differentiation and mineralization in chick chondrocytes maintained in a high cell density culture:...
TRANSCRIPT

In Vitro Cell. Dev. Biol. 31:288-294, April 1995 © 1995 Society for In Vitro Biology 1071-2690/95 $02.50+0.00
DIFFERENTIATION AND MINERALIZATION IN CHICK MAINTAINED IN A HIGH CELL DENSITY CULTURE:
ENDOCHONDRAL OSSIFICATION
CHONDROCYTES A MODEL FOR
COLIN FARQUHARSON A~D COLIN C. WHITEHEAD
Rostin Institute (Edinburgh), Roslin, Midlothian, EH25 9PS, Scotland, United Kingdom
(Received 21 April 1994; accepted 14 September 1994)
SUMMARY
Chondrocytes isolated from the proliferative and differentiating zones of 3-wk-old chick growth plates were cultured in the presence of 10% fetal bovine serum (FBS) and ascorbic acid for up to 21 d in a high cell density culture within Eppendorf tubes. The proliferative, differentiating, and calcification properties of the chondrocytes were examined by immunolocalization and by enzyme histochemical and biochemical methods. The cells maintained a chondrocyte pheno- type throughout culture: they were round in shape and synthesized both collagen type II and proteoglycans. The expres- sion of a hypertrophic phenotype was evident by Day 3 of culture and from this time onwards characteristics of terminal differentiation were observed. The cells were positive for both alkaline phosphatase (ALP) activity and c-myc protein and the surrounding matrix stained strongly for collagen type X. Small foci of mineralization associated with individual chondrocytes were first evident by Day 6 and more widespread areas of mineralization occupying large areas of matrix were present by Day 15. Mineralization occurred without the addition of exogenous phosphate to the medium. This culture system displays characteristics that are similar in both morphological and developmental terms to that of chick chondro- cyte differentiation and calcification in vivo and therefore offers an excellent in vitro model for endochondral ossification.
Key words: chondrocytes; high cell density culture; differentiation; cartilage; endochondral ossification.
INTRODUCTION
The epiphyseal growth plate of long bones is a specialized hyaline cartilage whose primary role is to regulate longitudinal bone growth through the process of endochondral ossification (36). The growth plate comprises both ehondrocytes and their extracellular matrix where proteoglycans and collagen type II predominate (16,23,38). The chondrocytes go through a well-defined and highly coordinated series of changes characterized by alterations in their proliferation rate, size and shape, and synthesis and deposition of extracellular matrix components (25). These events lead to a differentiated phe- notype and the synthesis by the hypertrophic chondrocytes of a matrix that mineralizes in the lower hypertrophic zone and through time is replaced by bone through the actions of the osteoclast and osteoblast (25,36).
The morphological changes associated with chondrocyte hyper- trophy are well defined and maturation results in the chondrocytes becoming more voluminous and spherical (9). Other phenotypic characteristics associated with chondrocyte maturation are the ex- pression of high levels of alkaline phosphatase (ALP) activity (41) and the synthesis of type X collagen (42), transforming growth factor-/~ (53), c-myc protein (15), and 1,25-dihydroxycholecalci- ferol [1,25(OH)2Dz] receptors (48). The exact role of these proteins in chondrocyte differentiation and mineralization remains uncertain although high ALP activity and type X collagen expression appear to be prerequisite for mineralization (42).
The regulation and coordination of the mechanisms involved in
the transition of ceils from a proliferative to a differentiating state are not fully understood. Most of the information available has been obtained from chondrocytes isolated from bovine (31,34), chick (34,40), rodent (27,43), or human (12,30) cartilage and cultured in monolayer. Under these conditions, chondrocytes have an unsta- ble phenotype and a tendency to dedifferentiate into a fibroblastic phenotype with the expression of type I collagen (37). In addition, monolayer cultures cannot mimic the spatial relationships that exist between the matrix and the chondrocytes in vivo, and in many miner- alizing cell culture systems calcification only occurs upon the addi- tion of supraphysiological concentrations of phosphate (20,34,50) resulting in a nonbiological deposition of mineral (49).
In order to study events associated with chondrocyte prolifera- tion, differentiation, and mineralization, a number of chondrocyte culturing systems have been developed to avoid the problems asso- ciated with chondrocyte monolayer cultures. A widely used method is the use of suspension cultures in which cell-to-surface attach- ments are prevented and the rounded phenotype is maintained. Such systems have employed culturing chondrocytes over agarose (10), within agarose (6), collagen gels (21), methyl cellulose (24), or alginate spheres (22). Chondrocyte aggregates that do not re- quire preformed gels of collagen or agarose to prevent monolayer formation have been previously described but at present are not fully characterized or widely used (29,39). Under these conditions mammalian chondrocytes first proliferate, then differentiate, and finally mineralization of the matrix takes place without the need for the addition of exogenous phosphate.
