development of a new cell-based assay for the ... · altex proceedings, 1/12, proceedings of wc8 89...
TRANSCRIPT

Altex Proceedings, 1/12, Proceedings of WC8 89
1 Introduction
FSH is an important pharmaceutical drug for the therapeutic control and support of female fertility. FSH is a heterodimeric glycoprotein of approximately 30 kDa synthesized in the ante-rior pituitary gland. It consists of a common α-subunit that is shared with other gonadotropins non-covalently linked to the activity bearing β-subunit (Baenziger and Green, 1988). For more than 40 years now, it has been used successfully for the treatment of female infertility. Besides purified FSH products retrieved from the urine of post-menopausal women, an increas-ing number of recombinant preparations from mammalian cell culture systems of higher specific activity and increased safety profile have been introduced in the past 20 years (Gleicher et al., 2003).
Until now, the only regulatory approved activity testing fol-lows an in vivo approach that was established more than half a century ago (Steelman and Pohley, 1953; USP, 2004; EP, 2011). This assay is based on FSH-induced gain in ovary weight of
premature rats. It has a number of limitations, which include its limited precision and cumbersome interpretation procedures. The assay is labor-intensive and time-consuming. Due to its de-pendency on experimental skills, testing services are limited to a few contract research organizations. Because the female rats need to be in a certain stage of development for this in vivo assay, this may involve delicate sample storage, e.g., for early material testing and in-process control samples with limited sta-bility, shipment logistics, and scheduling. In addition, the assay is ethically questionable due to it requiring a large number of animals. In line with the latter, the european Commission for enterprise and industry formed industrial associations with the pharmaceutical, chemical, cosmetics, and biotechnology sec-tors promoting its aim to reduce animal testing in the process of product development and release (Balls, 2009). A testing ban on finished cosmetic products and ingredients already is applied stepwise as of 2003, with a final cut-off date in March 2013, irrespective of the availability of alternative non-animal tests (EC, 2003).
Development of a New Cell-Based Assay for the Determination of FSH ActivityJan Rohde 1⁕, Christian D. Demmler 2⁕, Frank Mueller 1, Yoshihiro Nishi 3, Toshihiko Yanase 3, and Christoph Giese 2
1R&D Department, Minapharm Pharmaceuticals, 10th of Ramadan City, Cairo, Egypt; 2Cell And tissue Services, ProBioGen AG, Berlin, Germany; 3Dept. of Medicine and Bioregulatory Science, Graduate School of Medical Sciences, Kyushu University, Fukuoka, Japan
SummaryFollicle stimulating hormone (FSH) plays a central role for in vitro fertilization therapy. Historically, FSH was extracted from urine of post-menopausal women. Now, most preparations used in Europe and North America are of recombinant origin. For the past several decades, the potency of FSH dosage has been determined by a single in vivo potency assay based on weight gain of rat ovaries upon hormonal treatment. This assay lacks precision, is cumbersome in interpretation, and sacrifices large numbers of rats.We herewith present the development and qualification of an alternative potency assay that will circumvent the aforementioned obstacles. The setup is based on a human ovary granulosa cell lines expressing the intact human FSH receptor on its surface. FSH binding results in induction of progesterone expression and secretion, which is subsequently quantitatively determined by a commercially available diagnostic ELISA. We demonstrate in this publication that the new assay features excellent precision and accuracy and has a wider working range than the in vivo assay. Analyses of commercial products of different origin using the in vitro assay confirm their stated potency. Further, it was demonstrated that the assay delivered comparable results to the Steelman/Pohley assay for a newly developed recombinant FSH product. The assay has been successfully validated for that product. Its accuracy, handling, and universal application for FSH preparations of different origin qualify the presented assay to complement and replace the Steelman/Pohley assay in the future.
Keywords: Follicle stimulating hormone (FSH), cell-based assay, Steelman-Pohley assay, potency testing, granulosa cell line, progesterone, cAMP
⁕ These authors contributed equally to this publication.

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC890
Due to the limitations of the in vivo assay, several physico-chemical methodologies such as size-exclusion chromatog-raphy and quantitative isoelectric focusing have been applied recently to marketed FSH products, thereby circumventing reg-ular in vivo testing (Driebergen and Baer, 2003; Mulders et al., 1997; Almeida et al., 2011). These approaches show a higher accuracy than the Steelman-Pohley assay but are limited due to their product specificity requiring intensive correlative studies versus the in vivo assay. Together with immune-specific binding assays, e.g., in the elISA-format, these analytical methods do not characterize bioactivity as recommended for other biologics (e.g., interferons and other cytokines) for batch release by the ICH guidelines or US and EU Pharmacopoeia (e.g., Chapter 5.6 “Assay of Interferons”).
We have designed a cell-based assay using a relevant human target cell to characterize the bioactivity of recombinant human FSH preparations. the maturation of primary ovarial follicles is induced by the gonadotropin FSH. Granulosa cells are the natural target cells in the mammalian ovary. FSH binds to the membrane based G-protein coupled FSH receptor. Intracellu-lar cAMP signaling induces steroid synthesis by increased gene expression and phosphorylation triggered activation of relevant
enzymes, e.g., aromatase (ARO). FSH-induced granulosa cells secrete progesterone and estradiol (E2). Progesterone induces uterus proliferation for embryonal nidation in uteral mucosa and suppresses the release of gonadotropin releasing hormone (GnRH) in the hypothalamus. Follicular granulosa cells are the first producers of progesterone. Later on, progesterone produc-tion is taken over by the corpus luteum. estradiol interacts in a positive feedback loop on hypothalamic GnRH release and the hypophyseal gonadotropins FSH and LH. (Fig.1). Monitor-ing one or more steps of the receptor binding signaling steroid synthesis or secretion seemed to be most appropriate for a cell-based assay with physiological relevance.
