
Human Cancer Biology
Tumor-Reactive CD8þ T Cells in Metastatic GastrointestinalCancer Refractory to Chemotherapy
Simon Turcotte1, Alena Gros1, Eric Tran1, Chyi-Chia R. Lee2, John R. Wunderlich1,Paul F. Robbins1, and Steven A. Rosenberg1
AbstractPurpose: To evaluate whether patients with metastatic gastrointestinal adenocarcinomas refractory to
chemotherapy harbor tumor-reactive cytotoxic T cells.
ExperimentalDesign:ExpansionofCD8þ tumor-infiltrating lymphocytes (TIL) and cancer cell lineswas
attempted from gastrointestinal cancer metastases in 16 consecutive patients for the study of antitumor
immune recognition. Retroviral transduction of genes encoding T-cell receptors (TCR) was used to define
HLA-restriction elements and specific reactivity.
Results: TIL were expanded frommetastases in all patients, and new tumor cell lines were generated in 5
patients. Autologous tumor recognition without cross-reactivity against allogeneic HLA-matched gastro-
intestinal tumors was found in CD8þ TIL from 3 of these 5 patients. In a patient with gastric cancer liver
metastases, the repertoire of CD8þ TIL was dominated by cytolytic sister clones reactive to 2 out of 4
autologous cancer cell lines restricted by HLA-C�0701. From the same patient, a rare CD8þ TIL clone with a
distinct TCR recognized all four cancer cell lines restricted byHLA-B�4901. In a patient with bile duct cancer,
two distinct antitumor cytolytic clones were isolated from a highly polyclonal CD8þ TIL repertoire. TCRs
isolated from these clones recognized epitopes restricted by HLA-A�0201. In a third patient, CD8þ TIL
reactivity was progressively lost against an autologous colon cancer cell line that displayed loss of HLA
haplotype.
Conclusions: This study provides a basis for the development of immunotherapy for patients with
advanced gastrointestinal malignancies by first establishing the presence of naturally occurring tumor-
reactive CD8þ TIL at the molecular level. Clin Cancer Res; 20(2); 331–43. �2013 AACR.
IntroductionGastrointestinal adenocarcinomas are among the 10
most common malignancies worldwide and the overallmortality associated with their high metastatic potentialhas not changed significantly over the last decades (1).Currentmultimodality treatments can slowdisease progres-sion but fail to cure patients with metastatic disease. Thusfar, immune-based therapies have not shown clinical effec-tiveness in patients with gastrointestinal cancers (2–5). Apositive association between the density of the tumor-infiltrating lymphocyte (TIL) infiltrate and better outcomeshas, nonetheless, been reported in patients with adenocar-cinomas arising in the esophagus, stomach, pancreas, liver,
bile ducts, gallbladder, colon, and rectum (6). Multipleimmune-escape mechanisms have however been proposedthat may contribute to the absence, the depletion, and thedysfunction of tumor-reactive TIL in solid tumors (7–9).Thus, the specific recognition of human metastatic gastro-intestinal cancers by naturally occurring, autologous, cyto-toxic CD8þ T cells harvested from the tumor site has notbeen defined at the T-cell clonal level with defined HLArestriction elements.
Results from studies in patients with metastatic melano-ma have stimulated us to investigate the antitumor reactiv-ity of CD8þ TIL in common epithelial malignancies.Indeed, TIL isolated frommelanomametastases can exhibitdirect in vitro tumor recognition of defined antigens pre-sented by specific class I HLAs (10–14), and tumor depositsseem to harbor antitumor T cells of sufficient avidity and insufficient numbers to respond to nonspecific systemicmod-ulation of immunity (15–18). In addition, as now reportedby multiple institutions, the adoptive cell transfer of autol-ogous TIL can mediate complete cancer regression inpatients with metastatic disease considered incurable withstandard therapy, with complete responders reported upto 10 years after treatment (19–23). The curative potentialof TIL-based immunotherapy in advanced melanoma
Authors' Affiliations: 1Surgery Branch and 2Laboratory of Pathology,National Cancer Institute, NIH, Bethesda, Maryland
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Corresponding Author: Steven A. Rosenberg, National Cancer Institute,10 Center Drive MSC 1201, CRC Room 3-3940, Bethesda, MD 20892.Phone: 301-496-4164; Fax: 301-402-1738; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-13-1736
�2013 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org 331
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

represents a paradigmatic shift on how solid cancer treat-ment is approached, and whether this strategy can beapplied for common metastatic epithelial malignanciesmerits active investigations.
In the current report, an in vitro analysis of TIL was carriedin 16 patients with metastatic gastrointestinal cancer.Detailed CD8þ TIL reactivity to autologous gastrointestinalcancer metastases was carried out in 5 patients fromwhom,13new cancer cell lineswere established. TIL from3of thesepatients exhibited specific immune reactivity against theirautologous metastatic cancer. By defining immune featuresof metastatic gastrointestinal cancer cells and TIL, ourfindings have direct relevance to efforts to develop immu-notherapies for patients with these malignancies.
Materials and MethodsPatients and tumor processing
Written informed consent was obtained from all patientsenrolled under protocols approved by the InstitutionalReview Board of the National Cancer Institute (NCI) andU.S. Food and Drug Administration. Single-cell suspen-sions were obtained from freshly resected tumors by inde-pendent enzymatic digestion and mechanical dispersion aspreviously described for melanoma specimens (24).
Primary human cancer cell cultures and culture ofother cancer cell lines
To develop cancer cell lines, 0.25e6 live nucleated cellswere plated in multiple 25-cm2 ultra-low attachment andstandard treated cantedneckflasks (Corning3815and3056)in RPMI 1640–based medium supplemented with 20% FBS(Defined; HyClone Laboratories), 25 mmol/L HEPES, 2mmol/L L-glutamine, 100U/mL penicillin, 100 mg/mL strep-
tomycin (all fromLife Technologies, Invitrogen), 1.25mg/mLamphotericin B (X-GEN Pharmaceuticals), and 10 mg/mLciprofloxacin (BedfordLaboratories). After 6 to12weeks, cellaggregates/tumor spheres (�200 mm in diameter) weretransferred into standard 25-cm2 flasks for propagationunder adherent conditions. For adherent conditions, fibro-blast overgrowthwas controlledbydifferential trypsinization(Trypsin–EDTA 1�, 0.05%; Gibco) andmechanical removal(17-mm blade cell scraper; Sarstedt), and cultures were fedweekly or according to need, and passaged into larger flaskswhen reaching confluence. The human cancer cell lines KatoIII, NCIN87, NCIH508, Colo205, HCT15, SK-CO-1, KM12,HT29, SW480, SW620, HCC2998, SW1463, Capan1, andPanc 02.03 were purchased from the American Type CultureCollection and grown under the vendor’s suggested condi-tions. Human melanoma cell lines, 3350 and 624, andhuman pancreatic cancer cell lines, 2596 and 2742-2, wereestablished in our laboratory.
The authenticity of all cell lines was confirmed by HLAtyping and testing expression of melanoma differentiationantigens (MART-1, Melan-A, and gp100) and gastrointes-tinal cancer antigens (CEA, pan-cytokeratins). Mycoplas-ma contamination was ruled out on all cell cultures byusing MycoAlert (Lonza) according to the manufacturer’sinstructions.
Generation of TILTIL establishment and expansion followed techniques
used for metastatic melanoma tumor deposits and arepresented in Supplementary Methods (24, 25).
Characteristics of fresh TIL and cultured linesStaining of paraffin-embedded tissue and cell pellets was
done at the NCI Clinical Research Center (NCI/CRC)Pathology and Cytology Laboratory, following standardprocedures with appropriate positive and isotype controls.Antibodies and staining conditions are presented in Sup-plementary Methods. For flow cytometry, the followingmonoclonal antibodies specific for human antigens andappropriate isotype controls were used: from BD Bios-ciences, APC-H7-conjugated anti-CD3 (SK7), APC anti-CD137/4-1BB (4B4-1); from Invitrogen: R-PE-Texas Red-conjugated anti-CD8 (3B5). Cell aggregates and dead cellswere excluded by forward and side scatter, and with pro-pidium iodide staining. Flow cytometry analysis was carriedout with FlowJo F7.5.5 software (TreeStar). HLA typing ofcell lines was done by the NCI/CRC HLA laboratory ongenomic DNA following standard procedures (Supplemen-tary Methods).