288

CHICK CHONDROCYTE DIFFERENTIATION 289
Chick chondrocytes are extensively used in in vitro exper iments (20 ,32 ,34 ,40 ) and are known to grow and behave differently in certain aspects from those obtained from mammal ian sources (30). It was, therefore, the purpose of this present study to examine the proliferative, differentiating, and calcification propert ies of chick chondrocytes grown in high cell density cultures in an at tempt to assess the suitability of this culture system for avian chondrocyte studies.
cedure of Farndale et al. (14). Chondroitin 4-sulphate (Sigma) was used as standard. Hydroxyproline concentrations were determined after hydrolysis of the pellet in 500 ttl of 6 M HC1 at 107 ° C for 18 h after which, the acid was removed by evaporation in vacuo. After resuspension in water, an aliquot was assayed for hydroxyproline (18) and the values obtained were used to calculate the total amount of collagen in each chondrocyte pellet.
Histological Procedures
MATERIALS AND METHODS
Isolation of Chondrocytes
Growth plates from proximal tibiotarsi of 3-wk-old chicks (male: Ross I strain) were dissected into the proliferating (upper two-thirds) and differen- tiating (bottom one-third) zones (4) and diced into small 2 - 4 mm cubes. The chondrocytes were isolated from their surrounding matrix by digestion in 0.1% collagenase (collagenase A, Boehringer Mannheim Co., Lewes, Sussex, England) in Dulbecco's modified Eagle's medium (DMEM; GIBCO, Paisley, Scotland) containing 10% fetal bovine serum (FBS; GIBCO), gluta- mine, and gentamycin for 18 h at 37t' C. Undigested material was removed by passing the digestion medium through a sieve (45 #m sieve size) and the resultant cell suspension was washed three times with DMEM and finally suspended in complete medium (DMEM containing 10% FBS, gentamycin, glutamine, and ascorbic acid at a concentration 50/~g/ml). Cell viability was assessed by trypan blue staining and was always greater than 95.0%.
One ml of complete medium containing 80 × 103 cells was added to an Eppendorf tube and centrifuged at 500 g for 5 rain to form a small clump of ceils at the bottom of the tube. The ceils were cultured at 37 ° C under an atmosphere of 95% air /5% CO 2 and the medium was changed every sec- ond day. Holes were made in the Eppendorf top with a sterile needle to aid gaseous exchange. In the first experiment, cultures containing proliferating or differentiating chondrocytes were maintained for 3, 6, 10, 15, and 21 d. In the second experiment cells isolated, as above, from the whole growth plate were cultured for 1, 2, 3, 6, and 9 d in order to investigate the initial events in matrix synthesis. In both experiments aliquots of cells (80 × 10 z) that had not been subjected to culturing conditions were also taken for biochemical analysis on Day 0. Triplicate cultures were set up to analyze each parameter at all of the above time points.
Analytical Procedures
Pellets of cartilagelike tissue from cultures were washed twice with phos- phate-buffered saline (PBS) and then homogenized at 4 ° C in 200 ttl of 0.9% NaC1, 0.2% Triton X-100. An aliquot (100 ,ul) was removed and centrifuged at 12 000 g for 15 min and the supernatant was assayed for ALP activity. The remaining 100/zl was assayed for both cellular protein and DNA content.
Measurement of ALP activity. Aliquots were assayed for ALP activity by measuring the cleavage of 10 mM p-nitrophenol phosphate (pNPP) in 1 M diethanolamine buffer (pH 9.8) and 0.2 mM MgCI2 at 37 ° C. The ALP activity was determined spectrophotometrically at 410 nm by comparison with standard solutions of p-nitrophenol. Total pellet ALP activity was cal- culated and expressed as nmoles pNPP/hydrolyzed/min/mg protein.
Determination of cellular protein and DNA. Proteins were measured using Bio-Rad protein assay reagent based on Bradford dye binding proce- dure (7) and gamma-globulin as standard. DNA concentration was deter- mined using the method of Brunk et al. (8), using the fluorescent enhance- ment of bisbenzimidazole (Hoechst 33258: Sigma Chemical Co., St. Louis, MO) complexed with DNA. Fluorescence was measured using a Hoefer TKO100 fluorimeter.
Determination of glycosaminoglycan and hydroxyproline concentra- tion. Pellets were digested in 100 ~ul of sodium acetate buffer (100 mM pH 6.0) containing papain (1.06 U/ml), 50 mM EDTA, and 10 mM cys- teine HCI for 18 h at 60 ° C. The enzyme was inactivated by heating to 100 ° C for 30 min. This procedure resulted in complete digestion of the pellet. Sulphated glycosaminoglycans were assayed using 1,9-dimethylmethylene blue (Aldrich Chemical Co., Poole, Dorset, England) according to the pro-
Pellets were rinsed in PBS and frozen attached to cork using OCT com- pound. The tissue was chilled to - 7 0 ° C in n-hexane (17) prior to cutting frozen sections of 10 gm thickness on a cryostat (Brights Instruments, Huntingdon, England.) The sections were either stained by hematoxylin and eosin for general morphology, safranin O for proteoglyeans, or yon Kossa for sites of mineralization. Other sections were prepared for enzyme- and immuno-histochemistry.