Various human granulosa-like cell lines are published and available for research and applications. Cell lines were either established from ovarian tumors like the HTOG, COV434, and KGN cell lines or developed through oncogenic transformation like the HGL5, HO-23, GCIa, and HGP53 cell lines. KGN and COV434 are naturally transformed primary cells described as responsive to FSH in physiologically relevant concentrations (Havelock et al., 2004).
Because of their primary capability of inducible steroid pro-duction and secretion in combination with well-established
Fig. 2: Steroid biosynthesis of granulosa cells (simplified)The production of steroids is under the control of cyclic adenosine monophosphate (cAMP). Binding of FSH to the FSH receptor (FSHr) triggers the adenylate cyclase (AC) induced of synthesis of cAMP and subsequently signaling cascades for cAMP regulated transcription of enzymes. Progesterone is synthesized from cholesterol. Estradiol is synthesized from androstenedione. The key enzyme aromatase (ARO) converts androstenedione to estrone, a precursor of estradiol. Since Theca cells synthesize androstenedione in vivo according to the two cells/two gonatropins theory, this precursor has to be supplemented to the cell culture medium for estradiol synthesis (Havelock et al., 2004; Parakh et al., 2006).
Fig. 1: The neuro-endocrine control of follicle maturation and ovulation during menses (simplified)Hypothalamic releasing hormone (GnRH) induces the secretion of the pituitary gonadotropins FSH and LH. Follicular granulosa cells produce progesterone and estradiol. Both gonadotropins have an inhibitory effect on GnRH secretion of pituitary in a negative feedback loop.

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC8 91
progesterone or estradiol analysis in culture supernatants by commercial available diagnostic ELISA kits, KGN and COV434 can be used for assay development without addi-tional modifications or genetic engineering, e.g., reporter gene implementation.
The cell-based assay needed to be designed for multi-well formats for the rapid automated analysis of large numbers of samples, e.g., for screening or HTS applications. A broad range of FSH concentrations in samples ensured dose-response curves for the determination of effective concentration levels (EC50) and state-of-the-art data analysis, e.g., by the application of multi-parametric fitting algorithms (4P/5P).
The cost effective, robust, and rapid assay format can be used for the analysis of in-process controls (IPCs), early up- and downstream material, e.g., for the selection of high-producer clones, production clone, or lead candidate characterization, and for mode-of-action analysis (MoA).
2 Material and methods
FSH preparationsRecombinant FSH drug products from CHO cell lines, as well as urinary-derived preparations, were purchased from local pharmacies. Two recombinant CHO derived FSH products, Gonal F® (Merck-Serono, Darmstadt, Germany, Lot-No. Y15A9223), which has a stated activity of 13.6 IU/µg, and Pur-egon® (MSD, Munich, Germany, Lot-No. 719756), which has a stated activity of 10 IU/µg were used. A urinary derived FSH product Fostimon® (Institue Biochimique SA, Manno, IBSA, Switzerland, Lot-No. 100451) had a stated activity of 6.0 IU/µg. Gonapure® batches were provided by Minapharm Pharma-ceuticals (Cairo, Egypt). The 1st international standard for FSH (For bioassay; 92/642) was purchased from the National Insti-tute for Biological Standards and Control (NIBSC; Potters Bar, UK) and has a stated activity of 13.8 IU/µg.
Assay cell linesThe KGN cell line was purchased from the Riken Cell Bank (Riken Institute, Japan) and was cultured in DMEM/F-12 (Invi-torgen, Carlsbad, CA) with 10% FCS (Biochrom, Berlin, Ger-many). KGN cells grow adherent in a homogeneous monolayer. The cells were split using 0.5 ml TypLE Express (Invitrogen) per 25 cm2 for 3 min at 37°C. The cell line was incubated at 37°C at 5% CO2 in a humidified incubator. A research cell bank of 100 vials was prepared, cryopreserved, and stored in the gas phase of liquid nitrogen according to the Good Cell Cul-ture Practice (GCCP) recommendations of ECVAM (European Centre for the Validation of Alternative Methods, Coecke et. al., 2005; Hartung et al., 2002).
The COV434 cell line was purchased from the European Cell Culture Collection (Porton Down, UK). The round shaped COV434 cell line grows in small, slightly adherent clusters. The cells were split using 0.5 ml/25 cm2 TrypLE Express for 5 min at 37°C. A research cell bank of 100 vials was produced as al-ready described.
Both cell lines were confirmed sterile (Steritest, Millipore, Billerica, MA, USA) and free from mycoplasma (PCR based test, Minerva Biolabs, Berlin, Germany). For each experiment, a new vial was revitalized and seeded at 2.5 viable cells/T150 cell culture flask 3-4 days prior to assay inoculation.
ImmunofluorescenceKGN were seeded at 5 x104 viable cells/well in DMEM/F-12 (Invitrogen, Grand Island, NY, USA) with 10% FCS (Bio-chrom, Berlin, Germany) on glass slides in 12-well plates 3 days prior to fixation. The slides then were fixed with methanol for 5 min on an iso-propanol ice bath and subsequently air dried. The slides then were blocked with PBS + 2% FCS + 0.25 mM CaCl2 (Merck, Darmstadt, Germany), incubated with Cy3-con-jugated mouse anti-vimentin (Sigma-Aldrich) and mouse anti-pan-cytokeratin (Dako, Glostrup, Denmark) primary antibody for 45 min, and washed 2 times with PBS. The anti-vimentin specimens were incubated with 1 µg/ml 4',6-diamidino-2-phe-nylindole (DAPI, Sigma-Aldrich) for 15 min and the anti-pan-cytokeratin specimens were incubated with AlexaFluor488-conjugated goat anti-mouse IgG heavy and light chain (H+L) secondary antibody with 1 µg/ml DAPI for another 45 min. All antibodies were used at a concentration of 5 µg/ml.