Cancer-cell recognition by T cells coculture assaysT-cell reactivity to cancer cell lineswas assessed after 24-to
36-hour coculture assays in flat-bottom 96-well plates (1e5T cells, 0.5e5 trypsinized cancer cells; final volume 200 mL).ELISA was carried out for measurement of IFN-g release insupernatant, and flow cytometry was used to quantifyCD137 (4-1BB) upregulation on T cells (SupplementaryMethods). Chromium-51 release 4- to 6- hour cytolysis
Translational RelevanceLittle is known about T-cell immune response of
patients to advanced gastrointestinal adenocarcinomas.How immunotherapy could be a relevant alternative tocurrent therapeutics for these common malignancies isunclear. Conversely, adoptive transfer of tumor-infiltrat-ing lymphocytes (TIL) can mediate durable completeregression of metastatic melanoma in patients withrefractory disease. By overcoming the difficulty of estab-lishing new cancer cell lines and by expanding TIL fromthe samepatients, this studyfirst provides evidence at themolecular level for the presence of cytolytic tumor-reac-tive CD8þ T cells in growing gastrointestinal cancermetastases. The difficulty in establishing tumor targetsto test for T-cell reactivity, the heterogeneity and defectsin antigen expression by cancer cell lines, the low fre-quency of tumor-reactive CD8þ TIL, and the lack ofshared reactivity across HLA-matched allogeneic tumorsseem as major challenges for the development of effec-tive T-cell based immunotherapy for patients withadvanced gastrointestinal cancer.
Turcotte et al.
Clin Cancer Res; 20(2) January 15, 2014 Clinical Cancer Research332
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

assays were used as described in previous studies (26, 27)and in Supplementary Methods. The percentage of specificlysis was calculated as: percent lysis ¼ (sample release �spontaneous release)/(maximal � spontaneous release) �100, average of triplicate samples.
Limiting dilution cloning of 4-1BBþ–enriched TILAfter stimulation assays with autologous cancer cell line,
CD8þ TIL that exhibited reactivity by high 4-1BBþ expres-sion were sorted by magnetic cell separation (anti-APCIMag; BD Biosciences) or fluorescence-activated cell sorting(FACSAria II; BDBiosciences). Briefly, between 1 and 20 TILwere plated in each well of a 96-well U-bottomed plate in200 mL of conditioned medium containing 3,000 U/mLrhIL-2, 30 ng/mL of OKT3, with 5� 104 irradiated (40 Gy)allogeneic peripheral bloodmononuclear cells (PBMC).Onday 5 and every 3 to 4 days thereafter, half of themedium ineach well was replaced with fresh medium containinginterleukin (IL)-2. About 10 to 14 days after culture initi-ation, wells in which cell growth was visibly apparent werescreened for reactivity to autologous and allogeneic cancercell line. Clone reactive by IFN-g secretion and/or 4-1BBexpression were subsequently expanded by Rapid Expan-sion Protocol (REP; ref. 24).
TCRsequencingandcloning, retroviral transductionofPBMCClonotypic analysis of the TCR repertoire was done from
the RNA purified from CD8þ TIL with an RNeasy mini kit(Qiagen) and reverse transcribed into cDNA for 50RapidAmplification of cDNA Ends (RACE) by SMARTer RACEcDNA Amplification Kit as per manufacturer instructions(Clontech) and as described with subcloning into a pCR2.1vector by TA cloning (Life Technologies; ref. 28). Consensusregions were determined using the international ImMuno-GeneTics information system (29) for sequence analysis of96 coloniespicked fromeach50 RACEproductof bothTCRaand b chains and their variable regions. Murinized humanTCRswere cloned intoMSGV1 retroviral vectors as describedin ref. (30) and detailed in Supplementary Methods. Trans-duction of PBMC was done as described in ref. (31), andefficiency was assessed after 1-week expansion by flow cyto-metry using an anti-mouse TCRb-chain antibody (H57-597;eBioscience). MSGV1 vector expressing the TCR recognizingthe MART-1:27-35 epitope (31) and GFP were used ascontrols in all experiments, as well as untransduced PBMC.
Statistical analysisStatistical analyses were performed on GraphPad Prism
software version 5.04. Variances of mean values are pre-sented as SEM. Two-tailed, nonparametric tests were usedand P values of �0.05 were considered significant.
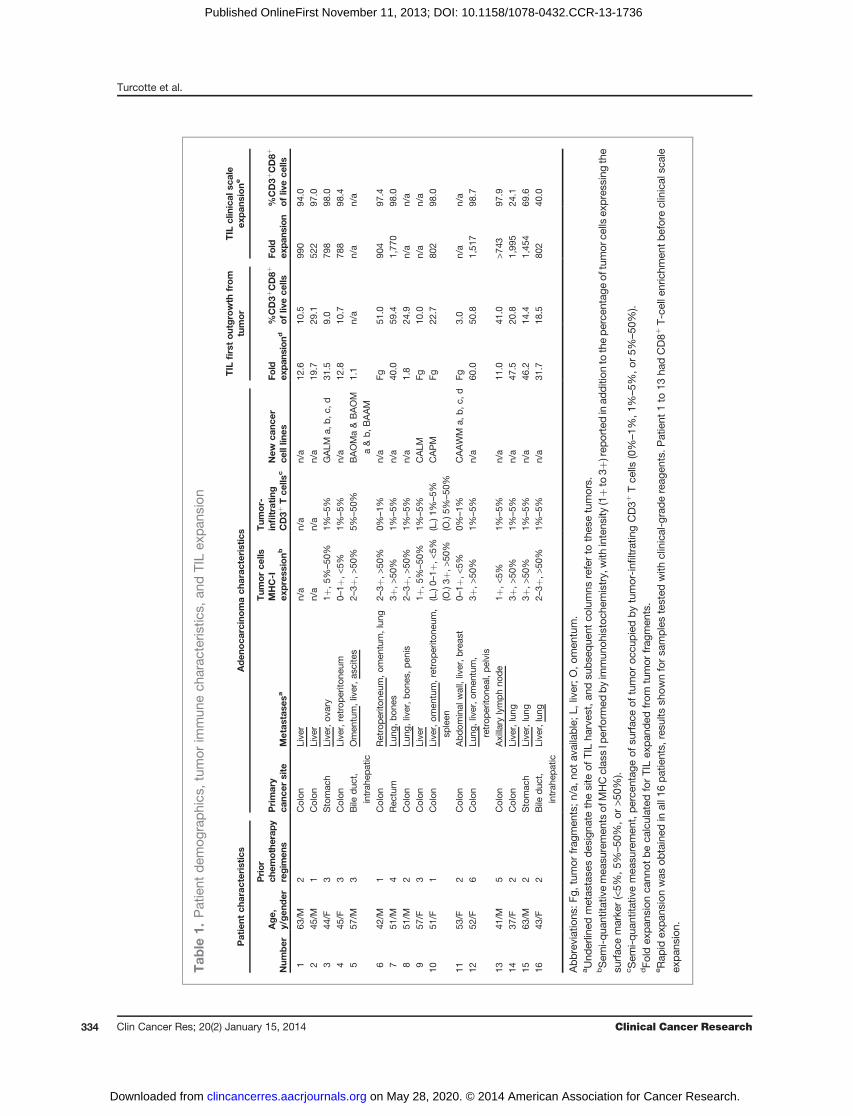
ResultsPatient and tumor clinicopathologic and immunologiccharacteristicsResected tumors from 16 patients withwidespread,mod-
erately to poorly differentiatedmetastatic adenocarcinomas
originating from the stomach, the bile duct, or the colonwere studied (Table 1). All patients had progressive diseaseafter receiving at least one standard first-line chemotherapyregimen (median, 2). The expression ofMHCclass-I (MHC-I)molecules, required for CD8þ T-cell recognition, was het-erogeneous across resected metastases and nearly undetect-able in 35% of lesions (Table 1 and Fig. 1). All metastaseswere poorly infiltrated by TIL, which represented less than5% of the tumor-cut surface on histologic assessment, withthe exception of omentalmetastases frompatients 5 and 10,which were more densely infiltrated by CD3þ T cells.Heterogeneity in antigenic expression by cancer cells withinthe same patient was exemplified in metastases resectedin patient 10, as MHC-I was faintly detected on the livermetastasis but strongly on the omental metastasis (Fig. 1).Patient 11 had a mucinous adenocarcinoma characterizedbyfine reticulatedbandsof tumor cells separatedby copiouspools of mucin and surrounded by a fibrotic stroma con-taining rare T cells (Fig. 1).
TIL expansion from gastrointestinal metastasesMinced tumor fragments or cell suspensions obtained
from gastrointestinal cancer metastases were cultured in IL-2–containing media. Overgrowth of tumor and otheradherent cells by TIL was observed between 16 and 29 daysfrom culture initiation (median, 21 days). An average of25.0% � 4.6% of the live cells in these cultures was CD3þ
CD8þ (range, 3.0%–59.4%). TIL were further expanded tolarge numbers from all samples using soluble anti-CD3 andirradiated allogeneic PBMC feeder cells for 14 days. Sepa-rated CD8þ T cells that were positively selected with mag-netic beads from the expanded bulk TIL cultures had amedian cumulative fold expansion of 853 using clinical-grade reagents (range, 522–1,770; Table 1). Without CD8þ
enrichment, TILwere similarly expanded frombulk culturesin patients 14, 15, and 16 with a median 1,454-fold expan-sion (range, 802–1,995). Overall, TIL from gastrointestinalcancer metastases in patients heavily pretreated with che-motherapy were found to have good in vitro proliferativepotential.
