
0
UNIVERSIDADE DE MOGI DAS CRUZES
MAUREEN COSTA FLORIAN SENEDEZE
EFEITOS DO PROCESSAMENTO E ARMAZENAMENTO
NAS CARACTERÍSTICAS QUÍMICAS, SENSORIAIS E
MICROBIOLÓGICAS DE COGUMELOS COMESTÍVEIS
Pleurotus ostreatus
Profª orientadora: Dra. Erna Elisabeth Bach
Mogi das Cruzes, SP 2009
Dissertação apresentada ao curso de Pós- graduação da Universidade de Mogi das Cruzes como parte dos requisitos para a obtenção do título de Mestre em Biotecnologia. Área de concentração: Biológica

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

1

2

3
AGRADECIMENTOS
Agora que estou no momento final deste trabalho, quero primeiramente agradecer a
Deus, pois mesmo com os obstáculos e dificuldades que existiram ao longo da realização
desta pesquisa, Ele me fez superá-los e me fortaleceu, possibilitando-me entender a
necessidade de tudo que estava acontecendo.
Hoje sei o real significado de ser uma pesquisadora e compreendo como é
fundamental ter uma orientadora dedicada e responsável, pois tive a honra de ter sido
orientada pela Profa. Dra. Erna Elisabeth Bach, que acreditou no meu projeto, apoiando-me e
ensinando tudo que se fazia necessário, com compreensão e amizade, estando presente e me
auxiliando na superação dos obstáculos encontrados. Obrigada pela sua amizade, pelos
ensinamentos e pela paciência.
Um grande trabalho nunca é realizado por uma só pessoa. Por isso, o grupo de
pesquisa da UNINOVE foi de primordial importância para o andamento do estudo. Em
especial, quero agradecer à Graziela, à Bruna, à Mariana e ao Fernando, alunos da graduação,
pela dedicação e trabalho em equipe.
Para a obtenção dos resultados desta pesquisa foram essenciais as doações de
cogumelos realizadas pelo Sr. Edison de Souza, proprietário da empresa Brasmicel. Agradeço
também à empresa Magnum, que ajudou para que os cogumelos pudessem ser embalados a
vácuo. Meu muito obrigada pelo apoio e atenção.
Agradeço aos mestres do curso de pós-graduação da Universidade de Mogi das
Cruzes pela contribuição ao meu aprendizado, crescimento e aperfeiçoamento profissional.
Às minhas verdadeiras e especiais amigas Lisméia, Fernanda e Ana Paula que
sempre farão parte da história de minha vida. Você Lisméia, por ter sido a maior
incentivadora para que eu me tornasse o que hoje tenho imenso prazer e grande orgulho de
ser: professora; pelas conversas e desabafos semanais, pelo incentivo e encorajamento. Você
Fernanda, pelo carinho, atenção, palavras sempre doces e certas para o momento, além das
necessárias e benéficas terapias à distância. E você Ana, pela paciência, por acreditar no meu
potencial e pela força em todos os momentos.
Não posso deixar de agradecer também a todos os demais amigos que souberam
entender a minha ausência, muitas vezes em ocasiões especiais e que toleraram os muitos
momentos de impaciência.

4
Durante a realização deste trabalho, principalmente na elaboração da dissertação,
compreendi o por que, na seção dos agradecimentos, sempre há uma homenagem especial aos
maridos. Não seria diferente para mim, que tenho muita gratidão ao meu amado e eterno
companheiro Rodrigo, por participar indiretamente da obtenção deste título, por ser meu
companheiro nas ausências nos encontros com os amigos, pela paciência, amizade e por
sempre me motivar, mesmo nas situações mais difíceis. Te amo!
Os laços sanguíneos fazem com que as pessoas que nascem em uma mesma família
sejam irmãos. Respeito, união, amizade e lealdade entre eles nem sempre é um privilégio de
todos. Eu, tendo esse privilégio, agradeço às minhas irmãs Monik, Marianne, Michelle e
Millena por me ensinarem a cada dia a ser uma pessoa mais humana, amiga e
verdadeiramente feliz. Amo muito vocês.
Como parte da minha família, agradeço aos meus cunhados Eduardo, Arthur e
Douglas e a Vanessa, minha mais nova cunhada, por torcerem pelo meu crescimento
profissional e acadêmico.
Não existe uma palavra para classificar uma pessoa que é ao mesmo tempo minha
segunda mãe e minha leal amiga. Obrigada Denice (minha sogra) por tudo o que você faz por
mim e pela importância que você tem em minha vida.
Escolhi para encerrar estes agradecimentos meus amados pais, Celso e Maria
Helena. No fim, não por serem menos importantes, mas por que eles representam o início de
tudo, pois me deram a oportunidade da vida. E se hoje estou aqui finalizando este trabalho,
devo muito a eles. Agradeço por me educarem, pelo carinho, orações e pelas palavras de
motivação. Por estarem sempre ao meu lado, por serem exemplos que tenho como referência
para a minha vida e por se tornaram grandes amigos, além de maravilhosos pais. Tenho um
imenso orgulho de tê-los como pais.

5
“Educar é semear com sabedoria e colher com paciência; é ser
um artesão da personalidade, um poeta da inteligência e um
semeador de idéias.
Bons professores educam para uma profissão e são mestres
temporários; professores fascinantes educam para a vida e são
mestres inesquecíveis.
Todos erram: a maioria usa os erros para se destruir; a minoria,
para se construir. Estes são os sábios”.
(Augusto Cury)

6
RESUMO
O cogumelo Pleurotus ostreatus é um produto perecível, com tempo de vida útil limitado, podendo sofrer contaminação por diversos micro-organismos se não for bem armazenado. Sua vida útil na prateleira pode variar entre 2 a 7 dias, em função da umidade relativa e temperatura de armazenamento. Assim, o objetivo do presente trabalho consistiu em avaliar componentes químicos e nutricionais do cogumelo P. ostreatus, tais como a atividade das enzimas PPO e POD, presença e quantificação de fenóis, vitamina C, proteína e carboidratos α e β-glucanas além das características microbiológicas e sensoriais quando submetidos a diferentes processos e tempos de acondicionamento. Para isto, cogumelos P. ostreatus, de tamanho grande e pequeno, foram colhidos junto à empresa Brasmicel (localizada em Suzano-SP) e transportados em geladeira de isopor até o laboratório e logo processados. Primeiramente, foi realizada a padronização do tempo para método de lavagem e fervura para posterior comparação dos resultados entre cogumelos de tamanho grande e pequeno. Os cogumelos in natura de ambos os tamanhos foram pesados e separados em grupos de amostras de 50 g. O grupo controle foi somente lavado em água corrente. As demais amostras foram submetidas a um pré-tratamento de branqueamento com ácido cítrico (0,25%) por 4 minutos, acondicionadas em embalagens plásticas, fechadas a vácuo ou por selagem simples e colocadas sob refrigeração durante 3, 7, 14 e 28 dias. Decorridos os períodos, as embalagens foram abertas, sendo avaliados: aspectos sensoriais (cor, odor, sabor e textura), comunidade microbiana das amostras além de extração e quantificação química e nutricional. Os resultados microbiológicos, para todos os tratamentos, demonstraram que nos líquidos exsudados dos cogumelos e avaliados em meios de cultura, somente foi detectada presença de leveduras. Quanto aos aspectos sensoriais, o de tamanho grande armazenado com vácuo, apresentou a textura mole, rompendo-se facilmente na mastigação. O cogumelo de tamanho pequeno armazenado a vácuo por até 14 dias apresentou todas as características sensoriais preservadas se comparado com a amostra armazenada sem vácuo pelo mesmo período de tempo. Quanto à análise bioquímica, foram detectados nos cogumelos presença de proteínas, fenóis, carboidratos alfa e beta e vitamina C. Portanto, cogumelos P. ostreatus podem ser submetidos ao branqueamento e embalados a vácuo aumentando assim a vida de prateleira e mantendo as características sensoriais, bioquímicas, microbiológicas e nutricionais por até 14 dias.
Palavra chave: Pleurotus ostreatus; aspectos químicos; características sensoriais; comunidade microbiana.

7
ABSTRACT
Pleurotus ostreatus is a perishable product, with limited time of life, being able to suffer contamination for diverse microorganism if well they will not be stored. His life on the shelf can vary from 2 to 7 days depending on humidity and temperature of storage. Thus, the objective of the present work was evaluate chemical and nutritional components of P. ostreatus, such as activity of enzymes PPO and POD, the presence and quantification of phenols, vitamin C, protein and carbohydrates α and β-glucans in addition to microbiological and sensory characteristics when submitted to different processes and time of packaging. For this, P. ostreatus, of large and small size, were collected from the company Brasmicel (located in Suzano-SP) and transported in the refrigerator (Styrofoam) to the laboratory and then processed. First, was standardized the time for washing and boiling method for later comparison of the results between large and small fungi. The fungi in nature of both sizes were weighed and separated into groups of samples of 50 g. Control group was only washed in tap water. The remaining samples were submitted to a pre-treatment of bleaching with citric acid (0.25%) for 4 minutes, packed in plastic, sealed under vacuum or by simply sealing and placed under refrigeration for 3, 7, 14 and 28 days. After the periods of time, the packages were opened, and evaluated the following: sensory aspects (color, odor, taste and texture), the microbial community of the sample and the extraction, the chemical and nutrition quantification. Microbiological results, for all the treatments, demonstrated that in liquid exudates of mushrooms and evaluated in culture way, only yeast was found. As to the sensory, the large size stored with vacuum, showed a soft texture, breaking up easily in mastication. The small size of fungi stored under vacuum for 14 days had preserved all the sensory characteristics compared to the stored sample without vacuum for the same period of time. As for the biochemical analysis, were detected in the mushrooms presence of protein, phenols, alpha and beta carbohydrates and vitamin C. Therefore, fungi of P. ostreatus can be laundered and packed under vacuum thereby increasing shelf life and maintaining the sensory characteristics, biochemical, microbiological and nutritional components for until 14 days.
Key words: Pleurotus ostreatus; chemical aspects; sensory characteristics; microbial characteristics.

8
LISTA DE ILUSTRAÇÕES
Figura 1: Desenho esquemático das principais estruturas do corpo de frutificação dos cogumelos ............................................................................................................... 18
Figura 2: Aspectos gerais do cogumelo Pleurotus ostreatus ................................................. 27
Figura 3: Etapas do cultivo de cogumelos Pleurotus spp ...................................................... 28
Figura 4: Diferentes estágios de crescimento do Pleurotus ostreatus ................................... 30
Figura 5: Produção mundial de cogumelos comestíveis, em porcentagem, por décadas....... 36
Figura 6: Etapas empregadas na padronização do método de lavagem do cogumelo Pleurotus ostreatus............................................................................................. 42
Figura 7: Etapas empregadas na padronização do tempo de fervura de cogumelo Pleurotus ostreatus....................................................................................... 43
Figura 8: Etapas empregadas no processamento do cogumelo Pleurotus ostreatus........ 45
Figura 9: Cogumelo Pleurotus ostreatus de tamanho pequeno preferencialmente comercializado. ....................................................................................................... 51
Figura 10 A: Diferentes estágios de desenvolvimento do Pleurotus ostreatus. Tamanho pequeno e médio. B: Pleurotus ostreatus de tamanho grande. Aspectos gerais no momento da colheita .......................................................................................... 52
Figura 11: Teores de proteína e fenol liberados para os líquidos de lavagem. Comparação entre as amostras controle e pré-tratadas de Pleurotus ostreatus de tamanho pequeno, médio e grande............................................................ 53
Figura 12: Enzimas polifenoloxidase e peroxidase liberadas para os líquidos de lavagem. Comparação entre as amostras controle e pré-tratadas de Pleurotus ostreatus de tamanho pequeno, médio e grande.......................................................................... 55
Figura 13: Teores de proteína e fenol em Pleurotus ostreatus. Comparação entre as amostras controle e pré-tratadas de tamanho pequeno, médio e grande. ................ 56
Figura 14: Atividade das enzimas polifenoloxidase (A) e peroxidase (B) em Pleurotus ostreatus. Comparação entre as amostras controle e pré-tratadas de tamanho pequeno, médio e grande. ....................................................................................... 58
Figura 15: Teores de proteína (mg SAB/g cogumelo fresco) e fenol (mg ác. clorogênico/g cogumelo fresco) e atividade enzimática da polifenoloxidase (Unidade PPO/minuto/g cogumelo fresco) em Pleurotus ostreatus. Comparação entre as amostras controle e as fervidas por 2 e 4 minutos.. .............. 59
Figura 16: Relação peso fresco/peso seco (g) e perda de água (%) do cogumelo Pleurotus. ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. ..................................................................................... 62
Figura 17: Perda de água (%) dos Pleurotus ostreatus após os tempos de armazenamento com relação ao peso pós-tratamento. Comparação entre os cogumelos de tamanhos pequeno e grande embalados com e sem vácuo. .................................... 64

9
Figura 18: Atividade da enzima polifenoloxidase (unidade de polifenoloxidase/g cogumelo fresco/minuto) em P.leurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento ................................. 66
Figura 19: Perda (%) da atividade enzimática da polifenoloxidase em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. .................................................................................................. 66
Figura 20: Atividade enzimática da peroxidase (unidade de peroxidase/minuto/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. .................................................. 69
Figura 21: Valor de vitamina C (mg vit. C/100g cog. fresco) em P. ostreatus,tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. .............. 70
Figura 22: Mudanças (%) no teor de vitamina C em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. .............. 72
Figura 23: Teor de proteína (mg SAB/100g de cog. fresco) em P. ostreatus, tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento...................................................................................... 73
Figura 24: Teor de fenóis (mg ác. clorogênico/100g de cog. fresco) em P. ostreatus, tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. ...................................................................................................... 75
Figura 25: Teor de glicose (mg de glicose ligação alfa/100g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. ..................................................................................... 77
Figura 26: Teor de β-glucana (mg de beta glucana solúvel/100g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento........................................................................... 78
Figura 27: Valor de pH do cogumelo Pleurotus ostreatus de tamanhos pequeno e grande e dos líquidos exsudados do alimento. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento......................................................................................... 80
Figura 28: Contagem de colônias de leveduras (UFC/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento................................................................................................... 82
Figura 29: Fotos do Pleurotus ostreatus tamanho grande embalados com e sem vácuo e armazenados por diferentes tempos ........................................................................ 86
Figura 30: Fotos do Pleurotus ostreatus tamanho pequeno embalados com e sem vácuo e armazenados por diferentes tempos ........................................................................ 87

10
LISTA DE TABELAS
Tabela 1: Características sensoriais do Pleurotus ostreatus de tamanhos pequeno e
grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. ................................ 84

11
LISTA DE QUADROS
Quadro 1: Ficha utilizada para a avaliação sensorial de cogumelos Pleurotus ostreatus...... 48 Quadro 2: Descrição dos meios de cultura e suas aplicações ................................................ 50

12
SUMÁRIO
1 INTRODUÇÃO ................................................................................................................... 15
1.1 OS FUNGOS .................................................................................................................. 15
1.1.1 Conceitos gerais ....................................................................................................... 15
1.1.2 Fisiologia e taxonomia ............................................................................................. 16
1.2 FUNGOS MACROSCÓPICOS ...................................................................................... 18
1.2.1 Cogumelos comestíveis ............................................................................................ 19
1.2.1.1 Propriedades físico-químicas e enzimáticas ......................................................... 20
1.2.1.2 Características sensoriais ..................................................................................... 22
1.2.1.3 Aspectos microbiológicos ...................................................................................... 24
1.3 CULTIVO DE Pleurotus ................................................................................................ 26
1.4 VANTAGENS NA ALIMENTAÇÃO HUMANA, VALOR NUTRICIONAL E MEDICINAL .................................................................................................................. 30
1.5 PRODUÇÃO, COMERCIALIZAÇÃO E CONSUMO DE COGUMELOS ................. 35
2 OBJETIVOS ........................................................................................................................ 40
3 MATERIAIS E MÉTODOS ............................................................................................... 41
3.1 COLETA DOS COGUMELOS Pleurotus ostreatus ...................................................... 41
3.2 PADRONIZAÇÃO DO MÉTODO DE TRATAMENTO (PRIMEIRA ETAPA) ........ 41
3.3 PADRONIZAÇÃO DO TEMPO DE FERVURA – COGUMELO DE TAMANHO GRANDE (SEGUNDA ETAPA) ................................................................................... 43
3.4 PROCESSAMENTO DO COGUMELO Pleurotus ostreatus (TERCEIRA ETAPA) .. 44
3.5 TEOR DE UMIDADE E PERDA DE ÁGUA DO COGUMELO Pleurotus ostreatus . 45
3.6 ANÁLISES BIOQUÍMICAS.......................................................................................... 46
3.6.1 Proteínas e fenóis ..................................................................................................... 46
3.6.2 Atividade enzimática da polifenoloxidase (PPO) .................................................... 46
3.6.3 Atividade enzimática da peroxidase (POD) ............................................................. 46
3.6.4 Quantificação da Vitamina C ................................................................................... 47
3.6.5 Beta e Alfa-glucana .................................................................................................. 47
3.7 AVALIAÇÃO DAS CARACTERÍSTICAS SENSORIAIS .......................................... 47
3.8 pH DO COGUMELO E DOS LÍQUIDOS EXSUDADOS DO ALIMENTO ............... 48

13
3.9 ANÁLISES MICROBIOLÓGICAS ............................................................................... 48
3.10 ANÁLISE ESTATÍSTICA ........................................................................................... 50
4 RESULTADOS E DISCUSSÃO ........................................................................................ 51
4.1 COLETA DOS COGUMELOS Pleurotus ostreatus ...................................................... 52
4.2 PADRONIZAÇÃO DO MÉTODO DE TRATAMENTO (PRIMEIRA ETAPA) ........ 53
4.3 PADRONIZAÇÃO DO TEMPO DE FERVURA – COGUMELO DE TAMANHO GRANDE (SEGUNDA ETAPA) ................................................................................... 58
4.4 PROCESSAMENTO DO COGUMELO Pleurotus ostreatus (TERCEIRA ETAPA) .. 60
4.4.1 Teor de umidade e perda de água do cogumelo Pleurotus ostreatus .......................... 61
4.4.2 Análises bioquímicas ............................................................................................... 64
4.4.2.1 Polifenoloxidase (PPO) ........................................................................................ 64
4.4.2.2 Peroxidase (POD) ................................................................................................. 67
4.4.2.3 Vitamina C ............................................................................................................ 70
4.4.2.4 Proteína e Fenol .................................................................................................... 73
4.4.2.5 Alfa e Beta glucana liberada ................................................................................. 76
4.4.3 pH do Pleurotus ostreatus e dos líquidos exsudados do alimento ........................... 79
4.4.4 Análises microbiológicas ......................................................................................... 80
4.4.5 Análise sensorial ...................................................................................................... 83
5 CONCLUSÕES E SUGESTÕES ....................................................................................... 89
6 REFERÊNCIAS .................................................................................................................. 90
ANEXO A- Teores de proteína (A) e fenol (B) liberados para os líquidos de lavagem. Comparação entre as amostras controle e pré-tratadas de Pleurotus ostreatus de tamanho pequeno, médio e grande............................................................... 107
ANEXO B- Relação peso fresco/peso seco (g) e perda de água (%) do cogumelo
Pleurotus. ostreatus de tamanhos grande e pequeno. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento......................................................................... 108
ANEXO C- Perda de água (%) do Pleurotus ostreatus após os tempos de armazenamento
com relação ao peso pós-tratamento. Comparação entre os cogumelos de tamanhos pequeno e grande embalados com e sem vácuo............................... 109
ANEXO D- Atividade da enzima polifenoloxidase (unidade de polifenoloxidase/
minuto/g cogumelo fresco) e erda (%) da atividade enzimática em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.......................................................................... 110

14
ANEXO E- Atividade enzimática da peroxidase (unidade de peroxidase/minuto/g
cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento..................................... 111
ANEXO F- Valor (mg vit. C/100g cogumelo fresco) e mudanças (%) no teor de
vitamina C em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento..................................... 112
ANEXO G- Teor de proteína (mg SAB/100g de cogumelo fresco) em Pleurotus
ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento................................................................................ 113
ANEXO H- Teor de fenóis (mg ác. clorogênico/100g de cogumelo fresco) em Pleurotus
ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento................................................................................ 114
ANEXO I- Contagem de colônias de leveduras (UFC/g cogumelo fresco) em Pleurotus
ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento................................................................................ 115

15
1 INTRODUÇÃO
1.1 OS FUNGOS
1.1.1 Conceitos gerais Os fungos pertencem ao reino Fungi e apresentam características particulares, as quais
os diferem das plantas, tais como: não sintetizam clorofila, sendo assim não realizam
fotossíntese; a parede celular não é formada por celulose, como nas plantas e sim por
substâncias quitinosas e armazenam glicose na forma de glicogênio e não de amido como as
espécies vegetais (GOMPERTZ et al., 2008).
São seres eucariotos, mononucleados como as leveduras, ou multinucleados, como os
fungos filamentosos (bolores) e na forma de cogumelos, também chamados de fungos
macroscópicos (BURTON; ENGELKIRK, 2005). O agrupamento de filamentos denomina-se
hifas, que formam um conjunto entrelaçado chamado micélio (BONONI et al., 1999).
O micélio pode se desenvolver no interior do substrato (micélio vegetativo), tendo a
função de sustentação e de absorção de nutrientes. Pode também se projetar para a superfície
e crescer acima do meio de cultura, sendo chamado de micélio aéreo. Esse, por sua vez, pode
se diferenciar e formar o micélio reprodutivo, onde são formados os esporos (GOMPERTZ et
al., 2008).
Acredita-se que existam, atualmente, cerca de 1,5 milhões de espécies diferentes de
fungos espalhados pela natureza, podendo ser encontrados em diversos lugares como no solo,
na água, nos vegetais, no homem e nos animais (BONONI et al., 1999; MUELLER;
SCHMIT, 2007). No seu habitat natural, esses micro-organismos, em condições ambientais e
de nutrição adequadas, multiplicam-se assexuada (por clivagem no interior dos esporângios
ou através dos conídios) ou sexuadamente (pela fusão de dois núcleos), de acordo com a
espécie e suas necessidades (BURTON; ENGELKIRK, 2005; GOMPERTZ et al., 2008).

16
1.1.2 Fisiologia e taxonomia Por não possuírem mecanismos químicos fotossintéticos ou autotróficos para a
produção de energia, a nutrição dos fungos depende exclusivamente de outros tecidos, sejam
estes vegetais, animais ou detritos orgânicos (SILVEIRA, 1995). Segundo Gompertz et al.
(2008) a nutrição dos fungos ocorre através do processo de absorção, onde enzimas
especializadas hidrolisam macromoléculas, as quais são transportadas, por osmose, por todo o
tecido vegetativo.
Por isso, os fungos são obrigados a viver em estado de saprofitismo, parasitismo ou
simbiose. O que difere estes três diferentes estilos de vida é a fonte de alimento, isto é, um
fungo saprofítico alimenta-se de matéria orgânica morta, já em decomposição; no parasitismo,
colonizam hospedeiros vivos, animais ou vegetais, com prejuízo para o hospedeiro (fungos
patogênicos) e os simbiontes, associam-se com outros organismos, vivendo em harmonia e
prestando ajuda mútua (KIMATI et al., 2005).
Para a germinação, crescimento e desenvolvimento dos fungos a presença ou não do
oxigênio, da água e da luminosidade assim como as condições do ambiente no que diz
respeito à temperatura e pH são fatores determinantes. Para a maioria dos fungos a presença
de oxigênio é primordial, contudo alguns podem metabolizar substâncias, de maneira lenta,
em meios com baixa concentração de oxigênio (SILVEIRA, 1995).
A temperatura ótima para o crescimento abrange uma larga escala, podendo então ser
as espécies classificadas como psicrófilas, mesófilas ou termófilas, isto é, organismos que
crescem melhor à temperatura de 15 a 20ºC, de 25 a 40ºC e acima de 45ºC, respectivamente
(PELCZAR JUNIOR; CHAN; KRIEG, 1997; GALVAGNO; FORCHIASSIN, 2004).
Os fungos também necessitam de água para o seu desenvolvimento, principalmente os
bolores. A quantidade de água disponível para que o micro-organismo se desenvolva e realize
suas atividades metabólicas, conhecida como atividade de água (Aa), deve ser de no mínimo
80%; abaixo de 60% de Aa os fungos não se reproduzem (GOMPERTZ et al., 2008).
De acordo com Silveira (1995) um pH em torno de 5,6 é considerado ótimo para o
desenvolvimento dos fungos. Os filamentosos podem crescer nos extremos da concentração
de íons hidrogênio, variando de 1,5 a 11; já as leveduras não toleram pH alcalino.
Quanto à luminosidade, no desenvolvimento vegetativo, os fungos preferem pouca
luz, diferentemente da fase reprodutiva, onde a presença de luz é importante. Porém, a fonte
de luz deve ser adequada e controlada uma vez que a luz solar direta, devido às radiações
ultravioletas, é considerada fungicida (GOMPERTZ et al., 2008).

17
A filogenia dos fungos, atualmente, está sendo auxiliada por técnicas de
sequenciamento moleculares modernas, principalmente na prova de PCR (Polymerase Chain
Reaction) e seleção de oligonucleotídeos com sondas específicas (GOMPERTZ et al., 2008).
Assim, os micro-organismos eucariontes são classificados em três reinos distintos:
Chromista, Protozoa, e Fungi. Este último reino está dividido em quatro filos: Ascomycota,
Zygomycota, Glomeromycota e Basidiomycota e em um grupo, dos Deuteromycetes
(GOMPERTZ et al., 2008; REDECKER, RAAB, 2006).
O filo Zygomycota é subdividido em 2 Classes, nas quais existem 173 gêneros e 1.056
espécies. Diferem dos demais fungos pelo fato dos esporos não apresentam flagelo e centríolo
e por produzirem um esporo de resistência, denominado zigósporo. Podem ser utilizados na
fabricação de produtos industriais como enzimas (amilase), pigmentos (β -caroteno) e ácidos
orgânicos assim como empregados no processo de fermentação de alguns alimentos
popularmente consumidos na Ásia (sufu e tempeh) (HAWKSWORTH et al., 1995 apud
LOGUERCIO-LEITE, 2004; HERRERA; ULLOA, 1990 apud LOGUERCIO-LEITE, 2004).
Segundo o “Dictionary of the fungi”, o maior número de espécies encontrada no reino
fungi constitui o filo Ascomycota, totalizando 32.200, distribuídas nas diferentes Ordens e
Classes (HAWKSWORTH et al., 1995 apud LOGUERCIO-LEITE, 2004). Apresentam
inúmeras vantagens nas atividades humanas tais como na destruição de produtos celulósicos,
como patógenos de plantações, biocontroladores, etc. (TRUFEM, 1998).
A divisão Basidiomycota possui cerca de 22.244 espécies, distribuídas em 3 Classes,
41 Ordens e 165 Famílias (HAWKSWORTH et al., 1995 apud LOGUERCIO-LEITE, 2004).
A estrutura característica distintiva desse grupo é o basídio, uma estrutura especializada na
produção de esporos exógenos, chamados basidiósporos. A maioria das espécies forma uma
estrutura macroscópica, chamada basidioma ou corpo de frutificação, constituída por píleo,
estipe e lamelas (Figura 1) (GUGLIOTTA; CAPELARI, 1998; NEVES, 2007).
Gugliotta e Capelari (1998) relatam que algumas espécies são úteis como produtoras
de substâncias antibióticas, outras como biorremediadoras de solos ou clareadoras de
efluentes industriais. É neste filo que se encontram a maioria das espécies consideradas
comestíveis, dentre elas o cogumelo Pleurotus ostreatus, amplamente cultivado e
comercializado no mundo inteiro.

