distinct form i, ii, iii, and iv rubisco proteins from the three kingdoms
TRANSCRIPT

Journal of Experimental Botany, Vol. 59, No. 7, pp. 1515–1524, 2008
doi:10.1093/jxb/erm361 Advance Access publication 16 February, 2008
SPECIAL ISSUE REVIEW PAPER
Distinct form I, II, III, and IV Rubisco proteins from thethree kingdoms of life provide clues about Rubiscoevolution and structure/function relationships
F. Robert Tabita1,2,3,*, Sriram Satagopan2, Thomas E. Hanson4, Nathan E. Kreel3, and Stephanie S. Scott2
1 Department of Microbiology, The Ohio State University, 484 West 12th Avenue, Columbus, Ohio 43210–1292,USA2 The Plant Molecular Biology Biology/Biotechnology Program, The Ohio State University, 484 West 12th Avenue,Columbus, Ohio 43210–1292, USA3 The OSU Biochemistry Program, The Ohio State University, 484 West 12th Avenue, Columbus,Ohio 43210–1292, USA4 College of Marine and Earth Studies, Delaware Biotechnology Institute, University of Delaware,127 DBI, 15 Innovation Way, Newark, DE 19711, USA
Received 22 October 2007; Revised 29 November 2007; Accepted 18 December 2007
Abstract
There are four forms of ribulose 1,5-bisphosphate
carboxylase/oxygenase (Rubisco) found in nature.
Forms I, II, and III catalyse the carboxylation and
oxygenation of ribulose 1,5-bisphosphate, while form
IV, also called the Rubisco-like protein (RLP), does not
catalyse either of these reactions. There appear to be
six different clades of RLP. Although related to bona
fide Rubisco proteins at the primary sequence and
tertiary structure levels, RLP from two of these clades
is known to perform other functions in the cell. Forms
I, II, and III Rubisco, along with form IV (RLP), are
thought to have evolved from a primordial archaeal
Rubisco. Structure/function studies with both archaeal
form III (methanogen) and form I (cyanobacterial)
Rubisco have identified residues that appear to be
specifically involved with interactions with molecular
oxygen. A specific region of all form I, II, and III
Rubisco was identified as being important for these
interactions.
Key words: CBB cycle, different forms, evolution, Rubisco,
structure/function.
Introduction
There are four known metabolic routes by which micro-organisms and plants reduce and assimilate carbon di-oxide into organic matter (Fuchs et al., 1987; Yoon et al.,2000). Paramount among these on a global scale is theCalvin–Benson–Bassham (CBB) reductive pentose phos-phate pathway. In this scheme, the enediol form of thesugar bisphosphate ribulose 1,5-bisphosphate (RuBP)accepts a molecule of CO2, with the enzyme RuBPcarboxylase/oxygenase (Rubisco) catalysing the actualprimary CO2 fixation reaction. Rubisco is found in mostautotrophic organisms, ranging from diverse prokaryotes,including photosynthetic and chemolithoautotrophic bac-teria and archaea, to eukaryotic algae and higher plants.All bona fide Rubisco proteins must first be activated orcarbamylated at the e-amino group of a specific lysineresidue (e.g. Lys-201 of the plant enzyme), with divalentcations bound to adjacent acidic residues serving tostabilize the carbamate. This carbamylated lysine orE-CO2-Me2+ ternary complex catalyses proton abstractionfrom the substrate RuBP, thus initiating the catalyticcycle. While the basic mechanistic details underlyingcatalysis are well understood (Cleland et al., 1998), therestill remain many perplexing issues about Rubisco. Notthe least of which are the fundamental reasons whystructurally superimposable proteins from different sources,with up to 90% sequence identity, often show vastly
* To whom correspondence should be addressed at the Department of Microbiology, The Ohio State University. E-mail: [email protected]
ª The Author [2008]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

different catalytic properties and distinct kinetic behav-iour (Tabita, 1999).
The active site of all Rubisco proteins is formed by theinteraction of two monomeric catalytic subunits, with theC-terminus of one monomer binding to the N-terminus ofa second monomer in a specific manner to form two activesites per dimer. Distinct residues from both monomericunits thus comprise the active site required for bothcarboxylation of CO2 and the fixation of molecularoxygen (Fig. 1), with the two gaseous substrates clearlycompeting for the same active site. The relative rate ofcarboxylation and oxygenation of RuBP (vc/vo) thusdefines the protein’s catalytic efficiency or ability toprovide the cell with needed carbon. The latter ratio may bedetermined after specific isolation of the reaction products,3- phosphoglyceric acid (3-PGA) and 2-phosphoglycolate(2-PG) (Fig. 1), which may be easily distinguished afterperforming the enzymatic reaction with [1-3H]-RuBP.Quantitation of [3H]-PGA or [3H]-2-PG from [1-3H]-RuBP,in the presence of both CO2 and O2, is a measure of therelative activity of the carboxylase and oxygenase reac-tions, respectively, under conditions where both reactionsmay be measured simultaneously. From the foregoing, itis apparent that efficient Rubisco catalysis is dependenton the inherent ability of the enzyme to discriminatebetween CO2 and O2 (the X or s value) at the relativeconcentration of CO2 and O2 employed in a particular
reaction. The rates of the two reactions are defined byvc/vo¼X[CO2]/[O2]. Thus, X¼vc [O2]/vo [CO2] andX¼VcKo/VoKc with Vc and Vo representing maximumvelocities for carboxylation and oxygenation, respectively,and Kc and Ko the relative Michaelis constants for CO2
and O2, respectively.
