.
Second, the protein might be an elongated or tapered mole-
cule, and the shape would allow it to penetrate the agarose
since we have recently repeated the gel filtration experiment
using a silica-based column eluted with 0.1 M sodium
phosphate in order to prevent protein adsorption. The ob-
served native molecular weight was less than 5,000 (data not
shown). Whether or not the apparent native molecularlecuarwighobtinedby uperse 2 sz xlso e
3
A B CFIG. 3. A 10 to 15% SDS-polyacrylamide gel of purified S.
roseiscleroticus xylanase (lane A), crude S. roseiscleroticus culturefiltrate (lane B), and Sigma SDS molecular weight standards (lane C)as follows: 1, bovine albumin; 2, egg albumin; 3, trypsinogen; and 4,lysozyme. During electrophoresis, the anode was located at thebottom of the gel.
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from
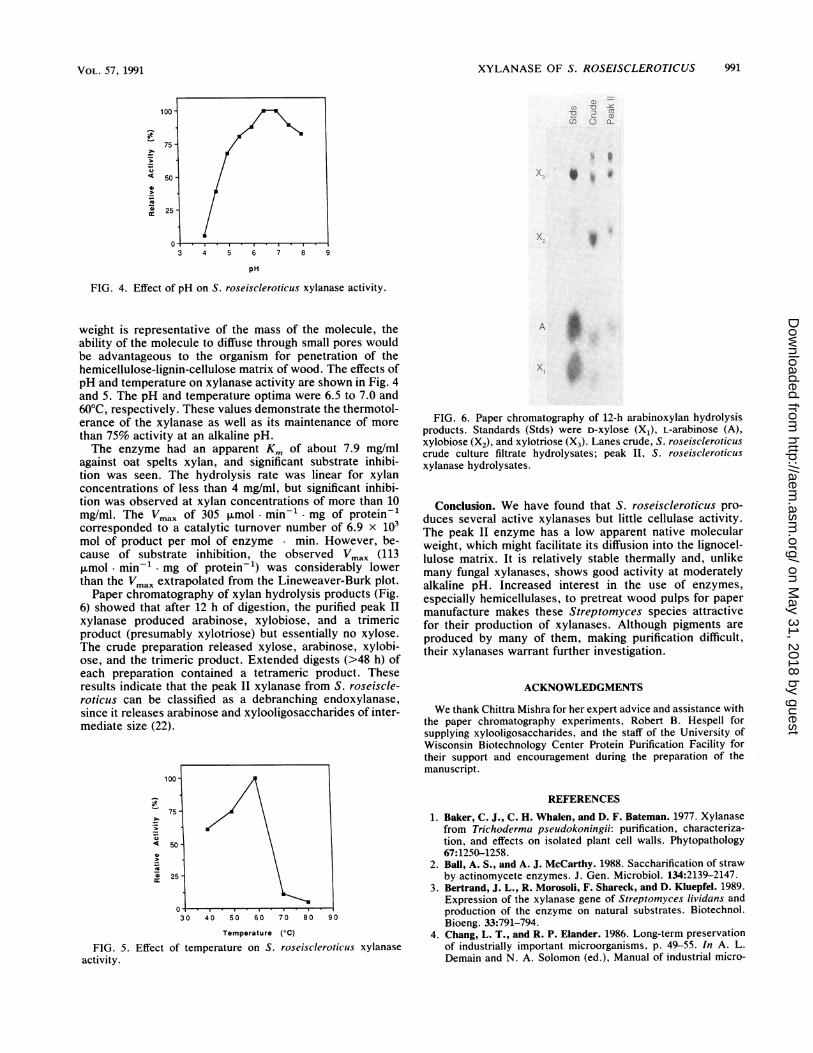
XYLANASE OF S. ROSEISCLEROTICUS 991
q.
75
50
25
0
3 4 5 6 7 8 9
pH
FIG. 4. Effect of pH on S. roseiscleroticus xylanase activity.
weight is representative of the mass of the molecule, theability of the molecule to diffuse through small pores wouldbe advantageous to the organism for penetration of thehemicellulose-lignin-cellulose matrix of wood. The effects ofpH and temperature on xylanase activity are shown in Fig. 4and 5. The pH and temperature optima were 6.5 to 7.0 and60°C, respectively. These values demonstrate the thermotol-erance of the xylanase as well as its maintenance of more
than 75% activity at an alkaline pH.The enzyme had an apparent Km of about 7.9 mg/ml
against oat spelts xylan, and significant substrate inhibi-tion was seen. The hydrolysis rate was linear for xylanconcentrations of less than 4 mg/ml, but significant inhibi-tion was observed at xylan concentrations of more than 10mg/ml. The Vmax of 305 ,umol. min-' mg of protein-'corresponded to a catalytic turnover number of 6.9 x 103mol of product per mol of enzyme min. However, be-cause of substrate inhibition, the observed Vmax (113,umol- min-' mg of protein-') was considerably lowerthan the Vmax extrapolated from the Lineweaver-Burk plot.Paper chromatography of xylan hydrolysis products (Fig.