18
Figura1: Desenho esquemático das principais estruturas do corpo de frutificação dos cogumelos. Fonte: Smiderle, 2008
1.2 FUNGOS MACROSCÓPICOS
Os fungos macroscópicos são aqueles que produzem corpos de frutificação visíveis a
olho nu. Todos os tipos de cogumelos são classificados como fungos do tipo macroscópico, os
quais são seres vivos primitivos que não obtém energia por meio de fotossíntese e, portanto,
extraem seus nutrientes do húmus (tecido parcialmente decomposto de formas vegetais mais
complexas) (EIRA, 2004).
Os fungos são utilizados na alimentação do homem desde os tempos mais remotos da
humanidade. Indiretamente, como agentes fermentativos na produção de bebidas e alimentos,
como, por exemplo, os queijos, e de maneira direta fazendo parte da dieta alimentar, através
dos cogumelos comestíveis (URBEN, 2002).
A relação do homem com os cogumelos é muito antiga o que é demonstrada através
dos registros arqueológicos que datam de 450 a.C. o início de seu uso. Diferentes eram as
suas aplicações e crenças entre as civilizações. A mitologia grega refere-se a eles como

19
“refinados” e “afrodisíacos” e os guerreiros achavam que lhes dariam forças e coragem
(HERRERA, 2001).
Para os Egípcios eram considerados um presente do Deus Osíris assim como os
romanos que os chamavam de “alimento dos deuses” e por isso deviam ser servidos apenas
em ocasiões especiais. Os chineses os consideravam e ainda acreditam ser um “elixir da vida”
e os índios mexicanos os utilizavam com propósitos terapêuticos e como alucinógenos
(MANZI et al.,1999).
Podem também ser venenosos e tóxicos ao homem e animais, causando desde simples
perturbações estomacais e intestinais até lesões irreversíveis nos rins e fígado ou mesmo a
morte (BONONI et al.,1999).
A toxicidade dos fungos varia de espécie para espécie. Entre as mais perigosas estão a
Amanita verna, que na Europa é conhecida como “Anjo Branco” ou “Anjo da morte”, a A.
phalloides, a A. virosa e a A. bisporigena. Aproximadamente 22 toxinas foram encontradas
nesse gênero, sendo que as mais perigosas afetam a síntese protéica por inibição da RNA
polimerase. Por isso, a inadequada identificação dos cogumelos pode levar a ingestão de
espécies tóxicas (TAVEIRA; NOVAES, 2007; BONONI et al., 1999).
1.2.1 Cogumelos comestíveis Segundo a literatura, entre as 45 mil espécies descritas de cogumelos, existem cerca de
2000 espécies comestíveis, sendo apenas 25 delas utilizadas na alimentação humana e um
número menor comercialmente cultivado (BONONI et al., 1999; COUTINHO, 2005).
De acordo com uma pesquisa realizada por Putzke entre 1990 e 1991, no Brasil são
descritos 136 gêneros e 1011 espécies de cogumelos (TAVEIRA; NOVAES, 2007). A
primeira espécie cultivada no Brasil foi o champignon de Paris (Agaricus bisporus) que é
também a mais conhecida. O início do cultivo em escala comercial, de acordo com os
registros nacionais, data os anos 50, entretanto a popularização no hábito alimentar do
brasileiro é mais recente (COUTINHO, 2005; FURLANI et al., 2005).
Há outras variedades de cogumelos comestíveis produzidas e comercializadas no
Brasil sendo o Shiitake (Lentinus edodes) e o caetetuba ou cogumelo gigante (Pleurotus
ostreatus) (BONONI et al., 1999).

20
1.2.1.1 Propriedades físico-químicas e enzimáticas
Enzimas são substâncias orgânicas, moléculas de proteínas, formadas no interior das
células vivas, portanto, encontradas em todos os alimentos. São capazes de reagir com o meio
extracelular, catalisando reações químicas a uma velocidade bastante rápida (LEHNINGER,
2006).
Os alimentos de origem vegetal sofrem transformações pós-colheita, desencadeadas
por processos bioquímicos e catalisadas por enzimas. Tais reações dependem da integridade
do tecido vegetal, isto é, no transporte, processamento, armazenamento e manipulação dos
cogumelos, frequentemente ocorrem rupturas dos seus tecidos, produzindo alterações
indesejáveis que comprometem principalmente a cor do produto. Estas reações de
escurecimento se devem, na grande maioria das vezes, às oxidações enzimáticas, sendo que a
intensidade do escurecimento tem sido atribuída a Polifenoloxidase (PPO) e Peroxidase
(POD) (VÁMOS-VIGYÁZO, 1981; MICHELS, 2005; ARAÚJO, et al., 2008).
As peroxidases são enzimas pertencentes ao grupo das oxiredutases e encontradas, na
maioria das frutas e legumes em diferentes formas moleculares, na forma solúvel e acoplada,
sendo que as propriedades catalíticas são influenciadas pelo cultivar, crescimento e fases
fisiológicas. A presença da PPO nos alimentos tem sido associada com a cadeia respiratória e
com a resistência das plantas à infecção de bactérias e vírus e a climas adversos. A POD é
encontrada na forma solúvel e iônica ligada à parede das células e está envolvida com
inúmeros processos fisiológicos na planta, tais como amadurecimento e desenvolvimento
(VÁMOS-VIGYÁZO, 1981).
Como produto final da oxidação das peroxidases forma-se a quinona, que pode se
polimerizar formando um pigmento escuro insolúvel, denominado de melanina, ou pode
reagir não enzimaticamente com outros compostos fenólicos, aminoácidos e proteínas,
formando o mesmo composto (ARAÚJO, 2008; LUPETTI et al., 2005).
As reações enzimáticas podem ser inibidas ou estimuladas pela concentração da
enzima, do substrato e dos íons hidrogênio e também pelas condições de temperatura. O
controle do teor de oxigênio é fundamental para minimizar as reações de oxidação. Por isso
que na indústria de alimentos, um método físico empregado para controlar o escurecimento
enzimático tem sido o fechamento a vácuo do produto, processo que retira quase que
totalmente o oxigênio do interior da embalagem (esse método será abordado ainda neste
capítulo) (ARAÚJO et al., 2008; LUPETTI et al., 2005). Esses mesmos autores explicam que
a refrigeração apenas diminui a atividade enzimática, enquanto que o calor tem sido o método

21
mais simples e usado para controlar as reações químicas. Normalmente, temperaturas em
torno de 75ºC, por 1 a 5 minutos, são suficientes para esta inativação.
Por isso que uma das técnicas usadas pelas indústrias de alimento para minimizar
esses efeitos tem sido o método de branqueamento, que consiste na imersão do alimento em
água aquecida, com temperatura e tempo específicos para cada um, e em seguida em água fria
pelo mesmo período de tempo previamente empregado. Tem por objetivo a redução da carga
microbiana superficial, inativação das enzimas e eliminação do oxigênio dos tecidos
(GOMES; SANTOS, 2006). Entretanto, a POD é uma enzima altamente estável ao tratamento
térmico e sua inativação tem sido frequentemente usada como índice da efetividade do
branqueamento (CLEMENTE, 1996).
Portanto, a participação das enzimas na produção e na tecnologia de alimentos pode
ser, ora vantajosa, quando melhoram processos e condições de armazenamento do produto,
ora indesejável, quando ocorre deterioração do mesmo (ARAÚJO et al., 2008).
Além das enzimas, os carboidratos, proteínas, vitaminas e compostos fenólicos são
considerados os principais componentes químicos presentes nos cogumelos. Dentre todas as
frutas e vegetais, os cogumelos apresentam uma maior taxa de respiração celular, o que
implica, mesmo após a colheita, na transformação dos carboidratos em energia, dióxido de
carbono e água proporcionando o processo de maturação do alimento (MAU et al., 1993).
A maior parte dos carboidratos encontrados nos alimentos ocorre na forma de
polissacarídeos, os quais desempenham funções biológicas de reserva ou estruturais de parede
celulares de vegetais superiores (BACH, 2006). Nos cogumelos, esses polissacarídeos são
formados por unidades de monossacarídeos unidos entre si por ligações glicosídicas alfa ou
beta, constituindo a celulose, não digerida pelas enzimas digestivas dos animais (SHARON;
LIS, 1993; FALCÃO, 2008). As β- glucanas da parede celular dos cogumelos estão ligadas à
quitina, um polissacarídeo estrutural, insolúvel em água e indigerível pelos seres humanos.
Poucos relatos são encontrados na literatura a respeito das modificações sofridas pela fibra e
pela quitina durante processos de cozimento e métodos de preservação (KALAC, 2009).
No caso do basidioma de P. ostreatus (YOSHIOKA et al., 1985) e de Flammulina
velutipes (SMIDERLE et al., 2006) foi caracterizado a mesma glucana, porém esta apresentou
somente ramificações em O-6 por terminais não redutores de beta-Glcp (beta-polímero de
glicose). Há evidências da ação benéfica das β- glucanas no sistema imunológico e
metabólico do organismo. Cogumelos Pleurotus contêm aproximadamente 55% de β -
glucanas insolúveis e de 10-15% na forma solúvel. São, portanto, considerados excelentes

22
fontes de fibras dietéticas com efeitos medicinais e nutricionais (BOBEK et al., 1997;
BROWN; GORDON, 2001).
Outro nutriente presente em grandes quantidades nos cogumelos, de acordo com
Sigrist e Chitarra (1982, 1998 apud MICHELS, 2005) é a proteína, considerada um dos
substratos das reações enzimáticas em cogumelos, processo este considerado inadequado uma
vez que diminui o valor nutricional e causa alterações na cor do alimento.
Em relação às vitaminas presentes no cogumelo, especialmente a vitamina C, essas
podem sofrer influência negativa com redução considerável no teor vitamínico do alimento
quando submetido a altas temperaturas, a manipulação, ao frio e ao armazenamento
prolongado (MICHELS, 2005).
Os cogumelos são, portanto, considerados alimentos frágeis, devido à sua delicada
estrutura, devendo ser submetidos a processos adequados no pós-colheita e protegidos por
embalagens apropriadas (MAU et al., 1993).
1.2.1.2 Características sensoriais
Segundo Simpsom et al. (1998) a análise sensorial é uma ciência utilizada para
evocar, avaliar, mensurar e interpretar as características de um alimento percebidas pelos
sentidos da visão, olfato, paladar, tato e audição.
Esse tipo de análise, assim como as físico-químicas e microbiológicas, é uma
importante técnica empregada para se conhecer e quantificar as qualidades de um alimento,
como intensidade, extensão, duração de características específicas e gosto e desgosto pelo
produto avaliado (MICHELS, 2005). Sendo assim, pode-se dizer que a qualidade de um
alimento está associada com a sensação que desperta, a qual pode ser prazerosa ou não
(GINANI; PINELLI, 2008).
De acordo com o Instituto Adolfo Lutz (2005) a análise sensorial tem sido
considerada uma análise subjetiva uma vez que depende do julgamento individual baseado
nos órgãos dos sentidos que pode ser influenciado pela experiência e capacidade do julgador,
assim como por fatores externos, dentre os quais as condições do ambiente onde se realiza a
análise, estado emocional e de saúde do julgador e condições e forma de apresentação das
amostras-testes são os que mais interferem.
Ao entrar em contato inicial com o alimento, o indivíduo emite respostas
desencadeadas pelos estímulos recebidos em seus sentidos, resultantes de reações fisiológicas.
A esse primeiro momento denomina-se sensação. Quando o indivíduo filtra, interpreta e

23
reconstrói as informações recebidas pelos sentidos, ocorre a percepção (GINANI; PINELLI,
2008; INSTITUTO ADOLFO LUTZ, 2005).
Segundo o Instituto Adolfo Lutz os atributos que podem ser avaliados de um
alimento ou produto são os seguintes:
Aparência: pode ser considerada a principal característica para que o alimento tenha
aceitação pelo consumidor, por ser o primeiro órgão sensorial estimulado. Refere-se
principalmente a propriedades visíveis como aspecto, cor, transparência, brilho, opacidade,
forma, tamanho, consistência e espessura.
Este atributo permite, por exemplo, identificar o grau de maturação de hortaliças e
frutas, quando há colorações intensas e também avaliar a inadequação para o consumo quando
alterações consequentes de traumas ou avançado estado de maturação se fazem presentes
(GINANI; PINELLI, 2008).
Textura oral e manual: refere-se às propriedades estruturais do alimento ou produto.
A definição da textura ocorre inicialmente pelo contato com a língua, cavidade bucal e com os
dentes; em seguida, a fase da mastigação pode transformar a textura original do alimento
devido ao contato com a saliva. Nos vegetais, a textura é dada pela rigidez dos tecidos,
enquanto que o endurecimento é associado ao metabolismo dos fenóis e à lignificação. A
perda desse atributo está relacionada a alterações na cor e sabor do alimento (PELEG, 2006).
Quanto à textura e aparência o alimento ou produto pode se apresentar, por exemplo:
congelado, cru ou cozido, cremoso, duro, firme ou mole, gorduroso, pegajoso, derretido,
quebradiço, manchado, áspero, etc. Em relação a cor pode ser acromático ou de tonalidades
primárias (azul, vermelho e amarelo), secundárias (laranja, verde e violeta) ou terciárias
(vinho, azul claro, verde-amarelado, cinza, marrom, lilás, bege, etc.) (INSTITUTO ADOLFO
LUTZ, 2005).
Odor e aroma: o odor é perceptível pelo órgão olfativo quando substâncias voláteis
são aspiradas e o aroma se desenvolve durante a mastigação e é dependente de sinais
desencadeados na via retronasal. Muitas vezes sentir o odor do alimento estimula mais o
apetite do que simplesmente visualizá-lo. Por isso que o odor sugere o contato direto e
desperta o prazer de comer (GINANI; PINELLI, 2008).
Embora os lipídeos estejam presentes nos cogumelos em concentrações baixas são
também responsáveis pelo aroma e flavor (sensação fisiológica da interação do paladar e
olfato). O típico flavor dos cogumelos depende dos produtos resultantes das reações
enzimáticas dos ácidos graxos insaturados (principalmente o ácido linoléico) assim como, dos
aminoácidos e substâncias não nitrogenadas (MAU et al., 1993).

24
Segundo os autores acima citados, o aroma mais característico pode ser atribuído a
uma série de compostos voláteis comuns a muitas espécies de cogumelos, com diferenças nas
concentrações e características particulares dos gêneros ou espécies.
Sabor e gosto: esses atributos sensoriais são percebidos pelos receptores gustativos
localizados na língua. Estudos recentes têm demonstrado que tais receptores são capazes de
diferenciar 6 distintos sabores e não somente 4 como previamente conhecido. A saber: o
salgado, pelos sais de cloreto de sódio, o ácido, através do ácido cítrico, málico, tartárico e
outros ácidos orgânicos, o amargo, representado pelo quinino e cafeína, o doce, pela presença
dos açúcares, sacarina, alcoóis, glicóis, o umami, através do glutamato e aspartame, e o sabor
metálico, quando na presença de sulfato heptahidratado de ferro II (DUTCOSKY, 2007).
Avaliados conforme esses dois últimos atributos um alimento ou produto pode ser:
salgado, doce, azedo, picante, adstringente, fétido, inodoro, pútrido, alterado ou característico,
etc. (INSTITUTO ADOLFO LUTZ, 2005).
Sabe-se que nos últimos anos a demanda por alimentos processados vem sofrendo
um crescente aumento em virtude de sua praticidade. Sobretudo, os consumidores esperam e
desejam que esses alimentos apresentem as características sensoriais originais, semelhantes à
sua versão in natura, seguros quanto à qualidade higiênico-sanitária e com adequados valores
nutricionais o que vem exigindo aperfeiçoamentos tecnológicos, tanto nos processos quanto
nas embalagens (MICHELS, 2005).
Logo, os consumidores esperam do alimento processado em relação ao produto in
natura o mesmo ou parecido atributo sensorial de cor, com uma aparência natural que o torne
atraente. Qualquer alteração nessas características pode torná-lo indesejável, acarretando
rejeição (MICHELS, 2005).
Além de influenciar na aceitação e na qualidade do produto alterações em quaisquer
uns dos atributos sensoriais acima descritos interfere no tempo de vida útil do alimento
gerando também prejuízos financeiros (MINIM, 2006).
1.2.1.3 Aspectos microbiológicos
Michels (2005) explica que a qualidade microbiológica dos alimentos está
relacionada com a presença de micro-organismos deterioradores e patogênicos que irão
contribuir com as alterações das características sensoriais do produto durante o período de
vida útil e/ou causar toxi-infecções alimentares no consumidor.

25
Por isso que a Agência Nacional de Vigilância Sanitária (ANVISA) regulamenta os
padrões microbiológicos toleráveis nos alimentos, através da Resolução- RDC nº12 de 2 de
janeiro de 2001. Quando contaminados são considerados em condições sanitárias
insatisfatórias sendo, portanto, impróprios para o consumo humano ou por trazerem risco à
saúde ou mesmo por apresentarem características sensoriais (cor, odor, sabor e textura)
alteradas, tornando-se rejeitados pelo consumidor (BRASIL, 2001).
Existem alguns fatores que aceleram o metabolismo dos micro-organismos
contaminantes dos alimentos que são conhecidas como condições extrínsecas, tais como
temperatura de armazenamento, falta de cuidados no processo de manipulação e métodos de
conservação, presença de gases (CO2 e O2) no ambiente, e condições intrínsecas, que são a
atividade de água do alimento (Aa), pH (condições ácidas ou básicas) e conteúdo de nutrientes
(composição do alimento) (SILVA JUNIOR, 2008).
Quando o alimento é constituído por elevada quantidade de água, com valores de Aa
acima de 0,9, sofrem facilmente contaminação microbiológica, contudo as enzimas presentes
nestes alimentos estão diluídas o que pode, dependendo do alimento, retardar a velocidade das
reações químicas e enzimáticas de deterioração (BOBBIO; BOBBIO, 2001).
Sendo os vegetais, de um modo geral, considerados altamente perecíveis por
apresentarem elevado teor de umidade (Aa >0,85) e um mínimo conteúdo de nutrientes, são
bastante suscetíveis ao crescimento bacteriano. Há entretanto pequena probabilidade de
sofrerem contaminação por fungos uma vez que este grupo de micro-organismos se
desenvolve preferencialmente em ambientes/alimentos com baixa Aa (0,60 a 0,65) (ARAÚJO
et al., 2008; MICHELS, 2005).
Quanto à acidez ou alcalinidade, para os fungos e bactérias são considerados como pH
ótimo para as atividades metabólicas valores de 5,5 e 6,5 respectivamente (SILVA JUNIOR,
2008).
Silva Junior (2008) relata que o crescimento e multiplicação dos micro-organismos
contaminantes de alimentos estão também associados à presença ou não de oxigênio no meio
em que se encontram, sendo que alguns são considerados aeróbios, pois dependem
obrigatoriamente da presença do oxigênio para se desenvolverem. Outros, no entanto, são
chamados anaeróbios, pois se desenvolvem na ausência do oxigênio.
Portanto, a modificação da atmosfera no interior da embalagem, como por exemplo
quando um alimento é embalado a vácuo, aumenta os riscos de contaminação por patógenos
anaeróbicos, uma vez que há redução de oxigênio, favorecendo o crescimento e proliferação

26
de bactérias como Clostridium botulinum, C. perfringens e Listeria monocytogenes
(PHILLIPS, 1996).
1.3 CULTIVO DE Pleurotus
Pleurotus, mais comumente chamado de cogumelo ostra por apresentar o píleo em
forma de concha, possui lamelas esbranquiçadas ou cinzas, estipe excêntrico ou lateral (de 10
mm a 50 mm) podendo alcançar 30 cm de altura (Figura 2) (THORN et al., 2000). É uma
espécie bastante difundida, sendo distribuída por todas as florestas tropicais do mundo,
inclusive no Brasil; é saprofítico, ou seja, cresce em restos de madeira, usualmente em árvores
mortas ou em toras caídas (COUTINHO, 2005; GUNDE-CIMERMAN, 1999; GUNDE-
CIMERMAN et al., 1993, 1995).
Mais de 1.000 espécies de Pleurotus spp foram descritas no mundo, no entanto apenas
50 destas são reconhecidas como válidas para este gênero. As dificuldades quanto à
identificação taxonômica, deve-se à grande variedade de espécies e sua ampla distribuição no
mundo. Os cogumelos Pleurotus ostreatus são popularmente chamados de cogumelo ostra,
pertencem à ordem Agaricales e à família Pleurotaceae. Suas espécies e linhagens oferecem
uma variedade de cores como cinza-escuro, branco, marrom, amarelo, salmão entre outras
(GUZMAN, 2000).
O gênero Pleurotus é um dos mais diversos grupos de cogumelos cultivados que tem
importância econômica e valor medicinal. A sua produção é a menos oneroso por ser de mais
fácil cultivo dentre todos os outros cogumelos comestíveis cultivados industrialmente, além
de crescerem em um número diferente de substratos (COHEN et al., 2002; GUNDE-
CIMERMAN, 1999; GUNDE-CIMERMAN et al., 1993, 1995; BEETZ; KUSTUDIA, 2004).
Segundo Kües e Liu (2000), por volta de 1900 o cultivo comercial de Pleurotus
iniciou-se nos Estados Unidos; nos anos conseguintes foi introduzido na Índia e na Europa.
Registros demonstram que as primeiras espécies do gênero cultivadas foram: P. eryngii. e P.
florida em 1958, P. flabellatus em 1962, P. cystidiosus em 1969, P. sajor-caju em 1974 e P.
citrinopileatus em 1981.

27
Figura 2: Aspectos gerais do cogumelo Pleurotus ostreatus. Fonte: Senedeze, 2009
Até os anos 70 o seu consumo dependia da colheita desses cogumelos diretamente
da natureza e a partir de então se iniciou o cultivo em escala industrial. No Brasil, entretanto,
o aumento da produção e consequente aumento de consumo só ocorreu na segunda metade da
década de 80, após intensos trabalhos de divulgação (BONONI et al., 1999).
Dentre as espécies mais conhecidas estão P. ostreatus (shimeji), P. ostreatoroseus
(salmão), P. ostreatus variedade florida (branco), P. eryngii (eryngii), P. citrinopileatus
(amarelo). Destas, o P. amarelo é utilizado na medicina e as demais são amplamente usadas
na culinária devido às suas texturas e sabores agradáveis (KÜES; LIU, 2000; SOUZA, 2006;
URBEN, 2002).
Esse cogumelo é conhecido pelos orientais como Hiratake e no caso de cogumelos
colhidos em pencas no estágio jovem (basidiomas entre 1 cm e 2 cm) e crescidos sob
condições de menor iluminação e ventilação, tem sido denominado Shimeji branco (BONONI
et al., 1999; EIRA, 2004).
O cultivo dos cogumelos do gênero Pleurotus pode ser dividido nas seguintes
etapas: compostagem, pasteurização, semeadura, incubação ou inoculação, produção e
colheita (Figura 3).

28
Figura 3: Etapas do cultivo de cogumelos Pleurotus spp. Fonte: Beetz, Kustudia, 2004. Tradução: Senedeze, 2009 A compostagem, etapa que apresenta 30 dias de duração, compreende o processo de
preparo do substrato ou composto onde o cogumelo vai crescer. O Pleurotus apresenta a
capacidade de crescer em diversos tipos de substratos sendo que no Brasil, o mais usual é o
bagaço de cana-de-açúcar, complementado com farelo de arroz ou soja para aumentar a
produtividade (BONONI et al., 1999; HERRERA, 2001).
Estudos recentes têm demonstrado algumas diferenças no conteúdo nutricional de P.
ostreatus e de P. sajor-caju quando cultivados em palha de arroz e banana (BONATTI, et al.,
2004). A palha de feijão pode ser considerada um bom resíduo para o cultivo de cogumelo P.
sajor-caju, quando comparada à palha de milho e a casca de café, pois apresenta melhor
eficiência biológica sem necessidade de enriquecimento, representando menor custo de
produção (DIAS et al., 2003).
Esterilização e preparo dos
meios de ágar
Isolamento dos micélios do cogumelo
Propagação da cultura pura
Inoculação da semente Esterilização da
semente inoculada
Inoculação da serragem
Produção do inóculo
Dispondo o inóculo na bandeja
Inserindo em toras
Cultura em tora
Cultura em tronco
Cultura em monte
Cultura em saco Cultura em coluna
Cultura em parede
Cultura em bandeja Inoculação do substrato

29
Portanto, os cogumelos Pleurotus podem se desenvolver em diversos tipos de
substratos, desde que estes contenham celulose, hemicelulose e lignina, além de outros
elementos químicos para o seu crescimento e desenvolvimento (BONONI et al., 1999).
A etapa seguinte compreende a pasteurização, que tem por finalidade eliminar
qualquer micro-organismo patogênico ou competidor pela colonização do substrato. Consiste
em um tratamento térmico, cujos intervalos variam de 55 a 90ºC, de 8 a 96 horas, feito pela
passagem de vapor de água através do composto. (BONONI et al., 1999; EIRA, 2004).
Após o processo de pasteurização o composto deve ser imediatamente resfriado e
semeado. Nesta etapa, realizada mais comumente em sacos de polietileno, os inócuos são
distribuídas uniformemente no composto para facilitar a colonização do substrato e garantir o
não crescimento de contaminantes (BONONI et al., 1999).
Bononi et al. (1999) descrevem que durante a fase de incubação (cerca de 15-20 dias),
há uma intensa atividade biológica, na qual o micélio degrada o composto, extraindo os
nutrientes e formando uma massa branca e compacta.
As condições de temperatura e iluminação devem ser controladas para que não haja
interferência na colonização do composto. Por isso, deve ser mantido em torno de 25-28ºC e
na ausência de luz para que assim os corpos de frutificação surjam entre 8 e 18 dias após este
processo (RAJARATHNAM; BANO, 1987). A Figura 4 retrata os diferentes estágios de
crescimento do P. ostreatus.
Para que os corpos de frutificação tenham a qualidade desejada e garantida a umidade
deve ser de 95% para evitar o ressecamento do composto e do cogumelo; a temperatura deve
ser a mesma da empregada na fase de inoculação; a luminosidade de 2000 lux/ hora por 12
horas ao dia e a troca gasosa deve ser de pelo menos 4 vezes por hora, para evitar acúmulo de
CO2. Por conseguinte, a primeira colheita ocorrerá entre 30 e 50 dias após a inoculação
(BONONI et al., 1999; EIRA, 2004; HERRERA, 2001).

