Download - Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 1/48
Page 1 of 48
Construction of growthdefective mutant strains of
Enterococcus faecium
BIOC3002 Research Project
Michael [email protected]
Supervisor: Dr. Sean Nair

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 2/48
Page 2 of 48
DECLARATIONOFOWNERSHIP
BIOC3003/27 / BIOL3004/3005 Academic Year 2009-2010
This submission is a result of my own work.
All help and advice, other than that received by tutors, has been acknowledged, and primary and secondary sources of information have been properly attributed.
Should this statement prove to be untrue, I recognise the right of the Board of Examinersto recommend what action should be taken in line with the University’s regulations.
I acknowledge that the Biological Sciences and Molecular Biosciences use the Turn ItIn® plagiarism detection system and that my work may be submitted to Turn It In®.
Name in block capitals ____________________________________
Signed _________________________________________________
Dated __________________________________________________

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 3/48
Page 3 of 48
TableofContents:
Abbreviations...................................................................................... 4
Abstract .............................................................................................. 5
Introduction......................................................................................... 6
Material and Methods....................................................................... 15
Results ............................................................................................. 23
Discussion and Future Directions..................................................... 42
References ....................................................................................... 47

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 4/48
Page 4 of 48
Abbreviations
KAN – Kanamycin
ERY – Erythromycin
MIC – Minimum Inhibitory Concentration
BHI – Brain Heart Infusion
BHA – Brain Heart Infusion (3.7%w/v) + Agar (1.2%w/v)
THB – Todd-Hewitt Broth
NaCl – Sodium Chloride
O/N – Overnight Culture
ATc – Anhydrotetracycline
PBS -- Phosphate Buffer Saline
PCR -- Polymerase Chain Reaction
dNTPs -- Deoxynucleotide triphosphates

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 5/48
Page 5 of 48
Abstract
Enterococcus faecium is currently one of the primary causes of nosocomial
infectionsi and has become a major burden to hospitals, due to the development
of resistance against the antimicrobials and antibiotics used to treat these
infectionsii. Research is now being directed at identifying new treatments and
approaches to prevent Enterococcal infections by using various genetic tools to
locate and identify genes that help develop and provide resistance to E. faecium.
In this study transposon mutagenesis was used to construct a library of random
mutants, which were screened for specific phenotypes such as loss of antibiotic
resistance or inability to grow on certain media incorporating stressors
encountered by the bacterium. The transposon delivery vector used in this study
was a mariner-based transposon system carried on two different plasmids, which
were introduced into the E. faecium strain E1162 via electroporation. A
transposition procedure was utilised to induce transposon mutagenesis. Once a
mutant had been identified attempts were made to locate the affected gene via
different gene amplification processes for potential sequencing and further
research. Several other experiments such as producing growth curves for E.
faecium different media and determining minimal inhibitory concentrations of
different antimicrobial agents were also carried out, to learn more about the
bacterium Enterococcus faecium.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 6/48
Page 6 of 48
Introduction
Enterococci were once thought to be harmless commensal bacteria found as
members of the human intestinal flora, which only very rarely showed any
pathogenic effectsiii. Today the species E. faecalis and E. faecium of the gram-
positive Enterococcus genus are emerging as two of the major causes of
nosocomial infections. Enterococci were first described and investigated at the
end of the 19th centuryiv, and have since become a source of major concern due
to the increase is the number of infections recorded and their ability to develop
different antimicrobial and antibiotic resistancesv. The discovery of a rise in
ampicillin and vancomycin resistant strains of E. faeciumvi, correlated with an
increase in nosocomial E. faecium infections recorded in hospitals throughout the
US and Europe. Enterococcal infections occur in the urinary tract and intra-
abdominal cavityvii but may also cause endocarditis, meningitis and sepsisviii in
patients with prolonged antibiotic treatment, long periods of hospitalisation and/or
weakened immune systems. The main treatment method to date has been the
use of different antibiotics, but treatments vary in their effectiveness due to
antibiotic resistance interfering with synergic therapy.
Naturally the main aim of research in this area should be to try and discover new
treatment methods against these infections, but before this can be achieved one
must first understand how E. faecium grows, survives and why it is pathogenic.
One way of achieving this is to identify specific genes, which allow certain strains

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 7/48
Page 7 of 48
of E. faecium to grow and survive in demanding environments, but also which
genes allow them to develop antibiotic resistance. Once these genes have been
identified, further research can be focused on how the gene operates, what it
encodes and if the gene or its products can be an affective target for treatment.
There are many different methods, which can be used to investigate the role of
certain genes in an organism. The technique used in this study is known as
transposon mutagenesis. Class II Transposons are sequences of DNA, which
are able to ‘jump’ from one location to another by using a ‘cut and paste’
mechanism thanks to the enzyme transposase. In the simplest cases only two
vital structures are needed for transposition, short inverted terminal repeats
(ITRs) found at either end of a DNA sequence and a functional tpase gene. The
expression of tpase leads to the production of the transposase enzyme, which
has the ability to target, cut and ligate sequences of DNA. It functions by
recognising the ITRs, cutting and then binding both ends of the transposon to
form a ‘loop structure’. This allows the whole transposon to migrate to a different
location (e.g. different gene or chromosome), where transposase cuts the
insertion site to form staggered ends so the transposon can be inserted (‘paste’)
and ligated to the target DNA. This mechanism is found naturally in many
different organisms (including humans) and acts as a way to form genetic
variation, but has also been adopted and modified for use in research as a
genetic tool known as transposon mutagenesis.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 8/48
Page 8 of 48
The idea is that transposons can be used to rapidly create a large library of
different mutants, where the transposon is cut from its original location and
inserted randomly into the host genome, causing mutations to host genes.
Mutants can then be investigated and screened for a certain growth defect or
phenotype (e.g. loss of antibiotic resistance). Due to the structure of transposons,
they can be genetically modified to carry small genes (e.g. in this study an
erythromycin resistance gene: ermC ), which can act as a selective marker for
artificial selection.
Different transposons have been used in different studies depending on their
overall goal and type of organism in question. In this study the transposon should
preferentially have a high rate of transposition, insertions should occur at random
(no bias towards certain sequences) to increase the chances of different mutants
and a high frequency of single insertions. Single insertions are important
because the process of determining which single gene mutation is responsible
for the change in phenotype is more straightforward. If there were a host of
random insertions into various gene loci it would be could become very
complicated to try and identify, which gene corresponds to which phenotype. For
these reasons the Himar1 mariner transposon delivery system integrated into
plasmids pMC38 and pMC39ix, was selected as a foundation. However in this
study several important genetic modifications to the before mentioned
transposon delivery vectors (pMC38/39) have been made. Plasmids pPZ4 and
pPZ114-5 (Zhang P., 2009, Eastman Dental Institute) are derivatives of pMC38,

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 9/48
Page 9 of 48
but expression of the tpase gene is now regulated by the anhydrotetracycline
(ATc) inducible Pxyl/tetO promoter from pKOR1x, consisting of a xyl promoter
and tet operator regulated by the Tet repressor. Point mutations to pPZ4’s
promoter have been used to prevent basal expression and increase ATc
selectivity to produce the vector pPZ114-5. Based on previous studies different
concentrations of ATc have to be used for different bacteria, due to the inhibitory
effect ATc has on bacterial growth. An excessive amount of ATc may increase
the level of transposon mutagenesis but no colonies will grow. On the other hand
too little ATc may not produce the desired rate of transposon mutagenesis, so a
balance has to be established between tpase expression and inhibiting E.
faecium growth.
Figure 1: Physical map of plasmid pPZ4/pP114-5 containing the transposon element (adapted
from Cao, et al., 2007).
pPZ4/pPZ114-5

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 10/48
Page 10 of 48
After transposon mutagenesis has taken place and a mutant strain has been
found, the number and loci of transposon inserts must be identified so the
mutated gene can be sequenced for further research. Two different methods
were used in this study, both using the polymerase chain reaction (PCR) to
amplify the transposon flanking sequences. The transposon number and location
can be determined by amplifying the DNA sequences either side of the
transposon rather than actually amplifying the transposon allows, which only
shows if there is a transposon present in the host genome. The first method
(Kwon YM and Ricke SC, 2000) requires a ligation step before the amplification
step, where ‘Y linkers’ are ligated to fragmented genomic DNA. The
endonuclease used in this study was Taq1α because it will frequently cut due to
its common recognition site, however it is known that Taq1α does not any part of
the transposon sequence. The Y linkers have a 3’ overhang and a non-
complementary sequence on the opposite strand at the 5’ regionxi to prevent the
linker annealing with itself. The Y linker primer used to amplify the transposon
flanking sequences consists of this region of DNA, which means that the primer
cannot actually anneal to the linker at first. A transposon specific primer (Tn) is
used to select for sequences of DNA, which have the transposon sequence. This
means that in the first cycle of PCR the only fragments containing the transposon
will be amplified and DNA synthesis occurs from the end region of the of the
transposon until the end of the Y linker regions, allowing the Y linker primer to
bind to the newly synthesised region, since the Y linker primer is complementary
to the extended sequence. In the second cycle, DNA synthesis takes place from