Alkaline phosphatase histochemistry. Sections were reacted for 2 min at 37 ° C in the following medium: 2 mM napthyl acid phosphate, magnesium chloride (2 mM) and Fast Blue RR (1 mg/ml) in 0.1 M barbitone buffer, pH 9.4. Sections were rinsed in 0.1 M acetic acid, washed in distilled water, and mounted in aqueous mountant (15,17).
Immunolocalization. Collagen types II and X and c-myc protein were localized in cryosections of the pellets using the following indirect peroxi- dase staining procedures.
1. Collagen type X. Sections were fixed in ice cold acetone for 5 min and air dried. The sections were pretreated with hyaluronidase (1 mg/ml; Sigma type l-S) in 0.15 M NaCI: 0.15 M sodium acetate, pH 7.2 for 1 h at 37 ° C in order to unmask epitopes (47). After blocking endogenous peroxidases by treatment with 3% H202 in methanol for 30 rain, the sections were rinsed in PBS and incubated in normal goat serum (diluted 1:5). A primary monoclonal antibody (MA3) to chick collagen type X diluted to 10 ttg/ml with PBS was added to the sections and left at 4 ° C for 18 h. After extensive washing in PBS, the tissue was incubated with peroxidase labeled goat anti-mouse IgG (Sigma) for 1 h, washed in PBS, and incubated in diamino- benzidine/HzO2 for 5 min. The tissue was finally dehydrated, cleared and mounted in DePeX.
2. Collagen type II. A primary monoclonal antibody (II-IIsB3) (35) to chick collagen type II (Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA) diluted to 2 #g IgG/ml was used in conjunction with an indirect peroxidase method similar to that described before to localize collagen type X.
3. C-mycprotein. Localizationwasaccomplisbedusingac-mycmonoclo- nal antibody (Cambridge Research Biochemicals, Cheshire, England) raised to a synthetic peptide consisting of residues 171-188 of the human p62 c-myc sequence. An indirect peroxidase method was used as has been previously described in detail (15). Suitable control procedures were per- formed in conjunction with all the above immunostaining protocols. This consisted of the substitution of the primary antibody with an appropriate dilution of normal mouse serum.
RESULTS
The first part of this study was designed to investigate the prolifer- ative and differentiating propert ies of the culture system. Similar results were obtained at any one time point from chondrocytes iso- lated from either the proliferating or the more mature differentiated zones of the growth plate. Therefore, in order to show the overall t rends in DNA and protein concentra t ions and ALP activity with t ime in culture, the data obtained from each of the two cell popula- tions were combined (n = 6). In addition, all microscopical results shown are representat ive of all cultures at any one particular t ime
point. Gross morphology. At Day 0, the cells formed a very small
loose aggregate a t tached to the side of the Eppendorf . By Day 3, the cell mass had increased in size and formed into a distinct pellet that could easily be picked up for histological processing. At the end of the culture period, the chondrocyte pellets had reached approxi-

290 FARQUHARSON AND WHITEHEAD
mately 2 mm in diameter. Without ascorbic acid, a compact chon- drocyte pellet was not formed.
Microscopical staining. From Day 3 onwards, most of the cells had the appearance of hypertrophic chondrocytes with the cells becoming more voluminous and spherical and containing an eccen- trically situated nucleus (Figs. 1 a and 1 b). At this stage, and at time points thereafter, the chondrocytes stained strongly for c-myc protein (Fig. 1 c). The newly synthesized matrix stained strongly with safranin O (Fig. 1 d) indicating the presence of proteoglycans. The matrix also stained positively with antibodies to type II and X collagen (Figs. le and 1 y'). Control sections reacted with normal mouse serum instead of the primary antibodies to collagen types II and X and c-myc protein displayed no significant staining. These findings agree with previous studies (15,35,47). Areas of mineral- ization were first apparent by Day 6 where they occupied small foci closely associated with individual chondrocytes (Fig. 1 g). In older culture (Day 15), mineralization sites were more widespread occu- pying large areas of matrix (Fig. 1 h).
DNA and cellular protein. DNA and cellular protein levels of Day 3 cultures were approximately 70% and 60% higher, respec- tively, than in Day 0 uncultured cells (80 000 cells) and thereafter remained relatively constant throughout the remainder of the cul- ture period (Figs. 2 a and 2 b).