30
1.4 VANTAGENS NA ALIMENTAÇÃO HUMANA, VALOR
NUTRICIONAL E MEDICINAL
Atualmente, os cogumelos comestíveis apresentam não somente uma importância
gastronômica, mas também medicinal tal como a espécie Agaricus blazei (Cogumelo do sol®)
que é considerado um alimento nutracêutico ou funcional fisiológico (FURLANI, 2004). No
mundo há cerca de 700 espécies consideradas farmacológicas com atividades comprovadas
(LULL et al., 2005).
Todavia, a maioria dos estudos realizados com cogumelos com a finalidade de
averiguar suas propriedades medicinais ainda ocorrem de maneira experimental ou in vitro.
Poucos são aplicados em seres humanos, pois ainda não se conhece muito claramente o
mecanismo de ação de seus elementos no metabolismo humano. Porém, estudos demonstram
que os cogumelos da espécie Pleurotus estimulam o sistema imunológico e têm se mostrado
Figura 4: Diferentes estágios de crescimento do Pleurotus ostreatus. A: fase inicial; B: intermediária; C: estágio final de crescimento e momento ideal para a colheita. Fonte: Senedeze, 2009
A
C
B

31
importantes aliados no tratamento complementar de doenças, como câncer, hepatite, HPV
(Vírus do Papiloma Humano), AIDS. Além de auxiliar no controle do diabetes mellitus,
hipercolesterolemia, hipertensão, diminuir riscos de doenças cardiovasculares e possuir
efeitos antioxidantes, anti-inflamatórios e de varredura dos radicais livres (AHN, et al., 2006;
NOVAES; FORTES, 2005; SMITH et al., 2002).
Tais pesquisas com cogumelos medicinais têm descoberto compostos, principalmente
as β-glucanas, que podem modular positivamente as respostas biológicas de células
imunológicas, estimulando as células T (tais como citotóxicos, macrófagos, monócitos,
neutrófilos, células natural killer) e intensificando reações químicas, elucidando os
mecanismos de ação (SMITH et al., 2002).
No caso dos Pleurotus spp., as lectinas, glicoproteínas de origem não imune e o
lavastatin, potente agente hipocolesterolêmico, isolados por Liener et al. em 1986, continuam
sendo alvo de inúmeros estudos devido aos efeitos medicinais associados (LI et al., 2008).
No Brasil, em 2005, Novaes e Fortes através de estudo clínico com 20 pacientes
acometidos de leucemia não-linfocítica aguda, divididos em grupo experimental, tratado com
20g de A. blazei, 3 vezes ao dia e grupo controle (apenas tratado com quimioterapia)
comprovou que no 1º grupo algumas células sanguíneas retornaram aos níveis normais entre 7
e 8 dias após todas as seções de quimioterapia, além de aumento nas quantidades de IgM. No
grupo controle não houve modificações nas taxas desses anticorpos e as células sanguíneas
retornaram a níveis normais, apenas em alguns pacientes, num período entre 14 e 21 dias.
Resultados semelhantes foram evidenciados por Gennari (2006), que revelou melhoras
no perfil imunológico de pacientes tratados com cogumelos Agaricus, evidenciando aumento
de 75,2% das células natural killer (NK), repercutindo, em certos casos, na remissão completa
da metástase tumoral.
Quanto ao valor nutricional dos cogumelos, vários estudos têm sido desenvolvidos a
fim de avaliar a composição destes e determinar o quanto contribuem em nutrientes na
alimentação do homem. O que se sabe é que a composição desses alimentos varia de acordo
com a origem, solo, clima e condições de cultivo (BONONI et al., 1999).
Quase 90% do cogumelo é água e dos 10% que restam há uma boa quantidade de
fibras, proteínas e vitaminas. É constituído por um baixo teor de lipídeo (menos de 5%) sendo
consequentemente um alimento pouco calórico (BANO; RAJARATHNAM, 1988; MANZI et
al., 1999; RIBEIRO, 2003). O Shiitake, Shimeji e o Cogumelo de Paris apresentam 33
Kcal/100g, 36 Kcal/100g e 30 Kcal/100g, respectivamente, podendo assim ser usados, sem

32
restrição, em dietas hipocalóricas, como por exemplo, para a redução de peso (FURLANI,
2004).
Considerando o elevado conteúdo protéico dos cogumelos frescos seu cultivo tem sido
uma alternativa para incrementar a oferta de proteínas às populações de países em
desenvolvimento e com alto índice de desnutrição e também utilizado pelos vegetarianos
como uma forma de suprir as necessidades diárias desse macronutriente (VILELA, 2006;
FURLANI, 2004).
Bonatti et al. (2004) estudaram o conteúdo nutricional de cogumelos Pleurotus spp.
obtendo valores protéicos entre 1,54-3,1%, teores estes similares a 100mL de leite de vaca
integral ou superior a 100g de vários vegetais porém inferior ao encontrado em uma porção de
ovos e carnes.
Quanto ao perfil de aminoácidos essenciais dos diferentes cogumelos da espécie
Pleurotus, Manzi et al. (1999) demonstraram que o ácido glutâmico, aspartato e arginina são
os componentes mais abundantes. Os limitantes são lisina e leucina, contudo, mesmo assim os
valores de proteínas destes cogumelos confirmam sua boa atividade biológica.
Analisando a composição nutricional dos cogumelos em relação às vitaminas não
foram encontradas grandes quantidades de vitamina B1 (0,07mg/100g), B12 (0,05µg/100g) e D
(0,02µg/100g), em Pleurotus ostreatus estudados por Matilla et al. (2001), na Finlândia.
Resultados semelhantes foram evidenciados por Furlani (2004) para as vitaminas B1 e B2,
que variam de 0,004 a 0,08mg/100g e 0,04 a 0,3mg/100g, respectivamente. A vitamina C
também pode ser encontrada em cogumelos, porém em valores inexpressíveis para a dieta
(FURLANI, 2004).
Porém, pode ser considerado uma boa fonte de riboflavina, niacina e folatos, pois
apresenta 0,20mg/100g, 5,2mg/100g e 5,1µg/100g, respectivamente (MATILLA et al., 2001).
Para ser um alimento considerado fonte de proteína, carboidratos, vitaminas e
minerais de acordo com a Portaria nº 27 de 1998, esses devem estar presentes e contemplar no
mínimo 15% da IDR (Ingestão Diária Recomendada). Categorizado como um alimento com
alto teor do nutriente, quantidades superiores a 30% da IDR devem estar presentes (BRASIL,
1998).
Portanto, os cogumelos não podem ser considerados alimentos fontes de vitaminas B1,
B2, B12, D e C.
Em uma pesquisa desenvolvida por Sturion e Ranzani (2000), com 7 espécies distintas
de Pleurotus, as autoras confirmaram que o macromineral mais abundante nestes é o potássio
(328mg/100g de cogumelo fresco), correspondendo a 16,4% da IDR e o fósforo (141mg/100g

33
de cogumelo fresco), cobrindo cerca de 17,65% das necessidades diárias. Corroborando esses
resultados Matilla et al. (2001) também encontraram como minerais mais abundantes nas
espécies A. bisporus branco, A. bisporus marrom, P. ostreatus e Lentinus edodes, o K e P,
respectivamente 364 e 98mg/100g, 359 e 101mg/100g, 298 e 111mg/100g e 224 e
73mg/100g, todos referentes ao peso fresco.
Em cogumelos frescos, o teor de sódio varia de 1 a 3,4mg/100g (MATILLA et al.,
2001) e de 0,52 a 3,8mg/100g. (STURION; RANZANI, 2000). São então considerados
alimentos que não contém sódio (exceto os em conserva) por apresentarem quantidades
inferiores a 5mg deste mineral/100g do alimento, atendendo às especificações da Portaria nº
27.
Sendo assim, os cogumelos devem ser incluídos nas dietas de pacientes hipertensos,
uma vez que a literatura recomenda para o tratamento dessa patologia, alimentos pobres em
sódio e ricos em potássio (KRUMMEL, 2005).
O cálcio não foi encontrado em quantidades significativas em nenhuma espécie de
cogumelo comestível estudada por Manzi et al. (1999), Matilla et al. (2001), Sturion e
Ranzani (2000).
Não há um consenso na literatura quanto a quantidade de ferro disponível nos
cogumelos comestíveis, pois os resultados dos estudos recentes divergem entre si e
contradizem a citação feita por Bano e Rajarathnam (1988) sobre a biodisponibilidade do
nutriente (MATILLA et al., 2001).
Tanto os cogumelos cultivados na Finlândia quanto no Brasil não podem ser
considerados fontes de ferro, pois quantidades muito pequenas foram encontradas, variando
de 0,22 a 0,43mg/100g (fresco) e 1,06 a 1,6 mg/100 g (fresco), respectivamente (MATILLA
et al., 2001; STURION; RANZANI, 2000). Atendem no máximo 11,4% da IDR para adultos
(14mg/dia) (BRASIL, 2005).
Contudo, cogumelos chilenos, também estudados por Sturion e Ranzani (2000),
podem ser considerados alimentos com alto teor de ferro, pois apresentam 5,73mg/100g de
peso fresco, suprindo cerca de 41% da IDR.
Na Turquia foram estudadas 30 espécies de cogumelos comestíveis, sendo que, por
grama de peso seco, foram obtidas quantidades de ferro variando entre 5-84,2mg
(GENÇCELEP et al., 2009).
Além da origem do cogumelo, o tipo de substrato e os compostos utilizados para a
suplementação durante o cultivo devem ser considerados. Pedra et al. (2009) citaram que P.
ostretaus cultivados em casca de coco e suplementados com farelo de arroz e de trigo

34
apresentaram teores de ferro que variaram entre 10 e 20 mg/100 g de amostra fresca.
Entretanto, no Brasil, a casca de coco não é usualmente empregada como substrato para o
cultivo de Pleurotus spp. o que pode interferir no teor de ferro do cogumelo (BONONI et al.,
1999; MODA; HORII; SPOTO, 2005a; BERNARDI et al., 2007).
Por isso, baseando-se nas informações acima descritas e por haver divergências nas
quantidades de ferro encontradas nos cogumelos cultivados no Brasil, eles não podem ser
considerados fonte de ferro na alimentação humana, porém podem ser utilizados como
coadjuvantes no tratamento e prevenção de anemia ferropriva.
Quanto à fibra, P. ostreatus crus e grelhados podem ser considerados fonte (mínimo
3g/100g do alimento), pois em 100 g de cogumelo há respectivamente 5,24g e 4,1g de fibra,
sendo que a maior fração é de fibra insolúvel (89% em ambos) (BRASIL, 1998; MANZI et
al., 2001).
A composição do cogumelo em fibra alimentar pode variar em relação as suas
estruturas e substratos cultivados. Em 1996, Cheung estudou a haste e o chapéu de cogumelos
da espécie L. edodes, L. shimeji e Pleurotus. Os resultados demonstraram valores diferentes
entre os cogumelos e suas estruturas, sendo que a espécie L. edodes apresentou maiores teores
deste nutriente tanto na haste quanto no chapéu (respectivamente 44,9% e 52,7%), enquanto
que o L. shimeji teve os menores valores no chapéu (39%) e a espécie Pleurotus na haste
(42,2%).
Resultados bastante inferiores foram relatados em 2003, por Bonatti et al., para as
espécies P. ostreatus, 9,41%, e P. sajor-caju, 8,13%, cultivados em folha de bananeira
demonstrando que o substrato utilizado pode neste caso ter interferido na quantidade de fibra
constituinte do cogumelo. Todos esses resultados são relativos à base seca.
Furlani (2004) encontrou em seu estudo 20,44g de fibra em 100g do alimento seco
para o champignon, resultado este semelhante ao de Cheung (1997) (18,2g/100g de base
seca). No shiitake, a quantidade de fibra é de 41,92g em 100g (base seca) e no shimeji de
39,62g/100g de base seca (FURLANI, 2004).
Esse componente presente nos cogumelos apresenta também propriedades funcionais,
pois as fibras solúveis estão associadas à redução dos níveis de colesterol sérico e prevenção
de câncer colo-retal. A fração insolúvel, relacionada ao aumento do bolo fecal pela grande
capacidade de absorção de água, previne a constipação e suas complicações (POURCHET-
CAMPOS, 1998; SILVA et al., 2002).
Outro benefício nutricional dos cogumelos nacionais é o alto teor de ácido fólico,
variando de 459 a 1431 microgramas/100g do alimento, dependendo do lote e da espécie. Por

35
ser a Ingestão Diária Recomendada para este nutriente de no mínimo 48 mcg para os lactentes
e de no máximo 355 mcg para as gestantes os cogumelos nacionais podem ser considerados
excelentes fontes de ácido fólico uma vez que atendem a 100% da recomendação diária
(BRASIL, 2005; FURLANI, 2004).
No organismo humano, em condições normais, são armazenados 5-20mg de folatos no
fígado e em outros tecidos (MASTERS, 2005). Esta vitamina é essencial para as reações
metabólicas específicas no meio celular e vital para o funcionamento normal do organismo.
Participa também de dois processos biológicos fundamentais: atua como cofator para as
enzimas implicadas na biossíntese de RNA e DNA e como cofator na formação da metionina
(VANNUCCHI; JORDÃO JUNIOR, 1998; RIBEIRO et al., 2002).
Santos e Pereira (2007) relatam o importante papel do ácido fólico na prevenção das
más formações fetais e congênitas (espinha bífida, anencefalia, lábio leporino e fenda
palatina), além de prevenir males cardíacos e doenças cardiovasculares, desordens mentais,
câncer e o mal de Alzheimer.
Portanto, o cogumelo P. ostreatus é uma ótima fonte de carboidratos simples, tem alto
conteúdo de fibra dietética, vitaminas e minerais, contém proteína de boa qualidade biológica
e a maioria dos aminoácidos essenciais ao metabolismo humano (GUNDE-CIMERMAN,
1999; GUNDE-CIMERMAN et al., 1993, 1995).
1.5 PRODUÇÃO, COMERCIALIZAÇÃO E CONSUMO DE
COGUMELOS
Em alguns países do mundo, como por exemplo na China e no Japão, a produção de
cogumelos comestíveis ocorre há aproximadamente 800 anos (FURLANI, 2004).
No Brasil, a produção de cogumelos comestíveis foi iniciada na década de 1970, com
um volume pouco expressivo (somente 700 toneladas no ano de 1975), enquanto que nesse
mesmo período, países como os Estados Unidos e a França produziram, respectivamente,
135.700 e 115,600 toneladas de cogumelos (BONONI et al., 1999).
O Brasil produz quantidades insignificantes no contexto mundial (0,12%) e
infelizmente, não vem crescendo na mesma escala, produzindo um volume de 1.600 toneladas
em 1988, aumentando apenas 6,6% em 8 anos. O maior percentual de produção nacional é de
champignon, representando cerca de 90% (VILELA, 2006; BONONI et al., 1999).

36
Nossa produtividade é extremamente baixa quando comparada a países da Europa e
América do Norte, que investem em tecnologia no cultivo e marketing para a comercialização
e divulgação do produto. Por exemplo, para a produção do Agaricus bisporus, utilizando-se
100 quilos de composto seco se obtém de 7 a 15 quilos de cogumelos frescos, enquanto que
países europeus produzem cerca de 30-35 quilos do produto fresco na mesma quantidade, em
peso seco, de substrato (EIRA, 2004).
O Pleurotus ostreatus apresenta cultivo mais produtivo do que o champignon, pois
para se produzir 1 quilo utiliza-se 2 quilos de composto seco, porém quantidades muito
limitadas se comparadas à produtividade dos Estados Unidos, que é de 1:1. Alguns fatores,
além da qualidade do substrato utilizado, podem explicar esta baixa produtividade, tais como,
pragas e doenças que afetam a cultura, métodos de cultivo na sua maioria ainda rudimentar,
baseado em cultura familiar, e pouco investimento e modernização dos processos de
fabricação dos cogumelos (BONONI et al., 1999).
Segundo a Organização das Nações Unidas para Agricultura e Alimentação - FAO,
em 2007, a produção mundial de cogumelos chegou a valores aproximados de 3,4 milhões de
toneladas (Figura 5), sendo que a Ásia e a Europa foram os continentes que mais produziram,
representando 54,13% e 30,28%, respectivamente (FAOSTAT, 2009).
Entre 1986 e 1991, segundo dados publicados por Chang (1993), citado por Royse
(1996), as espécies de Pleurotus apresentaram um crescimento de aproximadamente 442,6%
em sua produção. Em 1991 ocupavam o 2º lugar entre as espécies produzidas no mundo,
depois do A. bisporus (37,2%) (BANO; RAJARATHNAM, 1988).
Figura 5: Produção mundial de cogumelos comestíveis, em toneladas, por décadas. Fonte: FAOSTAT, 2009

37
O estado de São Paulo é o maior produtor nacional de cogumelos comestíveis. O
município de Mogi das Cruzes é responsável pelo cultivo de pelo menos 70% dos cogumelos
comercializados no Brasil (VILELA, 2006).
De acordo com um estudo de caso realizado em 2008 por Vieira, na região do Alto
Tietê, os cogumelos comestíveis representam, no cenário nacional, 55,76% dentre as culturas
agrícolas produzidas e 68,04% de participação no estado de São Paulo.
No que se refere a comercialização os cogumelos podem ser vendidos in natura, em
conserva, congelados, desidratados picados, fatiados ou em pó. Na sua grande maioria,
empregados na alta gastronomia, compondo pratos de renomados gourmets. O champignon é
normalmente comercializado fresco e cozido a granel ou fatiado; o shiitake e hiratake são
encontrados frescos ou desidratados. O cogumelo do sol®, mais usado como medicinal, é
vendido na forma desidratada, fatiado ou em pó (VILELA, 2006).
O shimeji (Pleurotus spp.), no mercado interno, é comercializado fresco. Em outros
países, de acordo com Stamets e Chilton (1996) citado por Apati (2004) o método de
conservação tradicionalmente utilizado é a desidratação. Todos os cogumelos desidratados
podem ser reconstituídos por imersão em água, por 1 a 2 horas, porém, as texturas originais
são modificadas.
O Brasil é um país com atividade muito maior de importação do que exportação deste
alimento. Apenas em 2000 é que passamos a exportar maiores quantidades de cogumelos em
conserva e in natura, se comparado com o volume de importação (VILELA, 2006).
Isto demonstra que o Brasil está sendo “obrigado” a aperfeiçoar as técnicas de cultivo,
colheita e comercialização visto que o mercado externo é extremamente exigente, não
somente no que se refere aos aspectos de qualidade e estética, mas também aos produtos
fungicidas, pesticidas, líquidos branqueadores e produtos conservantes empregados nos
processos de fabricação (BONONI et al., 1999).
Por isso que a indústria de alimentos está cada vez mais se preocupando com a
preservação dos produtos que comercializa, buscando inovações e adequados processos que
mantenham o sabor, a aparência, o valor nutritivo e previna a formação de compostos tóxicos
em alimentos in natura ou processados (LUPETTI et al.,2005).
Segundo a Agência Nacional de Vigilância Sanitária (2004), a vida de prateleira ou
shelf life de um produto é determinada pelas alterações nos aspectos químicos, físicos e
microbiológicos, isto é, o tempo máximo que ele pode ser conservado em determinadas
condições de temperatura, umidade, luz, etc. sofrendo pequenas alterações que ainda o torne
aceitável para o consumo humano.

38
Para Monteiro (2008), o shelf life auxilia na avaliação sensorial garantindo segurança
microbiológica e preservação das características químicas, físicas e nutricionais dos
alimentos.
A estabilidade de um produto, no que concerne às características sensoriais, conteúdo
nutricional e suas propriedades físico-químicas, sofrem influência de inúmeros fatores tais
como tempo entre a colheita e o consumo final, temperatura de transporte e armazenamento,
presença de luz e oxigênio, umidade relativa do ar, tipo e permeabilidade da embalagem e
reações químicas no alimento (ANJOS, et al., 2003; ANVISA, 2004).
A desidratação, congelamento, embalagens com controle e modificações na atmosfera,
preparo de líquidos acidificados como conserva são alguns dos processos mais comumente
empregados pela indústria de alimentos para conservar e aumentar o tempo de vida útil dos
cogumelos (BANO; RAJARATHNAM, 1988; MANZI et al., 2004; WALDE et al., 2006).
Os cogumelos são considerados alimentos com alta perecibilidade, perdendo suas
características de frescor horas após a colheita. Bano e Rajarathnam (1988) descreveram que
Pleurotus armazenados a 21-25ºC apresentaram sinais de deterioração 12 horas depois de
colhidos.
Cogumelos frescos da espécie Pleurotus, embalados sob atmosfera modificada e
mantidos à temperatura de 4ºC aumentou a vida útil para 10 dias, mantendo aparência de
produto fresco (SAPATA et al., 2005). Por isso que a refrigeração, associada a outros
processos tecnológicos, tem sido empregada na conservação de cogumelos.
O processo de embalar os alimentos a vácuo o qual consiste na retirada do ar presente
no interior da embalagem através de uma bomba de vácuo, cria um ambiente anaeróbico que
previne o crescimento de bactérias aeróbicas e fungos, além de preservar a cor, evitar a
rancificação e ressecamento do alimento (SILVA JUNIOR, 2008).
Baruffaldi e Oliveira (1998) explicam que esse procedimento, usualmente empregado
pelas indústrias para a maioria dos alimentos, tem por finalidade, não somente retirar o
oxigênio em contato com o alimento, o qual pode ser danoso por modificações oxidativas e
proliferação microbiana, mas também reduzir o volume da embalagem e eliminar a umidade
do ar interno. As características e temperatura do produto são fatores que interferem no
processo de fechamento a vácuo das embalagens.
Dessa forma, estudos sobre a composição de nutrientes em cogumelo Pleurotus spp. e
identificação do melhor método para o processamento desse alimento tornam-se
fundamentais, a fim de prevenir e/ou minimizar perdas de nutrientes, além de garantir
qualidade sensorial e microbiológica desse alimento.

39
Por ser o Pleurotus um cogumelo de sabor bem agradável e por haver disponibilidade
deste no mercado, o consumo aumentou significativamente nestes últimos anos e, hoje em
dia, ocupa o segundo lugar da produção mundial dentre os cogumelos comestíveis (BONONI
et al., 1999; VILELA, 2006).
De 1968 até hoje, o consumo anual aumentou de 20 toneladas para cinco mil
toneladas, mas o consumo per capita no país ainda é muito baixo, apenas 30 gramas por ano,
contra 2 quilos na França, 1,3 na Itália e 1,9 no Reino Unido. Cerca de 60% da produção
nacional são consumidos in natura e os 40% restantes são utilizados pela indústria de
conserva (DUPRAT; SOUZA, 2003; CLARKE, 2007).
Em uma pesquisa conduzida por Duprat e Souza (2003) a fim de conhecer a opinião e
os hábitos dos consumidores de cogumelos comestíveis, evidenciou-se que 75,23% deles
eram do sexo feminino e mais da metade apresentava nível de escolaridade concentrada nas
faixas de 2º grau e superior completos, indicando que o consumo pode estar relacionado ao
grau de conhecimento à cerca do produto e de suas propriedades.
A falta de informação sobre o produto, o custo um tanto quanto elevado, o
desconhecimento das técnicas de preparo e formas de utilização dos cogumelos foram
também resultados encontrados por Duprat e Souza (2003) para a não aquisição do produto.
No México, um estudo bastante semelhante ao realizado no Brasil, Mayett et al.
(2006) diagnosticaram que cerca de 50% dos consumidores urbanos compravam cogumelos,
independente da classe social. As razões para não comprá-los e consumi-los, segundo relato
dos próprios consumidores, foram: não gostar, desconhecimento inclusive dos métodos de
preparo, alimento prejudicial e impuro. O preço foi considerado como moderado e muito caro,
sendo também uma das justificativas para a não aquisição. O consumo médio anual, de acordo
com essa pesquisa, foi de 902g per capita.
Portanto, fica evidente que, apesar dos cogumelos, há séculos, serem utilizados e
conhecidos quanto as suas vantagens e benefícios nutricionais e medicinais, os brasileiros
ainda não possuem o hábito de consumi-lo por desconhecimento.

40
2 OBJETIVOS
a) Geral
Avaliar qual o melhor método para conservação do Pleurotus ostreatus quanto ao aspecto nutricional, químico e físico, levando em conta o período de armazenamento.
b) Específicos
- Averiguar, por análises bioquímicas, a presença e quantidade de proteínas, fenóis, α e β-glucanas e atividade enzimática da peroxidase e polifenoloxidase tanto na água de tratamento quanto no cogumelo. - Determinar e quantificar a vitamina C existente no cogumelo para assim classificá-lo como um alimento fonte desta vitamina. - Avaliar a influência do tempo de armazenamento e o método de conservação nas características sensoriais do P. ostreatus. - Analisar a comunidade microbiana presente no Pleurotus ostreatus.

41
3 MATERIAIS E MÉTODOS
A presente pesquisa foi dividida em três diferentes etapas. Na primeira procurou-se
avaliar qual o melhor método de lavagem para preparação do cogumelo, a fim de manter ou
melhorar os aspectos bioquímicos e minimizar as perdas nutricionais. Na segunda,
confirmado o método de lavagem, foi observado o melhor tempo de tratamento para
conservação do Pleurotus ostreatus. Na última etapa se aplicou as metodologias escolhidas
como padronização a dois tamanhos de cogumelos mais consumidos no mercado interno
brasileiro, a fim de se obter os objetivos propostos.
3.1 COLETA DOS COGUMELOS Pleurotus ostreatus
Os cogumelos foram doados pela empresa Brasmicel (localizada em Suzano- São
Paulo). A coleta ocorreu no mês de agosto, no período da manhã, de maneira manual e com a
utilização de luvas. Após a colheita, os cogumelos eram colocados em sacos plásticos e
transferidos para geladeira de isopor, com gelo no fundo, até a Universidade Nove de Julho
(Uninove). Todo o material coletado foi de um único lote.
O substrato, à base de palha de milho, bagaço de cana-de-açúcar e capim (fórmula em
sigilo), tem sido preparado por esta empresa e foi utilizado em todos os lotes. O tamanho dos
cogumelos coletados foi, quando pequeno (Figura 4B) e quando grande (Figura 4C), sempre
de vários sacos e repetido por diferentes meses. É importante frisar que os sacos são mantidos
em estufa fechada, em ambiente escuro e umidade controlada mantendo os cogumelos com
pouca água.
3.2 PADRONIZAÇÃO DO MÉTODO DE TRATAMENTO (PRIMEIRA
ETAPA)
P. ostretaus de tamanhos pequeno e grande foram separados em grupos, com 20
gramas cada, sendo submetidos a dois tipos de lavagem e preparação. Todas as amostras
foram lavagem em água corrente por 1 minuto.

42
Em seguida, as amostras controle foram imersa em água destilada estéril contendo
0,25% de ácido cítrico, utilizado como antioxidante, por um período de 4 minutos.
Posteriormente, foram realizadas as análises bioquímicas para a quantificação de proteínas e
fenóis e avaliação da atividade das enzimas peroxidase (POD) e polifenoloxidase (PPO). As demais amostras foram fervidas, por 4 minutos, em água destilada mais 0,25% de
ácido cítrico. As mesmas análises bioquímicas citadas anteriormente foram realizadas. Ainda nesta etapa, para averiguação das perdas nutricionais e químicas dos
cogumelos, 10mL da água utilizada na fervura e na imersão das amostras controle foram
guardadas para os testes bioquímicos envolvendo quantificação de proteínas e fenóis e
avaliação da atividade das enzimas peroxidase (POD) e polifenoloxidase (PPO).
Para análise dos pedaços dos cogumelos, um grama foi triturado em 40mL de água
destilada, mantido em geladeira por 1 hora sendo posteriormente submetido à filtração e
mantido em freezer -20ºC, até a realização dos testes enzimáticos, quantificação de proteínas
e fenóis.
Para a realização dos testes enzimáticos, foram realizadas diluições em tampão
fosfato, segundo Srivastava (1987), Southerton e Deverall (1990).
A figura 6 esquematiza os procedimentos empregados nesta etapa.
Figura 6: Etapas empregadas na padronização do método de lavagem do cogumelo Pleurotus ostreatus.

43
3.3 PADRONIZAÇÃO DO TEMPO DE FERVURA – COGUMELO DE
TAMANHO GRANDE (SEGUNDA ETAPA)
Cogumelo P. ostreatus de tamanho grande foi separado em 3 grupos de amostras de
20 gramas cada e lavados em água corrente por 1 minuto. A amostra controle foi armazenada
por 4 minutos sob refrigeração (em geladeira). Uma amostra foi submetida à fervura por 2
minutos e a outra fervida por 4 minutos. Durante todos esses processos, foi utilizada água
destilada estéril contendo 0,25% de ácido cítrico.
Ao final do processo de fervura, ambas as amostras foram imediatamente resfriadas
em água gelada.
Posteriormente, um grama de cogumelo foi triturado em 40mL de água destilada,
mantido em geladeira por 1 hora sendo em seguida submetido à filtração e mantido em freezer
-20ºC. Foi então submetido aos testes bioquímicos (quantificação de proteínas, fenóis e
atividade enzimática da POD e da PPO).
Para a realização dos testes enzimáticos, foram realizadas diluições em tampão
fosfato, segundo Srivastava (1987), Southerton e Deverall (1990).
A figura 7 esquematiza os procedimentos empregados nesta etapa.
Figura 7: Etapas empregadas na padronização do tempo de fervura de cogumelo Pleurotus ostreatus.

44
3.4 PROCESSAMENTO DO COGUMELO Pleurotus ostreatus
(TERCEIRA ETAPA)
Cogumelos de tamanho grande e pequeno foram separados em 6 grupos contendo 50g
cada um e em seguida lavados em água corrente por 1 minuto. Todas as análises foram
realizadas em duplicatas com 3 repetições
Os grupos foram categorizados da seguinte maneira 1) controle (in natura); 2) pré-
tratado; 3) pré-tratado e armazenado por 3 dias; 4) pré-tratado e armazenado por 7 dias; 5)
pré-tratado e armazenado por 14 dias; 6) pré-tratado e armazenado por 28 dias.
Amostras de cogumelos in natura depois de lavadas em água corrente foram
submetidas aos testes bioquímicos para quantificação de proteínas, fenóis, vitamina C, alfa e
beta glucanas, para avaliação da atividade enzimática da POD e da PPO além de serem
analisadas sensorial e microbiologicamente. Estas foram consideradas as amostras controle.
O grupo 2 de amostras, identificado como pré-tratado, referiu-se a cogumelos fervidos
por 4 minutos em solução de água destilada e ácido cítrico (0,25%) e resfriados
imediatamente em água gelada por 2 minutos. As mesmas análises bioquímicas, sensoriais e
microbiológicas realizadas nas amostras controle foram também realizadas nesse grupo.
As amostras do grupo 3, 4, 5 e 6 passaram pelo mesmo pré-tratamento descrito acima
e em seguida foram acondicionadas em sacos plásticos transparentes. Metade das amostras
foram fechadas hermeticamente sem vácuo e a outra metade selada a vácuo, através de um
equipamento industrial.
Todas as amostras embaladas com e sem vácuo foram mantidas em geladeira por um
período pré-determinado correspondente a 3, 7, 14 e 28 dias. Transcorridos os tempos de
armazenamento, os sacos foram abertos e os cogumelos foram submetidos à pesagem do peso
fresco e a avaliação do peso seco. Testes bioquímicos, análises microbiológicas e sensoriais
também foram efetuadas, assim como realizadas nas amostras do grupo 1 e 2.
A figura 8 esquematiza os procedimentos empregados nesta etapa.