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 11/48
Page 11 of 48
the Y linker primer to where the transposon specific primer, producing dsDNA of
the transposon flanking sequences. The advantage of using this method is that
many different independent transposon-flanking sequences can be amplified
from a mutant. The different steps involved in this protocol are summarised in the
figure below (Figure 2).
Figure 2: Figure shows steps involved in amplifying transposon-flanking sequences using Y
linkers, Y linker primer and a transposon-specific primer (Tn). (Adapted from Kwon YM, Ricke
SC., (2000) Efficient amplification of multiple transposon-flanking sequences. J Microbiol
Methods. 2000 Aug;41(3):197.)
The second method used in this study was developed by Cao M, et al . (2007),
where a form of nested PCR (2 PCR runs, where the second round amplifies the
products from the first cycle) was used to try and identify the transposon insertion
sites by again amplifying the transposon flanking sequences. For both rounds a
combination of an arbitrary primer and a transposon specific primer was used to

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 12/48
Page 12 of 48
amplify both ends of the fragment. The combination of different sets of primers
used is this study are shown in Figure 3 B. Arbitrary primers are oligonucleotides
synthesised to have a region with a random sequence (shown in Table 2 as N –
random nucleotide), allowing these random primers to bind at different sites on
the chromosome. These arbitrary primers point in the opposite direction to the
transposon specific primers, which means that if a combination of the two
primers are close enough together double stranded PCR product will be
synthesised. The second round of PCR is designed to only amplify the products
from the first containing transposon flanking sequences. This is achieved by
using another arbitrary primer (in this study Arb3), which consists of the same
known sequence as the arbitrary primers used in the first round (STAPHArb1 and
STAPHArb2). This means that Arb3 will most likely bind to the complementary
strands produced in the first round, which should contain the transposon flanking
sequence. The transposon specific primer points in the opposite direction to
Arb3, again allowing the amplification of double stranded PCR product, which
can be visualised by using gel electrophoresis. Through this mechanism
transposon flanking sequences are copied and amplified however this method is
thought to be not as accurate as the Y linker procedure because many products
of different lengths, which may or may not contain the transposon flanking
sequence may be produced.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 13/48
Page 13 of 48
A
Figure 3: A - Physical map of transposon element and transposon specific primers used to
amplify transposon-flanking sequences. (Adapted from Cao, et al., 2007). B – Different
combinations of arbitrary and transposon specific primers used in both rounds of PCR to test for
optimal results.
For selection purposes in other parts of this study it was necessary to examine
different growth properties of the species E. faecium, such as determine the
concentrations of the antibiotic kanamycin and common salt (NaCl) needed to
inhibit growth of E. faecium. The concentration at which no bacterial growth is
observed is known as the minimal inhibitory concentration. Sodium chloride
interferes with bacterial growth by increasing the osmotic stress the bacterium
has to withstandxii. Studies on other bacteria have shown that bacteria respond to
this osmotic stress by inducing the expression of genes, which result in the
synthesis of compatible solutesxiii, which protect the organism. Kanamycin on the
other hand is aminoglycoside antibiotic, which inhibits growth by interfering with
bacterial translation mechanismsxiv. The way the growth curve of E. faecium
A
B

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 14/48
Page 14 of 48
E1162 changed in different media was also studied and the two types of media
used were BHI broth and BHI + NaCl broth.
The main aim of this study was to create a library of E. faecium E1162 mutants
using transposon mutagenesis via the Himar1 mariner transposon delivery
system, where mutants were screened for any growth defects or phenotypes on
specific growth media (e.g. BHA+NaCl) and loss of specific antimicrobial
resistance (e.g. kanamycin). Once a growth defect had been identified, attempts
were made to locate and sequence the non-functioning or mutated gene in the
mutant for use in further research.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 15/48
Page 15 of 48
MaterialandMethods
Bacterial strains, plasmids, and growth conditions
The strain of Enterococcus faecium used in this study was E1162, grown in
either brain heart infusion (3.7%w/v; Oxoid) or Todd-Hewitt (3.64%w/v; Oxoid)
broth or agar (1.2%w/v; Fluka Analytical) at 30°C, unless stated otherwise. The
antibiotics, kanamycin (Melford K0126) and erythromycin (Sigma-Aldrich E5389)
were used at concentrations of 1mg/ml and 40µg/ml respectively, unless stated
otherwise. The growth of Enterococci was determined by measuring the optical
density at 600nm (OD600) using 1ml cuvettes in a PharmaciaBiotech Ultraspec
2000 spectrophotometer.
Table 1: Bacterial Strains and Plasmids used in this study.
Strain / Plasmid Relevant Characteristics Reference
E. coli DH5α Cloning host Lab Stock, UCLEastman Dental Institute
E. faecium E1162 Clinical isolate of the epidemic clonal complex 17 Heikens, et al., 2008
pPZ4 pM38 derivative, promoter replaced with Pxyl/tetO
promoter from pKOR1
P. Zhang, Unpub., UCL
Eastman Dental Institute
pPZ114-5 pPZ4 derivative, higher ATc selectivity due to point
mutations (tagagt in -10 sequence replaced with
tataat)
P. Zhang, Unpub., UCL
Eastman Dental Institute
Plasmids used in this study were pPZ4 and pPZ114-5, derived from pMC38/39
(Cao, et al ., 2007). Both have the anhydrotetracycline (ATc) inducible promoter
Pxyl/tetO from pKOR1 (Bae and Schneewind, 2006), regulating transposase
gene expression. Point mutations (tagagt in -10 sequence replaced with tataat) in
pPZ114-5’s promoter region have been used to prevent basal expression and

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 16/48
Page 16 of 48
increase ATc selectivity. This means that the plasmid pPZ114-5 should have
produced a higher number of mutants compared to pPZ4, which has a ‘leaky’
promoter. Plasmids were purified from E. coli DH5α cells using NBS Biologicals
Spin Column Miniprep kit according to the manufacturers protocol. To confirm
both plasmids were successfully purified, restriction enzyme digests using
endonuclease XbaI (New England BioLabs) were carried out following the
manufacturer’s protocol and the resulting fragments were separated via gel
electrophoresis and observed under UV light by using the Alphamanager (Alpha
Innotech). All gel electrophoresis procedures used a 1% agarose gel (Sigma-
Aldrich) with 0.1 µg/ml ethidium bromide, 2 µl of 1x loading buffer (0.05%
bromophenol blue, 5% glycerol in ddH2O) per sample and 1x Tris-Borate-EDTA
(TAE) buffer (40mM Tris-borate and 1mM EDTA) was applied at ~100 V and
100mA for 30 to 40 minutes. 1 Kb plus DNA ladder (Invitrogen; 10787-018) and
1kb DNA ladder (Promega; G5711) were used as a size reference.
Plasmid Electroporation into E1162
E. faecium cells were grown in THB at 37°C and once in stationary phase (no
change in OD600 at ~1.00) were spun down at 5000rpm for 15 min at 4°C using
the SORVALL RC5B PLUS centrifuge, resuspended in 10% glycerol and stored
at -70°C. Plasmids pPZ4 and pPZ114-5 were transformed into these
electrocompetent E1162 cells via electroporation using chilled 1ml cuvettes and
the BIO-RAD Gene Pulser II set at a voltage of 2.5 kV, capacitance of 2 µF and
resistance of 400-500 Ω. Pulsed cells were immediately resuspended in 1ml of

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 17/48
Page 17 of 48
BHI broth and incubated at room temperature for 30 minutes and then spread
onto BHA + KAN (1mg/ml) + ERY (40µg/ml) plates to select for cells that had
taken up either plasmid. Plates were incubated at 30°C for 7 days due to slow
growth. To confirm either pPZ4 or pPZ114-5 had been successfully taken up, a
plasmid preparation was carried out using the NBS Biologicals Spin Column
Miniprep kit and appropriate protocol. Additional enzymes were used to
hydrolyse the E. faecium cell wall include 10µl of lysozyme (50mg/ml), 10µl of
lysostaphin (20mg/ml) and 10µl of mutanolysin (50mg/ml). The purification
products were then digested with the endonuclease XbaI and run on an agarose
gel. The fragments were visualized under UV light and compared to a positive
control (pPZ4+XbaI or pPZ114-5+XbaI).
Transposon Mutagenesis
Transposon mutagenesis was used to produce mutant strains of E. faecium
strain E1162. Different concentrations of anhydrotetracycline (ATc) were used to
induce the expression of promoter Pxyl/tetO. The transposition protocol was
adapted for E. faecium E1162 from Cao, et al. (2007) and Zhang (unpublished,
Eastman Dental Institute, 2009). E. faecium containing either pPZ4 or pPZ114-5
were inoculated into 10ml BHI broth, 40µg/ml erythromycin and 1mg/ml
kanamycin and incubated at 30°C overnight (O/N culture). The overnight cultures
were used to inoculate fresh BHI at a ratio of 1 in 100 into 15ml of BHI broth,
40µg/ml ERY and different ATc concentrations (1.0µg/ml, 1.5µg/ml and
2.0µg/ml). The culture was incubated at 30°C until the OD600 had reached 0.2