Flow cytometryThe distribution of human FSH receptor (hFSHr) on KGN cells was determined by flow cytometry. KGN were revitalized 4 days prior to flow cytometric analysis and detached with TypLE Express for 5 min at 37°C prior to staining. 1 x106 viable cells were stained with 5 µg/ml anti-FSHr (AbD Serotec, Oxford, UK) for 20 min (RT), washed twice with PBS and subsequently stained with AlexaFluor488-conjugated donkey anti-sheep IgG (H+L) secondary antibody for 20 min (RT) in the dark. The sam-ples were washed again and immediately analyzed (10,000 cells/gate R1 FSC-SSC dot-plot) using a CyFlow space flow cytometer (Partec, Münster, Germany). Unspecific binding was determined by staining with secondary anti-sheep antibody alone.
Steelman-Pohley in vivo assay The Steelman-Pohley-Assay was performed by Bioassay – Lab-oratory for biological analytics GmBH (Heidelberg, Germany) according to the European Pharmacopoeia monograph (EP, 2011). Female, immature Lewis rats aged 22-24 days were used for in vivo testing (six rats per treatment group). The animals were weighed prior to the first administration and after the end of the study. Differences in individual body weight at the com-mencement of the study were within 10 g.
FSH from NIBSC (08/282) was used as reference material. The lyophilisate was re-suspended in 1 ml of phosphate-albu-min buffered saline solution, pH 7.2. The stock solution was diluted to the final concentrations and 1.78 IU/ml, using the re-suspension buffer. Three different groups of test animals were treated with three different dose levels on 3 consecutive days by subcutaneous injection (0.5 ml per animal and day). On day 4 of FSH treatment the animals were sacrificed. Ovaries were excised, cleaned, and weighed.

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC892
Cell-based assay optimized for progesterone secretionThe KGN cells were revitalized 3-4 days prior to assay inoc-ulation and seeded at a density of 2.5 x106 viable cells/T150 flask in 20 ml DMEM/F-12 + 10% FCS. FSH samples were dispensed in serial dilutions of 1:3 on a separated non-binding plate. KGN cells were seeded at 5 x104, 1.0 x104, and 2.0 x105
viable cells/well with 50 µl/well of the respective FSH dilution in a cellbind 96-well plate. The cells were incubated for 24, 48, 72, and 96 h at 37°C and 5% CO2.
Progesterone Enzyme ImmunoassayInduced progesterone synthesis was measured using a pro-gesterone EIA kit (Beckman Coulter, Brea, CA, USA). Therefore, 50 µl of each well was transferred into the corresponding well of the pre-coated and equilibrated EIA plate, incubated for 5 min on a shaker, mixed with 200 µl enzyme conjugate, and incubated for another 60 min. The plate was then washed and 200 µl substrate solution was added for 15 min until color de-velopment stopped. Absorptions were analyzed within 15 min after addition of the stop solution at 450 nm using a Tecan Infi-nite F200 multiwell-plate reader (Tecan).
Qualification experimentsThe geometric means (G) were used to calculate the mean of the sample size according to equation [1] using log-trans-formed activity data. The standard deviations (s) were calcu-lated as follows [2] using log-transformed activity data. the geometric coefficient of variation (GCV) was calculated ac-cording to equation [3]. The confidence intervals (CI) were cal-culated using equation [4]. The inter-serial precision (PI) of the method was calculated according to equation [5]. An ANOVA test was performed using Origin 8.1 (OriginLab Cooperation, North Hampton, MA). The accuracy of the test method was determined by dilution experiments of the test sample where the sample was pre-diluted to a theoretical sample potency of 50%, 75%, 100%, and 200%. The relative bias was calculated as stated in equation [6]. The 90% confidence interval of cal-culated activity data gives information about the accuracy of
Metabolic activity4 x103 viable cells/well were cultivated in serum supplement-ed media DMEM/F-12 + 10% FCS or serum reduced media (DMEM/F-12 + 1% FCS + 5 µg/ml insulin + 500 ng/ml hy-drocortisone + 5 µg/ml transferrin) in 96-well plates and incu-bated with serial dilutions of FSH (range of 2.6 pg/ml to 1 µg/ml) in the respective cell culture medium for 3, 4, and 5 days. 4 h before the end of the incubation time, 10 µl 10x resazurin solution (Sigma-Aldrich) was added to each well to determine the metabolic activity of the cells. Fluorescence intensity was then read using an Infinite 200 series reader (Tecan, Männedorf, Switzerland) at 535 nm excitation and 580 nm for emission.
Cyclic Adenosine Monophosphate (cAMP)Induced cAMP was analyzed using a cAMP ELISA (R&D Sys-tems, Minneapolis, MN, USA) according to the manufacturer’s protocol. KGN were inoculated at 1.5 x106 viable cells/well in 6-well plates with 3 ml DMEM/F-12 + 10% FCS + 0.1 mM IBMX (Sigma-Aldrich). For cell permeabilization the superna-tant was discharged and the cells were frozen at -20 °C in 150 µl cell lysis buffer. In total, 3 freeze-thaw cycles were performed. Samples were subsequently centrifuged at 600 x g for 10 min and assayed immediately.