45
Figura 8: Etapas empregadas no processamento do cogumelo Pleurotus ostreatus.
3.5 TEOR DE UMIDADE E PERDA DE ÁGUA DO COGUMELO
Pleurotus ostreatus
O teor de umidade das amostras foi determinado em estufa a vácuo, a 60ºC, por 24
horas, com a amostra em triplicata, até peso constante (AOAC, 1990). Os frascos de vidros
foram mantidos na estufa por 24 horas, colocados em dessecador e pesados. Após a pesagem,
foi transferido 1g de cogumelo fresco no frasco e mantido por 24 horas, sendo depois passado
para dessecador e após frio, pesado, voltando por mais 15 minutos para a estufa, repetindo a
pesagem até peso constante.
Os cogumelos retirados dos sacos plásticos, após os tempos de armazenamento,
foram submetidos à pesagem total para comparação com o grupo controle em relação à perda
de água.

46
3.6 ANÁLISES BIOQUÍMICAS
3.6.1 Proteínas e fenóis Para a quantificação de proteínas foi utilizado o método de Lowry, em equivalentes de
SAB/mL (SORO ALBUMINA BOVINA) (LOWRY, 1951). O total de fenóis foi analisado
através do método de Folin-Ciocalteau, sendo os resultados expressos em mg de equivalentes
de ácido clorogênico por mL (mg Eq. ácido. clorogênico/mL) (SWAIN; HILLIS, 1959).
3.6.2 Atividade enzimática da polifenoloxidase (PPO) A atividade da polifenoloxidase, tirosinase, catecolase foi determinada medindo-se a
variação de absorbância em 495 nm da ortoquinona formada na reação enzimática. Nessa
determinação foram usados 2mL do extrato em 1mL da solução de catecol 0,01 mol L-1 em
tampão fosfato 0,1 mol L-1 (pH 6,5), a 37°C. Uma unidade de atividade (unidades/mL) foi
definida como a quantidade de enzima que causa o aumento de 0,001 unidade de absorbância
por minuto nas condições citadas segundo Srivastava (1987). O potencial antioxidante em 5
minutos de reação ou perda de atividade da enzima entre os tratamentos foi transformada em
porcentagem para os tratamentos posteriores.
3.6.3 Atividade enzimática da peroxidase (POD) A atividade da peroxidase foi determinada medindo-se a variação de absorbância do
tetraguaiacol formado na reação enzimática em espectrofotômetro computadorizado Beckman
DU 650 com comprimento de onda de 470nm. Nessa determinação foram usados 0,05mL do
extrato em 2,95mL da solução de guaiacol 0,9 µmol L-1 e 0,36 µmol L-1 da solução de
peróxido de hidrogênio em tampão fosfato 0,1 mol L-1 (pH 7,0), a 37°C segundo
Moerschbacher et al. (1986). A absorbância foi lida em 5 minutos e uma unidade de atividade
foi expressa como aumento na absorbância de 0,001/min/mL (SOUTHERTON;
DEVERALL,1990).

47
3.6.4 Quantificação da Vitamina C O teor de vitamina C foi avaliado segundo técnica recomendada pelo Instituto Adolfo
Lutz (2005) para alimentos in natura, por titulação com solução de iodeto de potássio até a
coloração azul.
3.6.5 Beta e Alfa-glucana A quantificação de beta e alfa-glucana presente no cogumelo foi realizada utilizando,
respectivamente, enzimas beta e alfa-glucanase (1-3), (1-4) e (1-6) e os métodos de Lever
(1972), Van Hoof et al. (1991) e McCleary et al. (1985).
3.7 AVALIAÇÃO DAS CARACTERÍSTICAS SENSORIAIS
A avaliação da qualidade sensorial foi realizada, de maneira voluntária, por uma
equipe de 7 pessoas e conduzida no laboratório de pesquisa da UNINOVE. Os participantes
foram orientados quanto ao tema e objetivos da pesquisa e assinaram o termo de
consentimento livre e esclarecido.
Os atributos julgados para análise sensorial dos cogumelos foram:
SABOR: bom, quando as características originais estavam preservadas; ruim, quando
alguma alteração no sabor foi detectada, porém ainda comestível; péssimo, quando alterações
percebidas tornam o cogumelo impróprio para o consumo em relação ao sabor desejado,
como por exemplo, azedo.
ODOR: bom, quando próprio e característico de cogumelo; ruim, quando alguma
modificação no odor característico do alimento foi detectada, porém ainda agradável;
péssimo, quando o cogumelo apresentou alterações bastante significativas, tais como odor
azedo e fermentado, tornando-o inadequado.
COR: variando de marrom escuro a amarelo claro.
TEXTURA: firme, quando o cogumelo apresentou resistência ao rompimento no
momento da mastigação; mole, quando o rompimento ocorreu facilmente.
Inicialmente, os provadores avaliaram as amostras de cogumelo in natura e pré-
tratada. Decorridos os tempos estipulados para o armazenamento as embalagens eram abertas
e imediatamente averiguados os atributos sensoriais. Suas opiniões eram registradas em fichas
padronizadas (Quadro 1).

48
Nos cogumelos onde as características textura e/ou odor e/ou cor estavam alteradas,
não era avaliado o atributo sabor.
Os resultados foram comparados com os das análises microbiológicas e bioquímicas.
Quadro 1: Ficha utilizada para a avaliação sensorial de cogumelos Pleurotus ostreatus.
3.8 pH DO COGUMELO E DOS LÍQUIDOS EXSUDADOS DO
ALIMENTO
Todos os sacos plásticos contendo cogumelos tratados foram abertos, decorrido o
prazo de armazenamento determinado, para avaliação do pH do cogumelo e do líquido
exsudado medido através da fita de pH universal (Merck).
3.9 ANÁLISES MICROBIOLÓGICAS
As análises microbiológicas foram realizadas tanto nos líquidos exsudados dos
cogumelos quanto do próprio P. ostreatus seguindo as metodologias e técnicas descritas por
Silva e Junqueira (1995, 1997) e Vanderzant e Splittstoesser (1996).
Deguste a amostra de cogumelo e expresse a sua opinião para cada atributo solicitado (apenas 1 resposta).
1- SABOR ( ) bom ( ) ruim ( ) péssimo
bom: características originais estavam preservadas; ruim: alguma alteração no sabor foi detectada, péssimo: alterações mais acentuadas, como por exemplo, azedo
2- ODOR ( ) bom ( ) ruim ( ) péssimo
bom: próprio e característico de cogumelo; ruim: alguma modificação no odor característico do alimento, porém ainda agradável; péssimo: alterações bastante intensas, tais como odor azedo, pútrido ou fermentado.
3- COR ( ) marrom escuro ( ) marrom claro ( ) marrom esverdeado
( ) amarelo escuro ( ) amarelo claro
4- TEXTURA ( ) firme ( ) mole
Firme: cogumelo apresenta resistência ao rompimento no momento da mastigação; mole: o rompimento ocorre facilmente.

49
De cada um dos tratamentos foi retirado 0,2mL do líquido exsudado e dividido em
dois tubos. Um deles continha caldo lactosado e o outro meio líquido de batata-dextrose.
Após 24 horas foi transferido 0,1mL para lâmina e observado em microscópio. Como não foi
verificada a presença de bactérias, a diluição foi plaqueada utilizando o meio de batata-ágar-
dextrose (BAD), mantendo a temperatura de 27ºC durante 48 horas e verificada a presença ou
não de colônias de bactérias ou leveduras. Para a análise da comunidade microbiana das amostras, 0,25g de cogumelos foram
transferidos para tubos contendo 2,25mL de água peptonada e homogeneizados com bastão de
vidro. Posteriormente, foram preparadas as diluições seriadas em tubos estéreis. Para averiguar a presença de leveduras, os tubos estéreis continham 4,5mL de água
peptonada até a diluição 10-4. Em seguida, cada diluição foi vertida em placas de Petri
juntamente com 9mL de meio batata-ágar-dextrose fundido e acidificado com ácido tartárico
a 10%, utilizando-se a técnica em profundidade. As placas foram incubadas a uma
temperatura de 25ºC durante 5 dias. Após esse tempo foi feita a leitura das placas sendo que
quando o crescimento das colônias era compatível com leveduras realizou-se a contagem do
número de Unidades Formadoras de Colônias (UFC) (BRASIL, 2001).
No caso de bactérias aeróbias mesófilas (micro-organismos que crescem em aerobiose
e em temperatura de incubação entre 15 e 40oC e uma temperatura média de 35oC) foram
realizadas as diluições seriadas em tubos com água peptonada até a diluição de 10-4, sendo
posteriormente transferidas para placas contendo 15mL de ágar-água (Difco) e mantidas
incubadas a 35-37°C por 48 horas. Foram consideradas para contagem somente as placas da
mesma diluição que apresentaram de 30 a 300 colônias, multiplicada a sua média aritmética
pelo respectivo fator de diluição e o resultado expressado em Unidades Formadoras de
Colônias/g de amostra (UFC/g) (SILVA; JUNQUEIRA, 1995, 1997; VANDERZANT;
SPLITTSTOESSER, 1996).
Na pesquisa por Salmonella spp. a amostra contida na solução salina peptonada foi
incubada a 37°C por 24 horas. Cada amostra foi semeada em placas de Petri com Ágar Verde
Brilhante (DIFCO) e incubada por 24 horas a 37°C. As colônias típicas obtidas nas placas
foram observadas. Foi realizado também plaqueamento em meio XLD (xilose-lisina
desoxicolato), meio Rappaport Vassiliadis e passado para caldo Verde brilhante.
O quadro 2 descreve os meios de cultura utilizados e suas respectivas aplicações.

50
Quadro 2: Descrição dos meios de cultura e suas aplicações.
Meio Aplicações Simples- água peptonada Meio básico para enriquecimento de bactérias ou
leveduras.
Seletivos ou enriquecidos
1-Caldo verde-brilhante ou
Ágar Verde brilhante
Favorece Bacilos Gram negativos da família
Enterobacteriaceae, podendo ter Salmonellas.
2-Batata-agar-dextrose (BAD) Desenvolvimento de fungos e leveduras
3-XLD Plaqueamento diferencial onde Salmonellas apresentam
colônias cor de rosa escuro. Cepas fortemente produtoras
de gás sulfídrico podem produzir colônias com centro preto
grande e brilhante.
4-Rappaport Enriquecimento de Salmonellas
5-Meio de ágar-água Desenvolvimento de bactérias aeróbias mesófilas
3.10 ANÁLISE ESTATÍSTICA
Para análise dos resultados foi realizado teste T (Student´s) e teste de variância
(ANOVA), utilizando-se o software ORIGIN. O nível de significância foi 5% (p<0,05) para
todos os testes realizados.

51
4 RESULTADOS E DISCUSSÃO
O Pleurotus ostreatus, comparado com outros cogumelos comestíveis, requer uma
tecnologia de produção relativamente simples e apresenta um ciclo produtivo reduzido,
características consideradas desejáveis e determinantes na viabilidade técnica e econômica de
um cultivo industrial (CHANG; MILES, 2004; DABA et al., 2008).
Segundo o Dr. Edison de Souza, proprietário da empresa Brasmicel, principal
produtora de Pleurotus spp. da região de Suzano, ultimamente os consumidores têm preferido
adquirir o cogumelo P. ostreatus fresco de tamanho pequeno (Figura 9) devido ao sabor e
aparência mais agradáveis. Entretanto, um dos inconvenientes e limitantes na produção deste
cogumelo é a perda do frescor e das características originais em um curto espaço de tempo o
que determina, então, a sua comercialização em até 5 dias após a colheita (APATI, 2004).
Figura 9: Cogumelo Pleurotus ostreatus de tamanho pequeno, preferencialmente comercializado.
O maior problema existente após a colheita dos cogumelos é a falta de tecnologia para
evitar ou minimizar o escurecimento enzimático causado pela ação indesejada das enzimas
polifenoloxidase e peroxidase, o que prejudica a comercialização do produto, reduzindo seu
preço e muitas vezes o tornando inaceitável pelo consumidor.
As enzimas tirosinase e catecolase são distinguidas por sua atividade em substratos
específicos e são responsáveis pelo processo de escurecimento enzimático nos cogumelos
(GAMACHE et al., 1993; KING; YOUNG, 1999; OWUSU-APENTEN, 2004; XIE et al.,
2007). O ácido cítrico atua bloqueando a atividade da tirosinase, quelando os íons de cobres

52
constituintes da enzima ou reduzindo sua ação enzimática pela diminuição do pH do meio
(MODA et al., 2005).
Assim, no presente trabalho, no momento da colheita, os basidiocarpos apresentaram-
se brancos amarelados, escurecendo ligeiramente durante o transporte para o laboratório,
apesar da manutenção e controle da temperatura através do gelo.
Como a cor é um fator importante para valorizar a qualidade de um alimento, segundo
Moda et al., (2005) para contornar o problema do escurecimento dos cogumelos depois da
colheita, os produtores rurais têm utilizado algumas substâncias antioxidantes, sendo o ácido
ascórbico e ácido cítrico os mais comumente empregados, principalmente para o champignon.
Por isso que nessa pesquisa, antes dos tratamentos, os cogumelos foram submetidos à
fervura com água destilada e ácido cítrico para promover a inativação, principalmente da
enzima polifenoloxidase, favorecendo a permanência da cor clara nos basidiocarpos.
4.1 COLETA DOS COGUMELOS Pleurotus ostreatus
O cogumelo P. ostreatus de tamanho grande foi colhido antes dos bordos iniciarem o
enrolamento (Figura 10B) momento este considerado adequado, pois em seguida, ocorre a
liberação de esporos que cairão sobre os cogumelos interferindo em suas aparências,
favorecendo o processo de envelhecimento e diminuindo seu valor comercial (BONONI et al.,
1999; EIRA, 2004).
Para os testes iniciais foram utilizados cogumelos nos estágios de desenvolvimento
pequeno, médio e grande (Figura 10A e 10B).
Figura 10 A: Diferentes estágios de desenvolvimento do Pleurotus ostreatus. Tamanho pequeno e médio. B: Pleurotus ostreatus de tamanho grande. Aspectos gerais no momento da colheita.
B A médio
pequeno

53
4.2 PADRONIZAÇÃO DO MÉTODO DE TRATAMENTO (PRIMEIRA
ETAPA)
Comercialmente, os cogumelos são lavados em água corrente antes do
acondicionamento nas embalagens para a venda. Por isso, neste estudo, os cogumelos P.
ostreatus foram imersos em água destilada estéril para verificar a perda de proteínas (expressa
em mg SAB/mL), fenóis (equivalente em mg do ácido clorogênico) e enzimas
polifenoloxidase (PPO) e peroxidase (POD) nos diferentes métodos de lavagem (Figura 11).
Figura 11: Teores de proteína (A) e fenol (B) liberados para os líquidos de lavagem. Comparação entre as amostras controle e pré-tratadas de Pleurotus ostreatus de tamanho pequeno, médio e grande.
A
B

54
Na Figura 11 é possível observar que todas as amostras in natura tiveram maior perda
de proteína para a água, quando comparadas com as pré-tratadas. No cogumelo de tamanho
grande, em ambos os métodos (in natura e pré-tratado), a perda percentual de proteína foi
significativamente maior do que nos de tamanhos médios e pequenos.
A liberação de fenol foi mais alta nas amostras fervidas (pré-tratadas), para todos os
tamanhos de cogumelos, com diferenças significativas entre elas. O ácido clorogênico, um
dos compostos fenólicos existentes nos alimentos, é um dos principais responsáveis pelo
escurecimento enzimático do cogumelo e sua inativação depende do tipo de tratamento no
qual o alimento é submetido (ARAÚJO, 2008).
Em relação às enzimas PPO e POD estas foram liberadas tanto na água das amostras
controle quanto nas fervidas, sendo que dentre estas últimas, o cogumelo de tamanho grande
apresentou maiores perdas (Figura 12).
Contudo, nota-se que o processo de fervura não foi capaz de inativar totalmente estas
enzimas visto que o tempo e a temperatura necessários para a completa inativação são
elevados e variam consideravelmente entre os produtos alimentícios o que poderia então,
comprometer a qualidade nutricional e sensorial do cogumelo. Além do mais, são enzimas
consideradas termoestáveis, especialmente a POD, que também apresenta grande capacidade
de regeneração (MARSHALL; KIM; WEI, 2000; SCHWEIGGERT; SCHIEBER; CARLE,
2005; BERBICZ; CLEMENTE, 2001).
Essas enzimas estão associadas com reações de escurecimento enzimático e não
enzimático, as quais são dependentes de substratos como fenóis e proteínas para a formação
da melanina, um pigmento escuro insolúvel (VÁMOS-VIGYÁZO, 1981; ARAÚJO, 2008).
Por isso que, uma maior atividade das enzimas PPO e POD está correlacionada com a
maior perda de fenóis e proteínas uma vez que estes compostos são utilizados como substrato
nas reações de oxidação.
Portanto, a liberação da PPO e da POD para a água de lavagem poderá minimizar a
ocorrência de reações oxidativas no P. ostreatus, evitando assim o escurecimento do alimento
e as perdas de proteína, preservando então a cor e a qualidade nutricional.

55
Figura 12: Enzimas polifenoloxidase (A) e peroxidase (B) liberadas para os líquidos de lavagem. Comparação entre as amostras controle e pré-tratadas de Pleurotus ostreatus de tamanho pequeno, médio e grande.
Em relação aos teores de proteína e fenol quantificados nos pedaços de cogumelos a
Figura 13 demonstra que todas as amostras submetidas à fervura por 4 minutos apresentaram
maiores quantidades destes componentes no alimento quando comparadas com as amostras
controle (somente mantidas em água).
A
B
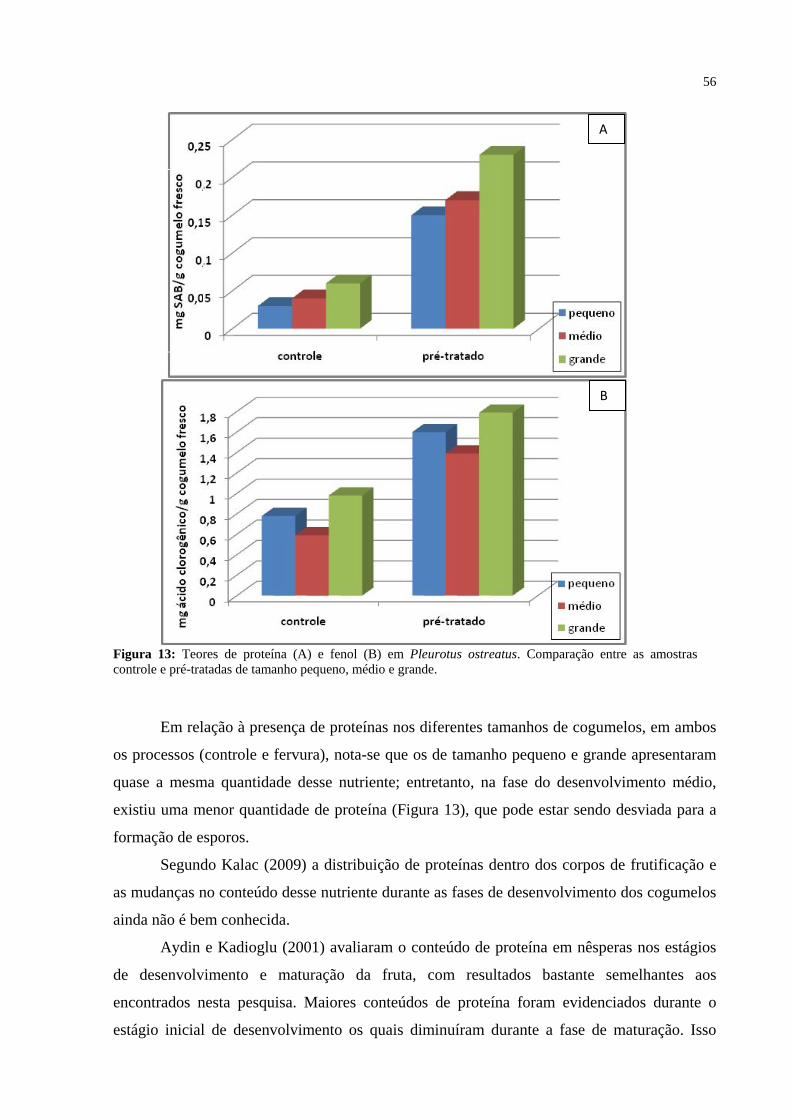
56
Figura 13: Teores de proteína (A) e fenol (B) em Pleurotus ostreatus. Comparação entre as amostras controle e pré-tratadas de tamanho pequeno, médio e grande.
Em relação à presença de proteínas nos diferentes tamanhos de cogumelos, em ambos
os processos (controle e fervura), nota-se que os de tamanho pequeno e grande apresentaram
quase a mesma quantidade desse nutriente; entretanto, na fase do desenvolvimento médio,
existiu uma menor quantidade de proteína (Figura 13), que pode estar sendo desviada para a
formação de esporos.
Segundo Kalac (2009) a distribuição de proteínas dentro dos corpos de frutificação e
as mudanças no conteúdo desse nutriente durante as fases de desenvolvimento dos cogumelos
ainda não é bem conhecida.
Aydin e Kadioglu (2001) avaliaram o conteúdo de proteína em nêsperas nos estágios
de desenvolvimento e maturação da fruta, com resultados bastante semelhantes aos
encontrados nesta pesquisa. Maiores conteúdos de proteína foram evidenciados durante o
estágio inicial de desenvolvimento os quais diminuíram durante a fase de maturação. Isso
A
B

57
provavelmente pode ser explicado por aumento na síntese de enzimas envolvidas na
maturação da fruta.
Estudos demonstraram que o estágio de maturação dos cogumelos, a espécie, suas
estruturas, os substratos onde são cultivados e os nutrientes disponíveis para crescimento
estão diretamente relacionados com a quantidade de nutrientes, dentre eles a proteína,
existentes e disponíveis no alimento (BARROS et al., 2007; CHEUNG, 1996; SILVA et al.,
2007).
Assim, a imersão dos cogumelos em água sem a aplicação do método de fervura, não
foi capaz de manter as proteínas do alimento intactas, podendo estas terem sofrido
desnaturação pela influência do fenol e pelas atividades enzimáticas presentes na constituição
do P. ostreatus.
Os cogumelos submetidos à fervura apresentaram na sua constituição bioquímica
menor atividade das enzimas PPO e POD quando comparados com as amostras controle
(Figura 14). Tal fato pode ser explicado pela inativação destas enzimas quando submetidas a
tratamento térmico, conforme já esclarecido anteriormente.
Nas amostras pré-tratadas, a PPO aumentou sua atividade durante os estágios de
desenvolvimento, contudo a POD apresentou maior atividade na amostra de tamanho médio.
Nas amostras controle, a PPO teve o mesmo comportamento que nas submetidas à
fervura e a atividade da enzima peroxidase foi maior nos estágios inicial e final de
desenvolvimento do cogumelo.
A atividade dessas mesmas enzimas estudadas em nêsperas apresentou um
comportamento semelhante aos encontrados nesta pesquisa sendo mais alta no início do
estágio de desenvolvimento e no final do processo de maturação, demonstrando que
provavelmente intensas atividades metabólicas ocorrem durante esses estágios (AYDIN;
KADIOGLU, 2001).
Nessa primeira fase, observou-se que o método de fervura auxiliou na manutenção das
propriedades do cogumelo, principalmente na liberação de enzimas oxiredutases,
minimizando assim as reações de oxidação e escurecimento dos cogumelos.

58
Figura 14: Atividade das enzimas polifenoloxidase (A) e peroxidase (B) em Pleurotus ostreatus. Comparação entre as amostras controle e pré-tratadas de tamanho pequeno, médio e grande.
4.3 PADRONIZAÇÃO DO TEMPO DE FERVURA – COGUMELO DE
TAMANHO GRANDE (SEGUNDA ETAPA)
Esta etapa teve como finalidade padronizar o tempo de fervura dos cogumelos uma
vez que, baseado nos resultados apresentados no item anterior, este foi o método escolhido
como padrão.
Conforme demonstrado na Figura 15, na amostra controle houve retenção da enzima
polifenoloxidase (PPO), de proteínas e de um pouco de fenol, entretanto após 2 minutos de
fervura, a quantidade de proteína preservada no alimento foi maior do que na amostra controle
podendo esta ser degradada pela PPO. No tempo de 4 minutos a proteína liberada foi menor
A
B

59
em relação à de 2 minutos e a atividade PPO foi inibida em 80%, não comprometendo assim a
qualidade do cogumelo.
Figura 15: Teores de proteína (mg SAB/g cogumelo fresco) e fenol (mg ác. clorogênico/g cogumelo fresco) e atividade enzimática da polifenoloxidase (Unidade PPO/minuto/g cogumelo fresco) em Pleurotus ostreatus. Comparação entre as amostras controle e as fervidas por 2 e 4 minutos.
Comparando a amostra controle com a submetida à fervura por 4 minutos, a maior
diferença está na atividade da PPO (menor após a fervura) diminuição esta favorável que pode
auxiliar na manutenção não só da cor como também na qualidade nutricional do alimento
(Figura 15).
Segundo Vámos-Vigyázo (1981) e Rios, Abreu e Corrêa (2002) as enzimas peroxidase
e polifenoloxidase e a presença de compostos fenólicos no cogumelo são os responsáveis pelo
escurecimento enzimático. A enzima PPO, responsável pela alteração da cor do alimento
quando ocorre o rompimento celular, na presença do oxigênio, utiliza como substrato as
quinonas, que são transformadas em substâncias escuras (melaninas), acarretando a redução
da qualidade sensorial. Esse é um dos principais problemas na industrialização de produtos
alimentícios (VIGYÁZO-VÁMOS, 1981; WATERMAN et al., 1994; MARSHALL; KIM;
WEI, 2000; FREITAS et al., 2008; ARAÚJO, 2008).
O branqueamento e o emprego de substâncias antioxidantes como o ácido cítrico
podem minimizar tais reações enzimáticas. Os antioxidantes auxiliam na prevenção do
escurecimento enzimático por inibição direta sobre as enzimas oxiredutases, atuando como
um redutor da quinona formada ou impedindo a participação de intermediários formados
durante a ação enzimática na oxidação e condensação adicionais (ARAÚJO, 2008).