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 18/48
Page 18 of 48
(2~3 hours) and then centrifuged at 5000rpm, using the Eppendorf centrifuge
5804R for 10 minutes at 21°C. Pellet was first resuspended in sterile PBS, spun
down again and then resuspended in BHI broth and 40µg/ml ERY. To remove
the plasmid from the cells the culture was then incubated at 45°C overnight,
since the origin of replication found in the plasmid ( pE194ts ori ) is temperature
sensitivexv. Temperatures above 37°C should prevent plasmid replication causing
a loss of the plasmids in E. faecium cells. The next day a 10-fold serial dilution
series was carried out, where different dilutions (neat to 10-9) were plated onto
BHA and BHA+ERY (40µg/ml) plates, which were incubated at 43°C overnight.
To check for plasmid retention single colonies were patched first onto BHA+KAN
(1mg/ml) and then BHA+ERY (40µg/ml) plates and incubated at 30°C overnight.
Identifying growth defective E. faecium clones
Replica plating was used to determine if any of the colonies from the
transposition showed any growth defective phenotype, when plated on
BHA+NaCl (7g of sodium chloride per 100ml of BHI broth) plates. A 10-fold
dilution series of the transposition culture was carried out with PBS and the 10-6
culture was plated out onto BHA plates and incubated at 37°C overnight. All
colonies from a BHA plate were then replicated onto a BHA+NaCl plate via the
Sigma Replica Plater. Once all plates were replicated, both BHA and BHA+NaCl
plates were incubated at 37°C overnight. Corresponding BHA and BHA+NaCl
plates were then compared to check for any colonies, which successfully grew on
the BHA plate but not on the BHA+NaCl plate.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 19/48
Page 19 of 48
Amplification of transposon-flanking sequences
Genomic DNA was obtained from potential growth defective mutant colonies by
using the Qiagen Pre-treatment for Gram-Positive Bacteria kit following the
corresponding protocol from DNeasy Blood/Tissue Handbook. The resulting DNA
concentration was measured by using a Nano-Drop Spectrophotometer ND-
1000. Restriction enzyme digests of genomic DNA were then carried out with the
endonuclease TaqIα (New England BioLabs) for 4 hours at 65°C. The digests
consisted of 2µl of TaqIα, 4µl of 10x NEBuffer 4 (New England Labs), 0.4µl of
100x Bovine Serum Albumin (BSA), 2.61µl of genomic DNA and 30.99µl of H2O.
Two different types of DNA amplification protocols were used in this experiment
via PCR using the Biometra T3000 ThermoCycler. All PCR products were run on
a 1% agarose gel to check for the presence of DNA.
Table 2: Oligonucleotides used in this study
Oligonucleotides Sequence Source
Y Linker 1 TTTCTGCTCGAATTCAAGCTTCTAACGATGTACGGGGACAT Sigma-Aldrich
Y Linker 2 CGATGTCCCCGTACATCGTTAGAACTACTCGTACCATCCACAT Sigma-Aldrich
Y linker primer CTGCTCGAATTCAAGCTTCTSigma-Aldrich
Tn Primer GGAATCATTTGAAGGTTGGTASigma-Aldrich
STAPH Arb1 GGCCACGCGTCGACTAGTACNNNNNNNNNNGTAAT Sigma-Aldrich
STAPH Arb2 GGCCACGCGTCGACTAGTACNNNNNNNNNNGATCA Sigma-Aldrich
Arb3 GGCCACGCGTCGACTAGTSigma-Aldrich
Maq 255 CAGTACAATCTGCTCTGATGCCGCATAGTT Sigma-Aldrich
Maq 269 GCTCTGATAAATATGAACATGATGAGTGAT Sigma-Aldrich
Maq 256 TAGTTAAGCCAGCCCCGACACCCGCCAACA Sigma-Aldrich
Maq 270 TGTGAAATACCGCACAGATGCGAAGGGCGA Sigma-Aldrich

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 20/48
Page 20 of 48
The first protocol devised by Kwon YM and Ricke SC (2000) required a ligation
step before amplification, where in a total volume of 20µl, 1µl of Y linker mixture
(Y linker 1 + Y linker 2) was ligated to 40ng of digested genomic DNA using 1µl
of T4 DNA ligase (New England BioLabs, 1unit/µl) and 2µl of 10x T4 DNA ligase
Buffer (New England BioLabs). The mixture was incubated at 14°C for a
minimum of 4 hours. PCR amplification mixture had a total volume of 50µl where,
5µl of 10x Taq PCR buffer (New England BioLabs), 1µl of Y linker primer
(350ng/µl), 1µl of Tn primer (350ng/µl), 3µl of a mix of dNTPs (10mM) and 2µl of
template DNA were initially incubated at 95°C for 2min. A hot-start was
performed where 1µl of Taq DNA polymerase (New England BioLabs; 5unit/µl)
was added at 80°C to prevent nonspecific primingxvi, followed by 30 DNA
amplification cycles of 95°C for 30s to denature the DNA, 58°C for 1 minute to
anneal primers to the template DNA and 70°C for 1 minute for the DNA
polymerase to extend the primers followed by an incubation at 70°C for 5
minutes.
The second DNA amplification protocol was designed by Cao et. al. (2007),
where two rounds of PCR are required. The first round had a total volume of 25µl
where 1µl of genomic DNA (~95 ng/µl), 1.25µl of either arbitrary primer (STAPH
Arb1 or STPAH Arb2), 1.25µl of either transposon primer (Maq255 or Maq 269),
0.5µl of dNTPs (5mM), 2.5µl of dimethyl sulphoxide (DMSO; Sigma-Aldrich),
0.25µl of Taq DNA polymerase (5unit/µl) and 2.5µl of 10x Taq PCR buffer was

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 21/48
Page 21 of 48
incubated at 95°C for 5 minutes, followed by 30 cycles of 95°C for 30s to
denature the DNA, 42°C for 45s to anneal primers to the template DNA and 72°C
for 1 minute for the DNA polymerase to extend the primers followed by a final
incubation at 72°C for 5 minutes. The second round of PCR had a total volume of
25µl where 5µl of template DNA (from 1st round), 1.25µl of arbitrary primer
(Arb3), 1.25µl of either transposon primer (Maq256 or Maq 270), 0.5µl of dNTPs
(5mM), 2.5µl of dimethyl sulphoxide (DMSO), 0.25µl of Taq DNA polymerase
(5unit/µl) and 2.5µl of 10x Taq PCR buffer were incubated at 95°C for 30s,
followed by 40 cycles of 95°C for 30s to denature the DNA, 45°C for 30s to
anneal primers to the template DNA and a final incubation at 72°C for 1 minute
for the DNA polymerase to extend the primers.
Determining Minimal Inhibitory Concentrations
Minimal inhibitory concentrations (MICs) of the antibiotic kanamycin (Sigma-
Aldrich) and sodium chloride (NaCl) for E. faecium were determined by using an
agar dilution method. Dilutions of kanamycin (32, 64, 128, 256, 512 and 1024
µg/ml) and NaCl (4, 5, 6, 7, 8, 9 and 10 g per 100ml of agar) were made in BHI
agar. Overnight cultures of E. faecium wild type E1162 (BHI+KAN+ERY) were
prepared and diluted 10-fold in sterile PBS and 50µl of the 10-6 dilution was
spread onto the agar plates with different kanamycin and NaCl concentrations,
along with BHI agar plates as a control. Plates were then incubated at 37°C for 3
days. Number and size of colonies was recorded and the lowest concentration

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 22/48
Page 22 of 48
plates that showed no visible colony growth were established as the minimal
inhibitory concentration.
Comparative Growth Curves
The culture to be examined was prepared in a 10ml BHI broth overnight culture
at 30°C. The next day 30ml of the specific growth media (BHI or BHI+NaCl) for
each sample was pre-warmed to 37°C. 1 in 20 dilutions were made of the O/N
culture, so the OD600 could be measured using the PharmaciaBiotech Ultraspec
2000 spectrometer. Culture was then inoculated into the corresponding pre-
warmed media sample so the initial OD600 reading was ~0.01. Samples were
incubated at 37°C and the OD600 was measured at certain time intervals (0, 90,
130, 170, 210, 250, 290 and 330 minutes).