6) showed that after 12 h of digestion, the purified peak II
xylanase produced arabinose, xylobiose, and a trimericproduct (presumably xylotriose) but essentially no xylose.The crude preparation released xylose, arabinose, xylobi-ose, and the trimeric product. Extended digests (>48 h) ofeach preparation contained a tetrameric product. Theseresults indicate that the peak II xylanase from S. roseiscle-roticus can be classified as a debranching endoxylanase,since it releases arabinose and xylooligosaccharides of inter-mediate size (22).
100 -
--e
21
zc
4)
4)cr
FIG. 5.activity.
30 40 50 60 70 80 90
Temperature (IC)
Effect of temperature on S. roseiscieroticus xylanase
X2
FIG. 6. Paper chromatography of 12-h arabinoxylan hydrolysisproducts. Standards (Stds) were D-xylose (X1), L-arabinose (A),xylobiose (X2), and xylotriose (X3). Lanes crude, S. roseiscieroticuscrude culture filtrate hydrolysates; peak II, S. roseiscieroticusxylanase hydrolysates.
Conclusion. We have found that S. roseiscieroticus pro-duces several active xylanases but little cellulase activity.The peak II enzyme has a low apparent native molecularweight, which might facilitate its diffusion into the lignocel-lulose matrix. It is relatively stable thermally and, unlikemany fungal xylanases, shows good activity at moderatelyalkaline pH. Increased interest in the use of enzymes,especially hemicellulases, to pretreat wood pulps for papermanufacture makes these Streptomyces species attractivefor their production of xylanases. Although pigments areproduced by many of them, making purification difficult,their xylanases warrant further investigation.
ACKNOWLEDGMENTS
We thank Chittra Mishra for her expert advice and assistance withthe paper chromatography experiments, Robert B. Hespell forsupplying xylooligosaccharides, and the staff of the University ofWisconsin Biotechnology Center Protein Purification Facility fortheir support and encouragement during the preparation of themanuscript.
REFERENCES
1. Baker, C. J., C. H. Whalen, and D. F. Bateman. 1977. Xylanasefrom Trichoderma pseudokoningii: purification, characteriza-tion, and effects on isolated plant cell walls. Phytopathology67:1250-1258.
2. Ball, A. S., and A. J. McCarthy. 1988. Saccharification of strawby actinomycete enzymes. J. Gen. Microbiol. 134:2139-2147.
3. Bertrand, J. L., R. Morosoli, F. Shareck, and D. Kluepfel. 1989.Expression of the xylanase gene of Streptomyces lividans andproduction of the enzyme on natural substrates. Biotechnol.Bioeng. 33:791-794.
4. Chang, L. T., and R. P. Elander. 1986. Long-term preservationof industrially important microorganisms, p. 49-55. In A. L.Demain and N. A. Solomon (ed.), Manual of industrial micro-
VOL. 57, 1991
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

992 GRABSKI AND JEFFRIES
biology and biotechnology. American Society for Microbiology,Washington, D.C.
5. Chauvet, J.-M., J. Comtat, and P. Noe. 1987. Assistance inbleaching of never-dried pulps by the use of xylanases, conse-quences on pulp properties, p. 325-327. In Proceedings of the4th International Symposium on Wood Pulp Chemistry, 27 to 30April 1987, Paris.
6. Dekker, R. F. H., and G. N. Richards. 1975. Purification,properties, and mode of action of hemicellulase I produced byCeratocystis paradoxa. Carbohydr. Res. 39:97-114.
7. Goodfellow, M., S. T. Williams, and G. Alderson. 1986. Transferof Chainia species to the genus Streptomyces with emendeddescription of species. Syst. Appl. Microbiol. 8:55-60.
8. lizuka, H., and T. Kawaminami. 1969. Isolation and selection ofxylanase-producing micro-organisms and the identification of anew species of Streptomyces. Agric. Biol. Chem. 33:1257-1263.
9. Johnson, K. G., B. A. Harrison, H. Schnider, C. R. MacKenzie,and J. D. Fontana. 1988. Xylan-hydrolysing enzymes fromStreptomyces spp. Enzyme Microb. Technol. 10:403-409.
10. Jurasek, L., and M. G. Paice. 1988. Biological bleaching of pulp,p. 11-13. In T. J. Salvo (ed.), International Pulp BleachingConference, Orlando, Fla. TAPPI proceedings. Technical As-sociation of the Pulp and Paper Industry, Atlanta.