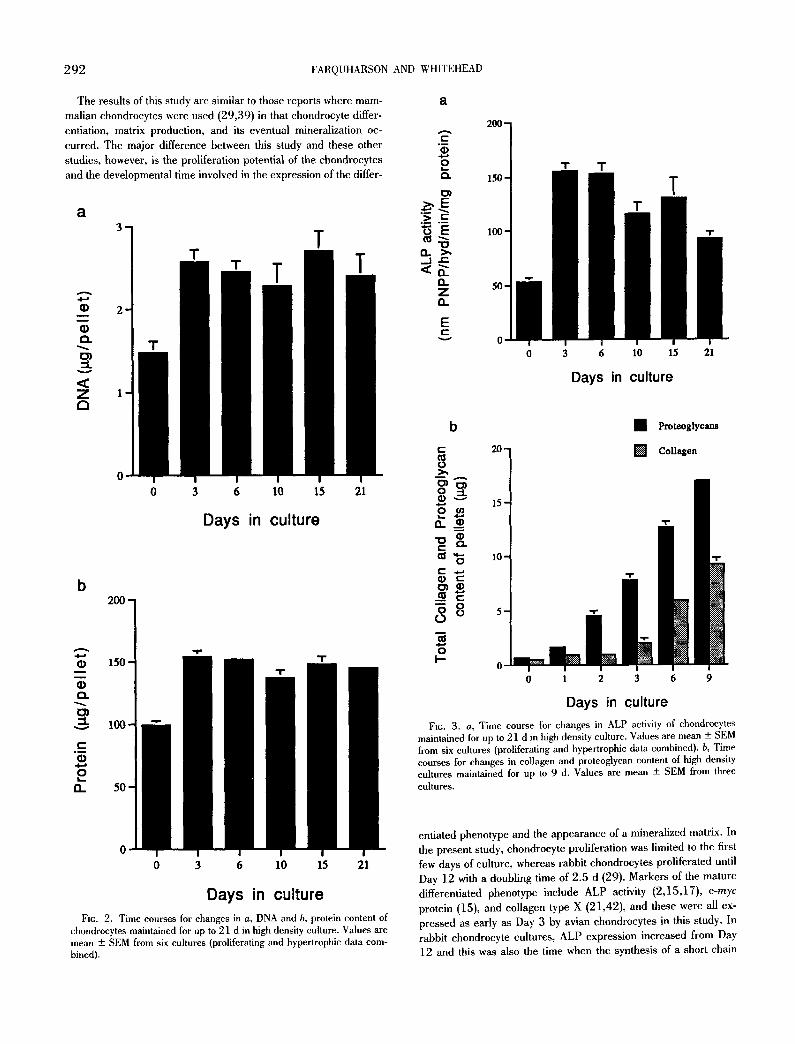
ALP activity. ALP activity was approximately 150% higher by Day 3 in comparison to Day 0 uncultured cells. This activity re- mained high at 6 d but thereafter declined with time (Fig. 3 a). This was also observed histochemically with more chondrocytes reacting intensely at Day 3 (Fig. 4 a) than at Day 21 (Fig. 4 b).
Proteoglycans and collagen. Matrix synthesis by the pelleted chondrocytes was detected at all time points, which agrees with the microscopy results. Both matrix components increased in amount with time in culture with the concentration of proteoglycans greater than that of collagen at all time points (Fig. 3 b).
DISCUSSION
It has long been recognized that chondrocytes grown in suspen- sion culture can maintain the major elements of the differentiated phenotype longer than those in monolayers (5). Suspension cultures have been used successfully by many workers (6,21,22,24) and under such conditions are capable of maintaining their rounded cell-shape and normal phenotype. As the supporting agarose slows the diffusion of newly synthesized matrix, the matrix components accumulate close to the cell surface resulting in the formation of aggregates of chondrocytes with a well-organized pericellular matrix similar to the in vivo organization (3). Inhibition of matrix diffusion also occurs in the high cell density cultures and results in the mainte- nance of the rounded phenotype, formation of a pericellular matrix, and the expression of differentiated characteristics, such as type II and X collagen, c-myc protein, and ALP activity.
The culture system described in this present study involved the formation of a clump of cells that were able to develop in close proximity to each other without adherence to the plastic surface. This system has several advantages over conventional suspension cultures. First, there is no requirement for preformed gels of colla- gen or agarose and, second, mineralization can take place without the addition of ~-glycerophosphate, which is necessary for the initia- tion of mineralization in collagen or agarose suspension cultures (50). In high density cultures, the intimate interactions and the
maintenance of spatial relationships between neighboring chondro- cytes and their matrix, which are known to influence the character- istics of chondrocytes (46), may be responsible for creating the ideal conditions for mineralization to occur. Terminal differentiation of the chondrocytes is regarded as a prerequisite for mineralization to proceed (27). Jikko et al. (27) recently demonstrated that chon- drocytes cultured in monolayer on type II collagen-coated surfaces are capable of mineralization. This provides further evidence that intimate matrix-cell interactions are necessary for mineralization, although in this case mineralization was not initiated until Day 20 of culture.