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 23/48
Page 23 of 48
Results
Determining Minimal Inhibitory Concentrations
It was important to first determine the minimal inhibitory concentration of
kanamycin and sodium chloride for wild type E. faecium E1162, because these
two antimicrobials were used to select for specific colonies during different
stages of the study, such as the selective plating of transposition colonies or
selecting cells that have taken up the plasmids after electroporation. For these
selection processes to be effective, the MIC for both KAN and NaCl must be
known. For example the concentration of kanamycin used for selecting colonies
that have taken up the plasmid was 1mg/ml. E. faecium wild type is known to
have a certain level of natural resistance to kanamycin, so if the wild type was
able to grow in these conditions the selection process would be ineffective, since
both cells with and without plasmid would grow. By determining the concentration
at which the wild type does not grow, but colonies with the selective marker (e.g.
KAN resistance gene found on plasmid) do, selection was carried out. The MIC
of kanamycin and sodium chloride for E1162 was determined by growing E.
faecium on agar containing increasing concentrations of both antimicrobials. The
results for both kanamycin (Figure 2) and sodium chloride (Figure 3) minimal
inhibitory concentrations are shown in the following figures.
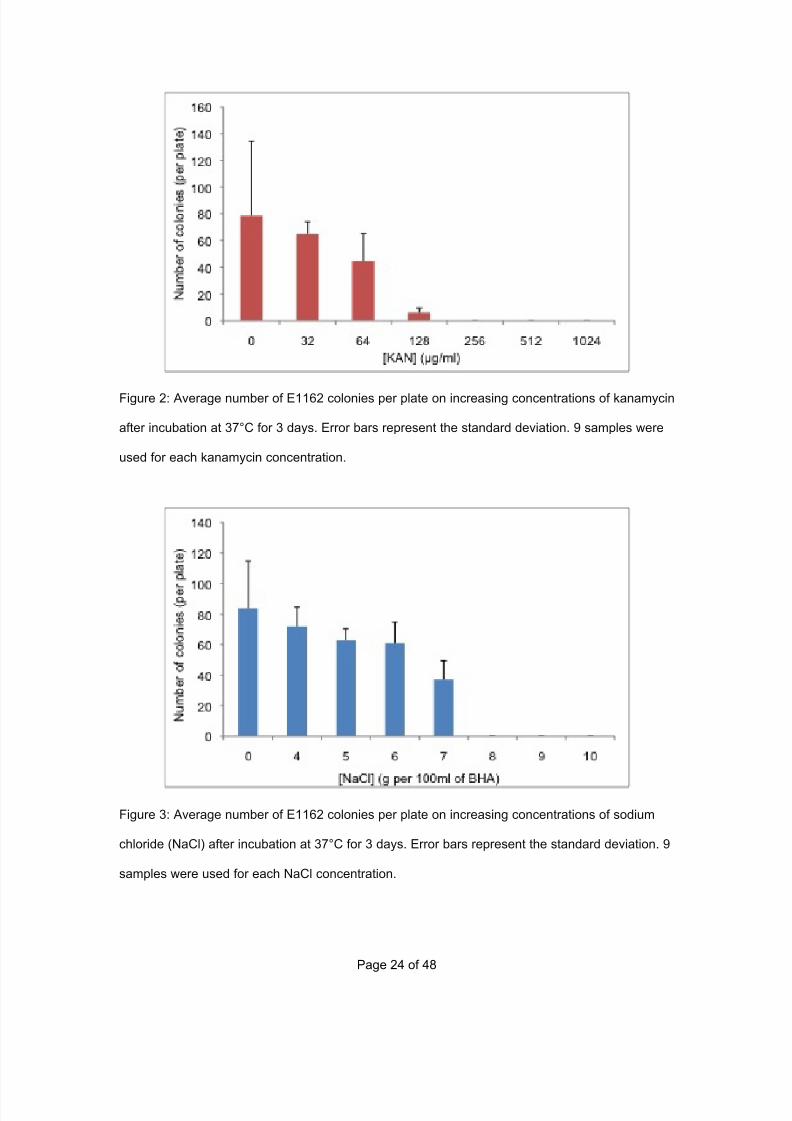
7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 24/48
Page 24 of 48
Figure 2: Average number of E1162 colonies per plate on increasing concentrations of kanamycin
after incubation at 37°C for 3 days. Error bars represent the standard deviation. 9 samples were
used for each kanamycin concentration.
Figure 3: Average number of E1162 colonies per plate on increasing concentrations of sodium
chloride (NaCl) after incubation at 37°C for 3 days. Error bars represent the standard deviation. 9
samples were used for each NaCl concentration.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 25/48
Page 25 of 48
The minimal inhibitory concentration is the concentration at which no E. faecium
colony growth is observed. This means that for E1162 wild type the MIC of
kanamycin is 256µg/ml and sodium chloride is 8g per 100ml of BHA. These
results can be used for example in the screening for uptake of plasmid, where
any concentration of kanamycin equal to or greater than 256µg/ml will inhibit the
growth of E1162 wild type.
Plasmid Electroporation into E1162
For transposon mutagenesis to be carried out it was necessary to introduce the
vectors carrying the transposon delivery system. Plasmids pPZ4 and pPZ114-5
were first purified from E. coli DH5α cells provided by Zhang P. (UCL Eastman
Dental Institute, 2009). Gel electrophoresis was then used to confirm that the
procedure was successful, by digesting the plasmids with the endonuclease XbaI
and comparing the resulting fragments to a positive control (pPZ4 MIDI + XbaI
provided by Wright J., UCL Eastman Dental Institute, 2010). By comparing the
fragments from the digestion products (Figure 4) with the positive control (Lane
6) I was able to confirm that the purified plasmids were pPZ4 (Lane 5) and
pPZ114-5 (Lane 4). Uncut plasmids (Lane 2 and 3) only form one band (larger
than 12,000bp), but when digested with XbaI both plasmids form 3 distinct bands
at ~5500bp, ~2200bp and ~1900bp.
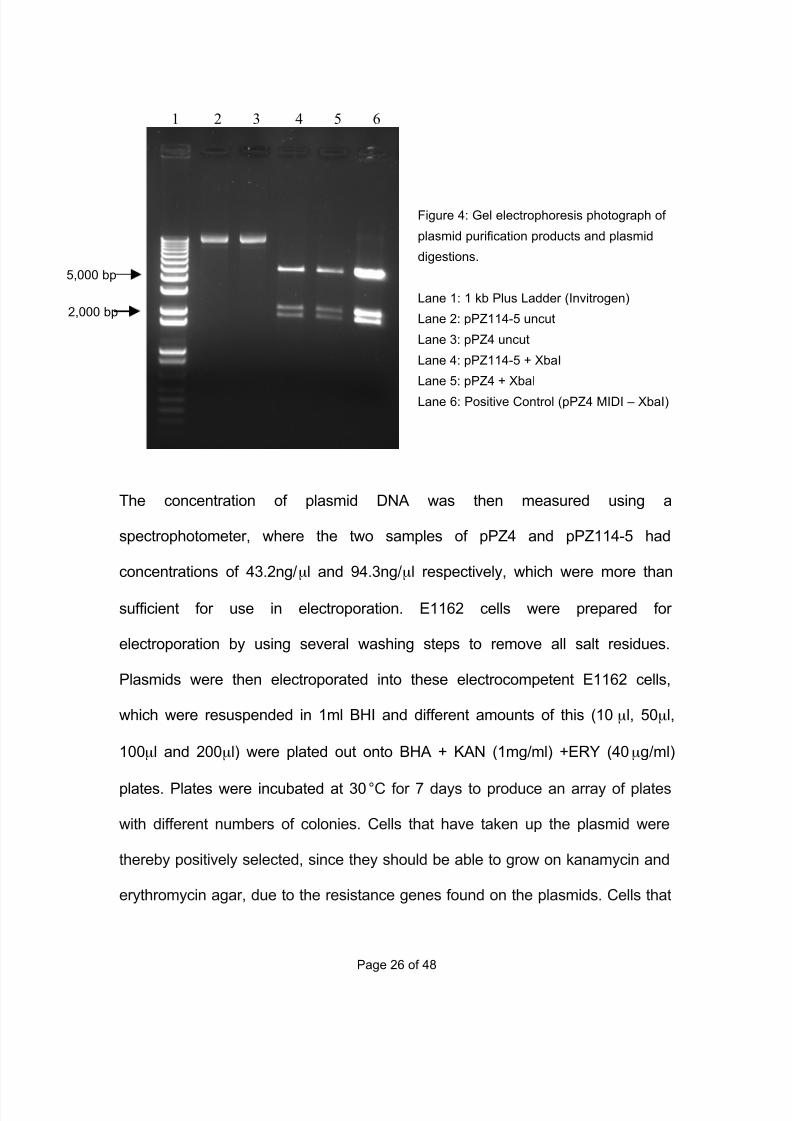
7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 26/48
Page 26 of 48
The concentration of plasmid DNA was then measured using a
spectrophotometer, where the two samples of pPZ4 and pPZ114-5 had
concentrations of 43.2ng/µl and 94.3ng/µl respectively, which were more than
sufficient for use in electroporation. E1162 cells were prepared for
electroporation by using several washing steps to remove all salt residues.
Plasmids were then electroporated into these electrocompetent E1162 cells,
which were resuspended in 1ml BHI and different amounts of this (10µl, 50µl,
100µl and 200µl) were plated out onto BHA + KAN (1mg/ml) +ERY (40µg/ml)
plates. Plates were incubated at 30°C for 7 days to produce an array of plates
with different numbers of colonies. Cells that have taken up the plasmid were
thereby positively selected, since they should be able to grow on kanamycin and
erythromycin agar, due to the resistance genes found on the plasmids. Cells that
5,000 bp
2,000 bp
Figure 4: Gel electrophoresis photograph of
plasmid purification products and plasmid
digestions.
Lane 1: 1 kb Plus Ladder (Invitrogen)
Lane 2: pPZ114-5 uncut
Lane 3: pPZ4 uncut
Lane 4: pPZ114-5 + XbaI
Lane 5: pPZ4 + XbaI
Lane 6: Positive Control (pPZ4 MIDI – XbaI)
1 2 3 4 5 6