The different forms of Rubisco
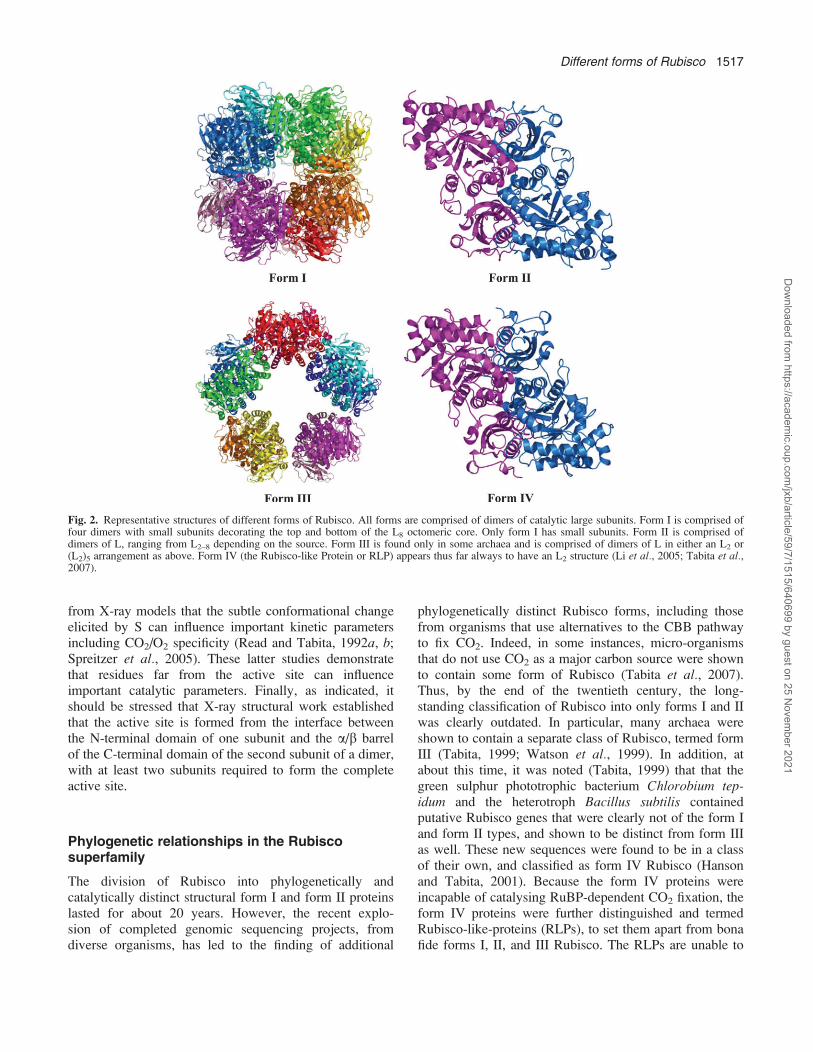
There are four known forms or types of Rubisco found innature, forms I, II, III, and IV (Tabita, 1999; Tabita et al.,2007), each of which is placed in a separate categorybased on differences in primary sequence of the constitu-ent ;50 kDa polypeptide. In addition, these four differentholoenzyme forms are often structurally unique, yet thefundamental unit common to all forms is the large(catalytic) subunit dimer. Of the four forms, form I is themost abundant. It is found in eukaryotes and bacteria, andis composed of eight large subunits and eight smallsubunits (L8S8) with 422 symmetry (Schneider et al.,1992). Many form I Rubisco structures from differentorganisms have been determined and show high similarity(Andersson and Taylor, 2003). In form I, the basicstructural motif, the dimer of L, is repeated four times toform a catalytic (L8) core of 8 L subunits, with smallsubunits on top and bottom of this core (Fig. 2).
The first Rubisco structure to be solved was that fromthe bacterium Rhodospirillum rubrum (Andersson et al.,1989; Schneider et al., 1990). This enzyme is a simpledimer of L subunits (Tabita and McFadden, 1974) thatshares only 25–30% identity to L subunits from type IRubisco. It has served for many years as the paradigm forstructure–function studies of Rubisco (Hartman andHarpel, 1994) and has provided the base-line informationrequired to determine the structure of the more complexform I enzyme. Thus, the structure of this type II or formII Rubisco closely resembles the structure of the basicdimer of the form I Rubisco despite some differences incatalytic properties. Further work in several laboratories(see Portis, 1992; Hartman and Harpel, 1994; Clelandet al., 1998; Spreitzer, 1999; Tabita, 1999; Spreitzer andSalvucci, 2002, for recent reviews), including our ownstudies, has led to: (i) the demonstration that L isdefinitely the catalytic subunit of the L8S8 enzyme andundergoes catalysis in the absence of S (Andrews, 1988;Lee and Tabita, 1990); (ii) the identification of residuesthat participate in various aspects of the catalytic mecha-nism including CO2/O2 specificity (Hartman and Harpel,1994; Cleland et al., 1998; Tabita, 1999; Spreitzer andSalvucci, 2002); (iii) the finding that effector sites are alsofound on L (Lee et al., 1991b); (iv) the finding that Saffects activity by influencing the correct conformation ofthe catalytic core of L (Andrews, 1988; Lee and Tabita,1990; Lee et al., 1991a); (v) verification of the prediction
Fig. 1. The fundamental dimeric unit of all forms of Rubisco. The basiccatalytic unit of all Rubisco molecules is a dimer of large catalyticsubunits in which each monomer interacts in a head to tail arrangementsuch that the C-terminus of one monomer interacts with the N-terminusof the second monomer. The active site thus contains residues fromprimarily the C-terminus of one monomer, but also includes residuesfrom the N-terminus of the second monomer, conferring two active sitesper dimer.