Establishment of new gastrointestinal cancer cell linesand MHC-I expression loss
In parallel to setting up cultures for TIL expansion, asmany cancer cell lines as possible were generated for eachpatient by initial culturing of tumor cell suspensions inultra-low attachment flasks in addition to standard techni-ques in adherent flasks. When tumor spheroids wereobtained, as in patients 3, 5, and 11, multiple cell lineswere initiated from distinct spheroids, and thus 13 cancercell lines were established in 5 patients (Table 1, Supple-mentary Fig. S1). Complete HLA genotyping of the cancercell lines and PBMCs confirmed the parenthood of the linesfor each patient (Supplementary Table S1). Loss of hetero-zygosity at the HLA loci (haplotype loss) was found in twoout of the 13 newly established cancer cell lines (patients 9and 10). Total loss ofMHC-I protein expression was seen inaddition to genomic HLA haplotype loss in the only one
T-Cell Recognition of Metastatic Gastrointestinal Cancer
www.aacrjournals.org Clin Cancer Res; 20(2) January 15, 2014 333
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736
Tab
le1.
Patient
dem
ographics
,tum
orim
mun
ech
arac
teris
tics,
andTILex
pan
sion
Patient
charac
teristics
Aden
oca
rcinomach
arac
teristics
TIL
firstoutgrowth
from
tumor
TIL
clinical
scale
expan
sione
Num
ber
Age,
y/gen
der
Prior
chem
otherap
yregim
ens
Primary
canc
ersite
Metas
tase
sa
Tum
orce
llsMHC-I
express
ionb
Tum
or-
infiltrating
CD3þ
Tce
llsc
New
canc
erce
lllin
esFo
ldex
pan
siond
%CD3þ
CD8þ
ofliv
ece
llsFo
ldex
pan
sion
%CD3þ
CD8þ
ofliv
ece
lls
163
/M2
Colon
Live
rn/a
n/a
n/a
12.6
10.5
990
94.0
245
/M1
Colon
Live
rn/a
n/a
n/a
19.7
29.1
522
97.0
344
/F3
Stomac
hLive
r,ov
ary
1þ,5
%–50
%1%
–5%
GALM
a,b,c
,d31
.59.0
798
98.0
445
/F3
Colon
Live
r,retrop
erito
neum
0–1þ
,<5%
1%–5%
n/a
12.8
10.7
788
98.4
557
/M3
Bile
duc
t,intrah
epatic
Omen
tum,liver,a
scite
s2–
3þ,>
50%
5%–50
%BAOMa&BAOM
a&b,B
AAM
1.1
n/a
n/a
n/a
642
/M1
Colon
Retroperito
neum
,om
entum,lung
2–3þ
,>50
%0%
–1%
n/a
Fg51
.090
497
.47
51/M
4Rec
tum
Lung
,bon
es3þ
,>50
%1%
–5%
n/a
40.0
59.4
1,77
098
.08
51/M
2Colon
Lung
,liver,b
ones
,pen
is2–
3þ,>
50%
1%–5%
n/a
1.8
24.9
n/a
n/a
957
/F3
Colon
Live
r1þ
,5%
–50
%1%
–5%
CALM
Fg10
.0n/a
n/a
1051
/F1
Colon
Live
r,om
entum,retroperito
neum
,sp
leen
(L.)0–
1þ,<
5%(L.)1%
–5%
CAPM
Fg22
.780
298
.0(O.)3þ
,>50
%(O
.)5%
–50
%11
53/F
2Colon
Abdom
inal
wall,liver,b
reas
t0–
1þ,<
5%0%
–1%
CAAWM
a,b,c
,dFg
3.0
n/a
n/a
1252
/F6
Colon
Lung
,liver,o
men
tum,
retrop
erito
neal,pelvis
3þ,>
50%
1%–5%
n/a
60.0
50.8
1,51
798
.7
1341
/M5
Colon
Axillary
lymphno
de
1þ,<
5%1%
–5%
n/a
11.0
41.0
>743
97.9
1437
/F2
Colon
Live
r,lung
3þ,>
50%
1%–5%
n/a
47.5
20.8
1,99
524
.115
63/M
2Stomac
hLive
r,lung
3þ,>
50%
1%–5%
n/a
46.2
14.4
1,45
469
.616
43/F
2Bile
duc
t,intrah
epatic
Live
r,lung
2–3þ
,>50
%1%
–5%
n/a
31.7
18.5
802
40.0
Abbreviations
:Fg,
tumor
frag
men
ts;n
/a,n
otav
ailable;L
,liver;O
,omen
tum.
aUnd
erlined
metas
tase
sdes
igna
tethesite
ofTILha
rves
t,an
dsu
bse
que
ntco
lumns
referto
thes
etumors.
bSem
i-qua
ntita
tivemea
suremen
tsof
MHCclas
sIp
erform
edbyim
mun
ohistoch
emistry,with
intens
ity(1þto
3þ)rep
ortedinad
dition
totheperce
ntag
eof
tumor
cells
express
ingthe
surfac
emarke
r(<5%
,5%–50
%,o
r>5
0%).
cSem
i-qua
ntita
tivemea
suremen
t,pe
rcen
tage
ofsu
rfac
eof
tumor
occu
piedbytumor-infi
ltratingCD3þ
Tce
lls(0%–1%
,1%–5%
,or5%
–50
%).
dFo
ldex
pan
sion
cann
otbeca
lculated
forTILex
pan
ded
from
tumor
frag
men
ts.
eRap
idex
pans
ionwas
obtained
inall1
6patients,
resu
ltssh
ownforsa
mples
tested
with
clinical-gradereag
ents.P
atient
1to
13ha
dCD8þ
T-ce
llen
richm
entbeforeclinical
scale
expan
sion
.
Turcotte et al.
Clin Cancer Res; 20(2) January 15, 2014 Clinical Cancer Research334
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

colon cancer cell line generated from patient 10 (colonadenocarcinoma peritoneal metastasis, CAPM; Fig. 1), ren-dering this cell line "invisible" for CD8þ T cells.
CD8þ TIL recognition of newly established autologouscancer cell linesAfter expansion, CD8þ TIL recognition of autologous
gastrointestinal cancer was evaluated by examining their
ability to secrete IFN-g and to upregulate expression of theinducible activation marker 4-1BB (CD137; ref. 32) inresponse to tumor stimulation in the 5 patients for whichnew cancer cell lines had been established (Table 2). Asexpected by lack of MHC-I expression, CAPM failed tostimulate CD8þ TIL in patient 10 (Supplementary TableS2). The rare fractionofCD8þ TIL expanded from fragmentsof mucinous abdominal wall colon cancer metastasis in
Figure 1. Cancer cell lines established from gastrointestinal cancer metastases with heterogeneous MHC-I expression and weak T-cell infiltration.Immunohistochemical staining of paraffin-embedded gastrointestinal cancermetastases (left) in 5 patients (Pt.) and representative corresponding cancer celllines (right). Pt. 3, gastric adenocarcinoma liver metastasis, adjacent normal liver, and derived cancer cell line (GALMa). Pt. 5, omental metastasis from anintrahepatic bile duct adenocarcinoma and derived cancer cell line (BAOMa). Pt. 9, colon adenocarcinoma liver metastasis, adjacent liver, and derivedcancer cell line (CALM). Pt. 10, colon adenocarcinoma liver metastasis, adjacent liver, and omental metastasis, with derived cancer cell line (CAPM). Pt. 11,abdominal wall metastasis from a mucinous colon adenocarcinoma, adjacent stroma, and derived cancer cell line (CAAWMa). All epithelial derivedcancer cells are stainedby thepan-cytokeratinmarkerAE1AE3,whichdelineate tumor (T) from theadjacent normal tissues (N).MHCclass I stains all nucleatedcells, with weak and heterogeneous expression found in liver metastases in vivo; however, most derived cancer cell lines express MHC class I inculture except CAPM. Tumor-infiltrating lymphocytes are revealed by the T-cell marker CD3, and in all cases represent less than 5% of cells in tumor mass.Representative fields selected from whole slide scan, measure bar, 100 mm.