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 27/48
Page 27 of 48
have not taken up the plasmid will not be able to grow because they lack these
resistance genes.
After the incubation a small number of colonies containing either pPZ4 or
pPZ114-5 were visible on some of the plates. Five colonies were selected at
random and prepared as overnight cultures in BHI + KAN (1mg/ml) +ERY
(40µg/ml), which were incubated at 30°C overnight. Of the five colonies, two
(named A1 and A2) were supposed to contain the plasmid pPZ4 and 3 (named
B1, B2 and B3) the plasmid pPZ114-5, however only 4 of the overnight cultures
successfully grew. Colony A1, which may have contained the plasmid pPZ4 did
not grow. To confirm if these clones had taken up either pPZ4 (A2) or pPZ114-5
(B1, B2 and B3), a plasmid preparation was carried out followed by a restriction
enzyme digest and gel electrophoresis (Figure 5).
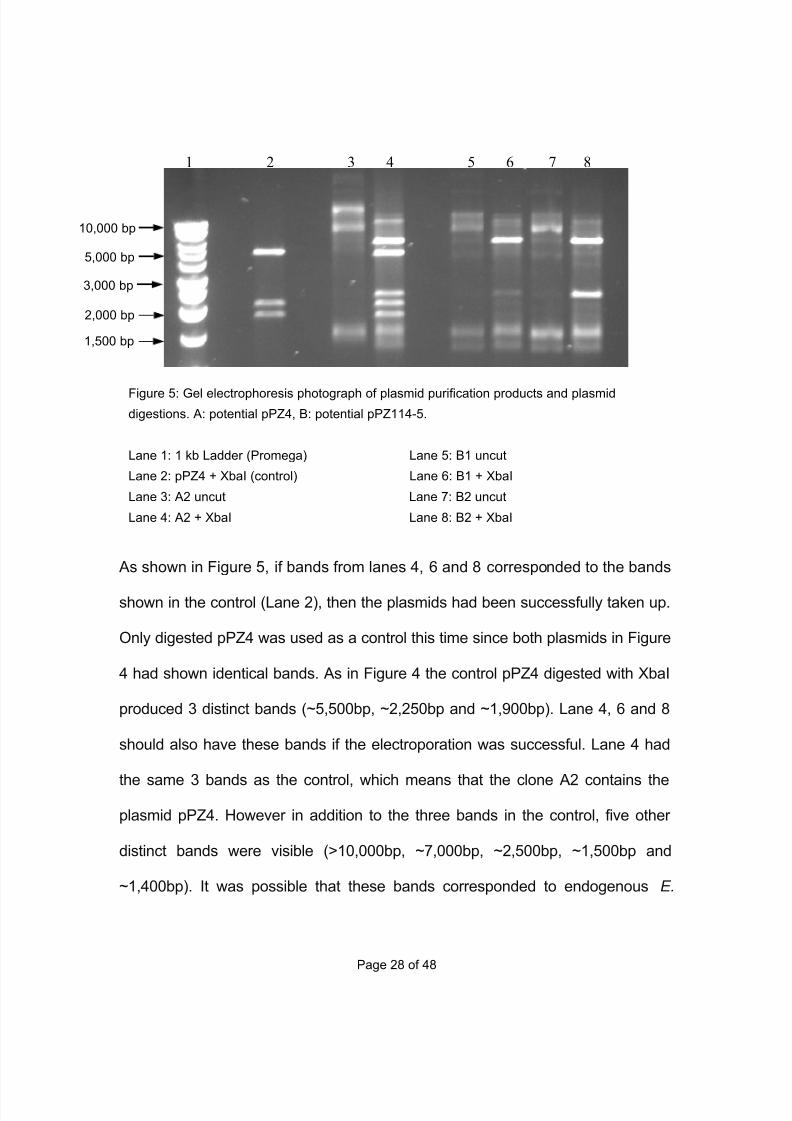
7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 28/48
Page 28 of 48
1 2 3 4 5 6 7 8
2,000 bp
5,000 bp
10,000 bp
As shown in Figure 5, if bands from lanes 4, 6 and 8 corresponded to the bands
shown in the control (Lane 2), then the plasmids had been successfully taken up.
Only digested pPZ4 was used as a control this time since both plasmids in Figure
4 had shown identical bands. As in Figure 4 the control pPZ4 digested with XbaI
produced 3 distinct bands (~5,500bp, ~2,250bp and ~1,900bp). Lane 4, 6 and 8
should also have these bands if the electroporation was successful. Lane 4 had
the same 3 bands as the control, which means that the clone A2 contains the
plasmid pPZ4. However in addition to the three bands in the control, five other
distinct bands were visible (>10,000bp, ~7,000bp, ~2,500bp, ~1,500bp and
~1,400bp). It was possible that these bands corresponded to endogenous E.
Figure 5: Gel electrophoresis photograph of plasmid purification products and plasmid
digestions. A: potential pPZ4, B: potential pPZ114-5.
Lane 1: 1 kb Ladder (Promega) Lane 5: B1 uncut
Lane 2: pPZ4 + XbaI (control) Lane 6: B1 + XbaI
Lane 3: A2 uncut Lane 7: B2 uncut
Lane 4: A2 + XbaI Lane 8: B2 + XbaI
3,000 bp
1,500 bp

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 29/48
Page 29 of 48
faecium plasmids, as the same bands were visible in Lane 6 and 8. Lane 6 and 8
did not show the 3 bands as in the control so both B1 and B2 clones didn’t taken
up the plasmid pPZ114-5. To check for any background plasmids in E1162 a
second plasmid preparation was carried out, on the wild type E1162, which had
not been electroporated, and another clone (B3) potentially containing pPZ114-5.
These preparations were digested with XbaI and run on an agarose gel (Figure
6).
E1162 digested with XbaI results in 4 distinct bands (~10,000bp, ~2,500bp,
~1,400bp and ~1,300bp). These bands most likely corresponded to some of the
bands seen in Lanes 4, 6 and 8 in Figure 5 and Lane 7 in this photograph, which
means that E. faecium does contain at least one background plasmid. When the
plasmid from clone B3 was digested three bands were visible as seen in the
1 2 3 4 5 6 7
Figure 6: Gel electrophoresis photograph of
plasmid purification products and plasmid
digestions. B: potential pPZ114-5
Lane 1: 1kb DNA ladder (Promega)
Lane 2: pPZ4 uncut
Lane 3: pPZ4 + XbaI
Lane 4: E1162 uncut
Lane 5: E1162 + XbaI
Lane 6: B3 uncut
Lane 7: B3 + XbaI
10,000 bp
3,000 bp
1,500 bp

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 30/48
Page 30 of 48
control (Lane 3). This indicated that the clone B3 had retained the plasmid
pPZ114-5. Now that two clones (A2 and B3) had been identified, which contain
pPZ4 and pPZ144-5 respectively, transposon mutagenesis was carried out.
Transposon Mutagenesis and Mutant Library
For the transposon to jump the tpase gene has to be expressed, which is
regulated by the Pxyl/tetO promoter, which can be induced by certain
concentrations of ATc. Several different trials using increased levels of ATc were
carried out to see what effect this tetracycline analogue had on growth of E.
faeciumxvii. The concentrations of ATc used in the three trials were 1.0µg/ml,
1.5µg/ml and 2.0µg/ml, based on previous transposition experiments. However
based on optical density measurements and the time required for the cultures to
reach these measurements it seems that increasing ATc levels does have an
affect on E. faecium growth rate but not to the same extend as in other bacteria.
By doubling the ATc concentration from 1µg/ml to 2µg/ml the time taken for the
culture to reach an OD600 of ~0.2 did increase but only by 35 minutes (145
minutes for 1µg/ml, 156 minutes for 1.5µg/ml and 180 minutes for 2µg/ml). These
observations may be due to E. faecium having a certain level of resistance
against ATc, causing the growth rate to remain relatively stable. When plasmids
with the Pxyl/tetO promoter were used in the bacterium Staphylococcus aureus
the minimal inhibitory concentration of ATc was found to be 2µg/mlxviii, showing
that ATc has a significant inhibitory effect on growth in other bacteria.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 31/48
Page 31 of 48
The 3 transposition experiments were carried out for the two clones A2 and B3
(containing pPZ4 and pPZ114-5 respectively) with increasing levels of ATc to
induce the expression of the tpase gene causing transposon mutagenesis to take
place. It was then necessary to try and remove the transformed plasmids from
the cells, once the transposon had been integrated into the host genome for two
reasons. Firstly, because the erythromycin resistance gene found in the
transposon was to be used as a positive marker for successful insertions. If the
plasmids were still present, then even the cells where transposon mutagenesis
had not occurred would still be able to grow on BHA + ERM (40µg/ml) plates,
since the ermC gene is found on the plasmid. The second reason was to help
determine which transposon delivery vector, pPZ4 or pPZ114-5, was more
successful by comparing the ratio of colonies that only grew on BHA + ERY with
the number of colonies that grew on both media. The transposition cultures were
incubated at 45°C overnight in an attempt to try and cure the cells of the
plasmids. 10-fold serial dilution series of the transposition cultures were then
carried out and 100µl of neat to 10-9 dilutions were spread onto BHA and
BHA+ERY (40µg/ml) plates, which were incubated at 43°C overnight. All the
colonies were counted for each plate and averages were calculated (Table 3).