Analysis of aromatase activityInduced aromatase was analyzed using an anti-human aro-matase ELISA (USCN Life Sciences, Wuhan, China) accord-ing to the manufacturer’s instructions. KGN were inoculated at 2 x105 viable cells/ml in 24-well plates in DMEM/F-12 + 10% FCS + 200 nM androstenedione and incubated for 46 h with serial dilutions of FSH (Range of 9.3 ng/ml to 250 ng/ml) at 37°C / 5% CO2. In order to break the cell walls, the supernatant was discharged and the cells were frozen at -20°C in 110 µl PBS. In total, 3 freeze-thaw cycles were performed. Samples were subsequently centrifuged at 5,000 x g for 10 min and as-sayed immediately.
Cell-based assay optimized for estradiol secretionthe cell lines were inoculated at 4 x103 viable cells/well in 96-well plates. Serial 1:3 dilutions of FSH samples were prepared in DMEM/F-12 + 10% FCS, DMEM/F-12 + 10% FCS + 100 nM androstenedione, and DMEM/F-12 + 1% FCS + 5 µg/ml insulin (Invitrogen) + 500 ng/ml hydrocortisone + 5 µg/ml transferrin + 1x defined lipid mix + 100 nM andros-tenedione (all from Sigma-Aldrich). The cells were incubated with serial dilutions of FSH (range of 2.6 pg/ml to 1 µg/ml) for 108 h at 37°C and 5% CO2.
Estrodiol Enzyme ImmunoassayInduced estradiol synthesis is measured using an estradiol eIA kit (Cayman, Ann Arbor, MI, USA) according to the manufac-turer’s protocol. Absorption values were read within 15 min after addition of the stop solution at 450 nm using a Tecan In-finite F200 multiwell-plate reader (Tecan). Curve fits were ac-complished using a 4-parameter logistic dose-response model with weighting of the standard deviations (Magellan Software, version 6.1, Tecan)
nnxxxxG ...321 [1]
1
)( 1
nGx
s [2]
%100)110( sGCV [3]
nstGCI n
1)/1( [4]
)110(100PrecisionteIntermedia )()( errorVARrunVAR [5]
whereas
2)()(
)(errorMSrunMS
runVAR
)()( errorMSerrorVAR
1100[%])(
potencytargetltheoreticapotencymeasuredmeanRBbiasrelativemean [6]

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC8 93
revitalization was analyzed in a batch kinetic and showed re-producible performance, as expected (data not shown). KGN and COV434 were tested free of mycoplasma and microbial contaminations.
Aromatase activitythe induction of aromatase expression was analyzed with dif-ferent FSH concentrations. The KGN cell line showed FSH dose-dependent aromatase expression when cultivated with an-drostenedione supplemented cell culture medium (Fig. 5).
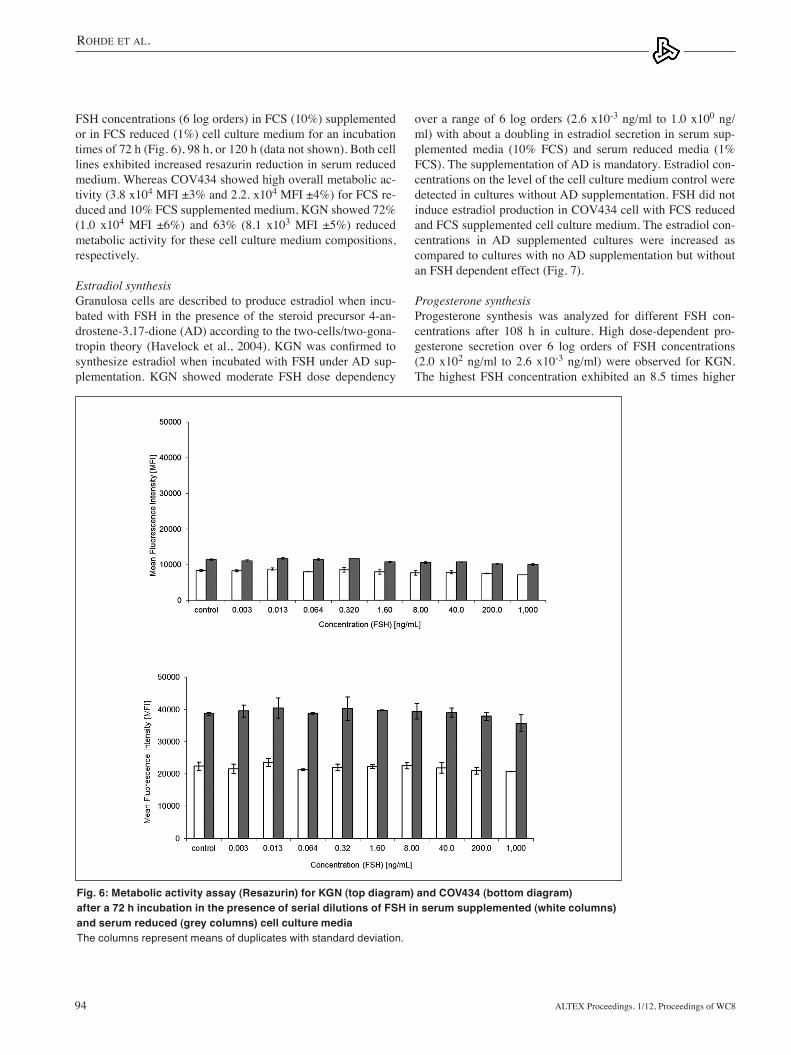
Metabolic activity The influence of FSH on the metabolic activity of COV454 and KGN was investigated using a resazurin assay. In both cell lines the metabolic activity was not changed over a broad range of
the method. Dose-response curves were accomplished using a 4-parameter logistic dose-response model with weighting of the standard deviations (Origin 8.1).