The necessity for ascorbic acid in the formation of high density aggregates is in accord with this vitamin's role as a cofactor for enzymatic hydroxylation of proline and lysine residues necessary for triple helical formation and secretion of collagen from the cells (11). Studies have indicated that the synthesis and deposition of a normal extracellular matrix and elevated ALP activity, which is me- diated by ascorbic acid, is essential for avian chondrocyte differen- tiation and matrix calcification (20,33,50). It is of interest that the decrease in ALP activity with increasing age of culture noted in this study may be a consequence of a reduction in its catalytic activity by the mineralization process (19). In contrast to this observation, hu- man chondrocyte development is inhibited by ascorbic acid, and hypertrophy and calcification of their matrix occurs in the absence of this vitamin (30). This species difference may be in part the result of increased susceptibility of human chondrocytes to toxic oxygen radicals of which ascorbate is a source (54).
Recent reports indicate that c-myc is essential in apoptosis (13,44) and it has been suggested that growth plate mineralization involves this process through the generation of matrix vesicles (1,28). The c-myc protein observed in the high cell density cultures and also in the hypertrophic region of mineralizing growth plates may be responsible for the generation of matrix vesicles. Such vesi- cles have been noted in mammalian high cell density cultures (29,39). This may explain the limited c-myc expression noted in the hypertrophic chondrocytes of Day 18 chick embryos (26) where little, if any, mineralization would be taking place.
Although the nature of the mineral formed in the cultures of this present study was not examined, it is unlikely that it was a conse- quence of cell necrosis as no evidence for the latter was observed. Others have also indicated that in mammalian high cell density cultures calcification was not secondary to cell death and the min- eral formed was composed of hydroxyapatite (29,39).
From previous studies (4), it was expected that chondrocytes isolated from different zones of the growth plate would, in culture, express phenotypic characteristics typical of the developmental stage at which they were isolated. This was not the case as all high density cultures expressed differentiated characteristics as early as Day 3 and calcification by Day 6. This is in contrast to the findings of Bashey et al. (4) where only monolayers of hypertrophic but not proliferative chondrocytes expressed type X collagen. This anomaly may be caused by the different culture systems used, with only the high density aggregates allowing maturation of the chondrocytes to their ultimate developmental fate. Alternatively, due to a combina- tion of the overnight digestion with collagenase to isolate the chon- drocytes and their fast turnover rate, the cells from the two pools may be at a similar maturational age at the start of the culture (Day 0).

CHICK CHONDROCYTE DIFFERENTIATION 291
FIe. 1. Sections of high density cultures stained with: a, hematoxylin and eosin, X162;b, hematoxylin and eosin, X642 (both 21-d cultures); c, antibodies to c-myc protein, X642 (3-d culture); d, safranin O, X642 (3-d culture); e, antibodies to type II collagen, X642 (3-d culture); f, antibodies to type X collagen, X1028 6-d culture); g, yon Kossa, X642 (6-d culture); h, von Kossa, X642 (15-d culture). Note, hypertrophic phenotype with eccentrically situated nucleus (arrows) in b, positive nuclear staining (arrows) in c, matrix staining (*) surrounding hypertrophic chondrocytes in d,e, andfand areas of mineralization (arrows) in g and h. a,b,e and f a r e cultures of chondrocytes isolated from the proliferating zone of the growth plate, whereas c,d,g and h are of chondrocytes isolated from the differentiating zone.
292 FARQUHARSON AND WHITEHEAD
The results of this study are similar to those reports where mam- malian chondrocytes were used (29,39) in that chondrocyte differ- entiation, matrix production, and its eventual mineralization oc- curred. The major difference between this study and these other studies, however, is the proliferation potential of the chondrocytes and the developmental time involved in the expression of the differ-
a
O
O
v
< Z a
3q m
0 0 3 6 10 15 21
Days in culture
b
• 1 m
O t-s
t - ° ~
O O
12_
"E" ° ~
O 4 . . t
o ~t.
.~,E ° ~
.> ._=
"U n >~ _ j r -
12. 13_ z n
E t,-
v
200-
150 -
100-
50-
0- 0 3 6 10 15 21
Days in culture
t" 20- O~ 0
o .-, 15- e ~
"~ 10-
"68 5- o
O J--
• Proteoglycans
l ~ Collagen
"11=
. LLN 0 1 2 3 6 9
Days in culture
Fro. 3. a, Time course for changes in ALP activity of chondrocytes maintained for up to 21 d in high density culture. Values are mean + SEM from six cultures (proliferating and hypertrophic data combined), b, Time courses for changes in collagen and proteoglycan content of high density cultures maintained for up to 9 d. Values are mean + SEM from three cultures,
0 3 6 10 15 21
Days in culture FIG. 2. Time courses for changes in a, DNA and b, protein content of
chondrocytes maintained for up to 21 d in high density culture. Values are mean -+ SEM from six cultures (proliferating and hypertrophic data com- bined).
entiated phenotype and the appearance of a mineralized matrix. In the present study, chondrocyte proliferation was limited to the first few days of culture, whereas rabbit chondrocytes proliferated until Day 12 with a doubling time of 2.5 d (29). Markers of the mature differentiated phenotype include ALP activity (2,15,17), c-myc
protein (15), and collagen type X (21,42), and these were all ex- pressed as early as Day 3 by avian chondrocytes in this study. In rabbit chondrocyte cultures, ALP expression increased from Day 12 and this was also the time when the synthesis of a short chain

CHICK CHONDROCYTE DIFFERENTIATION 293
the gift of the collagen type X monoclonal antibody from Dr. Alvin Kwan, University of Manchester, Manchester, England. The type II collagen mono- clonal antibody developed by Professor T. F. Linsenmayer was obtained from the Developmental Studies Hybridoma Bank maintained by the Depart- ment of Pharmacology and Molecular Sciences, Johns Hopkins University School of Medicine, Baltimore, MD, and the Department of Biological Sciences, University of Iowa, Iowa City, IA, under contract N01-HD-2- 3144 from the NICHD.
FlG. 4. Sections of high density suspension cultures reacted for ALP activity, a, 3-d culture )<642; b, 21-d culture, )<642. Both micrographs are of chondrocytes isolated from the proliferating zone of the growth plate.
collagen (presumed to be collagen type X) was first detected, Matrix synthesis by the avian chondrocytes was considerable and the first signs of its mineralization was detected histologically at Day 6, with more extensive areas of mineralization noted at Day 15. Mamma- lian chondrocyte matrix was sparse up to 7 d in culture (39) but had developed extensively by Day 16 (29). Mineralization was first noted by Day 15 (29) and 29 (39) in high density mammalian culture. It is possible to speculate that the differences in the develop- mental time between the species may be the result of built-in genetic control of the chondrocytes. The longitudinal growth rate of the chick tibiotarsi is approximately four- and fivefold greater than that of the rabbit and rat, respectively (45,51,52). The rapid movement of chondrocytes (approx. 21 h: 52) through the chick growth plate is in line with the development rate of the high density cultures of this present study.
In conclusion, high cell density chondrocyte cultures described in this study display characteristics that are morphologically similar to those of chondrocyte maturation and differentiation in vivo. Al- though cell proliferation is limited, recognized markers of terminal differentiation (ALP activity, c-myc protein, and collagen type X) are expressed early in culture. In addition, the abundant matrix synthesized by this system shows evidence of mineralization. This culture system, which is suitable for both biochemical and morpho- logical studies, offers an excellent in vitro model for endochondral ossification.
ACKNOWLEDGMENTS
This study was funded by the Ministry of Agriculture, Fisheries and Food and CEC Directorate-General for Agriculture. The authors are grateful for
REFERENCES
1. Akiska, T.; Gay, C. V. The plasma membrane and matrix vesicles of the mouse growth plate chondrocytes during differentiation as re- vealed in freeze fracture replicas. Am. J. Anat. 173:269-286; 1985.
2. All, S. Y.; Sajdera, S. W.; Anderson, H. C. Isolation and characteriza- tion of calcifying matrix vesicles from epiphyseal cartilage. Proc. Natl. Acad. Sci. USA 67:1513-1520; 1970.
3. Aydelotte, M. B.; Kuettner, K. E. Differences between sub-populations of cultured articular chondrocytes. 1. Morphology and cartilage ma- trix production. Connect Tissue Res. 18:205-222; 1988.
4. Bashey, R. I.; Iannotti, J. P.; Ran, V. H., et al. Comparison of morpho- logical and biochemical characteristics of cultured chondrocytes iso- lated from proliferative and hypertrophic zones of bovine growth plate cartilage. Differentiation 46:199-207; 1991.
5. Benya, P. D. Introduction and survey of techniques for chondrocyte culture. In: Maroudas, A.; Kuettncr, K., eds. Methods in cartilage research. London: Academic Press; 1990:85-89.
6. Benya, P. D.; Shaffer, J. D. Dedifferentiated chondrocytes reexpress the differentiated collagen phenotype when cultured in agarose gels. Cell 30:215-224; 1982.
7. Bradford, M. M. Rapid and sensitive method for quantification of mi- crogram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254; 1976.
8. Brunk, C. F.; Jones, K. C.; James, T. W. Assay for nanogram quanti- ties of DNA in cellular homogenates. Anal. Biochem. 92:497-500; 1979.
9. Buckwalter, J. A.; Mower, D.; Ungar, J., et at. Morphometric analysis of chondrocyte hypertrophy. J. Bone J. Surg. Am. Vol. 68A:243- 255; 1986.
10. Castagnola, P.; Dozin, B.; Morn, G., et at. Changes in the expression of collagen genes show two stages in chondrocyte differentiation in vitro. J. Cell Biol. 106:461-467; 1988.
11. Chan, D.; Lamande, S. R.; Cole, W,, et at. Regulation of procollagen synthesis and processing during ascorbate-induced extracellular matrix accumulation in vitro. Biochem. J. 269:175-181; 1990.
12. Elima, K.; Vuorio, E. Expression of mRNAs for collagen and other matrix components in dcdifferentiating and redifferentiating human condrocytes in culture. FEBS Lett. 258:195-198; 1989,
13. Evan, G.; Wyllie, A. H.; Gilbert, C. S., et al. Induction of apoptosis in fibroblasts by c-myc protein. Cell 69:119-128; 1992.
14. Farndale, R. W.; Buttle, D. J.; Barrett, A. J. Improved quantification and discrimination of sulphated glycosaminoglycans by use of di- methylene blue. Biochim. Biophys. Acta 883:173-177; 1986.
15, Farquharson, C.; Hesketh, J. E,; Loveridge, N. The proto-oncogene c-myc is involved in cell differentiation as well as cell proliferation: studies on growth plate chondrocytes in situ. J. Cell Physiol. 152:135-144; 1992.
16. Farquharson, C.; Whitehead, C. C.; Loveridge, N. Alterations in glycos- aminoglycan concentration and sulfation during chondrocyte matura- tion. Calcif. Tissue Int. 54:296-303; 1994.
17. Farquharson, C.; Whitehead, C. C.; Rennie, S., et at. Cell proliferation and enzyme activities associated with the development of avian tibial dyschondrnplasia: an in situ biochemical study. Bone 13:59-67; 1992.
18. Firschein, H. E.; Schill, J. P. The determination of total hydroxypro- line in urine and bone extracts. Anal. Biochem. 14:296-304; 1987.
19. Genge, B. R.; Saner, G. R.; Licia, N. Y., etah Correlation between loss of alkaline phosphatase activity and accumulation of calcium during matrix vesicle-mediated mineralization. J. Biol. Chem. 263:18513- 18519; 1988.

294 FARQUHARSON AND WHITEHEAD
20. Gerstenfeld, L. C.; Landis, W. J. Gene expression and extracellular matrix uhrastructure of a mineralizing chondrocyte cell culture sys- tem. J. Cell Biol. 112:501-513; 1991.
21. Gibson, G. J.; Schor, S. L.; Grant, M. E. Effects of matrix macromole- cules on chondrocyte genc expression: synthesis of a low molecular weight collagen species by cells cultured within collagen gels. J. Cell Biol. 93:767-774; 1982.
22. Grandolfo, M.; D Andrea, P.; Paaletti, M., et al. Culture and differen- tiation of chondrocytes entrapped in alginate gels. Calcif. Tissue Int. 52:42-48; 1993.
23. Heinegard, D. Polydispersity of cartilage proteoglycans. J. Biol. Chem. 252:1980-1989; 1977.
24. Horton, W.; Hassel, J. R. Independence of cell shape and loss of cartilage matrix production during retinoic acid treatment of cul- tured chondrocytes. Dev. Biol. 115:392-397; 1986.
25. Hunziker, E. B.; Schenk, R. K. Physiological mechanisms adopted by chondrocytes in regulating longitudinal bone growth in rats. J. Phys- iol. 414:55-71; 1989.
26. lwamoto, M.; Yagami, K.; Lu Valle, P., et al. Expression and role of c-myc in chondrocytes undergoing endochondral ossification. J. Biol. Chem. 268:9645-9652; 1993.
27. Jikko, A.; Aoba, T.; Murakami, H., et al. Characterization of the miner- alization process in cultures of rabbit growth plate chondrocytes. Dev. Biol. 156:372-380; 1993.
28. Kardos, T. B.; Hubbard, M. J. Are matrix vesicles apoptotic bodies? In: Dixon, A. D.; Sarnat, B. G., eds. Factors and mechanisms in- fluencing bone growth. New York: Alan R. Liss; 1982:45-62.
29. Kato, Y.; lwamoto, M.; Koike, T., et al. Terminal differentiation and calcification in rabbit chondrocyte cultures grown in centrifuge tubes: regulation by transforming growth factor b and serum factors. Proc. Natl. Acad. Sci. USA 85:9552-9556; 1988.
30. Kirsch, T.; Swoboda, B.; yon der Mark, K. Ascorbate independent differentiation of human chondrocytes in vitro: simultaneous expres- sion of types I and X collagen and matrix mineralization. Differen- tiation 52:89-100; 1992.
31. Kuettner, K. E.; Pauli, B. U.; Gall, G., et al. Synthesis of cartilage matrix by mammalian chondrocytes in vitro. 1. Isolation, culture characteristics, and morphology. J. Biol. Chem. 93:743-750; 1982.
32. Leach, R. L.; Roselot, G. E. The use of avian epiphyseal chondrocytes for in vitro studies of skeletal metabolism. J. Nutr. 122:802-805; 1992.
33. Leboy, P. S.; Vaias, L.; Uschmann, B. Ascorbic acid induces alkaline phosphatase, type X collagen, and calcium deposition in cultured chick chondrocytes. J. Biol. Chem. 264:17281-17286; 1989.
34. Lian, J. B.; McKee, M. D.; Todd, A. M., et al. Induction of bone-re- lated proteins, osteocalein and osteopontin, and their matrix ultra- structural localization with development of chondrocyte hypertrophy in vitro. J. Cell. Biochem. 52:206-219; 1993.
35. Linsenmayer, T. F.; Henrix, M. J. C. Monoclonal antibodies to connec- tive tissue macromolecules type II collagen. Biochem. Biophys. Res. Commun. 92:440-446; 1980.
36. Loveridge, N.; Thomson, B. M.; Farquharson, C. Bone growth and
turnover. In: Whitehead, C. C., ed. Bone biology and skeletal dis- orders in poultry. Poultry Science Symposium 23. Abingdon, En- gland: Carfax Publishing Co.; 1992:3-17.
37. Mayne, R.; Vail, M. S.; Mayne, P. M., et al. Changes in the type of collagen synthesized as clones of chick chondrocytes grow and even- tually lose division capacity. Proc. Natl. Acad. Sci. USA 73:1674- 1678; 1976.
38. Miller, E. J. Biochemical characteristics and biological significance of the genetically distinct collagens. Mot. Cell Biochem. 13:165-191; 1976.
39. Nakagawa, Y.; Shimizu, K.; Hamamoto, T., et al. Electron microscopy of calcification during high-density suspension culture of chondro- cytes. Calcif. Tissue Int. 53 :127-134; 1993.
40. O Keefe, R. J.; Rosier, R. N.; Puzas, J. E. Differential expression of effects of maturationally distinct subpopulations of growth plate cbondrocytes. Connect. Tissue Res. 24 :53-66; 1990.
41. Robinson, R. The possible significance of hexosephospheric esters in ossification in vitro. Biochem. J. 17:286-293; 1923.
42. Schmid, T. M.; Bonen, D. K.; Luchene, L., et at. Late events in chon- drocyte differentiation: hypertrophy, type X collagen synthesis and matrix calcification. In Vivo 5 :553-540; 1991.
43. Schwartz, Z.; Boyan, B. The effects of vitamin D metabolites on phos- pholipase A a activity of growth zone and resting zone cartilage cells in vitro. Endocrinology 122:2191-2198; 1988.
44. Shi, Y.; Glynn, J. M.; Guilbert, L. J., et al. Role of c-myc in activation- induced apoptotic cell death in T cell hybridomas. Science 257:212-214; 1992.
45. Sissons, H. A. Experimeutaldetermination of rate of longitudinal bone growth. J. Anat. 87 :228-237; 1953.
46. Sommarin, Y.; Larsson, T.; Heinegard, D. Chondrocyte matrix inter- actions. Exp. Cell Res. 184:181-192; 1989.
47. Stephens, M.; Kwan, A. P.; Bayliss, M. T., et al. Human articular surface chondrocytes initiate alkaline phosphatase and type X colla- gen synthesis in suspension culture. J. Cell Sci. 103:1111-1116; 1992.
48. Suda, S.; Takahashi, N.; Shinki, T., et al. 1, alpha-25-dihydroxyvita- min D 3 receptors and their action in embryonic chick chondrocytes. Calcif. Tissue Int. 37:82-90; 1985.
49. Suzuki, F.; Takase, T.; Takigawa, M., et al. Stimulation of the initial stage of endocbondral ossification: in vitro sequential culture of growth cartilage cells and bone marrow cells. Proc. Natl. Acad. Sci. USA 78:2368-2372; 1981.
50. Tacchetti, C.; Quarto, R.; Campanile, G., et at. Calcification of in vitro developed hypertrophic cartilage. Dev. Biol. 132:442-447; 1989.
51. Tapp, E. Tetracycline labelling methods of measuring the growth of bones in the rat. J. Bone J. Surg. Br. Vol. 48 :517-525; 1966.
52. Thorp, B. H. Relationship between the rate oflongitudinal bone growth and physeal thickness in the growing fowl. Res. Vet. Sci. 45:83-85; 1988.
53. Thorp, B. H.; Anderson, I.; Jakowlew, S. B. Transforming growth factor bl, -b2 and -b3 in cartilage and bone cells during endochon- dral ossification in the chick. Development 114:907-911; 1992.
54. Tschan, T.; Hoerler, I.; Houze, Y., et al. Resting chondrocytes in culture survive without growth factors, but are sensitive to oxygen metabolites. J. Cell Biol. 111:257-260; 1990.