1516 Tabita et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021
from X-ray models that the subtle conformational changeelicited by S can influence important kinetic parametersincluding CO2/O2 specificity (Read and Tabita, 1992a, b;Spreitzer et al., 2005). These latter studies demonstratethat residues far from the active site can influenceimportant catalytic parameters. Finally, as indicated, itshould be stressed that X-ray structural work establishedthat the active site is formed from the interface betweenthe N-terminal domain of one subunit and the a/b barrelof the C-terminal domain of the second subunit of a dimer,with at least two subunits required to form the completeactive site.
Phylogenetic relationships in the Rubiscosuperfamily
The division of Rubisco into phylogenetically andcatalytically distinct structural form I and form II proteinslasted for about 20 years. However, the recent explo-sion of completed genomic sequencing projects, fromdiverse organisms, has led to the finding of additional
phylogenetically distinct Rubisco forms, including thosefrom organisms that use alternatives to the CBB pathwayto fix CO2. Indeed, in some instances, micro-organismsthat do not use CO2 as a major carbon source were shownto contain some form of Rubisco (Tabita et al., 2007).Thus, by the end of the twentieth century, the long-standing classification of Rubisco into only forms I and IIwas clearly outdated. In particular, many archaea wereshown to contain a separate class of Rubisco, termed formIII (Tabita, 1999; Watson et al., 1999). In addition, atabout this time, it was noted (Tabita, 1999) that that thegreen sulphur phototrophic bacterium Chlorobium tep-idum and the heterotroph Bacillus subtilis containedputative Rubisco genes that were clearly not of the form Iand form II types, and shown to be distinct from form IIIas well. These new sequences were found to be in a classof their own, and classified as form IV Rubisco (Hansonand Tabita, 2001). Because the form IV proteins wereincapable of catalysing RuBP-dependent CO2 fixation, theform IV proteins were further distinguished and termedRubisco-like-proteins (RLPs), to set them apart from bonafide forms I, II, and III Rubisco. The RLPs are unable to
Fig. 2. Representative structures of different forms of Rubisco. All forms are comprised of dimers of catalytic large subunits. Form I is comprised offour dimers with small subunits decorating the top and bottom of the L8 octomeric core. Only form I has small subunits. Form II is comprised ofdimers of L, ranging from L2–8 depending on the source. Form III is found only in some archaea and is comprised of dimers of L in either an L2 or(L2)5 arrangement as above. Form IV (the Rubisco-like Protein or RLP) appears thus far always to have an L2 structure (Li et al., 2005; Tabita et al.,2007).
Different forms of Rubisco 1517
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

catalyse CO2 fixation because of key substitutions ofmany essential active-site residues (Hanson and Tabita,2001). The basic properties of Rubiscos and RLPs aredescribed in Table 1.
With respect to the function of the RLP, an RLP geneknockout strain of C. tepidum was shown to exhibita pleiotropic phenotype, including the deposition ofelemental sulphur into the surrounding media, as well asdistinct effects on autotrophic growth (Hanson and Tabita,2001). This was shown to be due to a specific effect onthiosulphate (and not sulphide) oxidation, leading toa general stress response. The stress response in C.tepidum was manifested by synthesis of stress-relatedproteins, including a thiol-specific hydroperoxidase andsuperoxide dismutase, resulting in resistance to high levelsof hydrogen peroxide (Hanson and Tabita, 2003). In B.subtilis, the specific function of its RLP (or YkrW/MtnW)was identified where it was shown to participate ina methionine salvage pathway. In this and relatedorganisms, RLP catalyses the enolization of the RuBPanalogue, 2,3-diketo-5-methylthiopentyl-1-P (Ashidaet al., 2003). Additional structure/function studies byImker et al. (2007), with a related protein from Geo-bacillus kaustophilus, clearly illustrated the similarities ofthe enolase reactions catalysed by this RLP and Rubisco.Moreover, these authors determined the structure of theactivated enolase and identified the base that catalysesproton abstraction.
In summary, phylogenetic analyses of Rubisco and RLPamino acid sequences indicate that there are at least threedistinct lineages of bona fide Rubisco (forms I, II, and III)and six distinct clades of RLP molecules (form IV)(Fig. 3). Beyond studies with the YkrW group or‘enolase’ RLPs from B. subtilis and G. kaustophilus, andphysiological indications that the C. tepidum RLP isinvolved in some way with thiosulphate oxidation,
representative RLPs from other clades have yet to beassigned a function. Furthermore, many of the RLP genesthat encode proteins of unknown function were unablegenetically to complement strains with knockouts in genesthat encode RLP proteins of known function (J Singh, FRTabita, unpublished results), suggesting that these un-known or undefined RLP proteins catalyse differentreaction(s).