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 32/48
Page 32 of 48
10-5
10-6
10-7
10-8
BHA 699.67 106.33 5.67 0.67A2
BHA+ERY 652.67 69.33 9 0
BHA 754 81.67 7.67 0B3
BHA+ERY 664 73.67 7.33 0
Table 3: Table shows average number of colonies on 10-fold transposition dilution plates from
10-5
to 10-8
for both BHA and BHA + ERY plates for clones A2 and B3. Three plates were used
for each dilution. (Neat – 10-4
and 10-9
dilution plate results not shown)
For the neat to 10-4 plates there was a lawn of bacteria, 10-5 and 10-6 plates had
a large amount of single colonies and 10-7
to 10-9
plates had either a small
amount of colonies or none at all. There were only a slight number of differences
in the number of colonies for both A2 and B3 clones between the BHA and
BHA+ERY plates, where there was an average of 40 colonies less on the BHA +
ERY plates.
Determining Growth defective E. faecium colonies
To check for plasmid retention even after the attempt to cure the cells, individual
A2 and B3 colonies were patched first onto BHA + KAN (1mg/ml) and then the
same colony was streaked onto BHA + ERY (40µg/ml) plates. Cells still
containing either plasmid were capable of growing on both KAN and ERY plates,
cells where there was no plasmid retention but transposon insertion did not occur
or the integrated transposon is not being expressed weren’t able to grow on both
types of plates. Cells in which the transposon had been successfully integrated
into the host genome and plasmids were not retained could grow on BHA + ERY

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 33/48
Page 33 of 48
plates but not on the BHA KAN plates, since they had lost the kanamycin
resistance gene. 300 colonies were patched onto both kanamycin and
erythromycin BHA plates and then incubated at 43°C overnight. All the plates
showed growth for each individual colony on both BHA + KAN and BHA + ERY
plates. This indicates that the attempt to cure the cells from the plasmids was
unsuccessful, since all the colonies grew on 1mg/ml kanamycin plates and wild
type E1162 cannot survive on concentrations of kanamycin higher than 128µg/ml
(Figure 2). As a result of all the screened cells retaining either plasmid I decided
to assume that transposon mutagenesis had successfully occurred and test for a
mutant strain of E. faecium E1162. Colonies from the transposition were
screened for a loss of sodium chloride resistance due to the transposon
interfering with the resistance genes. Due to there being no significant difference
in number of colonies on the transposition dilution plates and the failure to
determine whether pPZ4 (culture A2) or pPZ114-5 (culture B3) had a higher
transposon mutagenesis rate I decided to carry out the rest of this study with the
clone A2 containing the plasmid pPZ4.
Colonies were screened for the loss of NaCl resistance by using replica plating.
A2 culture from the third transposition was chosen due to the high concentration
of ATc used (2µg/ml), which may have increased the rate of transposon
mutagenesis. A 10-fold dilution series was carried out and 100µl of the 10-6
dilution was spread onto 30 BHA plates and at 37°C overnight. The 10 -6 dilution
was used due to the results in table 3, where there were an average number of

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 34/48
Page 34 of 48
106.33 colonies on BHA plates, which is ideal for replica plating. These 30 plates
represent the mutant library created by the third transposition. The library
consisted of ~1921 colonies distributed onto the 30 BHA plates. All the colonies
from each BHA plate were replicated onto BHA + NaCl (7g per 100ml of BHI
agar) plates via filter paper and both sets of plates were incubated overnight at
37°C to let the colonies grow back. The concentration of sodium chloride used for
screening (7g per 100ml of BHI agar) was chosen due to the results from the
NaCl MIC experiment (Figure 3), where it was determined that wild type E1162
cannot grow on NaCl concentrations 8g or higher. A colony that had grown on
the BHA plate but did not grow on the NaCl plate indicated that the colony had a
genetic defect causing the colony to loose some of its sodium chloride resistance
and was a mutant strain. Out of the 1912 colonies replicated only one colony was
found that grew on BHA but not on the BHA + NaCl plate. To confirm that this
specific clone named A2-28α had lost some of its NaCl resistance, the colony
was streaked onto a BHA + NaCl and then a BHA plate and incubated at 37°C
overnight. A2-28α successfully grew on BHA but not on the BHA + NaCl plate
confirming that this mutant had lost some of its NaCl resistance. To test for pPZ4
plasmid retention, the mutant A2-28α was streaked onto a BHA + ERY (40µg/ml)
and BHA + KAN (1mg/ml) plate and incubated at 37°C overnight. Both plates
showed a significant amount of colony growth, which confirmed that the plasmid
had been retained. Even though the plasmid was still present in the mutant the
next step was to try and determine how many inserts had occurred and the
location of these inserts via different DNA amplification processes.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 35/48
Page 35 of 48
5000 bp
200 bp
1 2 3
Amplification of transposon-flanking sequences
Genomic DNA was extracted from A2-28α (mutant strain) and the concentrations
of the two samples were measured (94.6ng/µl and 153.4ng/µl), confirming the
presence of DNA. The products from genomic DNA preparation were then
digested with the frequent cutting endonuclease TaqIα to produce a large
amount of DNA fragments with staggered ends. The two samples were run on an
agarose gel (Figure 7) to confirm the presence of DNA and determine the size of
the fragments.
When A2-28α was cut with TaqIα (Lane 2) there was a continuous band between
400bp and 200bp, which could have been due to the endonuclease TaqIα cutting
the genome very frequently because of its common recognition site. However
more bands were expected because of the size of A2-28α’s genome. Uncut A2-
28α (Lane 3) produced, an array of bands. Again a continuous band could be
seen between 500bp and 200bp. Two bands (~1,450bp and ~5,000bp) were also
visible, which may correspond to plasmid DNA that had come through the
Figure 7: Gel electrophoresis photograph of
A2-28α genomic preparation products and
digestions.
Lane 1: 1kb PLUS ladder (Invitrogen)
Lane 2: A2-28α + TaqIα
Lane 3: A2-28α uncut
12000 bp
1650 bp

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 36/48
Page 36 of 48
5,000 bp
200 bp
1 2 3 4 5
genomic DNA preparation and clean up, since previous results confirmed there
were endogenous E1162 plasmids. The final band is larger than 12,000bp, which
probably corresponded to the uncut mutant genomic DNA. To test if some of the
bands from Lane 3 corresponded to plasmid DNA, a second preparation was
carried out where A2-28α DNA and pPZ114-5 DNA were digested with the
endonuclease XbaI and run on an agarose gel (Figure 8).
As in Figure 4, when pPZ114-5 was digested with XbaI (Lane3), three bands
were visible. The largest and brightest of the three bands (~5500bp) was also
visible when A2-28α was digested with XbaI (Lane 4), which may point towards
the hypothesis that pPZ4 plasmid DNA had come through the genomic
preparation. However the other two bands (~2200bp and ~1900bp) from Lane 2
are not visible in Lane 4, which may disprove this theory. Unlike in Figure 4 uncut
pPZ114-5 (Lane 2) produced 3 distinct bands, which may have corresponded to
different plasmid configurations. There are several different types if plasmid
Figure 8: Gel electrophoresis photograph of A2-28α genomic preparation products and
pPZ114-5 digested with XbaI.
Lane 1: 1kb PLUS ladder (Invitrogen)
Lane 2: pPZ114-5 uncut
Lane 3: pPZ114-5 + XbaI
Lane 4: A2-28α + XbaI
Lane 5: A2-28α uncut
12,000 bp
2,000 bp

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 37/48
Page 37 of 48
configurations, which are known to affect their migration in gel electrophoresisxix.
Thawing and refreezing of the pPZ114-5 DNA sample could have caused the
plasmids to change their topological structure and the three bands could have
corresponded to the same plasmid in linear, supercoiled and relaxed circular
conformations or the same plasmid with different degrees of supercoiling. One of
the conformations produced a band larger than 12,000bp, which matched the
uncut pPZ114-5 band seen in Figure 4 Lane 2. Uncut A2-28α (Lane 5) produced
two bands at (~8,000bp and ~1,200kb) and a continuous band between 400bp
and 200bp just like in Lane 4 and Lanes 2 and 3 in Figure 7. The band at
~8,000kb may have corresponded to one of the uncut pPZ114-5 conformations,
however the other two bands aren’t visible. This means that the results obtained
from Figure 8 were inconclusive as to whether any pPZ4 DNA had come through
the genomic prep.
To test whether there was a transposon element in any of the fragments, and if
so, attempt to try determine the number and locations of transposons, a DNA
amplification protocol utilising Y linkers was carried out using PCR. First a
ligation step was required where a mixture of Y linkers (Y linker 1 + 2 shown in
Table 2) was ligated to the fragmented A2-28α DNA using the enzyme T4 DNA
ligase. PCR was then carried out using a Y linker primer and the transposon
specific Tn primer (Table 2) to try and amplify the transposon flanking
sequences. Gel electrophoresis was carried out for the DNA amplification
products to confirm if DNA had been amplified. However the gel photograph (not