T-Cell Recognition of Metastatic Gastrointestinal Cancer
www.aacrjournals.org Clin Cancer Res; 20(2) January 15, 2014 335
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736
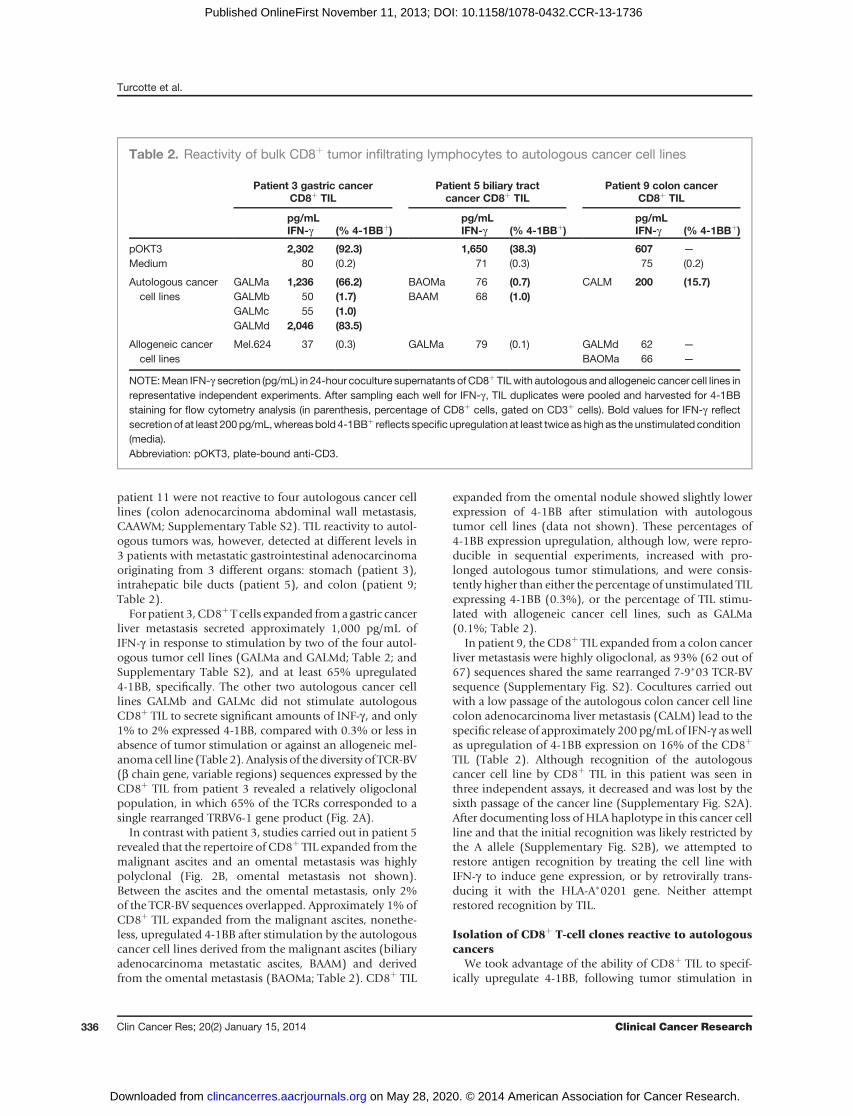
patient 11 were not reactive to four autologous cancer celllines (colon adenocarcinoma abdominal wall metastasis,CAAWM; Supplementary Table S2). TIL reactivity to autol-ogous tumors was, however, detected at different levels in3 patients with metastatic gastrointestinal adenocarcinomaoriginating from 3 different organs: stomach (patient 3),intrahepatic bile ducts (patient 5), and colon (patient 9;Table 2).
For patient 3, CD8þT cells expanded froma gastric cancerliver metastasis secreted approximately 1,000 pg/mL ofIFN-g in response to stimulation by two of the four autol-ogous tumor cell lines (GALMa and GALMd; Table 2; andSupplementary Table S2), and at least 65% upregulated4-1BB, specifically. The other two autologous cancer celllines GALMb and GALMc did not stimulate autologousCD8þ TIL to secrete significant amounts of INF-g , and only1% to 2% expressed 4-1BB, compared with 0.3% or less inabsence of tumor stimulation or against an allogeneic mel-anoma cell line (Table 2). Analysis of the diversity of TCR-BV(b chain gene, variable regions) sequences expressed by theCD8þ TIL from patient 3 revealed a relatively oligoclonalpopulation, in which 65% of the TCRs corresponded to asingle rearranged TRBV6-1 gene product (Fig. 2A).
In contrast with patient 3, studies carried out in patient 5revealed that the repertoire of CD8þ TIL expanded from themalignant ascites and an omental metastasis was highlypolyclonal (Fig. 2B, omental metastasis not shown).Between the ascites and the omental metastasis, only 2%of the TCR-BV sequences overlapped. Approximately 1% ofCD8þ TIL expanded from the malignant ascites, nonethe-less, upregulated 4-1BB after stimulation by the autologouscancer cell lines derived from the malignant ascites (biliaryadenocarcinoma metastatic ascites, BAAM) and derivedfrom the omental metastasis (BAOMa; Table 2). CD8þ TIL
expanded from the omental nodule showed slightly lowerexpression of 4-1BB after stimulation with autologoustumor cell lines (data not shown). These percentages of4-1BB expression upregulation, although low, were repro-ducible in sequential experiments, increased with pro-longed autologous tumor stimulations, and were consis-tently higher than either the percentage of unstimulated TILexpressing 4-1BB (0.3%), or the percentage of TIL stimu-lated with allogeneic cancer cell lines, such as GALMa(0.1%; Table 2).
In patient 9, the CD8þ TIL expanded from a colon cancerliver metastasis were highly oligoclonal, as 93% (62 out of67) sequences shared the same rearranged 7-9�03 TCR-BVsequence (Supplementary Fig. S2). Cocultures carried outwith a low passage of the autologous colon cancer cell linecolon adenocarcinoma liver metastasis (CALM) lead to thespecific release of approximately 200 pg/mL of IFN-g aswellas upregulation of 4-1BB expression on 16% of the CD8þ
TIL (Table 2). Although recognition of the autologouscancer cell line by CD8þ TIL in this patient was seen inthree independent assays, it decreased and was lost by thesixth passage of the cancer line (Supplementary Fig. S2A).After documenting loss of HLA haplotype in this cancer cellline and that the initial recognition was likely restricted bythe A allele (Supplementary Fig. S2B), we attempted torestore antigen recognition by treating the cell line withIFN-g to induce gene expression, or by retrovirally trans-ducing it with the HLA-A�0201 gene. Neither attemptrestored recognition by TIL.
Isolation of CD8þ T-cell clones reactive to autologouscancers
We took advantage of the ability of CD8þ TIL to specif-ically upregulate 4-1BB, following tumor stimulation in
Table 2. Reactivity of bulk CD8þ tumor infiltrating lymphocytes to autologous cancer cell lines
Patient 3 gastric cancerCD8þ TIL
Patient 5 biliary tractcancer CD8þ TIL
Patient 9 colon cancerCD8þ TIL
pg/mLIFN-g (% 4-1BBþ)
pg/mLIFN-g (% 4-1BBþ)
pg/mLIFN-g (% 4-1BBþ)
pOKT3 2,302 (92.3) 1,650 (38.3) 607 —
Medium 80 (0.2) 71 (0.3) 75 (0.2)
Autologous cancercell lines
GALMa 1,236 (66.2) BAOMa 76 (0.7) CALM 200 (15.7)GALMb 50 (1.7) BAAM 68 (1.0)GALMc 55 (1.0)GALMd 2,046 (83.5)
Allogeneic cancercell lines
Mel.624 37 (0.3) GALMa 79 (0.1) GALMd 62 —
BAOMa 66 —
NOTE:Mean IFN-g secretion (pg/mL) in 24-hour coculture supernatants of CD8þ TILwith autologous and allogeneic cancer cell lines inrepresentative independent experiments. After sampling each well for IFN-g, TIL duplicates were pooled and harvested for 4-1BBstaining for flow cytometry analysis (in parenthesis, percentage of CD8þ cells, gated on CD3þ cells). Bold values for IFN-g reflectsecretion of at least 200pg/mL,whereasbold 4-1BBþ reflects specific upregulation at least twice as high as the unstimulated condition(media).Abbreviation: pOKT3, plate-bound anti-CD3.
Turcotte et al.