The diversity and evolution of Rubisco and itshomologue, the Rubisco-like protein
It is our contention that different molecular forms ofRubisco, from organisms that fix CO2 under highlydivergent conditions in many distinct environments, mayprovide clues relative to the molecular basis for Rubiscosubstrate specificity and other kinetic properties, whilealso shedding light as to how the active site of this proteinmay have evolved from primordial ancestors. Certainly inlight of efforts to improve the catalytic properties ofRubisco, studying its evolution ensures that one does notrepeat experiments that nature has already performed. Ina comprehensive analysis of all available microbial andplant Rubisco and RLP sequences, the different lineageswere evaluated by various methods including phylo-genetic reconstruction with multiple models on a moreextensive sequence set, structural homology searches,gene conservation (both local and genome wide), con-certed variation in active site substitution patterns, andothers (Tabita et al., 2007). In all instances, it wasconcluded that the most likely scenario was that a formIII Rubisco, arising within the Methanomicrobia, was theultimate source of all Rubisco and RLP lineages (Fig. 4).Indeed, from these analyses, the euryarchaea appear toharbour the deepest branching Rubisco and RLP
Table 1. Properties and distribution of currently defined lineages of Rubisco and RLP
Lineage Subunit composition Rubiscoactivity
Rubiscoactive site
Phylogenetic distribution
I-A L8S8 + + a, b, c-Proteobacteria, Cyanobacteria, Prochlorales, Sargasso Seametagenome, GOS metagenome
I-B L8S8 + + Cyanobacteria, Prochlorales, Eukaryotes-Viridiplantae (Streptophyta,Chlorophyta), Euglenozoa, Sargasso Sea metagenome
I-C L8S8 + + a, b -Proteobacteria, ChloroflexiI-D L8S8 + + a, b, c-Proteobacteria, Eukaryotes-stramenopiles, Rhodophyta,
HaptophyceaeII L2 and Ln + + a, b, c-Proteobacteria, Eukaryotes-Alveolata (Dinophyceae)III L2 and (L2)5 (Ln?) + + Methanogenic and thermophilic crenarchaeota, thermophilic and
halophilic euryarchaeotaIV-NonPhoto ? ? - a, b, c-Proteobacteria, ChloroflexiIV-DeepYkr ? - - a-Proteobacteria, Clostridia, Non-methanogenic euryarchaeota,
Eukaryotes-Ostreococcus tauriIV-AMC ? ? - Acid mine drainage microbial consortiumIV-GOS ? ? - GOS sequence collectionIV-Photo L2 - - a, b -Proteobacteria, ChlorobiaIV-YkrW L2 - - Firmicutes, acid mine drainage microbial consortium
1518 Tabita et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

sequences and therefore are the most likely candidates forthe evolutionary root of the Rubisco and RLP superfam-ily. We have thus proposed that a single transfer of RLPfrom a methanogenic euryarchaeon into an ancestor of theFirmicutes, Proteobacteria, and Chlorobia, with sub-sequent lateral transfer to the Chloroflexi, followed bygene losses could account for the distribution of most ofthe RLP lineages (Fig. 4). Likewise, lateral transfer ofa form III Rubisco from a euryarchaeon to a commonancestor of Cyanobacteria and Proteobacteria, witheukaryote Rubiscos being acquired via subsequent endo-symbiotic events, could account for the currently observeddistribution of bona fide Rubisco lineages. From these
considerations, the likely evolutionary development of thelarge subunit of Rubisco and RLP probably follows themodel illustrated (Fig. 4). The scenario presented here islargely an extension of prior work by Delwiche andPalmer (1996) and Tabita (1999), but incorporates formIII and form IV sequences that were not known at thetime. In addition, a significant number of Rubisco andRLP sequences were recently reported from the globalocean sampling (GOS) expedition (Yooseph et al., 2007)and the authors purport to have discovered a number ofnew sub-families of Rubisco and RLP. However, our ownexamination of the same data suggests that this claim isoverstated as will be detailed elsewhere.
Fig. 3. Phylogenetic tree illustrating the three classes of Rubisco and the six clades of RLP. For a detailed description of the different Rubisco andRLP molecules, see Tabita et al. (2007). Basic properties of each clade are presented in Table 1. The tree was constructed from a CLUSTALWalignment of a non-redundant set of Rubisco and RLP sequences culled from publicly accessible sequence databases, primarily GenBank. The non-redundant set was selected such that each sequence was no more than 76% identical at the amino acid level to any other sequence in the alignment.The tree was produced by Neighbor–Joining in the MEGA 4.0 software suite using the p-distance model and pairwise gap deletion. Bootstrap valuesare the percentage of 1000 trials in which a given node was present.