3 Results
Cell line characterizationKGN was found to be homogeneously positive for vimentin, whereas pan-cytokeratin expression was not detected (Fig. 3). These findings are consistent with the literature for granulo-sa-like tumor cell lines (Czernobilsky et al., 1987). The cell line was characterized for human FSH receptor (hFSHr) by flow cytometry, and it showed homogeneous expression of this receptor (Fig. 4). Furthermore, the growth behavior after
Fig. 3: Immunofluorescence staining of KGN for pan-cytokeratin (I) and vimentin (II) and the respective counterstained DAPI images (WU filter, Olympus)KGN are negative for cytokeratin (I) and positive for vimentin (II). Cytokeratin was labeled using an AlexaFluor488 conjugated anti-mouse IgG secondary antibody. The anti-vimentin antibody was Cy3 conjugated (NB and WG filters, respectively; with 100x magnification; Olympus IX-50).
Fig. 4: hFSHr expression of KGN (black line) and the according isotype control (gray line)
Fig. 5: FSH dose dependent aromatase expression levels (under androstenedione supplementation)The columns represent means of triplicates with standard deviation.
Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC894
over a range of 6 log orders (2.6 x10-3 ng/ml to 1.0 x100 ng/ml) with about a doubling in estradiol secretion in serum sup-plemented media (10% FCS) and serum reduced media (1% FCS). The supplementation of AD is mandatory. Estradiol con-centrations on the level of the cell culture medium control were detected in cultures without AD supplementation. FSH did not induce estradiol production in COV434 cell with FCS reduced and FCS supplemented cell culture medium. the estradiol con-centrations in AD supplemented cultures were increased as compared to cultures with no AD supplementation but without an FSH dependent effect (Fig. 7).
Progesterone synthesisProgesterone synthesis was analyzed for different FSH con-centrations after 108 h in culture. High dose-dependent pro-gesterone secretion over 6 log orders of FSH concentrations (2.0 x102 ng/ml to 2.6 x10-3 ng/ml) were observed for KGN. The highest FSH concentration exhibited an 8.5 times higher
FSH concentrations (6 log orders) in FCS (10%) supplemented or in FCS reduced (1%) cell culture medium for an incubation times of 72 h (Fig. 6), 98 h, or 120 h (data not shown). Both cell lines exhibited increased resazurin reduction in serum reduced medium. Whereas COV434 showed high overall metabolic ac-tivity (3.8 x104 MFI ±3% and 2.2. x104 MFI ±4%) for FCS re-duced and 10% FCS supplemented medium, KGN showed 72% (1.0 x104 MFI ±6%) and 63% (8.1 x103 MFI ±5%) reduced metabolic activity for these cell culture medium compositions, respectively.
Estradiol synthesisGranulosa cells are described to produce estradiol when incu-bated with FSH in the presence of the steroid precursor 4-an-drostene-3,17-dione (AD) according to the two-cells/two-gona-tropin theory (Havelock et al., 2004). KGN was confirmed to synthesize estradiol when incubated with FSH under AD sup-plementation. KGN showed moderate FSH dose dependency
Fig. 6: Metabolic activity assay (Resazurin) for KGN (top diagram) and COV434 (bottom diagram) after a 72 h incubation in the presence of serial dilutions of FSH in serum supplemented (white columns) and serum reduced (grey columns) cell culture mediaThe columns represent means of duplicates with standard deviation.

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC8 95
Fig. 7: Estradiol concentrations in KGN (top diagram) and COV434 (bottom diagram) culture supernatants after 108 h of FSH stimulation (cultures in duplicates) with different media supplementations (white columns: serum supplemented cell culture media; grey columns: serum supplemented cell culture media with AD supplementation; black columns: serum reduced cell culture media with AD supplementation)The control was cultured without FSH supplementation. The columns represent means of duplicates including standard deviation.
Fig. 8: Progesterone concentrations in KGN (top diagram) and COV434 (bottom diagram) culture supernatants after 108 h of FSH stimulation (cultures in duplicates) with different media supplementations (white columns: serum supplemented cell culture media; grey columns: serum supplemented cell culture media with AD supplementation; black columns: serum reduced cell culture media with AD supplementation)The control was cultured without FSH supplementation. The columns represent means of duplicates with standard deviation.

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC896
termined by dilution experiments of 92/642. The relative bias and the 90% confidence interval for the calculated activities are displayed in Table 1. The error bars indicate the 90% confidence interval of the mean sample potencies. this data shows a rela-tive bias of ≤20% for the 90% CI for the 50%, 100%, and 200% sample potencies. The 90% CI for these sample potencies lay within ±20% of the actual sample potency. Therefore, a work-ing range from 50% to 200% sample potency was regarded as accurate in this assay. the potency of the 1st standard for FSH was determined with 132 IU/ml (Reference value 138 IU/ml). This reflects a relative bias of -4%. The 95% confidence interval for this standard material ranges from 119 IU/ml to 147 IU/ml. Subsequently, an internal Gonapure® reference standard was calibrated against the 1 international standard for FSH and the assay was validated with results comparable to the preliminary qualification (data not shown).
Comparison of the stated specific activity of commercial
progesterone secretion as compared to the cell culture medium control without FSH when FCS-reduced medium was used. A 5.7 time increase was observed for KGN when 10% FCS-supplemented medium was used. COV434 did not show FSH dependent progesterone synthesis. Over 5 log orders of FSH concentrations, ranging from 3.2 x10-2 ng/ml to 5.0 x102 ng/ml, the detected progesterone concentrations were on the level of the cell culture medium control (Fig. 8).