Clin Cancer Res; 20(2) January 15, 2014 Clinical Cancer Research336
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

patients 3 and 5 to enrich and isolate reactive T cells. Forpatient 3, CD8þ TIL expressing 4-1BB after stimulation withGALMa were enriched by magnetic beads before limitingdilution cloning (Fig. 2C). Expansionoccurred in16%of the
platedmicrocultures, allowing evaluation of the reactivity of154 clonal T cells. Specific upregulation of 4-1BB expressionwas observed in 19 microcultures (12.3%). After reexpan-sion with soluble anti-CD3 and irradiated allogeneic PBMC
Figure 2. Reactivity of CD8þTIL clones to autologous gastric and bile duct cancers. A, diversity of the TCR repertoire of CD8þ TIL expanded frompatient (Pt.) 3liver metastasis, in which distinct TCR variable b chains sequences are labeled by their TCR germline V consensus regions (TRBV). B, highly polyclonalTCRBV repertoire of TIL found in malignant ascites from a bile duct cancer in pt. 5. C, for Pt. 3, FACS plot of 4-1BB expression upregulation in bulk CD8þ TILafter stimulation by autologous GALMa that allowed enrichment of the 4-1BBþ cells for limiting dilution cloning (red rectangle). Inferior to the FACS plot,the bar graphs represent the reactivity of 19 CD8þ TRBV6-1 TIL clones to the autologous GALMa, but not to the autologous GALMb cancer cell line, anonreactive (NR) clone (TRBV12-4), and themelanoma (F5) CD8þ TIL specificity control. FACS plot (middle) of the approximately 4%4-1BB upregulation (redrectangle) seen in bulk CD8þ TIL from pt.3 after stimulation by autologous GALMb. The bar graphs on the right side of the dotted line represent the lowreactivity found in two expanded clones (1�, TRBV9; 2�, unsequenced) to GALMa and GALMb, the bulk CD8þ TIL, a NR clone, and the F5 specificitycontrol by 4-1BB upregulation; none of these clones secreted significant amount of IFN-g . D, for Pt.5, FACS plot of the approximately 1%4-1BB upregulationseen in the bulk CD8þ TIL. Limiting dilution cloning allowed isolation of two clones (TRBV28 and TRBV3-1) reactive to two autologous cancer celllines established from an omental metastasis BAOMa and the malignant ascites BAAM. For bar graphs, left axis, IFN-g ng/mL; right axis, %4-1BBþ ofCD3þCD8þ. Unstim., unstimulated.
T-Cell Recognition of Metastatic Gastrointestinal Cancer
www.aacrjournals.org Clin Cancer Res; 20(2) January 15, 2014 337
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

feeders, each of the 19 clones still upregulated 4-1BB expres-sion after stimulation by GALMa, but only six of the 19clones secreted significant amounts of IFN-g . The TCR of 18reactive clones were sequenced and all shared the samedominant TRBV6-1�01 b chain rearrangement sequence(Supplementary Table S3 for complete TCR a and b chains).As expected, the TRBV6-1 clones were only stimulated byGALMa and GALMd, but not by GALMb and GALMc,reflecting the reactivity pattern initially found from the bulkCD8þ (Fig. 2C left bar graphs; TRBV6-1 clone reactivityagainst GALMd, nonreactivity against GALMc, and alloge-neic specificity controls are not shown).
In an attempt to isolate rare CD8þ TIL clones reactive toGALMb, FACS sorting was carried out to enrich a popula-tion corresponding to approximately 4% of the bulk TILthat upregulated 4-1BB expression in response to stimula-tion by this autologous cancer cell line (Fig. 2C, right FACSplot). Clonal growth efficiency was 2.1% of the sorted TILthat were plated at 1 and 2 cells per well and led toexpansion of 32 clones. Five clones could successfully bereexpanded, two of which demonstrating approximately10%4-1BBupregulation following coculturewith all gastricadenocarcinoma livermetastasis (GALM) cell lines, withoutsignificant IFN-g secretion capacity (Fig. 2C, right bargraphs; clones 1� and 2� reactivity to GALMc and GALMdare not shown). The TCR of the CD8þ T-cell clones mostreactive to GALM cell lines was sequenced and found to bederived from the TRBV9 germline gene (SupplementaryTable S3). We estimated that the frequency of the TRBV9TCR was of less than 1.3% of CD8þ TIL expanded from themetastasis, as this specific TCR sequence did not match anyof the one initially analyzed (n ¼ 76) from the bulk CD8þ
TIL (Fig. 2A).Thus, from the gastric cancer liver metastasis of patient 3,
multiple TRBV6-1 sister clones constituting a dominant T-cell clonotype in CD8þ TIL recognized two out of fourautologous cancer cell lines (GALMa andGALMd), whereasa genetically distinct TIL clone found at low frequencyrecognized all four autologous cell lines with lower reac-tivity. Of note, the TRBV6-1 sister clones, with geneticallyidentical TCRs, upregulated 4-1BB to similar levels upontumor recognition but differed in their capacity to secreteIFN-g , implying distinct functional attributes regulated atthe epigenetic level. Our findings also supported distinctantigen expression profiles found in four autologous cancercell lines derived from a single liver metastasis constitutedby heterogeneous cancer cells.
For patient 5 with bile duct cancer, approximately 4,0004-1BBþCD8þ TIL from themalignant ascites and the omen-tal metastasis were FACS-sorted after stimulation withautologous cell lines (Fig. 2D). Overall, 604 clones grewat a clonal efficiency of 9%, 17%, and 32%whenplated at 1,2, and 5 cells perwell. A preliminary assessment of the T-cellclone reactivity by stimulation with autologous tumorindicated that three clones (0.5%)were reactive by standardcriteria for IFN-g release and 10 (1.6%) clones possessedrelatively low reactivity with IFN-g secretion at least twotimes higher than theunstimulated condition, however, not
reaching the 200 pg/mL cutoff. Among 32 microculturesassessed for 4-1BB upregulation, four clones (12.5%) upre-gulated the surface marker by at least 20%. After reexpan-sion, two out of the 17 clones showed signs of specific TCRengagement with autologous BAOMa andBAAMcancer celllines by 4-1BB upregulation, but only one clone secretedsignificant amounts of IFN-g (Fig. 2D bar graphs; non-reactivity to allogeneic targets not shown). The two clonesdid not share the same TCR (TRBV28 and TRBV3-1;Supplementary Table S3) and were found at a frequencyof less than 1.8% of the expanded CD8þ TIL. Again, thedifferential reactivity of these clones to autologous cancercell lines here, established from distinct body compart-ments (omental metastases and malignant ascites), sug-gested a heterogeneous expression of immune epitopes bythese cell lines.
Importantly, all of the CD8þ TIL clones isolated from thePatient 3 and 5 lysed autologous tumors with specificity,irrespective of their ability to specifically secrete IFN-g(Fig. 3). For patient 3, the TRBV6-1 sister clonesmaintainedthe same pattern of reactivity noted by 4-1BB upregulationby not lysing the autologous GALMc line, whereas theTRBV9 clone could lyse both cell lines GALMa and GALMc(Fig. 3A). For patient 2, similarly, the 4-1BB upregulationseen by the TRBV28 and TRBV3-1 clones after stimulationby BAOMa and BAAM was associated with cytolysis oftumor lines from patient 5 but not from patient 3. Thus,CD8þ TIL clones with specific lytic capacity toward autol-ogous cancer cells were isolated from a metastatic gastricadenocarcinoma and a cholangiocarcinoma.
TCR-transduced lymphocytes redirected to recognizegastric and bile duct cancers restricted to theautologous context by common HLA
Wenext synthesized the genes encoding the TCRs derivedfrom reactive TIL clones isolated from patients 3 and 5, aswell as the dominant TCR found in patient 9 (Fig. 4A), andtransduced these genes into PBMC. TCR-transduced PBMCsfrom patient 3 and 5 were cocultured with autologous celllines in the presence of absence of HLA-blocking antibodiesto define the HLA restriction elements mediating tumorrecognition (Fig. 4B). For patient 3, the TRBV6-1 andTRBV9TCRs seemed to recognize tumors in the context of theHLA-C�0701 and HLA-B�4901 class I alleles, respectively. Forpatient 5, TRBV28 and TRBV3-1 TCRs both seemed torecognize autologous tumor cells in the context of HLA-A�0201.
To define the specificity of the TCRs derived from thereactive clones and to investigate shared reactivities acrosscancer cell lines, large coculture assays were carried out toassess IFN-g responses against a variety of targets, includ-ing autologous cancer cell lines, normal autologous orHLA-matched B cells, and a panel of 17 commerciallyavailable gastrointestinal cancer cell lines that were HLAgenotyped (Supplementary Table S2). The reactivityobserved from patient 3 bulk CD8þ TIL was recapitulatedwith the TRBV6-1 TCR-engineered T cells, specificallyrecognizing the autologous GALMa and GALMd, but not
Turcotte et al.