60
A utilização do ácido cítrico como um inibidor das reações de oxidação apresenta
vantagens também para outros alimentos, como a guariroba, que apresentou redução em 8,5%
na atividade da PPO quando imersa em solução de ácido cítrico entre 10 e 20 minutos
(CARNEIRO et al., 2003).
Manzi et al. (2001) avaliaram a composição de proteínas em cogumelos P. ostreatus
que foram grelhados (calor seco) por 10 minutos, sem a adição de outras substâncias, e
concluíram que o processamento interfere significativamente na composição nutricional,
aumentando os teores de proteína de 1,61 para 2,53g/100g de cogumelo fresco.
O calor seco promove uma diminuição no conteúdo de água constituinte do cogumelo,
através do processo de evaporação, o que consequentemente concentra os nutrientes. Do
contrário, a diminuição na quantidade de nutrientes pode ser explicada pelas interações entre
diferentes componentes, reações químicas e/ou degradações térmicas (Manzi et al., 2004).
Diante do descrito, os resultados obtidos nos experimentos mostraram que em 4
minutos a enzima PPO foi inibida em aproximadamente 80%, embora não totalmente
degradada, podendo ainda estar ativa no alimento caso não seja bem acondicionado.
Entretanto, a proteína foi preservada e não liberada totalmente para a água de fervura
demonstrando que o procedimento tem validade na sua aplicação.
4.4 PROCESSAMENTO DO COGUMELO Pleurotus ostreatus
(TERCEIRA ETAPA)
Os cogumelos em geral são descritos como produtos muito perecíveis, com tempo de
vida útil limitado quando conservados à temperatura ambiente (MAU et al., 1993). Durante o
armazenamento podem ocorrer vários processos de alteração de comportamento que se
manifestam na constituição bioquímica, interferindo no sabor, odor, textura e aparência do
alimento (SAPATA et al., 2005). Baseando-se nos resultados demonstrados nas etapas 1 e 2,
adotou-se o método de lavagem em água corrente e o tratamento sob fervura com água e
ácido cítrico 0,25% durante 4 minutos como o mais apropriado para as experimentações com
o cogumelo P. ostreatus. Com o objetivo de diminuir o oxigênio dentro das embalagens plásticas, minimizando
assim as reações de oxidação, metade das amostras foram fechadas a vácuo. As do grupo
controle foram apenas seladas, sem modificação da atmosfera interna da embalagem. Todos

61
os sacos foram mantidos em geladeira a fim de auxiliar na manutenção das características
sensoriais, nutricionais e físico-químicas do cogumelo. Primeiramente foi realizado um teste prévio, onde foram colocados os cogumelos in
natura em sacos plásticos, vedados sem vácuo e mantidos em geladeira por períodos de 0, 3 e
7 dias. No período de 3 dias apresentaram um cheiro não agradável, o qual se agravou no 7º
dia de armazenamento. Isto veio provar que o cogumelo, quando fresco, tem que ser colhido e
vendido para ser usado como comestível no prazo máximo de 7 dias.
Quando vedados a vácuo, sem aplicação do processo de branqueamento, após 3 dias
os cogumelos apresentaram aspecto marrom escuro com bolhas de ar no interior dos sacos
plásticos, sendo que após 6 dias foram descartados.
Os resultados até aqui apresentados demonstraram que para o cogumelo in natura a
preservação em sacos plásticos, sem vácuo, não foi ideal e que sem o processo de
branqueamento, reações químicas, tais como respiração e fermentação, ainda ocorrem. Quando os cogumelos in natura passaram pelo pré-tratamento (branqueamento) e
foram acondicionados em sacos plásticos fechados com vácuo, foi observado que os corpos
de frutificação apresentaram-se com cor mais clara (de amarelado a esbranquiçado) e sem
alterações no cheiro. Quanto ao aspecto visual e a textura, os cogumelos encontravam-se
firmes, com lamelas bem visíveis, sem sinais de deterioração. Assim, foi montado o esquema
de tratamento prévio e acondicionamento em sacos com vácuo e sem vácuo por período de
tempo variável.
4.4.1 Teor de umidade e perda de água do cogumelo Pleurotus ostreatus A umidade de algumas amostras foi determinada por secagem em estufa a 105ºC,
entretanto, nessa temperatura os cogumelos não somente se desidratavam por completo, como
também se apresentaram queimados. Sendo assim, passou-se a usar a temperatura de 60ºC
para avaliar o peso seco, temperatura esta usada pelos produtores na secagem de cogumelos e
também citada na AOAC (1990).
A relação entre peso fresco e peso seco traduz a perda de água do cogumelo, sendo
que os in natura de ambos os tamanhos apresentaram menor perda de água do que os pré-
tratados, diferenças estas significativas (Figura 16).
Nos de tamanho pequeno, a perda de água nos embalados sem vácuo foi próxima a dos
empacotados a vácuo (Figura 16), mesmo que em porcentagem isso seria desprezível.

62
Figura 16: Relação peso fresco/peso seco (g) e perda de água (%) do cogumelo Pleurotus. ostreatus de tamanhos grande (A) e pequeno (B). Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: a: cogumelos embalados a vácuo; b: cogumelos embalados sem vácuo.
Em relação aos cogumelos de tamanho grande, embalados com e sem vácuo, o
percentual de perda de água foi muito próximo entre os tratamentos. Esses resultados devem
estar atrelados à estrutura das glucanas, que por estarem mais abertas no tamanho grande
apresentam maior quantidade de água que é retirada no vácuo; no cogumelo pequeno foi mais
difícil retirar a água, pois as estruturas provavelmente estavam mais fechadas (Figura 16).
O teor médio de umidade dos P. ostreatus estudados nesta pesquisa foi de 89% para os
de tamanho pequeno e de 90% para os grandes. Estes resultados estão dentro da média
encontrada nos estudos com Pleutorus spp. realizados por Sturion; Ranzani (2000), Manzi et
al. (1999) e Yang et al. (2001) que foi de 85,24 a 94,70%, o que demonstra ser o P. ostreatus
um alimento favorável ao crescimento microbiano e de fácil deterioração.
A
B

63
Mesmo tendo sido desenvolvido em outro substrato, os resultados obtidos no presente
trabalho estão de acordo com os encontrados por Manzi et al., (1999) que obtiveram um valor
médio de 90,7% no teor de água em cogumelos deste mesmo gênero cultivados em palha de
trigo. Quando utilizada a palha de bananeira, P. ostreatus apresentaram um teor de umidade
igual a 87,8%, demonstrando que o tipo de substrato utilizado no cultivo dos cogumelos pode
também interferir na umidade (BONATTI et al., 2003).
Entre as espécies L. edodes e Agaricus spp o conteúdo de água não apresenta
diferenças significativas, ficando entre 91,6%, no e 92,3%, respectivamente (MATTILA et
al., 2001).
Após o processo branqueamento (pré-tratamento), os cogumelos de tamanho pequeno
perderam em média 4,73% de água, quando comparado com o alimento cru; os de tamanho
grande reduziram cerca de 24,9% em relação ao seu peso inicial. Estes dados demonstram que
o processo de cozimento dos cogumelos interfere diretamente no teor de água constituinte do
alimento.
Esses resultados são superiores aos encontrados por Manzi et al. (2001), que
compararam o teor de água constituinte do cogumelo P. ostreatus cru e após a cocção através
do calor seco por 10 minutos e obtiveram o menor valor nos cogumelos cozidos, redução esta
de cerca de 3,5% em relação ao alimento cru. Nos diferentes gêneros estudados, a menor
perda foi de 2,05% no Agaricus em conserva e a maior foi no Boletus spp. desidratado,
3,97%.
Conforme observado na Figura 17, cogumelos grandes e pequenos embalados a vácuo
por 14 e 28 dias respectivamente, apresentaram maiores perdas de água decorridos os tempos
de armazenamento. O contrário ocorreu com os cogumelos, de ambos tamanhos, armazenados
sem vácuo por apenas 3 dias. A perda média de água, após o 28º dia de armazenamento, foi
de 9,93% para os cogumelos de tamanho pequeno e de 11,34% para os grandes, reduções
estas consideradas estatisticamente significativas.

64
Figura 17: Perda de água (%) do Pleurotus ostreatus após os tempos de armazenamento com relação ao peso pós-tratamento. Comparação entre os cogumelos de tamanhos pequeno e grande embalados com e sem vácuo. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
A água constituinte dos alimentos encontra-se dentro de células e está fortemente
ligada a proteínas. Aproximadamente 25% de toda água encontra-se no estado livre,
fracamente ligada a outras moléculas e esta água pode exsudar do alimento quando submetido
à pressão (como no processo de empacotamento a vácuo) ou outro processo tecnológico que
altere sua constituição química (MATHIAS, 2008).
Por apresentar elevado teor de umidade, o cogumelo pode facilmente exsudar líquidos,
quando submetidos ao processo de fechamento a vácuo.
Por isso, estes resultados indicaram que, em relação à perda de água, o processo de
embalar o P. ostreatus a vácuo não é satisfatório, uma vez que ocasionou maiores perdas no
conteúdo de água no decorrer dos tempos de armazenamento, em ambos os tamanhos de
cogumelos.
4.4.2 Análises bioquímicas
4.4.2.1 Polifenoloxidase (PPO)
Essa tem sido uma das enzimas responsáveis pelo processo de escurecimento
enzimático nos cogumelos, quando cortados e expostos ao ar, uma vez que a ação dela com os
compostos fenólicos naturais e o oxigênio atmosférico forma a 0-quinona. Assim, a prevenção
da ruptura dos tecidos ou a inativação da PPO pelo calor ou pela utilização de antioxidantes

65
pode reduzir o escurecimento enzimático dos cogumelos pelo retardo ou inibição da oxidação
do substrato (MARSHALL; KIM; WEI, 2000; ARAÚJO, 2008).
A enzima polifenoloxidase foi encontrada em grande atividade em cogumelos in
natura tanto no tamanho grande como pequeno. Após o tratamento pelo branqueamento a
enzima foi parcialmente inativada, mas ainda persiste, parte por apresentar alta
termoestabilidade, o que explica a perda de qualidade do produto após um período de tempo
de 3 dias quando in natura, não podendo mais ser comercializado (Figura 18).
O branqueamento foi capaz de reduzir a atividade da PPO nos cogumelos de tamanhos
pequeno e grande em cerca de 99,48% e 60%, respectivamente. No decorrer dos tempos de
armazenamento as amostras de cogumelos fechadas a vácuo e mantidas em geladeira
apresentaram menor atividade da enzima do que quando armazenadas sob refrigeração em
sacos fechados sem vácuo (Figura 18 e na Figura 19). Isso vem explicar que a presença do
oxigênio do ar acelera a reação de degradação desta enzima e que, quando fechado a vácuo
isso não ocorre com a mesma rapidez.
Esses resultados demonstraram a alta termoestabilidade da enzima perante os
tratamentos, o que também foi evidenciado por Freitas et al. (2008) em extratos enzimáticos
de uvas submetidas a tratamento térmico com temperatura de 85ºC por 10 minutos, enzima
esta que diminuiu sua atividade com o tratamento, porém não foi totalmente inibida.
Sabe-se que o escurecimento dos cogumelos causado por um longo tempo ou
inapropriadas condições de armazenamento diminui a qualidade e o valor comercial do
produto (MAU et al., 1993; MARSHALL; KIM; WEI, 2000).
No cogumelo de tamanho pequeno, a atividade se manteve constante ao longo dos
tempos de armazenamento para os cogumelos fechados a vácuo, não havendo diferença
significativa até o 14º dia. Nas embalagens sem vácuo a atividade diminui com o tempo, com
diferença estatisticamente significativa entre os tratamentos. Isso pode ser observado na
Figura 18 e na Figura 19.
O pico da atividade da PPO nos cogumelos grandes foi no 28º dia de armazenamento
em ambos os tratamentos. Para os de tamanho pequeno, nas amostras fechadas sob vácuo, o
pico foi evidenciado no 28º dia de armazenamento, enquanto que dentre as amostras sem
vácuo foi no 3º dia. Esses resultados sugerem que no armazenamento convencional dos
cogumelos, ou seja, embalados sem vácuo, as reações de oxidação ocorrem precocemente,
pois principalmente a enzima POD está mais ativa, desencadeando processos de deterioração
por contaminação microbiana e por alterações sensoriais do alimento.

66
Figura 18: Atividade da enzima polifenoloxidase (unidade de polifenoloxidase/minuto/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
Modificações na composição e na pressão da atmosfera de armazenamento dos
cogumelos podem influenciar nas reações de escurecimento. Níveis mais altos de dióxido de
carbono e baixos de oxigênio parecem aumentar esse processo, embora a atividade da
polifenoloxidase esteja praticamente inativada (ADAMS, 1991; SAPATA et al., 2005).
Figura 19: Perda (%) da atividade enzimática da polifenoloxidase em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.

67
A atividade da PPO parece estar associada com a quantidade de água disponível no
alimento, isto é, quanto maior o teor de água livre presente no cogumelo menor é a atividade
da enzima polifenoloxidase. Esta associação se deve ao fato da água ser considerada um
solvente para as reações químicas. A temperatura também pode interferir significativamente
na atividade desta enzima tendo em vista que quando acima de 40ºC a inativação é quase que
completa. Diferentemente da POD, a PPO não apresenta capacidade de regeneração e é por
isso que em cogumelos desidratados a atividade dela é inexistente (VÁMOS-VIGYÁZO,
1981; OWUSU-APENTEN, 2004).
De acordo com Lupetti et al. (2005) a prevenção da oxidação em vegetais pode ser
realizada através de procedimentos físicos ou químicos, como desidratação, armazenamento a
baixas temperaturas, tratamento térmico, retirada do oxigênio do meio e controle da
luminosidade, empregando técnicas e procedimentos adequados, como embalagens a vácuo e
utilização de antioxidantes.
Por isso que brócolos branqueados a 100ºC por 2 minutos reduziram em 18% a
atividade da PPO (LOPES; CLEMENTE, 2002). Maçãs tratadas termicamente a 75ºC por 10
minutos tiveram decréscimo quase contínuo da atividade de polifenoloxidase, chegando à
total inativação na casca da maçã (cultivar Gala) (VALDERRAMA et al., 2001).
Pineli et al. (2005) pesquisou em batata “Ágata”, minimamente processada, o efeito do
empacotamento a vácuo parcial e sob condições modificadas de atmosfera, empregando
diferentes concentrações de CO2+O2+N2. Os resultados demonstraram que o vácuo parcial
garantiu menores índices de escurecimento e uma diferença de cerca de 75% na atividade da
PPO em relação às demais amostras analisadas nos diferentes tempos.
Sendo assim, os resultados evidenciados neste presente estudo indicam que, em
relação à atividade da PPO, o processo de embalar o P. ostreatus a vácuo é satisfatório, uma
vez que ocasionou maiores perdas da atividade enzimática ao longo dos tempos de
armazenamento, em ambos os tamanhos de cogumelos.
4.4.2.2 Peroxidase (POD)
A peroxidase, como anteriormente explicado, catalisa, através do peróxido de
hidrogênio, a oxidação de substratos como mono e difenóis, polifenóis, aminofenóis, entre
outros. É conhecida como uma enzima termoestável que pode ter sua atividade regenerada
após tratamento térmico (GAMACHE et al., 1993; BRAVO, 1998; CROFT, 1998; KING;
YOUNG, 1999; CHILAKA et al., 2002; CLEMENTE, 2002).

68
O pré-tratamento do P. ostreatus promoveu redução da atividade da POD em 69,23%
nos cogumelo de tamanho pequeno e 75% nos grandes, diferença esta estatisticamente
significativa quando comparada com as amostras in natura. Em relação aos tratamentos,
observou-se que na embalagem sem vácuo a atividade foi maior do que na embalagem com
vácuo (Figura 20). Portanto, o empacotamento a vácuo poderá auxiliar na manutenção da cor
e nas qualidades nutricionais dos cogumelos embalados a vácuo.
Uma das características dessa enzima é a alta resistência à desnaturação térmica. Por
isso que os processos térmicos convencionais objetivando a sua inativação são considerados
ineficazes, uma vez que o tratamento na temperatura de desnaturação da POD levaria ao
comprometimento da textura e do sabor do produto (HEMEDA; KLEIN, 1991; FANG;
JIANG; ZHANG, 2008).
No cogumelo de tamanho grande embalado sem vácuo a atividade da POD foi menor
somente no 7º dia de tratamento com aumento significativo após 14 e 28 dias de
armazenamento. Nas amostras fechadas a vácuo, para o mesmo tamanho de cogumelo, a
atividade foi significativamente menor (Figura 20).
O percentual médio de regeneração da POD, no 28º dia de armazenamento, foi de
150% e 100% nos cogumelos pequenos embalados com e sem vácuo, respectivamente. No P.
ostreatus de tamanho grande, para o mesmo tempo de armazenamento, a regeneração foi
menor na amostra embalada a vácuo. Estes resultados demonstram que mudanças no teor de
oxigênio no interior da embalagem evitam a regeneração da POD retardando assim as reações
químicas e as alterações sensoriais dos cogumelos.
Resultados semelhantes a estes foram encontrados em suco de laranja com uma
regeneração média de 4 a 8% da peroxidase após inativação parcial por 1 minuto
(CLEMENTE, 1996).
A Figura 20 demonstra que nas amostras tratadas sem vácuo o pico da POD ocorreu
no 28º dia para os cogumelos grandes e pequenos. Quando fechados a vácuo, somente no
tamanho grande é que a mais alta atividade enzimática foi evidenciada no 28º dia de
armazenamento, diferentemente dos cogumelos pequenos, nos quais a atividade enzimática
não variou significativamente.
Em carambolas, o grau de maturação da fruta não influenciou na atividade dessa
enzima, contudo houve perda de aproximadamente 85% na atividade enzimática após 10
minutos de tratamento térmico a 85ºC (LAURENTI; CLEMENTE, 2005). Os mesmos
resultados foram encontrados por Valderrama et al. (2001) para maçãs e extratos de uvas das
variedades Rubi e Benitaka (FREITAS et al., 2008).
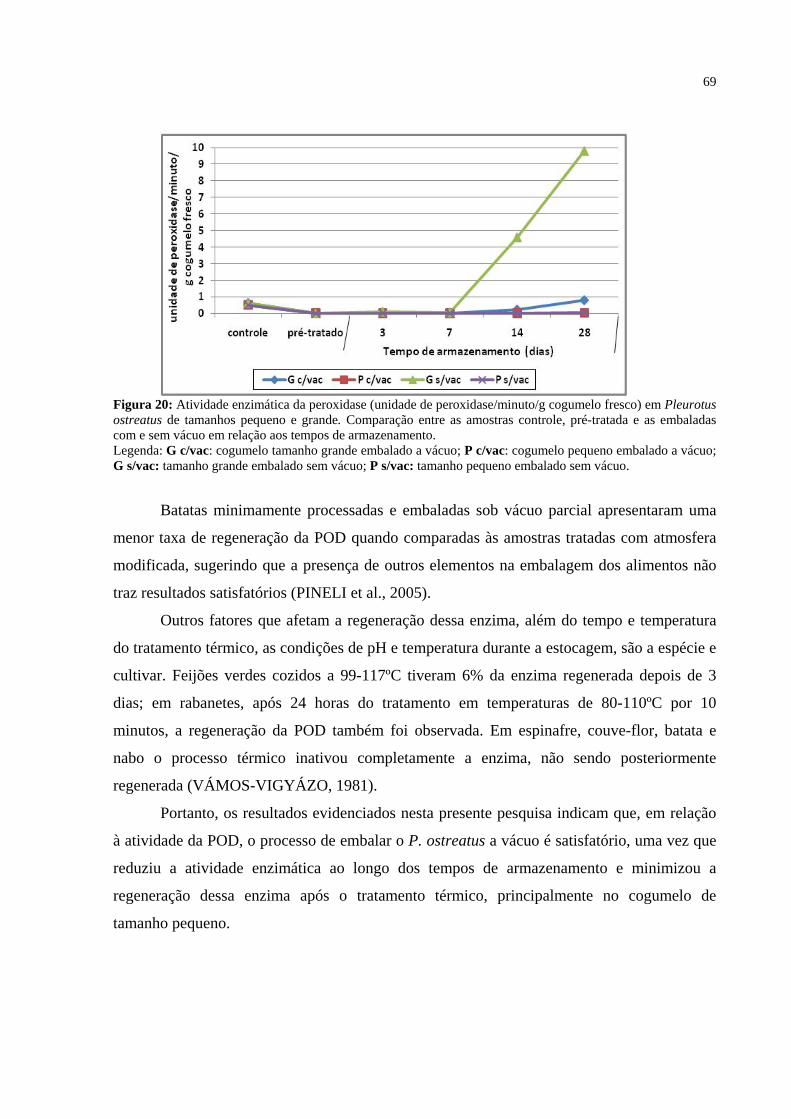
69
Figura 20: Atividade enzimática da peroxidase (unidade de peroxidase/minuto/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
Batatas minimamente processadas e embaladas sob vácuo parcial apresentaram uma
menor taxa de regeneração da POD quando comparadas às amostras tratadas com atmosfera
modificada, sugerindo que a presença de outros elementos na embalagem dos alimentos não
traz resultados satisfatórios (PINELI et al., 2005).
Outros fatores que afetam a regeneração dessa enzima, além do tempo e temperatura
do tratamento térmico, as condições de pH e temperatura durante a estocagem, são a espécie e
cultivar. Feijões verdes cozidos a 99-117ºC tiveram 6% da enzima regenerada depois de 3
dias; em rabanetes, após 24 horas do tratamento em temperaturas de 80-110ºC por 10
minutos, a regeneração da POD também foi observada. Em espinafre, couve-flor, batata e
nabo o processo térmico inativou completamente a enzima, não sendo posteriormente
regenerada (VÁMOS-VIGYÁZO, 1981).
Portanto, os resultados evidenciados nesta presente pesquisa indicam que, em relação
à atividade da POD, o processo de embalar o P. ostreatus a vácuo é satisfatório, uma vez que
reduziu a atividade enzimática ao longo dos tempos de armazenamento e minimizou a
regeneração dessa enzima após o tratamento térmico, principalmente no cogumelo de
tamanho pequeno.

70
4.4.2.3 Vitamina C
Também conhecida como ácido ascórbico na sua forma reduzida e como ácido
deidroascórbico na sua forma oxidada é uma vitamina hidrossolúvel, termosensível e
facilmente oxidada pela presença da luz. Por isso, o processamento e exposição das frutas e
hortaliças a tais condições podem resultar em perdas significativas de vitamina C no alimento
(VANNUCCHI; JORDÃO JUNIOR, 1998).
Esta vitamina desempenha inúmeras e importantes funções no organismo humano,
como a redução do ferro férrico a ferroso, propiciando aumento na absorção deste nutriente
ingerido nos alimentos, participa da síntese de colágeno pela hidroxilação de prolina, melhora
as respostas do sistema imunológico, além de ser um excelente antioxidante, podendo estar
associada à prevenção e tratamento de câncer, diminuição do risco de doenças
cardiovasculares, tratamento de hipertensão e redução da incidência de outras patologias
(GALLAGHER, 2005).
A Figura 21 demonstra que a temperatura empregada no pré-tratamento dos
cogumelos não oxidou completamente a vitamina C. No caso dos cogumelos grandes,
comparando a quantidade no alimento in natura e pré-tratado, houve um aumento no teor da
vitamina podendo ser devido a maleabilidade da estrutura e liberação da vitamina C que
estava presa no interior das células.
Figura 21: Valor de vitamina C (mg vit. C/100g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.

71
Todos os alimentos, quando submetidos a processos de cocção e armazenamento por
períodos prolongados, estão sujeitos a perdas de ácido ascórbico (CAMPOS et al., 2008). Em
um estudo realizado com espinafres frescos cozidos, a perda de vitamina C para a água de
fervura foi de até 60% provando que a o processamento térmico prolongado oxida a vitamina
(GIL et al., 1999).
Kabasakalis et al. (2000) avaliaram o conteúdo dessa vitamina em sucos de laranja
industrializados e armazenados em diferentes condições de tempo e temperatura. Perdas de
ácido ascórbico, variando entre 29-41% foram evidenciadas em sucos acondicionados em
recipientes fechados mantidos em temperatura ambiente por 4 meses. Maiores foram as
perdas (60-67%) quando os sucos foram armazenados em recipientes abertos, por 31 dias, sob
refrigeração. Em 10 dias, quando armazenados em temperatura ambiente e em refrigeração,
com a embalagem aberta, as perdas foram de 12,5 e 9%, respectivamente.
Estes resultados comprovam que o armazenamento de alimentos, como por exemplo
os cogumelos, em baixas temperaturas reduz a perda de vitamina C, mantendo assim suas
propriedades nutricionais no que diz respeito ao ácido ascórbico.
As maiores perdas de vitamina C ocorreram nos cogumelos embalados sem vácuo,
principalmente no 28º dia de armazenamento onde a redução, em comparação com a amostra
pré-tratada, foi de 44,42 e 50,29% nos de tamanho grande e pequeno respectivamente.
Quando empacotados a vácuo, os cogumelos de ambos os tamanhos não perderam ácido
ascórbico (Figura 22).
O processo de fechamento a vácuo, portanto, auxiliou na preservação da vitamina C,
pois comparando-se o teor médio encontrado nos cogumelos de tamanhos grande e pequeno,
fechados com e sem vácuo os resultados foram, respectivamente, 39,4mg, 32,8mg, 25,2mg e
18mg/100g de cogumelo fresco.
O P. ostreatus estudado apresentou teores superiores ou cerca de 50% da quantidade
de vitamina C encontrada em alimentos que são considerados fontes desta vitamina, tais como
maracujá (20mg/100g alimento cru), mexerica (22mg/100g), espinafre (5mg/100g alimento
refogado), laranja (77mg/100g de produto fresco) e couve manteiga refogada (77mg/100g)
(LIMA et al., 2006).
Sabe-se que para o adequado funcionamento do metabolismo humano é
imprescindível a presença da vitamina C em quantidades específicas conforme a idade e
condições fisiológicas. Por isso, a Ingestão Diária Recomendada (IDR) dessa vitamina para
adultos é de 45 mg. Para crianças varia entre 25 e 35 mg e para gestantes e lactentes é de 55 e
70 mg, respectivamente (BRASIL, 2005). Sendo assim, os cogumelos P. ostreatus estudados

72
neste trabalho podem ser considerados alimentos fonte e com alto teor de vitamina C, pois
atendem às recomendações da Portaria nº27 de 1998.
A composição de vitamina C nos cogumelos pode variar de acordo com o tipo de
substrato utilizado no cultivo. Rios-Hurtado et al. (2003) analisaram P. ostreatus cultivados
em diferentes compostos, encontrando para vitamina C valores que variaram de 1,19 a 194,14,
sendo os valores expressos em mg/100g de parte comestível (não foi especificado se esses
resultados são em base seca ou úmida). Quando esta mesma espécie de cogumelo foi
cultivada em substrato à base de casca de coco, o qual foi suplementado com farelo de arroz e
trigo, a quantidade da vitamina ficou entre 24-40mg de vitamina/100 g de cogumelo fresco
(PEDRA et al., 2009).
Dentre os 3 gêneros mais consumidos, o shiitake é o que apresentou maiores
quantidades de ácido ascórbico (7,19mg/100g em base úmida), seguido do shimeji
(6,5mg/100g de cogumelo fresco) e do champignon (6,3mg/100g) (FURLANI; GODOY,
2007), valores esses que superam os descritos por Mattila et al. (2001) de 2,1; 1,6;
1,45mg/100g por peso fresco, respectivamente para os gêneros acima citados.
Figura 22: Mudanças (%) no teor de vitamina C em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado avácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
Todas as amostras armazenadas sem vácuo perderam maiores quantidades de vitamina
C quando comparadas com as fechadas sob vácuo para o mesmo período de tratamento
(Figura 22), sendo estas diferenças estatisticamente significativas. Esses resultados

73
demonstraram que a quantidade de oxigênio presente na embalagem e de substâncias
presentes como fenol podem oxidar a vitamina C, degradando-a muito rapidamente.
O congelamento é um outro fator que pode reduzir a concentração de vitamina C
presente nos cogumelos o que foi provado por Jaworska, Bernas (2009) quando estudaram P.
ostreatus mantidos a temperatura de -25ºC por um período de 12.
Além disso, a perda da estabilidade da vitamina C pode também ser consequente do
rompimento celular por dano ao tecido, corte ou moedura (KLEIN, 1987; CAMPOS et al.,
2008).
Então, os resultados apresentados e discutidos em relação ao teor de vitamina C no P.
ostreatus de tamanhos pequeno e grande sugerem que o processo de embalá-los a vácuo é
satisfatório, uma vez que promoveu a retenção dessa vitamina no alimento e minimizou as
perdas ao longo dos tempos de armazenamento.
4.4.2.4 Proteína e Fenol
O teor médio de proteína encontrado no P. ostreatus de tamanhos pequeno e grande
foi de 6,8 e 4,39mgSAB/100g de cogumelo fresco, respectivamente. No decorrer dos tempos
de armazenamento, todos os cogumelos embalados com vácuo perderam menor quantidade de
proteína solúvel (média de 9,85%) se comparados com os empacotados sem vácuo (média de
43,35%) (Figura 23).
Figura 23: Teor de proteína (mg SAB/100g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.