Establishment of an FSH cell-based assay for induced progesterone synthesisKGN cells were seeded at 5 x104, 1 x105, and 2 x105 viable cells/ml and incubated for 24 h, 48 h, 72 h, and 96 h with 100 ng/ml and without FSH. An inoculation cell density of 5 x104 viable cells/ml resulted in a dynamic range of 5%, 5%, 6%, and 21% for an incubation time of 24 h, 48 h, 72 h, and 96 h, respectively. An inoculation cell density of 1 x105 viable cells/ml resulted in a dynamic range of 7%, 13%, 22%, and 28%, respectively for these incubations times and an inocula-tion cell density of 2 x105 viable cells/ml resulted in a dynamic range of 20%, 31%, 31%, and 29%, respectively. In order to re-duce the assay time, an incubation time of 48 h was selected. An inoculation cell density of 2 x105 viable cells/ml was selected in order to maximize the dynamic range of the assay.
Qualification of an FSH cell-based assaythe 1st international WHO standard for FSH (Recombinant; NIBSC code 92/642) was used as a standard and sample in the course of the qualification. This preparation has a unitage of 138 IU/ampoule with a nominal content of 10 µg per ampoule. The cell-based assay was qualified regarding its inter-serial precision, accuracy, and robustness when conducted by two different operators. the underlying method has an inter-serial precision of IP ≤16.7% when performed by different operators and on different days. the linearity of the test method was de-
Tab. 1: Summary of the qualification experiments
200% sample 100% sample 50% sample potency potency potency
Theoretical sample 27.6 13.8 6.9 potency [IU/µg]
G [µg/ml] 25.6 13.5 6.6
GCV [%] 21% 8% 16%
RP [%] -7% -2% -4%
CI90% (RB) [%] 8% 7% 5%
CI90% (RB) [%] -20% -14% -12%
Fig. 9: Comparison of the dynamic range at different inoculation cell densities (5 x104, 1 x105, and 2 x105 viable cells/ml) and incubation times (24 h, 48 h, 72 h, and 96 h)The columns represent means of duplicates including standard deviation.

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC8 97
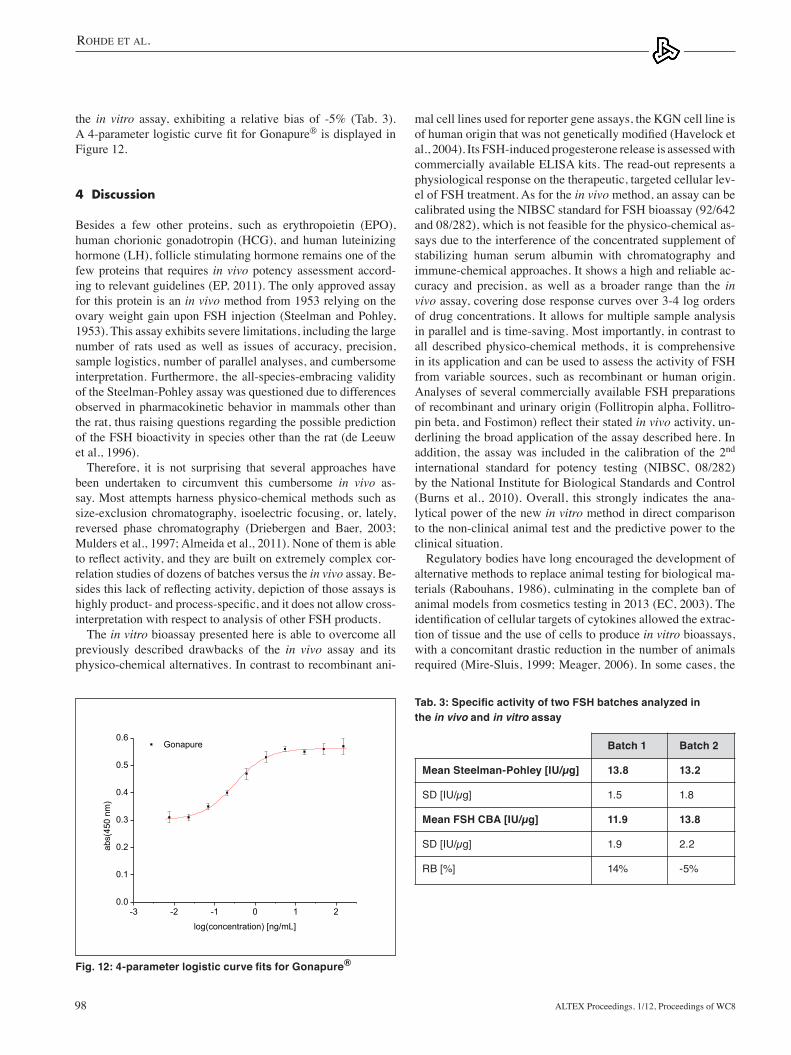
and FSH CBATwo Gonapure® batches were analyzed in the Steelman- Pohley assay and FSH CBA. The activity data are summa-rized in Table 2. The derived specific activity of batch 1 is 13.8 IU/µg ±1.5 IU/µg in the in vivo assay and 11.9 IU/µg ±1.9 IU/µg in the in vitro assay, exhibiting a relative bias of +14%. The derived specific activity of batch 2 is 13.2 IU/µg ±1.6 IU/µg in the in vivo assay and 13.8 IU/µg ±2.2 IU/µg in
FSH products with the calculated specific activity derived from the FSH CBAThe stated specific activity given by the manufacturers of three different FSH products was compared to the FSH CBA derived specific activity. A specific activity of Gonal F (Merck-Serono) was determined with 12.9 IU/µg. This reflects a rela-tive bias of -5% when compared to the stated specific activ-ity of 13.6 IU/µg. For Fostimon (IBSA) a specific activity of 4.1 IU/µg was determined, which reflects a relative bias of -31% when compared to the stated value of 6.0 IU/µg. For Puregon (Schering Plough) a specific activity of 12.9 IU/µg was deter-mined, and this reflects a relative bias of 29% when compared to the 10.0 IU/µg, which is stated by the manufacturer (Tab. 2). 4-parameter logistic curve fits for these FSH products are displayed in Figure 11.