Different forms of Rubisco 1519
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

Recent structure/function studies
The archaeal form III Rubisco
It is clear from the discussion above that many distinctforms of Rubisco are found in nature and are available forstructure–function studies. In many instances, these di-verse proteins exhibit distinctive catalytic properties suchthat their study provides useful insights as to how allRubisco eenzymes function (Tabita, 1999). For example,the discovery of the form III enzymes, obtained fromorganisms that never see molecular oxygen, offer tantaliz-ing possibilities to learn more about how the active site ofRubisco might have evolved. This is especially relevantsince it was found that several archaeal enzymes arehighly sensitive to molecular oxygen and have extremelypoor capabilities to discriminate between CO2 and O2
(Watson et al., 1999; Finn and Tabita, 2003; Kreel andTabita, 2007), due in part to an extremely high affinity ofthese enzymes for O2 (Kreel and Tabita, 2007). In thisrecent study, the basis for the high affinity to O2 wassought, with the enzyme from Archaeoglobus fulgidusused as a model system. Bioinformatic and structuralcomparisons of all sequenced archaeal form III Rubiscospointed towards unique residues that were positioned tointeract with the active site. One of these residues, Met-295, was changed by site-directed mutagenesis and theresultant M295D protein was shown to recover from
oxygen exposure much more effectively than the wild-type A. fulgidus enzyme. This was consistent with anincrease in the Ko for oxygen from 5 lM in the wild-typeenzyme to about 24 lM for the M295D enzyme. Inaddition, there was an unprecedented 3-fold increase inthe substrate specificity factor (X) of the M295D enzymecompared with the wild-type protein. Structural analysesindicated that Met-295 was situated in a hydrophobicpocket created by residues along the active site and inclose proximity to a highly conserved active-site residue,Arg-279 found in all other forms of Rubisco and shown tobe necessary for substrate (RuBP) binding (Knight et al.,1990). In the wild-type A. fulgidus enzyme (RbcL2), thereis no hydrogen bond to the Arg-279 residue (Fig. 5A).However, the model structure suggests that a mutation toan aspartate residue at the Met-295 position would allowfor an ionic interaction between one of the hydroxyl sidechains of the aspartate residue with one of the side chainnitrogen atoms of Arg-279 (Fig. 5B). Interestingly, thereis definite hydrogen bonding to the equivalent Arg residuein all other form I and form II Rubisco structures; forexample, originating from the oxygen atom of thecarbonyl group of His-324 from the peptide backbone ofthe Synechococcus PCC 6301 enzyme (Kreel and Tabita,2007).
Further investigation into the above localized structuralchange led us to another amino acid, Ser-363, which was
Fig. 4. A simplified scheme for the evolution of major Rubisco/RLP lineages and their distribution by both vertical (solid arrows) and horizontal(dashed arrows) gene transfer. Also depicted are likely endosymbiotic events (dotted arrows). For a more detailed view, see Tabita et al. (2007).
1520 Tabita et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

predicted to have similar effects on oxygen sensitivity ofthe A. fulgidus enzyme. Ser-363 is situated in whatappears to be a hydrophobic pocket that surrounds oneside of the active site (Fig. 5C). In addition, the modelstructure shows an interaction of the side chain of Ser-363with two highly conserved and catalytically importantresidues, Gly-313 and Thr-314. Gly-313 and Thr-314,found in all forms of Rubisco, show no interactions withthe amino acid residue equivalent to Ser-363 of RbcL2 inform I and form II enzymes. Thus, this unique interactionand positioning of Ser-363 in a key hydrophobic pocket ofRbcL2, similar to Met-295, suggested that Ser-363 ofRbcL2 might be a likely candidate for further investiga-tion by site-directed mutagenesis. Ser-363 was thuschanged to residues found in equivalent positions in formI and form II enzymes (alanine and isoleucine, respec-tively, and valine). Much like the M295D enzymes, theS363I and S363V enzymes retained much higher levels ofactivity (about 45–50%) when exposed to oxygen com-pared to the wild-type enzyme (about 10–15% activity)(Kreel and Tabita, 2007). Moreover, double mutants(M295DS363I or M295DS363V) exhibited an apparentadditive effect and recovered nearly 90% activity afterbeing exposed to oxygen. Recent studies further indicatethat the Ko for the M295DS363I double mutant has beenraised to over 400 lM, very close to values obtained forform I and form II Rubisco proteins (KE Kreel and FRTabita, unpublished results). Clearly, the above studiespoint to the importance of hydrophobic regions forinteractions with oxygen, with the oxygen-sensitive A.fulgidus form III enzyme providing a potential templatefor examining other forms of Rubisco.
Random mutagenesis and bioselection of mutantcyanobacterial form I Rubisco
Prokaryotic bioselection after random mutagenesis hasnow become feasible and adapted for the isolation ofmutant forms of Rubisco (Smith and Tabita, 2003; Greenet al., 2007). It is convenient to use Rhodobactercapsulatus, with its endogenous form I and form IIRubisco genes deleted, as a host strain for whatever
Fig. 5. Predicted side-chain interactions with Met-295 in wild-type A.fulgidus RbcL2 (A), the mutant M295D enzyme (B), and thehydrophobic pocket surrounding Ser-363 (C). Side chains shown areamino acids Met-295 (A) and Asp-295 (B), as well as conserved aminoacids found in all other forms of Rubisco. In A. fulgidus RbcL2 and themutant M295D enzyme, His-278, Arg-279, and His-311, are illustrated
as they are necessary for catalysis and binding of the five carbonsubstrate, RuBP. The model structure predicts no ionic interactionsbetween Arg-279 and Met-295 in the wild-type form of the enzyme (A).In the M295D mutant, the model predicts an ionic interaction betweenthe hydroxyl group of the Asp-295 residue and the amino group of theArg-279 residue (dashed purple line). In (C), the ribbon structure isremoved to illustrate more clearly the formation of a hydrophobicpocket where Ser-363 resides. Ser-363 is coloured yellow and interactsto form a triad of hydrogen bonds with highly conserved yet non-catalytically important residues Gly-313 and Thr-314, coloured blue.Highly conserved and catalytically important residues Ser-364 and Gly-366, which are necessary for the binding of the five carbon substrate,RuBP, are coloured white. All other hydrophobic residues that surroundSer-363 forming the pocket are coloured red.