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 38/48
Page 38 of 48
650 bp
1 2 3 4 5
shown) did not produce any bands, which indicated that the DNA amplification
procedure was unsuccessful, even after a second trial.
Since the Y linker DNA amplification method did not produce any results a
second technique was used. The procedure used a form of nested PCR where in
two rounds of PCR a combination of arbitrary and transposon specific primers
were used to try and amplify the transposon flanking sequences. In the first
round of PCR the product from the mutant genomic digest were mixed with a
combination of one arbitrary primer: STAPHArb1 or STAPH Arb2 and one
transposon specific primer: Marq 255 or Marq 269 (see Figure 3 B for
combinations used). 30 cycles of DNA amplification were then carried out and
the PCR products were examined using gel electrophoresis (Figure 9).
Figure 9 shows the results for the 4 different combinations of primers used. The
PCR product using the combination of primers STAPHArb1 and Marq269
produced the only visible band (~600bp). All the other combinations did not
produce a single band, however this didn’t mean that the DNA amplification had
Figure 9: Gel electrophoresis photograph of
products from first PCR round using the
arbitrary + transposon specific primer method.
Lane 1: 1kb PLUS ladder (Invitrogen)
Lane 2: STAPH Arb1 + Marq 255
Lane 3: STAPH Arb1 + Marq 269
Lane 4: STAPH Arb2 + Marq 255
Lane 5: STAPH Arb2 + Marq 269

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 39/48
Page 39 of 48
650 bp
Figure 10: Gel electrophoresis photograph of
products from second PCR round using the
arbitrary + transposon specific primer method.
Lanes 2 to 5 correspond to the samples from
the first PCR round Figure 9, Lanes 2 to 5.
Lane 1: 1kb PLUS ladder (Invitrogen)
Lane 2: Arb3 + Marq 256
Lane 3: Arb3 + Marq 270
Lane 4: Arb3 + Marq 256
Lane 5: Arb3 + Marq 270
300 bp
1 2 3 4 5
been unsuccessful. IT could have been possible that there simply wasn’t a large
enough quantity of DNA present after the first round to produce a visible band.
So for each sample the second round of PCR was carried out, which was
designed to amplify the products from the first round, potentially containing the
transposon flanking sequence. Again a combination of the arbitrary primer Arb3
and either transposon specific primer Marq 256 or Marq 270 was used and the
products were examined by gel electrophoresis (Figure 10).
Lanes 2, 3 and 4 produced an array of visible bands, however Lane 5 (containing
1st: STAPH Arb2 + Marq 269 2
nd: Arb3 + Marq 270) did not produce any bands,
which could indicate that there either there wasn’t enough DNA, which is unlikely
since there were a total of 70 DNA amplification cycles, or more likely that the
combination of primers used didn’t produce any double stranded PCR products,
possibly due to primer locations for fragment length. All the bands produced in
Lanes 2, 3 and 4 could have potentially been sequences of DNA containing the
transposon flanking sequence, but by just inspecting the gel there was no way to
know without further research.
7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 40/48
Page 40 of 48
Comparative Growth Curves
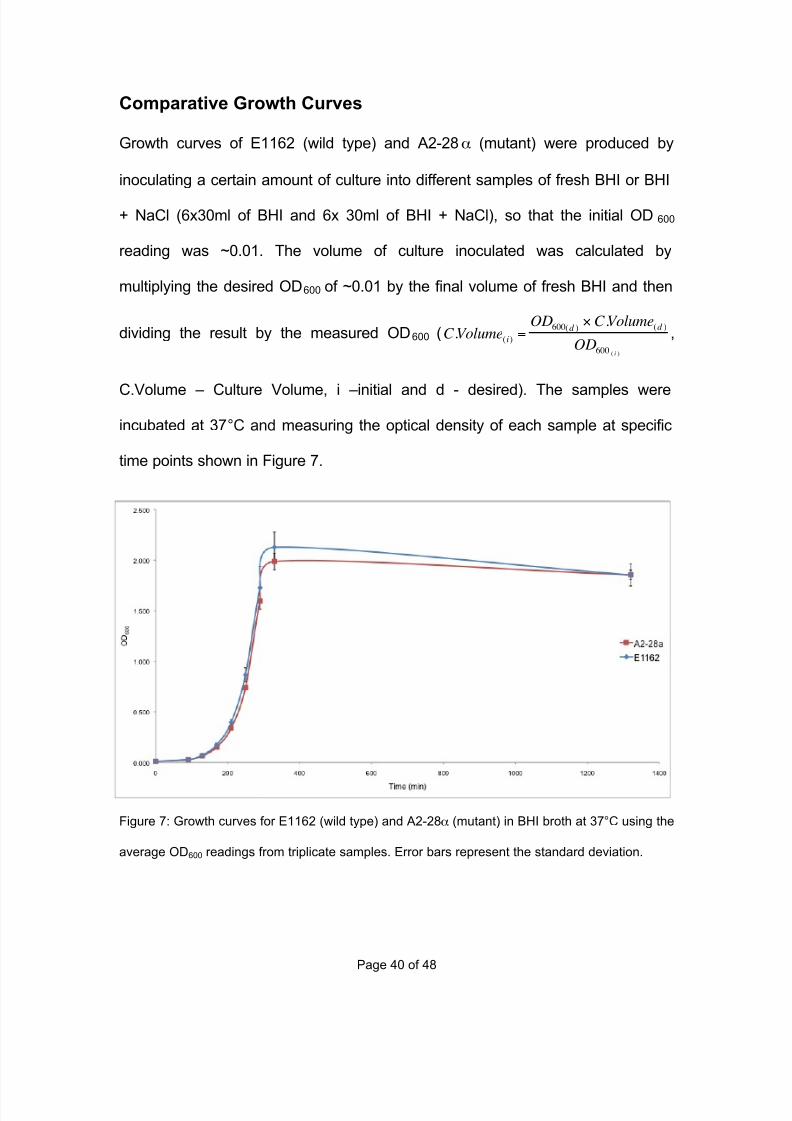
Growth curves of E1162 (wild type) and A2-28α (mutant) were produced by
inoculating a certain amount of culture into different samples of fresh BHI or BHI
+ NaCl (6x30ml of BHI and 6x 30ml of BHI + NaCl), so that the initial OD600
reading was ~0.01. The volume of culture inoculated was calculated by
multiplying the desired OD600 of ~0.01 by the final volume of fresh BHI and then
dividing the result by the measured OD600 (
C .Volume(i)=
OD600(d )×C .Volume(d )
OD600 ( i )
,
C.Volume – Culture Volume, i –initial and d - desired). The samples were
incubated at 37°C and measuring the optical density of each sample at specific
time points shown in Figure 7.
Figure 7: Growth curves for E1162 (wild type) and A2-28α (mutant) in BHI broth at 37°C using the
average OD600 readings from triplicate samples. Error bars represent the standard deviation.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 41/48
Page 41 of 48
The four main phases of a bacterial growth curve are clearly shown in both
samples. The lag phase from 0 to 150 minutes where there is very little increase
in optical density, while the bacteria adjust to their new environmentxx. During the
exponential phase (150 to 350 min) the bacteria start to double at a constant
ratexxi, causing an exponential growth in the number of bacteria. Slowly the
growth rate will decrease and stop altogether, which is known as the stationary
phase (~350-370 min). Eventually the optical density will start to decrease as the
number of bacteria decreases, which is known as the death phase. There is very
little difference in growth rate and phase times between E1162 and A2-28α
,
which indicates that the mutation is not affecting the mutant’s growth in BHI
broth.
When the two cultures were grown in BHA + NaCl (7g per 100ml of BHI broth)
broth the lag phase lasted so long that no further measurements were made. For
example, even after 330 minutes incubation at 37°C the OD600 for E1162 had
only risen from 0.009 to 0.017.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 42/48
Page 42 of 48
DiscussionandFutureDirections
Construction of E. faecium mutant A2-28
In this study transposon mutagenesis was used to create a library of E. faecium
E1162 mutants, by using the Himar1 mariner transposon delivery, which had
been integrated into the two plasmids pPZ4 and pPZ114-5. These two plasmids
were successfully electroporated into the E. faecium wild type (Figures 5 and 6)
and gel electrophoresis revealed that E1162 contains endogenous plasmids.
Research into these plasmids as well as genomic DNA and their potential
functions could reveal what gives this strain some of its characteristics, such as
resistance to external stressors or antimicrobial agents. Once the two clones A2
and B3 containing pPZ4 and pPZ114-5 respectively, had been identified a
transposition procedure was used to produce the library with mutants. However
the size of library created in this study was relatively small, with only around 1921
clones. This meant that the amount of specific growth defective mutants
produced was likely to be very small, since the rate of transposon insertion is
thought to be very low added with the small probability of the transposon actually
being inserted into the gene affecting the screened phenotype. A larger mutant
library could have yielded more growth defective mutants and more reliable
results.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 43/48
Page 43 of 48
To identify the concentration of the antibiotic kanamycin needed for colony
selection and the level of salt needed to inhibit E1162 wild type growth, the
minimal inhibitory concentration for both substances was successfully
determined. As reported in previous studies wild type E. faecium is resistant to
very high concentrations of kanamycin (concentrations equal or smaller than
128µg/ml) and can successfully grow in environments containing high levels of
NaCl. However the method used to determine the MIC in this study differs from
the protocol used in many other studiesxxii. The time and temperature, at which
the bacteria were incubated and left to grow was altered, which may affect the
final results.
Selective plating of transposon mutagenesis colonies showed that the high
temperature incubation step designed to remove the plasmids from the cells by
inhibiting their replication, was unsuccessful. This meant that the rate of
transposon insertion could not be determined or compared between pPZ4 and
pPZ114-5 because selective plating would have been ineffective, due to the
plasmids carrying the selective marker used to select for a transposon insert.
Despite the plasmids having been retained by the cells the library was screened
for colonies that showed a loss in the ability to grow on media containing high
concentrations of salt. The library could not be screened for the loss of
kanamycin resistance since the plasmids, which carry a kanamycin resistance
gene, could not be removed from the cells. Attempts should be made to try and
optimize the transposition procedure for E. faecium E1162, especially the