Comparison of results from the Steelman-Pohley assay
Fig. 10: Mean and relative bias of the specific activity at different sample potencies on the basis of a dilution experimentFor the 1st international standard for FSH the method is accurate (90%CI ≤ ±20%) in the range from 50% to 200% sample potency.
Fig. 11: 4-parameter logistic curve fits for different FSH products. Gonal F® (top), Puregon® (middle) and Fostimon® (bottom)
Tab. 2: Comparison of the by the manufacturer stated and in the FSH CBA determined specific activity of commercially available FSH products
Gonal F® Fostimon® Puregon® (Merck (IBSA) (MSD) Serono)
FSH-CBA Mean specific 12.9 4.1 12.9 activity [IU/µg]
Stated specific activity 13.6 6.0 10.0 [IU/µg]
Relative bias [%] -5% -31% +29%
Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC898
mal cell lines used for reporter gene assays, the KGN cell line is of human origin that was not genetically modified (Havelock et al., 2004). Its FSH-induced progesterone release is assessed with commercially available ELISA kits. The read-out represents a physiological response on the therapeutic, targeted cellular lev-el of FSH treatment. As for the in vivo method, an assay can be calibrated using the NIBSC standard for FSH bioassay (92/642 and 08/282), which is not feasible for the physico-chemical as-says due to the interference of the concentrated supplement of stabilizing human serum albumin with chromatography and immune-chemical approaches. It shows a high and reliable ac-curacy and precision, as well as a broader range than the in vivo assay, covering dose response curves over 3-4 log orders of drug concentrations. It allows for multiple sample analysis in parallel and is time-saving. Most importantly, in contrast to all described physico-chemical methods, it is comprehensive in its application and can be used to assess the activity of FSH from variable sources, such as recombinant or human origin. Analyses of several commercially available FSH preparations of recombinant and urinary origin (Follitropin alpha, Follitro-pin beta, and Fostimon) reflect their stated in vivo activity, un-derlining the broad application of the assay described here. In addition, the assay was included in the calibration of the 2nd international standard for potency testing (NIBSC, 08/282) by the National Institute for Biological Standards and Control (Burns et al., 2010). Overall, this strongly indicates the ana-lytical power of the new in vitro method in direct comparison to the non-clinical animal test and the predictive power to the clinical situation.
Regulatory bodies have long encouraged the development of alternative methods to replace animal testing for biological ma-terials (Rabouhans, 1986), culminating in the complete ban of animal models from cosmetics testing in 2013 (EC, 2003). The identification of cellular targets of cytokines allowed the extrac-tion of tissue and the use of cells to produce in vitro bioassays, with a concomitant drastic reduction in the number of animals required (Mire-Sluis, 1999; Meager, 2006). In some cases, the
the in vitro assay, exhibiting a relative bias of -5% (Tab. 3). A 4-parameter logistic curve fit for Gonapure® is displayed in Figure 12.
4 Discussion
Besides a few other proteins, such as erythropoietin (EPO), human chorionic gonadotropin (HCG), and human luteinizing hormone (LH), follicle stimulating hormone remains one of the few proteins that requires in vivo potency assessment accord-ing to relevant guidelines (EP, 2011). The only approved assay for this protein is an in vivo method from 1953 relying on the ovary weight gain upon FSH injection (Steelman and Pohley, 1953). This assay exhibits severe limitations, including the large number of rats used as well as issues of accuracy, precision, sample logistics, number of parallel analyses, and cumbersome interpretation. Furthermore, the all-species-embracing validity of the Steelman-Pohley assay was questioned due to differences observed in pharmacokinetic behavior in mammals other than the rat, thus raising questions regarding the possible prediction of the FSH bioactivity in species other than the rat (de Leeuw et al., 1996).
therefore, it is not surprising that several approaches have been undertaken to circumvent this cumbersome in vivo as-say. Most attempts harness physico-chemical methods such as size-exclusion chromatography, isoelectric focusing, or, lately, reversed phase chromatography (Driebergen and Baer, 2003; Mulders et al., 1997; Almeida et al., 2011). None of them is able to reflect activity, and they are built on extremely complex cor-relation studies of dozens of batches versus the in vivo assay. Be-sides this lack of reflecting activity, depiction of those assays is highly product- and process-specific, and it does not allow cross-interpretation with respect to analysis of other FSH products.
the in vitro bioassay presented here is able to overcome all previously described drawbacks of the in vivo assay and its physico-chemical alternatives. In contrast to recombinant ani-
-3 -2 -1 0 1 20.0
0.1
0.2
0.3
0.4
0.5
0.6 Gonapure
abs(
450
nm)
log(concentration) [ng/mL]
Fig. 12: 4-parameter logistic curve fits for Gonapure®
Tab. 3: Specific activity of two FSH batches analyzed in the in vivo and in vitro assay
Batch 1 Batch 2
Mean Steelman-Pohley [IU/µg] 13.8 13.2
SD [IU/µg] 1.5 1.8
Mean FSH CBA [IU/µg] 11.9 13.8
SD [IU/µg] 1.9 2.2
RB [%] 14% -5%

Rohde et al.
Altex Proceedings, 1/12, Proceedings of WC8 99
Driebergen, R. and Baer, G. (2003). Quantification of follicle stimulating hormone (follitropin alfa): Is in vivo bioassay still relevant in the recombinant age? Curr. Med. Res. Opin. 19, 41-46.
EC (2003). Directive 2003/15/EC of the European Parliament and of the Council of 27 February 2003 amending Council Directive 76/768/EEC on the approximation of the laws of the Member States relating to cosmetic products. Official J. EU L66, 26-35.