74
Os tratamentos térmicos, extremos de pH, oxigênio do ar, substâncias oxidantes, ação
da luz e atividade de água do alimento são fatores que interferem diretamente no valor
biológico das proteínas, pois causam desnaturação e influenciam nas reações de degradação,
oxidação e decomposição (LAJOLO; TIRAPEGUI, 1998). A alta atividade da enzima
polifenoloxidase também é responsável pelo decréscimo de proteínas nos cogumelos
(TAVEIRA; NOVAES, 2007).
Os resultados encontrados nas amostras de cogumelos grandes e pequenos embalados a
vácuo, os quais não apresentaram diferenças estatisticamente significativas, podem ser
explicados pela redução do oxigênio no interior das embalagens e pela baixa atividade da
enzima polifenoloxidase.
Em 2004, Manzi et al. estudaram o efeito do calor seco na composição de proteína de
P. eryngii e observaram um aumento na quantidade desse nutriente no cogumelo grelhado
(3,1g/100g do peso fresco), em relação ao in natura (2,2g/100g do peso fresco) fato esse
explicado pela diminuição do conteúdo de água do alimento durante o processo térmico. Os
mesmos resultados já haviam sido encontrados em 2001, por Manzi et al., para P. ostreatus.
Nestas pesquisas, a determinação de proteínas foi através do método da AOAC.
Os cogumelos podem ser considerados ótimas fontes de proteína na alimentação
humana, com conteúdos similares aos encontrados nos alimentos de origem vegetal.
Entretanto, P. ostreatus não pode ser considerado um alimento fonte de proteínas de alto valor
biológico, como os de origem animal, pois são deficientes nos aminoácidos essenciais leucina
e treonina (YANG et al., 2001).
Em média, P. ostreatus apresentam 2,55% de proteína (base úmida) quantidades muito
superiores de outras espécies como o L. edodes (1,53% em base úmida) (MANZI et al., 1999).
O champignon é constituído por cerca de 28,45% de proteína (base seca) seguido do shimeji
com 22,22% e do shiitake com 18,98% (FURLANI, 2004). Resultados superiores foram
descritos por Yang et al. (2001) para o shimeji (23,9%) e para o shiitake (20,1%) e por Manzi
et al. (1999) somente para o shimeji (23,55%). Quantidades estas expressas em base seca.
Uma das razões para a variação no conteúdo de proteína nos cogumelos é o tipo de
substrato utilizado para o cultivo, o que foi provado por Bonatti et al. (2004) para P. ostreatus
e P. sajor-caju cultivados em palha de arroz (1,89 e 1,54g/100 g de massa fresca) e de banana
(2,01 e 3,10g/100 g de massa fresca).
Portanto, os resultados apresentados e discutidos em relação ao teor de proteína no P.
ostreatus de tamanhos pequeno e grande sugerem que o processo de embalá-los a vácuo é

75
satisfatório, uma vez que reduziu a perda deste nutriente ao longo dos tempos de
armazenamento.
Em relação ao conteúdo de fenol, de acordo com a Figura 24 todas as amostras de
cogumelos embalados sem vácuo apresentaram maior quantidade de fenol (média de 0,77mg
ácido clorogênico/100g cogumelo fresco), em todos os tempos de armazenamento, quando
comparadas com as armazenadas sob vácuo (média de 0,53mg/100g cogumelo fresco).
Figura 24: Teor de fenóis (mg ác. clorogênico/100g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
O pré-tratamento dos cogumelos promoveu perda estatisticamente significativa do
conteúdo de compostos fenólicos constituinte do alimento, sendo esta foi maior nos
cogumelos de tamanho pequeno (-45,74%) do que nos grande (-27,58%). Tal fato pode ser
explicado pela capacidade de solubilização dos compostos fenólicos em água (CAMPOS et
al., 2008).
Assim como esta pesquisa, um estudo recente investigou a influência de diferentes
métodos de cocção por calor úmido sobre o conteúdo de compostos fenólicos em 6 tipos de
hortaliças. Os resultados evidenciaram que o conteúdo de fenóis em repolho, couve-flor e
nabo foi afetado pela cocção em micro-ondas, com perdas de cerca de 70%. Após cocção em
água por 5 minutos houve redução entre 15-60% e após refogar (por 5 minutos) as perdas
foram de 8 a 58%, para todas as hortaliças analisadas (SULTANA; ANWAR; IQBAL, 2008).
Em geral, o cozimento por tempos menores preserva o conteúdo de compostos
fenólicos, o que foi comprovado por Ismail, Marjan e Foong (2004) num estudo realizado

76
com hortaliças folhosas cozidas por 1 minuto em água fervente onde houve perda no conteúdo
de fenóis de apenas 12% em couve, 14% em espinafre e 20% em repolho.
No presente estudo, o tempo empregado no branqueamento do P. ostreatus foi de 4
minutos, seguido do processo imediato de resfriamento, sugerindo que este tempo não foi
suficiente para reduzir completamente os compostos fenólicos existentes no alimento.
Resultados como esses foram também encontrados em sucos, néctar, purê e vinho
fabricados a partir de morangos. Os compostos fenólicos diminuíram com o avançar do tempo
e do estágio da produção, sendo que a pasteurização teve influência significativamente maior
na redução dos fenóis do que as demais etapas do processo (KLOPOTEK et al., 2005).
Em 2004, Manzi et al. publicaram um estudo cujo objetivo foi avaliar a qualidade
nutricional e os efeitos do cozimento em diferentes espécies de cogumelos. Em P. eryngii
observaram um aumento na quantidade de fenóis no cogumelo grelhado (70,4mg/100g do
peso fresco) em relação ao in natura (51,4mg/100g do peso fresco), resultados esses
contrários aos evidenciados neste estudo. Os autores explicam que o aumento da quantidade
de compostos fenólicos nos cogumelos pode ser devido à evaporação da água durante a
cocção pelo calor seco, o que concentra tais elementos.
Outro processo que preserva o teor de compostos fenólicos em alimentos e que deve
ser mencionado é o armazenamento sob refrigeração (CAMPOS et al., 2008). Todavia,
brócolos frescos, embalados em sacos de polietileno de baixa densidade e estocados por 7 dias
a 1ºC e, em seguida, por 3 dias a 15ºC tiveram perdas superiores a 70% de compostos
fenólicos (VALLEJO; TOMÁS-BARBERÁN; GARCÍA-VIGUERA, 2003).
Segundo Araújo (2008) os compostos fenólicos podem interferir na qualidade
nutricional dos alimentos, pois reduzem a biodisponibilidade de minerais e vitaminas, como o
zinco e o ácido ascórbico, além de se combinar com aminoácidos, tornando-os indisponíveis
para o organismo. Estão também associados ao escurecimento enzimáticos nos alimentos.
Por conseguinte, a diminuição no conteúdo de fenóis nos cogumelos armazenados a
vácuo se comparado com os embalados sem vácuo e com os in natura é favorável para
garantir a qualidade nutricional e sensorial do alimento.
4.4.2.5 Alfa e Beta glucana liberada
A alfa e beta glucanas são polissacarídeos que nos cogumelos desempenham função de
reserva dos carboidratos, na forma de glicogênio, e estrutural, constituindo a parede celular
(FALCÃO, 2008).

77
A Figura 25 demonstra que, no extrato aquoso das amostras de cogumelos, houve
aumento na quantidade de α-glucana ao longo dos tempos de tratamento. O valor médio desse
constituinte nos cogumelos de tamanho grande com e sem vácuo foi, respectivamente, 223,34
e 291,88mg de glicose ligação alfa/g cogumelo fresco. Valores menores foram encontrados
nos cogumelos pequenos embalados com vácuo (156,09 mg de glicose ligação alfa/g
cogumelo fresco) e sem vácuo (e 222,86mg de glicose ligação alfa/g cogumelo fresco).
Esses resultados podem estar relacionados com o grau de maturação do cogumelo,
pois o de tamanho pequeno ainda não desenvolveu completamente as substâncias e nutrientes
correspondentes a sua constituição.
Ao final do período de estocagem, verificou-se uma elevação no teor de glicose em
todas as amostras de P. ostreatus, principalmente nas empacotadas sem vácuo (Figura 26).
Tal fato indica uma possível transformação do glicogênio em glicose, por vias metabólicas ou
pela ação dos micro-organismos presentes no cogumelo.
Um estudo realizado com o intuito de quantificar a alfa glucana em cogumelos do
gênero Pleurotus evidenciou que as diferentes estruturas do fungo não interferem na
quantidade desta substância, isto é, o píleo e a haste apresentam quantidades parecidas de
α-glucana, sendo de 3,4 a 7,9% e de 3 a 7,6%, respectivamente. Valores esses expressos em
peso seco (SYNYTSYA, 2008).
Figura 25: Teor de glicose (mg de glicose ligação alfa/g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.

78
Referente à beta glucana, no extrato aquoso das amostras de cogumelos observa-se
somente a sua fração solúvel, pois a insolúvel foi descartada e não quantificada. No decorrer
dos tempos de estocagem o conteúdo desse componente diminuiu, tanto nos embalados a
vácuo quanto nos sem vácuo, em ambos os tamanhos. O pré-tratamento não interferiu na
quantidade de β-glucana do P. ostreatus (Figura 26).
O teor médio de beta glucana nos cogumelos de tamanho grande com e sem vácuo foi,
respectivamente, 35,73 e 18,31mg de glicose ligação beta/g cogumelo fresco. Valores maiores
foram encontrados nos cogumelos pequenos embalados a vácuo e sem, sendo 41,72 e
24,06mg de glicose ligação beta/g cogumelo fresco, respectivamente.
Figura 26: Teor de β-glucana (mg de beta glucana solúvel/g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
Uma das condições que interfere no conteúdo de beta glucana é a espécie do
cogumelo, sendo que P. ostreatus frescos apresentam maior quantidade desse componente
(139,24mg/100g de cogumelo fresco) quando comparado com o A. bisporus (1,4mg/100g de
cogumelo fresco). Quando grelhados (calor seco) a quantidade de beta glucana aumenta em
ambas as espécies o podem estar relacionado ao conteúdo de água eliminado do cogumelo
durante o processo de cocção (MANZI et al., 2001).
Synytsya et al. (2008) sugerem que esse aumento na quantidade de β-glucana
encontrado no estudo realizado por Manzi et al (2001 e 2004) provavelmente se deve também
à presença de substâncias inertes na fibra residual, talvez um insolúvel complexo quitina-
glucana, o qual não é totalmente degradado no processo de cocção.

79
Processos de industrialização, principalmente a desidratação e a conserva enlatada,
podem reduzir significativamente a quantidade de nutrientes, inclusive de beta glucana,
presentes nos coguemlos, interferindo nas propriedades nutricionais desse alimento (MANZI
et al., 2001, 2004).
Muitos estudos têm reportado a relação e ação da β -glucana como um ativador do
sistema imunológico e modificador da resposta celular, demonstrando atividades
anticarcinogênicas, prevenindo a oncogênese e metástase (AKRAMIENĖ et al., 2007).
Portanto, os resultados apresentados e discutidos em relação aos valores de α-glucana
no P. ostreatus de tamanhos pequeno e grande sugerem que o processo de embalá-los a vácuo
é satisfatório, uma vez que promoveu o aumento de glicose no alimento no decorrer dos
tempos de armazenamento. Para a β-glucana, o empacotamento a vácuo foi eficaz apenas para
o P. ostreatus de tamanho pequeno pois manteve estável os níveis desse elemento até o 28º
dia de estocagem.
4.4.3 pH do Pleurotus ostreatus e dos líquidos exsudados do alimento Ao longo dos períodos de tratamento, houve diminuição no valor do pH dos líquidos
exsudados do Pleurotus ostreatus de tamanhos pequeno e grande embalados sem vácuo. Nos
empacotados com vácuo, a concentração de íons hidrogênio nos líquidos exsudados
permaneceu constante quando comparados com as amostras pré-tratadas (Figura 27).
Em relação ao pH dos cogumelos, dentre todas as amostras analisadas nota-se que o P.
ostreatus de tamanho grande empacotado sem vácuo teve o maior aumento na concentração
de íons hidrogênio, no decorrer dos tempos de armazenamento (Figura 27).
Tal fato pode estar correlacionado com a presença de fungos e leveduras que
apresentam a capacidade de se desenvolverem em condições de pH ácido, em temperaturas de
refrigeração e em Aa elevada (TANIWALKI et al., 1999; SILVA JUNIOR, 2008).
Esses resultados corroboram os evidenciados por Martins et al. (2006) que estudaram
fruta-pão minimamente processada e armazenada em diferentes situações: sem embalagem,
embaladas com filme de PVC e em sacos de polietileno sob vácuo. Ao final de 12 dias de
estocagem a 5ºC houve um acentuado declínio nos valores de pH da fruta embalada sob
vácuo.

80
Figura 27: Valor de pH do cogumelo Pleurotus ostreatus de tamanhos pequeno e grande e dos líquidos exsudados do alimento. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
Comparando o pH das amostras controle e das estocadas por 28 dias nota-se que as
maiores diferenças ocorreram tanto nos líquidos exsudados quanto nos cogumelos embalados
sem vácuo, de ambos os tamanhos (Figura 27), demonstrando que a retirada do oxigênio do
interior das embalagens e o armazenamento sob refrigeração minimizam alterações
metabólicas no P. ostreatus.
Sendo assim, a manutenção do pH nos P. ostreatus armazenados a vácuo é favorável
para garantir a qualidade nutricional, sensorial e microbiológica do alimento.
4.4.4 Análises microbiológicas Os cogumelos são produtos perecíveis, com tempo de vida útil limitado, podendo
sofrer contaminação por diversos micro-organismos se não forem bem armazenados.
Condições extrínsecas aos alimentos, tais como temperatura de armazenamento, falta de
cuidados no processo de manipulação e métodos de conservação, além das condições
intrínsecas, que são a atividade de água, pH e nutrientes, aceleram o metabolismo dos micro-
organismos contaminantes (SILVA JUNIOR, 2008).
A Agência Nacional de Vigilância Sanitária (ANVISA) tem sido o órgão
regulamentador dos padrões microbiológicos toleráveis nos alimentos, sendo que quando
contaminados são considerados impróprios para o consumo humano ou por trazerem risco à

81
saúde ou mesmo por apresentarem características sensoriais (cor, odor, sabor e textura)
alteradas, tornando-se rejeitados pelo consumidor.
Nas análises microbiológicas realizadas dos líquidos exsudados dos cogumelos,
decorridos os tempos de armazenamento, somente foi observada a presença de leveduras
quando examinados em lâminas ao microscópio.
No meio de água peptonada onde os pedaços dos cogumelos foram colocados,
somente foi evidenciada a presença de uma nata de leveduras para algumas amostras. Nos
meios contendo caldo verde-brilhante, ágar verde-brilhante, XLD, Rappaport e ágar-água não
foram detectadas presenças de bactérias gram negativas ou do tipo Salmonella.
A inibição do crescimento bacteriano pelos fungos comestíveis também foi encontrada
numa pesquisa realizada por Wisbeck; Robert e Furlan (2002) com diferentes espécies de
fungos do gênero Pleurotus. Os autores confirmaram a existência de agentes antimicrobianos
contra Bacillus subtilis, Staphylococcus aureus e Candida albicans.
Outros estudos ratificam a existência de agentes antimicrobianos em cogumelos,
inclusive no gênero Pleurotus, com ação inibitória para Escherichia coli, S. aureus, C.
albicans e Pseudomonas aeroginosa (OPIGE et al., 2006; SANTOYO et al., 2009).
Tanto na forma fresca quanto na desidratada, os corpos de frutificação e o micélio de
P. ostreatus demonstram ação antimicrobiana, segundo Wolff et al. (2008). Os corpos de
frutificação frescos e desidratados assim como o micélio fresco mostraram mais de 50% de
inibição contra C. albicans; o micélio desidratado apresentou alto grau de ação contra E. coli
(87,5%) e B. subtilis (57,5%).
Logo, estas evidências demonstraram que o cogumelo P. ostreatus é constituído por
compostos antimicrobianos, o que permite que sobrevivam em seus ambientes naturais
(LINDEQUIST; NIEDERMEYER; JÜLICH, 2005).
Em relação aos pedaços de cogumelos plaqueados no meio de cultura BAD foi
observada presença de leveduras cujas colônias foram submetidas à contagem e expressas em
Unidades Formadoras de Colônias (UFC).
A maior contagem de leveduras foi identificada na amostra de cogumelo de tamanho
grande, empacotada sem vácuo e armazenada por 28 dias (1,853x104 UFC/g cogumelo
fresco), uma diferença de 38,9% na contagem total de leveduras se comparada com a amostra
embalada sob vácuo e estocada pelo mesmo tempo (Figura 28).
Na amostra de cogumelo de tamanho pequeno, fechada sem vácuo e armazenada por
28 dias foram encontradas 3,85x103 UFC/g cogumelo fresco, uma diferença de 92,2% no total

82
de leveduras quando comparada com a amostra embalada com vácuo e estocada pelo mesmo
tempo (Figura 28).
A presença de leveduras nas amostras de cogumelos, de ambos os tamanhos,
embalados com e sem vácuo apresentou um comportamento crescente ao longo dos tempos de
armazenamento, com diferenças estatisticamente significativas entre elas.
No 28º dia de estocagem os P. ostreatus de tamanho grande demonstraram as mais
elevadas contagens de colônias (Figura 28) sugerindo que o desenvolvimento dos cogumelos
pode favorecer o crescimento de leveduras.
Relacionando a presença de leveduras nos P. ostreatus com o pH anteriormente
discutido nota-se que em todas as amostras a concentração de íons hidrogênio foi maior no
28º dia de armazenamento, assim como a contagem de leveduras.
Figura 28: Contagem de colônias de leveduras (UFC/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento. Legenda: G c/vac: cogumelo tamanho grande embalado a vácuo; P c/vac: cogumelo pequeno embalado a vácuo; G s/vac: tamanho grande embalado sem vácuo; P s/vac: tamanho pequeno embalado sem vácuo.
Além da retirada do oxigênio do interior da embalagem o tipo de material utilizado
pode também interferir na comunidade microbiana dos cogumelos. Isto foi provado por
Kuyper et al. (1993) que determinaram os efeitos de diferentes embalagens e suas
permeabilidades na qualidade microbiológica de cogumelos embalados a vácuo e
armazenados por 9 dias em temperatura de 5ºC. Pseudomonas e contagem total de bactérias
foram maiores nas embalagens com mais alta permeabilidade ao O2 do que nas amostras
embaladas com polietileno de baixa permeabilidade o que pode ser resultante das condições
aeróbicas e da elevada umidade presente no interior dos sacos plásticos.

83
Resultados semelhantes foram encontrados por Sapata et al. (2005) utilizando
embalagem de filme permeável com modificação passiva da atmosfera (20,9% O2 e 0% CO2)
o que aumentou para 10 dias de armazenamento o tempo de vida útil dos cogumelos P. sajor-
caju.
Batatas minimamente processadas e embaladas a vácuo também apresentaram
aumento na quantidade de bolores e leveduras na ordem de 7,31x103 UFC/g após 6 dias de
estocagem a 10ºC; contudo as armazendas sem vácuo apresentaram diferença muito maior
(6,50x107 UFC\g) (MICHELS, 2005).
Uma outra condição capaz de inibir a atividade fúngica e o crescimento de bolores e
leveduras em cogumelos, segundo pesquisa realizada por Braga et al. (2005) é o processo de
desidratação. Sendo assim, o crescimento de micro-organismo patogênicos e deteriorantes
podem ser inibidos ou retardados pelo emprego de embalagem e métodos de preservação
adequados.
Não há na legislação brasileira limites de tolerância para bolores e leveduras em
cogumelos, entretanto sabe-se que a presença destes nos alimentos é indesejável, pois
alteram características sensoriais (cor, odor, sabor e textura) o que inviabiliza a venda e
consumo do produto.
Então, os resultados apresentados e discutidos em relação a comunidade microbiana
do P. ostreatus de tamanhos pequeno e grande sugerem que o processo de embalá-los a
vácuo é satisfatório, uma vez que retardou o crescimento de colônias de leveduras no
decorrer dos tempos de armazenamento.
4.4.5 Análise sensorial Concentrações de dióxido de carbono e oxigênio nas embalagens de cogumelos têm
sido responsabilizadas, em parte, pelas mudanças sensoriais ocorridas na pós-colheita, o que
interfere na qualidade do produto (MAU et al., 1993).
Por retirar quase que totalmente o oxigênio presente no interior da embalagem, o
processo de empacotamento a vácuo minimiza a taxa de respiração celular dos cogumelos
bem como retarda as reações químicas dependentes de oxigênio, garantindo melhores
propriedades sensoriais do P. ostreatus.
Na tabela 1 observa-se que o sabor dos cogumelos in natura foi considerado ruim por
ser um produto que precisa sofrer cocção para ser consumido, fato esse que pode ser
comprovado pela boa aceitação das amostras pré-tratadas.

84
Tabela 1: Características sensoriais do Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
Legenda- B:bom; R, ruim; P, péssimo; ME: marrom escuro; MEs: marrom esverdeado; MC: marrom claro; AE: amarelo escuro; AC: amarelo claro; F: firme; M: mole * referente ao píleo (cogumelo pequeno) e a lamela (cogumelo grande) ** não foi possível degustar devido a alterações encontradas nos atributos textura e/ou odor e/ou cor.
No cogumelo de tamanho grande embalado sem vácuo, até o 3º dia de armazenamento
o sabor permaneceu bom, porém depois desse tempo de tratamento não foi mais possível
degustá-lo devido a alterações acentuadas no odor e na textura. O sabor do P. ostreatus
empacotado com vácuo foi avaliado como bom até o 28º dia de armazenamento (Tabela 1).
Nas amostras de cogumelo pequeno fechadas sem vácuo, durante todos os tempos de
tratamento, o sabor foi considerado péssimo, diferentemente dos embalados com vácuo que
foram avaliados como alterados somente após o 14º dia de armazenamento (Tabela 1).
O odor dos cogumelos grandes fechados sem vácuo e mantidos por até 3 dias na
geladeira foi avaliado como bom, tornando-se péssimo (azedo e fermentado) após este tempo
de armazenamento. Os empacotados com vácuo mantiveram o odor bom até o 28º dia de
tratamento (Tabela 1).
Em todas as amostras de cogumelo grande embaladas com vácuo a textura se alterou
no 3º dia de estocagem, ficando mole e rompendo-se facilmente quando na mastigação. Nos
sem vácuo, a textura foi considerada firme até o 7º dia, amolecendo nos tempos posteriores
(Tabela 1). Esses resultados vêm ao encontro com os relatados por Bano e Rajarathnam
(1988).
Nas amostras de cogumelos de tamanho pequeno empacotados com vácuo, somente a
partir do 14º dia de armazenamento é que houve alteração na textura, na cor e no odor do
alimento. Os P. ostreatus embalados com vácuo tiveram todos os atributos sensoriais
preservados até o 28º dia de tratamento (Tabela 1).
Amostras Com vácuo Sem vácuo Sabor Odor Cor* Textura Sabor Odor Cor* Textura
Cogumelo grande
Controle R B AE F R B AE F Pré-tratado B B AC F B B AC F 3dias B B AE M B B ME F 7dias B B AC M ** P AC M 14dias B B AC M ** P ME M 28dias ** R AC M ** P ME M
Cogumelo pequeno
Controle R B ME F R B ME F Pré-tratado B B ME F B B ME F 3dias B B ME F P B ME F 7dias B B ME F P B ME F 14dias B B ME F P R MEs M 28dias P P MC F P P MEs M

85
A textura é considerada uma das mais importantes qualidades dos cogumelos frescos
que até a colheita apresentam-se firmes e tenros. Após esse processo sofrem alterações como
amolecimento e enrijecimento, o que pode ser resultante da síntese de quitina na parede
celular ou da diminuição do turgor celular, ambos por mudança na permeabilidade da
membrana (MAU et al., 1993).
A textura amolecida do cogumelo pode ser um sinal inicial de deterioração do
produto, pelas reações enzimáticas naturais que degradam sua parede celular. Depois, reações
oxidativas passam a acontecer ocorrendo alterações no sabor e aroma, além de perdas no valor
nutricional (ORNELAS, 2007).
Cogumelos in natura possuem coloração amarelo escuro, quando grandes, e marrom
escuro quando pequenos. Após o tratamento, cogumelos de tamanho grande alteram a cor,
tornando-se amarelo claro, adquirindo novamente a tonalidade amarela escura após 3 dias de
armazenamento, tanto nas amostras embaladas a vácuo quanto nas sem. Nos tempos seguintes
de armazenamento (7, 14 e 28 dias), em ambos os processos de acondicionamento, a
tonalidade evidenciada foi amarela clara e marrom escuro (Figura 29).
Uma das estratégias comumente empregadas para manter o frescor dos cogumelos é o
armazenamento em temperaturas frias (cerca de 5ºC), pois ocorre supressão da maturação,
retardo nas atividades metabólicas e redução da atividade das enzimas oxiderdutases (BANO;
RAJARATHNAM, 1988; ADAMS, 1991).

86
H
E
G
C
B
F
D
D
Figura 29: Fotos do Pleurotus ostreatus tamanho grande armazenados por: A- 3 dias com vácuo; B- 3 dias sem vácuo; C- 7 dias com vácuo; D- 7 dias sem vácuo; E- 14 dias com vácuo; F- 14 dias sem vácuo; G- 28 dias com vácuo; H- 28 dias sem vácuo; I- in natura. Setas indicam líquido esbranquiçado no interior da embalagem.
I
A

87
A amostra de cogumelo pequeno embalado sem vácuo, a partir do 14º dia, apresentou
uma coloração alterada (marrom esverdeado) tornando-o indesejado para o consumo (Figura
30).
Relacionando a cor das amostras de cogumelo com a presença das enzimas
polifenoloxidase (PPO) e peroxidase (POD) pode-se afirmar que na amostra de cogumelo
pequeno, embalado sem vácuo, a atividade da enzima PPO foi maior no 3º dia de
armazenamento, o que explica a coloração marrom escura. Esta mesma associação não pode
A B
C D
Figura 30: Fotos do Pleurotus ostreatus tamanho grande armazenados por: A- 14 dias com vácuo; B- 14 dias sem vácuo; C- 28 dias com vácuo; D- 28 dias sem vácuo; E- in natura. Setas indicam líquido esbranquiçado no interior da embalagem.
E

88
ser realizada para a amostra de cogumelo grande que apresentou a mais alta atividade
enzimática da PPO somente no 28º dia de armazenamento.
Estes resultados podem ser explicados pelos danos mecânicos (colheita e o corte)
causados durante o processamento o que pode aumentar a concentração de dióxido de
carbono, etileno e água livre, desencadeando alterações no sabor e no aroma do P. ostreatus,
bem como elevando a atividade das enzimas oxiredutoras, causando o escurecimento do
cogumelo (MORETTI, 2002).
Segundo Freitas et al. (2008) a enzima POD é capaz de catalisar um grande número de
reações oxidativas, por pertencer ao grupo das oxidoredutases, usando peróxido como
substrato ou oxigênio como aceptor de hidrogênio. Em cogumelos, esta enzima induz a
mudanças negativas de sabor e cor durante a estocagem.
Comparando as características sensoriais com os resultados microbiológicos nota-se
que a contagem de leveduras foi maior nas amostras de cogumelos de tamanhos grande
(1,853x104 UFC/g cogumelo fresco) e pequeno (3,85x103 UFC/g cogumelo fresco),
embaladas sem vácuo e armazenadas por 28 dias, as quais também apresentavam odor
péssimo, coloração amarelada e marrom escura e textura mole, alterações estas que
impossibilitaram a degustação da amostra para avaliação do sabor. Além disso, observou-se
também nestas amostras um líquido esbranquiçado, indicando provavelmente a presença de
leveduras (Figura 29 e 30).
Segundo Mau et al. (1993), os cogumelos frescos são altamente perecíveis e
preservam suas características por cerca de 2 a 3 dias depois da colheita a temperatura
ambiente. Sob temperatura de refrigeração (10-13ºC) este tempo pode ser estendido, pois
reações pós-colheita são retardadas.
Por conseguinte, o processo de empacotamento a vácuo do P ostreatus pode ser
considerado favorável em relação aos aspectos sensoriais, pois o cogumelo de tamanho
grande armazenado por até 14 dias apresentou o sabor, odor e cor preservados. O cogumelo
pequeno fechado a vácuo pelo mesmo período de tempo demonstrou todas as características
sensoriais preservadas.

89
5 CONCLUSÕES E SUGESTÕES
Os resultados evidenciados nesta pesquisa permitiram concluir que:
1) O pré-tratamento dos cogumelos, por um tempo de 4 minutos, favoreceu a perda dos
compostos fenólicos e a diminuição da atividade da PPO e da POD, principalmente em
cogumelos de tamanho grande. Quantidades menores de proteínas foram perdidas para a água
de lavagem quando submetidos à fervura.
2) Comparando a perda de água dos cogumelos frescos e pré-tratados, para todos os tempos
de tratamento, as diferenças foram significativas, sendo maiores nos cogumelos grandes.
3) Pleurotus ostreatus, tanto de tamanho pequeno quanto grande, podem ser armazenados por
até 14 dias, sob refrigeração, quando embalados a vácuo, pois apresentaram características
sensoriais preservadas, menor contagem de colônias de leveduras, maior teor de proteína e
vitamina C, presença diminuída de compostos fenólicos e atividades reduzidas das enzimas
polifenoloxidase e peroxidase quando comparados com cogumelos armazenados por 28 dias.
4) Cogumelos pequenos demonstraram melhores resultados na maioria das análises
realizadas, comprovando que este tamanho, atualmente comercializado, tem sido o mais
favorável.
5) Pode ser considerado um alimento fonte e com alto teor de vitamina C, pois apresentaram
em média 36,1mg em 100g de cogumelo.
6) Quanto a comunidade microbiana, nenhuma espécie de bactéria se desenvolveu no P.
ostreatus. As amostras apresentaram somente colônias de leveduras, que aumentaram no
decorrer dos tempos de armazenamento, principalmente nos embalados sem vácuo.
7) As propostas para trabalhos futuros serão investigar outros compostos nutricionais e
averiguar a viabilidade econômica da implantação em escala comercial do processo de
fechamento a vácuo dos cogumelos Pleutorus ostreatus.