Clin Cancer Res; 20(2) January 15, 2014 Clinical Cancer Research338
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

GALMb and GALMc, and T cells transduced with theTRBV9 TCR recognizing all four GALM tumor lines (Fig.4C, top: orange bar, TRBV6-1; yellow bar, TRBV9; and redbar, bulk CD8þ). The TRBV6-1 lymphocyte reactivity wasentirely specific to the autologous tumor, despite the factthat the restriction element HLA-C�0701 allele was poten-tially expressed by 13 other allogeneic tumor targets(Supplementary Table S1). For patient 5, the TRBV28TCR-transduced PBMCs recognized the autologous celllines established from this patient’s omental nodule(BAOMa and BAOMb) and malignant ascites (BAAM;Fig. 4C, middle, lighter green bar). The reactivity of theTRBV3-1 was weaker but specific to the autologous tumor(Fig. 4C, dark green bar). Again, although 13 cancer celllines potentially expressed HLA-A�0201, there was noreactivity suggestive of shared antigen recognition by Tcells transduced with TRBV28 and TRBV3-1.In addition, other CD8þ TIL derived from HLA-A�0201
patients or with known HLA-A�0201–restricted epitopes(Patient 9, Patient 10, F5melwithMART1 reactivity, ref. 31;CEA, ref. 33; and MAGE-A3, ref. 27 transduced T cells)failed to recognize biliary adenocarcinoma omentalmetas-tasis (BAOM) and BAAM tumor cell lines. PBMCs engi-neered to express the dominant TCR from patient 9(TRBV7) failed to recognize CALM, even when the cancercell line was transduced to expressed HLA-A�0201, theexpression that it had lost. Finally, native CD8þ TIL frompatients 3, 5, 9, 10, and 11 failed to recognize allogeneictumor cell lines.Altogether, these results showed that it was possible to
redirect PBMC specificity toward gastrointestinal cancerantigens presented by distinct HLA class I alleles, and thatthe antigens recognized in the autologous settingwere likelyunique to each patient.
DiscussionWhether common human epithelial malignancies such
as gastrointestinal cancers harbor tumor-reactive T cells hasbeen debated for decades, with arguments mainly relyingon associative findings (6) rather than direct demonstra-tion. Suggestive evidence has supported the idea that CD8þ
TIL reactive to tumor cell suspensions could be expandedfrom metastatic gastrointestinal cancers (34, 35), but thelack of well-characterized cancer cell lines precluded assaysnecessary to define tumor reactivity and specificity at theTCR-HLAmolecular level. Ten years ago, a colon cancer cellline established from a liver metastasis was used to identifyCD4þ TIL clones reactive to a self-epitope restricted byHLA-DRb1, but required engineering of the cancer cell line forexpression of MHC Class II molecules (36). Here, weestablish the presence of naturally occurring CD8þ TIL ableto specifically recognize autologous metastatic gastric, bileduct, and colon adenocarcinomas by generating new cancercell lines and TIL cultures. Three main observations can bemade fromourfindings: first, tumor-reactiveCD8þTILwerefound at low frequency; second, metastatic gastrointestinalcancer recognition by CD8þ TIL was seen only in theautologous setting, in absence of shared recognition acrossallogeneic cancer cell lines; and third, autologous cancer celllines generated from given patients were not equally rec-ognized by T cells, and in other patients were deficient inMHC-I expression.
Unlike melanoma, only anecdotal responses to systemicimmunomodulation have been reported in patients withmetastatic gastrointestinal cancers (2–5), implying that thesetumors lacked tumor-reactive T cells in sufficient numbers orwith the quality necessary tomediate tumor regression. Oneof the factors that may contribute to the low frequency oftumor-reactive TIL in gastrointestinal cancer is the relatively
Figure 3. Specific CD8þ TIL clonelysis of autologous gastric and bileduct cancer cell lines. Chromium-51 release assays testing cancercell line lysis by the TRBV6-1 andTRBV9 CD8þ TIL clones frompatient (Pt.) 3, and TRBV28 andTRBV3-1 clones from patient 5 indifferent experiments. A, Pt. 3TRBV9 CD8þ TIL clone lysedautologous GALMa and GALMc,whereas the TRBV6-1 sister CD8þ
clones only lysed autologousGALMa andGALMd (B). B, fromPt.5, TRBV28 and TRBV3-1 CD8þ TILclones lysed two autologouscancer cell lines (BAOMa andBAAM). For all clones, reactivitywas specific to autologous tumorcell lines.
T-Cell Recognition of Metastatic Gastrointestinal Cancer
www.aacrjournals.org Clin Cancer Res; 20(2) January 15, 2014 339
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

low number of exomic mutations those tumors generallycarry (14). Whole-exome sequencing studies have reportedan average of approximately 200 mutations per melanomacompared with approximately 55 mutations in gastrointes-tinal cancers, representing less potential opportunities for
antitumor CD8þ T-cell recognition (37). In silico-basedepitope prediction analysis has suggested that 2 to 17HLA-A�0201–mutated epitopes may be generated per coloncancer (38). In addition, even if a productive antitumorimmune response occurred against mutated epitopes, it is
Figure 4. Retroviral transduction of lymphocyteswith four TCRs and assessment of their HLA restriction elements and specificity. A, design of gammaretroviralTCR construct used for transduction of PBMCs with four TCRs derived from patient (Pt.) 3 and Pt. 5 cytolytic TIL clones. The human constant chainswere replaced with mouse constant chains (mTCRa and mTCRb) to reduce mispairing of transduced a and b chains with endogenous TCR chains. Aribosomal skipping motif (2A) is inserted between the a and b chains for separate transcription and expression of the two TCR chains. B, determination of therestriction elements by which TCR react to autologous cancers (Supplementary Table S1 for HLA listing). For TCRs in Pt. 3, TRBV6-1 TCR recognizes itsepitope by HLA-C�0701, whereas TRBV9 is restricted by HLA-B�4901, blocked by Bw4 and not by Bw6, the latter having the potential to block HLA-B�0801.The reactivity of both TCRs in Pt. 5 are not blocked by the anti-BC antibody, and are restricted by HLA-A�0201. The specificity of the Class I and Class IIblocking antibodies was demonstrated in the same experiment by the selective block of known Class I (F5 mel) and Class II (Tyr 450) lymphocyte lines.C, reactivity of native TIL and PBMCs transduced with TCRs from Pt. 3 (top), TCRs from Pt. 5 (middle), TCR that recognize MART1, MAGE A3, andCEA presented by HLA-A�020101, to 17 targets (Supplementary Table S2 for all target tested): 1, GALMa; 2, GALMb; 3, GALMc; 4, GALMd; 5, BAOMa; 6,BAOMb; 7, BAAM; 8, CALM; 9, CALM transduced to express HLA-A�020101; 10, CAPM; 11, CAAWMa; 12, NCI H508 colon cancer (Ca.); 13, SK-CO-1colon Ca.; 14, 624 melanoma TC; 15, SW1463 rectal Ca; 16, KATO III gastric Ca; 17, HCT15 colon Ca; Pt. 3 and Pt. 5 TCR reactivity are restricted to theautologous setting.
Turcotte et al.
Clin Cancer Res; 20(2) January 15, 2014 Clinical Cancer Research340
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

possible that over time, a natural selection of the leastimmunogenic cancer cells occurs, as suggested in animalmodels (39, 40).Here, our ability to establishnewcancer celllines in a limited number of patients with advanced gastro-intestinal cancers consistently supported a low frequency oftumor-reactiveCD8þTIL,butmore samples shouldbe testedto further examine this phenomenon. Higher frequencies ofgastrointestinal tumor-reactive TIL may be found in earlierstages of disease, or in gastrointestinal cancer subsets, such asthose with a high mutation rate due to mismatch repair–gene deficiencies.Our study thus demonstrates that tumor-reactive T cells
with proliferative potential may be isolated from advancedgastrointestinal cancer, provided that suitable tumor targetsare available for testing. Thus far, CD137 (4-1BB), a cellsurface marker of recent TCR engagement, had been usedfor isolation of precursor T cells derived from PBMCs, usingpeptides from known tumor or viral antigens as stimulators(32, 41, 42). Without knowing the antigens recognized bypolyclonal TIL, the use of 4-1BB expression here allowed toisolate polyclonal cytolytic CD8þ T cells, independent oftheir capacity to secrete IFN-g (Figs. 2 and 3). Because thedifficulty in generating cancer cell lines has limited the studyof the immune recognition of epithelial cancers in vitro, itseems critical that new methods be developed to increasethe yield of new cell line establishment, as shown recentlyby the use of a ROCK inhibitor with stromal cells (43, 44).As clinical-grade flow cytometry cell sorting becomes avail-able for enriching cell products in tumor-reactive T cells(45), it may be possible to design adoptive cell transferimmunotherapy for patients with tumors that harbor asmall fraction of tumor-reactive T cells that can be expandedin vitro.By defining the reactivity of CD8þ T cells against gastro-
intestinal cancers at the TCR and the HLA molecular leveland by testing allogeneic reactivity against a comprehensivepanel of HLA-genotyped gastrointestinal cancer cell lines,our study highlighted an additional difference with mela-noma, which is the absence of shared recognition acrossallogeneic tumors (Supplementary Table S2 and Fig. 4). TILin melanoma not only recognize mutated epitopes, butmany self-epitopes, such as cancer testis antigens (MAGE,NY-ESO1, etc.) and melanocyte-differentiation antigens(gp100, MART-1, etc.; refs. 10–13). Although ongoingstudies are aimed at identifying the genes that encode theantigens recognized by gastrointestinal TIL, conceivably,thesemay consist ofmutation-generated epitopes unique toeach tumor, or self-antigens overexpressed by the tumor.Mining exomic mutation expression in this context couldrepresent a powerful tool for defining new tumor antigensrecognized by T cells (14). Thus, for advanced gastrointes-tinal cancers, T-cell based immunotherapy may have to relyon high throughput screening of unique reactive TIL, espe-cially considering the infrequent expression of cancer-testisantigens (46) by gastrointestinal cancers and the potentialtoxicity seen in trial targeting shared self-differentiationantigens such as CEA (47). Further studies should also aimat clarifying whether tumor-reactive CD8þ TIL represent
dominant clonotypes in freshly resected tumors, and ifcurrent in vitro TIL expansion protocols lead to overgrowthof nontumor reactive bystander rather than tumor-reactiveT cells.