EP – European Pharmacopoeia (2011). Urophollitropin monog-raphy 0958, version 7.0, 4711.
Gervais, A. Hammel, Y., Pelloux, S., et al. (2003). Glycosyla-tion of human recombinant gonadotrophins: characterization and batch-to-batch consistency. Glycobiology 13, 179-189.
Gleicher, N., Vietzke, M., and Vidali, A. (2003). Bye-Bye gona-dotropins? – Recombinant FSH: A real progress in ovulation induction and IVF? Hum. Reprod. 18, 476-482.
Hartung, T., Balls, M., Bardouille, C., et al. (2002). Good Cell Culture Practice. ECVAM Good Cell Culture Practice Task Force Report 1. ATLA 30, 407-414.
Havelock, J. C., Rainey, W. E., and Carr, B. R. (2004). Ovarian granulosa cell lines. Mol. Cell. Endocrinol. 228, 67-78.
Meager, A. (2006). Measurement of cytokines by bioassays: theory and application. Methods 38, 237-252.
Mire-Sluis, A.R. (1999). The development of non-animal-based bioassays for cytokines and growth factors. Dev. Biol. Stand. 101, 169-175.
Mulders, J. W. M., Derksen, M., Swolfs, A., and Maris, F. (1997). Prediction of the in vivo biological activity of human recombinant follicle stimulating hormone using quantitative isoelectric focusing. Biologicals 25, 269-281.
Olijve, W., de Boer, W., Mulder, J. W. M., and van Wezenbeek, P. M. G. F. (1996). Molecular biology and biochemistry of human recombinant follicle stimulating hormone (Puregon). Mol. Hum. Reprod. 2, 371-382.
Parakh, T. N., Hernandez, J. A., Grammer, J. C., et al. (2006). Follicle-stimulating hormone/cAMP regulation of aromatase gene expression requires beta-catenin. Proc. Natl. Acad. Sci. USA 103, 12435-12440.
Rabouhans, M. L. (1986). Reduction of animal usage: British Phar-macopoeia Commission policy. Dev. Biol. Stand. 64, 11-16.
Steelman, S. L. and Pohley, F. M. (1953). Assay of the follicle stimulating hormone based on the augmentation with human chorionic gonadotropin. Endocrinoloy 53, 604-616.
USP – United States Pharmacopeia (2004). United States Phar-macopeia Convention (USP 28-NF), 2274-2286.
Correspondence toChristoph Giese, PhDCell and tissue ServicesGoethestr. 5413086 BerlinGermanyPhone: +49 30 924006 24e-mail: [email protected]
in vitro potency tests now are specified in the European Pharma-copoeia (e.g., Chapter 5.6 “Assays of Interferons”). However, the validation of in vitro alternatives for glycoproteins such as FSH is considered challenging due to its heterogenous structure (Bristow and Charlton, 2002). The degree of essential sializa-tion of the proteins is influenced by the cultivation conditions, resulting in the formation of various isoforms (Olijve et al., 1996; Gervais et al., 2003). These isoforms differ in potency and pharmacokinetics, e.g., clearance rates (D’Antonio et al., 1999). The cell-based potency assay presented here accommo-dates these structural diversities. The KGN granulosa cell line used represents a most natural substrate for modeling physi-ological effects of FSH in vitro, exhibiting the original receptor on its cell surface. the steroid production, as a natural response to FSH exposure, further reflects the similarity to the human organism and the designated therapeutic treatment.
We believe that the herein described new in vitro bioassay for FSH with all its advantages is a valid candidate to com-plement and eventually substitute the in vivo assay for FSH potency testing.
ReferencesAlmeida, B. E., Oliveira, J. E., Damiani, R., et al. (2011). A
pilot study on potency determination of human follicle stimu-lating hormone: A comparison between reversed-phase high-performance liquid chromatography method and the in vivo bioassay. J. Pharm. Biomed. Anal. 54, 681-686.
Baenziger, J. U. and Green, E. D. (1988). Pituitary glycoprotein hormone oligosaccharides: structure, synthesis and function of the asparagine-linked oligosaccharides on lutropin, follitro-pin and thyrotropin. Biochim. Biophys. Acta 947, 287-306.
Balls, M. (2009). The three Rs and the humanity criterion. An abridged version of “The principles of humane experimental techniques” by W. M. S. Russel and R. L. Burch. Nottingham, UK: Frame.
Bristow, A. F. and Charton, E. (2002). Progress and problems in the replacement of in vivo pharmacopoeial bioassays for hormones. Dev. Biol. 111, 79-88.
Burns, C., Tiplady, R., Moore, M., and Rigsby, P. (2010). WHO International Collaborative Study of the proposed 2nd Inter-national Standard for Follicle-Stimulating Hormone, human, recombinant, for bioassay. WHO Press, Geneva, Switzerland.
Coecke, S., Balls, M., Bowe, G., et al. (2005). Guidance on good cell culture practice. A report of the second ECVAM Task Force on Good Cell Culture Practice. ATLA 33, 261-287.
Czernobilsky, B., Moll, R., Leppien, G., et al. (1987). Desmo-somal plaque-associated vimentin filaments in human ovarian granulosa cell tumors of various histologic patterns. Am. J. Pathol. 126, 476-86.
D’Antonio, M., Borrelli, F., Datola, A., et al. (1999). Biological characterization of recombinant human follicle stimulating hormone isoforms. Hum. Reprod. 14, 1160-1167.
de Leeuw, R., Mulders, J., Voortman, G., et al. (1996). Struc-ture-function relationship of recombinant follicle stimulating hormone (Puregon). Mol. Hum. Reprod. 2, 361-369.