90
6 REFERÊNCIAS
ADAMS, J. B. Significance of enzymes in individual vegetables. In: FOX, P. F. Food Enzymology, London: Elsevier, 1991. p. 499-537.
AKRAMIENĖ, D. et al. Effects of b-glucans on the immune system. Medicina, v. 43, n. 8, p. 597-606, 2007.
AOAC: Association of Official Analytical Chemists. Official Methods of Analysis, 15. ed. Washington, D.C., v. 1, 1990.
APATI, G. P. Secagem e resfriamento à vácuo de cogumelos comestíveis da espécie Pleurotus ostreatus DSM 1833. 2004. 90f. Dissertação (Mestrado em Engenharia de Alimentos). Departamento de Engenharia Química de Alimentos, Universidade de Santa Catarina, Florianópolis, 2004.
AHN, W. S. et al. Natural killer cell activity and quality of life were improved by consumption of a mushroom extract, Agaricus blazei Murril Kyowa, in gynecological cancer patients undergoing chemotherapy. In: III SIMPÓSIO INTERNACIONAL DE COGUMELOS NO BRASIL E II SIMPÓSIO NACIONAL SOBRE COGUMELOS COMESTÍVEIS, 2006, Brasília. Anais...Brasília: EMBRAPA, 2006, p. 89- 96.
ANJOS, V. D. de A. et al. Estabilidade do purê de tomate em embalagens de consumo: aspectos sensoriais. Brazilian Journal of Food Technology, v. 6, n. 2, p. 171-177, jul./dez. 2003.
ANVISA- Agência Nacional de Vigilância Sanitária. Guia de estabilidade de produtos cosméticos. Brasília: ANVISA, 2004. p. 11-24.
ARAÚJO, W. M. C.; BORGO, L. A.; ARAÚJO, H. M. C. Aspectos da química e da funcionalidade das substâncias químicas presentes nos alimentos.. In: ARAÚJO, W. M. C et al. (Org.). Alquimia dos alimentos. 2. ed. Brasília: Senac- DF, 2008. p. 128- 130.
ARAÚJO, J. M. A. Química dos alimentos: teoria e prática. 4. ed. Viçosa: UFV, 2008. 596p.

91
AYDIN, N.; KADIOGLU, A. Changes in the chemical composition, polyphenol oxidase and peroxidase activities during development and ripening of medlar fruits (Mespilus germanica L.). Bulgarian Journal of Plant Physiology, v. 27, n. 3–4, p. 85–92, 2001.
BACH, E. E. Características de polissacarídeos estraídos de Agaricus no Brasil. In: III SIMPÓSIO INTERNACIONAL DE COGUMELOS NO BRASIL E II SIMPÓSIO NACIONAL SOBRE COGUMELOS COMESTÍVEIS, 2006, Brasília. Anais...Brasília: EMBRAPA, 2006, p. 82-86.
BANO, Z.; RAJARATHNAM, S. Pleurotus mushrooms. Parte II. chemical, composition, nutritional value, post- harvest physiology, preservation and role as human food. Critical Reviews in Food Science and Nutrition, v. 27, n. 2, p. 87-158, 1988.
BARROS, L. et al. Effect of fruiting body maturity stage on chemical composition and antimicrobial activity of Lactarfius sp. mushrooms. Journal of Agricultural and Food Chemistry, v. 55, n. 21, p. 8766-8771, out. 2007.
BARUFFALDI, R.; OLIVEIRA, M. N. Fundamentos de tecnologia de alimentos. v. 3, São Paulo; Atheneu, 1998. p. 254
BEETZ, A.; KUSTUDIA, M. Mushroom cultivation and marketing. National Sustainable Agriculture Information Service. 2004. Disponível em: <http://www.attra.ncat.org>. Acesso em: 11 mar. 2008.
BERBICZ, F.; CLEMENTE, E. Avaliação da termoestabilidade e da regeneração da atividade da peroxidase extraída de laranja (Citrus spp.). Acta Scientiarum, Maringá, v. 23, n. 5, p. 1239-1242, 2001.
BERNARDI, E.; DONINI, L. P.; MINOTTO E.; NASCIMENTO, J. S. Utilização de diferentes substratos para a produção de inóculo de Pleurotus ostreatoroseus Sing. Ciência Agronômica, v. 38, n. 1, p. 84-89, 2007.
BOBEK, P.; OZDIN, L.; KUNIAK, L. Effect of oyster mushroom and isolated beta-glucan and lipid peroxidation and on the activities of antioxidative enzymes in rats fed the cholesterol diet. Journal of Nutritional Biochemistry, v. 8, n. 8, p. 469-471, ago. 1997.
BOBBIO, P. A.; BOBBIO, F. O. Química do processamento de alimentos. 3. ed. São Paulo: Varela, 2001. p. 5-8.

92
BONATTI, M. et al. Evaluation of Pleurotus ostreatus and Pleurotus sajor-caju nutritional characteristics when cultivated in different lignocellulosic wastes. Food Chemistry, v. 88, n. 3, p. 425-428, dez. 2004.
BONATTI, M. et al. Estudo da comparação de cogumelos das espécies Pleurotus ostreatus e Pleurotus sajor-caju cultivados em palha de bananeira. Revista Saúde e Ambiente, v. 4, n. 1, p. 31-35, jun. 2003.
BONONI, V. L. et al. Cultivo de cogumelos comestíveis. 2. ed. São Paulo: Ícone, 1999. 206 p.
BRAGA, G. C. et al. Variações de cor e atividade de água em Agaricus blazei desidratado e armazenado em diferentes embalagens plásticas. Acta Scientiarum: Technology, Maringá, v. 27, n. 1, p.83-87, jan./jun. 2005.
BRASIL. Portaria nº 27 de 13 de janeiro de 1998. Dispõe sobre o regulamento técnico referente à informação nutricional complementar. Brasília: Agência Nacional de Vigilância Sanitária- ANVISA, 1998. Disponível em: <http://e-legis.anvisa.gov.br/leisref/public/showAct.php?id=97>. Acesso em: 15 jan. 2007.
BRASIL. Resolução RDC - nº 12 de 02 de janeiro de 2001. Dispõe sobre os princípios gerais para o estabelecimento de critérios e padrões microbiológicos para alimentos. Brasília: Agência Nacional de Vigilância Sanitária – ANVISA, 2001. Disponível em: <http://www.anvisa.gov.br/legis/resol/12_01rdc.htm>. Acesso em: 18 dez. 2006.
BRASIL. Resolução RDC nº. 269, de 22 de setembro de 2005. Dispõe sobre o regulamento técnico sobre a ingestão diária recomendada (IDR) de proteína, vitaminas e minerais. Brasília. Agência Nacional de Vigilância Sanitária- ANVISA, 2005. Disponível em: <http://e-legis.anvisa.gov.br/leisref/public/showAct.php?id=18828&word>. Acesso em: 19 dez. 2006.
BRAVO, L. Polyphenols: chemistry, dietary sources, metabolism and nutrition significance. Nutrition Reviews, New York, v. 56, n. 11, p. 317-333, nov. 1998.
BROWN, G. D.; GORDON, S. Immune recognition: a new receptor for β- glucans. Nature, v. 413, n. 6851, p. 36-37, 2001.
BURTON, G. R. W.; ENGELKIRK, P. G. Microbiologia para as ciências da saúde. 7. ed. Rio de Janeiro: Guanabara Koogan, 2005. p. 95- 97.

93
CAMPOS, F. M.; MARTINO, H. S. D.; SABARENSE, C. M.; PINHEIRO-SANT’ANA, H. M. Estabilidade de compostos antioxidantes em hortaliças processadas: uma revisão. Alimentos e Nutrição, Araraquara, v. 19, n. 4, p. 481-490, out./dez. 2008.
CARNEIRO, C. E. A.; ROLIM, H. M. V.; FERNANDES, K. F. Estudo das atividades de peroxidades e polifenoloxidase de guariroba (Syagrus oleracea Becc) sob a ação de diferentes inibidores. Acta Scientiarum: Biological Sciences, Maringá, v. 25, n. 1, p. 189-193, 2003.
CHANG, S.; MILES, P. G. Mushrooms: cultivation, nutritional value, medicinal effect, and environmental impact. 2. ed. New York: CRC Press, 2004. 482 p.
CHEUNG, P. C. K. Dietary fiber content and composition of some cultivated edible mushroom fruiting bodies and mycelia. Journal of Agricultural and Food Chemistry, v. 44, n. 2, p. 468-471, fev. 1996.
CHEUNG, P. C. K. Dietary fibre content and composition of some edible fungi determined by two methods of analysis. Journal of Agricultural and Food Chemistry, v. 73, n. 2, p. 255-260, fev. 1997.
CHILAKA, F. C.; EZE, S.; ANYADIEGWU, C.; UVERE, P. O. Browning in processed yams: peroxidase or polyphenol oxidase? Journal of the Science of Food and Agriculture, v. 82, n. 8, p. 899-903, 2002.
CLARKE, J. Mushrooms: the new superfood. Disponível em: <http://www.mushroom-uk.com>. Acesso em: 23 set. 2007.
CLEMENTE, E. Isolamento, purificação e termoestabilidade da isoperoxidase do suco de laranja. Ciência e Tecnologia de Alimentos, Campinas, v. 16,n . 1, p. 1-5, jan./maio 1996.
CLEMENTE, E. Peroxidase from oranges (Citrus sinenses (L.) Osbeck. European Food Research and Technology, v. 215, n. 2, ago. 2002.
COHEN, R.; PERSKY, L.; HADAR, Y. Biotechnological applications and potential of wood-degrading mushrooms of the genus Pleurotus. Applied Microbiology Biotechnology, v. 58, n. 5, p. 582-594, abr. 2002.
COUTINHO, L. N. Cultivo de espécies de cogumelo comestíveis. Instituto Biológico de São Paulo: 2005. Disponível em: <http://www.portaldoagronegocio.com.br/conteudo.php?id=23083>. Acesso em: 21 dez. 2006.

94
CROFT, K. D. The chemistry and biological effects of flavonoids and phenolic acids. Annals of the New York Academy of Science, New York, v. 854, p. 435-442, nov. 1998.
DABA, A. S.; KABEIL, S. S, BOTROS, W. A.; EL-SAADANI, M. A. Production of Mushroom (Pleurotus ostreatus) in Egypt as a Source of Nutritional and Medicinal Food. World Journal of Agricultural Sciences, v. 4, n. 5, p. 630-634, 2008.
DIAS, E. S. et al. Cultivo do cogumelo Pleurotus sajor-caju em diferentes resíduos agrícolas. Ciência e Agrotecnologia, Lavras, v. 27, n. 6, p. 1363-1369, nov./dez. 2003.
DUPRAT, L. A.; SOUZA, J. V. Análise da comercialização e do consumo de cogumelos comestíveis no mercado do Distrito Federal e entorno. Boletim de Pesquisa e Desenvolvimento, Brasília, ano 48, 34 p., abr. 2003. Disponível em: <http://www. cenargen.embrapa.br/publica/trabalhos/bp048.doc>. Acesso em: 24 jul. 2006.
DUTCOSKY, S. D. Análise de alimentos. 2. ed. Curitiba: Champagnat, 2007. 221p.
EIRA, A. F. Fungos comestíveis: In: ESPOSITO, E.; AZEVEDO, J. L. (Org). Fungos: uma introdução à biologia, bioquímica e biotecnologia. Caxias do Sul: Educs, 2004. p. 379-448.
FALCÃO, S. I. D. M. Avaliação da actividade electroquímica em cogumelos silvestres comestíveis. 2008. 91f. Dissertação (Mestrado em Química). Faculdade de Ciências, Universidade do Porto, Portugal, 2008.
FANG, L.; JIANG, B.; ZHANG, T. Effect of combined high pressure and thermal treatment on kiwifruit peroxidase. Food Chemistry, v. 109, v. 4, p. 802- 807, 2008.
FAOSTAT data, 2009. FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS. Agricultural Production - Crops Primary. Disponível em: <http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor>. Acesso em: 21 abr. 2009.
FREITAS, A. A. et al. Atividades das enzimas peroxidase (POD) e polifenoloxidase (PPO) nas uvas das cultivares benitaka e rubi e em seus sucos e geléias. Ciência e Tecnologia de Alimentos, Campinas, v. 28, n. 1, p. 172-177, jan./mar. 2008.
FURLANI, R. P. Z. Valor nutricional de cogumelos cultivados no Brasil. 2004. 99f. Tese (Doutorado em Ciência de Alimentos). Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Campinas, 2004.

95
FURLANI, R. P. Z.; GODOY, H. T. Valor nutricional de cogumelos comestíveis: uma revisão. Revista do Instituto Adolfo Lutz, São Paulo, v. 64, n. 2, p.149-154, jul./dez. 2005.
FURLANI, R. P. Z.; GODOY, H. T. Valor nutricional de cogumelos comestíveis. Ciência e Tecnologia de Alimentos, Campinas, v. 27, n.1, p. 154-157, jan./mar. 2007.
GALLAGHER, M. L. Vitaminas. In: MAHAN, L. K.; ESCOTT- STUMP, S. Krause: alimentos, nutrição e dietoterapia. 11. ed. São Paulo: Roca, 2005. p. 108-109.
GALVAGNO, M. A.; FORCHIASSIN, F. Fisiologia dos fungos: crescimento, morfologia e diferenciação. In: ESPOSITO, E.; AZEVEDO, J. L. (Org). Fungos: uma introdução à biologia, bioquímica e biotecnologia. Caxias do Sul: Educs, 2004. p. 91
GAMACHE, P., RYAN, E., ACWORTH, I. N. Analysis of phenolic and flavonoid compounds in juice beverages using high-performance liquid chromatography with coulometric array detection. Journal of Chromatography, Amsterdam, v. 635, n. 1, p. 143-150, abr. 1993.
GENÇCELEP, H. et al. Determination of mineral contents of wild-grown edible mushrooms. Food Chemistry, v. 113, n. 4, p. 1033-1036, abr. 2009.
GENNARI, J. L. Cogumelos medicinais como prevenção e tratamento de enfermidades. In: III SIMPÓSIO INTERNACIONAL DE COGUMELOS NO BRASIL E II SIMPÓSIO NACIONAL SOBRE COGUMELOS COMESTÍVEIS, 2006, Brasília. Anais...Brasília: EMBRAPA, 2006, p. 98-101.
GIL, M. I.; FERRERES, F.; TOMÁS-BARBERÁN, F. A. Effect of postharvest storage and processing on the antioxidant constituents (flavonoids and vitamin C) of fresh-cut spinach. Food Chemistry, v. 47, n. 6, p. 2213-2217, 1999.
GINANI, V. C.; PINELLI, L. L. O. A estética do gosto. In: ARAÚJO, W. M. C. et al. (Org.). Alquimia dos alimentos. 2. ed. Brasília: Senac- DF, 2008. p. 61- 88.
GOMPERTZ, O. F. et al. Características gerais dos fungos. In: TRABULSI, L. R.; ALTERTHUM, F. (Ed). Microbiologia. São Paulo: Atheneu, 2008. p. 479- 489.
GOMES, C. E. T.; SANTOS, E. C. Estudo da legislação e método de conservação dos alimentos. In: CANDIDO, Cynthia Cavalini et al. Nutrição: guia prático. São Paulo: Iátria, 2006. p. 161-174.

96
GUGLIOTTA, A. de M.; CAPELARI, M. Taxonomia de Basidiomicetos. In: BONONI, V. L. R. (Org.). Zigomicetos, Basidiomicetos e Deuteromicetos: noções básicas de taxonomia e aplicações biotecnológicas. São Paulo: Instituto de Botânica- Secretaria de Estado do Meio Ambiente, 1998. p. 69-102.
GUNDE-CIMERMAN, N.; PLEMENITAS, A.; CIMERMAN, A. Pleurotus fungi produce mevinion, and inhibitor of HMG CoA redutase. FEMS Microbiology Letters, v. 133, n. 3, p. 333-338, nov. 1993.
GUNDE-CIMERMAN, N.; CIMERMAN, A. Pleurotus fruiting bodies contain the inhibitor of 3-Hydroxy-3-Methylglutaryl-Coenzyme A reductase-lovastatin. Experimental Mycology, v. 19, n. 1, p. 1-6, mar. 1995.
GUNDE-CIMERMAN, N. Medicinal Values of the Genus Pleurotus. International Journal of Medicinal Mushrooms, New York, v. 1, p. 69-80, 1999.
GUZMAN, G. Genus Pleurotus (Jacq.: Fr.) P. Kumm. (Agaricomycetideae): diversity, taxonomic problems, and cultural and traditional medicinal uses. Internacional Journal of Medicinal Mushrooms, v. 2, n. 2, p. 95-124, 2000.
HEMEDA, H. M.; KLEIN, B. P. Inactivation and regeneration of peroxidase activity in vegetable extracts treated with antioxidants. Journal of Food Science, Chicago, v. 56, n. 1, p. 68-71, jan. 1991.
HERRERA, O. M. Produção, economicidade e parâmetros energéticos do cogumelo Agaricus blazei: um enfoque na cadeia produtiva. 2001. 183f. Tese (Doutorado em Ciências Agronômicas). Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, 2001.
INSTITUTO ADOLFO LUTZ (SÃO PAULO). Métodos químicos e físicos para análise de alimentos. 4. ed. São Paulo: Instituto Adolfo Lutz, 2005. p. 666-672.
ISMAIL, A.; MARJAN, Z. M.; FOONG, C. H. Total antioxidant activity and phenolic content in selected vegetables. Food Chemistry, v. 87, n. 4, p. 581-586, 2004.
JAWORSKA, G; BERNAS, E. Qualitative changes in Pleurotus ostreatus (Jacq.: Fr.) Kumm. mushrooms resulting from different methods of preliminary processing and periods of frozen storage. Journal of the Science of Food and Agriculture, v. 89, n. 6, p. 1066-1075, mar. 2009.

97
KALAC, P. Chemical composition and nutritional value of European species of wild growing mushrooms: a review. Food Chemistry, v. 113, n. 1, p. 9-16, mar. 2009.
KABASAKALIS, V.; SIOPIDOU, D.; MOSHATOU, E. Ascorbic acid content of commercial fruit juices and its rate of loss upon storage. Food Chemistry, v. 70, n. 3, p. 325-328, ago. 2000.
KIMATI, H. et al. Manual de fitopatologia: doenças das plantas cultivadas. 4. ed. São Paulo: Agronômica Ceres, v. 2, 2005, 663 p.
KING, A.; YOUNG, G. Characteristics and occurrence of phenolic phytochemicals. Journal of the American Dietetic Association, Chicago, v. 99, n. 2, p. 213-218, fev. 1999.
KLEIN, B. P. Nutritional consequences of minimal processing of fruits and vegetables. Journal of Food Quality, v.10, n. 3, p. 179-93, 1987.
KLOPOTEK, Y.; OTTO, K.; BÖHM, V. Processing strawberries to different products alters contents of vitamin C, total phenolics, total anthocyanins, and antioxidant capacity, Journal of Agricultural and Food Chemistry, v. 53, n. 14, p. 5640-5646, jul. 2005.
KRUMMEL, D. Nutrição na hipertensão. In: MAHAN, L. K.; ESCOTT- STUMP, S. Tradução: Andrea Favano. Krause: alimentos, nutrição e dietoterapia. 11. ed. São Paulo: Roca, 2005. p. 859-876.
KÜES, U.; LIU, Y. Fruiting body production in basidiomycetes. Applied Microbiology and Biotechnology, v. 54, n. 2, p.141-152, ago. 2000.
KUYPER, L.; WELNERT, I. A. C.; MCGILL, A. E. J. The effect of modified atmosphere packaging and addition of calcium hypochloride on the atmosphere composition colour and microbial quality of mushrooms. Lebensmittel-Wissenschaft und Technonlogie, London, v.26, p. 14-20, 1993.
LAJOLO, F. M.; TIRAPEGUI, J. Proteínas e aminoácidos. In: DUTRA-de-OLIVEIRA, J. E.; MARCHINI, J. S. Ciências nutricionais. São Paulo: Sarvier, 1998. p. 58-60.
LAURENTI, C.; CLEMENTE, E. Avaliação da atividade da peroxidase em carambola (Oxalidacia averrhoa) em diferentes estágios de maturação. Acta Scientiarum: Agronomy, Maringá, v. 27, n. 1, p. 159-163, jan./mar. 2005.

98
LEHNINGER, A. L. Princípios de bioquímica. 4. ed. São Paulo: Sarvier, 2006. p.192-193.
LEVER, M. A new reaction for colorimetric determination of carbohydrates. Analytical Biochemistry, Academic Press, v. 47, n. 1, p. 273-279, maio 1972.
LI, Y. R. et al. A novel lectin with potent antitumor, mitogenic and HIV-1 reverse transcriptase inhibitory activities from the edible mushroom Pleurotus citrinopileatus. Biochimica et Biophysica Acta: BBA. General Subjects, v. 1780, n. 1, p. 51-57, jan. 2008.
LIMA D. M. et al. Tabela brasileira de composição de alimentos / NEPA-UNICAMP.- Versão II. - Campinas: NEPA-UNICAMP, 2006. p. 27-35.
LINDEQUIST, U.; NIEDERMEYER, T. H. J.; JÜLICH, W. The Pharmacological Potential of Mushrooms. Evidence-based Complementary and Alternative Medicine, v. 2, n. 3, p. 285-299, 2005. Disponível em: <http://ecam.oxfordjournals.org/cgi/content/full/2/3/285>. Acesso em: 26 jul. 2009.
LOGUERCIO-LEITE, C. Taxonomia dos fungos: In: ESPOSITO, Elisa; AZEVEDO, João Lúcio de (Org). Fungos: uma introdução à biologia, bioquímica e biotecnologia. Caxias do Sul: Educs, 2004. p. 48-88.
LOPES, A. S.; CLEMENTE, E. Isolamento e caracterização de peroxidase (POD) e polifenoloxidase (PPO) em brócolos (Brassica oleracea L. Cv. italica). In: XI ENCONTRO ANUAL DE INICIAÇÃO, 2002, Paraná: Universidade Federal do Paraná, 2002. Anais… Disponível em: <http://www.ppg.uem.br/Docs/pes/eaic/XI_EAIC/trabalhos/arquivos/11-0168-0.pdf>. Acesso em: 3 fev. 2009.
LOWRY, O. H.; ROSEBROUGH, N. J.; FARR, A. L.; RANDALL, R. J. Protein measurement with the Folin phenol reagent. Journal Biological Chemistry, Baltimore, v. 193, p. 265-275, nov. 1951.
LULL, C.; WICHERS, H. J; SAVELKOUL, H. F. J. Antiinflammatory and immunomodulating properties of fungal metabolites. Mediators of Inflammation, v. 2, p. 63-80, 2005. Disponível em: <http://www.hindawi.com/GetArticle.aspx?doi=10.1155/MI.2005.63>. Acesso em: 31 mar. 2009.
LUPETTI, K. O. et al. Análise de imagem em química analítica: empregando metodologias simples e didáticas para entender e prevenir o escurecimento de tecidos vegetais. Química Nova, v. 28, n. 3, p. 548-554, maio/jun. 2005.

99
MANZI, P. et al. Nutrients in edible mushrooms: an inter- species comparative study. Food Chemistry, v. 65, n. 4, p. 477-482, jun. 1999.
MANZI, P.; AGUZZI, A.; PIZZOFERRATO, L. Nutritional value of mushrooms widely consumed in Italy. Food Chemistry, v. 73, n. 3, p.321-325, maio 2001.
MANZI, P.; MARCONI, S.; AGUZZI, A.; PIZZOFERRATO, L. Commercial mushrooms: nutritional quality and effect of cooking. Food Chemistry, v. 84, n. 2, p. 201-206, fev. 2004.
MARSHALL, M. R.; KIM, J.; WEI, C. Enzymatic browning in fruits, vegetables and seafoods, 2000. Disponível em: <http://www.fao.org/ag/ags/agsi/ENZYMEFINAL/Enzymatic%20Browning.html>. Acesso em: 21 jul. 2009.
MARTINS, L. P. et al. Embalagens na conservação de fruta-pão minimamente processada. In: VI SEMINÁRIO DA AGROINDÚSTRIA e III JORNADA NACIONAL DA AGROINDÚSTRIA, 2006, Bananeiras. Anais...Bananeiras, 2006. Disponível em: <http://www.seminagro.com.br/trabalhos_publicados/1jornada/02_ciencia_e_tecnologia_de_alimentos/12cta.pdf>. Acesso em: 9 maio 2009.
MASTERS, S. B. Fármacos utilizados no tratamento de doenças do sangue, inflamação e gota. Fármacos utilizados nas anemias; fatores de crescimento hematopoiéticos. In: KATZUNG, B. G. Farmacologia básica e clínica. 9. ed. Rio de Janeiro: Guanabara Koogan, 2005. p. 446-450.
MATHIAS, S. P. Avaliação físico-química, microbiológica e sensorial do presunto de peru submetido à tecnologia de alta pressão hidrostática. 2008. 82f. Dissertação (Mestrado em Ciência e Tecnologia de Alimentos). Instituto de Tecnologia, Universidade Federal Rural do Rio de Janeiro, Seropédica, 2008.
MATTILA, P. et al. Contents of vitamins, mineral elements, and some phenolic compounds in cultivated mushrooms. Journal of Agricultural and Food Chemistry, v. 49, n. 5, p. 2343-2348, abr. 2001.
MAU, J. L.; MILKLUS, M. B.; BEELMAN, R. B. The shelf life of Agaricus mushrooms. In: CHARALAMBOUS, George (Ed.). Shelf life studies of foods and beverages. New York: Elsevier, 1993. p. 255-283.
MAYETT, Y. et al. Consumption trends of edible mushrooms in developing countries: the case of Mexico. Journal of International Food & Agribusiness Marketing, v. 18, n. 1, p. 151-176, 2006.

100
MCCLEARY, B. V., GLENNIE-HOLMES, M. Enzymic quantification of (1- 3)(1 -4)-b-D-glucan in barley and malt. Journal of the Institute of Brewing, Londres, v. 91, p. 285–295, 1985.
MICHELS, I. Aspectos tecnológicos do processamento mínimo de tubérculos de yacon (Polymnia sonchifolia) armazenados em embalagens com atmosfera modificada. 2005. 107f. Dissertação (Mestrado em Tecnologia de Alimentos). Setor de Tecnologia, Universidade Federal do Paraná, Curitiba, 2005.
MINIM, V. P. R. Análise Sensorial - estudos com consumidores. Viçosa: UFV, 2006. 225 p.
MODA, E. M.; SPOTO, M. H. F.; HORII, J.; ZOCCHI, S. S. Uso de peróxido de hidrogênio e ácido cítrico na conservação de cogumelos Pleurotus sajor-caju in natura. Ciência e Tecnologia de Alimentos, Campinas, v. 25, n. 2, p. 291-296, abr./jun. 2005.
MODA, E. M.; HORII J.; SPOTO M. H. F. Edible mushroom Pleurotus sajor-caju production on washed and supplemented sugarcane bagasse. Scientia Agricola, Piracicaba, v. 62, n. 2, p. 127-132, mar./abr. 2005a.
MOERSCHBACHER, B. M.; HECK, B.; KOGEL, K. H.; OBST, O.; REISNER, H. J. An elicitor of the hypersensitive lignification response in wheat leaves isolated from the rust fungus Puccinia graminis f. sp. tritici. Zeitschrift für Naturforschung - Section C: Biosciences, Zürich, v. 41, n. 9-10, p. 32-37, 1986.
MONTEIRO, C. S. Desenvolvimento de molho de tomate Lycopersicon esculentum Mill formulado com cogumelo Agaricus brasiliensis. 2008. 176f. Tese (Doutorado em Tecnologia de Alimentos). Setor de Tecnologia de Alimentos, Universidade Ferderal do Paraná, Curitiba, 2008.
MORETTI, C. L. Vegetable crops production In: Guidelines for Good Agricultural Practices. Brasília: Embrapa, 2002, v.1, p. 65-97.
MUELLER, G. M.; SCHMIT, J. P. Fungal biodiversity: what do we know? What can we predict? Biodiversity and Conservation, Netherlands, v. 16, n. 1, p. 1-5, jan. 2007.
NEVES, J. C. S. Caracterização estrutural dos polissacarídeos obtidos do basidioma de Pleurotus ostreatus variedade florida. 2007. 79f. Dissertação (Mestrado em Ciências Bioquímica). Departamento de Bioquímica e Biologia Molecular, Universidade Federal do Paraná, Paraná, 2007.