Different forms of Rubisco 1521
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

prokaryotic Rubisco we wish to study (Finn and Tabita,2003; Smith and Tabita, 2003, 2004). Both R. sphaeroides(Falcone and Tabita, 1991), but particularly R. capsulatus(Paoli et al., 1998) with its more rapid growth, especiallyunder aerobic chemolithoautotrophic conditions, offersa host system with a built-in reductive pentose phosphatepathway for easy complementation under a variety ofgrowth conditions, including conditions where CO2 maybe used as the sole carbon source in both aerobic andanaerobic atmospheres. Recovery of mutants is also madesimple by the fact that CO2 fixation is dispensable in theseorganisms and strains may be grown under aerobicheterotrophic conditions as with Escherichia coli. Clearly,growth at various levels and ratios of CO2 and O2
provides intriguing selective pressures to isolate poten-tially useful mutant forms of Rubisco, in an intracellularenvironment where Rubisco normally functions. Thissystem thus far appears inherently more stable thanartificial systems recently developed (Parikh et al., 2006),however, the potential for all such systems is great.Moreover, the R. capsulatus system offers a simple wayto isolate specific mutations in either the large (RbcL/CbbL) or small subunit (RbcS/CbbS). This may beaccomplished by using a two-plasmid system wherebyspecific mutations in either subunit may be obtained aftermating mutated rbcL/cbbL or rbcS/cbbS genes into theRubisco negative host containing the opposite, unmuta-genized gene carried on a separate plasmid (Fig. 6). Asillustrated for cyanobacterial Rubisco (Fig. 6), it wasfound that expression of rbcL or rbcS on separateplasmids results in the reconstitution of an active form IL8S8 holoenzyme in vivo from newly synthesized largeand small subunits. Thus, specific alterations on eachsubunit may be assessed after normal protein assembly inE.coli, followed by purification of the recombinant proteinas with the one-plasmid system.
An important hyrophobic pocket identified uponrandom mutagenesis and bioselection
Several interesting mutant cyanobacterial (Synechococcussp. strain PCC 6301) rbcL and rbcS genes have beenisolated upon error-prone random mutagenesis using theR. capsulatus system (Smith and Tabita, 2003, 2004; SSScott and FR Tabita, unpublished results). Of particularinterest were residues that were distal to the active sitethat were localized at monomer–monomer or dimer–dimerinterfaces and appeared to influence important kineticconstants, particularly the Kc. One of these mutantproteins, D103V, was shown not to be able to supportgrowth in R. capsulatus SBI/II–, presumably by virtue ofcausing a rise in the Kc value (Smith and Tabita, 2003).Asp-103 is localized on the surface of one monomer ofa dimeric pair, where it contacts Ser-367 of a monomerfrom a second dimeric pair of large subunits. To gain an
understanding as to how Asp-103, which is far from theactive site, might influence catalysis, an attempt was madeto isolate specific internal suppressor mutations which, itwas surmised, would overcome the effect of D103V,provide insights into residues that might interact withAsp-103, and subsequently allow growth in the R.capsulatus SBI/II– background. Over several years, multi-ple investigators in our laboratory were all able to isolatea single suppressor mutation that allowed growth in the R.capsulatus system. This suppressor or compensatory pro-tein, a D103VA375V double mutant, was found topossess an improved Kc. Furthermore, it appears that bothAsp-103 and Ala-375 exert their influence on Ser-376, animportant active-site residue required for binding the P2
Fig. 6. One-plasmid (A) and two-plasmid (B) vectors used to identifymutations in Synechococcus PCC 6301 Rubisco. To ensure constantlevels of expression, the rbcLrbcS genes are under the control of thetranscriptional regulator CbbR and the cbbM promoter from R. rubrum(Smith and Tabita, 2003). The plasmids offer blue/white screening andare resistant to tetracyline (TcR). The two-plasmid system (B) may beused to target rbcL or rbcS specifically for error-prone randommutagenesis, The rbcL gene may be independently expressed via thecbbR/cbbM promoter sequence on plasmid pRPS1-rbcL. The rbcS geneis independently expressed on plasmid pMCS5-rbcS. The two plasmidsare resistant to different antibiotics and belong to two differentincompatability groups. Both plasmids can be co-expressed in theRubisco-deficient strain of R. capsulatus (SBI/II-) to support photo-autotrophic growth comparable to cells containing plasmid pRPSMCS3-RbcLS (A).
1522 Tabita et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

phosphate of the substrate ribulose 1,5-bisphospate (SSatagopan et al., unpublished results). Most importantly,however, Ala-375 was subsequently shown to be localizedin an interesting hydrophobic region near the active site.Indeed, all forms of Rubisco, including form I (Anders-son, 1996, form II (Schneider et al., 1990), and form IIIRubisco (Kitano et al., 2001) contain this hydrophobicpocket (Fig. 7).
Why might this hydrophobic region be important? Ala-375 of the Synechococcus form I Rubisco is equivalent toSer-363 of the A. fulgidus enzyme. As discussed above,Ser-363 of the archaeal A. fulgidus enzyme appears toplay some role in the ability of this enzyme to interactwith oxygen (Kreel and Tabita, 2007), and this residue,along with Met-295, appears able to influence the Ko.Further studies with the equivalent Ala-375 residue ofform I Rubisco have also recently shown that this residueinfluences the ability of the cyanobacterial enzyme tosupport aerobic chemolithoautotrophic growth in the R.capsulatus system. Moreover, as expected, this residuealso directly affects the Ko value (S Satagopan et al.,unpublished results).