11. Kantelinen, A., M. Ratto, J. Sundquist, M. Ranua, L. Viikari,and M. Linko. 1988. Hemicellulases and their potential role inbleaching, p. 1-4. In T. J. Salvo (ed.), International PulpBleaching Conference, Orlando, Fla. TAPPI proceedings. Tech-nical Association of the Pulp and Paper Industry, Atlanta.
12. Kawaminami, T., and H. Iizuka. 1969. Studies on xylanase frommicroorganisms. Part III. Production of xylanase by Streptomy-ces xylophagus nov. sp. Agric. Biol. Chem. 33:1787-1789.
13. Kluepfel, D., F. Shareck, F. Mondou, and R. Morosoli. 1986.Characterization of cellulase and xylanase activities of Strepto-myces lividans. Appl. Microbiol. Biotechnol. 24:230-234.
14. Koyama, H., M. Ujiie, H. Taniguchi, and T. Sasaki. 1990.Purification and some properties of xylan-hydrolysing enzymesfrom Robillarda sp. Y-20. Enzyme Microb. Technol. 12:218-224.
15. Miller, G. L. 1959. Use of dinitrosalicylic acid reagent fordetermination of reducing sugars. Anal. Chem. 31:426-429.
16. Mora, F., J. Comtat, F. Barnoud, F. Pla, and P. Noe. 1986.Action of xylanases on chemical pulp fibers. Part I. Investiga-tions on cell-wall modifications. J. Wood Chem. Technol.6:147-165.
17. Morosoli, R., J.-L. Bertrand, F. Mondou, F. Shareck, and D.Kluepfel. 1986. Purification and properties of a xylanase fromStreptomyces lividans. Biochem. J. 239:587-592.
18. Nakanishi, K., H. Arai, and T. Yasui. 1984. Purification and
some properties of xylanase from Cryptococcus flavus. J. Fer-ment. Technol. 62:361-369.
19. Nelson, N. 1944. A photometric adaptation of the Somogyimethod for the determination of glucose. J. Biol. Chem. 153:375-380.
20. Noe, P., J. Chevalier, F. Mora, and J. Comtat. 1986. Action ofxylanases on chemical pulp fibers. Part II. Enzymatic beating. J.Wood Chem. Technol. 6:167-184.
21. Paice, M. G., R. Bernier, Jr., and L. Jurasek. 1988. Viscosity-enhancing bleaching of hardwood kraft pulp with xylanase froma cloned gene. Biotechnol. Bioeng. 32:235-239.
22. Reilly, P. J. 1981. Xylanases: structure and function, p. 111-129.In A. Hollander (ed.), Trends in the biology of fermentations.Plenum Publishing Corp., New York.
23. Roybt, J. F., and W. H. Whelan. 1972. Reducing value methodsfor maltodextrins. 1. Chain length dependence of alkaline 3,5-dinitrosalicylate and chain-length independence of alkaline cop-per. Anal. Biochem. 45:510-516.
24. Royer, J. C., and J. P. Nakas. 1989. Xylanase production byTrichoderma longibrachiatum Enzyme Microb. Technol. 11:405-410.
25. Somogyi, M. 1952. Notes on sugar determination. J. Biol. Chem.195:19-23.
26. Srinivasan, M. C., H. G. Vartak, V. K. Powar, M. V. Rele, andK. B. Bastawde. 1984. Studies on xylan degrading enzyme fromChainia. Biotechnol. Lett. 6:715-718.
27. Tan, L. U. L., K. K. Y. Wong, E. K. C. Yu, and J. N. Saddler.1985. Purification and characterization of two D-xylanases fromTrichoderma harzianum. Enzyme Microb. Technol. 7:425-430.
28. Tan, L. U. L., E. K. C. Yu, G. W. Louis-Seize, and J. N. Saddler.1987. Inexpensive, rapid procedure for bulk purification ofcellulase-free P-1,4-D-xylanase of high specific activity. Bio-technol. Bioeng. 30:96-100.
29. Tien, M., and T. K. Kirk. 1988. Lignin peroxidase of Phanero-chaete chrysosporium. Methods Enzymol. 161:238-249.
30. Trevelyn, W., P. D. Procter, and J. S. Harrison. 1950. Detectionof sugars on paper chromatograms. Nature (London) 166:444-445.
31. Viikari, L., A. Kantelinen, K. Poutanen, and M. Ranua. 1990.Characterization of pulps treated with hemicellulolytic enzymesprior to bleaching, p. 145-151. In T. K. Kirk and H.-M. Chang(ed.), Biotechnology in pulp and paper manufacture. Butter-worth-Heinemann, Stoneham, Mass.
32. Walseth, C. S. 1952. Occurrence of cellulases in enzyme prep-arations from microorganisms. TAPPI J. 35:228-233.
33. Wolin, E. A., M. J. Wolin, and R. S. Wolfe. 1963. Formation ofmethane by bacterial extracts. J. Biol. Chem. 238:2882-2886.
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from