Although the genetic heterogeneity found in a giventumor and across distinct metastases in a given patient isnow well established using second generation genomicsequencing (37, 48, 49), our data support that this hetero-geneity can translate into the generation of various cell lineswith the distinct potential of being recognized by autolo-gous CD8þ TIL. For example, in the case of a gastric cancermetastatic to the liver (patient 3), two of four cancer celllines seemed to express the gene encoding an epitoperecognized by a TCR (TRBV6-1) that dominated theCD8þ TIL repertoire and was restricted by the HLA-C�0701(Fig. 4B and C). A second epitope, restricted by HLA-B�4901, was present on the four autologous cancer celllines and recognizedby adistinct TCR(TRBV9) expressedbyless than 1.3% of the CD8þ bulk TIL expanded from thetumor. Adding to the complexity of in vitro assessment oftumor recognition by T cells, the loss of HLA expression bycancer cells—a well-known mechanism of tumor immune-escape (9, 50)—was seen in 2 of the 5 patients inwhich newcancer cell lines were established, and in four of the 17commercially available gastrointestinal cancer cell linestested. MHC expression on paraffin-embedded gastrointes-tinal cancer metastases further suggested frequent deficientexpression in vivo (Fig. 1 and Table 1). Implications of thesefindings for advanced gastrointestinal cancer immunother-apy are 2-fold: first, a nonpolyclonal immunologic app-roach that only targets one tumor antigen is unlikely tomediate sustained tumor regression. Second, the antigenpresentation capacity of cancer cells should be evaluated asa potential biomarker of response to immunotherapy,while strategies that aim at restoring antigen presentationhave to be developed to broaden the use of T-cell basedimmunotherapy.
In conclusion, this study defines at the clonal andmolecular level, the existence of naturally occurring cyto-lytic CD8þ TIL specifically reactive to autologous meta-static gastrointestinal cancers. These findings propose ave-nues for the development of T-cell based immunothera-pies for gastrointestinal cancers by pointing to the mainchallenges to be addressed, such as the low frequency oftumor-reactive TIL, the absence of shared antigen recog-nition across allogeneic tumors, the difficulty in establish-ing suitable tumor targets for the selection of reactive TIL,the heterogeneity of antigen expression by tumors, andthe loss of MHC-I expression by immune-escape tumorvariants.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: S. Turcotte, A. Gros, S.A. RosenbergDevelopment of methodology: S. Turcotte, P.F. Robbins, S.A. RosenbergAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): S. Turcotte, C.-C.R. Lee, J.R. Wunderlich, S.A.Rosenberg
T-Cell Recognition of Metastatic Gastrointestinal Cancer
www.aacrjournals.org Clin Cancer Res; 20(2) January 15, 2014 341
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

Analysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): S. Turcotte, A. Gros, E. Tran, C.-C.R. Lee,S.A. RosenbergWriting, review, and/or revision of the manuscript: S. Turcotte, A. Gros,E. Tran, S.A. RosenbergAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): A. Gros, S.A. RosenbergStudy supervision: S.A. Rosenberg
AcknowledgmentsThe authors thank P. Fetsch and A.C. Filie for cytoimmunohistochemisty
staining and interpretation, L.T. Ngo for technical support, K. Hogan forexpertise in TIL expansion, Q.J. Wang and Y.F. Li for TCR repertoire assess-ment and cloning, A. Mixon and S. Farid for cell sorting by flow cytometry,
S. Adams forMHC genotyping, S.A.Williams for high-resolution whole slidescanning, and N.P. Restifo for thoughtful discussions on T-cell biology andeditorial review.
Grant SupportThis work was supported by the Intramural Research Program of the NIH,
NCI, Center for Cancer Research.The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received June 26, 2013; revised October 23, 2013; accepted November 4,2013; published OnlineFirst November 11, 2013.
References1. JemalA,BrayF,CenterMM, Ferlay J,WardE, FormanD.Global cancer
statistics. CA Cancer J Clin 2011;61:69–90.2. Brahmer JR, DrakeCG,Wollner I, Powderly JD, Picus J, SharfmanWH,
et al. Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacody-namics, and immunologic correlates. J Clin Oncol 2010;28:3167–75.
3. ChungKY,Gore I, Fong L, VenookA, BeckSB,Dorazio P, et al. Phase IIstudy of the anti-cytotoxic T-lymphocyte-associated antigen 4 mono-clonal antibody, tremelimumab, in patients with refractory metastaticcolorectal cancer. J Clin Oncol 2010;28:3485–90.
4. RosenbergSA, LotzeMT,Yang JC,AebersoldPM,LinehanWM,SeippCA, et al. Experience with the use of high-dose interleukin-2 in thetreatment of 652 cancer patients. Ann Surg 1989;210:474–84.
5. Royal RE, Levy C, Turner K, Mathur A, Hughes M, Kammula US, et al.Phase 2 trial of single agent Ipilimumab (anti-CTLA-4) for locallyadvanced or metastatic pancreatic adenocarcinoma. J Immunother2010;33:828–33.
6. Fridman WH, Pages F, Sautes-Fridman C, Galon J. The immunecontexture in human tumours: impact on clinical outcome. Nat RevCancer 2012;12:298–306.
7. Mantovani A,RomeroP,PaluckaAK,Marincola FM. Tumour immunity:effector response to tumour and role of the microenvironment. Lancet2008;371:771–83.
8. Baitsch L, Fuertes-Marraco SA, Legat A, Meyer C, Speiser DE. Thethree main stumbling blocks for anticancer T cells. Trends Immunol2012;33:364–72.
9. Khong HT, Restifo NP. Natural selection of tumor variants in thegeneration of "tumor escape" phenotypes. Nat Immunol 2002;3:999–1005.
10. van der BruggenP, Traversari C, ChomezP, LurquinC, DePE, Van denEynde B, et al. A gene encoding an antigen recognized by cytolytic Tlymphocytes on a human melanoma. Science 1991;254:1643–7.
11. Kawakami Y, Eliyahu S, Delgado CH, Robbins PF, Rivoltini L, TopalianSL, et al. Cloning of the gene coding for a shared human melanomaantigen recognized by autologous T cells infiltrating into tumor. ProcNatl Acad Sci U S A 1994;91:3515–9.
12. Wang RF, Appella E, Kawakami Y, Kang X, Rosenberg SA. Identifi-cation of TRP-2 as a human tumor antigen recognized by cytotoxic Tlymphocytes. J Exp Med 1996;184:2207–16.
13. Guillaume B, Stroobant V, Bousquet-Dubouch MP, Colau D, ChapiroJ, ParmentierN, et al. Analysis of theprocessing of seven human tumorantigens by intermediate proteasomes. J Immunol 2012;189:3538–47.
14. Robbins PF, Lu YC, El-Gamil M, Li YF, Gross C, Gartner J, et al. Miningexomic sequencing data to identify mutated antigens recognized byadoptively transferred tumor-reactive T cells. Nat Med 2013;19:747–52.
15. AtkinsMB, LotzeMT, Dutcher JP, Fisher RI, Weiss G,Margolin K, et al.High-dose recombinant interleukin 2 therapy for patients with meta-static melanoma: analysis of 270 patients treated between 1985 and1993. J Clin Oncol 1999;17:2105–16.
16. Hodi FS, O'Day SJ, McDermott DF, Weber RW, Sosman JA, HaanenJB, et al. Improved survival with ipilimumab in patients with metastaticmelanoma. N Engl J Med 2010;363:711–23.
17. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDer-mott DF, et al. Safety, activity, and immune correlates of anti-PD-1antibody in cancer. N Engl J Med 2012;366:2443–54.
18. Wolchok JD, Kluger H, Callahan MK, Postow MA, Rizvi NA, LesokhinAM, et al. Nivolumab plus ipilimumab in advancedmelanoma. N Engl JMed 2013;369:122–33.
19. Rosenberg SA, Yang JC, Sherry RM, Kammula US, Hughes MS, PhanGQ, et al. Durable complete responses in heavily pretreated patientswith metastatic melanoma using T-cell transfer immunotherapy. ClinCancer Res 2011;17:4550–7.