101
NOVAES, M. R. C. G.; FORTES, R. C. Efeitos antitumorais de cogumelos comestíveis da família Agaricaceae. Nutrição Brasil, v. 4, n. 4, p. 207-217, jul./ago. 2005.
OPIGE, M.; KATEYO, E.; KABASA, J. D.; OLILA, D. Antibacterial activity of extracts of selected indigenous edible and medicinal mushrooms of eastern Uganda. International Journal of Tropical Medicine, v. 1, n. 3, p. 111-116, 2006.
ORNELAS, L. H. Técnica dietética: seleção e preparo de alimentos. 8. ed. São Paulo: Atheneu, 2007. p. 158.
OWUSU-APENTEN, R. K. Introduction to Food Chemistry. New York: CRC Press, 2004. p. 199-202.
PEDRA, W. N. et al. Análise química e sensorial de Pleurotus ostreatus cultivado em casca de coco suplementada com farelo de trigo e/ou de arroz. Arquivos do Instituto Biológco, São Paulo, v. 76, n. 1, p. 91-98, jan./mar. 2009.
PELCZAR JUNIOR, M. J.; CHAN, E .C. S.; KRIEG, N. R. Microbiologia: conceitos e aplicações. 2. ed. São Paulo: Pearson Education do Brasil, 1997. p. 459-469.
PELEG, M. On fundamental issues in texture evaluation and texturization - A view. Food Hydrocolloids, v. 20, n. 4, p. 405-414, jun. 2006.
PHILLIPS, C. A. Review: Modified Atmosphere Packaging and its effects on the microbiology quality and safety of products. International Journal of Science and Tecnology, v. 31, n. 6, p. 463-479, dez. 1996.
PINELI, L. L. O. et al. Caracterização química e física de batatas ‘Ágata’ minimamente processadas, embaladas sob diferentes atmosferas modificadas ativas. Pesquisa Agropecuária Brasileira, Brasília, v. 40, n. 10, p. 1035-1041, out. 2005.
POURCHET-CAMPOS, M. A. Fibra dietética. In: DUTRA-de-OLIVEIRA, José E.; MARCHINI, J. S. Ciências nutricionais. São Paulo: Sarvier, 1998. p. 209-215.
RAJARATHNAM, S.; BANO, Z. Pleurotus mushrooms. Parte I: morfology, life cycle, taxonomy, breeding and cultivation. Critical Reviews in Food Science and Nutrition, v. 26, n. 2, p. 157-223, 1987.

102
REDECKER, D.; RAAB, P. Phylogeny of the glomeromycota (Arbuscular mycorrhizal fungi): recent developments and new gene markers. Mycologia, v. 98, n. 6, p. 885-895, 2006.
RIBEIRO, L. C. et al. Ácido fólico: sua importância em situações fisiológicas do ciclo vital. São Paulo: Editora de Projetos Médicos, 2002. 18 p.
RIBEIRO, V. L. Produção de Agaricus blazei Murril no Brasil. In: SIMPÓSIO INTERNACIONAL SOBRE COGUMELOS, ALIMENTAÇÃO, SAÚDE, TECNOLOGIA E MEIO AMBIENTE NO BRASIL, 1., 2003, Brasília. Anais...Brasília: EMBRAPA, 2003, 71-76.
RIOS, A. O.; ABREU, C. M. P.; CORRÊA, A. D. Efeitos da época de colheita e do tempo de armazenamento no escurecimento do tegumento do feijão (Phaseolus vulgaris, L.). Ciências e Agrotecnlogia, Lavras, v. 26, n. 3, p. 550-558, mai./jun. 2002.
RIOS-HURTADO, A.; TORRES-TORRES, M. G.; MEDINA-RIVAS, M. A. Caracterización bromatológica de la seta (Pleurotus sajor-caju) producida em cuatro sustratos orgánicos. Alimentaria, n. 349, p. 85-89, 2003.
ROYSE, D. J. Specialty mushrooms. In: JANICK, J. (ed.). Progress in new crops. New Crops Resource Online Program. 1996. Disponível em: <http://www.hort.purdue.edu/newcrop/proceedings1996/v3-464.html#Table 1>. Acesso em: 20 abr. 2009.
SANTOS, L. M. P.; PEREIRA, M. Z. Efeito da fortificação com ácido fólico na redução dos defeitos do tudo neural. Caderno de Saúde Pública, Rio de Janeiro, v. 23, n. 1, p. 17-24, jan. 2007.
SANTOYO, S.; RAMÍREZ-ANGUIANO, A. C.; REGLERO, G.; SOLER-RIVAS, C. Improvement of the antimicrobial activity of edible mushroom extracts by inhibition of oxidative enzymes. International Journal of Food Science & Technology, v. 44, n. 5, p. 1057-1064, maio 2009.
SAPATA, M. M. et al. Conservação de cogumelos frescos do gênero Pleurotus em atmosfera modificada. In: VII CONGRESSO LUSO-GALAICO DE MACROMICOLOGIA, MACROFUNGOS: DIVERSIDADE E BIOTECNOLOGIA, 2005, Portugal. Anais...Portugal: UTAD, 2005, p. 14. Disponível: <http://www.iniap.min-agricultura.pt/resultado_descritivo_2.aspx?id_publicacao=2954&uni=9>. Acesso em: 15 ago. 2008.

103
SCHWEIGGERT, Ute; SCHIEBER, A.; CARLE, R. Inactivation of peroxidase, polyphenoloxidase, and lipoxygenase in paprika and chili powder after immediate thermal treatment of the plant material. Innovative Food Science & Emerging Technologies, v. 36, n. 4, p. 403-411, dez. 2005.
SHARON, N.; LIS, H. Carbohydrates in cell recognition. Scientific American Magazine, p. 74-81, jan. 1993.
SILVA, N.; JUNQUEIRA, V. C. A. Métodos de análise microbiológica de alimentos. Campinas: ITAL, 1995. 228 p.
SILVA, N.; JUNQUEIRA, V. C. A.; SILVEIRA, N. F. de A. Manual de métodos de análise microbiológica de alimentos. São Paulo: Varela, 1997. 295 p.
SILVA, S. O.; COSTA, S. M. G.; CLEMENTE, E. Chemical composition of Pleurotus pulmonarius (Fr.) Quél., substrates and residue after cultivation. Brazilian Archives of Biology and Technology, v. 45, n. 4, p. 531- 535, dez. 2002.
SILVA, E. G. et al. Análise química de corpos de frutificação de Pleurotus sajor-caju cultivado em diferentes concentrações de nitrogênio. Ciência e Tecnologia de Alimentos, Campinas, v. 27, n. 1, p. 72-75, jan./mar. 2007.
SILVA JUNIOR, E. A. Manual de controle higiênico-sanitário em serviços de alimentação. 6. ed. São Paulo: Varela, 2008. p. 21-35.
SILVEIRA, V. D. Micologia. 5. ed. Rio de Janeiro: Âmbito Cultural, 1995. 351 p.
SIMPSOM, S. J.; PICGOT, J. R.; WILLIAMS, S. A. R. Sensory analysis. International Journal of Food Science and Technology, v. 33, n. 1, p.7-18, fev. 1998.
SMIDERLE, F. R. Caracterização estrutural de alguns polissacarídeos presentes no basidioma de Pleurotus pulmonarius e aplicações 2008. 71f. Dissertação (mestrado em Ciências Bioquímicas). Departamento de Bioquímica e Biologia Molecular, Universidade Federal do Paraná, Paraná, 2008.
SMIDERLE, F.; CARBONERO, E. R.; MELLINGER, C. G.; SASSAKI, G. L.; GORIN, P. A. J.; IACOMINI, M. Structural characterization of a polysaccharide and a beta-glucan isolated from an edible mushroom Flammulina velutipes. Phytochemistry. v. 67, p. 2189-2196, 2006.

104
SMITH, J. E.; ROWAN, N. J.; SULLIVAN, R. Medicinal mushrooms: their therapeutic properties and current medical usage with special emphasis on cancer treatments. University of Strathclyde. 2002. Disponível em: <http:// www.ecancermedicalscience.com/cache/pdf/news-52.pdf>. Acesso em: 21 mar. 2009.
SOUTHERTON, S. G.; DEVERALL, B. J. Changes in phenylalanine ammonia-lyase and peroxidase activities in wheat cultivars expressing resistance to the leaf-rust fungus. Plant Pathology, v. 39, n. 2, p. 223-230, jun. 1990.
SOUZA, E. Método de cultivo de cogumelos comestíveis no Brasil. In: III SIMPÓSIO INTERNACIONAL DE COGUMELOS NO BRASIL E II SIMPÓSIO NACIONAL SOBRE COGUMELOS COMESTÍVEIS, 2006, Brasília. Anais...Brasília: EMBRAPA, 2006, p. 25-36.
SRIVASTAVA, S. K. Peroxidase and poly-phenol oxidase in Brassica juncea plants infected with Macrophomina phaseolina (Tassai) Goid, and their implication in disease resistence. Journal of Phytopathology, v. 120, n. 3, p. 249-254, nov. 1987.
STURION, G. L.; RANZANI, M. R. T. C. Composição em minerais de cogumelos comestíveis cultivados no Brasil - Pleurotus spp e outras espécies desidratadas. Archivos Latinoamericanos de Nutrición, Caracas, v.50, n.1, p.102-108, mar. 2000.
SULTANA, B.; ANWAR, F.; IQBAL, S. Effect of different cooking methods on the antioxidant activity of some vegetables from Pakistan. International Journal of Food Science & Technology, v. 43, p. 560-567, 2008.
SYNYTSYA, A. et al. Mushrooms of genus Pleurotus as a source of dietary fibres and glucans for food supplements. Czech Journal of Food Sciences, v. 26, n. 6, p. 441–446, 2008.
SWAIN, R.; HILLIS, W. E. The phenolic constituents of Prunus domestica. I.- the quantitative analysis of phenolic constituents. Journal of the Science of Food and Agriculture, Oxford, v. 10, n. 1, p. 63-68, jan. 1959.
TANIWALKI, M. H.; IAMANAKA, B. T.; BANHE, A. A. Comparision of culture media to recorver fungi from flour and tropical fruit pulp. Journal of Food Mycology, v. 2, p. 291-302, 1999.
TAVEIRA, V. C.; NOVAES, M. R. C. G. Consumo de cogumelo na nutrição humana: uma revisão da literatura. Comunicação em Ciências da Saúde, v. 18, n. 4, p. 315-322, out./nov. 2007.

105
THORN, G. R.; MONCALVO, J.; REDDY, C. A.; VILGALYS, R. Phylogenetic analyses and distribution of nematophagy support a monophyletic pleurotaceae within the polyphyletic pleuroid-lentinoid fungi. Mycologia, v. 92, n .2, p. 241-252, mar. 2000.
TRUFEM, S. F. B. Taxonomia de zigomicetos. In: BONONI, Vera Lúcia Ramos (Org.). Zigomicetos, Basidiomicetos e Deuteromicetos: noções básicas de taxonomia e aplicações biotecnológicas. São Paulo: Instituto de Botânica- Secretaria de Estado do Meio Ambiente, 1998. p. 19-67.
URBEN, A. F. Importância do uso de cogumelos: aspectos nutricionais e funcionais. In: ENCONTRO FRANCO BRASILEIRO DE BIOCIÊNCIA E BIOTECNOLOGIA, 2002, Brasília. Palestras. Disponível em: <http://www. cenargen.embrapa.br/publica/trabalhos/doc085.pdf>. Acesso em: 24 jul. 2006.
VALDERRAMA, P.; MARANGONI, F.; CLEMENTE, E. Efeito do tratamento térmico sobre a atividade de peroxidase (POD) e polifenoloxidase (PPO) em maçã (Mallus comunis). Ciência e Tecnologia de Alimentos, Campinas, v. 21, n, 3, p. 321-325, set./dez. 2001.
VALLEJO, F.; TOMÁS-BARBERÁN, F.; GARCÍA-VIGUERA, C. Health-Promoting compounds in broccoli as influenced by refrigerated transport and retail sale period. Journal of Agricultural and Food Chemistry, v. 51, n. 10, p. 3029-3034, 2003.
VÁMOS-VIGYÁZO, L. Polyphenol oxidase and peroxidase in fruits and vegetables. Critical Reviews in Food Science and Nutrition, v. 15, n. 1, p. 49-127, set. 1981.
VAN HOOF, A.; LEYMAM, J.; SCHEFFER, H. J.; WALTON, J. D. A single beta-1,3-glucanase secreted by the maize pathogen C. carbonum acts by an exolytic mechanism. Physiological and Molecular Plant Pathology, Michigan, v. 39, n. 4, p. 259-267, out. 1991.
VANDERZANT, C.; SPLITTSTOESSER, D. F. Compendium of methods for the microbiological examination of foods. 3. ed. Washington: American Public Health Association, 1996. 873 p.
VANNUCCHI, H.; JORDÃO JUNIOR, A. A. Vitaminas hidrossolúveis. In: DUTRA-de-OLIVEIRA, J. E.; MARCHINI, J. S. Ciências nutricionais. São Paulo: Sarvier, 1998. p. 191-207.
VIEIRA, E. M. Estruturação de redes de cooperação para o desenvolvimento territorial rural: estudo de caso do programa Agroalt na região do Alto Tietê. 2008. 193f. Dissertação (Mestrado em Desenvolvimento Econômico). Instituto de Economia, Universidade Estadual de Campinas, Campinas, 2008.

106
VILELA, P. S. Cogumelos: mercado e comercialização. Belo Horizonte, 2006. Disponível em: <http:// www.faemg.org.br/Content.aspx?Code=353&ParentPath=None;13>. Acesso em: 26 jul. 2006.
WALDE, S. G. et al. Effects of pretreatments and drying methods on dehydration of mushroom. Journal of Food Engineering, v. 74, n. 1, p. 108-115, maio 2006.
WATERMAN, P. G.; MOLW, S.; MOLE, S. Analysis of phenolic plant metabolites, analysis of phenolic plant metabolites (Methods in Ecology). Oxford: Blackwell Scientific Publications, 1994. p. 238-239.
WISBECK, E.; ROBERT, A. P.; FURLAN, S. A. Avaliação da produção de agentes antimicrobianos por fungos do gênero Pleurotus. Health and Environment Journal, v. 3, n. 2, p. 7-10, 2002.
WOLFF, E. R. S. et al. Antimicrobial and antineoplasic activity of Pleurotus ostreatus. Applied Biochemistry and Biotechnology, v. 151, n. 2-3, p. 402-410, dez. 2008.
YANG, J.; LIN, H.; MAU, J. Non-volatile taste components of several commercial mushrooms. Food Chemistry, v. 72, n. 4, p. 465-471, mar. 2001.
YOSHIOKA, Y.; TABETA, R.; SAITO, H.; UEHARA, N.; FUKUOKA, F. Antitumor polysaccharides from P. ostreatus (Fr) Quél.: isolation and structure of a beta-glucan. Carbohydate Research, v. 140, p. 93-100, 1985.
XIE, J. et al. Inhibitory effects of substrate analogues on enzyme activity and substrate specificities of mushroom tyrosinas. Food Chemistry, v. 103, n. 4, p. 1075-1079, 2007.

107
ANEXO A
Teores de proteína (A) e fenol (B) liberados para os líquidos de lavagem. Comparação entre as amostras controle e pré-tratadas de Pleurotus ostreatus de tamanho pequeno, médio e grande.
Amostras mg SAB/mL (A)* mg ácido clorogenico/mL (B)*
Controle pequeno 0,80 +/-0,001c 0,020a médio 0,75 +/- 0,001d 0,035a grande 1,50 +/- 0,002e 0,040b
pré-tratado (fervura)
pequeno 0,23 +/- 0,05ª 0,850c médio 0,28 +/- 0,01a 0,560d grande 0,35 +/- 0,01b 0,960e
*Números são médias de 3 repetições. +/- corresponde a erro entre repetições para proteínas, enquanto para fenóis quase foi inexistente. Letras diferentes nas colunas, para cada grupo (in natura e fervido), são significativamente diferentes entre si ao nível de 0,05% (teste t Student). Letras iguais nas colunas, para cada grupo (in natura e fervido), não são significativamente diferentes entre si ao nível de 0,05% (teste t Student).

108
ANEXO B
Relação peso fresco/peso seco (g) e perda de água (%) do cogumelo Pleurotus. ostreatus de tamanhos grande e pequeno. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
*Média de 3 repetições com erro. Letras iguais entre si, dentro das colunas, entre cogumelos de mesmo tamanho, são significativamente iguais ao nível de 0,05% (teste t Student). Letras diferentes entre si, dentro das colunas, entre cogumelos de mesmo tamanho, são significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Com vácuo Sem vácuo
1gpf = peso seco* % perda de água 1gpf = peso seco* % perda
de água
Cogumelo grande
controle 0,1027 +/- 0,0001a 89,73 0,1027 +/- 0,0001a 89,73 pré-tratado 0,0998 +/- 0,0002b 90,02 0,0998 +/- 0,0002b 90,02 3dias 0,0996 +/- 0,0002c 90,04 0,0991 +/- 0,0003d 90,09 7dias 0,0998 +/- 0,0001e 90,02 0,0981 +/- 0,0005e 90,19 14dias 0,0984 +/- 0,0001f 90,16 0,0983 +/- 0,0004f 90,17 28dias 0,0991 +/- 0,0005g 90,09 0,0988 +/- 0,0001h 90,12
Cogumelo pequeno
controle 0,1079 +/- 0,0002a 89,21 0,1079 +/- 0,0002a 89,21 pré-tratado 0,1019 +/- 0,0003b 89,81 0,1019 +/- 0,0003b 89,81 3dias 0,1050 +/- 0,0002c 89,50 0,1052 +/- 0,0002c 89,48 7dias 0,1037 +/- 0,0003d 89,63 0,1043 +/- 0,0002e 89,57 14dias 0,1016 +/- 0,0002f 89,84 0,1016 +/- 0,0002f 89,84 28dias 0,1021 +/- 0,0004g 89,79 0,1026 +/- 0,0001h 89,74

109
ANEXO C
Perda de água (%) do Pleurotus ostreatus após os tempos de armazenamento com relação ao peso pós-tratamento. Comparação entre os cogumelos de tamanhos pequeno e grande embalados com e sem vácuo.
*Média de 3 repetições com erro. Letras iguais entre si, entre as colunas e linhas, para cada tamanho de cogumelo, são significativamente iguais ao nível de 0,05% (teste t Student). Letras diferentes entre si, entre as colunas e linhas, para cada tamanho de cogumelo, significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Com vácuo Sem vácuo
Peso pré-tratamento (g)*
Peso pós-drenagem (g)*
Peso pré-tratamento (g)*
Peso pós-drenagem (g)*
Cogumelo grande
3dias 39,89 +/- 0,012a 35,59 +/- 0,190a1 41,41 +/- 0,010b 38,29 +/- 0,032b1 7dias 38,77 +/- 0,020c 33,77 +/- 0,068c1 42,03 +/- 0,015d 38,70 +/- 0,200d1 14dias 41,81 +/- 0,026e 34,22 +/- 0,025e1 41,32 +/- 0,035f 37,30 +/- 0,021f1 28dias 38,85 +/- 0,025g 32,68 +/- 0,020g1 40,24 +/- 0,020h 36,61 +/- 0,141h1
Cogumelo pequeno
3dias 50,45 +/- 0,021a 44,40 +/- 0,200a1 52,02 +/- 0,015b 48,55 +/- 0,040b1 7dias 54,61 +/- 0,020c 48,60 +/- 0,015c1 55,99 +/- 0,027d 51,67 +/- 0,015d1 14dias 50,98 +/- 0,027e 44,40 +/- 0,200e1 52,91 +/- 0,031f 49,16 +/- 0,076f1 28dias 51,66 +/- 0,015g 44,54 +/- 0,040g1 50,65 +/- 0,010h 46,46 +/- 0,042h1

110
ANEXO D
Atividade da enzima polifenoloxidase (unidade de polifenoloxidase/minuto/g cogumelo fresco) e erda (%) da atividade enzimática em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
*Média de 3 repetições. Números seguidos de letras iguais entre as colunas, dentro do mesmo tamanho de cogumelo, não são significativamente diferentes ao nível de 0,05% (teste t Student, Anova). Números seguidos de letras diferentes entre as colunas, dentro do mesmo tamanho de cogumelo, são significativamente diferentes ao nível de 0,05% (teste t Student, Anova).
Amostras Com vácuo Sem vácuo
UnidadePolif/ gcog/min*
perda enzima (%)
UnidadePolif/ gcog/min*
perda enzima (%)
Cogumelo grande
controle 2,40a 100,00 2,40a 100,00 Pré-tratado 0,96b -60,00 0,96b -60,00 3dias 0,16c -93,34 0,32f -86,60 7dias 0,24d -90,00 0,38g -84,17 14dias 0,24d -90,00 0,40h -83,33 28dias 0,32e -86,66 0,52i -78,33
Cogumelo pequeno
controle 15,52a 100,00 15,52a 100,00 pré-tratado 0,08b -99,48 0,08b -99,48 3dias 0,08b -99,48 6,80d -56,18 7dias 0,08b -99,48 2,40e -84,50 14dias 0,08b -99,48 0,56f -96,39 28dias 0,10c -99,35 0,63g -95,94

111
ANEXO E
Atividade enzimática da peroxidase (unidade de peroxidase/minuto/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
* Média de 3 repetições. Números seguidos de letras iguais entre as colunas, dentro do mesmo tamanho de cogumelo, não são significativamente diferentes ao nível de 0,05% (teste t Student). Números seguidos de letras diferentes entre as colunas, dentro do mesmo tamanho de cogumelo, são significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Com vácuo* Sem vácuo*
Cogumelo grande
controle 0,640a 0,640a pré-tratado 0,160b 0,160b 3dias 0,016c 0,120f 7dias 0,016c 0,080g 14dias 0,240d 4,600h 28dias 0,800e 9,800i
Cogumelo pequeno
controle 0,520a 0,520a pré-tratado 0,160b 0,160b 3dias 0,016c 0,040e 7dias 0,016c 0,032f 14dias 0,016c 0,040e 28dias 0,040d 0,080g

112
ANEXO F
Valor (mg vit. C/100g cogumelo fresco) e mudanças (%) no teor de vitamina C em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
*Média de 3 repetições com erro. Letras iguais nas colunas, no mesmo tamanho de cogumelo, não são significativamente diferentes ao nível de 0,05% (teste t Student). Letras diferentes nas colunas, no mesmo tamanho de cogumelo, são significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Com vácuo Sem vácuo
mg vit. C/ 100g cogumelo*
Diferença de vit. C (%)
mg vit. C/ 100g cogumelo*
Diferença de vit. C (%)
Cogumelo grande
controle 28,1a +/- 0,21a 0,00 28,1a +/- 0,21a 0,00 pré-tratado 39,4b +/- 0,03b 40,20 39,4c +/- 0,03b 40,20 3dias 39,4b +/- 0,29b 40,21 33,8d +/- 0,46c -14,30 7dias 39,4b +/- 0,03b 40,21 22,5e +/- 0,28e -42,90 14dias 39,4b +/- 0,01b 40,21 22,5f +/- 0,28f -42,90 28dias 39,4b +/- 0,03b 40,21 21,9g +/- 0,05g -44,42
Cogumelo pequeno
controle 28,1a +/- 0,05a 0,00 28,1a +/- 0,05a 0,00 pré-tratado 34,0b +/- 0,38a 21,00 34,0b +/- 0,38a 21,00 3dias 35,5b +/- 0,38a 4,41 21,1c +/- 0,20b -37,94 7dias 35,5b +/- 1,15d 4,41 16,9d +/- 0,14c -50,29 14dias 36,0b +/- 0,57e 5,88 16,9d +/- 0,14c -50,29 28dias 35,5b +/- 0,34f 4,41 16,9d +/- 0,14c -50,29

113
ANEXO G
Teor de proteína (mg SAB/100g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
*Média de 3 repetições com erro. Letras iguais nas colunas, no mesmo tamanho de cogumelo, não são significativamente diferentes ao nível de 0,05% (teste t Student). Letras diferentes nas colunas, no mesmo tamanho de cogumelo, são significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Com vácuo* Sem vácuo*
Cogumelo grande
controle 3,4e +/- 0,057 3,4e +/- 0,057a pré-tratado 5,3f +/- 0,088b 5,3f +/- 0,088b 3dias 4,9d +/- 0,120d 3,9g +/- 0,120c 7dias 5,1d +/- 0,088f 4,3h +/- 0,057e 14dias 5,2d +/- 0,145h 3,8i +/- 0,202g 28dias 4,8d +/- 0,152j 2,2j +/- 0,133i
Cogumelo pequeno
controle 4,3a +/- 0,1201 4,3a +/- 0,1201a pré-tratado 3,9a +/- 0,1453b 3,9a +/- 0,1453b 3dias 3,8a +/- 0,1530d 3,3a +/- 0,1855c 7dias 3,8a +/- 0,8810f 3,5a +/- 0,1201e 14dias 3,6a +/- 0,8810h 2,8b +/- 0,1666g 28dias 3,5a +/- 0,5700j 2,8c +/- 0,8810i

114
ANEXO H
Teor de fenóis (mg ác. clorogênico/100g de cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
*Média de 3 repetições com erro. Letras iguais nas colunas, no mesmo tamanho de cogumelo, não são significativamente diferentes ao nível de 0,05% (teste t Student). Letras diferentes nas colunas, no mesmo tamanho de cogumelo, são significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Com vácuo* Sem vácuo*
Cogumelo grande
controle 0,87i +/- 0,014a 0,87i +/- 0,014a pré-tratado 0,63j +/- 0,006b 0,63j +/- 0,006b 3dias 0,50k +/- 0,029d 0,65h +/- 0,005c 7dias 0,56m +/- 0,011f 1,02l +/- 0,014e 14dias 0,69h +/- 0,005h 1,01n +/- 0,016g 28dias 0,87p +/- 0,006ª 1,27o+/- 0,018i
Cogumelo pequeno
controle 0,94b +/- 0,0120a 0,94b +/- 0,012a pré-tratado 0,51c +/- 0,037b 0,51c +/- 0,037b 3dias 0,46a +/- 0,0880d 0,62d +/- 0,023c 7dias 0,39e +/- 0,0880f 0,55a +/- 0,017e 14dias 0,36f +/- 0,0088g 0,55a +/- 0,025e 28dias 0,43g +/- 0,0088i 0,47a +/- 0,015h

115
ANEXO I
Contagem de colônias de leveduras (UFC/g cogumelo fresco) em Pleurotus ostreatus de tamanhos pequeno e grande. Comparação entre as amostras controle, pré-tratada e as embaladas com e sem vácuo em relação aos tempos de armazenamento.
*Média de 3 repetições com erro. Letras iguais nas colunas, no mesmo tamanho de cogumelo, não são significativamente diferentes ao nível de 0,05% (teste t Student). Letras diferentes nas colunas, no mesmo tamanho de cogumelo, são significativamente diferentes ao nível de 0,05% (teste t Student).
Amostras Contagem UFC/g Com vácuo* Sem vácuo*
Cogumelo grande
controle 0i 0i pré-tratado 0i 0i 3dias 2,50x101b +/- 1,201 2,30x102a +/-3,055 7dias 2,38x103d +/- 2,081 5,82x103c +/- 1,001 14dias 2,83x103f +/- 2,720 8,23x103e +/- 2,081 28dias 1,13x104h +/- 3,750 1,85x104g +/- 0,880
Cogumelo pequeno
controle 0i 0i pré-tratado 0i 0i 3dias 2a +/- 0,577 1,36x103b +/- 1,76 7dias 2a +/- 0,333 1,50x103c +/- 20,81 14dias 2,8x102f +/- 21,660 2,50x103e +/- 11,54 28dias 3,0x102h +/- 5,770 3,85x103g+/- 12,01

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo