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 44/48
Page 44 of 48
mechanism by which the bacteria should be cured from the cells, which was
important for so many parts of this study. In the whole library only one growth
mutant was identified to have lost a certain degree of its NaCl resistance, which
may have indicated that very few transposon insertions were made by this
transposon delivery system, since a high insertion rate may have produced a
larger percentage of mutants. However this is only speculation due to the before
mentioned plasmid retention and further work needs to be carried out to
determine the insertion rate for this system in E. faecium.
Identification and amplification of Transposon
A DNA amplification procedure utilising Y linkers was unsuccessful in trying to
locate the position of the transposon in the genome and determine how many
inserts have taken place. Although the main cause could not be determined one
simple reason could have been incomplete digestion of the mutant genome. The
Y linker method does not produce any results if the fragment containing the
transposon is to long. When investigated by gel electrophoresis A2-28α genomic
DNA showed a lack of predicted bands. This may however have been due to the
high cutting frequency of the endonuclease used, which would invalidate the
previous hypothesis. Previous experiments using the Himar1 mariner transposon
delivery have confirmed that the endonuclease TaqIα does not interfere with
amplification because most if not all of the transposon sequences are left uncut.
The protocol could be optimised for E. faecium by carrying out different PCR

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 45/48
Page 45 of 48
trails several times with different annealing and extension temperatures or maybe
a different DNA polymerase to try and produce some PCR products.
A second procedure was used to try and amplify the transposon flanking
sequences. A combination of arbitrary primers and transposon specific primers in
two rounds of PCR produced an array of different bands, which may or may not
have contained the desired transposon flanking sequences. It is possible for
different bands to have contained the transposon flanking sequence since the
arbitrary primers used would have bound to different locations on mutant
genome. If several of these primers were close enough to the transposon specific
primer then several different strands of DNA will be produced but all containing
the transposon flanking sequence. Of course it was also possible that both types
of primers actually had annealed to the plasmid and amplified the transposon
flanking sequences from pPZ4, since previous results confirmed that pPZ4 had
been retained in the cells. Some of the bands may not have even contained the
desired sequence if the primers annealed to sequences, which were not 100%
complementary. Even though this is extremely unlikely there were a total of 70
DNA amplification cycles, which increases the chance of this occurring. Even if it
only happened once, that undesired sequence would then be amplified and
shown on the gel. Most likely there was a combination of all these possible
scenarios to produce all the different types of bands. Due to not knowing, which
band definitely contained the transposon flanking sequence the PCR products
could not be sequenced. This means the exact location of the transposon

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 46/48
Page 46 of 48
causing the growth defect in A2-28α could not be determined and the gene
responsible for at least some of E. faecium’s ability to deal with high salt levels
could not be located without further research. The method used in this study
used unusually low annealing temperatures, which may have had some effects
on the PCR products. Again by carrying out several trials with different annealing
and extension temperatures, more reliable results may have been produced.
This study has shown that the Himar1 mariner transposon delivery can
successfully be used to carry out transposon mutagenesis and create a library of
E. faecium mutants. However the tools and methods used to screen for the
location of these inserts need to optimised for the bacterium in question, so that
further research can be carried out into which genes produced the screened
phenotype.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 47/48
Page 47 of 48
References
iChou YY, Lin TY, Lin JC, Wang NC, Peng MY, Chang FY. (2008) Vancomycin-resistant
enterococcal bacteremia: comparison of clinical features and outcome betweenEnterococcus faecium and Enterococcus faecalis., J Microbiol Immunol Infect. 2008 Apr;41(2):124-9.
iiMurdoch DR, Mirrett S, Harrell LJ, Monahan JS, Reller LB. (2002) Sequential emergence of
antibiotic resistance in enterococcal bloodstream isolates over 25 years. Antimicrob AgentsChemother. 2002 Nov;46(11):3676-8.
iiiMoellering RC. (1992) Emergence of Enterococcus as a significant pathogen. Clin Infect
Dis 1992; 14: 1173–178.
ivMaccallum WG, Hastings TW. (1899) A case of acute endocarditis caused by micrococcus
zymogens (NOV. Spec), with a description of the microorganism. J Exp Med. 1899 Sep
1;4(5-6):521-534.
vWillems RJ, Top J, van Santen M, Robinson DA, Coque TM, Baquero F, Grundmann H, Bonten
MJ. (2005) Global spread of vancomycin-resistant Enterococcus faecium from distinctnosocomial genetic complex. Emerg Infect Dis. 2005 Jun;11(6):821-8.
viIwen PC, Kelly DM, Linder J, et al. (1997) Change in prevalence and anti- biotic resistance
of Enterococcus species isolated from blood cultures over an 8-year period . Antimicrob Agents Chemother 1997;41:494-5.
viiBonten MJ, Willems R, Weinstein RA. Vancomycin-resistant enterococci: why are they
here, and where do they come from? Lancet Infect Dis. 2001 Dec;1(5):314-25.
viiiMurray BE. (2000) Vancomycin-resistant enterococcal infections. N Engl J Med. 2000 Mar
9;342(10):710-21.
ixCao M, Bitar AP, Marquis H. (2007) A mariner-based transposition system for Listeria
monocytogenes. Appl Environ Microbiol. 2007 Apr;73(8):2758-61. Epub 2007 Feb 16.
xBae T, Schneewind O. (2005) Allelic replacement in Staphylococcus aureus with inducible
counter-selection. Plasmid. 2006 Jan;55(1):58-63. Epub 2005 Jul 26.
xiKwon YM, Ricke SC., (2000) Efficient amplification of multiple transposon-flanking
sequences. J Microbiol Methods. 2000 Aug;41(3):195-9.
xiiVijaranakul U, Nadakavukaren MJ, Bayles DO, Wilkinson BJ, Jayaswal RK. (1997)
Characterization of an NaCl-sensitive Staphylococcus aureus mutant and rescue of theNaCl-sensitive phenotype by glycine betaine but not by other compatible solutes. ApplEnviron Microbiol. 1997 May;63(5):1889-97.

7/29/2019 Undergraduate Dissertation: Construction of growth defective mutant strains of Enterococcus faecium
http://slidepdf.com/reader/full/undergraduate-dissertation-construction-of-growth-defective-mutant-strains 48/48
xiiiVijaranakul U, Nadakavukaren MJ, Bayles DO, Wilkinson BJ, Jayaswal RK. (1997)
Characterization of an NaCl-sensitive Staphylococcus aureus mutant and rescue of theNaCl-sensitive phenotype by glycine betaine but not by other compatible solutes. ApplEnviron Microbiol. 1997 May;63(5):1889-97.
xiv
Xiong XP, Wang C, Ye MZ, Yang TC, Peng XX, Li H. (2010) Differentially Expressed Outer Membrane Proteins of Vibrio alginolyticus in Response to Six Types of Antibiotics. Mar Biotechnol (NY). 2010 Mar 9.
xvVillafane R, Bechhofer DH, Narayanan CS, Dubnau D. (1987) Replication control genes of
plasmid pE194. J Bacteriol. 1987 Oct;169(10):4822-9.
xviKwon YM, Ricke SC. (2000) Efficient amplification of multiple transposon-flanking
sequences. J Microbiol Methods. 2000 Aug;41(3):195-9.
xviiBae T, Schneewind O. (2005) Allelic replacement in Staphylococcus aureus with
inducible counter-selection. Plasmid. 2006 Jan;55(1):58-63. Epub 2005 Jul 26.
xviiiBae T, Schneewind O. (2005) Allelic replacement in Staphylococcus aureus with
inducible counter-selection. Plasmid. 2006 Jan;55(1):58-63. Epub 2005 Jul 26.
xixSummers K. (1996) The biology of Plasmids, Blackwell Science Ltd., p.15
xxSingleton P. (2004) Bacteria in Biology, Biotechnology and Medicine, 6
thed., John Wiley
and Sons, p.63-65
xxiSingleton P. (2004) Bacteria in Biology, Biotechnology and Medicine, 6
thed., John Wiley
and Sons, p.63-65
xxiiWiegand I, Hilpert K, Hancock RE. (2008) Agar and broth dilution methods to determine
the minimal inhibitory concentration (MIC) of antimicrobial substances.
Nat Protoc. 2008;3(2):163-75.