Conclusions
It is apparent that Rubisco, and its homologue, theRubisco-like protein, share an interesting history andundoubtedly present a classic example of divergentevolution. The different forms of Rubisco found in nature,some of which must function in very extreme orinhospitable environments, almost by definition, havemade structural adaptations to allow catalysis to occur.Investigating these structural adaptations is very useful, assuch studies provide a framework for understanding moreabout Rubisco structure and function in general.
Acknowledgements
This work was supported by grant GM24497 from the NationalInstitutes of Health and grants DE-FG02-01ER63241 and DE-FG02-91ER20033 from the offices of Biological and EnvironmentalResearch (Genomics: GTL Program) and Energy Biosciences,respectively, of the US Department of Energy.
References
Andersson I. 1996. Large structures at high resolution: the 1.6 Acrystal structure of spinach ribulose-1,5-bisphosphate carboxy-lase/oxygenase complexed with 2-carboxyarabinitol bisphosphate.Journal of Moecular Biology 259, 160–174.
Andersson I, Knight S, Schneider G, Lindqvist Y, Lundqvist T,Branden C-I, Lorimer GH. 1989. Crystal structure of theactive site of ribulose-bisphosphate carboxylase. Nature 337,229–234.
Andersson I, Taylor TC. 2003. Structural framework for catalysisand regulation in ribulose-1,5-bisphosphate carboxylase/oxygen-ase. Archives of Biochemistry and Biophysics 414, 130–140.
Andrews TJ. 1988. Catalysis by cyanobacterial ribulose bisphos-phate carboxylase large subunits in the complete absence ofsmall subunits. Journal of Biological Chemistry 263, 12213–12219.
Ashida H, Saito Y, Kojima C, Kobayashi K, Ogasawara N,Yokota A. 2003. A functional link between rubisco-like proteinof Bacillus and photosynthetic rubisco. Science 302, 286–290.
Cleland WW, Andrews JT, Gutteridge S, Hartman FC,Lorimer GH. 1998. Mechanism of Rubisco: the carbamate asgeneral base. Chemical Reviews 98, 549–561.
Delwiche CF, Palmer JD. 1996. Rampant horizontal transfer andduplication of rubisco genes in eubacteria and plastids. Molecularand Biological Evolution 13, 873–882.
Falcone DL, Tabita FR. 1991. Expression of endogenous andforeign ribulose 1,5-bisphosphate carboxylase-oxygenase(Rubisco) genes in a Rubisco deletion mutant of Rhodobactersphaeroides. Journal of Bacteriology 173, 2099–2108.
Finn MW, Tabita FR. 2003. Synthesis of catalytically active formIII ribulose 1,5-bisphosphate carboxylase/oxygenase in archaea.Journal of Bacteriology 185, 3049–3059.
Fig. 7. Comparison of hydrophobic pockets surrounding Ala-375 (green) in the Synechococcus Rubisco (A; 1RBL; form I) and the equivalentresidues, Ile-367 in Rhodospirillum rubrum form II enzyme (B; 5RUB) and Ser-366 in the Pyrococcus kodakaraensis form III enzyme (C; 1GEH).The adjacent residue, which is an active-site Ser is coloured in all the three structures. The active-site ligands, i.e. CABP in the SynechococcusRubisco and sulphate ions in the Pyrococcus enzyme are indicated in black. The side chains of residues coloured in yellow are within 4 A of the Ala-375 or the equivalent residue in at least one of the three structures.
Different forms of Rubisco 1523
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021

Fuchs G, Lange S, Rude E, Schaefer S, Schauder R, Scholtz R,Stupperich E. 1987. Autotrophic CO2 fixation in chemotrophicanaerobic bacteria. In: Van Verseveld HW, Duine JA, eds.Microbial growth on C1 compounds. Dordrecht, The Netherlands:Martinus Nijhoff, 39–43.
Green DM, Whitney SM, Matsumura I. 2007. Artificially evolvedPCC6301 Rubisco variants exhibit improvements in folding andcatalytic efficiency. Biochemical Journal 404, 517–524.
Hanson TE, Tabita FR. 2001. A ribulose-1,5-bisphosphatecarboxylase/oxygenase (Rubisco)-like protein from Chlorobiumtepidum that is involved with sulfur metabolism and the responseto oxidative stress. Proceedings of the National Academy ofSciences, USA 98, 4397–4402.
Hanson TE, Tabita FR. 2003. Insights into the stress response andsulfur metabolism revealed by proteome analysis of a Chlorobiumtepidum mutant lacking the Rubisco-like protein. PhotosynthesisResearch 78, 231–48.
Hartman FC, Harpel MR. 1994. Structure, function, regulationand assembly of D-ribulose-1,5-bisphosphate carboxylase/oxygen-ase. Annual Review of Biochemistry 63, 197–234.
Imker HJ, Fedorov AA, Fedorov EV, Almo SC, Gerlt JA. 2007.Mechanistic diversity in the Rubisco superfamily: the ‘enolase’ inthe methionine salvage pathway in Geobacillus kaustophilus.Biochemistry 46, 4077–4089.
Kitano K, Maeda N, Fukui T, Atomi H, Imanaka T, Miki K.2001. Crystal structure of a novel-type archaeal rubisco withpentagonal symmetry. Structure 9, 473–481.
Kreel NE, Tabita FR. 2007. Substitutions at methionine 295 ofArchaeoglobus fulgidus ribulose-1,5-bisphosphate carboxylase/oxygenase affect oxygen binding and CO2/O2 specificity. Journalof Biological Chemistry 282, 1341–1351.