20. Besser MJ, Shapira-Frommer R, Treves AJ, Zippel D, Itzhaki O,Hershkovitz L, et al. Clinical responses in a phase II study usingadoptive transfer of short-term cultured tumor infiltration lymphocytesin metastatic melanoma patients. Clin Cancer Res 2010;16:2646–55.
21. Ellebaek E, Iversen TZ, Junker N, Donia M, Engell-Noerregaard L, MetO, et al. Adoptive cell therapy with autologous tumor infiltratinglymphocytes and low-dose Interleukin-2 in metastatic melanomapatients. J Transl Med 2012;10:169–80.
22. Radvanyi LG, Bernatchez C, ZhangM, Fox P, Miller P, Chacon J, et al.Specific lymphocyte subsets predict response to adoptive cell therapyusing expanded autologous tumor-infiltrating lymphocytes in meta-static melanoma patients. Clin Cancer Res 2012;18:6758–70.
23. Pilon-Thomas S, Kuhn L, Ellwanger S, JanssenW, Royster E, MarzbanS, et al. Efficacy of adoptive cell transfer of tumor-infiltrating lympho-cytes after lymphopenia induction for metastatic melanoma. J Immun-other 2012;35:615–20.
24. Dudley ME, Wunderlich JR, Shelton TE, Even J, Rosenberg SA.Generationof tumor-infiltrating lymphocyte cultures for use in adoptivetransfer therapy for melanoma patients. J Immunother 2003;26:332–42.
25. Dudley ME, Gross CA, Somerville RP, Hong Y, Schaub NP, Rosati SF,et al. Randomized selection design trial evaluating CD8þ-enrichedversus unselected tumor-infiltrating lymphocytes for adoptive celltherapy for patients with melanoma. J Clin Oncol. 2013 Jun 10. [Epubahead of print].
26. Topalian SL, Solomon D, Rosenberg SA. Tumor-specific cytolysis bylymphocytes infiltrating human melanomas. J Immunol 1989;142:3714–25.
27. Chinnasamy N, Wargo JA, Yu Z, Rao M, Frankel TL, Riley JP, et al. ATCR targeting the HLA-A�0201-restricted epitope of MAGE-A3 recog-nizes multiple epitopes of the MAGE-A antigen superfamily in severaltypes of cancer. J Immunol 2011;186:685–96.
28. Wang QJ, Hanada K, Robbins PF, Li YF, Yang JC. Distinctive featuresof the differentiated phenotype and infiltration of tumor-reactive lym-phocytes in clear cell renal cell carcinoma. Cancer Res 2012;72:6119–29.
29. Brochet X, Lefranc MP, Giudicelli V. IMGT/V-QUEST: the highly cus-tomized and integrated system for IG and TR standardized V-J and V-D-J sequence analysis. Nucleic Acids Res 2008;36:W503–8.
30. Cohen CJ, Zhao Y, Zheng Z, Rosenberg SA, Morgan RA. Enhancedantitumor activity of murine-human hybrid T-cell receptor (TCR) inhuman lymphocytes is associated with improved pairing and TCR/CD3 stability. Cancer Res 2006;66:8878–86.
Turcotte et al.
Clin Cancer Res; 20(2) January 15, 2014 Clinical Cancer Research342
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

31. Johnson LA, Heemskerk B, Powell DJ Jr, Cohen CJ, Morgan RA,Dudley ME, et al. Gene transfer of tumor-reactive TCR confers bothhigh avidity and tumor reactivity to nonreactive peripheral bloodmononuclear cells and tumor-infiltrating lymphocytes. J Immunol2006;177:6548–59.
32. Wolfl M, Kuball J, Ho WY, Nguyen H, Manley TJ, Bleakley M, et al.Activation-induced expression of CD137 permits detection, isolation,and expansion of the full repertoire of CD8þ T cells responding toantigen without requiring knowledge of epitope specificities. Blood2007;110:201–10.
33. Parkhurst MR, Joo J, Riley JP, Yu Z, Li Y, Robbins PF, et al. Charac-terization of genetically modified T-cell receptors that recognize theCEA:691-699 peptide in the context of HLA-A2.1 on human colorectalcancer cells. Clin Cancer Res 2009;15:169–80.
34. Hom SS, Rosenberg SA, Topalian SL. Specific immune recognition ofautologous tumor by lymphocytes infiltrating colon carcinomas: anal-ysis by cytokine secretion. Cancer Immunol Immunother 1993;36:1–8.
35. Turcotte S, Gros A, Hogan K, Tran E, Hinrichs CS,Wunderlich JR, et al.Phenotype and function of T cells infiltrating visceral metastases fromgastrointestinal cancers and melanoma: implications for adoptive celltransfer therapy. J Immunol 2013;191:2217–25.
36. Maccalli C, Li YF, El-Gamil M, Rosenberg SA, Robbins PF. Identifi-cation of a colorectal tumor-associated antigen (COA-1) recognized byCD4(þ) T lymphocytes. Cancer Res 2003;63:6735–43.
37. Vogelstein B, Papadopoulos N, Velculescu VE, Zhou S, Diaz LA Jr,Kinzler KW. Cancer genome landscapes. Science 2013;339:1546–58.
38. Segal NH, Parsons DW, Peggs KS, Velculescu V, Kinzler KW, Vogel-stein B, et al. Epitope landscape in breast and colorectal cancer.Cancer Res 2008;68:889–92.
39. DuPage M, Mazumdar C, Schmidt LM, Cheung AF, Jacks T. Expres-sion of tumour-specific antigens underlies cancer immunoediting.Nature 2012;482:405–9.
40. Matsushita H, Vesely MD, Koboldt DC, Rickert CG, Uppaluri R,Magrini VJ, et al. Cancer exome analysis reveals a T-cell-dependentmechanism of cancer immunoediting. Nature 2012;482:400–4.
41. Watanabe K, Suzuki S, Kamei M, Toji S, Kawase T, Takahashi T, et al.CD137-guided isolation and expansion of antigen-specific CD8 cellsfor potential use in adoptive immunotherapy. Int J Hematol 2008;88:311–20.
42. Han S, Huang Y, Liang Y, Ho Y, Wang Y, Chang LJ. Phenotype andfunctional evaluation of ex vivo generated antigen-specific immuneeffector cells with potential for therapeutic applications. J HematolOncol 2009;2:34.
43. Liu X, Ory V, ChapmanS, YuanH, AlbaneseC, KallakuryB, et al. ROCKinhibitor and feeder cells induce the conditional reprogramming ofepithelial cells. Am J Pathol 2012;180:599–607.
44. Yuan H, Myers S, Wang J, Zhou D, Woo JA, Kallakury B, et al. Use ofreprogrammed cells to identify therapy for respiratory papillomatosis.N Engl J Med 2012;367:1220–7.
45. Jaye DL, Bray RA, Gebel HM, Harris WA, Waller EK. Translationalapplications of flow cytometry in clinical practice. J Immunol 2012;188:4715–9.
46. Simpson AJ, Caballero OL, Jungbluth A, Chen YT, Old LJ. Cancer/testis antigens, gametogenesis and cancer. Nat Rev Cancer 2005;5:615–25.
47. Parkhurst MR, Yang JC, Langan RC, Dudley ME, Nathan DA, FeldmanSA, et al. T cells targeting carcinoembryonic antigen can mediateregression of metastatic colorectal cancer but induce severe transientcolitis. Mol Ther 2011;19:620–26.
48. Campbell PJ, Yachida S, Mudie LJ, Stephens PJ, PleasanceED, Stebbings LA, et al. The patterns and dynamics of genomicinstability in metastatic pancreatic cancer. Nature 2010;467:1109–13.
49. Gerlinger M, Rowan AJ, Horswell S, Larkin J, Endesfelder D,Gronroos E, et al. Intratumor heterogeneity and branched evolutionrevealed by multiregion sequencing. N Engl J Med 2012;366:883–92.
50. Aptsiauri N, Cabrera T, Garcia-Lora A, Lopez-Nevot MA, Ruiz-CabelloF, Garrido F. MHC class I antigens and immune surveillance intransformed cells. Int Rev Cytol 2007;256:139–89.
T-Cell Recognition of Metastatic Gastrointestinal Cancer
www.aacrjournals.org Clin Cancer Res; 20(2) January 15, 2014 343
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736

2014;20:331-343. Published OnlineFirst November 11, 2013.Clin Cancer Res Simon Turcotte, Alena Gros, Eric Tran, et al. Refractory to Chemotherapy
T Cells in Metastatic Gastrointestinal Cancer+Tumor-Reactive CD8
Updated version
10.1158/1078-0432.CCR-13-1736doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2013/11/11/1078-0432.CCR-13-1736.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/20/2/331.full#ref-list-1
This article cites 49 articles, 22 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/20/2/331.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/20/2/331To request permission to re-use all or part of this article, use this link
on May 28, 2020. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 11, 2013; DOI: 10.1158/1078-0432.CCR-13-1736