Knight S, Andersson I, Branden CI. 1990. Crystallographicanalysis of ribulose 1,5-bisphosphate carboxylase from spinach at2.4 A resolution. Subunit interactions and active site. Journal ofMolecular Biology 215, 113–160.
Lee B, Berka R, Tabita FR. 1991a. Mutations in the small subunitof cyanobacterial ribulose bisphosphate carboxylase/oxygenasethat modulate interactions with large subunits. Journal of Bio-logical Chemistry 266, 7417–7422.
Lee B, Read BA, Tabita FR. 1991b. Catalytic properties ofrecombinant octameric, hexadecameric, and heterologous cyano-bacterial/bacterial ribulose-1,5 bisphosphate carboxylase/oxygen-ase. Archives of Biochemistry and Biophysics 291, 263–269.
Lee B, Tabita FR. 1990. Purification of recombinant ribulose-1,5-bisphosphate carboxylase/oxygenase large subunits suitablefor reconstitution and assembly of active L8S8 enzyme. Bio-chemistry 29, 9352–9357.
Li H, Sawaya MR, Tabita FR, Eisenberg D. 2005. Crystalstructure of a novel Rubisco-like protein from the green sulfurbacterium Chlorobium tepidum. Structure 13, 779–789.
Parikh MR, Greene DH, Woods KK, Matsumuara I. 2006.Directed evolution of Rubisco hypermorphs through geneticselection in engineered E. coli. Protein Engineering Design andSelection 19, 113–119.
Paoli GC, Vichivanives P, Tabita FR. 1998. Physiological controland cbb gene regulation in Rhodobacter capsulatus. Journal ofBacteriology 180, 4258–4269.
Portis Jr AR. 1992. Regulation of ribulose 1,5-bisphosphatecarboxylase/oxygenase activity. Annual Review of Plant Physiol-ogy 43, 415–437.
Read BA, Tabita FR. 1992a. Amino acid substitutions in thesmall subunit of ribulose-1,5carboxylase/oxygense that in-fluence catalytic activity of the holoenzyme. Biochemistry 31,519–525.
Read BA, Tabita FR. 1992b. A hybrid ribulose bisphosphatecarboxylase/oxygenase enzyme exhibiting a substantial increasein substrate specificity factor. Biochemistry 31, 5554–5560.
Schneider G, Lindqvist Y, Branden CI. 1992. RUBISCO:structure and mechanism. Annual Review of Biophysical andBiomolecular Structure 21, 119–143.
Schneider G, Lindqvist Y, Lundqvist T. 1990. Crystallographicrefinement and structure of ribulose-1,5-bisphosphate carboxylasefrom Rhodospirillum rubrum at 1.7 A resolution. Journal ofMolecular Biology 211, 989–1008.
Smith SA, Tabita FR. 2003. Positive and negative bioselection ofmutant forms of prokaryotic (cyanobacterial) ribulose-1,5-bisphosphate carboxylase/oxygenase. Journal of MolecularBiology 331, 557–569.
Smith SA, Tabita FR. 2004. Glycine 176 affects catalyticproperties and stability of the Synechococcus sp. strain PCC6301 ribulose 1,5-bisphosphate carboxylase/oxygenase. Journalof Biological Chemistry 279, 25632–25637.
Spreitzer RJ. 1999. Questions about the complexity of chloroplastribulose-1, 5-bisphosphate carboxylase/oxygenase. Photosynthe-sis Research 60, 29–42.
Spreitzer RJ, Salvucci ME. 2002. Rubisco: structure, regulatoryinteractions, and possibilities for a better enzyme. Annual Reviewof Plant Biology 53, 449–475.
Spreitzer RJ, Srinivasa RP, Satagopan S. 2005. Phylogeneticengineering at an interface between large and small subunitsimparts land-plant kinetic properties to algal Rubisco.Proceedings of the National Academy of Sciences, USA 102,17225–17230.
Tabita FR. 1999. Microbial ribulose 1,5-bisphosphate carboxylase/oxygenase: a different perspective. Photosynthesis Research 60,1–28.
Tabita FR, Hanson TE, Li H, Satagopan S, Singh J, Chan S.2007. Function, evolution, and structure of the Rubisco-likeproteins and their Rubisco homologs. Microbiology and Molecu-lar Biology Reviews 71, 576–599.
Tabita FR, McFadden BA. 1974. D-Ribulose 1, 5-diphosphatecarboxylase from Rhodospirillum rubrum. Il. Quaternary struc-ture, composition, catalytic and immunological properties. Jour-nal of Biological Chemistry 249, 3459–3464.
Watson GM, Yu JP, Tabita FR. 1999. Unusual ribulose 1,5-bisphosphate carboxylase/oxygenase of anoxic archaea. Journalof Bacteriology 181, 1569–1575.
Yooseph S, Sutton G, Rusch DB, et al. 2007. The Sorcerer IIglobal ocean sampling expedition: expanding the universe ofprotein families. PLoS Biology 5, e16.
Yoon KS, Hanson TE, Gibson JL, Tabita FR. 2000. AutotrophicCO2 metabolism. In: Lederburg J, ed. Encyclopedia of micro-biology, 2nd edn. San Diego, CA: Academic Press Inc.,349–358.
1524 Tabita et al.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/59/7/1515/640699 by guest on 25 N
ovember 2021